Фототроф • ru.knowledgr.com
Террестриальные и водные фототрофы: растения растут на упавшем бревне, плавающем в богатой алгами воде
Фототрофы (Гр: ф, ф Фототрофами могут быть либо автотрофы, либо гетеротрофы. Если их электрон и гидрогены являются неорганическими соединениями (например, как в некоторых le sulfur teria, или, как в некоторых зеленых sulfur teria), они также могут называться отрофами, и так, некоторые фотоавтотрофы также называются фотототрофами. Примерами фототрофных организмов являются: Rhodobacter capsul , Chromatium, Chlorobium и т.д.
История
Первоначально использовавшийся с другим значением, термин взял своё нынешнее определение после Lwoff и коллаборационистов (1946).
Фотоавтопроф
Большинство хорошо известных фототрофов являются автотрофными, также известными как фототрофы, и могут фиксировать углерод. Они могут быть с хемотрофами, которые получают свою энергию за счет окисления доноров в окружающей их среде. Фотоавтотрофы способны синтезировать собственную пищу из неорганических веществ, используя свет в качестве источника энергии. Зелёные растения и фотосинтетические терии являются фотоавтотрофами. Фотоавтотрофные организмы иногда называют голофитными. Такие организмы снижать свою энергию для синтеза пищи из света и способны использовать диоксид углерода в качестве своего основного источника углерода.
Они могут быть с хемотрофами, которые получают свою энергию за счет окисления доноров в окружающей их среде. Фотоавтотрофы способны синтезировать собственную пищу из неорганических веществ, используя свет в качестве источника энергии. Зелёные растения и фотосинтетические терии являются фотоавтотрофами. Фотоавтотрофные организмы иногда называют голофитными. Такие организмы снижать свою энергию для синтеза пищи из света и способны использовать диоксид углерода в качестве своего основного источника углерода.
Оксигенные фотосинтетические организмы используют хлорофилл для улавливания световой энергии и оксидизации воды, » превращая» его в молекулярный оксиген. Напротив, аноксигенные фотосинтетические терии имеют вещество, называемое териохлорофилл — которое абсорбирует преимущественно при неоптических вавельенгетах — для захвата световой энергии, живут в водной среде, в виде сернистой воды, а не будет, а будет.
Экология
В экологическом контексте фототрофы часто являются источником пищи для соседней гетеротрофной жизни. В террестриальной среде растения являются преобладающим разнообразием, в то время как водные среды включают ряд фототрофных организмов, таких как алги (например, ламинария), другие протисты (такие как эуглена), фитопланктон и терии (такие как цианобактерия). Глубина, на которую солнечный свет или искусственный свет может проникать в воду, так что может происходить фотосинтез, известна как фотическая зона.
В террестриальной среде растения являются преобладающим разнообразием, в то время как водные среды включают ряд фототрофных организмов, таких как алги (например, ламинария), другие протисты (такие как эуглена), фитопланктон и терии (такие как цианобактерия). Глубина, на которую солнечный свет или искусственный свет может проникать в воду, так что может происходить фотосинтез, известна как фотическая зона.
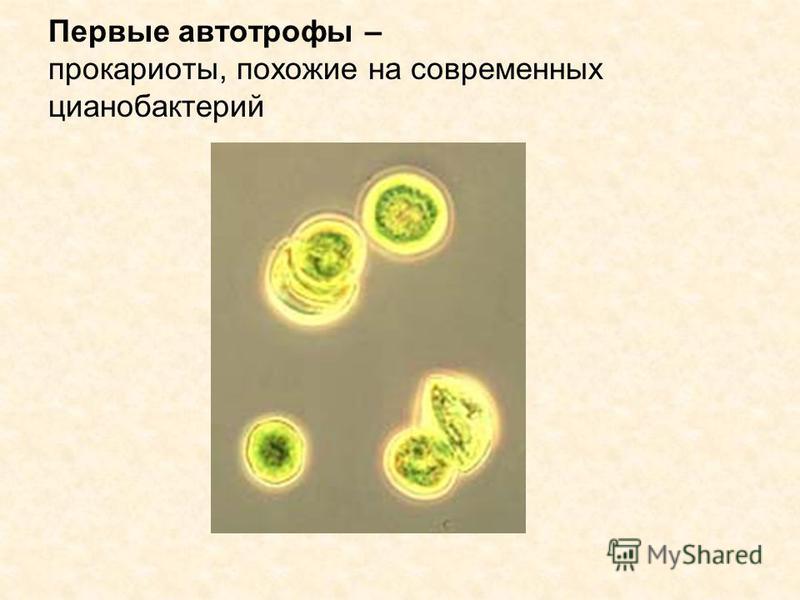
Cyanobacteria, которые являются прокариотическими организмами, которые осуществляют оксигенный фотосинтез, занимают многие условия окружающей среды, включая пресную воду, море, почву и лихен. Cyanobacteria осуществляют растительный фотосинтез, потому что органелла в растениях, которые осуществляют фотосинтез, выделена из эндосимбиотического cyanobacterium. Этот терий может использовать воду в качестве источника электронов для проведения реакций восстановления CO2. Эволюционно способность cyanobacteria в оксигенных состояниях, которые считаются токсичными для большинства анаэробных teria, могла бы дать teria адаптивное преимущество, которое могло бы позволить cyanobacteria заселяться более эффективно.
Фототототрофом является автотрофный организм, который использует световую энергию, и неорганический донор (например, h3O, h3, h3S) и CO2 в качестве источника углерода. Примеры включают растения.
Фотогетеротроф
В отличие от фотоавтотрофов, фотогетеротрофы представляют собой организмы, которые зависят исключительно от света в отношении их энергии и главным образом от органических соединений в отношении их углерода. Фотогетеротрофы продуцируют АТФ посредством фотофосфорилирования, но используют полученные в окружающей среде органические соединения для построения структур и других биомольекул.
Блок-схема
Блок-схема для определения, является ли вид автотрофом, гетеротрофом или подтипом
См. также
Растения Автотрофы Фотосинтетики Редко миксотрофы или гетеротрофы Клеточная
Растения Автотрофы. Фотосинтетики. Редко миксотрофы или гетеротрофы Клеточная стенка из целлюлозы Запасающее вещество – крахмал Рост в течение всей жизни Регуляция процессов в организме с помощью фитогормонов (гуморальная) Подвижность ограничена Тело ветвится Чередование поколений (спорофит и гаметофит)
Строение клетки растений biology. ru
ru
Растения Низшие Высшие • Тело не расчленено на органы • Нет тканей • Тело называется таллом (слоевище) • Есть органы • Ткани сложно дифференцированы Водоросли Споровые Моховидные, Плауновидные, Хвощевидные, Папоротниковидные Семенные Голосеменные, Покрытосеменные
ВОДОРОСЛИ
Группа отделов Водоросли Обитают в основном в воде, редко на поверхности почвы, влажным камнях, коре деревьев Есть одноклеточные, многоклеточные, колониальные У некоторых одноклеточных водорослей есть светочувствительный глазок (стигма) Эвглена зеленая
Отдел Зеленые водоросли Хроматофор – хлоропласт водорослей Пигменты: хлорофилл Зеленый цвет таллома Организация: Одноклеточные Колониальные Многоклеточные (в том числе нитчатые)
Представители Верхний ряд, слева направо: хламидомонада, хлорелла, микрастериас, сценедесмус двуформенный, вольвокс. Нижний ряд, слева направо: спирогира, улотрикс, ульва, каулерпа, кладофора
Одноклеточные зеленые водоросли Хламидомонада Хлорелла
Хламидомонада Строение клетки Жизненный цикл
Хлорелла Строение клетки митоз Освобождение дочерних клеток Жизненный цикл
Колониальная зеленая водоросль Вольвокс
Нитчатые зеленые водоросли спирогира улотрикс
Конъюгация спирогиры
Улотрикс. Жизненный цикл
Жизненный цикл
Многоклеточные зеленые водоросли Ульва
Значение очистка сточных вод пища для животных могут вступать в симбиоз с животными (губками, кишечнополостными) и грибами формирование сапропеля (ила) изготовления бумаги (кладофора) вызывают цветение воды корм для домашних животных используются в пищу
Отдел Красные водоросли (Багрянки) В основном морские, могут обитать на больших глубинах (до 200) Пигменты: хлорофиллы каротиноиды фикобилины Окраска от ярко-красной до голубовато-зеленой и желтой Нитевидные, пластинчатые, цилиндрические, корковидные, в виде шнуров, разветвлённых кустиков Прикреплены к грунту с помощью ризоидов
Представители Пальмария (Palmaria palmata) Devaleraea ramentacea Родомела (Rhodomela lycopodioides)
Представители порфира лопастная немалион червеобразный полиидес округлый литотамний кораллина целебная а — внешний вид, б — отдельная ветвь, увел.
Значение В природе Для человека Наряду с кораллами участвуют в рифообразовании Пища для морских животных Получение агара Пищевая и медицинская промышленность
Отдел Бурые водоросли Приемущественно морские Одноклеточные (неск. мм) и многоклеточные (десятки метров) Пигменты: Желтовато-бурая окраска Запасающее вещество – ламинарин хлорофиллы a и с бета-каротин фукоксантин нитевидные корковидные шаровидные пластинчатые кустообразные У некоторых есть газовые пузыри для поддержания таллома в вертикальном состоянии Слоевище из нескольких слоев клеток Прикреплены к грунту с помощью ризоидов
Представители Верхний ряд, слева направо: фукус постелсия пальмовидная Макроцистис саргассум Нижний ряд, слева направо: ламинария аналипус японский пельвеция пучковатая цистозейра
Представители хорда нитевидная хорда опушенная ламинария сахаристая ламинария пальчаторассечен ная алярия Деляпиля
Ламинария
Значение Место обитания, укрытие, питание для многих водных организмов Альгинаты используются для производства консервов, соков, красящих и клеящих веществ, смазок паст, мазей Фармацевтика Парфюмерия Пища Медицина (заменитель сахара и кровезаменитель — маннит) Корм для животных Получение йода и других микроэлементов
Фототроф
Организм использует энергию света в обменных процессах
Эта статья о фототрофизме, получении энергии от света. Для тропизм который управляет ростом к источнику света или от него, см. Фототропизм. Наземные и водные фототрофные растения растут на упавшем бревне, плавающем в воде, богатой водорослями.
Для тропизм который управляет ростом к источнику света или от него, см. Фототропизм. Наземные и водные фототрофные растения растут на упавшем бревне, плавающем в воде, богатой водорослями.Фототрофы (Gr: φῶς, φωτός = свет, τροϕή = питание) организмы которые осуществляют захват фотонов для производства сложных органических соединений (например, углеводов) и получения энергии. Они используют энергия из свет осуществлять различные клеточные метаболические процессы. Распространенное заблуждение, что фототрофы обязательно фотосинтетический. Многие, но не все, фототрофы часто фотосинтезируют: они анаболически конвертировать углекислый газ в органический материал, который будет использоваться структурно, функционально или в качестве источника для дальнейшего использования катаболический процессы (например, в виде крахмала, сахаров и жиров). Все фототрофы используют цепи переноса электронов или прямой протонная накачка для установления электрохимического градиента, который используется АТФ-синтаза, чтобы обеспечить валюту молекулярной энергии для клетки.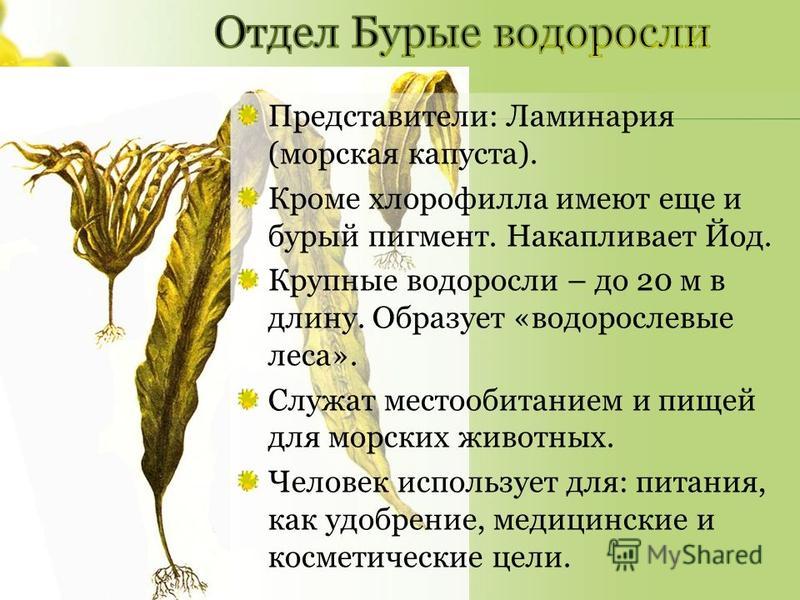 Фототрофы могут быть как автотрофы или же гетеротрофы. Если их доноры электронов и водорода являются неорганическими соединениями (например, Na
Фототрофы могут быть как автотрофы или же гетеротрофы. Если их доноры электронов и водорода являются неорганическими соединениями (например, Na
2S
2О
3, как в некоторых фиолетовые серные бактерии, или же ЧАС
2S, как в некоторых зеленые серные бактерии) их также можно назвать литотрофы, поэтому некоторые фотоавтотрофы еще называют фотолитоавтотрофами. Примеры фототрофных организмов: Rhodobacter capsulatus, Chromatium, Хлоробий и т.п.
История
Первоначально использовавшийся в другом значении, термин получил свое текущее определение после Lwoff и сотрудники (1946).[1][2]
Фотоавтотроф
Большинство известных фототрофов автотрофный, также известный как фотоавтотрофы, и может исправить углерод. Их можно противопоставить хемотрофы которые получают свою энергию окисление из доноры электронов в их среде. Фотоавтотрофы способны синтезировать собственную пищу из неорганических веществ, используя свет в качестве источника энергии. Зеленые растения и фотосинтезирующие бактерии — фотоавтотрофы. Фотоавтотрофные организмы иногда называют голофитный.[3] Такие организмы получают энергию для синтеза пищи из света и способны использовать углекислый газ в качестве основного источника углерода.
Зеленые растения и фотосинтезирующие бактерии — фотоавтотрофы. Фотоавтотрофные организмы иногда называют голофитный.[3] Такие организмы получают энергию для синтеза пищи из света и способны использовать углекислый газ в качестве основного источника углерода.
Кислородные фотосинтезирующие организмы используют хлорофилл для улавливания световой энергии и окисления воды, «расщепление» это в молекулярный кислород. Напротив, аноксигенные фотосинтезирующие бактерии имеют вещество, называемое бактериохлорофилл — который поглощает преимущественно на неоптических длинах волн — для захвата световой энергии, живет в водной среде и, используя свет, окисляет химические вещества, такие как сероводород а не вода.
Экология
В экологический В контексте, фототрофы часто являются источником пищи для соседней гетеротрофной жизни. В земных условиях, растения являются преобладающим разнообразием, в то время как водная среда включает ряд фототрофных организмов, таких как водоросли (например. , ламинария), Другой протисты (такие как эвглена), фитопланктон, и бактерии (такие как цианобактерии). Глубина, на которую солнечный или искусственный свет может проникать в воду, что может вызвать фотосинтез, известна как фотическая зона.
, ламинария), Другой протисты (такие как эвглена), фитопланктон, и бактерии (такие как цианобактерии). Глубина, на которую солнечный или искусственный свет может проникать в воду, что может вызвать фотосинтез, известна как фотическая зона.
Цианобактерии, которые являются прокариотическими организмами, осуществляющими оксигенный фотосинтез, обитают во многих условиях окружающей среды, включая пресную воду, моря, почва, и лишайник. Цианобактерии осуществляют фотосинтез, подобный растениям, потому что органелла в растениях, осуществляющих фотосинтез, происходит из[4] эндосимбиотические цианобактерии.[5] Эта бактерия может использовать воду в качестве источника электроны для выполнения CO2снижение реакции. Эволюционно способность цианобактерий выживать в оксигенный условия, которые считаются токсичными для большинства анаэробный бактерии, возможно, дали бактериям адаптивное преимущество, которое позволило бы цианобактериям более эффективно заселять их. Кэмпбелл, Нил А .; Рис, Джейн Б.; Урри, Лиза А .; Каин, Майкл Л .; Вассерман, Стивен А .; Минорский, Петр V .; Джексон, Роберт Б. (2008). Биология (8-е изд.). п. 564. ISBN 978-0-8053-6844-4.
Кэмпбелл, Нил А .; Рис, Джейн Б.; Урри, Лиза А .; Каин, Майкл Л .; Вассерман, Стивен А .; Минорский, Петр V .; Джексон, Роберт Б. (2008). Биология (8-е изд.). п. 564. ISBN 978-0-8053-6844-4.
Ламинария, бурая водоросль – Inanimate Life
Like Focus ( каменистое растение ) Laminaria является представителем интересной группы организмов, которые обычно встречаются в приливной зоне и выбрасываются на пляжи. Большинство организмов, называемых «морскими водорослями», представляют собой бурые водоросли, хотя некоторые из них являются красными водорослями, а некоторые — зелеными водорослями. Как и большинство (но не все) бурых водорослей, ламинария представляет собой крупный многоклеточный организм, хорошо приспособленный к жизни в приливных и мелководных прибрежных водах, чаще всего в относительно прохладных водах.
Таксономия и филогения Бурые водоросли представляют собой небольшую группу эукариот, которых традиционно относили к их собственному типу Phaeophyta, иногда вместе с двумя группами преимущественно одноклеточных водорослей, золотыми водорослями и желто-зелеными водорослями, на основании сходства пигментации и других факторов. . Большинство современных методов лечения не возводят эту группу на уровень типа, а объединяют ее с другими «гетероконтами», группой, определяемой наличием двух характерных жгутиков, один длиннее другого.Гетероконты (иногда называемые Stramenopiles) также включают диатомовые водоросли и гетеротрофные водные формы (Oomycota). Пигментация фотосинтезирующих страменопилей аналогична пигментации гаптофитов, таких как
. Большинство современных методов лечения не возводят эту группу на уровень типа, а объединяют ее с другими «гетероконтами», группой, определяемой наличием двух характерных жгутиков, один длиннее другого.Гетероконты (иногда называемые Stramenopiles) также включают диатомовые водоросли и гетеротрофные водные формы (Oomycota). Пигментация фотосинтезирующих страменопилей аналогична пигментации гаптофитов, таких как
Типична для большинства бурых водорослей Ламинария действительно многоклеточная и имеет три различных многоклеточных органа: фиксатор, который прикрепляет организм к субстрату, широкую плоскую пластинку, которая осуществляет основную часть фотосинтеза, и ножку (стебель ), который соединяет лезвие с держателем и имеет достаточную длину, чтобы лезвие могло получать свет.
Большинство бурых водорослей являются половыми и демонстрируют чередование поколений. «Доминирующей» (т. е. более крупной и заметной) стадией обычно является стадия спорофита (диплоидная), но есть некоторые бурые водоросли, у которых наблюдается изоморфное чередование поколений (спорофит и гаметофит выглядят одинаково), а у некоторых доминирует стадия гаметофита.Другая бурая водоросль, каменная водоросль ( Фукус) , демонстрирует жизненный цикл, подобный человеческому, без чередования поколений и где единственными гаплоидными клетками являются гаметы.
Ламинария является типичным фотоавтотрофом, использующим энергию солнечного света для синтеза углеводов из углекислого газа, а затем использующим углеводы в качестве источника энергии для клеточного дыхания и в качестве строительного материала для синтеза различных биомолекул.Бурые водоросли производят особую форму полисахарида для хранения углеводов, называемого ламинараном, состоящего из единиц глюкозы, соединенных связью бета 1-3, а не связью альфа ,1-4, обнаруженной в крахмале. Они также имеют высокие концентрации маннита, который служит транспортным углеводом, роль которого в большинстве растений выполняет сахароза.
Они также имеют высокие концентрации маннита, который служит транспортным углеводом, роль которого в большинстве растений выполняет сахароза.
Ламинария является важным представителем прохладных мелководных прибрежных вод.Наряду с другими представителями бурых водорослей они образуют «леса ламинарии», обеспечивая пищу для ряда организмов и среду обитания для других. Роквей ( Фукус ) особенно важен в приливной зоне по тем же причинам.
Является ли водоросль автотрофом? – Restaurantnorman.com
Является ли водоросль автотрофом?
Водоросли, наряду с растениями и некоторыми бактериями и грибами, являются автотрофами. Автотрофы являются производителями в пищевой цепи, то есть они создают свои собственные питательные вещества и энергию.
Водоросли автотрофы или гетеротрофы Почему?
 Гетеротрофы, наоборот, питаются другими организмами и органическими материалами в своих…
Гетеротрофы, наоборот, питаются другими организмами и органическими материалами в своих…Зеленые водоросли гетеротрофны или автотрофны?
