
перевод, произношение, транскрипция, примеры использования
Как во множественном числе будет слово «mouse» (мышь)?
До меня доносился писк мыши. / Я слышал, как пищит мышь.
The cat laid a dead mouse at my feet.
Кот положил у моих ног дохлую мышь.
A witch had changed him into a mouse.
Одна ведьма превратила его в мышь.
To open the program, point at the icon and click the left mouse button.
Чтобы открыть программу, наведите указатель на значок и щёлкните левой кнопкой мыши.
A mouse scampered across the floor.
По полу опрометью пронеслась мышь.
The snake’s venom paralyzed the mouse.
Змеиный яд парализовал мышь.
He moved the mouse to click on the icon.
Он переместил курсор мыши,чтобы нажать на иконку.
The hawk gripped the mouse in its talons.
Ястреб схватил мышь когтями.
The house was infested with mice and rats.
В доме было полно мышей и крыс.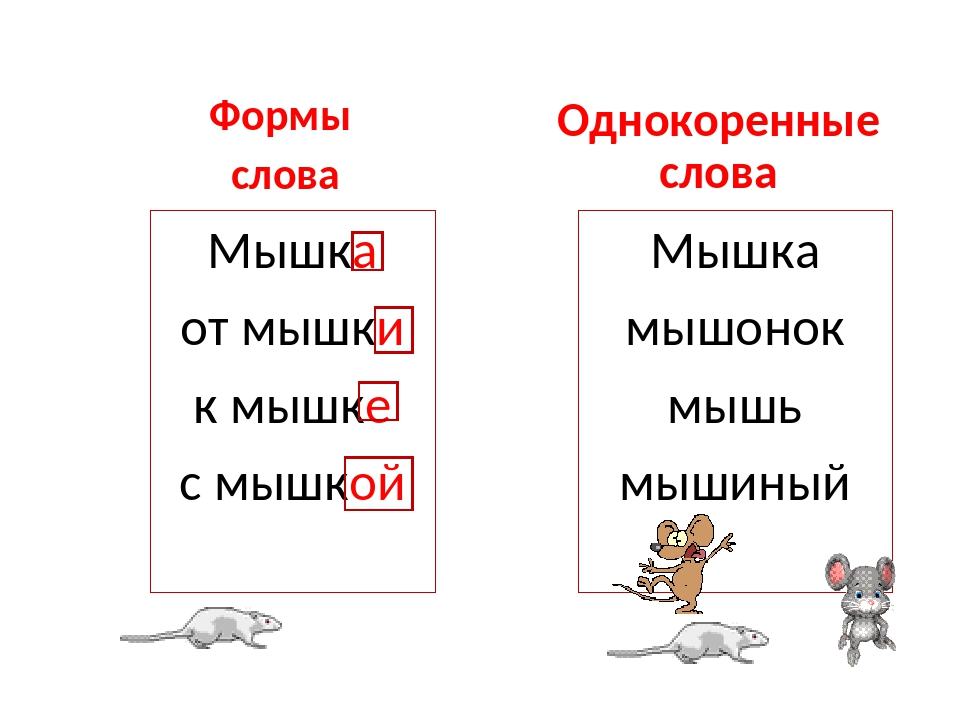
I know there’s a mouse under there somewhere.
Я точно знаю, что где-то там, внизу — мышь.
The company is just a Mickey Mouse operation.
Эта компания — просто шарашкина контора.
A mouse takes much more room than a trackball.
Мышь занимает гораздо больше места, чем шаровой указатель.
Click on the icon with the right mouse button.
Нажмите на иконку правой кнопкой мыши.
This game works with either a mouse or joystick.
Эта игра работает или с мышью, или при помощи джойстика.
The eagle was carrying a mouse in its sharp claws.
Орел нес мышь в своих острых когтях.
Cheese is very good for luring a mouse into a trap.
Сыр очень хорошо подходит для того, чтобы заманить мышь в мышеловку.
The mountain (in labour) has brought forth a mouse.
Гора родила мышь.
The computer gets its input from a keyboard or mouse.
Компьютер получает входные данные от клавиатуры или мыши.
I’m sure that was a mouse I saw scuttling across the mat.
Я уверена, что видела мышь, которая пробежала по ковру.
Select the printer icon and then click the left mouse button.
Выберите значок принтера, а затем щёлкните левой кнопкой мыши.
She squeaked out a cry of fear as the mouse ran across the floor.
Она завизжала от страха, когда по полу пробежала мышка.
The police played an elaborate game of cat and mouse to trap him.
Полицейские затеяли сложную игру в кошки-мышки, чтобы заманить его в ловушку.
The cat sat absolutely still, watching as the mouse began to make its way across the floor.
Кот сидел абсолютно неподвижно, наблюдая, как мышь начала пробираться по полу.
Морфологический разбор слова «мышь»
Часть речи: Существительное
МЫШЬ — слово может быть как одушевленное так и неодушевленное, смотрите по предложению в котором оно используется.
Начальная форма слова: «МЫШЬ»
| Слово | Морфологические признаки |
|---|---|
| МЫШЬ |
|
| МЫШЬ |
|
Все формы слова МЫШЬ
МЫШЬ, МЫШИ, МЫШЬЮ, МЫШЕЙ, МЫШАМ, МЫШАМИ, МЫШАХ
Разбор слова по составу мышь
| Основа слова | мышь |
|---|---|
| Корень | мышь |
| Нулевое окончание |
Разбор слова в тексте или предложении
Если вы хотите разобрать слово «МЫШЬ» в конкретном предложении или тексте, то лучше использовать
морфологический разбор текста.
Примеры предложений со словом «мышь»
1
Пусть это и усиленно сознающая мышь, но все-таки мышь, а тут человек, а следственно…, и проч.
Записки из подполья, Федор Достоевский, 1864г.2
если петух – так вылитый петух, если мышь – так вылитая мышь. Одно слово стоит тысячи, Лю Чжэньюнь, 2009г.3
Сплю я чутко и слышу даже когда мышь на мышь ползёт и от удовольствия попискивает.
4
Мышь разбегается и прыгает к нему на колени, маленькая, уютная, кротко свернувшаяся в клубок
5
Мышь, мышь, зубки мелкие-мелкие.
Ларочка, Михаил Попов, 2014г.Найти еще примеры предложений со словом МЫШЬ
Вывучаем беларускую » Мышка
МЫШКА
МЫШКА
(наз. , ж.р.)
, ж.р.)
(сущ., ж.р.)
1. прыстасаванне, якое ўжываецца падчас працы з графічным інтэрфейсам камп’ютара.
1. приспособление,которое употребляется во время работы с графическим интерфейсом компьютера.
2.маленькая шэрая жывёла.
2. маленькое серое животное.
Граматычныя формы
Грамматические формы
АДЗ.Л. ЕД.Ч. | МН.Л. МН.Ч. | |
| N | Мышка мышка | Мышкі мышки |
| N2 | няма мышкі нет мышки | Мышак мышек |
| N3 | спатрэбіцца мышцы понадобится мышке | Мышкам мышкам |
| N4 | бачыць мышку видеть мышку | Мышкі мышки |
| N5 | карыстацца мышкай пользоваться мышкой | Мышкамі мышками |
| N6 | размаўляць аб мышцы разговаривать о мышке | аб мышках о мышках |
| Чаргаванне К-Ц Чередование К-Ц |
Лексічная спалучальнасць
Лексическая сочетаемость
Pron. + N :мая / ваша мышка
+ N :мая / ваша мышка
Pron. + N: моя / ваша мышка
Adj. + N: аптычная,механічная.
Adj. + N: оптическая, механическая.
N + N2: функцыі мышкі, асаблівасці мышкі
N + N2: функции мышки, особенности мышки
N + Prep.+ N2 : мышка для работы
N + Prep.+ N2: мышка для работы
V + N4: набыць/купіць мышку
V + N4: приобрести/купить мышку
Prep.+ N2+ N2 :з дапамогай мышкі
Prep.+ N2+ N2: с помощью мышки
Аднакаранёвыя словы і іх спалучальнасць
Однокоренные слова и их сочетаемость
Мышыны (Adj. )— мышыны хвост
)— мышыны хвост
Мышиный (Adj.)— мышиный хвост
Ужыванне ў мове
Употребление в речи
— Камп’ютарную мышку кладуць з правага боку.
— Компьютерную мышку кладут с правой стороны.
— Сучасным вынаходніцтвам з’яўляецца аптычная мышка.
— Современным изобретением является оптическая мышка.
Ищем однокоренное слово
Однокоренными называются слова, сходные по смыслу и имеющие общую часть. Нетрудно догадаться, что этой общей частью должен быть корень (или один из корней, если слово сложное). Следовательно, в первую очередь следует уяснить то, что такое корень. Правило русского языка за 1 класс гласит, что корнем называется общая часть родственных слов, заключающая в себе их основное значение.
Подобно дереву, слово «вырастает» из корня.
Умение правильно подбирать родственные слова (они же – однокоренные) позволяет нам грамотно писать. Это касается такой орфограммы, как безударная гласная в корне (например, в слове «река» безударную «е» проверяет однокоренное слово «речка», в котором «е» находится под ударением).
Дети часто путают родственные связи с разными формами одного и того же слова. Так, для слова «мышка» — «мышь» – это однокоренное слово, а «мышки» — другая форма одного и того же. Формы слова меняются с изменением его окончания, а родственные слова образуются с помощью приставок и суффиксов. Причем приставки могут изменять смысл слова даже на противоположный. К примеру, антонимы ПРИезд и ОТъезд являются в то же время однокоренными словами.
Еще одна распространенная ошибка в определении однокоренных слов – принимать за родственные слова те, у которых корень совпадает по написанию и звучанию, но не по значению. Такие корни называют омонимичными. Например, в словах «носить», «поднос», «носильщик» – корень -нос-, в словах «нос», «носок», «переносица» – тоже -нос-, но нет ничего общего в их значениях. Слова с омонимичными корнями не являются однокоренными. Обратную ситуацию можно наблюдать в словах очень близких по смыслу, но не имеющих общего корня. Например: собака – щенок, лошадь — жеребенок и т.д. Русский язык не только богат, но и весьма непредсказуем!
Слова с омонимичными корнями не являются однокоренными. Обратную ситуацию можно наблюдать в словах очень близких по смыслу, но не имеющих общего корня. Например: собака – щенок, лошадь — жеребенок и т.д. Русский язык не только богат, но и весьма непредсказуем!
Однокоренные слова могут являться как к одной частью речи, так и разными (бег, бегун, бегунок – это существительные; бегать, бежать – глаголы; беговой, беглый – прилагательные; бегущий — причастие).
Интересным моментом в словообразовании является чередование звуков в корнях родственных слов, причем, как гласных (заря – зори, повесить – висел), так и согласных (друг-друзья-дружить, сушить – сохнуть). Возможны варианты, когда согласные чередуются с сочетаниями согласных (водить – вождение, ловить — ловля), а гласные — с «нулевым звуком», так называемой беглой гласной (отец – отца, сон – сна).
Однокоренное слово может иметь два и более корня. Такие слова называются сложными. Один корень в этом случае будет родственным, а другой послужит основой для образования нового по значению слова.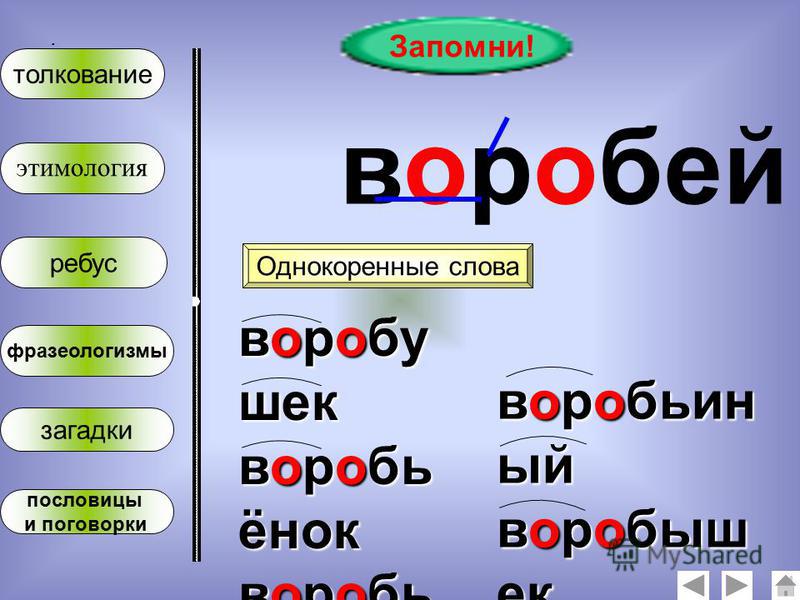 Например: зелень – вечнозеленый, земля – землепашец, электричество – гидроэлектростанция.
Например: зелень – вечнозеленый, земля – землепашец, электричество – гидроэлектростанция.
Все родственные слова, упорядоченные в отношении их производности, в совокупности образуют словообразовательное гнездо. В основании гнезда – вершина (исходное слово), от которой образуются производные однокоренные слова. Примеры: петь — певец, запеть – запевать – запевала; старый – стареть — устареть – устаревший. Словообразовательное гнездо может быть слаборазвернутым (до двух производных) и сильноразвернутым (вершина и более трех производных). Примером сильноразвернутого гнезда могут служить однокоренные слова к слову «дом»: домик, домишка, домище, домовой, придомовой, домашний, домовитый, домушник, надомный, бездомный, домосед, домочадцы, домовладелец, домоводство, домострой, домостроение, детдом, дурдом.