Зеленые водоросли автотрофны.
Все ли водоросли фотосинтезируют?
Не все водоросли имеют хлоропласты и фотосинтез. «Бесцветные» водоросли могут получать энергию и пищу, окисляя органические молекулы, которые они поглощают из окружающей среды или переваривают из поглощенных частиц.
Какие 3 типа водорослей существуют?
Макроводоросли подразделяются на три основные группы: бурые водоросли (Phaeophyceae), зеленые водоросли (Chlorophyta) и красные водоросли (Rhodophyta).
Почему водоросли не растения?
Почему водоросли считаются растительноподобными? Основная причина в том, что они содержат хлоропласты и производят пищу посредством фотосинтеза. Однако у них отсутствуют многие другие структуры настоящих растений. Например, у водорослей нет ни корней, ни стеблей, ни листьев.
Водоросли — это растение или животное?
Водоросли иногда считают растениями, а иногда считают «протистами» (категория обычно отдаленно родственных организмов, сгруппированных на основании того, что они не являются животными, растениями, грибами, бактериями или археями).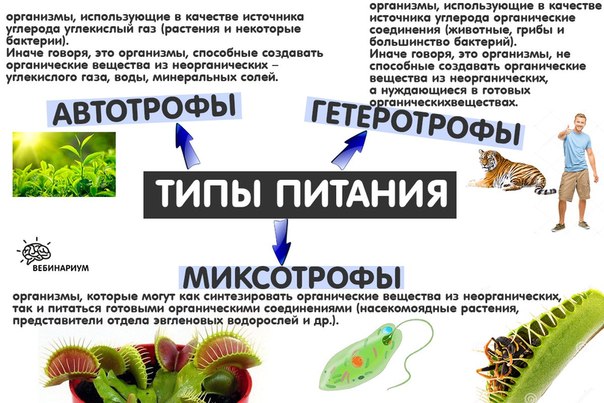
Где чаще всего встречаются водоросли?
Водоросли – это водные растительноподобные организмы. Они включают множество простых структур, от одноклеточного фитопланктона, плавающего в воде, до крупных морских водорослей (макроводорослей), прикрепленных к дну океана 2. Водоросли можно найти в океанах, озерах, реках, прудах и даже в снегу, где угодно на земле.
Водоросли бактерии или эукариоты?
Наконец, водоросли встречаются в различных водных средах обитания, как пресноводных, так и соленых.В силу этих характеристик общий термин «водоросли» включает прокариотические организмы — цианобактерии, также известные как сине-зеленые водоросли, — а также эукариотические организмы (все другие виды водорослей).
Является ли водоросль водорослью?
Несмотря на то, что морские водоросли являются одним из видов водорослей, они имеют серьезные различия с точки зрения истории, разнообразия видов, мест обитания и даже клеточной структуры. В то время как водоросли растут в морских водоемах, море и даже в пресных водоемах, морские водоросли растут только в море.
Морские водоросли — это растение или животное?
Вопреки тому, что мы можем предположить, водоросли не являются растениями. Это может выглядеть так, но у растений есть корни, а у водорослей нет. Морские водоросли — это водоросли, поэтому другие названия морских водорослей включают «морские водоросли». Морские водоросли растут в океанах, озерах и реках.
Какое животное питается водорослями?
КАКИЕ ЖИВОТНЫЕ ЕДЯТ ВОДОРОСЛИ? Многие беспозвоночные едят морские водоросли, такие как медузы, крабы, ракообразные, морские ежи, тюлени, морские черепахи, омары, раки, мокрицы и многие другие.
Едят ли креветки водоросли?
Едят ли креветки водоросли? Креветки не очень часто едят морские водоросли; Однако, если он доступен, они могут решить его потреблять. В частности, если водоросли достигают дна воды и никакие другие животные их не съели, креветки могут съесть их при необходимости.
Едят ли морские ежи водоросли?
Морские ежи питаются с помощью устройства, называемого фонарем Аристотеля. Морские ежи едят почти все, что проплывает мимо. Его острые зубы могут соскребать водоросли с камней и перемалывать планктон, ламинарию, барвинок, а иногда даже ракушек и мидий.
Морские ежи едят почти все, что проплывает мимо. Его острые зубы могут соскребать водоросли с камней и перемалывать планктон, ламинарию, барвинок, а иногда даже ракушек и мидий.
Какое травоядное ест водоросли?
В экосистеме океана многие виды рыб и черепах являются травоядными, питающимися водорослями и водорослями. В ламинариевых лесах водоросли, известные как гигантские ламинарии, обеспечивают убежище и пищу для всей экосистемы. Морские ежи являются основными потребителями водорослей в ламинариях.
Каковы 3 примера хищников?
Примеры плотоядных животных Одними из самых смертоносных хищных животных являются рыси, ястребы, горные львы, рыжие лисы, рыбаки, росомахи, койоты, орлы и многие другие.Кроме того, наиболее узнаваемыми являются тигры, гепарды, львы и гиены. Важно понимать, что все хищники не обязательно млекопитающие.
Является ли черепаха травоядным?
Некоторые черепахи являются плотоядными, в то время как другие придерживаются строго вегетарианской диеты. Однако большинство черепах всеядны, они едят как животных, так и растения. Зеленые морские черепахи (Chelonia myadis), с другой стороны, являются травоядными, которые питаются водорослями и водорослями.
Однако большинство черепах всеядны, они едят как животных, так и растения. Зеленые морские черепахи (Chelonia myadis), с другой стороны, являются травоядными, которые питаются водорослями и водорослями.
Что черепахи могут есть из человеческой пищи?
Измельченная морковь, кабачки и цуккини — отличная еда, которую могут есть и черепахи.Вы также можете использовать съедобную водную растительность, такую как водяной салат, водяной гиацинт и ряску. «Из фруктов рассмотрите тертые яблоки и дыни, а также нарезанные ягоды», — рекомендует доктор
.Какие животные едят черепах?
Плотоядные млекопитающие
- Еноты. Еноты, в частности, наслаждаются возможностью совершить набег на черепашье гнездо или перекусить свежими вылупившимися черепахами.
- Опоссумы. Опоссумы — еще одно распространенное оппортунистическое всеядное животное, которое часто поедает молодых черепах и черепашьи яйца.
- Скунсы и другие куньи.
- Лисы и другие псовые.

- Ящерицы.
- Змеи.
Как долго могут жить черепахи?
Черепахи и черепахи являются одними из самых долгоживущих представителей семейства рептилий. Даже мелкие виды, которых обычно держат в качестве домашних животных, например коробчатые черепахи и морские черепахи, живут от 30 до 40 лет, если их держат здоровыми. По оценкам, более крупные виды, такие как морские черепахи, живут около 80 лет.
Какое животное живет дольше всех?
Гренландский кит
Могут ли черепахи жить до 500 лет?
Некоторые черепахи могут прожить в неволе до 80 лет, но Общество охраны черепах говорит, что, возможно, некоторые более крупные черепахи тоже живут сотни лет.
Сколько лет самой старой черепахе?
около 189 лет
холобионтов морских водорослей: понимание взаимодействия водорослей и бактерий | Обзоры микробиологии FEMS
 Эти инженеры экосистем вносят значительный вклад в глобальную первичную продукцию и являются основными формирователями среды обитания на скалистых берегах в водах с умеренным климатом, обеспечивая пищу и убежище для водных организмов.Как и другие эукариотические организмы, макроводоросли содержат богатое разнообразие ассоциированных микроорганизмов, функции которых связаны со здоровьем и защитой хозяина. В частности, сообщается, что эпифитные бактериальные сообщества необходимы для нормального морфологического развития водоросли-хозяина, а бактерии с противообрастающими свойствами, как считается, защищают химически незащищенные макроводоросли от вредной вторичной колонизации другими микроскопическими и макроскопическими эпибиотами. Эта тесная связь предполагает, что макроводоросли и эпифитные бактерии взаимодействуют как единая функциональная сущность или холобионт, аналогично ранее предполагаемой взаимосвязи у кораллов.Кроме того, учитывая, что воздействие болезней на морские экосистемы, по-видимому, увеличивается, понимание роли бактерий как сапрофитов и патогенов в сообществах морских водорослей может иметь важные последствия для стратегий управления морской средой.
Эти инженеры экосистем вносят значительный вклад в глобальную первичную продукцию и являются основными формирователями среды обитания на скалистых берегах в водах с умеренным климатом, обеспечивая пищу и убежище для водных организмов.Как и другие эукариотические организмы, макроводоросли содержат богатое разнообразие ассоциированных микроорганизмов, функции которых связаны со здоровьем и защитой хозяина. В частности, сообщается, что эпифитные бактериальные сообщества необходимы для нормального морфологического развития водоросли-хозяина, а бактерии с противообрастающими свойствами, как считается, защищают химически незащищенные макроводоросли от вредной вторичной колонизации другими микроскопическими и макроскопическими эпибиотами. Эта тесная связь предполагает, что макроводоросли и эпифитные бактерии взаимодействуют как единая функциональная сущность или холобионт, аналогично ранее предполагаемой взаимосвязи у кораллов.Кроме того, учитывая, что воздействие болезней на морские экосистемы, по-видимому, увеличивается, понимание роли бактерий как сапрофитов и патогенов в сообществах морских водорослей может иметь важные последствия для стратегий управления морской средой. В этом обзоре сообщается о последних достижениях в понимании взаимодействий между макроводорослями и бактериями со ссылкой на разнообразие и функциональную роль эпифитных бактерий в поддержании здоровья водорослей с акцентом на концепцию холобионтов.
В этом обзоре сообщается о последних достижениях в понимании взаимодействий между макроводорослями и бактериями со ссылкой на разнообразие и функциональную роль эпифитных бактерий в поддержании здоровья водорослей с акцентом на концепцию холобионтов. , 2008; Egan и др. ., 2008 г.; Кроуфорд и Кларди, 2011 г.; Wahl и др. ., 2012).
, 2008; Egan и др. ., 2008 г.; Кроуфорд и Кларди, 2011 г.; Wahl и др. ., 2012).
, 2008; Egan и др. ., 2008 г.; Кроуфорд и Кларди, 2011 г.; Wahl и др. ., 2012). Морские водоросли или морские макроводоросли представляют собой сидячие многоклеточные фотосинтезирующие эукариоты, которые отличаются от растений отсутствием специализированных тканей (например, корневой системы и сосудистых структур) (Graham & Wilcox, 1999). Записи окаменелостей макроводорослей датируются более чем 1200 миллионами лет, предшествуя эволюции наземных растений, а в случае красных водорослей Bangiomorpha sp.представляют собой древнейший таксономически разрешенный многоклеточный организм (Butterfield, 2000). Сегодня макроводоросли играют важную роль в создании экосистем на скалистых берегах в прибрежной морской среде с умеренным климатом. Здесь они вносят большой вклад в первичную продуктивность и определяют физическую структуру среды обитания (Schiel & Foster, 2006). Они позволяют поддерживать местное биоразнообразие (Schiel, 2006; Schiel & Lilley, 2007), служат рассадниками и защитными убежищами для многих видов беспозвоночных и обеспечивают необходимое пространство для эпибионтов, от бактерий до макробеспозвоночных (Wilson et al .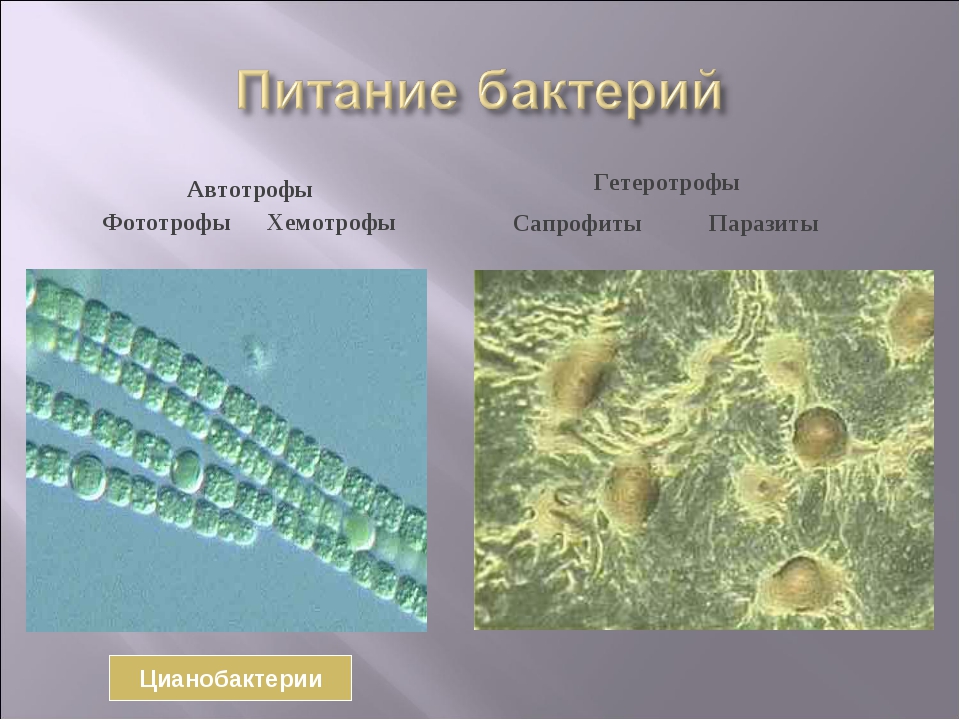 , 1990; Bulleri и др. ., 2002). В коммерческом контексте аквакультура макроводорослей за последние несколько лет расширилась, в частности, для азиатского продовольственного рынка и в качестве сырья для производства биотоплива (Neori, 2009; Borines et al ., 2011).
, 1990; Bulleri и др. ., 2002). В коммерческом контексте аквакультура макроводорослей за последние несколько лет расширилась, в частности, для азиатского продовольственного рынка и в качестве сырья для производства биотоплива (Neori, 2009; Borines et al ., 2011).
В основе этого обзора лежит утверждение о том, что макроводоросли, функционирующие как в экологических, так и в промышленных условиях, невозможно понять без учета взаимодействия с ассоциированным с ними микробиомом. Имеются убедительные лабораторные доказательства того, что здоровье, производительность и устойчивость макроводорослей функционально регулируются и частично поддерживаются эпифитными бактериями.Эта функциональная помощь подразумевает, что макроводоросли и вся связанная с ними микробиота образуют единую сущность или холобионт (рис. 2), в соответствии с тем, что было предложено для кораллового холобионта (Rosenberg et al ., 2007; Bourne et al . , 2009). На самом деле Barott и др. . (2011) недавно предположили, что это взаимодействие может быть настолько важным для тропических рифовых водорослей, что они аналогичным образом предложили концепцию водорослей-холобионтов для этих систем.
. (2011) недавно предположили, что это взаимодействие может быть настолько важным для тропических рифовых водорослей, что они аналогичным образом предложили концепцию водорослей-холобионтов для этих систем.
Концепция холобионта предполагает необходимость коллективного взгляда на все взаимодействия и действия внутри и между хозяином и всеми связанными с ним организмами.Знания о многих отдельных аспектах этих взаимодействий быстро расширились за последние несколько лет (последние обзоры см. в Gachon et al ., 2010; Goecke et al ., 2010; Hollants et al ., 2012; Wahl ). et al ., 2012), включая химическое взаимодействие между бактериями и водорослями-хозяевами (Goecke et al ., 2010; Wahl et al ., 2012), бактериальное разнообразие (Hollants et al ., 2012), и микробные заболевания водорослей (Gachon et al ., 2010). Здесь мы сосредоточимся на современных знаниях о разнообразии и взаимодействии бактерий, связанных с морскими макроводорослями. В частности, мы рассмотрим, какие бактерии могут способствовать «холобионту» и какие факторы окружающей среды влияют на поддержание, стабильность и установление таких взаимодействий. Затем мы обсудим функциональные результаты этих взаимодействий и то, как стресс окружающей среды может привести к потере функции холобионта. Наконец, мы обращаемся к потенциальной роли небактериальных представителей холобионта морских водорослей и обсуждаем будущие направления и возможности исследований.
В частности, мы рассмотрим, какие бактерии могут способствовать «холобионту» и какие факторы окружающей среды влияют на поддержание, стабильность и установление таких взаимодействий. Затем мы обсудим функциональные результаты этих взаимодействий и то, как стресс окружающей среды может привести к потере функции холобионта. Наконец, мы обращаемся к потенциальной роли небактериальных представителей холобионта морских водорослей и обсуждаем будущие направления и возможности исследований.
 , 2000). ; Бенгтссон и др. ., 2010). Анализ изображения микробного сообщества, связанного с поверхностью Ulva australis , показывает, что плотность бактерий увеличивается на порядок от кончиков слоевища (10 6 клеток см -2 ) к основанию водорослей (10 7 клетки см -2 ) (Tujula, 2006; рис. 1). Еще в 1970-х годах исследования, основанные на культивировании и микроскопии, выявили четкие различия между микробным составом, связанным с макроводорослями, и составом окружающей морской воды, между разными видами водорослей, в разные времена года, а также между разными участками слоевища макроводорослей (Cundell и др. ., 1977; Bolinches и др. ., 1988). Эти наблюдения специфичности хозяина, а также временных и пространственных вариаций были дополнительно уточнены рядом недавних исследований, независимых от культуры (см. Вспомогательную информацию, Таблица S1).
, 2000). ; Бенгтссон и др. ., 2010). Анализ изображения микробного сообщества, связанного с поверхностью Ulva australis , показывает, что плотность бактерий увеличивается на порядок от кончиков слоевища (10 6 клеток см -2 ) к основанию водорослей (10 7 клетки см -2 ) (Tujula, 2006; рис. 1). Еще в 1970-х годах исследования, основанные на культивировании и микроскопии, выявили четкие различия между микробным составом, связанным с макроводорослями, и составом окружающей морской воды, между разными видами водорослей, в разные времена года, а также между разными участками слоевища макроводорослей (Cundell и др. ., 1977; Bolinches и др. ., 1988). Эти наблюдения специфичности хозяина, а также временных и пространственных вариаций были дополнительно уточнены рядом недавних исследований, независимых от культуры (см. Вспомогательную информацию, Таблица S1).
Рисунок 1
Бактериальное поверхностное сообщество макроводоросли-хозяина. Микроскопические изображения, показывающие бактериальное сообщество на дистальных концах (а), срединном слоевища (б) и основании (в) срезов зеленой водоросли Ulva australis .Бактерии выявляли с помощью конфокальной микроскопии с использованием CARD-FISH. Все бактерии (зеленый), Alphaproteobacteria (красный). Масштабные полосы представляют собой длину 10 мкм. Эти изображения были сделаны в Центре морских биоинноваций Университета Нового Южного Уэльса, Австралия, доктором Нииной Туджулой.
Микроскопические изображения, показывающие бактериальное сообщество на дистальных концах (а), срединном слоевища (б) и основании (в) срезов зеленой водоросли Ulva australis .Бактерии выявляли с помощью конфокальной микроскопии с использованием CARD-FISH. Все бактерии (зеленый), Alphaproteobacteria (красный). Масштабные полосы представляют собой длину 10 мкм. Эти изображения были сделаны в Центре морских биоинноваций Университета Нового Южного Уэльса, Австралия, доктором Нииной Туджулой.
Рисунок 1
Бактериальное поверхностное сообщество макроводоросли-хозяина. Микроскопические изображения, показывающие бактериальное сообщество на дистальных концах (а), срединном слоевища (б) и основании (в) срезов зеленой водоросли Ulva australis .Бактерии выявляли с помощью конфокальной микроскопии с использованием CARD-FISH. Все бактерии (зеленый), Alphaproteobacteria (красный). Масштабные полосы представляют собой длину 10 мкм. Эти изображения были сделаны в Центре морских биоинноваций Университета Нового Южного Уэльса, Австралия, доктором Нииной Туджулой.
Специфичность к хозяину относится к появлению определенного набора бактериальных эпифитов на одном типе водорослей, которые отсутствуют (или встречаются в очень небольшом количестве) на других видах водорослей. В поддержку специфичности хозяина дактилоскопия бактериального сообщества (электрофорез в денатурирующем градиентном геле — DGGE) различных макроводорослей в разных местах показала, что модели сообщества больше похожи на таковые у конспецифических макроводорослей из другого географического происхождения, чем на другие виды макроводорослей или морскую воду из того же места. среды (Lachnit и др. ., 2009). Аналогичные закономерности наблюдались для активных сообществ, связанных с красной водорослью Laurencia dendroidea , где транскриптомное профилирование обнаружило небольшие различия в таксономическом составе сообщества на разных участках выборки (de Oliveira et al ., 2012). Такая специфичность хозяина может также относиться к бактериям, живущим в клетках водорослей. Несмотря на то, что они были описаны более 40 лет назад (Colombo, 1978), эндофитные сообщества сифонных зеленых водорослей, таких как Caulerpa sp.и Bryopsis sp., только недавно было показано, что они стабильны во времени (Meusnier et al ., 2001; Hollants et al ., 2011b) и действительно отличаются от эпифитного сообщества той же водоросли (Hollants и др. ., 2011а).