Теперь вы знаете, что такое однокоренное слово!
«Под мышкой» или «подмышкой» — как правильно?
Какими такими «мышками» мы носим предметы?
И как грамотно их носить: «под мышкой» или «подмышкой»?
Откуда в человеческом организме «мышки»?
Интересный факт: в русском языке слова «мышь» и «мускул» — однокоренные (так же, как и «бык» с «пчелой»).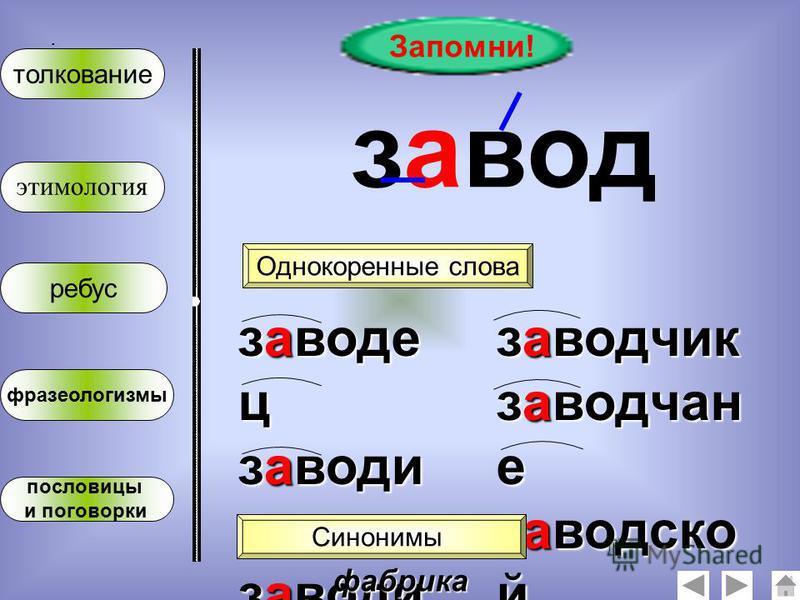 Оба имеют общие корни с латинским musculus — мускул, мышца, мышонок. Однако как часть тела может быть связана с животным?
Оба имеют общие корни с латинским musculus — мускул, мышца, мышонок. Однако как часть тела может быть связана с животным?
Чтобы это понять, придется немного напрячь фантазию. Нашим предкам игра мускулов под кожей чем-то напоминала суетливую мышиную возню, как будто там происходит не сокращение тканей, а забег спринтеров среди грызунов.
Почесали предки голову, да и придумали слово musculus от mus образовать (от «мыши», значит). С тех времен эта мышь-mus и понеслась во все европейские языки, где счастливо пребывает до сих пор.
Хитрое слово «под/мышки»
Не менее интересна история слова «под/мышки». «Под/мышка» — это просто то, что находится под мышцей. Но такая форма, видимо, русским людям показалась недостаточно милой, и «мышца» получила уменьшительную форму — «мышку».
А теперь проверьте себя: слитно или раздельно вы написали бы слово в этих примерах?
— Это платье мне жмет в под?мышках.
— Папа схватил ребенка под?мышки и перенес через лужу.
— Девушка сунула папку с отчетом под?мышку и побежала к начальнику.
— В этом костюме хорошо скроена под?мышка пиджака.
По ходу статьи вы узнаете правильные ответы. Переходим к главному.
Когда пишем слитно
«Подмышка» — это внутренняя часть плечевого сгиба или часть одежды, приходящаяся на это место (Большой толковый словарь русского языка, С. А. Кузнецов).
Если перед нами существительное «подмышка», то слово пишется слитно и изменяется по числам и падежам.
В творительном падеже слово приобретает форму «подмышкой» и тоже пишется слитно. В указанных выше примерах слитно также напишем:
— Это платье мне жмет в подмышках (= во внутренней части плечевого сгиба).
— В этом костюме хорошо скроена подмышка пиджака (= часть одежды, приходящаяся на подмышки).
Когда пишем раздельно
В русском языке есть так называемые застывшие формы существительных с предлогами, которые стали наречиями и пишутся раздельно. Например, на память, под утро, на карачках, не с руки. Сюда же относятся и наречия под мышкой, под мышки, из-под мышек, под мышку, под мышками.
Например, на память, под утро, на карачках, не с руки. Сюда же относятся и наречия под мышкой, под мышки, из-под мышек, под мышку, под мышками.
Важно понимать, что все перечисленные наречия — это разные слова, а не одно и то же слово, измененное по падежам и числам. Как узнать, что это именно тот случай, если возникло затруднение?
Подобные наречия, как правило, зависят от глагола-сказуемого и отвечают на обстоятельственные вопросы. В примерах выше раздельно следует написать:
— Папа схватил ребенка под мышки и перенес через лужу.
— Девушка сунула папку с отчетом под мышку и побежала к начальнику.
Признавайтесь, во всех 4 примерах ответили правильно?
https://zen.yandex.ru/media/berech_rech/pod-myshkoi-ili-podmyshkoi—kak-pravilno-i-pochemu-vdrug-u-cheloveka—myshki-5df9d445bb892c00affe3b9e
«Под мышкой» или «подмышкой» — как правильно? И почему вдруг у человека — «мышки» | Беречь речь
Какими такими «мышками» мы носим предметы? И как грамотно их носить: «под мышкой» или «подмышкой»? Ответы на эти вопросы ждут вас в статье.
Откуда в человеческом организме «мышки»?
Вот вам интересный факт: в русском языке слова «мышь» и «мускул» — однокоренные (так же, как и «бык» с «пчелой»). Оба имеют общие корни с латинским musculus — мускул, мышца, мышонок. Однако как часть тела может быть связана с животным?
Чтобы это понять, придется немного напрячь фантазию. Нашим предкам игра мускулов под кожей чем-то напоминала суетливую мышиную возню, как будто там происходит не сокращение тканей, а забег спринтеров среди грызунов.
Почесали предки голову, да и придумали слово musculus от mus образовать (от «мыши», значит). С тех времен эта мышь-mus и понеслась во все европейские языки, где счастливо пребывает до сих пор. (Источник: этимологический словарь Л.В. Успенского)
Хитрое слово «под/мышки»
Не менее интересна история слова «под/мышки». «Под/мышка» — это просто то, что находится под мышцей. Но такая форма, видимо, русским людям показалась недостаточно милой, и «мышца» получила уменьшительную форму — «мышку».
Но такая форма, видимо, русским людям показалась недостаточно милой, и «мышца» получила уменьшительную форму — «мышку».
А теперь проверьте себя: слитно или раздельно вы написали бы слово в этих примерах?
- Это платье мне жмет в под?мышках.
- Папа схватил ребенка под?мышки и перенес через лужу.
- Девушка сунула папку с отчетом под?мышку и побежала к начальнику.
- В этом костюме хорошо скроена под?мышка пиджака.
По ходу статьи вы узнаете правильные ответы. Переходим к главному.
Когда пишем слитноМой соавторМой соавтор
«Подмышка» — это внутренняя часть плечевого сгиба или часть одежды, приходящаяся на это место (Большой толковый словарь русского языка, С. А. Кузнецов).
Если перед нами существительное «подмышка», то слово пишется слитно и изменяется по числам и падежам.
В творительном падеже слово приобретает форму «подмышкой» и тоже пишется слитно. В указанных выше примерах слитно также напишем:
- Это платье мне жмет в подмышках (= во внутренней части плечевого сгиба).
- В этом костюме хорошо скроена подмышка пиджака (= часть одежды, приходящаяся на подмышки).
Когда пишем раздельно
Удивителен русский язык!Удивителен русский язык!
В русском языке есть так называемые застывшие формы существительных с предлогами, которые стали наречиями и пишутся раздельно. Например, на память, под утро, на карачках, не с руки. Сюда же относятся и наречия под мышкой, под мышки, из-под мышек, под мышку, под мышками. (Источник: «Русский язык. Орфография и пунктуация», Д.Розенталь)
Важно понимать, что все перечисленные наречия — это разные слова, а не одно и то же слово, измененное по падежам и числам. Как узнать, что это именно тот случай, если возникло затруднение?
Как узнать, что это именно тот случай, если возникло затруднение?
Подобные наречия, как правило, зависят от глагола-сказуемого и отвечают на обстоятельственные вопросы. В примерах выше раздельно следует написать:
- Папа схватил ребенка под мышки и перенес через лужу.
- Девушка сунула папку с отчетом под мышку и побежала к начальнику.
Признавайтесь, во всех 4 примерах ответили правильно?
Поставьте, пожалуйста, «палец вверх» и подпишитесь на канал – это очень поможет развитию проекта.
С уважением, автор канала «Беречь речь» – здесь пишут о русском языке с юмором и любовью.
Генная терапия с использованием Т-клеточных рецепторов человека и мыши опосредует регрессию рака и нацелена на нормальные ткани, экспрессирующие родственный антиген | Кровь
Антигены, ассоциированные с опухолью, были идентифицированы в широком спектре раковых заболеваний человека. Многие из этих антигенов являются нормальными, немутировавшими собственными белками, избирательно экспрессируемыми или сверхэкспрессируемыми при раке. 1 Антигены, такие как MART-1 и gp100, экспрессируются на меланомах и нормальных меланоцитах кожи, глаз и ушей. 2,3 Другие антигены, ассоциированные с раком, такие как карциноэмбриональный антиген (СЕА), Her2/neu и Muc-1, экспрессируются на низких уровнях в некоторых нормальных тканях, в то время как антигены, такие как NY-ESO-1 и семейство MAGE белки экспрессируются в тканях плода и семенниках взрослых, но не в других нормальных тканях взрослых. 4 Присутствие этих нормальных белков во время развития плода приводит к центральной толерантности, основанной на негативной селекции в тимусе клонов лимфоцитов, несущих высокоаффинные аутореактивные Т-клеточные рецепторы (TCR).Иногда лимфоциты, несущие высокоаффинный TCR, избегают делеции тимуса; и в этих случаях механизмы периферической толерантности могут подавлять их активность.
Многие из этих антигенов являются нормальными, немутировавшими собственными белками, избирательно экспрессируемыми или сверхэкспрессируемыми при раке. 1 Антигены, такие как MART-1 и gp100, экспрессируются на меланомах и нормальных меланоцитах кожи, глаз и ушей. 2,3 Другие антигены, ассоциированные с раком, такие как карциноэмбриональный антиген (СЕА), Her2/neu и Muc-1, экспрессируются на низких уровнях в некоторых нормальных тканях, в то время как антигены, такие как NY-ESO-1 и семейство MAGE белки экспрессируются в тканях плода и семенниках взрослых, но не в других нормальных тканях взрослых. 4 Присутствие этих нормальных белков во время развития плода приводит к центральной толерантности, основанной на негативной селекции в тимусе клонов лимфоцитов, несущих высокоаффинные аутореактивные Т-клеточные рецепторы (TCR).Иногда лимфоциты, несущие высокоаффинный TCR, избегают делеции тимуса; и в этих случаях механизмы периферической толерантности могут подавлять их активность. 5
5
Терапия переноса клеток появилась как инструмент для преодоления ограничений, налагаемых как центральной, так и периферической толерантностью. 6-9 Перенос противоопухолевых Т-клеток мышам с истощением лимфоузлов может опосредовать отторжение крупных васкуляризированных опухолей, 10 , а введение встречающихся в природе антимеланомных опухоле-инфильтрирующих лимфоцитов (TIL) может опосредовать объективную регрессию рака в 51–72 % случаев. % пациентов с лимфодеплецией и метастатической меланомой. 7,8,11
Основным препятствием для широкого применения методов переноса клеток является трудность идентификации человеческих Т-клеток с противоопухолевым распознаванием. Только примерно половина меланом воспроизводимо дает противоопухолевые TIL, 12 и другие типы рака лишь изредка содержат идентифицируемые опухолереактивные лимфоциты. Альтернативой нахождению этих естественных опухоле-реактивных клеток у каждого пациента является перенос в нормальные лимфоциты опухоле-реактивных генов TCR, распознающих общие опухолевые антигены.
Альтернативой нахождению этих естественных опухоле-реактивных клеток у каждого пациента является перенос в нормальные лимфоциты опухоле-реактивных генов TCR, распознающих общие опухолевые антигены.
В предыдущем исследовании мы идентифицировали TCR (MSGV1AIB, здесь обозначаемый как DMF4), который распознавал меланомный/меланоцитарный антиген MART-1, клонированный из TIL резецированного очага меланомы. 13 Мы пролечили 31 пациента с помощью аутологичных лимфоцитов периферической крови (PBL), трансдуцированных генами, кодирующими этот рецептор (ранее сообщалось о 17), 14 , и у 4 пациентов (13%) наблюдалась объективная регрессия метастатической меланомы.Ни у одного из 31 пациента не было кожной сыпи или обычной токсичности меланоцитов в глазах или ушах. Рецептор DMF4 обладал лишь умеренной способностью распознавать ограниченные количества антигена, и мы предположили, что более высокореактивный TCR может быть клинически более эффективным в распознавании злокачественных клеток, экспрессирующих антиген-мишень.