Несмотря на то, что они были описаны более 40 лет назад (Colombo, 1978), эндофитные сообщества сифонных зеленых водорослей, таких как Caulerpa sp.и Bryopsis sp., только недавно было показано, что они стабильны во времени (Meusnier et al ., 2001; Hollants et al ., 2011b) и действительно отличаются от эпифитного сообщества той же водоросли (Hollants и др. ., 2011а).
В отличие от этой специфичности некоторых растений-хозяев существует возможность существования универсальных эпифитов, общих для всех или многих макроводорослей, или, альтернативно, что некоторые макроводоросли могут не содержать сильно специфичных для хозяина сообществ (Burke et al ., 2011а,б). Действительно, общие таксоны были идентифицированы на поверхности макроводорослей, хотя в основном на уровне типа. Например, бактерии, принадлежащие к типам Planctomycetes и Verrucomicrobia , широко распространены на норвежских водорослях ( Laminaria hyperborea ) (Bengtsson & Øvreås, 2010; Bengtsson et al . , 2010) и на Балтийское море (Lachnit et al ., 2011). Однако эти типы заметно отсутствовали у родственных видов водорослей [ Saccharina latissima (ранее Laminaria saccharina )] из Балтийского и Северного морей (Staufenberger et al ., 2008), либо в результате специфичности хозяина для этих типов, либо в результате биогеографии. Что касается других макроводорослей, общие эпифитные бактерии включают членов Alphaproteobacteria , Gammaproteobacteria , Bacteroidetes и Cyanobacteria с небольшими различиями в этих более высоких таксономических рангах между сообществами, связанными с различными группами водорослей (т. е. красными, зелеными и бурые водоросли) (см. также Таблицу S1 и Holllants et al ., 2012). Интересно, что во многих случаях сходство на более высоких таксономических рангах (т.е. тип или класс) не наблюдаются на более низких рангах (т. е. родах или видах). Возможно, что ограничения имеющихся в настоящее время наборов данных (как обсуждается ниже) препятствуют идентификации родов, видов или даже штаммов, которые являются общими для всех поверхностей макроводорослей, и, следовательно, может быть слишком рано предлагать конкретные группы бактерий, которые могут быть обнаружены.
, 2010) и на Балтийское море (Lachnit et al ., 2011). Однако эти типы заметно отсутствовали у родственных видов водорослей [ Saccharina latissima (ранее Laminaria saccharina )] из Балтийского и Северного морей (Staufenberger et al ., 2008), либо в результате специфичности хозяина для этих типов, либо в результате биогеографии. Что касается других макроводорослей, общие эпифитные бактерии включают членов Alphaproteobacteria , Gammaproteobacteria , Bacteroidetes и Cyanobacteria с небольшими различиями в этих более высоких таксономических рангах между сообществами, связанными с различными группами водорослей (т. е. красными, зелеными и бурые водоросли) (см. также Таблицу S1 и Holllants et al ., 2012). Интересно, что во многих случаях сходство на более высоких таксономических рангах (т.е. тип или класс) не наблюдаются на более низких рангах (т. е. родах или видах). Возможно, что ограничения имеющихся в настоящее время наборов данных (как обсуждается ниже) препятствуют идентификации родов, видов или даже штаммов, которые являются общими для всех поверхностей макроводорослей, и, следовательно, может быть слишком рано предлагать конкретные группы бактерий, которые могут быть обнаружены. считаются «типичными» или «основными» и потенциально уникальными эпифитами из макроводорослей.
считаются «типичными» или «основными» и потенциально уникальными эпифитами из макроводорослей.
Сообщества макроводорослей также испытывают пространственные и временные сдвиги, которые могут быть отражением меняющихся местных условий, физиологии хозяина или химических и физических параметров.Например, Lachnit и др. . (2011) обнаружили воспроизводимые сезонные сдвиги в бактериальных сообществах трех различных одновременно встречающихся водорослей-хозяев, при этом определенный состав зимнего и летнего бактериального сообщества повторяется в течение последовательных лет. На наблюдаемые различия и сходства также могут влиять методологические ограничения. Эти ограничения иллюстрируются исследованиями космополитической зеленой водоросли Ulva lactuca (также называемой U. australis ), где анализ на основе DGGE предположил существование основного сообщества, стабильного в пространстве и времени (Longford et номер ., 2007; Tujula и др. . , 2010). Напротив, обширное секвенирование гена 16S рРНК бактериального сообщества U. australis не смогло обнаружить основное сообщество, поскольку только шесть видов бактерий из общего числа 528 были общими для шести отдельных водорослей (Burke et al ., 2011a). ). Эти кажущиеся противоречивыми результаты, вероятно, являются отражением методов с более высоким разрешением, используемых Burke et al . (2011a), тем не менее, результаты этих более продвинутых методов отличаются от более конкретных сообществ, описанных выше.
, 2010). Напротив, обширное секвенирование гена 16S рРНК бактериального сообщества U. australis не смогло обнаружить основное сообщество, поскольку только шесть видов бактерий из общего числа 528 были общими для шести отдельных водорослей (Burke et al ., 2011a). ). Эти кажущиеся противоречивыми результаты, вероятно, являются отражением методов с более высоким разрешением, используемых Burke et al . (2011a), тем не менее, результаты этих более продвинутых методов отличаются от более конкретных сообществ, описанных выше.
Различия в специфичности микробных сообществ на разных водорослях-хозяевах могут быть устранены путем рассмотрения микробного функционирования, а не филогении, как недавно было продемонстрировано для бактериального сообщества U. australis (Burke et al ., 2011b). С помощью метагеномного секвенирования эпифитного сообщества водорослей можно было определить набор основных функций, которые постоянно присутствовали у особей U.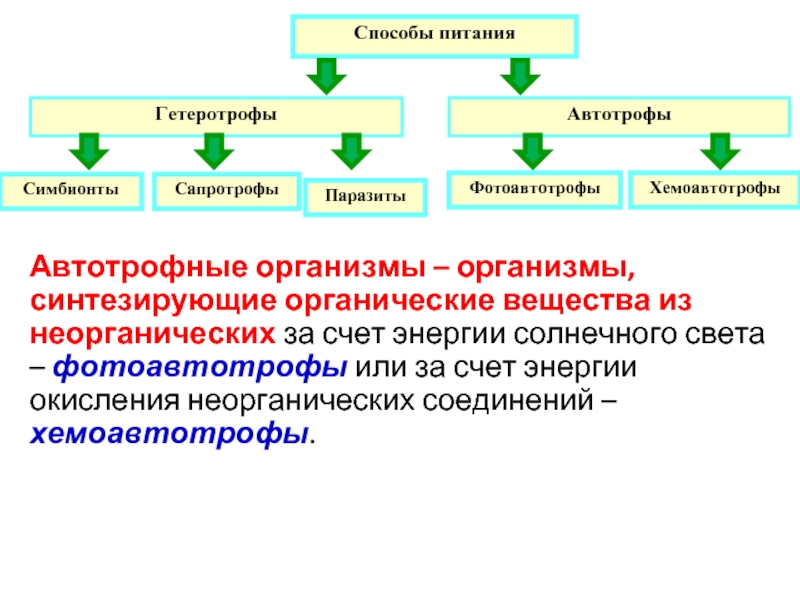 australis , несмотря на отсутствие общности в таксономическом составе на более низких уровнях (т.е. ниже семьи). Эти основные функции соответствовали концептуальному пониманию экологии бактериального сообщества, связанного с водорослями или поверхностью. Например, функции, связанные с обнаружением и перемещением к поверхности хозяина, а также прикреплением и образованием биопленки, были более многочисленными в сообществе U. australis , чем в планктонных сообществах. Другие чрезмерно представленные функции связаны с реакцией на среду водорослей-хозяев, защитой и латеральным переносом генов.Последняя функция представляет собой один из возможных механизмов, создающих функциональное сходство у филогенетически различных бактерий на поверхности U. australis (Burke et al ., 2011b).
australis , несмотря на отсутствие общности в таксономическом составе на более низких уровнях (т.е. ниже семьи). Эти основные функции соответствовали концептуальному пониманию экологии бактериального сообщества, связанного с водорослями или поверхностью. Например, функции, связанные с обнаружением и перемещением к поверхности хозяина, а также прикреплением и образованием биопленки, были более многочисленными в сообществе U. australis , чем в планктонных сообществах. Другие чрезмерно представленные функции связаны с реакцией на среду водорослей-хозяев, защитой и латеральным переносом генов.Последняя функция представляет собой один из возможных механизмов, создающих функциональное сходство у филогенетически различных бактерий на поверхности U. australis (Burke et al ., 2011b).
Данные метагенома U. australis предполагают, что состав сообщества в значительной степени определяется функцией, а не таксономической идентичностью. Поверхности макроводорослей часто недавно заселяются бактериями из планктона, которые, вероятно, содержат много видов с эквивалентной функциональностью, которая позволила бы им стать частью сообщества, связанного с поверхностью. Если первоначальная колонизация происходит случайно («лотерея») набором функционально эквивалентных планктонных бактерий («гильдия»), то окончательный состав сообщества не будет иметь узнаваемой таксономической модели, но будет постоянно содержать все признаки, необходимые для эпифитного вида. сообщества для функционирования (Burke et al ., 2011b). Такой сценарий может быть не только ограничен поверхностями макроводорослей, но также применим к морским беспозвоночным или даже к ряду других микробиомов, например, из желудочно-кишечного тракта человека, где ассоциаты бактериального сообщества рекрутируются из окружающей среды (т.е. горизонтальный захват). В этой модели филогенетическая специфичность (или ее отсутствие) определяется степенью, в которой филогенез сопоставляется с функцией, которая в случае U. australis была плохой.
Если первоначальная колонизация происходит случайно («лотерея») набором функционально эквивалентных планктонных бактерий («гильдия»), то окончательный состав сообщества не будет иметь узнаваемой таксономической модели, но будет постоянно содержать все признаки, необходимые для эпифитного вида. сообщества для функционирования (Burke et al ., 2011b). Такой сценарий может быть не только ограничен поверхностями макроводорослей, но также применим к морским беспозвоночным или даже к ряду других микробиомов, например, из желудочно-кишечного тракта человека, где ассоциаты бактериального сообщества рекрутируются из окружающей среды (т.е. горизонтальный захват). В этой модели филогенетическая специфичность (или ее отсутствие) определяется степенью, в которой филогенез сопоставляется с функцией, которая в случае U. australis была плохой.
Будущие исследования таксономического и филогенетического состава сообщества с использованием методов высокого разрешения необходимы, чтобы пролить свет на возможность существования основного сообщества бактерий, связанных с морскими водорослями. Здесь мы утверждаем, что функциональные исследования (например, метагеномика, транскриптомика, протеомика, метаболомика и т.) следует проводить параллельно со стандартными филогенетическими анализами, если это вообще возможно. Однако, несмотря на разнообразие макроводорослей-хозяев и изменчивость среды, в которой они живут, вполне вероятно, что взаимодействия между макроводорослями и бактериями будут столь же разнообразными и варьируются от специализированных до универсальных. Поэтому важно получить представление о биологических, физических и химических факторах, влияющих на эпифитное сообщество отдельных видов макроводорослей.
Здесь мы утверждаем, что функциональные исследования (например, метагеномика, транскриптомика, протеомика, метаболомика и т.) следует проводить параллельно со стандартными филогенетическими анализами, если это вообще возможно. Однако, несмотря на разнообразие макроводорослей-хозяев и изменчивость среды, в которой они живут, вполне вероятно, что взаимодействия между макроводорослями и бактериями будут столь же разнообразными и варьируются от специализированных до универсальных. Поэтому важно получить представление о биологических, физических и химических факторах, влияющих на эпифитное сообщество отдельных видов макроводорослей.
 Параметры, определяющие поверхностную среду макроводорослей, включают метаболиты водорослей, существующее постоянное микробное сообщество с его пулом вторичных метаболитов микробного происхождения и физико-химические условия на поверхности слоевища, такие как кислород и углекислый газ, которые могут дополнительно модулировать рН поверхности (рис.2). Многие из этих параметров подвержены ежедневным (Спиллинг и др. , 2010), Фишер и др. , неопубликованные) или сезонным модуляциям (Хеллио и др. , 2004). Таким образом, бактерии, вступающие в устойчивую ассоциацию с макроводорослями-хозяевами, должны обладать адаптивными чертами, отражающими эти нишевые условия.
Параметры, определяющие поверхностную среду макроводорослей, включают метаболиты водорослей, существующее постоянное микробное сообщество с его пулом вторичных метаболитов микробного происхождения и физико-химические условия на поверхности слоевища, такие как кислород и углекислый газ, которые могут дополнительно модулировать рН поверхности (рис.2). Многие из этих параметров подвержены ежедневным (Спиллинг и др. , 2010), Фишер и др. , неопубликованные) или сезонным модуляциям (Хеллио и др. , 2004). Таким образом, бактерии, вступающие в устойчивую ассоциацию с макроводорослями-хозяевами, должны обладать адаптивными чертами, отражающими эти нишевые условия.
Рисунок 2
Холобионт морских водорослей и факторы, которые, по прогнозам, влияют на бактериальную колонизацию макроводорослей-хозяев. Рисунок 2
 Таким образом, фотосинтез хозяина позволил бы аэробным процессам происходить в ситуациях, когда в противном случае кислород мог бы быть ограничен. Триас и др. . (2012) специально проверили эту идею, выдвинув гипотезу о том, что поверхность глубоководных макроводорослей может представлять собой избирательную среду обитания для требующего кислорода процесса окисления аммония. С помощью количественной ПЦР было обнаружено, что количество окисляющих аммоний бактерий было относительно высоким (1% от общего количества бактерий) на поверхности водорослей по сравнению с тем, что ранее было продемонстрировано для других морских местообитаний [e.г. 0,1% для морских губок (Bayer et al ., 2008)]. Однако кислород также может нанести вред бактериальным эпифитам, особенно если он приводит к образованию вредных активных форм кислорода (АФК). На самом деле, макроводоросли могут быстро высвобождать большое количество АФК, таких как ионы супероксида и перекиси водорода (так называемые «окислительные взрывы»), чтобы защитить себя от бактериальной атаки [рассмотрено в (Weinberger, 2007)].
Таким образом, фотосинтез хозяина позволил бы аэробным процессам происходить в ситуациях, когда в противном случае кислород мог бы быть ограничен. Триас и др. . (2012) специально проверили эту идею, выдвинув гипотезу о том, что поверхность глубоководных макроводорослей может представлять собой избирательную среду обитания для требующего кислорода процесса окисления аммония. С помощью количественной ПЦР было обнаружено, что количество окисляющих аммоний бактерий было относительно высоким (1% от общего количества бактерий) на поверхности водорослей по сравнению с тем, что ранее было продемонстрировано для других морских местообитаний [e.г. 0,1% для морских губок (Bayer et al ., 2008)]. Однако кислород также может нанести вред бактериальным эпифитам, особенно если он приводит к образованию вредных активных форм кислорода (АФК). На самом деле, макроводоросли могут быстро высвобождать большое количество АФК, таких как ионы супероксида и перекиси водорода (так называемые «окислительные взрывы»), чтобы защитить себя от бактериальной атаки [рассмотрено в (Weinberger, 2007)]. В свою очередь, чтобы защитить себя, резидентные бактерии могут экспрессировать пероксидазу, каталазу и другие оксидазы, которые разрушают АФК и, следовательно, минимизируют ущерб.Хотя важность этих защитных механизмов еще не установлена напрямую, следует отметить, что геномы нескольких бактерий, связанных с макроводорослями, микробный метагеном U. australis, и транскриптом микробного сообщества, связанного с L. dendroidea, содержат множество генов, связанных с реакцией на окислительный стресс (Thomas et al ., 2008; Burke et al ., 2011b; Fernandes et al ., 2011; de Oliveira et al ., 2012).
В свою очередь, чтобы защитить себя, резидентные бактерии могут экспрессировать пероксидазу, каталазу и другие оксидазы, которые разрушают АФК и, следовательно, минимизируют ущерб.Хотя важность этих защитных механизмов еще не установлена напрямую, следует отметить, что геномы нескольких бактерий, связанных с макроводорослями, микробный метагеном U. australis, и транскриптом микробного сообщества, связанного с L. dendroidea, содержат множество генов, связанных с реакцией на окислительный стресс (Thomas et al ., 2008; Burke et al ., 2011b; Fernandes et al ., 2011; de Oliveira et al ., 2012). Компоненты клеточной стенки макроводорослей могут представлять собой источник питательных веществ для бактерий, способных утилизировать эти биополимеры. В поддержку этого есть несколько исследований, демонстрирующих способность конкретных морских бактерий разлагать различные полимеры макроводорослей.Обзор специфической ферментативной активности, обнаруженной у соответствующих морских бактерий, которые разрушают клеточные стенки макроводорослей, дан в Goecke et al . (2010).
Компоненты клеточной стенки макроводорослей могут представлять собой источник питательных веществ для бактерий, способных утилизировать эти биополимеры. В поддержку этого есть несколько исследований, демонстрирующих способность конкретных морских бактерий разлагать различные полимеры макроводорослей.Обзор специфической ферментативной активности, обнаруженной у соответствующих морских бактерий, которые разрушают клеточные стенки макроводорослей, дан в Goecke et al . (2010).
Компоненты клеточной стенки макроводорослей могут представлять собой источник питательных веществ для бактерий, способных утилизировать эти биополимеры. В поддержку этого есть несколько исследований, демонстрирующих способность конкретных морских бактерий разлагать различные полимеры макроводорослей.Обзор специфической ферментативной активности, обнаруженной у соответствующих морских бактерий, которые разрушают клеточные стенки макроводорослей, дан в Goecke et al . (2010). Деградация полимера (например, клеточных стенок или материалов для хранения), очевидно, может оказать пагубное воздействие на хозяина, если ее не контролировать. Таким образом, устойчивые или долговременные бактериальные ассоциаты макроводорослей могут не обладать способностью к начальной деградации полимера (или должны жестко контролировать ее). Это подтверждается наблюдением, что у обычного бактериального эпифита макроводорослей Pseudoalteromonas tunicata отсутствуют ферменты, необходимые для гидролиза полимеров клеточной стенки макроводорослей, но все же он содержит структуры, участвующие в связывании полимера (например,г. частичная целлюлосома) (Thomas et al ., 2008). Таким образом, бактерии с признаками разрушения полимеров могут представлять условно-патогенные микроорганизмы или сапрофиты, а не комменсальные или мутуалистические симбионты макроводорослей. Однако, как только происходит повреждение хозяина, безвредные ассоциаты могут способствовать деградации хозяина или в полной мере использовать высвободившиеся питательные вещества. Например, P. tunicata сохраняет способность использовать мономеры, полученные в результате разложения типичных полимеров макроводорослей, таких как целлюлоза и ксилан (Thomas et al ., 2008), и это пойдет на пользу организму, когда его хозяин будет скомпрометирован. Подобный сдвиг в поведении недавно наблюдался и у бактериального симбионта микроводоросли Emiliania huxleyi . Здесь симбионт Phaeobacter gallaeciensis вырабатывал сильнодействующий альгицид в ответ на продукт распада водорослей (п-кумаровую кислоту), тем самым способствуя дальнейшему разрушению своего стареющего хозяина (Seyedsayamdost et al .
частичная целлюлосома) (Thomas et al ., 2008). Таким образом, бактерии с признаками разрушения полимеров могут представлять условно-патогенные микроорганизмы или сапрофиты, а не комменсальные или мутуалистические симбионты макроводорослей. Однако, как только происходит повреждение хозяина, безвредные ассоциаты могут способствовать деградации хозяина или в полной мере использовать высвободившиеся питательные вещества. Например, P. tunicata сохраняет способность использовать мономеры, полученные в результате разложения типичных полимеров макроводорослей, таких как целлюлоза и ксилан (Thomas et al ., 2008), и это пойдет на пользу организму, когда его хозяин будет скомпрометирован. Подобный сдвиг в поведении недавно наблюдался и у бактериального симбионта микроводоросли Emiliania huxleyi . Здесь симбионт Phaeobacter gallaeciensis вырабатывал сильнодействующий альгицид в ответ на продукт распада водорослей (п-кумаровую кислоту), тем самым способствуя дальнейшему разрушению своего стареющего хозяина (Seyedsayamdost et al . , 2011).
, 2011).