Таким образом, мы получили TCR с высокой авидностью из Т-клетки человека, которая распознала эпитоп MART-1:27-35 (обозначаемый здесь как DMF5). 15 Эпитоп gp100:154-162 из меланомно-меланоцитарного антигена gp100 представляет собой наиболее высокоэкспрессируемый пептид этого белка, экспонируемый на клеточной поверхности. Попытки создать TCR человека с высокой авидностью против этого эпитопа не увенчались успехом. Однако нам удалось создать высокоактивный TCR против этого эпитопа у мышей, трансгенных по человеческому лейкоцитарному антигену-A2 (HLA-A2). Эти анти-MART-1 и анти-gp100 TCR использовались для лечения пациентов с метастатической меланомой.
Тридцать шесть пациентов с метастатической меланомой проходили лечение в Хирургическом отделении Национального института рака в период с июля 2007 г. по март 2008 г. в соответствии с протоколами, одобренными Институциональным наблюдательным советом и Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов, 20 с TCR, признающим HLA-A*02. –рестриктированный антиген меланомы MART-1 (DMF5) 15 и 16 с TCR, распознающим HLA-A*02-рестриктированный антиген меланомы gp100(154). 16 Все пациенты дали информированное согласие на лечение в соответствии с Хельсинкской декларацией. Пациенты были HLA-A*02 + , в возрасте 18 лет и старше, имели измеримые метастатические меланомы и статус 0 или 1 Восточной кооперативной онкологической группы. У всех пациентов наблюдалось прогрессирование после предшествующего лечения интерлейкином-2 (ИЛ-2)/ Альдеслейкин и имели опухоли, которые экспрессировали соответствующий антиген (MART-1/Melan-A или gp100/HMB45). Противопоказания были следующими: сопутствующие серьезные медицинские заболевания, любая форма первичного или вторичного иммунодефицита, тяжелая гиперчувствительность к любому из агентов, использованных в этом исследовании, противопоказания для введения высоких доз ИЛ-2, лечение системными стероидами в течение 30 дней до лечения и нелеченные внутричерепные метастазы более 1.
по март 2008 г. в соответствии с протоколами, одобренными Институциональным наблюдательным советом и Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов, 20 с TCR, признающим HLA-A*02. –рестриктированный антиген меланомы MART-1 (DMF5) 15 и 16 с TCR, распознающим HLA-A*02-рестриктированный антиген меланомы gp100(154). 16 Все пациенты дали информированное согласие на лечение в соответствии с Хельсинкской декларацией. Пациенты были HLA-A*02 + , в возрасте 18 лет и старше, имели измеримые метастатические меланомы и статус 0 или 1 Восточной кооперативной онкологической группы. У всех пациентов наблюдалось прогрессирование после предшествующего лечения интерлейкином-2 (ИЛ-2)/ Альдеслейкин и имели опухоли, которые экспрессировали соответствующий антиген (MART-1/Melan-A или gp100/HMB45). Противопоказания были следующими: сопутствующие серьезные медицинские заболевания, любая форма первичного или вторичного иммунодефицита, тяжелая гиперчувствительность к любому из агентов, использованных в этом исследовании, противопоказания для введения высоких доз ИЛ-2, лечение системными стероидами в течение 30 дней до лечения и нелеченные внутричерепные метастазы более 1. 0 см в диаметре.
0 см в диаметре.
В общей сложности от 5 до 10 × 10 8 PBL пациентов были получены с помощью лейкафереза и стимулированы in vitro в концентрации 10 6 /мл с 50 нг/мл анти-CD3 mAb OKT-3 в полной среде AIMV (Invitrogen) с добавлением с 5% сывороткой человека (Surgery Branch, National Cancer Institute) и 300 МЕ IL-2 (среда TCR). Два дня спустя ретровирусный супернатант, кодирующий TCR, быстро оттаивали, разбавляли 1:1 в среде TCR и добавляли в планшеты, которые в течение ночи покрывали 10 мкг/мл ретронектина (Takara Bio Inc).Супернатант наносили на планшеты путем центрифугирования в течение 2 часов при 2000 г g при 32°С. Стимулированные PBL промывали и ресуспендировали в концентрации от 0,25 до 0,5 × 10 6 /мл в среде TCR, и добавляли от 1 до 2 × 10 6 PBL на лунку планшетов с ретровирусом. Планшеты вращали при 1000 g при 32°C в течение 10 минут и инкубировали в течение ночи при 37°C, 5% CO 2 . На следующий день (3-й день) PBL переносили в свежеприготовленные 6-луночные планшеты, покрытые ретровирусом, как и на 2-й день.На следующий день трансдуцированные PBL промывали, ресуспендировали в свежей среде TCR и переносили в колбы при 37°C, 5% CO 2 . На 9-12 день клетки размножались или не размножались еще 9-14 дней в 6000 МЕ IL-2 с 50 нг/мл анти-CD3 mAb OKT-3 и 100-кратным избытком облученных 5 Гр аллогенных фидерных клеток PBL. Перед лечением TCR-трансдуцированные PBL от всех пациентов оценивали на предмет экспрессии соответствующего TCR с помощью окрашивания тетрамером и проточного цитометрического анализа, а функцию клеток оценивали с помощью ночного совместного культивирования с родственными антиген-несущими клетками-мишенями (1 × 10 5 :1 × 10 5 ) и измерение с помощью иммуноферментного анализа (ELISA) (Pierce Endogen) интерферона-γ (IFN-γ), продуцируемого в супернатанте культуры.Лечебные клетки промывали физиологическим раствором перед внутривенной инфузией пациентам.
На следующий день (3-й день) PBL переносили в свежеприготовленные 6-луночные планшеты, покрытые ретровирусом, как и на 2-й день.На следующий день трансдуцированные PBL промывали, ресуспендировали в свежей среде TCR и переносили в колбы при 37°C, 5% CO 2 . На 9-12 день клетки размножались или не размножались еще 9-14 дней в 6000 МЕ IL-2 с 50 нг/мл анти-CD3 mAb OKT-3 и 100-кратным избытком облученных 5 Гр аллогенных фидерных клеток PBL. Перед лечением TCR-трансдуцированные PBL от всех пациентов оценивали на предмет экспрессии соответствующего TCR с помощью окрашивания тетрамером и проточного цитометрического анализа, а функцию клеток оценивали с помощью ночного совместного культивирования с родственными антиген-несущими клетками-мишенями (1 × 10 5 :1 × 10 5 ) и измерение с помощью иммуноферментного анализа (ELISA) (Pierce Endogen) интерферона-γ (IFN-γ), продуцируемого в супернатанте культуры.Лечебные клетки промывали физиологическим раствором перед внутривенной инфузией пациентам.
Регистрационные номера клинических испытаний и утвержденные регистрационные названия следующие: NCI-07-C-0175, Фаза II исследования метастатической меланомы с использованием лимфоистощающего кондиционирования с последующей инфузией анти-MART-1 F5 TCR-генно-инженерных лимфоцитов; NCI-07-C-0174, Фаза II исследования метастатической меланомы с использованием лимфоистощающего кондиционирования с последующей инфузией анти-gp100:154-162 лимфоцитов, сконструированных с помощью TCR-гена.
Перед лечением трансдуцированными PBL пациентам была проведена временная лимфоаблация с использованием немиелоаблативной лимфодеплетирующей схемы, как описано ранее, 8 путем внутривенного введения циклофосфамида 60 мг/кг в течение 2 дней с последующим введением флударабина 25 мг/м 2 в течение 5 дней. Через день после завершения курса лимфодеплетации пациентам внутривенно вводили трансдуцированные лимфоциты с последующим введением высоких доз (720 000 ЕД/кг) ИЛ-2 (Aldesleukin; Chiron Corp) каждые 8 часов до достижения толерантности. Пять пациентов с DMF5 (пациенты 7-11) и 4 пациента gp100(154) (пациенты 7-10) получали TCR-трансдуцированные клетки на 10-12 день после стимуляции. Остальные 15 пациентов с ДМФ5 и 12 пациентов с gp100(154) получили большее количество TCR-трансдуцированных клеток, которые выращивали в течение дополнительных 9-14 дней после второй стимуляции ОКТ-3.
Пять пациентов с DMF5 (пациенты 7-11) и 4 пациента gp100(154) (пациенты 7-10) получали TCR-трансдуцированные клетки на 10-12 день после стимуляции. Остальные 15 пациентов с ДМФ5 и 12 пациентов с gp100(154) получили большее количество TCR-трансдуцированных клеток, которые выращивали в течение дополнительных 9-14 дней после второй стимуляции ОКТ-3.
Пациенты прошли исходную компьютерную томографию (КТ) и/или магнитно-резонансную томографию перед лечением, а также прошли обследование глаз и аудиологию до и после лечения.Размер опухоли оценивали ежемесячно с помощью КТ, магнитно-резонансной томографии или фотографировали кожные/подкожные поражения. Измерения опухоли и реакцию пациента определяли в соответствии с Критериями оценки ответа при солидных опухолях (RECIST). 17 Образцы PBL и сыворотки пациентов были взяты после инфузии трансдуцированных PBL. Уровни цитокинов в сыворотке измеряли с помощью анализа ELISA (Pierce Endogen).
Были получены биоптаты кожи и опухолей и иммуногистохимически окрашены на наличие CD4 и CD8 Т-клеток.У одного пациента с увеитом глазную жидкость оценивали на наличие трансдуцированных противоопухолевых Т-клеток с помощью проточной цитометрии. Активность клеток оценивали путем совместного культивирования PBL пациентов с родственным антигеном на клетках-мишенях T2 или HLA-совместимых и несовместимых меланомах mel526, mel624 (HLA-A*02 + ) или mel888 и mel938 (не-HLA-A*02). . IL-2 и IFN-γ измеряли с помощью ELISA (Pierce Endogen), ELISPOT (реагенты от Mabtech Inc, Millipore Corp, Invitrogen, BD Biosciences PharMingen и Kirkegaard & Perry) или внутриклеточного окрашивания (mAb от eBiosciences) и проточной цитометрии.Лизис оценивали с помощью анализа высвобождения хрома клетками-мишенями 51 . После персистенции перенесенных клеток в крови проводили окрашивание тетрамера тетрамером HLA-A2/MART-1:27-35 или HLA-A2/gp100:154-162 (Beckman Coulter Immunotech).
Статистическую значимость оценивали с помощью парного теста t .
Мы использовали 2 высокореактивных TCR, способных распознавать антигены меланоцитов MART-1 или gp100, сверхэкспрессированные на меланомах.Чтобы преодолеть проблему толерантности к центральным делециям лимфоцитов, экспрессирующих высокоаффинные противоопухолевые TCR, мы создали высокореактивный TCR, называемый gp100(154), против эпитопа gp100:154-162 меланоцитов человека путем иммунизации трансгенных мышей HLA-A*0201 этим пептидом. которая отличается от мышиной последовательности одной аминокислотой. Кроме того, обширный скрининг более 600 клонов TIL от нескольких пациентов выявил клон лимфоцитов, называемый DMF5, с гораздо большей реактивностью, чем ранее идентифицированный DMF4, в отношении пептидного эпитопа MART-1:27-35. 15 Были выделены гены TCR из клеток мыши и из клона DMF5 человека, и эквивалентные количества РНК были созданы in vitro и перенесены как в Jurkat, так и в донорские PBL.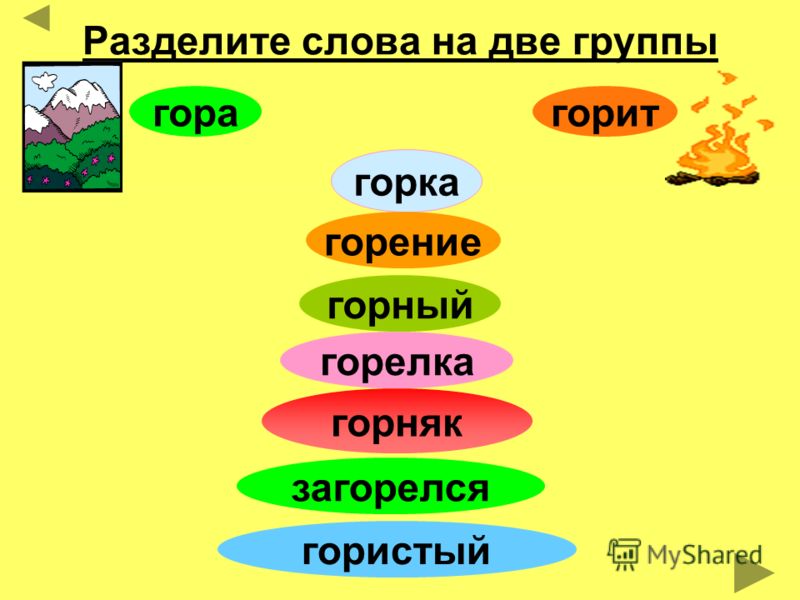 После переноса того же количества экзогенного TCR (оцененного по поверхностной экспрессии CD3 в клетках Jurkat) 15 TCR с более высокой авидностью DMF5 и gp100(154) придавали более высокую реактивность донорским PBL, чем ранее идентифицированный DMF4 при распознавании опухолевого антигена (рис. 1А). Каждый из генов, кодирующих альфа- и бета-цепи этих двух рецепторов, gp100(154) и DMF5, клонировали в бицистронные гамма-ретровирусные векторы с использованием IRES или фурин-2-А пикорнавирусоподобной последовательности расщепления, соответственно, для управления экспрессией второй ген (рис. 1В).Эти ретровирусные векторы использовали для трансдукции нормальных периферических лимфоцитов человека (рис. 2). Было продемонстрировано, что ДМФ4 слабо связывает тетрамер, недооценивая количество поверхностного TCR, 14,15 результат, также наблюдаемый здесь (рис. 2А). Используя аналогично подготовленные ретровирусные супернатанты в той же процедуре трансдукции донорских клеток, DMF4 на поверхности трансдуцированных CD8 + PBL связывал только 2% тетрамера MART-1, а трансдуцированные CD4 + клетки не связывали тетрамера.