Недавние геномные и метагеномные данные ассоциатов макроводорослей выявили дополнительные доказательства поглощения питательных веществ, таких как использование фосфора, азота и железа (Thomas et al ., 2008; Берк и др. ., 2011b; Fernandes и др. ., 2011). Члены клады Roseobacter обычно выделяют из макроводорослей (Brinkhoff et al ., 2008), и их геномы кодируют функции, позволяющие использовать осмолиты водорослей, такие как путресцин, таурин (Kalhoefer et al ., 2011). ), креатин, саркозин (Thole et al ., 2012) и диметилсульфониопропионат (DMSP). Помимо того, что они распространены в фитопланктоне (Curson и др. ., 2011), ДМСП часто продуцируется макроводорослями (например, Polysiphonia и Fucus ) (Malin & Erst, 1997; Saha et al ., 2012), и связанный с водорослями метаболизм этого соединения может играть еще неизученную роль в глобальный круговорот серы.
В качестве последнего комментария по этой теме, хотя бактериальная деградация ткани макроводорослей вредна для хозяина, этот процесс имеет решающее значение для глобального круговорота углерода и питательных веществ. Более того, при правильном управлении такая деградация потенциально может быть использована для облегчения эффективного разложения, необходимого для преобразования полимеров макроводорослей в биотопливо (Wargacki et al ., 2012), область, представляющая растущий коммерческий интерес.
Более того, при правильном управлении такая деградация потенциально может быть использована для облегчения эффективного разложения, необходимого для преобразования полимеров макроводорослей в биотопливо (Wargacki et al ., 2012), область, представляющая растущий коммерческий интерес.
 , 2010; Снид и Понерт, 2011). Экспериментальная система, предназначенная для моделирования выброса метаболитов макроводорослей с искусственной поверхности, использовалась для измерения воздействия метаболитов макроводорослей на бактериальную колонизацию при экологически реалистичных концентрациях. Основываясь на анализе отпечатков пальцев сообщества, состав тестовых образцов отличался от контрольных образцов, но был аналогичен составу естественной поверхности макроводорослей (Lachnit et al ., 2010), что свидетельствует о том, что метаболиты водорослей сами по себе являются сильным фактором отбора для состава сообщества. .Воздействие метаболитов макроводорослей также может выходить за пределы поверхности хозяина: несколько исследований демонстрируют, как макроводоросли могут влиять на структуру бактериального сообщества планктона (Lam & Harder, 2007; Lam et al ., 2008; Sneed & Pohnert, 2011). .
, 2010; Снид и Понерт, 2011). Экспериментальная система, предназначенная для моделирования выброса метаболитов макроводорослей с искусственной поверхности, использовалась для измерения воздействия метаболитов макроводорослей на бактериальную колонизацию при экологически реалистичных концентрациях. Основываясь на анализе отпечатков пальцев сообщества, состав тестовых образцов отличался от контрольных образцов, но был аналогичен составу естественной поверхности макроводорослей (Lachnit et al ., 2010), что свидетельствует о том, что метаболиты водорослей сами по себе являются сильным фактором отбора для состава сообщества. .Воздействие метаболитов макроводорослей также может выходить за пределы поверхности хозяина: несколько исследований демонстрируют, как макроводоросли могут влиять на структуру бактериального сообщества планктона (Lam & Harder, 2007; Lam et al ., 2008; Sneed & Pohnert, 2011). .
В дополнение к неочищенным экстрактам макроводорослей также были идентифицированы специфические метаболиты, которые, как было показано, влияют на состав и/или рост бактериального сообщества на макроводорослях-хозяевах (таблица 1). Зная о локализации, скорости доставки и влиянии специфических метаболитов макроводорослей на колонизирующие бактерии, было начато несколько исследований, посвященных их экологической значимости (Dworjanyn et al ., 1999; Nylund и др. ., 2010; Перссон и др. ., 2011; Саха и др. ., 2011). Одним из примеров является красная водоросль Delisea pulchra, , которая вырабатывает ряд галогенированных фуранонов, препятствующих поверхностному обрастанию микро- и макроорганизмами и поддерживающих здоровье и репродуктивную функцию этой макроводоросли (Campbell et al ., 2011). Фураноны локализуются в центральных везикулах клеток железы и постоянно высвобождаются на поверхность, где достигают поверхностных концентраций приблизительно 100 нг см -2 (Dworjanyn et al ., 1999). Концентрация фуранона уменьшается по мере удаления от дистальных концов макроводоросли; тем не менее, эти более низкие концентрации остаются достаточными для сдерживания экологически значимой эпибиоты и изменения состава бактериального сообщества (Maximilien et al .
Зная о локализации, скорости доставки и влиянии специфических метаболитов макроводорослей на колонизирующие бактерии, было начато несколько исследований, посвященных их экологической значимости (Dworjanyn et al ., 1999; Nylund и др. ., 2010; Перссон и др. ., 2011; Саха и др. ., 2011). Одним из примеров является красная водоросль Delisea pulchra, , которая вырабатывает ряд галогенированных фуранонов, препятствующих поверхностному обрастанию микро- и макроорганизмами и поддерживающих здоровье и репродуктивную функцию этой макроводоросли (Campbell et al ., 2011). Фураноны локализуются в центральных везикулах клеток железы и постоянно высвобождаются на поверхность, где достигают поверхностных концентраций приблизительно 100 нг см -2 (Dworjanyn et al ., 1999). Концентрация фуранона уменьшается по мере удаления от дистальных концов макроводоросли; тем не менее, эти более низкие концентрации остаются достаточными для сдерживания экологически значимой эпибиоты и изменения состава бактериального сообщества (Maximilien et al . , 1998; Campbell et al ., 2011).
, 1998; Campbell et al ., 2011).
Макроалгал метаболиты, влияющие на бактериальную и грибковую колонизацию при экологически чистых условиях
| MacroaLGA | Algal Metabolite | Ссылки | | Delisea Pulchra | Галогенированный фураноны | Maximilien et al .(1998) и Dworjanyn и др. . (1999) | Lobophora variegata | Циклический лактон – лобофоролид | Кубанек и др. . (2003) | Asparagopsis armata | Бромоформ | Paul и др. . (2006) | Asparagopsis armata | Дибромуксусная кислота | Paul et al . (2006) | Bonnemaisonia hamifera | Полигалогенированный 2-гептанон | Nylund et al .(2008) | Callophycus serratus | Бромфиколлиды | Lane et al . | 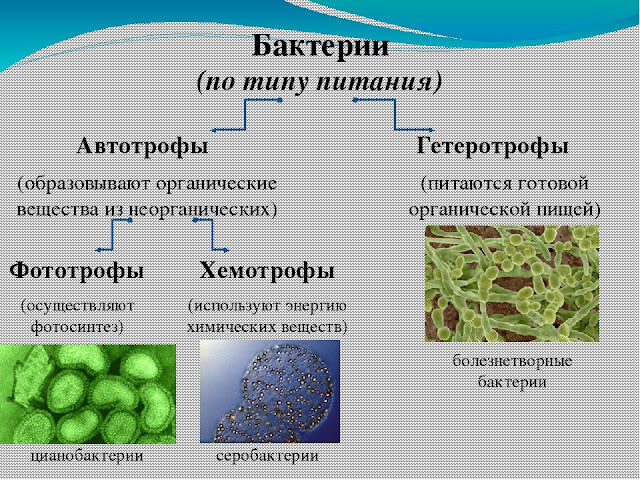 (2009) (2009) Fucus vesiculosus | Фукоксантин | Саха и др. . (2011) | |
 (2009)
(2009)макроводорослей метаболитов, которые влияют на бактериальную и грибковую колонизацию при экологически соответствующих условиях
| макроводоросль | Algal метаболитом | Ссылок |
| Delisea Pulchra | галогенированных фуранонами | Максимилиан и др. .(1998) и Dworjanyn и др. . (1999) |
| Lobophora variegata | Циклический лактон – лобофоролид | Кубанек и др. . (2003) |
| Asparagopsis armata | Бромоформ | Paul и др. . (2006) |
| Asparagopsis armata | Дибромуксусная кислота | Paul et al . (2006) |
| Bonnemaisonia hamifera | Полигалогенированный 2-гептанон | Nylund et al . (2008) (2008) |
| Callophycus serratus | Бромфиколлиды | Lane et al . (2009) |
| Fucus vesiculosus | Фукоксантин | Саха и др. . (2011) |
 (2008)
(2008) Другим примером является красная водоросль Bonnemaisonia hamifera , основной биоактивный метаболит которой – 1,1,3,3-тетрабром-2-гептанон – хранится в локализованных на поверхности клетках железы и может достигать концентрации до 4 мкг см -2 (Nylund и др. ., 2008). Этот метаболит, нанесенный в соответствующей концентрации на полевые панели, изменяет плотность, разнообразие и состав бактериального сообщества (Persson et al ., 2011). Кроме того, бурая водоросль Fucus vesiculosus продуцирует пигмент фукоксантин, который в экологически реалистичных концентрациях (0,7–0,9 мкг см -2 ) предотвращает прикрепление бактериальных изолятов из сопутствующих макроводорослей (Saha et al . , 2011). ). Напротив, бактерии, выделенные из самой водоросли, остаются относительно нечувствительными к действию фукоксантина (Saha et al ., 2011). Последующее исследование показало, что, хотя наблюдалось значительное сокращение числа бактериальных колонизаторов, фукоксантин мало влиял на общий состав бактериального сообщества (Lachnit et al., , неопубликовано). Следовательно, в отличие от фуранонов из D. pulchra или полигалогенированных 2-гептанонов из B. hamifera, фукоксантин кажется менее селективным, действуя как общий ингибитор прикрепления бактерий, а не как специфический ингибитор роста бактерий, влияющий на состав сообщества. .Тем не менее, переменная чувствительность отдельных бактерий к специфическим метаболитам макроводорослей, вероятно, является общей темой, влияющей на состав сообществ эпифитных бактерий. Например, Саха и др. . (2012) недавно показали, что распространенные метаболиты макроводорослей, такие как ДМСП и аминокислоты пролин и аланин, ингибируют прикрепление к поверхности определенных бактерий (например, Cytophaga sp), в то же время способствуя прикреплению других (например, Rheinheimera baltica ).
, 2011). ). Напротив, бактерии, выделенные из самой водоросли, остаются относительно нечувствительными к действию фукоксантина (Saha et al ., 2011). Последующее исследование показало, что, хотя наблюдалось значительное сокращение числа бактериальных колонизаторов, фукоксантин мало влиял на общий состав бактериального сообщества (Lachnit et al., , неопубликовано). Следовательно, в отличие от фуранонов из D. pulchra или полигалогенированных 2-гептанонов из B. hamifera, фукоксантин кажется менее селективным, действуя как общий ингибитор прикрепления бактерий, а не как специфический ингибитор роста бактерий, влияющий на состав сообщества. .Тем не менее, переменная чувствительность отдельных бактерий к специфическим метаболитам макроводорослей, вероятно, является общей темой, влияющей на состав сообществ эпифитных бактерий. Например, Саха и др. . (2012) недавно показали, что распространенные метаболиты макроводорослей, такие как ДМСП и аминокислоты пролин и аланин, ингибируют прикрепление к поверхности определенных бактерий (например, Cytophaga sp), в то же время способствуя прикреплению других (например, Rheinheimera baltica ).
Тот факт, что вторичные метаболиты макроводорослей часто продуцируются и высвобождаются специфическими клетками, вероятно, приводит к сильному локальному воздействию на бактериальные эпифиты. Недавние достижения в методах аналитической химии теперь позволяют проводить мелкомасштабную прямую оценку метаболитов на нативных поверхностях в условиях окружающей среды. Лейн и др. . (2009) впервые применили визуализирующий метод масс-спектрометрии (масс-спектрометрия с ионизацией десорбционным электрораспылением) на нативной поверхности красной водоросли Callophycus serratus для визуализации и измерения группы противогрибковых метаболитов водорослей – бромфиколидов.Этот подход выявил пятнистое распределение противогрибковых метаболитов по поверхности макроводорослей, предполагая, что поверхности макроводорослей неоднородны по отношению к биоактивным метаболитам. Корреляция этих мелкомасштабных градиентов состава метаболитов с анализом состава бактериального сообщества с высоким пространственным разрешением (например, флуоресцентная гибридизация in situ ) является мощным инструментом для оценки прямого влияния химии поверхности макроводорослей на микробное разнообразие, связанное с хозяином. На самом деле, вполне вероятно, что крутые локальные градиенты метаболитов макроводорослей создадут множество специфических микрониш (аналогично почвенной среде), что потенциально повлияет на общее микробное разнообразие и состав хозяина макроводорослей.
На самом деле, вполне вероятно, что крутые локальные градиенты метаболитов макроводорослей создадут множество специфических микрониш (аналогично почвенной среде), что потенциально повлияет на общее микробное разнообразие и состав хозяина макроводорослей.
Механизмы крепления и поверхностные соревнования
Бактерии используют ряд придатков для прикрепления к поверхности, что может опосредовать специфичность хозяина (Klemm & Schembri, 2000). Например, лектины представляют собой белки, связывающие сахар, которые могут способствовать прикреплению бактерий ко многим биологическим поверхностям (Rudiger & Gabius, 2001; Ogawa et al )., 2011). Однако роль лектин-опосредованного связывания с поверхностью макроводорослей остается неизученной. На самом деле очень мало исследований, в которых экспериментально проверялась роль специфических адгезинов в прикреплении к поверхности макроводорослей. Маннозочувствительные гемагглютининовые (MSHA)-пили участвуют в прикреплении P. tunicata к U. lactuca . Тем не менее, этот организм может дополнять MSHA-опосредованное связывание несколькими другими механизмами адгезии, включая курли-белковые волокна, которые, как известно, важны для колонизации растений в Е.coli (Thomas et al ., 2008), липопротеин (LipL32), участвующий в адгезии к волокнам общего внеклеточного матрикса (ECM) (Hoke et al ., 2008), и несколько дивергентных белков пилей (Thomas et al. ., 2008). Важность прикрепления к поверхности также отражена в геномах других бактериальных эпифитов, включая Nautella sp . R11 и P. gallaeciensis, , каждый из которых кодирует ряд известных и гипотетических адгезинов и внеклеточных полимеров, участвующих в формировании биопленки (Fernandes et al ., 2011; Thole и др. ., 2012). Кроме того, транскрипты, соответствующие генам, участвующим в производстве бактериальных внеклеточных полисахаридов, были чрезмерно представлены в микробиоме красной водоросли L.
tunicata к U. lactuca . Тем не менее, этот организм может дополнять MSHA-опосредованное связывание несколькими другими механизмами адгезии, включая курли-белковые волокна, которые, как известно, важны для колонизации растений в Е.coli (Thomas et al ., 2008), липопротеин (LipL32), участвующий в адгезии к волокнам общего внеклеточного матрикса (ECM) (Hoke et al ., 2008), и несколько дивергентных белков пилей (Thomas et al. ., 2008). Важность прикрепления к поверхности также отражена в геномах других бактериальных эпифитов, включая Nautella sp . R11 и P. gallaeciensis, , каждый из которых кодирует ряд известных и гипотетических адгезинов и внеклеточных полимеров, участвующих в формировании биопленки (Fernandes et al ., 2011; Thole и др. ., 2012). Кроме того, транскрипты, соответствующие генам, участвующим в производстве бактериальных внеклеточных полисахаридов, были чрезмерно представлены в микробиоме красной водоросли L. dendroidea (de Oliveira et al ., 2012).
dendroidea (de Oliveira et al ., 2012).
Прикрепившись, бактерии должны конкурировать с другими микробными эпифитами за питательные вещества и пространство в поверхностной биопленке макроводорослей. В такой ситуации производство антагонистических химических метаболитов (например, антибиотиков) было бы выгодным.Экологическая важность этого подтверждается частым выделением бактериальных штаммов, продуцирующих биологически активные вещества, с поверхности макроводорослей [рассмотрено в (Egan et al. ., 2008) и обсуждается ниже]. В бактериальном сообществе, связанном с U. australis, нерибосомных пептидных синтетаз, которые часто продуцируют биологически активные вещества, и помпы множественного оттока лекарств, как правило, широко распространены, что еще раз подтверждает роль химически опосредованного антагонизма и противодействующих защитных процессов в таких средах (Burke и др. ., 2011б). Также наблюдалась повышенная экспрессия антимикробных метаболитов в биопленке, что может дополнительно улучшить способность этих бактерий конкурировать на поверхности хозяина (Matz et al . , 2008).
, 2008).
Экологическая важность химического антагонизма, вытекающая из приведенных выше наблюдений, также подтверждается экспериментальными исследованиями в лаборатории. Например, P. tunicata и P. gallaeciensis являются лучшими конкурентами других совместно встречающихся эпифитных бактерий для заселения на U.australis , однако мутантные штаммы, не продуцирующие антибиотики [AlpP и троподитиетиновую кислоту (TDA) соответственно], значительно менее конкурентоспособны (Rao et al ., 2005). Интересно, что, хотя это показывает преимущество продукции антагонистических метаболитов во время раннего формирования естественного эпифитного сообщества, значение для последующей бактериальной колонизации еще предстоит определить. На самом деле, предварительно созданные естественные эпифитные сообщества могут быть устойчивыми к интродукции новых членов, поскольку P.tunicata и P. gallaeciensis , как недавно было показано, плохо проникают в предварительно созданные биопленки как на искусственных поверхностях, так и на поверхностях макроводорослей (Rao et al . , 2010).
, 2010).
В целом, множество факторов хозяина, микробных ассоциатов и условий окружающей среды, вероятно, играют роль в формировании состава микробного сообщества морских макроводорослей. Улучшение понимания того, в какой степени эти различные факторы влияют на связанный с поверхностью микробиом in situ , будет иметь решающее значение для прогнозирования потенциального воздействия микробных симбионтов на их хозяина с точки зрения здоровья и функций, как обсуждается в следующем разделе.
Функциональные результаты взаимодействия водорослей и бактерий
Хотя макроводоросли представляют собой ниши с уникальными и избирательными свойствами, они также испытывают ряд полезных и вредных взаимодействий со своим бактериальным симбиотическим сообществом. Учитывая экологическое и прикладное значение макроводорослей, возрастает интерес к определению результатов этих взаимодействий.
Бактерии поставляют основные питательные вещества и необходимы для нормального морфологического развития морских макроводорослей
Эпифитные гетеротрофные бактерии не только обеспечивают CO 2 для фотоавтотрофии макроводорослей, но в некоторых случаях также обеспечивают фиксированный азот (Penhale & Capone, 1981; Phlips & Zeman, 1990). Действительно, недавно было обнаружено, что азотфиксирующие цианобактерии входят в число доминирующих активных членов микробного сообщества, связанного с L. dendroidea (de Oliveira et al ., 2012). Эпифитные бактерии также могут способствовать или дополнять первичную продукцию макроводорослей-хозяев, поскольку бентосные виды макроводорослей часто изобилуют автотрофными цианобактериями (Barott et al ., 2011).
Действительно, недавно было обнаружено, что азотфиксирующие цианобактерии входят в число доминирующих активных членов микробного сообщества, связанного с L. dendroidea (de Oliveira et al ., 2012). Эпифитные бактерии также могут способствовать или дополнять первичную продукцию макроводорослей-хозяев, поскольку бентосные виды макроводорослей часто изобилуют автотрофными цианобактериями (Barott et al ., 2011).
Кроме того, бактерии положительно влияют на морфологическое развитие некоторых видов макроводорослей.Возможно, наиболее изученным примером являются ранние наблюдения того, что некоторые зеленые макроводоросли не развивают нормальную морфологию в отсутствие местных бактериальных сообществ (Provasoli & Pintner, 1980). В частности, выращенный аксеном U. lactuca развил аномальную «подушкообразную» морфологию, которая могла быть восстановлена до типичного листовидного слоевища при повторной инокуляции бактериальными штаммами, выделенными из водоросли. Аналогичные эффекты были зарегистрированы для других видов зеленых водорослей, в том числе Ulva linza , Ulva Compressa (формально Enteromorpha linza и Enteromorpha Compressa ) (Fries, 1975), Ulva pertusa (Nakanishi et al 9), 1999), Ulva fasciata (Singh et al ., 2011) и Monostroma oxyspermum (Matsuo et al ., 2003). Хотя в каждом случае нормальная морфология может быть восстановлена «повторной инокуляцией» соответствующими бактериями, механизмы этого взаимодействия, по-видимому, различаются между макроводорослями-хозяевами. И Наканиши , и др. . (1996) и Marshall и др. . (2006) представили доказательства того, что бактерии из ряда типов, включая представителей Proteobacteria, Bacteroidetes и Firmicutes, способны индуцировать нормальный морфогенез у видов Ulva и что для восстановления нормальной морфологии макроводорослей может потребоваться прикрепление бактерий к хозяину.