После переноса того же количества экзогенного TCR (оцененного по поверхностной экспрессии CD3 в клетках Jurkat) 15 TCR с более высокой авидностью DMF5 и gp100(154) придавали более высокую реактивность донорским PBL, чем ранее идентифицированный DMF4 при распознавании опухолевого антигена (рис. 1А). Каждый из генов, кодирующих альфа- и бета-цепи этих двух рецепторов, gp100(154) и DMF5, клонировали в бицистронные гамма-ретровирусные векторы с использованием IRES или фурин-2-А пикорнавирусоподобной последовательности расщепления, соответственно, для управления экспрессией второй ген (рис. 1В).Эти ретровирусные векторы использовали для трансдукции нормальных периферических лимфоцитов человека (рис. 2). Было продемонстрировано, что ДМФ4 слабо связывает тетрамер, недооценивая количество поверхностного TCR, 14,15 результат, также наблюдаемый здесь (рис. 2А). Используя аналогично подготовленные ретровирусные супернатанты в той же процедуре трансдукции донорских клеток, DMF4 на поверхности трансдуцированных CD8 + PBL связывал только 2% тетрамера MART-1, а трансдуцированные CD4 + клетки не связывали тетрамера. Напротив, от 30% до 60% лимфоцитов CD4 + и CD8 + , трансдуцированных улучшенными генными конструкциями, кодирующими высокореактивный DMF5 или gp100(154) TCR, эффективно связывали тетрамер (рис. 2А). Эти новые конструкции TCR придавали высокую экспрессию TCR, которая была независимой от корецепторов PBL донора. Сравнение функциональной реактивности донорских лимфоцитов, трансдуцированных соответствующими гамма-ретровирусами, кодирующими эти DMF4, DMF5 или gp100(154) TCR, показано на рисунке 2B и C.Клетки, экспрессирующие либо высокореактивный DMF5, либо gp100(154) TCR, распознают в 100 раз более низкие концентрации пептида, продуцируют больше IFN-γ и лизируют мишени меланомы более эффективно, чем клетки, экспрессирующие DMF4.
Напротив, от 30% до 60% лимфоцитов CD4 + и CD8 + , трансдуцированных улучшенными генными конструкциями, кодирующими высокореактивный DMF5 или gp100(154) TCR, эффективно связывали тетрамер (рис. 2А). Эти новые конструкции TCR придавали высокую экспрессию TCR, которая была независимой от корецепторов PBL донора. Сравнение функциональной реактивности донорских лимфоцитов, трансдуцированных соответствующими гамма-ретровирусами, кодирующими эти DMF4, DMF5 или gp100(154) TCR, показано на рисунке 2B и C.Клетки, экспрессирующие либо высокореактивный DMF5, либо gp100(154) TCR, распознают в 100 раз более низкие концентрации пептида, продуцируют больше IFN-γ и лизируют мишени меланомы более эффективно, чем клетки, экспрессирующие DMF4.
Рисунок 1
Опухолереактивный DMF5 или gp100(154) альфа- и бета-цепь TCR РНК, электропорированная в PBL, придает высокую реактивность к опухолевым антигенам меланомы . ( A ) Десятидневные анти-CD3-стимулированные донорские PBL подвергали электропорации с транскрибированной in vitro РНК, кодирующей парные альфа- и бета-цепи TCR DMF4, DMF5 или gp100 (154) или контроль GFP.Клетки совместно культивировали в течение 18 часов с клетками Т2, обработанными пептидом, HLA-A*02 + меланомы mel624 + или mel526 + , или HLA-A*02 — меланомы mel888 — . IFN-γ в супернатанте определяли с помощью ELISA. (B) Структура γ-ретровирусных векторов DMF4 и gp100(154) на основе MSGV, включающих IRES и DMF5 с последовательностью рибосомного пропуска фурина 2A, обеспечивающей экспрессию двойного гена.
( A ) Десятидневные анти-CD3-стимулированные донорские PBL подвергали электропорации с транскрибированной in vitro РНК, кодирующей парные альфа- и бета-цепи TCR DMF4, DMF5 или gp100 (154) или контроль GFP.Клетки совместно культивировали в течение 18 часов с клетками Т2, обработанными пептидом, HLA-A*02 + меланомы mel624 + или mel526 + , или HLA-A*02 — меланомы mel888 — . IFN-γ в супернатанте определяли с помощью ELISA. (B) Структура γ-ретровирусных векторов DMF4 и gp100(154) на основе MSGV, включающих IRES и DMF5 с последовательностью рибосомного пропуска фурина 2A, обеспечивающей экспрессию двойного гена.
Рисунок 1
Опухолереактивный DMF5 или gp100(154) альфа- и бета-цепь TCR РНК, электропорированная в PBL, придает высокую реактивность к опухолевым антигенам меланомы .( A ) Десятидневные анти-CD3-стимулированные донорские PBL подвергали электропорации с транскрибированной in vitro РНК, кодирующей парные альфа- и бета-цепи TCR DMF4, DMF5 или gp100 (154) или контроль GFP. Клетки совместно культивировали в течение 18 часов с клетками Т2, обработанными пептидом, HLA-A*02 + меланомы mel624 + или mel526 + , или HLA-A*02 — меланомы mel888 — . IFN-γ в супернатанте определяли с помощью ELISA. (B) Структура γ-ретровирусных векторов DMF4 и gp100(154) на основе MSGV, включающих IRES и DMF5 с последовательностью рибосомного пропуска фурина 2A, обеспечивающей экспрессию двойного гена.
Клетки совместно культивировали в течение 18 часов с клетками Т2, обработанными пептидом, HLA-A*02 + меланомы mel624 + или mel526 + , или HLA-A*02 — меланомы mel888 — . IFN-γ в супернатанте определяли с помощью ELISA. (B) Структура γ-ретровирусных векторов DMF4 и gp100(154) на основе MSGV, включающих IRES и DMF5 с последовательностью рибосомного пропуска фурина 2A, обеспечивающей экспрессию двойного гена.
Рисунок 2
Ретровирусные конструкции DMF5 и gp100(154) TCR придавали донорским PBL большую противоопухолевую реактивность, чем исходный рецептор DMF4 . (A) Донорские PBL стимулировали анти-CD3 mAb OKT-3 и разделяли на популяции CD4 и CD8 перед ретровирусной трансдукцией конструкциями TCR DMF4, DMF5 или gp100(154). Экспрессию TCR анализировали через 7 дней с помощью окрашивания тетрамерами и проточной цитометрии. (B) Донорские PBL, трансдуцированные ретровирусными конструкциями TCR, совместно культивировали с клетками T2, обработанными пептидом MART-1:27-35 или gp100:154-162, и секрецию IFN-γ измеряли с помощью ELISA. (C) Трансдуцированные PBL совместно культивировали с меланомами mel624 + , и лизис опухоли-мишени оценивали с помощью анализа высвобождения Cr 51 . Клетки не лизировали HLA-несовместимые опухоли (данные не показаны).
(B) Донорские PBL, трансдуцированные ретровирусными конструкциями TCR, совместно культивировали с клетками T2, обработанными пептидом MART-1:27-35 или gp100:154-162, и секрецию IFN-γ измеряли с помощью ELISA. (C) Трансдуцированные PBL совместно культивировали с меланомами mel624 + , и лизис опухоли-мишени оценивали с помощью анализа высвобождения Cr 51 . Клетки не лизировали HLA-несовместимые опухоли (данные не показаны).
Рисунок 2
Ретровирусные конструкции DMF5 и gp100(154) TCR придавали донорским PBL большую противоопухолевую реактивность, чем исходный рецептор DMF4 .(A) Донорские PBL стимулировали анти-CD3 mAb OKT-3 и разделяли на популяции CD4 и CD8 перед ретровирусной трансдукцией конструкциями TCR DMF4, DMF5 или gp100(154). Экспрессию TCR анализировали через 7 дней с помощью окрашивания тетрамерами и проточной цитометрии. (B) Донорские PBL, трансдуцированные ретровирусными конструкциями TCR, совместно культивировали с клетками T2, обработанными пептидом MART-1:27-35 или gp100:154-162, и секрецию IFN-γ измеряли с помощью ELISA. (C) Трансдуцированные PBL совместно культивировали с меланомами mel624 + , и лизис опухоли-мишени оценивали с помощью анализа высвобождения Cr 51 .Клетки не лизировали HLA-несовместимые опухоли (данные не показаны).
(C) Трансдуцированные PBL совместно культивировали с меланомами mel624 + , и лизис опухоли-мишени оценивали с помощью анализа высвобождения Cr 51 .Клетки не лизировали HLA-несовместимые опухоли (данные не показаны).
Для исследования in vivo активности аутологичных клеток, трансдуцированных этими высокореактивными TCR, 36 пациентам с серьезно предлеченной прогрессирующей метастатической меланомой были введены трансдуцированные клетки (20 пациентов с ДМФ5 и 16 с gp100(154)), после лимфодепилирующего подготовительного режима для истощения эндогенные циркулирующие лимфоциты. 8 Демографические данные пациентов и сведения о лечении представлены в таблицах 1 и 2. Пациентам внутривенно вводили IL-2, начиная с 8 часов после адоптивного переноса клеток и продолжая каждые 8 часов в течение 3 дней. Все пациенты были невосприимчивы к предшествующему лечению ИЛ-2, а у 42% и 33% наблюдалось прогрессирование в результате предшествующей химиотерапии и лучевой терапии соответственно.
DMF5
| Пациент №.. | Возраст, пол . | Предварительное лечение* . | Очаги заболевания† . | Количество ячеек × 10 9 . | Процент CD4/8 . | Процент Тет . | Процент IC IFN-γ‡ . | ЭЛИСПОТ . | Дозы ИЛ-2 . | Токсичность¶ (кожа/глаза/уши) . | Опухолевой ответ, мес** . | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ил-2§ . | ИФН-γ‖ . | |||||||||||||
| 1 |  «> 36 / F «> 36 / F | R, S, LU | BO, Li, Lu | 22.2 | 17/83 | 73 | 52 | 95 | 173 | 10 | 1/2/3 | NR | ||
| 2 | 43/F | S,I | lu | 10.5 | 15/85 | 46 | 37 | 37 | 166 | 5 | 2/2/3 | PR (17+) | PR (17+) | |
| 3 | 60 / F | S, I | BO, LN, SC | 6.5 | 7/93 | 56 | 40 | 1 | 56 | 3 | 0/0/0 | NR | ||
| 4 | 38 / м | S,C,I | глаз, sc | 12.0 | 22/78 |  «> 22/78 «> 22/78 | 65 | 38 | 24 | > 112 | 8 | 2/2/1 | Nr | |
| 5 | 47 / F | R, S, I | SC, CU | 23.39999 | 37/63 | 37/63 | 61 | 21 | 91 | 53 | 5 | 1,1 † / 2,2 † Детк / 0,3 †† | PR ( 17+) | |
| 6 | 57/М | Р,С,К,И | Лн, Бо, Сбн, Лу | 17.6 | 27/73 | 27/73 | 69 | 29 | 38 | 52 | 7 | 1/0/0 | NR | |
| 7 | ||||||||||||||
| 7 | 33 / M | S, I | Ln Ли, Li | 1.5 | 11/89 |  «> 33 «> 33 | 19 | 68 | 132 | 10 | 1/0/0 | NR | ||
| 8 | 46 / млн. | с I | пер, бр, лу | 5.7 | 32/68 | 49 | 49 | 17 | 26 | 65 | 7 | 0/2/0 | PR (16+) | PR (16+) |
| 9 | 54 / м | с | LN, SP, SC | 3.8 | 28/72 | 69 | 28 | 860 | 164 | 5 | 1/0/0 | PR (9) | ||
| 10 | 35/М | Р,С,И | пер, л, сб | 2.0 | 61/39 | 48 | 7 | 1200 9 |  «> 1200 9 «> 1200 9 | 3 9 | 11 | 11 | 1/2/0 | NR |
| 11 | 60 / F | R, S, I | BR, SC, LU | 3.0 | 27/0 | 27/73 | 60 | 19 | 310 | 152 | 15 | 1/0/0 | NR | |
| 12 | 49 / F | S,I | ln, ki, li, lu, sp | 4.8 | 2/98 | 2/98 | 85 | 61 | 66 | 34 | 4 | 0/0/0 | NR | NR |
| 13 | 54 / M | R, S, I | В, SC | 38.0 | 6/94 | 91 | 70 91 | 7099 |  «>> 292 «>> 292 | > 292 | 9 | 0/2/3 | PR (4) | |
| 14 | 24 / F | Р,С,И | пер, сбн | 80.0 | 10/90 | 75 | 64 | 4767 | > 349 | > 349 | 6 | 1/1/3 | NR | |
| 15 | 56 / M | R, S, I | IP, Li, Lu | 30.5 | 13/87 | 95 | 72 | 990 92 | 990 | > 191 | 9 | 1/1/1 | NR | |
| 16 | 54 / м | S,C,I | пер, sc | 29.1 | 4/96 | 89 | 54 | 54 | > 270 |  «>> 270 «>> 270 | > 253 | 7 | 0/0/0 | NR |
| 17 | 37 / M | S, I | Adr, Ln, Lar, SC | 9.9 | 2/98 | 77 | 50 | 83 | 27 | 10 | 0/0/0 | NR | ||
| 18 | 56 / F | С,И | Лн, Сб | 48.2 | 21/79 | 92 | 92 | 68 | 1410 | > 157 | 3 | 1/2/3 | PR (3) | |
| 19 | 31 / F | R, S , C, I | BR, SC | 73.7 | 13/60 | 92 | ND | 166 | > 332 | 8 | 2/2/3 | NR | ||
| 20 |  «> 56/M «> 56/M | S,C,I | adr, ln, lu, im, panc | 107.0 | 33/66 | 33/66 | 91 | ND | 1500 | > 376 | > 376 | 11 | 1/0/3 | NR |
| Пациент №. . | Возраст, пол . | Предварительное лечение* . | Очаги заболевания† . | Количество ячеек × 10 9 . | Процент CD4/8 . | Процент Тет . | Процент IC IFN-γ‡ . | ЭЛИСПОТ . | Дозы ИЛ-2 . | Токсичность¶ (кожа/глаза/уши)
. | Опухолевой ответ, мес** . | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ил-2§ . | ИФН-γ‖ . | |||||||||||||
| 1 | 36 / F | R, S, LU | BO, Li, Lu | 22.2 | 17/83 | 73 | 52 | 95 | 173 | 10 | 1/2/3 | NR | ||
| 2 | 43/F | S,I | lu | 10.5 | 15/85 | 46 | 37 | 37 | 166 | 5 | 2/2/3 | PR (17+) | PR (17+) | |
| 3 | 60 / F | S, I | BO, LN, SC | 6.5 | 7/93 | 56 | 40 | 1 | 56 |  «> 3 «> 3 | 0/0/0 | NR | ||
| 4 | 38 / м | S,C,I | глаз, sc | 12.0 | 22/78 | 22/78 | 65 | 38 | 24 | > 112 | 8 | 2/2/1 | Nr | |
| 5 | 47 / F | R, S, I | SC, CU | 23.39999 | 37/63 | 37/63 | 61 | 21 | 91 | 53 | 5 | 1,1 † / 2,2 † Детк / 0,3 †† | PR ( 17+) | |
| 6 | 57/М | Р,С,К,И | Лн, Бо, Сбн, Лу | 17.6 | 27/73 | 27/73 | 69 | 29 | 38 | 52 | 7 |  «> 1/0/0 «> 1/0/0 | NR | |
| 7 | ||||||||||||||
| 7 | 33 / M | S, I | Ln Ли, Li | 1.5 | 11/89 | 33 | 19 | 68 | 132 | 10 | 1/0/0 | NR | ||
| 8 | 46 / млн. | с I | пер, бр, лу | 5.7 | 32/68 | 49 | 49 | 17 | 26 | 65 | 7 | 0/2/0 | PR (16+) | PR (16+) |
| 9 | 54 / м | с | LN, SP, SC | 3.8 | 28/72 | 69 | 28 | 860 | 164 | 5 | 1/0/0 | PR (9) | ||
| 10 |  «> 35/М «> 35/М | Р,С,И | пер, л, сб | 2.0 | 61/39 | 48 | 7 | 1200 9 | 1200 9 | 3 9 | 11 | 11 | 1/2/0 | NR |
| 11 | 60 / F | R, S, I | BR, SC, LU | 3.0 | 27/0 | 27/73 | 60 | 19 | 310 | 152 | 15 | 1/0/0 | NR | |
| 12 | 49 / F | S,I | ln, ki, li, lu, sp | 4.8 | 2/98 | 2/98 | 85 | 61 | 66 | 34 | 4 | 0/0/0 | NR | NR |
| 13 |  «> 54 / M «> 54 / M | R, S, I | В, SC | 38.0 | 6/94 | 91 | 70 91 | 7099 | > 292 | > 292 | 9 | 0/2/3 | PR (4) | |
| 14 | 24 / F | Р,С,И | пер, сбн | 80.0 | 10/90 | 75 | 64 | 4767 | > 349 | > 349 | 6 | 1/1/3 | NR | |
| 15 | 56 / M | R, S, I | IP, Li, Lu | 30.5 | 13/87 | 95 | 72 | 990 92 | 990 | > 191 | 9 | 1/1/1 | NR | |
| 16 | 54 / м | S,C,I | пер, sc |  «> 29.1 «> 29.1 | 4/96 | 89 | 54 | 54 | > 270 | > 270 | > 253 | 7 | 0/0/0 | NR |
| 17 | 37 / M | S, I | Adr, Ln, Lar, SC | 9.9 | 2/98 | 77 | 50 | 83 | 27 | 10 | 0/0/0 | NR | ||
| 18 | 56 / F | С,И | Лн, Сб | 48.2 | 21/79 | 92 | 92 | 68 | 1410 | > 157 | 3 | 1/2/3 | PR (3) | |
| 19 | 31 / F | R, S , C, I | BR, SC | 73.7 | 13/60 |  «> 92 «> 92 | ND | 166 | > 332 | 8 | 2/2/3 | NR | ||
| 20 | 56/M | S,C,I | adr, ln, lu, im, panc | 107.0 | 33/66 | 91 | 91 | ND | 1500 | > 376 | 11 | 1/0/3 | NR | |
gp100(154)
| Пациент №. . | Возраст, пол . | Предварительное лечение* . | Очаги заболевания† . | Количество ячеек × 10 9 . | Процент CD4/8
. | Процент Тет . | ЭЛИСПОТ . | Дозы ИЛ-2 . | Токсичность‖ (кожа/глаза/уши) . | Опухолевой ответ, мес¶ . | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ил-2‡ . | ИФН-γ§ . | ||||||||||||
| 1 | 31/F | S,C,I | In, ip, li | 3.7 | 35/65 | 6099 | 60 | 7100 | > 137 | 5 | 1/0/0 | NR | |
| 9 | |||||||||||||
| 2 | 32 / M | S, C, I | Ln, IM, SC | 10/54 | 36/54 | 78 | 5300 | 53009 | 5 962 | 5 | 2/0/0 | NR | |
| 3 |  «> 54 / M «> 54 / M | S, I | SC | 9.8 | 13/82 | 13/82 | 87 | 2590 | > 302 | 4 | 1/0/0 | NR | |
| 4 | 50 / M | S, I | пер, гб | 4.6 | 11/78 | 97 | 97 | 12 050 | > 409 | 2 909 | 2 | 1/0/0 | NR |
| 5 | 49 / F | S, I | Ln, Лу , SC | 9.9 | 21/80 | 91 | 91 | 11 683 | > 451 | 2 | 2/0/0 | NR | |
| 6 * | 36 / F | R, S , C, i | ln, bo, li, sp | 5.8 |  «> 6/89 «> 6/89 | 77 | 2370 | > 336 | 5 | 1/0/0 | NR | ||
| 7 | 60/M | S,I | In, bo, li, lu, sc | 1.8 | 40/56 | 83 | 6117 | > 213 | > 213 | 4 | 1/0/0 | NR | |
| 8 | 50 / F | S, C, I | ST Ли, Лу, INT, SC | 19.4 | 44/53 | 55 | 2900 | 294 | 5 | 2/2/0 | NR | ||
| 9 | 25 / м | с , Я | Ln, Bo, IP | 2.3 | 44/56 | 84 | ND | 46 | 9 | 0/0/0 | NR | ||
| 10 |  «> 40 / F «> 40 / F | S,C,I | ш, пер, им | 2.7 | 24/69 | 24/69 | 59 | 6700 | > 445 | 10 | 1/0/0 | NR | |
| 11 ** | 50 / м | R, S, C, I | BR, LN | 68.8 | 5/96 | 97 | 8167 | > 495 | 8 | 1/2/3 | CR (14+) | ||
| 12 | 62 / м | S, C, I | LU, LN | 902 | 20/82 | 85 | 14 583 | > 526 | 7 | 1/2/2 | NR | ||
| 13 * * | 44/F | R,S,C,I | li, ln | 54.0 | 2/97 | 9/97 | 90 |  «> 8700 «> 8700 | > 399 | 13 | 1/2/1 | NR | |
| 14 | 51 / F | S, I | Ln, Li, Лу, SP | 110.0 | 11/88 | 92 | 11 817 | > 421 | > 421 | 6 | 1/0/2 | PR (4) | |
| 15 | 33 / м | S, I | LN | 94.1 | 16/78 | 89 | 6833 | > 389 | 6 | 2/0/0 | NR | ||
| 16 | 41 / м | с ,я | пер | 39.1 | 18/79 | 9099 | 90 | 10 967 | > 485 | 7 | 1/0/1 | PR (3) | PR (3) |
Пациент №. . . | Возраст, пол . | Предварительное лечение* . | Очаги заболевания† . | Количество ячеек × 10 9 . | Процент CD4/8 . | Процент Тет . | ЭЛИСПОТ . | Дозы ИЛ-2 . | Токсичность‖ (кожа/глаза/уши) . | Опухолевой ответ, мес¶ . | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ил-2‡ . | ИФН-γ§ . | ||||||||||||
| 1 | |||||||||||||
| 1 | 31 / F | S, C, I | LN, IP, Li | 3.7 | 35/65 | 60209 | 7100 | > 137 | 5 | 1 /0/0 | NR | ||
| 2 |  «> 32/M «> 32/M | S,C,I | ln, im, sc | 10.0 | 36/54 | 36/54 | 78 | 5300 | > 362 | 5 | 2/0/0 | NR | |
| 3 | 54 / M | S, I | SC | 9.8 | 13/82 | 0/82 | 87 | 2590 | > 302 | 4 902 | 4 | 1/0/0 | NR |
| 4 | 50 / M | S, I | LN, GB | 4.6 | 11/78 | 97 | 97 | 12 050 | > 409 | 2 9099 | 1/0/0 | NR | |
| 5 | 49 / F | S, I | Ln, Lu , сбн | 9.9 |  «> 21/80 «> 21/80 | 91 | 91 | 11 683 | > 451 | 2 | 2/0/0 | NR | |
| 6 * | 36 / F | R, S, C, I | Ln, Bo, Li, SP | 5.8 | 6/89 | 77 | 2370 | > 396 | 5 | 1/0/0 | NR | ||
| 7 | 60 / M | S,I | In, bo, li, lu, sc | 1.8 | 40/56 | 83 | 6117 | > 213 | > 213 | 4 | 1/0/0 | NR | |
| 8 | 50 / F | S, C, I | ST Ли, Лу, INT, SC | 19.4 | 44/53 | 55 | 2900 | 294 |  «> 5 «> 5 | 2/2/0 | NR | ||
| 9 | 25 / м | с , Я | Ln, Bo, IP | 2.3 | 44/56 | 84 | ND | 46 | 9 | 0/0/0 | NR | ||
| 10 | 40 / F | S,C,I | ш, пер, им | 2.7 | 24/69 | 24/69 | 59 | 6700 | > 445 | 10 | 1/0/0 | NR | |
| 11 ** | 50 / м | R, S, C, I | BR, LN | 68.8 | 5/96 | 97 | 8167 | > 495 | 8 | 1/2/3 | CR (14+) | ||
| 12 | 62 / м | S, C, I | LU, LN | 902 |  «> 20/82 «> 20/82 | 85 | 14 583 | > 526 | 7 | 1/2/2 | NR | ||
| 13 * * | 44/F | R,S,C,I | li, ln | 54.0 | 2/97 | 9/97 | 90 | 8700 | > 399 | 13 | 1/2/1 | NR | |
| 14 | 51 / F | S, I | Ln, Li, Лу, SP | 110.0 | 11/88 | 92 | 11 817 | > 421 | > 421 | 6 | 1/0/2 | PR (4) | |
| 15 | 33 / м | S, I | LN | 94.1 | 16/78 | 89 | 6833 | > 389 | 6 |  «> 2/0/0 «> 2/0/0 | NR | ||
| 16 | 41 / м | с ,я | пер | 39.1 | 18/79 | 9099 | 9 | 10 967 | > 485 | 7 | 1/0/1 | PR (3) | |
Основываясь на окрашивании тетрамерами, средняя эффективность трансдукции клеток, введенных этим 36 пациентам, составила 71% и 82% для DMF5 и gp100(154) TCR соответственно (таблицы 1–2). Все обработанные клетки показали высокие уровни специфической реактивности против родственных антиген-содержащих опухолевых мишеней, что оценивалось окрашиванием внутриклеточных цитокинов и анализом ELISPOT (как IFN-γ, так и IL-2; таблицы 1–2), а также высвобождением IFN-γ (таблица 3). и лизис клеток-мишеней (данные не показаны).