Аналогичные эффекты были зарегистрированы для других видов зеленых водорослей, в том числе Ulva linza , Ulva Compressa (формально Enteromorpha linza и Enteromorpha Compressa ) (Fries, 1975), Ulva pertusa (Nakanishi et al 9), 1999), Ulva fasciata (Singh et al ., 2011) и Monostroma oxyspermum (Matsuo et al ., 2003). Хотя в каждом случае нормальная морфология может быть восстановлена «повторной инокуляцией» соответствующими бактериями, механизмы этого взаимодействия, по-видимому, различаются между макроводорослями-хозяевами. И Наканиши , и др. . (1996) и Marshall и др. . (2006) представили доказательства того, что бактерии из ряда типов, включая представителей Proteobacteria, Bacteroidetes и Firmicutes, способны индуцировать нормальный морфогенез у видов Ulva и что для восстановления нормальной морфологии макроводорослей может потребоваться прикрепление бактерий к хозяину. (Наканиши и др. ., 1999). Напротив, при скрининге более 50 изолятов Singh et al . (2011) обнаружили только пять штаммов, принадлежащих либо к Marinomonas, либо к sp. или Bacillus sp. которые смогли вызвать нормальное развитие аксеничных U. fasciata . Более того, исследования с M. oxyspermum также предполагают, что индукция морфогенеза ограничена определенными группами бактерий ( Cytophaga – Flavobacterium – Bacteroides ) (Matsuo et al ., 2003) и возникает в ответ на секретируемый фактор морфогенеза, называемый таллюзином (Matsuo et al ., 2005). Таллузин эффективен в низких концентрациях (диапазон фг мл 90 265 -1 90 266), но со временем активность теряется, что позволяет предположить, что макроводоросль может полагаться на постоянное поступление индуктора из эпифитной бактерии. Интересно, что как бактерия-продуцент, так и чистый таллюзин смогли восстановить нормальную морфологию других зеленых водорослей, что позволяет предположить, что это универсальный сигнал для морфогенеза зеленых водорослей (Matsuo et al .
(Наканиши и др. ., 1999). Напротив, при скрининге более 50 изолятов Singh et al . (2011) обнаружили только пять штаммов, принадлежащих либо к Marinomonas, либо к sp. или Bacillus sp. которые смогли вызвать нормальное развитие аксеничных U. fasciata . Более того, исследования с M. oxyspermum также предполагают, что индукция морфогенеза ограничена определенными группами бактерий ( Cytophaga – Flavobacterium – Bacteroides ) (Matsuo et al ., 2003) и возникает в ответ на секретируемый фактор морфогенеза, называемый таллюзином (Matsuo et al ., 2005). Таллузин эффективен в низких концентрациях (диапазон фг мл 90 265 -1 90 266), но со временем активность теряется, что позволяет предположить, что макроводоросль может полагаться на постоянное поступление индуктора из эпифитной бактерии. Интересно, что как бактерия-продуцент, так и чистый таллюзин смогли восстановить нормальную морфологию других зеленых водорослей, что позволяет предположить, что это универсальный сигнал для морфогенеза зеленых водорослей (Matsuo et al . , 2003, 2005).
, 2003, 2005).
Бактерии, связанные с макроводорослями, способствуют защите хозяина от нежелательной колонизации и биологического обрастания
Существуют многочисленные лабораторные исследования, демонстрирующие, что эпифитные бактерии обладают ингибирующей активностью в отношении обычных организмов, вызывающих биообрастание (см. обзор Holmström et al ., 2002; Dobretsov et al ., 2006; Egan et al ., 2008). Например, водные экстракты и биопленки макроводоросли Vibrio sp.и Pseudoalteromonas sp. подавляют расселение и метаморфоз полихет Hydroides elegans (Добрецов и Цянь, 2002). Также штаммов Pseudoalteromonas из U. lactuca как в умеренных (Egan et al ., 2001), так и в тропических водах (Kumar et al ., 2010) обладают активностью против различных обрастающих организмов (бактерий, диатомей, диатомей, .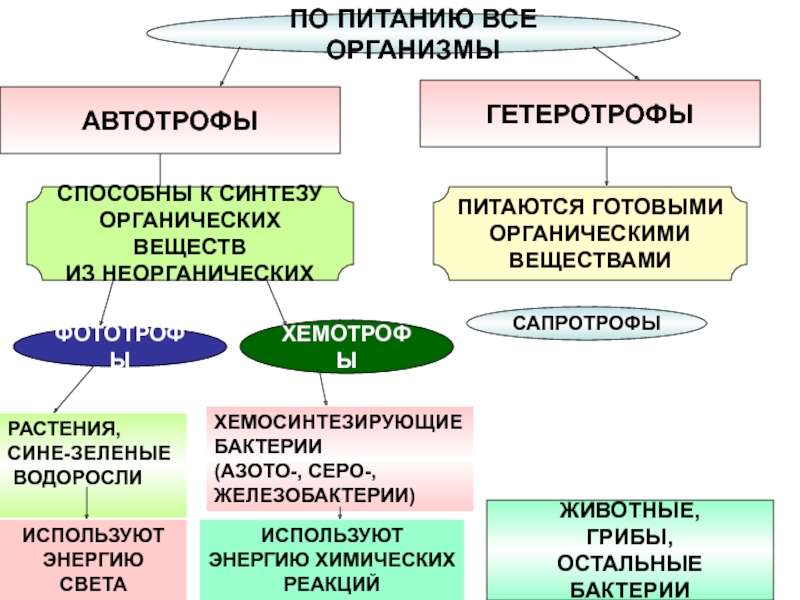 ). Фактически, видов Pseudoalteromonas обычно выделяют с поверхности водорослей и регулярно проявляют противообрастающие свойства.В частности, P. tunicata стал модельным организмом для борьбы с обрастанием, поскольку он обладает активностью против ряда целевых организмов, включая споры водорослей, личинок беспозвоночных, донных диатомей, различные бактерии, грибы (Bowman, 2007; Egan et al. . , 2008), протисты (Matz et al ., 2008) и нематоды (Ballestriero et al ., 2010). Примечательно, что в то время как P. tunicata и близкородственные виды P. ulvae, проявляются в относительно низкой плотности (10 3 клеток см -2 ) на макроводорослях-хозяевах ( U.lactuca и Ulvaria fusca) в полевых условиях (Skovhus et al ., 2007), эти плотности по-прежнему достаточны для подавления обрастания спорами макроводорослей, морскими грибами и личинками беспозвоночных (Rao et al .
). Фактически, видов Pseudoalteromonas обычно выделяют с поверхности водорослей и регулярно проявляют противообрастающие свойства.В частности, P. tunicata стал модельным организмом для борьбы с обрастанием, поскольку он обладает активностью против ряда целевых организмов, включая споры водорослей, личинок беспозвоночных, донных диатомей, различные бактерии, грибы (Bowman, 2007; Egan et al. . , 2008), протисты (Matz et al ., 2008) и нематоды (Ballestriero et al ., 2010). Примечательно, что в то время как P. tunicata и близкородственные виды P. ulvae, проявляются в относительно низкой плотности (10 3 клеток см -2 ) на макроводорослях-хозяевах ( U.lactuca и Ulvaria fusca) в полевых условиях (Skovhus et al ., 2007), эти плотности по-прежнему достаточны для подавления обрастания спорами макроводорослей, морскими грибами и личинками беспозвоночных (Rao et al . , 2007) . Аналогичные наблюдения были зарегистрированы для противообрастающих свойств P. gallaeciensi s 2,10 (Rao et al ., 2007).
, 2007) . Аналогичные наблюдения были зарегистрированы для противообрастающих свойств P. gallaeciensi s 2,10 (Rao et al ., 2007).
Противообрастающая и антимикробная активность обнаружена в широком диапазоне таксономических групп бактерий. Например, бурая ламинария S.Latissima (ранее Laminaria Saccharina ) Harbors более 100 различных антимикробных штаммов, покрывающих Phyla протеабактерии , бактериоиды , 70009 и актинобактерий (Wiese et al ., 2009). В другом исследовании было идентифицировано 30 штаммов с антимикробной активностью из D. pulchra и U. lactuca . Хотя у них была одна и та же широкая таксономическая классификация, между двумя макроводорослями-хозяевами было мало совпадений на уровне видов или родов (Penesyan et al ., 2009). Большинство исследований, направленных на оценку противообрастающих/противомикробных свойств эпифитных бактерий, были сосредоточены на культивируемых штаммах и, следовательно, вероятно, упустили потенциал некультивируемой части сообщества. Действительно, это мнение было недавно подтверждено результатами функционального метагеномного скрининга микробных сообществ, связанных с морскими губками и макроводорослями, в которых были обнаружены новые классы антибактериальных белков (Yung et al ., 2011).
Действительно, это мнение было недавно подтверждено результатами функционального метагеномного скрининга микробных сообществ, связанных с морскими губками и макроводорослями, в которых были обнаружены новые классы антибактериальных белков (Yung et al ., 2011).
Нарушение холобионта макроводорослей бактериальными патогенами
Микроорганизмы получают все большее признание в связи с их этиологической ролью в качестве возбудителей болезней морских животных, растений и водорослей. Этот интерес к микробным заболеваниям в морских экосистемах отчасти обусловлен опасениями, что стресс, связанный с изменением климата, на формирующих морскую среду обитания (кораллы, макроводоросли и т. д.) и связанный с ними микробиом сделает их более восприимчивыми к потенциальным условно-патогенным микроорганизмам (Harvell ). и др. ., 1999). Патогены макроводорослей разнообразны и включают вирусы, эукариотических паразитов и бактерий.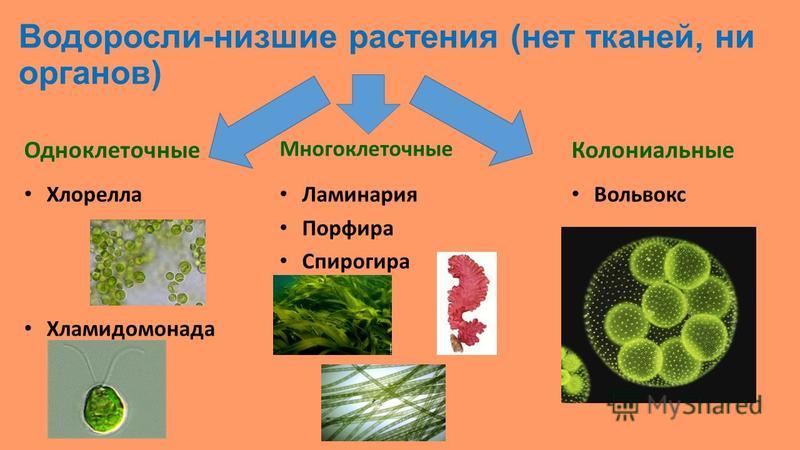 Здесь мы сосредоточимся на современных знаниях о взаимодействии макроводорослей с бактериальными патогенами. Для получения подробной информации о других микробных патогенах читатели могут обратиться к недавнему обзору Gachon et al . (2010), в котором преимущественно обсуждаются эукариотические и вирусные патогены и их роль в качестве движущих сил функционирования экосистемы и эволюции макроводорослей.
Здесь мы сосредоточимся на современных знаниях о взаимодействии макроводорослей с бактериальными патогенами. Для получения подробной информации о других микробных патогенах читатели могут обратиться к недавнему обзору Gachon et al . (2010), в котором преимущественно обсуждаются эукариотические и вирусные патогены и их роль в качестве движущих сил функционирования экосистемы и эволюции макроводорослей.
Изучение бактериальных возбудителей макроводорослей все еще находится в зачаточном состоянии.Отличие возбудителей от других условно-патогенных бактерий остается одним из основных препятствий к разграничению механизмов вирулентности от сапрофитных процессов. Однако одним из примеров, когда был достигнут существенный прогресс в понимании этих аспектов, является болезнь обесцвечивания D. pulchra . Здесь было показано, что два патогена ( Nautella italic sp. R11 и P. galleciensis LSS9) колонизируют и инфицируют D. pulchra в лабораторных условиях, что приводит к обесцвечиванию слоевища, аналогичному наблюдаемому в полевых условиях (случай и др. ., 2011; Fernandes и др. ., 2011). Эти две бактерии таксономически принадлежат к морской кладе Roseobacter , а другие представители этой группы вызывают галлоподобные опухоли у красной водоросли Prionitis (Ashen & Goff, 2000).
., 2011; Fernandes и др. ., 2011). Эти две бактерии таксономически принадлежат к морской кладе Roseobacter , а другие представители этой группы вызывают галлоподобные опухоли у красной водоросли Prionitis (Ashen & Goff, 2000).
Доступность культивируемых патогенов для болезни обесцвечивания D. pulchra открыла путь к определению молекулярных механизмов патогенности. Сравнительная геномика N. italic sp. R11 и P. galleciensis LSS9 с 18 близкородственными непатогенными бактериями выявили наличие у этих штаммов нескольких предполагаемых генов вирулентности (Fernandes et al ., 2011). Было обнаружено, что один ген, уникальный для обоих патогенов, кодирует активатор транскрипции типа Lux-R, аналогичный тем, которые участвуют в AHL-опосредованном ощущении кворума (QS), которое, как известно, у нескольких хорошо охарактеризованных патогенов регулирует колонизацию и вирулентность (Venturi, 2006). ; Barnard et al .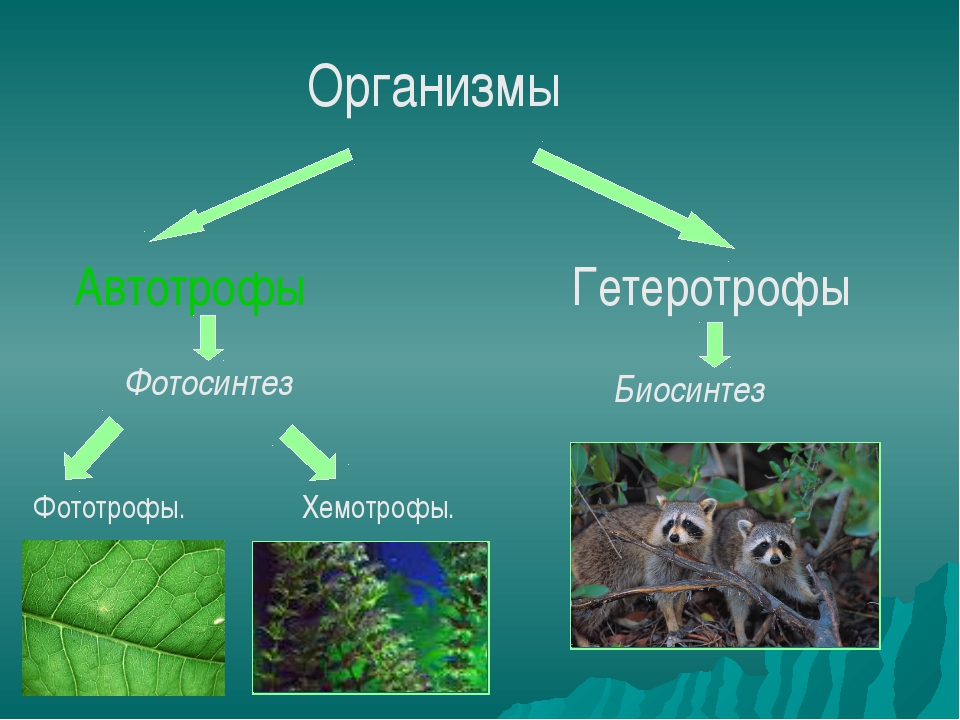 , 2007; Charkowski, 2009).
, 2007; Charkowski, 2009).
Патогенность, основанная на системе QS типа AHL, обеспечивает экологическую связь с химической защитой D. pulchra (см. выше), основанной на фуранонах, которые действуют как блокаторы QS (Givskov et al ., 1996). Таким образом, здоровый химически защищенный D. pulchra может обладать способностью подавлять экспрессию генов вирулентности (и, следовательно, заболевание) с помощью N. italic sp. R11 и P. galleciensis LSS9. Интересно, что в летние месяцы, когда D. pulchra теряет свои фураноны, наблюдается более высокая частота обесцвечивания (Campbell et al ., 2011). Эти наблюдения согласуются с моделью, согласно которой N. italic sp. R11 и P. galleciensis LSS9 переходят от комменсальных признаков к патогенным посредством активации механизмов вирулентности на основе QS.
Если такая модель верна, то поверхности макроводорослей могут содержать другие бактериальные патогены, которые экспрессируют гены вирулентности только при определенных условиях или когда хозяин скомпрометирован (например, условно-патогенные микроорганизмы). Действительно, отпечатки пальцев сообщества (t-RFLP и DGGE) и библиотеки клонов генов 16S рРНК подтвердили, что многие представители бактерий различаются как по численности, так и по наличию/отсутствию между здоровыми и больными D. pulchra (Campbell et al ., 2011, Fernandes). и др., неопубликовано). Например, бактерии, принадлежащие к таксонам Colwelliaceae, Thalassomonas, Rhodobacteraceae, и Celluophaga , были в большом количестве в обесцвеченных тканях и отсутствовали или их количество было снижено в здоровых тканях. Кроме того, метагеномный анализ выявил изменения в функциональности, при этом сообщество пораженной ткани было обогащено продукцией вторичных метаболитов, транспортными системами, хемотаксисом и генной регуляцией.
Действительно, отпечатки пальцев сообщества (t-RFLP и DGGE) и библиотеки клонов генов 16S рРНК подтвердили, что многие представители бактерий различаются как по численности, так и по наличию/отсутствию между здоровыми и больными D. pulchra (Campbell et al ., 2011, Fernandes). и др., неопубликовано). Например, бактерии, принадлежащие к таксонам Colwelliaceae, Thalassomonas, Rhodobacteraceae, и Celluophaga , были в большом количестве в обесцвеченных тканях и отсутствовали или их количество было снижено в здоровых тканях. Кроме того, метагеномный анализ выявил изменения в функциональности, при этом сообщество пораженной ткани было обогащено продукцией вторичных метаболитов, транспортными системами, хемотаксисом и генной регуляцией.
Обогащение некоторыми бактериями также наблюдалось при гниении водорослей (см. Gachon et al ., 2010). Например, Ван и др. . (2008) культивировали большое количество бактерий из талломов Laminaria japonica , у которых проявлялись симптомы дырявой болезни, и обнаружили поразительное количество Pseudoalteromonas sp. и Vibrio sp. Хотя повторное заражение ткани ламинарии этими штаммами действительно приводило к наблюдаемым симптомам, не было предпринято никаких попыток повторно выделить потенциальный патоген (т.е. продемонстрировать постулаты Коха), и, таким образом, неясно, действительно ли эти штаммы являются истинными возбудителями болезни.Действительно, вполне вероятно, что некоторые из бактерий, обнаруженных на пораженной ткани макроводорослей (в том числе бактерии на D. pulchra ), являются вторичными колонизаторами, которые потенциально действуют как сапрофиты или разрушители. Как культуральные, так и более поздние геномные исследования показали, что многие бактерии, связанные с макроводорослями, содержат ферменты для деградации сложных полисахаридов, составляющих клетки макроводорослей (Sakai et al ., 2003; Kalhoefer et al ., 2011). Таким образом, вполне вероятно, что определенные бактериальные эпифиты могут быть комменсальными, но в условиях инфекции или стресса макроводоросли-хозяина они становятся преимущественно сапрофитными.
и Vibrio sp. Хотя повторное заражение ткани ламинарии этими штаммами действительно приводило к наблюдаемым симптомам, не было предпринято никаких попыток повторно выделить потенциальный патоген (т.е. продемонстрировать постулаты Коха), и, таким образом, неясно, действительно ли эти штаммы являются истинными возбудителями болезни.Действительно, вполне вероятно, что некоторые из бактерий, обнаруженных на пораженной ткани макроводорослей (в том числе бактерии на D. pulchra ), являются вторичными колонизаторами, которые потенциально действуют как сапрофиты или разрушители. Как культуральные, так и более поздние геномные исследования показали, что многие бактерии, связанные с макроводорослями, содержат ферменты для деградации сложных полисахаридов, составляющих клетки макроводорослей (Sakai et al ., 2003; Kalhoefer et al ., 2011). Таким образом, вполне вероятно, что определенные бактериальные эпифиты могут быть комменсальными, но в условиях инфекции или стресса макроводоросли-хозяина они становятся преимущественно сапрофитными.
Болезнь морских макроводорослей отмечается уже много лет; однако наблюдения и модели, полученные из D. pulchra и других макроводорослей, теперь указывают на сложное взаимодействие между хозяином и микробным сообществом, которое ранее не оценивалось. Более того, работа над D. pulchra показала, что секвенирование генома патогенов макроводорослей и сравнительный метагеномный анализ больных и здоровых макроводорослей могут быстро дать представление об экологии и функциях болезней.Эти знания, применяемые в рамках существующей морской и химической экологии, обеспечивают мощный инструмент системной биологии для создания и последующей проверки новых гипотез.