Таблица 3Продукция интерферона-γ трансдуцированными TCR инфузионными клетками
Номер пациента. . . | Клеточная линия меланомы . | Клетки Т2 + мкМ пептид* . | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Нет . | А2- . | А2+ . | Отрицательный . | Положительный . | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 888 . | 938 . | 526 . | 624 . | Нет . | 1,0 . | 1,0 . | 0,1 . | 0,01 . | 0,001 . | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| | | 1 |  «> 0 «> 0 6 | 49 | 62 300 | 111 200 | 102 | 232 | 132 400 | 132 400 | 56 700 | 12 330 9009 9 | 1139 | 2 † | 7 | 133 | 175 | 16 800 | 99 400 | 159 | 141 | 117 400 | 53 700 | 6910 | 1182 | 0 | 125 | 163 | 3830 | 27 700 | 37 | 64 | 50 5009 909 | 15 000 | 984 | 241 | 4 | 9021 0 0 1 | 0 | 983 |  «> 983 «> 983 7 | 7 | 4 | 45 000 | 11 760 | 798 | 139 9999 | 5 † | 0 | 2 | 3 | 7760 | 7760 | 180 800 | 20 | 21 | 34 000 | 17 000 | 1076 | 298 | 6 | 1 | 3 | 4 | 3020 9 | 2980 | 13 | 17 | 9580 | 9580 | 4810 | 960 | 9 9 | 16 | 41 | 8420 | 39 700 9 | 102 |  «> 134 «> 134 50 400 | 32 200 | 11 950 | 1556 | 8† | 744 | 3 9020 | 10201 9021 7 50009 88 000 | 1299 | 1299 | 1254 | 113 100 | 14 510 | > 16 460 | > 3278 | 9 † | 0 | 1 | 6 | 7440 | 13 400 | 176 | 176 | 290 | 54 800 | 54 800 | > 1649 | > 1649 | 10 | 51 | 80210 51 | 155 | 102 3009 | 119 200 | 527 | 857 |  «> 93 400 «> 93 400 93 400 | 104 800 | > 5226 | 9005 | 11 | 9 | 8 | 97 | 9 | 6460 97 | 6460 | 26 700 | 221 | 321 | 42 300 | 36 200 | 12 860 | 1576 | 1576 | | |||||||||||||||
| 12 | 0 | 0 | 8 | 1300 | 90 210 19 70024 7009 | 90 | 16 150 | 4720 | 4720 | 338 | 124 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 13 † | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 13 † | 0 | 6 | 0 | 3120 | 117 9009 | 78 | 156 |  «> 137 900 «> 137 900 | 41 900 | 41 900 | 1750 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 9 | 9 | 0 | 9 | 31 | > 131 600 | 426 600 | 84 | 352 | > 283 400 | 38 800 | 7950 | 9950 | 956 | 956 | 9|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 9 | 5 | 91 | 152 | 9400 | 47 200 | 54 | 295 | 69 400 | 71 400 | > 1575 | 271 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 16 | Н/П | 40 | 15 | 5410 | 2 н/п | 17247 | 52 650 | 20 225 |  «> 6370 «> 6370 | 6370 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 17 | 0 | 9 118 | 7100 | 63 200 | 46 | 394 | 70 200 9 | 3260 | 118 | 73 | 93 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 42 | 56 | 71 800 | 328 900 | 651 9009 | 1400 | > 283 400 | 62 800 | 11 450 | 47 510 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 19 | 2 | 39 | 82 | > 142 700 | 275 590 | 275 590 | 829 | 1155 | 329 600 | > 184 200 | 7100 |  «>> 2470 «>> 2470 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 20 | н/д | 17 | 9 | 9995 | 24 900 | 8902 902 902 | 22 750 | 6710 | 6710 | 590 | 590 | GP100 (154)||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1 | 46 | 25 | 32 | 14 460 | 74 900 | 74 900 | 292 | 275 | 9540 | 9100 | 4210 | 3900 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2 | 0 | 26 | 42 | 88 300 | 199 9 | 199 400 |  «> 9 «> 9 | 37 | 2080 | 40 900 | > 19 200 | 1620 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3 | 15 | 80210 9 | 80999 | 185 | 90 500 | 209 800 | 87 | 163 | 15 490 | 14 870 | 8970 902 09 | 10 040 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4 | 0 | 9 9 | 94 | 101 | 90 300 | 97 9009 | 97 | 80210 9 9 | 3000 | 70 500 | > 19 7709999 | 829 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5 | 29 | 0 | 15 | 40 000 | 287 800 | 253 | 43 | 142 400 |  «> 114 500 «> 114 500 | 11 540 | 4540 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6 | 0 | 38 | 31 | 83 400 | 163 200 | 163 | 57 | 14 160 | 11 750 | 7160 | 6640 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 7 | 23 | 923 | 169 | 82 | 4670 | 18 350 | 23 | 212 | 10 700 | 7370 | 6060 | 9009 908 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8 | 166 | 102 | 120 | 98 600 | 179 400 | 97 400 | 87 | 777 | 59 200 | 55 600 | 16 070209 |  «> 5050 «> 5050 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 9 | 2 | 36 | 23 | 14 420 | 91 800 | 91 800 | 96 | 32 | 39 600 | 52 500 | 11 890 | 2440 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 10 | 200 | 20 | 245 | 13 640 | 148 800 | 148 800 | 21 | 29 | 64 300 | 115 900 | > 30 880 | 4140 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 11 † | 71 | 0 | 3 | 141 700 | 143 600 | 29 | 265 | 161 900 | 58 700 | 4850 | 790 |  «> 790 «> 790 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 12 | 87 | 87 | 10 | 48 | 14 720 | 8 | 8 | 210 | 115 700 | 69 500 9| 12 500 | 849 | 9 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 13 | Na | 14 | 30 | 102 600 | 172 800 | 172 800 | Na | 30 | 130 900 | 59 800 | 8295 | 651 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 14 † | Na | 314 | 151 | 62 750 | 85 400 | Na | 142 | 135 700 | 58 700 | 11 3600 9009 | 4390 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Na | 43 | 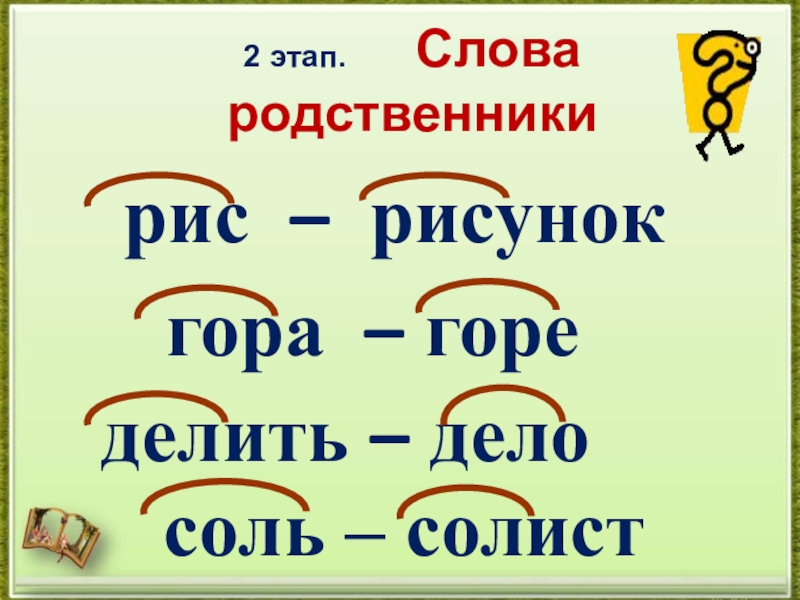 «> 0 «> 0 | 2455 | 21 300 | NA | 149 | 7420 | 3855 | 2360 | 974 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 20 16† 9 NA 90 16† 9 20 0 16† 9 09 | 86 | 113 | 23 050 | 110 350 | Na | 50 | 58 800 | 225 500 | 225 500 | 3840 | 1436 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| У пациента нет.. | Клеточная линия меланомы | . Клетки Т2 + мкМ пептид* | . Нет
. | А2- | . А2+ | . Отрицательный | .  Положительный | . 888
. | 938
. | 526
. | 624
. | Нет
. | 1,0
. | 1,0
. | 0,1
. | 0,01
. | 0,001
. | | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| | | 1 | 0 | 6 | 49 | 62 300 | 111 200 | 102 | 232 | 132 400 | 132 400 | 56 700 | 12 330 9009 9 | 1139 | 2 † | 7 |  «> 133 «> 133 175 | 16 800 | 99 400 | 159 | 141 | 117 400 | 53 700 | 6910 | 1182 | 0 | 125 | 163 | 3830 | 27 700 | 37 | 64 | 50 5009 909 | 15 000 | 984 | 241 | 4 | 9021 0 0 1 | 0 | 983 | 983 | 7 | 7 | 4 | 45 000 | 11 760 | 798 | 139 9999 | 5 † | 0 | 2 | 3 | 7760 | 7760 |  «> 180 800 «> 180 800 20 | 21 | 34 000 | 17 000 | 1076 | 298 | 6 | 1 | 3 | 4 | 3020 9 | 2980 | 13 | 17 | 9580 | 9580 | 4810 | 960 | 9 9 | 16 | 41 | 8420 | 39 700 9 | 102 | 134 | 50 400 | 32 200 | 11 950 | 1556 | 8† | 744 | 3 9020 | 10201 9021 7 50009 88 000 | 1299 | 1299 | 1254 |  «> 113 100 «> 113 100 14 510 | > 16 460 | > 3278 | 9 † | 0 | 1 | 6 | 7440 | 13 400 | 176 | 176 | 290 | 54 800 | 54 800 | > 1649 | > 1649 | 10 | 51 | 80210 51 | 155 | 102 3009 | 119 200 | 527 | 857 | 93 400 | 93 400 | 104 800 | > 5226 | 9005 | 11 | 9 | 8 | 97 | 9 | 6460 97 | 6460 | 26 700 | 221 |  «> 321 «> 321 42 300 | 36 200 | 12 860 | 1576 | 1576 | | |||||||||||||||
| 12 | 0 | 0 | 8 | 1300 | 90 210 19 70024 7009 | 90 | 16 150 | 4720 | 4720 | 338 | 124 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 13 † | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 13 † | 0 | 6 | 0 | 3120 | 117 9009 | 78 | 156 | 137 900 | 41 900 | 41 900 | 1750 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 9 | 9 | 0 | 9 | 31 | > 131 600 | 426 600 | 84 | 352 |  «>> 283 400 «>> 283 400 | 38 800 | 7950 | 9950 | 956 | 956 | 9|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 9 | 5 | 91 | 152 | 9400 | 47 200 | 54 | 295 | 69 400 | 71 400 | > 1575 | 271 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 16 | Н/П | 40 | 15 | 5410 | 2 н/п | 17247 | 52 650 | 20 225 | 6370 | 6370 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 17 | 0 | 9 118 | 7100 | 63 200 | 46 | 394 | 70 200 9 | 3260 | 118 |  «> 73 «> 73 | 93 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 42 | 56 | 71 800 | 328 900 | 651 9009 | 1400 | > 283 400 | 62 800 | 11 450 | 47 510 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 19 | 2 | 39 | 82 | > 142 700 | 275 590 | 275 590 | 829 | 1155 | 329 600 | > 184 200 | 7100 | > 2470 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 20 | н/д | 17 | 9 | 9995 | 24 900 | 8902 902 902 | 22 750 | 6710 | 6710 | 590 | 590 | GP100 (154)||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 1 |  «> 46 «> 46 | 25 | 32 | 14 460 | 74 900 | 74 900 | 292 | 275 | 9540 | 9100 | 4210 | 3900 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2 | 0 | 26 | 42 | 88 300 | 199 9 | 199 400 | 9 | 37 | 2080 | 40 900 | > 19 200 | 1620 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3 | 15 | 80210 9 | 80999 | 185 | 90 500 | 209 800 | 87 | 163 | 15 490 | 14 870 | 8970 902 09 | 10 040 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4 | 0 |  «> 9 9 «> 9 9 | 94 | 101 | 90 300 | 97 9009 | 97 | 80210 9 9 | 3000 | 70 500 | > 19 7709999 | 829 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5 | 29 | 0 | 15 | 40 000 | 287 800 | 253 | 43 | 142 400 | 114 500 | 11 540 | 4540 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 6 | 0 | 38 | 31 | 83 400 | 163 200 | 163 | 57 | 14 160 | 11 750 | 7160 | 6640 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 7 | 23 | 923 | 169 | 82 | 4670 |  «> 18 350 «> 18 350 | 23 | 212 | 10 700 | 7370 | 6060 | 9009 908 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 8 | 166 | 102 | 120 | 98 600 | 179 400 | 97 400 | 87 | 777 | 59 200 | 55 600 | 16 070209 | 5050 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 9 | 2 | 36 | 23 | 14 420 | 91 800 | 91 800 | 96 | 32 | 39 600 | 52 500 | 11 890 | 2440 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 10 | 200 | 20 | 245 | 13 640 | 148 800 | 148 800 |  «> 21 «> 21 | 29 | 64 300 | 115 900 | > 30 880 | 4140 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 11 † | 71 | 0 | 3 | 141 700 | 143 600 | 29 | 265 | 161 900 | 58 700 | 4850 | 790 | 790 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 12 | 87 | 87 | 10 | 48 | 14 720 | 8 | 8 | 210 | 115 700 | 69 500 9| 12 500 | 849 | 9 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 13 | Na | 14 | 30 | 102 600 | 172 800 | 172 800 | Na |  «> 30 «> 30 | 130 900 | 59 800 | 8295 | 651 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 14 † | Na | 314 | 151 | 62 750 | 85 400 | Na | 142 | 135 700 | 58 700 | 11 3600 9009 | 4390 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Na | 43 | 0 | 2455 | 21 300 | NA | 149 | 7420 | 3855 | 2360 | 974 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 20 16† 9 NA 90 16† 9 20 0 16† 9 09 | 86 | 113 | 23 050 | 23 050 | 110 350 | Na | 50 | 58 800 | 225 500 | 3840 | 1436 |  «> «> | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
В предыдущих исследованиях пациентов, получавших немодифицированные TIL, персистенция перенесенных клеток in vivo сильно коррелировала с противоопухолевым ответом. 18 Таким образом, мы оценили персистенцию TCR-трансдуцированных клеток DMF5 человека и мышиного gp100(154) с использованием связывания тетрамера и анализов ELISPOT. У всех пациентов были измеримые уровни (≥ 1%) тетрамер-позитивных Т-клеток в их кровообращении через 1 месяц после лечения (рис. 3A-B). Не было никакой разницы в персистентности клеток, несущих человеческий TCR DMF5 (22% ± 6%) или мышиный gp100(154) TCR (22% ± 5%; P = 0,4; фигура 3B). Чтобы измерить функциональное распознавание опухолевых клеток, PBL после лечения совместно культивировали с меланомами, совпадающими или несовместимыми с основным комплексом гистосовместимости, а реактивность клеток оценивали с использованием анализов ELISPOT и внутриклеточного флуоресцентно-активированного клеточного сортера (FACS) (рис. 3).Анализ IFN-γ ELISPOT показал персистенцию (> 20 специфических пятен/100 000 клеток) опухоле-реактивных трансдуцированных клеток у 11 из 20 пациентов с DMF5 и у 7 из 16 пациентов с gp100 (154) (рис. 3B). Точно так же 11 и 7 пациентов с DMF5 и gp100 (154), соответственно, продемонстрировали персистенцию активных клеток с помощью анализов IL-2 ELISPOT PBL через один месяц после лечения (рис. 3B). В тот же самый 1-месячный период времени специфическое внутриклеточное окрашивание IFN-γ наблюдалось у 12 из 20 пациентов с ДМФ5 (рис. 3А). Таким образом, как было определено в нескольких анализах, мышиные и человеческие клетки с модифицированным геном TCR сохранялись в кровотоке через 1 месяц у большинства пациентов.При сравнении пациентов, ответивших на лечение, и пациентов, не ответивших на лечение, все пациенты, ответившие на лечение, имели высокоперсистентные опухоле-реактивные клетки, превышающие или равные 10% тетрамер-позитивных Т-клеток в крови через 1 месяц после лечения. Однако у некоторых неответивших пациентов также были высокие уровни активных персистентных клеток, что позволяет предположить, что персистенция может быть необходимой, но недостаточной, чтобы вызвать регрессию опухоли у пациентов.