Другие микробные представители голобионта морских водорослей
На сегодняшний день подавляющее большинство исследований, связанных с микробиомом макроводорослей, сосредоточены на бактериях. Интересно, что в значительной степени историческое внимание к бактериям согласуется с недавним анализом метагенома и транскриптома, который указывает на то, что бактерии действительно доминируют в этих сообществах (Burke et al . , 2011б; de Oliveira и др. ., 2012). Тем не менее, имея в виду концепцию холобионтов, важно также учитывать роль других микробов, связанных с хозяином (например, архей, эукариотических протистов и вирусов).
, 2011б; de Oliveira и др. ., 2012). Тем не менее, имея в виду концепцию холобионтов, важно также учитывать роль других микробов, связанных с хозяином (например, архей, эукариотических протистов и вирусов).
Мезофильные Crenarchaeota были обнаружены во многих морских местообитаниях, в том числе у сидячих беспозвоночных, таких как губки, где, как считается, они играют ключевую роль в окислении аммиака (Taylor et al ., 2007; Turque et al ., 2010). Археи, окисляющие аммоний, также были обнаружены на некоторых макроводорослях-хозяевах, однако, в отличие от других морских местообитаний, они недостаточно представлены по сравнению со своими бактериальными аналогами (Trias et al ., 2012). Более того, археи составляют лишь незначительную часть эпифитного микробиома U. lactuca (Burke et al ., 2011b). Хотя эти наблюдения подразумевают, что археи играют второстепенную роль в голобионте морских водорослей, необходимы дополнительные исследования, чтобы определить их потенциал в качестве эпифитов макроводорослей.
В ряде исследований сообщается о численности и разнообразии различных групп эукариотических микробов, в том числе динофлагеллят (Armstrong et al ., 2000; Porto et al ., 2008), инфузорий (Armstrong et al . , 2000), диатомовые водоросли (Armstrong et al ., 2000), амебы (Rogerson, 1991; Armstrong et al ., 2000) и грибы (Zuccaro et al ., 2008). Анализ последовательностей гена 18S рРНК из микробиома U. lactuca дополнительно выявил присутствие простейших, включая инфузорию Ephelota sp., гриб Tremisus helvelloides, и диатомовая водоросль Asterionellopsus glacialis (Burke et al ., 2009). Эпифитные эукариоты могут иметь патогенные или сапрофитные взаимодействия со своим хозяином, такие как грибковая инвазия и некроз ткани водорослей (Kawamura et al ., 2005), заражение ткани водорослей оомицетами (водяными плесенями) (Grenville-Briggs et al ., 2011) и образование опухолей у крупных водорослей (Goecke et al . , 2012). Морские эпифитные и эндофитные грибы также являются источником природных защитных соединений, которые можно использовать в качестве новых терапевтических средств (Rateb & Ebel, 2011), что может свидетельствовать о положительной или защитной роли грибов в холобионте.Однако по большей части экологическая роль эукариотических микроорганизмов в здоровье и функционировании водоросли-хозяина носит спекулятивный характер и остается в значительной степени неизвестной.
, 2012). Морские эпифитные и эндофитные грибы также являются источником природных защитных соединений, которые можно использовать в качестве новых терапевтических средств (Rateb & Ebel, 2011), что может свидетельствовать о положительной или защитной роли грибов в холобионте.Однако по большей части экологическая роль эукариотических микроорганизмов в здоровье и функционировании водоросли-хозяина носит спекулятивный характер и остается в значительной степени неизвестной.
Вирусы широко распространены в морской среде, и их роль в круговороте питательных веществ в океане тщательно изучалась в планктоне (см. (Suttle, 2005) и ссылки там). Что касается вирусов макроводорослей, вирусы нитчатой бурой водоросли Ectocarpus sp. возможно, являются наиболее изученными и наиболее разнообразными (Van Etten et al ., 2002; Дуниган и др. ., 2006). Эти крупные ДНК-вирусы заражают свободноживущие гаметы или споры, затем интегрируются в геном хозяина, где они остаются латентными в вегетативных частях водоросли, но становятся активными в репродуктивных клетках водорослей (Van Etten et al . , 2002). . Недавний анализ генома Ectocarpus siliculosus показал, что инфицировано до 50% естественной популяции водорослей (Cock et al ., 2010), что позволяет предположить, что вирусы могут сильно влиять на эволюцию и экологию макроводорослей.
, 2002). . Недавний анализ генома Ectocarpus siliculosus показал, что инфицировано до 50% естественной популяции водорослей (Cock et al ., 2010), что позволяет предположить, что вирусы могут сильно влиять на эволюцию и экологию макроводорослей.
Перспектива
Морские макроводоросли являются важными инженерами экосистемы, однако до недавнего времени было мало что известно о разнообразии и функциях связанного с ними бактериального сообщества. Эпифитные бактериальные сообщества, вероятно, состоят как из универсальных, так и из специализированных популяций и в значительной степени зависят от вида водорослей-хозяев, а также от географического положения. Хотя исследования разнообразия показали, что основные типы ( Proteobacteria , Cyanobacteria , Bacteroidetes ), которые являются обычными членами сообществ водорослей, существует мало доказательств, подтверждающих идею о том, что отдельные виды бактерий специфичны для хозяина. Скорее, возможно, что рекрутирование бактерий на поверхность водорослей (и, следовательно, специфичность хозяина) основано на выборе специфических функциональных признаков, таких как те, которые обсуждались выше.
Скорее, возможно, что рекрутирование бактерий на поверхность водорослей (и, следовательно, специфичность хозяина) основано на выборе специфических функциональных признаков, таких как те, которые обсуждались выше.
Независимо от механизма, сохранение определенных бактериальных групп и/или их функциональных признаков, вероятно, будет отражать их пользу для хозяина. В конечном итоге это взаимодействие приведет к развитию тесных отношений между водорослями и связанным с ними микробиомом, что поддержит концепцию холобионтов морских водорослей.Более того, данные о том, что бактерии и их вторичные метаболиты (например, АГЛ) являются важными сигналами для высвобождения спор водорослей (Weinberger et al ., 2007) и заселения [рассмотрено в (Joint et al ., 2007)], подчеркивают роль бактерий на ранних стадиях жизни макроводорослей, что выходит за рамки концепции холобионтов и направлено на колонизацию новых поверхностей.
Несмотря на то, что наука прошла долгий путь от первых наблюдений о том, что нативные бактерии необходимы для нормального морфологического развития макроводорослей, очевидно, что предстоит еще много работы, и, возможно, наиболее важным является сопоставление функциональных лабораторных исследований с результатами в природные сообщества. Детальное понимание механизма и функциональной роли всех микробных членов, будь то бактерии, археи, вирусы или эукариоты, в холобионте морских водорослей и их экологической роли в жизненном цикле водорослей было бы полезно для управления морскими водорослями как в естественных, так и в антропогенных условиях. -установки для аквакультуры. Роль микроорганизмов в болезнях водорослей вызывает растущий интерес, и будущая работа в этой области должна пролить свет не только на конкретные патогены водорослей, но и на потенциальный пробиотический эффект микробиома хозяина.В каждом случае остается проблема получения информации не только о механизмах этих специфических взаимодействий, но и об их экологической значимости. Чтобы достичь этого, будущие исследования должны отойти от преимущественно лабораторных экспериментов и сосредоточиться на получении надежных данных из манипулятивных исследований, проводимых в полевых условиях. Наконец, несмотря на то, что технологические достижения будут по-прежнему предоставлять инструменты для продвижения этих исследований, именно взаимодействие ученых с дополнительными навыками, включая химиков, экологов и микробиологов, обеспечит максимальное использование этих возможностей.
Детальное понимание механизма и функциональной роли всех микробных членов, будь то бактерии, археи, вирусы или эукариоты, в холобионте морских водорослей и их экологической роли в жизненном цикле водорослей было бы полезно для управления морскими водорослями как в естественных, так и в антропогенных условиях. -установки для аквакультуры. Роль микроорганизмов в болезнях водорослей вызывает растущий интерес, и будущая работа в этой области должна пролить свет не только на конкретные патогены водорослей, но и на потенциальный пробиотический эффект микробиома хозяина.В каждом случае остается проблема получения информации не только о механизмах этих специфических взаимодействий, но и об их экологической значимости. Чтобы достичь этого, будущие исследования должны отойти от преимущественно лабораторных экспериментов и сосредоточиться на получении надежных данных из манипулятивных исследований, проводимых в полевых условиях. Наконец, несмотря на то, что технологические достижения будут по-прежнему предоставлять инструменты для продвижения этих исследований, именно взаимодействие ученых с дополнительными навыками, включая химиков, экологов и микробиологов, обеспечит максимальное использование этих возможностей.
Благодарности
Авторы благодарят Австралийский исследовательский совет и Центр морских биоинноваций за поддержку исследования. Выражаю благодарность д-ру Ниине Туджула за использование ее микрофотографий и д-ру Шэрон Лонгфорд за графику и редактирование рукописи.
Каталожные номера
& (
2000
)Численность гетеротрофных протистов , связанных с литоральными водорослями
.Шельф эстуарного побережья Sci
50
:415
–424
.& (
2000
)Молекулярные и экологические доказательства видовой специфичности и коэволюции в группе морских водорослей-бактериальных симбиозов
.Appl Environ Microbiol
66
:3024
–3030
.& (
2010
)Идентификация соединений с биологической активностью против нематод Caenorhabditis elegans с помощью скрининга на основе функциональной геномики морской бактерии Pseudoalteromonas tunicata D2
.
Appl Environ Microbiol
76
:5710
–5717
.& (
2007
)Чувство кворума, вирулентность и продукция вторичных метаболитов у бактерий мягкой гнили растений
.Philos Trans R Soc Lond B Biol Sci
362
:1165
–1183
.& (
2011
)Микробное разнообразие, связанное с четырьмя функциональными группами бентических рифовых водорослей и рифообразующих кораллов Montastraea annularis
.Environ Microbiol
13
:1192
–1204
.& (
2008
)Физиология, филогения и in situ доказательства наличия бактериальных и архейных нитрификаторов в морской губке Aplysina aerophoba
.Environ Microbiol
10
:2942
–2955
.& (
2010
)Планктомицеты преобладают в биопленках на поверхности водорослей Laminaria hyperborea
.
BMC Microbiol
10
:261
–273
.& (
2010
)Сезонная динамика бактериальных биопленок на ламинарии Laminaria hyperborea
.Акват Микроб Экол
60
:71
–83
.& (
1988
)Популяционная динамика гетеротрофных бактериальных сообществ, связанных с Fucus vesiculosus и Ulva Rigida в эстуарии
.Микроб Экол
15
:345
–357
.& (
2011
)Комплексное производство макроводорослей для устойчивого биоэтанола, аквакультуры и сельского хозяйства в тихоокеанских островных государствах
.Биотопливо, Bioprod Biorefin
5
:599
–608
.& (
2009
)Микробные заболевания и голобионт кораллов
.Trends Microbiol
17
:554
–562
.(
2007
)Синтетическая способность биоактивных соединений и экологическое значение морских бактерий рода Pseudoalteromonas
.
Mar Наркотики
5
:220
–241
.& (
2008
)Разнообразие, экология и геномика клады Roseobacter: краткий обзор
.Arch Microbiol
189
:531
–539
.& (
2002
)Влияние полога водорослей на вертикальное распределение прибрежных комплексов на скалистых побережьях северо-западного Средиземноморья
.J Exp Mar Biol Ecol
267
:89
–106
.& (
2009
)Селективная экстракция бактериальной ДНК с поверхности макроводорослей
.Appl Environ Microbiol
75
:252
–256
.& (
2011a
)Сборка бактериального сообщества на основе функциональных генов , а не видов
.P Natl Acad Sci USA
108
:14288
–14293
.& (
2011b
)Состав, уникальность и изменчивость эпифитного бактериального сообщества зеленой водоросли Ulva australis
.
ISME J
5
:590
–600
.(
2000
)Bangiomorpha pubescens n. ген., н. sp.: значение для эволюции пола, многоклеточности и мезопротерозойской/неопротерозойской радиации эукариот
.Палеобиология
26
:386
–404
.& (
2011
)Изменение климата и болезни: обесцвечивание химически защищенных водорослей
.Glob Change Biol
17
:2958
–2970
.& (
2011
)Индуцированная температурой бактериальная вирулентность и обесцвечивание химически защищенных морских макроводорослей
.Environ Microbiol
13
:529
–537
.(
2009
)Сигналы разложения: поможет ли понимание связи между бактериями и растениями контролировать мягкую гниль?
Curr Opin Biotechnol
20
:178
–184
. и др. . (
. (
2010
)Геном Ectocarpus и независимая эволюция многоклеточности бурых водорослей
.Природа
465
:617
–621
.(
1978
)Присутствие эндофитных бактерий в сифонных водорослях
.Phycologia
17
:148
–151
.& (
2011
)Бактериальные симбионты и натуральные продукты
.Химическая коммуна
47
:7559
–7566
.& (
1977
)Микробные популяции , связанные с поверхностью бурых водорослей Ascophyllum nodosum
.Микроб Экол
4
:81
–91
.& (
2011
)Катаболизм диметилсульфониопропионата: микроорганизмы, ферменты и гены
.Nat Rev Microbiol
9
:849
–859
.& (
2002
)Воздействие бактерий, связанных с зеленой водорослью Ulva reticulata , на морское микро- и макрообрастание
.
Биообрастание
18
:217
–228
.и др. . (
2012
)Транскриптомный анализ красных морских водорослей Laurencia dendroidea (Florideophyceae, Rhodophyta) и их микробиома
.BMC Genomics.
13
:487
.& (
2006
)Ингибирование биообрастания морскими микроорганизмами и их метаболитами
.Биообрастание
22
:43
–54
.& (
2008
)Симбиотическое разнообразие морских животных: искусство использования хемосинтеза
.Nat Rev Microbiol
6
:725
–740
.& (
2006
)Фикоднавирусы: взгляд на генетическое разнообразие
.Virus Res
117
:119
–132
.& (
1999
)Локализация и поверхностный количественный анализ вторичных метаболитов в красных водорослях Delisea pulchra
.
Мар Биол
133
:727
–736
.& (
2001
)Ингибирование прорастания спор водорослей морской бактерией Pseudoalteromonas tunicata
.FEMS Microbiol Ecol
35
:67
–73
.& (
2008
)Раскрытие разнообразия и биотехнологического потенциала микробных сообществ , связанных с морской поверхностью
.Curr Opin Microbiol
11
:219
–225
.& (
2011
)Геномы и факторы вирулентности новых бактериальных патогенов, вызывающих обесцвечивание морских красных водорослей Delisea pulchra
.PLoS ONE
6
:e27387
.(
1975
)Некоторые наблюдения по морфологии Enteromorpha linza (L) J. Ag. и Enteromorpha Compressa (L.) Grev. в аксенической культуре
.Бот март
18
:251
–253
.
& (
2010
)Болезни водорослей: в центре внимания черный ящик
.Trends Plant Sci
15
:633
–640
.& (
2005
)Молекулярное разнообразие и экология микробного планктона
.Природа
437
:343
–348
.и др. . (
1996
)Эукариотическое вмешательство в передачу сигналов прокариот, опосредованную гомосеринлактоном
.J Бактериол
178
:6618
–6622
.& (
2010
)Химические взаимодействия между морскими макроводорослями и бактериями
.Mar Ecol Prog Ser
409
:267
–300
.& (
2012
)Новый фитомиксовый паразит, ассоциированный с галлами ламинарии быка Durvillaea antarctica (Chamisso) Hariot
.PLoS ONE
7
:e45358
.& (
1999
)Водоросли
.
Прентис-Холл
,Аппер-Сэдл-Ривер, Нью-Джерси
.& (
2011
)Молекулярный взгляд на войну водорослей и оомицетов: анализ кДНК Ectocarpus siliculosus , инфицированных базальным оомицетом Eurychasma dicksonii
.PLoS ONE
6
:e24500
.и др. . (
1999
)Обзор: Морская экология – возникающие морские болезни – климатические связи и антропогенные факторы
.Наука
285
:1505
–1510
.& (
2004
)Сезонные колебания противообрастающей активности морских водорослей на побережье Бретани (Франция)
.Мар Биотехнолог
6
:67
–82
.& (
2008
)LipL32 представляет собой взаимодействующий с внеклеточным матриксом белок Leptospira spp.и Pseudoalteromonas tunicata
.Infect Immun
76
:2063
–2069
.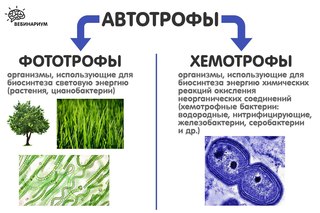
& (
2011a
)Жизнь без клеточной мембраны: вызов специфичности бактериальных эндофитов в пределах Bryopsis (Bryopsidales, Chlorophyta)
.BMC Microbiol
11
:255
.& (
2011b
)Кто там? Исследование эндофитных бактерий в сифонных зеленых водорослях Bryopsis (Bryopsidales, Chlorophyta)
.PLoS ONE
6
:e26458
.& (
2012
)Чему мы можем научиться у суши: обзор ассоциаций морских водорослей и бактерий
.FEMS Microbiol Ecol
83
:1
–16
.& (
2002
)Противообрастающая активность, выраженная морскими поверхностями Pseudoalteromonas видов
.FEMS Microbiol Ecol
41
:47
–58
.& (
2007
)Передача сигналов между царствами: использование молекул, чувствительных к бактериальному кворуму, зелеными водорослями Ulva
.
Philos Trans R Soc Lond B Biol Sci
362
:1223
–1233
.и др. . (
2011
)Сравнительный анализ генома и физиологический анализ на основе генома Roseobacter litoralis
.BMC Genomics
12
:324
.& (
2005
)Распространение Pythium porphyrae , возбудителя красной гнили Porphyrae spp., в море Ариаке
.Jpn Plant Dis
89
:1041
–1047
.& (
2000
)Бактериальные адгезины: функция и структура
.Int J Med Microbiol
290
:27
–35
.& (
2003
)Устойчивость морских водорослей к микробной атаке: целенаправленная химическая защита от морских грибков
.P Natl Acad Sci USA
100
:6916
–6921
.& (
2010
)Антидиатомовая и антибактериальная активность эпифитных бактерий , выделенных из Ulva lactuca в тропических водах
.
World J Microbiol Biotechnol
27
:1543
–1549
.& (
2009
)Специфические эпибактериальные сообщества макроводорослей: филогения важнее среды обитания
.Аква Биол
5
:181
–186
.& (
2010
)Выделенные соединения, связанные со талломом макроводоросли Fucus vesiculosus опосредуют бактериальную колонизацию поверхности в полевых условиях, аналогичную колонизации естественной водоросли
.Биообрастание
26
:247
–255
.& (
2011
)Образцы эпибактериальных сообществ морских макроводорослей зависят от хозяина, но меняются во времени
.Environ Microbiol
13
:655
–665
.& (
2007
)Морские макроводоросли влияют на численность и богатство сообщества бактериопланктона в непосредственной близости
.J Phycol
43
:874
–881
.
& (
2008
)Планктонные бактерии и грибы избирательно уничтожаются при контакте с морскими макроводорослями в непосредственной близости
.FEMS Microbiol Ecol
63
:283
–291
.и др. . (
2009
)Десорбционная масс-спектрометрия с ионизацией электрораспылением выявляет поверхностно-опосредованную противогрибковую химическую защиту тропических водорослей
.P Natl Acad Sci USA
106
:7314
–7319
.и др. . (
2007
)Сравнение разнообразия бактериальных сообществ , связанных с тремя сидячими морскими эукариотами
.Акват Микроб Экол
48
:217
–229
.& (
1997
)Производство диметилсульфида водорослями и его роль в атмосфере
.J Phycol
33
:889
–896
.& (
2006
)Влияние изолятов морских бактерий на рост и морфологию аксеничных проростков зеленой водоросли Ulva linza
.
Микробиол Экол
52
:302
–310
.& (
2003
)Выделение и филогенетическая характеристика бактерий , способных вызывать дифференцировку зеленых водорослей Monostroma oxyspermum
.Environ Microbiol
5
:25
–35
.& (
2005
)Выделение индуктора морфогенеза водорослей из морской бактерии
.Наука
307
:1598
.и др. . (
2008
)Морские биопленочные бактерии избегают эукариотического хищничества с помощью целенаправленной химической защиты
.PLoS ONE
3
:e2744
.и др. . (
1998
)Химическое опосредование бактериальной колонизации поверхности вторичными метаболитами красной водоросли Delisea pulchra
.Акват Микробиал Экол
15
:233
–246
.
& (
2001
)Филогенетический анализ Caulerpa Taxifolia (Chlorophyta) и связанной с ней бактериальной микрофлоры позволяет понять происхождение средиземноморской интродукции
.Мол Экол
10
:931
–946
.& (
1996
)Бактерии, вызывающие морфогенез у Ulva pertusa (Chlorophyta), выращенных в аксенических условиях
.J Phycol
32
:479
–482
.& (
1999
)Необходимое морфологическое взаимодействие для прикрепления между Ulva pertusa (Chlorophyta) и симбиотическими бактериями
.Мар Биотехнолог
1
:107
–111
.(
2009
)Существенная роль выращивания морских водорослей в интегрированных мультитрофических аквакультурных хозяйствах для глобального распространения марикультуры: анализ
.Девятнадцатый международный симпозиум по морским водорослям
, Vol.
2
(& , ред.), стр.117
–120
.Springer
,Нидерланды
.& (
2008
)Защита морских водорослей от бактерий: полибромированный 2-гептанон из красных водорослей Bonnemaisonia hamifera ингибирует бактериальную колонизацию
.Mar Ecol Prog Ser
369
:39
–50
.& (
2010
)Красная водоросль Bonnemaisonia asparagoides регулирует численность эпифитных бактерий и состав сообщества посредством химической защиты
.FEMS Microbiol Ecol
71
:84
–93
.& (
2011
)Разнообразные лектины, связывающие углеводы, из морских ресурсов
.Аминокислоты J
2011
:838914
.& (
2006
)Химическая защита красных водорослей от бактерий Asparagopsis armata : связующая структура с функцией
.Mar Ecol Prog Ser
306
:87
–101
.
& (
2009
)Антимикробная активность , наблюдаемая у культивируемых морских эпифитных бактерий , отражает их потенциал как источника новых лекарств
.FEMS Microbiol Ecol
69
:113
–124
.& (
1981
)Первичная продуктивность и азотфиксация в двух ассоциациях макроводорослей и цианобактерий
.Bull Mar Sci
31
:164
–169
.& (
2011
)Экологическая роль вторичного метаболита морских водорослей для колонизирующего бактериального сообщества
.Биообрастание
27
:579
–588
.& (
1990
)Фотосинтез, рост и фиксация азота эпифитными формами нитчатых цианобактерий из пелагического саргассума
.Bull Mar Sci
47
:613
–621
.& (
2008
)Связанные с макроводорослями динофлагелляты, принадлежащие к роду Symbiodinium на карибских рифах
.
PLoS ONE
3
:e2160
.& (
1980
)Индуцированный бактериями полиморфизм в аксеничном лабораторном штамме Ulva lactuca (Chlorophyceae)
.J Phycol
16
:196
–201
.& (
2005
)Конкурентные взаимодействия в смешанных биопленках, содержащих морскую бактерию Pseudoalteromonas tunicata
.Appl Environ Microbiol
71
:1729
–1736
.& (
2007
)Низкая плотность эпифитных бактерий из морских водорослей Ulva australis подавляет расселение обрастающих организмов
.Appl Environ Microbiol
73
:7844
–7852
.& (
2010
)Способность Pseudoalteromonas tunicata колонизировать естественные биопленки и ее влияние на структуру микробного сообщества
.FEMS Microbiol Ecol
73
:450
–457
.
& (
2011
)Вторичные метаболиты грибов из морских местообитаний
.Nat Prod Rep
28
:290
–344
.(
1991
)О численности морских голых амеб на поверхности пяти видов макроводорослей
.FEMS Microbiol Lett
85
:301
–312
.& (
2007
)Роль микроорганизмов в здоровье, болезнях и эволюции кораллов
.Nat Rev Microbiol
5
:355
–362
.& (
2001
)Лектины растений: возникновение, биохимия, функции и применение
.Glycoconj J
18
:589
–613
.& (
2011
)Ассоциированный с поверхностью фукоксантин опосредует заселение бактериальными эпифитами на камнеломке Fucus vesiculosus
.Биообрастание
27
:423
–433
.& (
2012
)Диметилсульфопропионат (ДМСП) и пролин с поверхности бурой водоросли Fucus vesiculosus ингибируют прикрепление бактерий
.
Биообрастание
28
:593
–604
.& (
2003
)Выделение и характеристика морской бактерии, разлагающей фукоидан
.Mar Biotechnol (NY)
5
:409
–416
.(
2006
)Заклепки или болты? При учете отдельных видов в функции сообществ скалистых рифов умеренного пояса
.J Exp Mar Biol Ecol
338
:233
–252
.& (
2006
)Популяционная биология крупных бурых водорослей: экологические последствия многофазных жизненных циклов в динамичных прибрежных средах
.Annu Rev Ecol Evol Syst
37
:343
–372
.& (
2007
)Градиенты нарушения водорослевого покрова и модификации литорального сообщества
.Mar Ecol Prog Ser
339
:1
–11
.& (
2011
)Химия Джекила и Хайда Phaeobacter gallaeciensis
.
Nat Chem
3
:331
–335
.& (
2011
)Выделение бактерий, ассоциированных с морскими водорослями, и их способность индуцировать морфогенез в аксенических культурах зеленых водорослей Ulva fasciata
.Аква Биол
12
:13
–21
.& (
2007
)Молекулярное исследование распространения, численности и разнообразия представителей рода Pseudoalteromonas в морских пробах
.FEMS Microbiol Ecol
61
:348
–361
.& (
2011
)Зеленая макроводоросль Dictyosphaeria ocellata влияет на структуру сообщества бактериопланктона посредством дифференцированного воздействия на отдельные бактериальные филотипы
.FEMS Microbiol Ecol
75
:242
–254
.& (
2010
)Микросенсорные измерения внешней и внутренней микросреды Fucus vesiculosus (Phaeophyceae)
.
J Phycol
46
:1350
–1355
.& (
2008
)Филогенетический анализ бактерий, ассоциированных с Laminaria saccharina
.FEMS Microbiol Ecol
64
:65
–77
.(
2005
)Вирусы в море
.Природа
437
:356
–361
.& (
2007
)Губкоассоциированные микроорганизмы: эволюция, экология и биотехнологический потенциал
.Microbiol Mol Biol Rev
71
:295
–347
.и др. . (
2012
)Phaeobacter gallaeciensis Геномы из глобально противоположных мест обнаруживают большое сходство адаптации к жизни на поверхности
.ИСМЭ J
.6
:2229
–2244
.и др. . (
2008
)Анализ генома Pseudoalteromonas tunicata выявляет свойства связанного с поверхностью образа жизни в морской среде
.
PLoS ONE
3
:e3252
.& (
2012
)Численность и состав эпифитных бактериальных и архейных окислителей аммиака морских красных и бурых макроводорослей
.Appl Environ Microbiol
78
:318
–325
.(
2006
)Анализ эпифитного бактериального сообщества зеленых водорослей Ulva australis
. Кандидатская диссертация,Университет Нового Южного Уэльса
,Сидней, Новый Южный Уэльс
.& (
2010
)Изменчивость и численность эпифитного бактериального сообщества , связанного с зеленой морской ульвовой водорослью
.ISME J
4
:301
–311
.и др. . (
2010
)Экологическое формирование сообществ архей , связанных с губками
.PLoS ONE
5
:e15774
.& (
2002
)Phycodnaviridae – крупные ДНК-вирусы водорослей
.
Арх Вирол
147
:1479
–1516
.(
2006
)Регулирование определения кворума в Pseudomonas
.FEMS Microbiol Rev
30
:274
–291
.& (
2012
)Вторая кожа: экологическая роль эпибиотических биопленок на морских организмах
.Front Microbiol
3
:292
.& (
2008
)Филогенетический анализ эпифитных морских бактерий на пораженных гнилью спорофитах Laminaria japonica
.J Appl Phycol
20
:403
–409
.и др. . (
2012
)Разработанная микробная платформа для прямого производства биотоплива из бурых макроводорослей
.Наука
335
:308
–313
.(
2007
)Патоген-индуцированная защита и врожденный иммунитет у макроводорослей
.
Биол Бык
213
:290
–302
.и др. . (
2007
)Высвобождение спор в Acrochaetium sp. (Rhodophyta) контролируется бактериями
.J Phycol
43
:235
–241
.& (
2009
)Разнообразие антибиотикоактивных бактерий, связанных с бурой водорослью Laminaria saccharina из Балтийского моря
.Мар Биотехнолог
11
:287
–300
.& (
1990
)Уровень хищничества молоди синих крабов в эстуарных местах нагула – свидетельство важности макроводорослей ( Ulva lactuca )
.Mar Ecol Prog Ser
58
:243
–251
.& (
2011
)Новые антибактериальные белки из микробных сообществ губки Cymbastela concentrica и зеленой водоросли Ulva australis
.Appl Environ Microbiol
77
:1512
–1515
.
и др. . (
2011
)Глобальные закономерности бактериального бета-разнообразия в экосистемах морского дна и морской воды
.PLoS ONE
6
:e24570
.& (
2008
)Обнаружение и идентификация грибов, тесно связанных с бурыми водорослями Fucus serratus
.Appl Environ Microbiol
74
:931
–941
.© 2013 Федерация европейских микробиологических обществ.
Рис – гетеротроф или автотроф? – Easyrwithpractice.com
Рис – гетеротроф или автотроф?
Однако эти растения сохраняют свои способности к фотосинтезу, и их даже можно успешно выращивать, не снабжая их съедобными насекомыми (рис). Согласно Rice (2007), это делает их лишь частично гетеротрофными, поскольку они по-прежнему получают большую часть своей энергии за счет фотосинтеза.
Что такое примеры автотрофов?
Растения, лишайники и водоросли являются примерами автотрофов, способных к фотосинтезу. Обратите внимание на их зеленый цвет из-за большого количества пигментов хлорофилла внутри их клеток. Этимология: от греческого autos, что означает «я», и trophe, что означает «питание». Синонимы: автофит; автотрофный организм; первичный производитель.
Обратите внимание на их зеленый цвет из-за большого количества пигментов хлорофилла внутри их клеток. Этимология: от греческого autos, что означает «я», и trophe, что означает «питание». Синонимы: автофит; автотрофный организм; первичный производитель.
Почему водоросли не растения?
Морские водоросли лишены сосудистой системы и корней растения; они могут поглощать воду и питательные вещества, в которых они нуждаются, непосредственно из окружающего их океана.Их лопасти (листообразные структуры) технически не являются листьями, поскольку у них нет жилок. Все эти водоросли можно найти на линии затонувших кораблей, особенно после шторма.
Полезны ли морские водоросли для собак?
Морские водоросли содержат широкий спектр питательных веществ, которые чрезвычайно полезны для вашей собаки. Среди них жирные кислоты Омега-3, железо, йод и магний. Было обнаружено, что водоросли помогают восстановить здоровье кожи и тканей вашей собаки.
Можно ли есть водоросли с пляжа?
Те водоросли, которые можно есть в сыром виде, можно есть либо свежими (из моря или с пляжа), либо сначала сушить, а затем жевать, как вяленое мясо.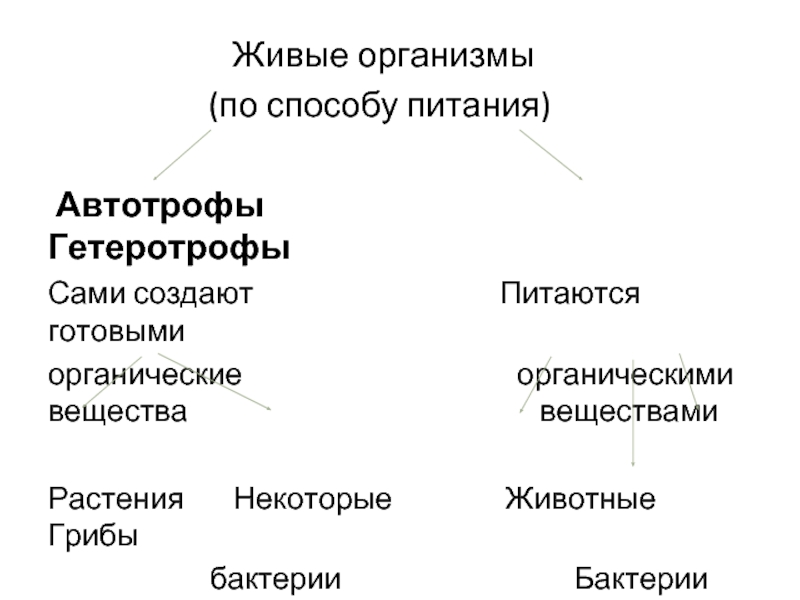 Кипячение предпочтительнее в некоторых случаях, когда водоросли полностью сухие.
Кипячение предпочтительнее в некоторых случаях, когда водоросли полностью сухие.
Что водоросли делают с водой?
Вода и питательные вещества Все части водорослей соприкасаются с водой, поэтому им не нужна внутренняя проводящая система, как наземным растениям; они поглощают то, что им нужно, непосредственно из воды через поверхностные ткани. Таким образом, вода вокруг водорослей должна постоянно двигаться, принося свежие запасы питательных веществ и газов.
Есть ли в водорослях рыбий помет?
Многие виды морских водорослей нуждаются в воде, богатой неорганическими питательными веществами, такими как азот и фосфор, чтобы выжить.«Рыбий помет, — говорит Яриш, — состоит именно из этих питательных веществ и обеспечивает питание для морских водорослей.
Полезно ли есть водоросли?
Морские водоросли содержат много антиоксидантов в виде определенных витаминов (А, С и Е) и защитных пигментов. Он содержит приличное количество йода, микроэлемента, жизненно важного для здоровья и функции щитовидной железы.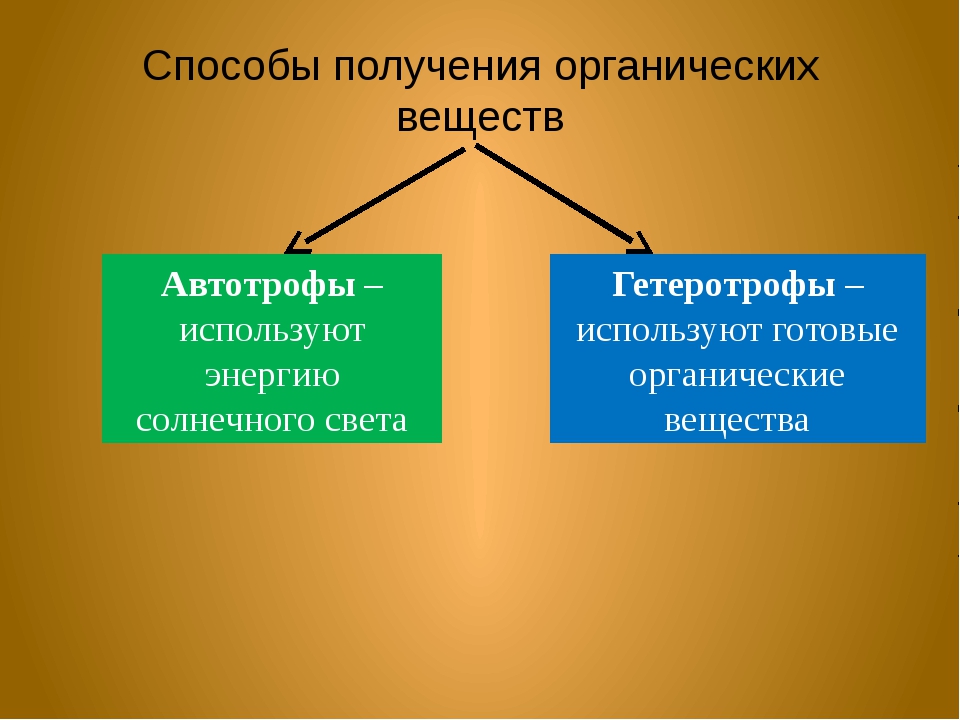 Некоторые морские водоросли, такие как пурпурная водоросль, также содержат большое количество B12.
Некоторые морские водоросли, такие как пурпурная водоросль, также содержат большое количество B12.
краеугольных камней | экология | Британика
Узнайте, как морские выдры поддерживают баланс экосистем водорослей и леса, поедая морских ежей
Узнайте о морской выдре ( Enhydra lutris ), ключевом виде в лесных экосистемах водорослей, обитающих вдоль тихоокеанского побережья Северной Америки.Основная его роль в лесу ламинарии состоит в том, чтобы контролировать популяции морских ежей.
Encyclopædia Britannica, Inc. Посмотреть все видео к этой статье ключевой вид , в экологии вид, оказывающий непропорционально большое влияние на сообщества, в которых он встречается. Такие виды помогают поддерживать местное биоразнообразие в сообществе, либо контролируя популяции других видов, которые в противном случае доминировали бы в сообществе, либо предоставляя важные ресурсы для широкого круга видов.Название краеугольных камней вида , придуманное американским зоологом Робертом Т. Пейном в 1969 году, произошло от практики использования клиновидного камня для поддержки вершины арки в мосту или другом сооружении. Точно так же, как другие камни в конструкции зависят от краеугольного камня для поддержки, другие виды в биологическом сообществе зависят от присутствия краеугольного камня для поддержания структуры сообщества.
Пейном в 1969 году, произошло от практики использования клиновидного камня для поддержки вершины арки в мосту или другом сооружении. Точно так же, как другие камни в конструкции зависят от краеугольного камня для поддержки, другие виды в биологическом сообществе зависят от присутствия краеугольного камня для поддержания структуры сообщества.
Морская звезда Pisaster ochraceus является краеугольным камнем каменистых морских литоральных сообществ у северо-западного побережья Северной Америки.Эта хищная морская звезда питается мидией Mytilus californianus и отвечает за сохранение большей части местного разнообразия видов в определенных сообществах. Когда морские звезды были удалены экспериментально, популяции мидий быстро увеличились и покрыли скалистые приливные берега настолько исключительно, что другие виды не могли прижиться. Следовательно, взаимодействие между Pisaster и Mytilus поддерживает структуру и видовое разнообразие этих сообществ. Однако в других сообществах, в которых встречается Pisaster , морская звезда в целом мало влияет на структуру сообщества. Следовательно, вид может быть ключевым видом в одних сообществах, но не в других.
Однако в других сообществах, в которых встречается Pisaster , морская звезда в целом мало влияет на структуру сообщества. Следовательно, вид может быть ключевым видом в одних сообществах, но не в других.
Подробнее по этой теме
Экология сообщества: ключевые виды
Однако даже полностью построенная пищевая сеть может дать лишь поверхностное и статичное представление о структуре биологических сообществ.Не…
В некоторых лесных сообществах тропической Америки инжир и несколько других растений действуют как краеугольные камни, но совершенно иначе, чем морская звезда Pisaster . Такие растения служат ключевыми пищевыми ресурсами. Инжир приносит плоды круглый год в некоторых из этих лесных сообществ, и большое количество птиц и млекопитающих в значительной степени зависят от этой небольшой группы видов растений в то время года, когда другие пищевые ресурсы скудны. Без инжира многие виды исчезли бы из сообщества.
Что такое водные растения и водоросли
Введение в водные растения и водоросли
Автотрофы — это организмы, которые могут производить себе пищу. Растение — известный пример автотрофа (рис. 2.2). Растения производят себе пищу в процессе фотосинтеза. Фотосинтез преобразует энергию света в химическую энергию и пищу. Фотосинтезирующих автотрофов также называют фотоавтотрофами. Этот термин отличает их от хемоавтотрофов, которые способны преобразовывать углекислый газ в химическую энергию и пищу без энергии света.