3).Анализ IFN-γ ELISPOT показал персистенцию (> 20 специфических пятен/100 000 клеток) опухоле-реактивных трансдуцированных клеток у 11 из 20 пациентов с DMF5 и у 7 из 16 пациентов с gp100 (154) (рис. 3B). Точно так же 11 и 7 пациентов с DMF5 и gp100 (154), соответственно, продемонстрировали персистенцию активных клеток с помощью анализов IL-2 ELISPOT PBL через один месяц после лечения (рис. 3B). В тот же самый 1-месячный период времени специфическое внутриклеточное окрашивание IFN-γ наблюдалось у 12 из 20 пациентов с ДМФ5 (рис. 3А). Таким образом, как было определено в нескольких анализах, мышиные и человеческие клетки с модифицированным геном TCR сохранялись в кровотоке через 1 месяц у большинства пациентов.При сравнении пациентов, ответивших на лечение, и пациентов, не ответивших на лечение, все пациенты, ответившие на лечение, имели высокоперсистентные опухоле-реактивные клетки, превышающие или равные 10% тетрамер-позитивных Т-клеток в крови через 1 месяц после лечения. Однако у некоторых неответивших пациентов также были высокие уровни активных персистентных клеток, что позволяет предположить, что персистенция может быть необходимой, но недостаточной, чтобы вызвать регрессию опухоли у пациентов.
Рисунок 3
Трансдуцированные TCR клетки от отвечающих на лечение пациентов сохранялись и проявляли противоопухолевую активность ex vivo .Образцы крови были взяты из клеток пациентов до и после инфузии TCR-трансдуцированных клеток. PBMC оценивали на персистенцию инфузированных клеток в периферической крови после обработки с помощью специфического окрашивания тетрамером, а также использовали непосредственно в анализах совместного культивирования с опухолями mel624 (MART1 + , gp100 + , HLA-A2 + ). Противоопухолевую активность оценивали с помощью IFN-γ и IL-2 ELISPOT, а также путем внутриклеточного окрашивания продукции IFN-γ. (A) Стойкость и активность клеток пациентов, получающих лечение DMF5, до, через 2 недели после и через 1 месяц после инфузии.Отвечающие (PR) и неотвечающие (NR) пациенты представлены сплошными и пунктирными линиями соответственно. (B) Сравнение PBMC от пациентов, получавших либо DMF5, либо gp100 (154) TCR-трансдуцированные клетки. Окрашивание тетрамером и анализ ELISPOT продукции IFN-γ и IL-2 у неответивших (NR) и ответивших (PR) пациентов через 1 месяц после лечения. Во всех образцах было менее 10 ELISPOT (на 100 000 РВМС) и менее 1% IFN-γ-положительных клеток соответственно против опухоли HLA-A * 02 — mel888 (данные не показаны).Пациенты с объективным клиническим ответом (PR) имели большее количество противоопухолевых клеток, секретирующих IFN-γ ( P = 0,02) и IL-2 ( P = 0,02), чем нереспондеры (NR).
(B) Сравнение PBMC от пациентов, получавших либо DMF5, либо gp100 (154) TCR-трансдуцированные клетки. Окрашивание тетрамером и анализ ELISPOT продукции IFN-γ и IL-2 у неответивших (NR) и ответивших (PR) пациентов через 1 месяц после лечения. Во всех образцах было менее 10 ELISPOT (на 100 000 РВМС) и менее 1% IFN-γ-положительных клеток соответственно против опухоли HLA-A * 02 — mel888 (данные не показаны).Пациенты с объективным клиническим ответом (PR) имели большее количество противоопухолевых клеток, секретирующих IFN-γ ( P = 0,02) и IL-2 ( P = 0,02), чем нереспондеры (NR).
Рисунок 3
TCR-трансдуцированные клетки от отвечающих на лечение пациентов сохранялись и проявляли противоопухолевую активность ex vivo . Образцы крови были взяты из клеток пациентов до и после инфузии TCR-трансдуцированных клеток. PBMC оценивали на персистенцию инфузированных клеток в периферической крови после обработки с помощью специфического окрашивания тетрамером, а также использовали непосредственно в анализах совместного культивирования с опухолями mel624 (MART1 + , gp100 + , HLA-A2 + ). Противоопухолевую активность оценивали с помощью IFN-γ и IL-2 ELISPOT, а также путем внутриклеточного окрашивания продукции IFN-γ. (A) Стойкость и активность клеток пациентов, получающих лечение DMF5, до, через 2 недели после и через 1 месяц после инфузии. Отвечающие (PR) и неотвечающие (NR) пациенты представлены сплошными и пунктирными линиями соответственно. (B) Сравнение PBMC от пациентов, получавших либо DMF5, либо gp100 (154) TCR-трансдуцированные клетки. Окрашивание тетрамером и анализ ELISPOT продукции IFN-γ и IL-2 у неответивших (NR) и ответивших (PR) пациентов через 1 месяц после лечения.Во всех образцах было менее 10 ELISPOT (на 100 000 РВМС) и менее 1% IFN-γ-положительных клеток соответственно против опухоли HLA-A * 02 — mel888 (данные не показаны). Пациенты с объективным клиническим ответом (PR) имели большее количество противоопухолевых клеток, секретирующих IFN-γ ( P = 0,02) и IL-2 ( P = 0,02), чем нереспондеры (NR).
Противоопухолевую активность оценивали с помощью IFN-γ и IL-2 ELISPOT, а также путем внутриклеточного окрашивания продукции IFN-γ. (A) Стойкость и активность клеток пациентов, получающих лечение DMF5, до, через 2 недели после и через 1 месяц после инфузии. Отвечающие (PR) и неотвечающие (NR) пациенты представлены сплошными и пунктирными линиями соответственно. (B) Сравнение PBMC от пациентов, получавших либо DMF5, либо gp100 (154) TCR-трансдуцированные клетки. Окрашивание тетрамером и анализ ELISPOT продукции IFN-γ и IL-2 у неответивших (NR) и ответивших (PR) пациентов через 1 месяц после лечения.Во всех образцах было менее 10 ELISPOT (на 100 000 РВМС) и менее 1% IFN-γ-положительных клеток соответственно против опухоли HLA-A * 02 — mel888 (данные не показаны). Пациенты с объективным клиническим ответом (PR) имели большее количество противоопухолевых клеток, секретирующих IFN-γ ( P = 0,02) и IL-2 ( P = 0,02), чем нереспондеры (NR).
Мы оценили фенотип клеток пациентов, получавших DMF5 MART-1-реактивный TCR, до, во время и после лечения с помощью проточной цитометрии для костимулирующих молекул CD27 и CD28 и маркеров клеточной дифференцировки CD45RA и CD45RO.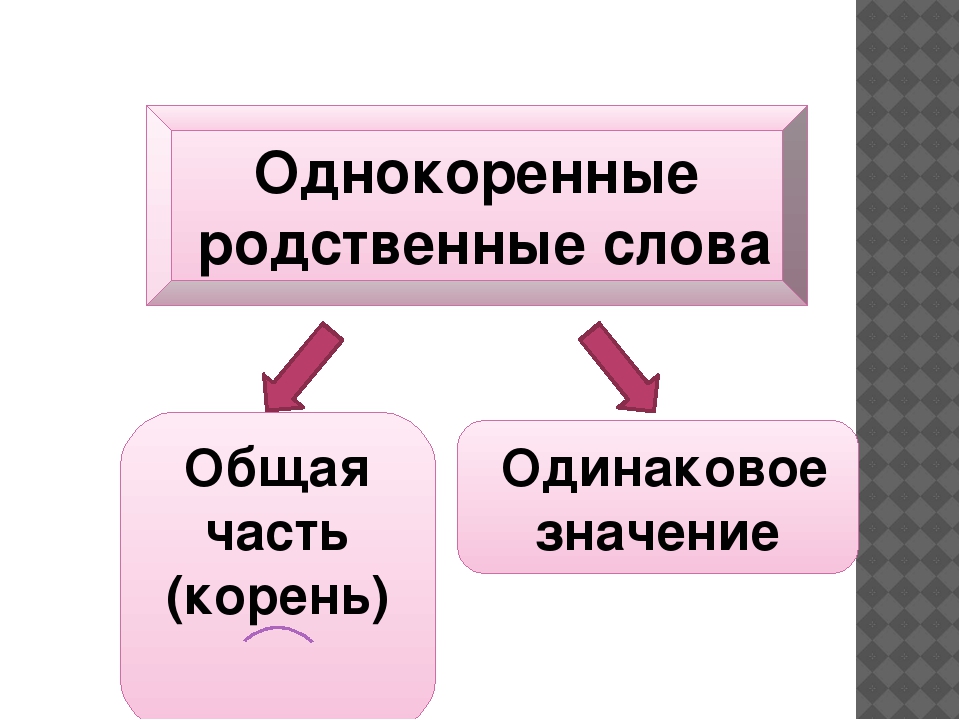 До модификации гена TCR в среднем (± стандартная ошибка среднего) периферические Т-лимфоциты пациентов состояли из 53 % плюс или минус 5 % CD27 + , 62 % плюс или минус 4 % CD28 + , 42 % плюс или минус 3 %. CD45RA + и 53% плюс или минус 3% клеток CD45RO + (фиг. 4). После экспансии ex vivo тетрамер-положительные инфузионные клетки демонстрировали меньшее количество CD27 (36% ± 6%), аналогичное количество CD28 (59% ± 6%), почти полную потерю CD45RA (3,5% ± 1,2%) и увеличение CD45RO. до 94% плюс-минус 2% (рис. 4).Оценка тетрамер-позитивных клеток в крови через 1 месяц после лечения показала уровни CD27, аналогичные инфузии (30% ± 5%), сниженные уровни CD28 (36% ± 5%) и, что интересно, уровни CD45RA увеличились, а CD45RO снизились. возвращаясь к уровням, аналогичным тем, которые наблюдались в образцах PBL до лечения (27% ± 4% CD45RA + , 66% ± 4% CD45RO + ; фигура 4). Это предполагает, что либо несколько тетрамер-позитивных CD45RA + клеток, присутствующих при инфузии, сохранялись и размножались, либо, в соответствии с ранее опубликованным клиническим отчетом о пациенте, получавшем аллогенный перенос клеток с маркировкой гена для лечения лейкемии, 19 клеток которые были CD45RA — и CD45RO + , превратились в CD45RA + и CD45RO — in vivo.