Фотосинтез — это процесс преобразования световой энергии солнца в химическую энергию. Фотосинтез начинается с захвата световой энергии в виде фотонов. Фотоны — это крошечные сгустки энергии. Как и все другие формы энергии, фотоны способны выполнять работу или перемещать предметы. Дополнительную информацию о фотосинтезе см. в разделе «Биогеохимические циклы».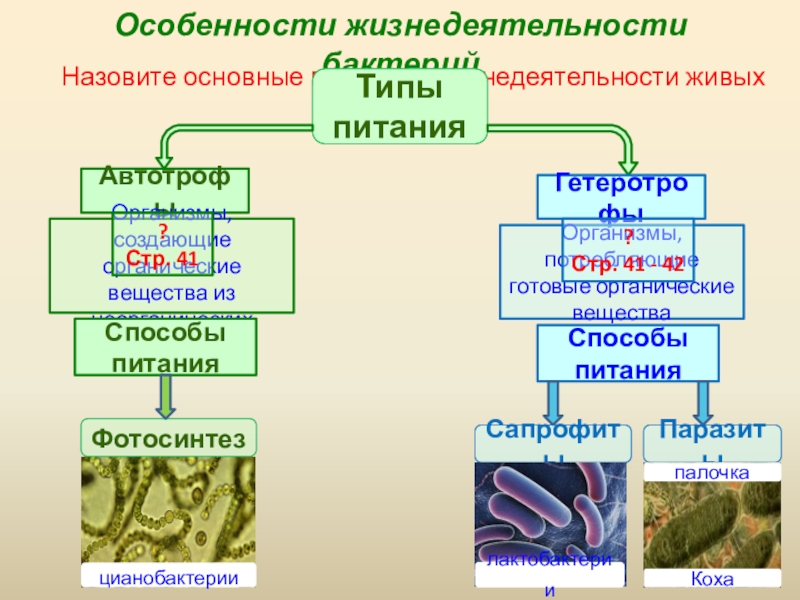
Фотоны поглощаются пигментами внутри автотрофных клеток. Пигменты — это химические соединения, которые кажутся красочными и поглощают свет, при этом приобретая определенный цвет. Пигменты встречаются во многих местах — цветы, кораллы и даже человеческая кожа имеют пигменты. Наиболее распространенным пигментом, участвующим в фотосинтезе, является зеленый пигмент хлорофилл.
Когда пигмент поглощает энергию фотонов, возбуждаются электроны молекул пигмента. Возбужденные электроны запускают ряд химических реакций, в результате которых образуются молекулы сахара на основе углерода, такие как глюкоза, из двуокиси углерода (CO 2 ) и воды (H 2 O).Химическая реакция фотосинтеза показана ниже:
| 6 CO 2 + | 6 Н 2 О | солнечный свет = | С 6 Н 12 О 6 + | 6 О 2 |
| двуокись углерода | вода | хлорофилл | глюкоза | кислород |
Этот процесс фотосинтеза преобразует энергию света в химическую энергию. Химическая энергия — это энергия, запасенная в химических связях. Распространенной формой химической энергии внутри живого организма является молекула сахара, называемая глюкозой (C 6 H 12 O 6 ). Глюкоза может использоваться для построения клеточных структур, таких как клеточные мембраны и клеточные стенки. Глюкоза также может быть объединена с другими питательными веществами для образования аминокислот и белков.
Химическая энергия — это энергия, запасенная в химических связях. Распространенной формой химической энергии внутри живого организма является молекула сахара, называемая глюкозой (C 6 H 12 O 6 ). Глюкоза может использоваться для построения клеточных структур, таких как клеточные мембраны и клеточные стенки. Глюкоза также может быть объединена с другими питательными веществами для образования аминокислот и белков.
Живые существа должны высвободить химическую энергию, хранящуюся в глюкозе, прежде чем они смогут использовать ее для роста, движения, размножения и других жизненных процессов. Клеточное дыхание — это процесс распада молекулы глюкозы с высвобождением накопленной химической энергии. Клеточное дыхание является обратным фотосинтезу.
В дополнение к производству глюкозы в процессе фотосинтеза также выделяется кислород. Кислород очень важен для жизни на Земле, потому что он необходим для клеточного дыхания. Фотосинтезирующие автотрофы делают возможной жизнь на Земле для других организмов. Интересно, что микроскопические автотрофы производят больше кислорода, чем водные и наземные растения вместе взятые.Они также несут ответственность за то, чтобы сделать землю пригодной для жизни. Первыми автотрофами были бактерии, называемые сине-зелеными водорослями или цианобактериями. Цианобактерии впервые появились на Земле более трех миллиардов лет назад, и они радикально изменили атмосферу планеты, увеличив количество кислорода.
Фотосинтезирующие автотрофы делают возможной жизнь на Земле для других организмов. Интересно, что микроскопические автотрофы производят больше кислорода, чем водные и наземные растения вместе взятые.Они также несут ответственность за то, чтобы сделать землю пригодной для жизни. Первыми автотрофами были бактерии, называемые сине-зелеными водорослями или цианобактериями. Цианобактерии впервые появились на Земле более трех миллиардов лет назад, и они радикально изменили атмосферу планеты, увеличив количество кислорода.
Водные растения
Растения являются известными примерами фотосинтезирующих автотрофов. Большинство видов растений встречаются в наземных местообитаниях. Люди в значительной степени зависят от наземных растений, таких как пшеница, кукуруза и помидоры, в качестве пищи.Многие растения также процветают в воде. Водные растения — это растения, обитающие в мелководных прибрежных зонах, водно-болотных угодьях, реках и озерах. Водные растения обеспечивают важную пищу и среду обитания для других организмов.
Прибрежные водные растения, такие как мангровые заросли (рис. 2.2 A) и болотные травы (рис. 2.2 D), могут переносить влажные условия, в которых наземные растения обычно тонут. Многие из этих прибрежных видов также приспособились к выживанию в соленой морской или солоноватой воде. Некоторые виды водных растений даже приспособились жить полностью погруженными в морскую воду (рис.2.2 Б) или пресноводных озер (рис. 2.2 В) вдали от необходимого для них солнечного света и углекислого газа.
Почти все виды растений развили сосудистую или жилкообразную ткань, которая переносит воду и питательные вещества по всему растению. Большинство растений также имеют отдельные корни, побеги и листья. Водные растения сохранили эти характеристики. Однако есть некоторые растения без сосудистых тканей — мохообразные виды, которые примерно напоминают некоторые морские водоросли или водоросли.
Водоросли
Термин водоросли (единственное число: водоросль ) относится к разнообразной группе фотосинтезирующих организмов, которые процветают в водной среде (рис. 2.3). Термин водорослей включает более 350 000 видов, представителей различных типов. Водоросли варьируются от одноклеточных микробов, плавающих в воде (рис. 2.3 A и 2.3 B), до высоких гигантских водорослей (рис. 2.3 D). Все водоросли и растения являются фотосинтезирующими автотрофами.
2.3). Термин водорослей включает более 350 000 видов, представителей различных типов. Водоросли варьируются от одноклеточных микробов, плавающих в воде (рис. 2.3 A и 2.3 B), до высоких гигантских водорослей (рис. 2.3 D). Все водоросли и растения являются фотосинтезирующими автотрофами.
Водоросли трудно определить, потому что этот термин описывает такое большое разнообразие организмов. Многие виды водорослей, такие как более крупные морские водоросли и гигантские ламинарии, кажутся похожими на растения (рис.2.3 С и D). Однако эти водоросли не являются настоящими растениями. У водорослей нет венозной сосудистой системы, которая есть у большинства растений.
Водоросли считаются самыми важными фотосинтезирующими организмами на Земле. Без водорослей жизнь в океане была бы совсем другой. Мелкие микроскопические водоросли служат основой большинства морских пищевых сетей (рис. 2.3 А и 2.3 В). Более крупные макроскопические водоросли обеспечивают пищу и трехмерную среду обитания для других организмов (рис. 2.3 C и 2.3 D).Эта роль водорослей в океане аналогична тому, как деревья обеспечивают пищу и среду обитания в лесу (рис. 2.3 D).
2.3 C и 2.3 D).Эта роль водорослей в океане аналогична тому, как деревья обеспечивают пищу и среду обитания в лесу (рис. 2.3 D).
Понимание эволюционных отношений между организмами важно для биологов, которые хотят знать, как эволюционировали сообщества видов. Молекулярная генетика помогла получить информацию о многих из этих взаимосвязей у водорослей. Однако остается еще много непонятного.Водоросли как группа имеют представителей в нескольких отдаленно родственных ветвях древа жизни (рис. 2.4). Например, цианобактерии или «сине-зеленые водоросли» — это одноклеточные организмы из домена Bacteria. Красные водоросли Rhodophyta — это многоклеточные организмы из домена Eukarya. А в области Eukarya водоросли распространены среди многих основных отделов. Для получения дополнительной информации о филогенетических деревьях см. тему «Классификация жизни».
Трудно определить и классифицировать виды водорослей.Большинство учебников и полевых справочников делят водоросли на широкие группы в зависимости от типа содержащихся в них фотосинтетических пигментов.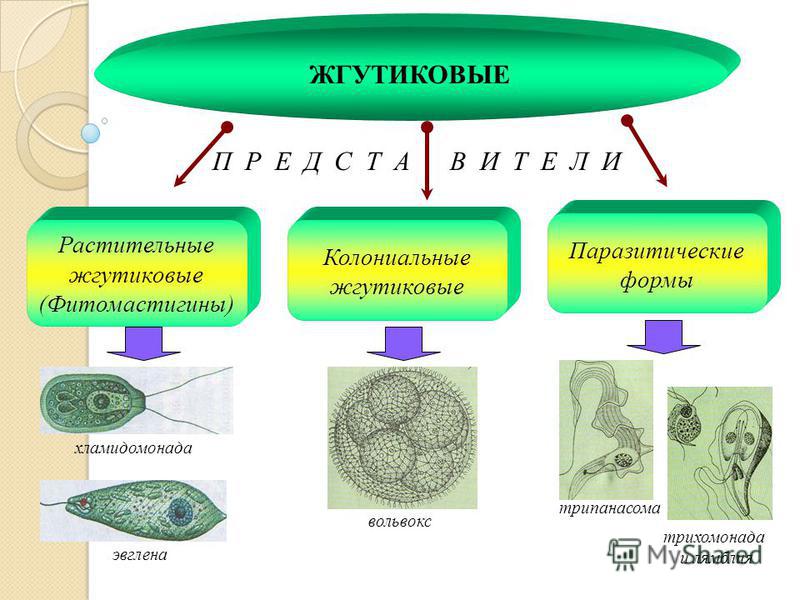 Крупные растительноподобные водоросли, или макроводоросли, обычно подразделяются на три группы: Chlorophyta (зеленые водоросли), Rhodophyta (красные водоросли) и Phaeophyceae (бурые водоросли). Микроскопические водоросли включают диатомовые водоросли и динофлагелляты. Цианобактерии (также называемые сине-зелеными водорослями) технически являются бактериями. Однако цианобактерии относятся к водорослям, потому что они фотосинтезируют и образуют большие колонии.
Крупные растительноподобные водоросли, или макроводоросли, обычно подразделяются на три группы: Chlorophyta (зеленые водоросли), Rhodophyta (красные водоросли) и Phaeophyceae (бурые водоросли). Микроскопические водоросли включают диатомовые водоросли и динофлагелляты. Цианобактерии (также называемые сине-зелеными водорослями) технически являются бактериями. Однако цианобактерии относятся к водорослям, потому что они фотосинтезируют и образуют большие колонии.
Прокариоты и эукариоты
Все живые организмы можно разделить на две широкие категории: прокариоты и эукариоты (рис. 2.5). К прокариотам относятся все организмы доменов Bacteria и Archaea (рис. 2.5). Прокариоты были первыми организмами, существовавшими на Земле. Они эволюционировали примерно 3,5 миллиарда лет назад.
Первые эукариотические организмы, для сравнения, эволюционировали намного позже — всего 1,6 к 2.2 миллиарда лет назад. Эукариоты включают все организмы из домена Eukaryota , а также растения, животных и грибы. Ученые подсчитали, что сегодня на Земле живет 8,7 миллиона видов эукариот, но пока официально описано только 1,2 миллиона видов.
Ученые подсчитали, что сегодня на Земле живет 8,7 миллиона видов эукариот, но пока официально описано только 1,2 миллиона видов.
Основные различия между прокариотами и эукариотами показаны в таблице 2.1. Клетки (термин клетка определен в теме «Свойства жизни») эукариот обычно более сложны, чем клетки прокариот (рис.2.6). Большинство водорослей являются эукариотами, хотя термин «водоросли» также используется для описания фотосинтезирующих прокариот, таких как цианобактерии.
| Прокариоты | Эукариоты | |
|---|---|---|
| Размер ячейки | 1–10 микрометров (мкм) | 10–100 мкм |
| Связанное с мембраной ядро | отсутствует | подарок |
| Связанные с мембраной органеллы | отсутствует | подарок |
| Клеточная мембрана | подарок | подарок |
| Клеточная стенка | обычно присутствует | присутствует в грибах, растениях, некоторых водорослях |
| Форма ДНК | часто круглая | хромосомы |
Основное различие между прокариотами и эукариотами заключается в их клеточном строении (рис. 2.6). Прокариоты — это одноклеточные организмы размером от одного до 10 микрометров (мкм). Как правило, они менее сложны, чем эукариотические клетки.
2.6). Прокариоты — это одноклеточные организмы размером от одного до 10 микрометров (мкм). Как правило, они менее сложны, чем эукариотические клетки.
Информационный бюллетень о каменистой траве — Признаки времен года: программа фенологии Новой Англии
Информационный бюллетень по камчатникам
Ascophyllum nodosum , более известный как Rockweed, представляет собой разновидность бурых водорослей или морских водорослей, встречающихся вдоль побережья Новой Англии. Он растет на доступных твердых поверхностях, включая камни, ракушки и сваи доков.Средняя длина каменной травы составляет от 20 до 30 дюймов, и она может расти длиннее там, где меньше волн, вызывающих поломку.
Почему именно этот вид?
Как член прибрежной среды обитания Новой Англии, Ascophyllum nodosum играет несколько важных ролей, влияющих на множество других морских видов. Во-первых, листья каменной травы создают защищенный навес для организмов. Эта защищенная среда обитания скрывает более мелкие организмы от хищников и может предотвратить высыхание литоральных видов, когда вода отступает во время отлива.
Эта защищенная среда обитания скрывает более мелкие организмы от хищников и может предотвратить высыхание литоральных видов, когда вода отступает во время отлива.
Менее очевидным является влияние каменной травы на химический состав морской воды. Увеличение концентрации двуокиси углерода (CO 2 ) в атмосфере из-за сжигания ископаемого топлива приводит к увеличению поглощения CO 2 океаном. Когда CO 2 растворяется в морской воде, образуется угольная кислота, и этот процесс называется «подкисление океана». Ряд химических реакций между CO 2 и морской водой снижает pH, делая воду более кислой.Это особенно проблематично для организмов, производящих раковины. Фотосинтезаторам, таким как каменные водоросли, требуется CO 2 , и они снижают его уровень, используя его для производства собственной пищи, тем самым также регулируя pH. Кроме того, кислород является полезным побочным продуктом фотосинтеза, который многие виды литоральных животных используют для дыхания.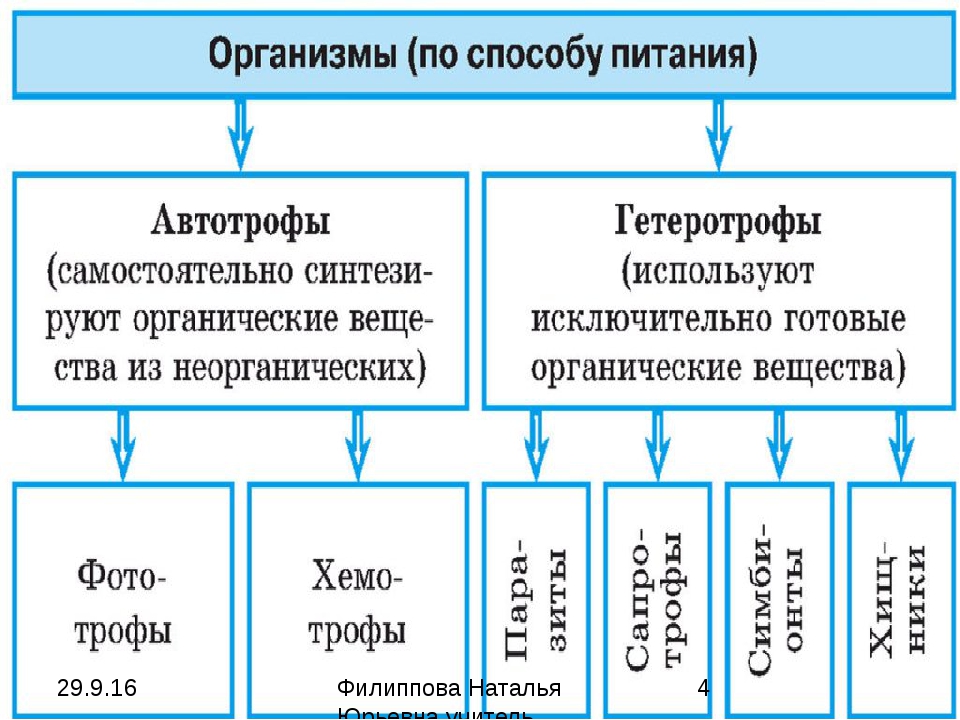 Регулируя эти растворенные газы, каменная трава помогает поддерживать качество воды вдоль побережья.
Регулируя эти растворенные газы, каменная трава помогает поддерживать качество воды вдоль побережья.
Наконец, каменные водоросли служат источником пищи на побережье.В то время как некоторые виды питаются самими листьями, другие могут извлечь выгоду из репродуктивных усилий каменных водорослей. Роквид размножается снаружи, выпуская половые клетки в воду. Такие организмы, как фильтраторы, могут потреблять эти клетки в качестве источника питательных веществ. В настоящее время ученые изучают роль репродуктивных клеток каменных водорослей в пищевой сети и могут ли литоральные организмы использовать эти питательные вещества в качестве источника энергии для своих собственных фенофазных изменений.
Морфология
Воздушный пузырь скальника внутри слоевища.Э. Коскела, UMaine Ext. Слоевище: Тело каменной травы, называемое слоевищем, состоит из длинных плоских листьев, лишенных средней жилки. Роквид показывает осевой рост, что означает, что ветвление происходит от центрального слоевища. Как новый, так и регенеративный рост происходит на концах листьев.
Как новый, так и регенеративный рост происходит на концах листьев.
Воздушные пузыри: Пузырьки, или воздушные пузыри, создают плавучесть для каменных водорослей. Они поднимают каменную траву к солнечному свету, необходимому для фотосинтеза. Воздушные пузыри располагаются отдельно вдоль слоевища и обычно формируются один раз в год.
Holdfast: Морские водоросли не имеют корней, потому что, в отличие от наземных растений, водоросли могут получать необходимые питательные вещества из окружающей морской воды. Хотя его внешний вид напоминает пучок корней, основная роль фиксатора заключается в том, чтобы закрепить каменную сорняк на твердой поверхности.
Сосуды: Размножение происходит весной или в начале лета, когда цветоложа, которые начали формироваться в предыдущем году, увеличиваются и развиваются. Изменение цвета и наличие выпуклых точек указывают на то, что цветоложе достигает зрелости.Когда окружающая морская вода достигает минимальной температуры, половые клетки (яйцеклетки и сперматозоиды) выбрасываются в воду для оплодотворения. Пустые сосуды отпадают от водорослей, когда размножение завершено.
Пустые сосуды отпадают от водорослей, когда размножение завершено.
Прирост: Начиная со второго года жизни, у каменной травы на кончике основной ветви появляется воздушный пузырь. Подсчет воздушных пузырей на самой длинной ветви может дать оценку возраста, хотя регенерация на кончике препятствует полной точности. Измерение расстояния между воздушными камерами показывает степень ежегодного роста, который составляет в среднем 3-4 дюйма вдоль побережья штата Мэн.
Адаптировано с сайта vector-eps Кэтлин Тенга-Гонсалес, Maine Sea Grant Коммерческое использование: Rockweed собирают для различных коммерческих целей. В качестве упаковочного материала для омаров или морских червей-наживок он обеспечивает необходимую влагу для живых организмов во время транспортировки. Rockweed добавляется непосредственно в сады в качестве удобрения и мульчи. При переработке он становится пищевой добавкой, обеспечивающей витамины и микроэлементы для корма для домашних животных и скота. Химическая экстракция удаляет соединение, называемое альгинатом, которое используется в качестве загустителя в продуктах питания, косметике и некоторых красках.
Химическая экстракция удаляет соединение, называемое альгинатом, которое используется в качестве загустителя в продуктах питания, косметике и некоторых красках.
Распространение: Роквид обычно встречается вдоль северного атлантического побережья Северной Америки. Его ареал простирается от Баффиновой Земли, Канада, на юг до Нью-Джерси.
Источники / Для получения дополнительной информации:
Исследовательский институт залива Мэн, родом из штата Мэн, Ascophyllum nodosum, http://www.gmri.org/upload/files/VS_Ascophyllum_nodosum.pdf
Департамент морских ресурсов штата Мэн, План управления промыслом каменной травы, январь 2014 г., https://www.maine.gov/dmr/science-research/species/rockweed/documents/DMRRockweedFMPJan2014.пдф
Maine SeaGrant, Rockweed: экология, промышленность и управление, https://eos.ucs.uri.edu/EOSWebOPAC/OPAC/Common/Pages/GetDoc.aspx?ClientID=EOSMAIN&MediaCode=15870
Уайт, Сьюзен. Полевой справочник по экономически важным водорослям северной Новой Англии .