До модификации гена TCR в среднем (± стандартная ошибка среднего) периферические Т-лимфоциты пациентов состояли из 53 % плюс или минус 5 % CD27 + , 62 % плюс или минус 4 % CD28 + , 42 % плюс или минус 3 %. CD45RA + и 53% плюс или минус 3% клеток CD45RO + (фиг. 4). После экспансии ex vivo тетрамер-положительные инфузионные клетки демонстрировали меньшее количество CD27 (36% ± 6%), аналогичное количество CD28 (59% ± 6%), почти полную потерю CD45RA (3,5% ± 1,2%) и увеличение CD45RO. до 94% плюс-минус 2% (рис. 4).Оценка тетрамер-позитивных клеток в крови через 1 месяц после лечения показала уровни CD27, аналогичные инфузии (30% ± 5%), сниженные уровни CD28 (36% ± 5%) и, что интересно, уровни CD45RA увеличились, а CD45RO снизились. возвращаясь к уровням, аналогичным тем, которые наблюдались в образцах PBL до лечения (27% ± 4% CD45RA + , 66% ± 4% CD45RO + ; фигура 4). Это предполагает, что либо несколько тетрамер-позитивных CD45RA + клеток, присутствующих при инфузии, сохранялись и размножались, либо, в соответствии с ранее опубликованным клиническим отчетом о пациенте, получавшем аллогенный перенос клеток с маркировкой гена для лечения лейкемии, 19 клеток которые были CD45RA — и CD45RO + , превратились в CD45RA + и CD45RO — in vivo. Существенных различий в клеточном фенотипе между ответившими и не ответившими пациентами не было.
Существенных различий в клеточном фенотипе между ответившими и не ответившими пациентами не было.
Рисунок 4
Фенотип клеток для лечения пациентов до и после инфузии . PBMC пациентов с DMF5 окрашивали тетрамерами для TCR-узнающих MART1: 27-35 и mAb для CD3 и маркеров активации и дифференцировки CD27, CD28, CD45RA и CD45RO. Клеточный фенотип оценивали с помощью проточной цитометрии, гейтировали клетки CD3 + до лечения и образцы CD3 + тетрамера + для инфузии (Rx) и через 1 месяц после обработки (1 мес).Столбики погрешностей указывают среднее значение ± стандартная ошибка среднего. Отвечающие пациенты (PR) представлены сплошными символами, а неответившие пациенты (NR) — незаштрихованными символами.
Рисунок 4
Фенотип клеток пациента до и после инфузии . PBMC пациентов с DMF5 окрашивали тетрамерами для TCR-узнающих MART1: 27-35 и mAb для CD3 и маркеров активации и дифференцировки CD27, CD28, CD45RA и CD45RO. Клеточный фенотип оценивали с помощью проточной цитометрии, гейтировали клетки CD3 + до лечения и образцы CD3 + тетрамера + для инфузии (Rx) и через 1 месяц после обработки (1 мес).Столбики погрешностей указывают среднее значение ± стандартная ошибка среднего. Отвечающие пациенты (PR) представлены сплошными символами, а неответившие пациенты (NR) — незаштрихованными символами.
Клеточный фенотип оценивали с помощью проточной цитометрии, гейтировали клетки CD3 + до лечения и образцы CD3 + тетрамера + для инфузии (Rx) и через 1 месяц после обработки (1 мес).Столбики погрешностей указывают среднее значение ± стандартная ошибка среднего. Отвечающие пациенты (PR) представлены сплошными символами, а неответившие пациенты (NR) — незаштрихованными символами.
Клиническое течение пациентов, получавших DMF5 с высокой опухолереактивностью и gp100(154) TCR-трансдуцированные клетки, сильно отличалось от такового в нашем предыдущем исследовании с рецептором DMF4. 14 В ходе текущего исследования в сыворотке пациентов были обнаружены повышенные уровни IFN-γ, достигшие максимума примерно через 3–5 дней после лечения (рис. 5А).Средние пиковые значения IFN-γ в сыворотке пациентов, получавших улучшенные генные конструкции DMF5 и gp100(154), составляли 128 пг/мл и 210 пг/мл соответственно по сравнению с 14 пг/мл у пациентов, ранее получавших DMF4 ( P = 0,03; рис. 5В). Поскольку IFN-γ является эффекторным цитокином, продуцируемым активированными Т-лимфоцитами, а эндогенные лимфоциты пациентов оставались истощенными в течение первой недели после препаративной лимфодеплеции, предполагается, что перенесенные клетки были источником продукции этого цитокина.
5В). Поскольку IFN-γ является эффекторным цитокином, продуцируемым активированными Т-лимфоцитами, а эндогенные лимфоциты пациентов оставались истощенными в течение первой недели после препаративной лимфодеплеции, предполагается, что перенесенные клетки были источником продукции этого цитокина.
Рисунок 5
Пациенты, получавшие высокореактивный ДМФ5 или gp100(154), но не ДМФ4 TCR, имели повышенные уровни IFN-γ в сыворотке после лечения . Уровни IFN-γ в сыворотке пациентов измеряли ежедневно до, во время и после инфузии клеток с помощью ELISA. (A) Уровни IFN-γ в сыворотке увеличились от исходного уровня до пика через 3-6 дней после инфузии клеток (показаны 3 репрезентативных пациента с DMF5). (B) Пиковые уровни IFN-γ в сыворотке после лечения пациентов, получавших TCR-трансдуцированные клетки DMF5, gp100(154) и DMF4.Только у пациентов, получавших высокореактивный DMF5 или gp100(154) TCR, наблюдалось повышение IFN-γ в сыворотке ( P = 0,001 и P < 0,001 соответственно, по сравнению с P = 0,09 для DMF4 TCR). .
.
Рис. 5
Пациенты, получавшие высокореактивный ДМФ5 или gp100(154), но не ДМФ4 TCR, имели повышенные уровни IFN-γ в сыворотке после лечения . Уровни IFN-γ в сыворотке пациентов измеряли ежедневно до, во время и после инфузии клеток с помощью ELISA.(A) Уровни IFN-γ в сыворотке увеличились от исходного уровня до пика через 3-6 дней после инфузии клеток (показаны 3 репрезентативных пациента с DMF5). (B) Пиковые уровни IFN-γ в сыворотке после лечения пациентов, получавших TCR-трансдуцированные клетки DMF5, gp100(154) и DMF4. Только у пациентов, получавших высокореактивный DMF5 или gp100(154) TCR, наблюдалось повышение IFN-γ в сыворотке ( P = 0,001 и P < 0,001 соответственно, по сравнению с P = 0,09 для DMF4 TCR). .
Во время этого всплеска цитокинов у 29 из 36 пациентов в текущем исследовании была обнаружена широко распространенная эритематозная кожная сыпь, которая при биопсии показала выраженный эпидермальный спонгиоз и некротические эпидермальные кератиноциты с плотным инфильтратом CD3 + Т-лимфоцитов (преимущественно CD8 + ; фигура 6А). Неожиданно было обнаружено разрушение эпидермальных меланоцитов во всех биопсиях, выполненных у 14 пациентов с ДМФ5 и 13 пациентов с gp100(154), начиная уже с 5-го дня после лечения (рис. 6А), хотя вокруг волосяных фолликулов иногда наблюдались редкие оставшиеся меланоциты. Эта потеря меланоцитов совпала с дермальной и эпидермальной инфильтрацией лимфоцитов, а также с признаками витилиго у пациентов при последующем наблюдении. Эта сыпь постепенно исчезала в течение нескольких дней без лечения у всех пациентов.Чтобы выяснить факторы, вызывающие сыпь и инфильтрацию лимфоцитов в кожу, мы исследовали 2 пациентов с ранее существовавшим пятнистым витилиго. У этих 2 пациентов сыпь появилась только на пигментированной, а не на витилигинозной коже (рис. 6В). Биопсия после лечения выявила диффузные инфильтраты клеток CD8 + в участках пигментированной кожи с небольшим или отсутствующим лимфоцитарным инфильтратом в витилигинозной коже (рис. 6В). Эти результаты, наряду с доказательствами разрушения меланоцитов в то время, когда эндогенные лимфоциты были истощены, убедительно свидетельствуют о том, что эпидермальные меланоциты были мишенями иммунной атаки со стороны генно-инженерных клеток.
Неожиданно было обнаружено разрушение эпидермальных меланоцитов во всех биопсиях, выполненных у 14 пациентов с ДМФ5 и 13 пациентов с gp100(154), начиная уже с 5-го дня после лечения (рис. 6А), хотя вокруг волосяных фолликулов иногда наблюдались редкие оставшиеся меланоциты. Эта потеря меланоцитов совпала с дермальной и эпидермальной инфильтрацией лимфоцитов, а также с признаками витилиго у пациентов при последующем наблюдении. Эта сыпь постепенно исчезала в течение нескольких дней без лечения у всех пациентов.Чтобы выяснить факторы, вызывающие сыпь и инфильтрацию лимфоцитов в кожу, мы исследовали 2 пациентов с ранее существовавшим пятнистым витилиго. У этих 2 пациентов сыпь появилась только на пигментированной, а не на витилигинозной коже (рис. 6В). Биопсия после лечения выявила диффузные инфильтраты клеток CD8 + в участках пигментированной кожи с небольшим или отсутствующим лимфоцитарным инфильтратом в витилигинозной коже (рис. 6В). Эти результаты, наряду с доказательствами разрушения меланоцитов в то время, когда эндогенные лимфоциты были истощены, убедительно свидетельствуют о том, что эпидермальные меланоциты были мишенями иммунной атаки со стороны генно-инженерных клеток.
Рисунок 6
Транспортировка тканей и разрушение клеток-мишеней у пациентов после инфузии трансдуцированных TCR клеток . (A) Биопсия воспаленной кожи пациента 2 с ДМФ5, 5-й день после лечения, демонстрирующая спонгиотические везикулы и некротические/дискератотические кератиноциты (стрелки), окрашенные гематоксилином и эозином и иммуногистохимически окрашенные на клетки CD8 + или антитела к мелан-А распознающий антиген MART-1.(Внизу слева) Окрашивание положительного контроля на антитело против MART-1 Melan-A, показывающее нормальные эпидермальные меланоциты и подкожные отложения меланомы (исходное увеличение ×20). (B) Через неделю после лечения, биопсия gp100(154) пациента с ранее существовавшим пятнистым витилиго, демонстрирующая клеточный инфильтрат CD8 + в эпидермисе пигментированной, но не витилигинозной кожи (исходное увеличение ×20). (C) Офтальмологическое обследование 5-го глаза пациента с ДМФ5 с помощью щелевой лампы, через 2 недели после лечения, демонстрирующее мутный клеточный инфильтрат передней камеры (вверху) и (внизу) с индуцированным расширением радужной оболочки через 6 месяцев после лечения TCR и введения стероидных глазных капель, демонстрирующее задние синехии (бессимптомный). (D) Клетки, присутствующие в глазной жидкости на панели C (вверху), проанализированы непосредственно с помощью окрашивания и проточной цитометрии. (E) Аудиологическое обследование пациента 2 с DMF5 до лечения TCR, через 13 дней после лечения, показывающее потерю слуха, и через 11 месяцев после лечения, показывающее восстановление слуха после внутритимпанального лечения стероидами.
(D) Клетки, присутствующие в глазной жидкости на панели C (вверху), проанализированы непосредственно с помощью окрашивания и проточной цитометрии. (E) Аудиологическое обследование пациента 2 с DMF5 до лечения TCR, через 13 дней после лечения, показывающее потерю слуха, и через 11 месяцев после лечения, показывающее восстановление слуха после внутритимпанального лечения стероидами.
Рисунок 6
Транспортировка тканей и разрушение клеток-мишеней у пациентов после инфузии трансдуцированных TCR клеток . (A) Биопсия воспаленной кожи пациента 2 с ДМФ5, 5-й день после лечения, демонстрирующая спонгиотические везикулы и некротические/дискератотические кератиноциты (стрелки), окрашенные гематоксилином и эозином и иммуногистохимически окрашенные на клетки CD8 + или антитела к мелан-А распознающий антиген MART-1.(Внизу слева) Окрашивание положительного контроля на антитело против MART-1 Melan-A, показывающее нормальные эпидермальные меланоциты и подкожные отложения меланомы (исходное увеличение ×20). (B) Через неделю после лечения, биопсия gp100(154) пациента с ранее существовавшим пятнистым витилиго, демонстрирующая клеточный инфильтрат CD8 + в эпидермисе пигментированной, но не витилигинозной кожи (исходное увеличение ×20). (C) Офтальмологическое обследование 5-го глаза пациента с ДМФ5 с помощью щелевой лампы, через 2 недели после лечения, демонстрирующее мутный клеточный инфильтрат передней камеры (вверху) и (внизу) с индуцированным расширением радужной оболочки через 6 месяцев после лечения TCR и введения стероидных глазных капель, демонстрирующее задние синехии (бессимптомный).(D) Клетки, присутствующие в глазной жидкости на панели C (вверху), проанализированы непосредственно с помощью окрашивания и проточной цитометрии. (E) Аудиологическое обследование пациента 2 с DMF5 до лечения TCR, через 13 дней после лечения, показывающее потерю слуха, и через 11 месяцев после лечения, показывающее восстановление слуха после внутритимпанального лечения стероидами.
(B) Через неделю после лечения, биопсия gp100(154) пациента с ранее существовавшим пятнистым витилиго, демонстрирующая клеточный инфильтрат CD8 + в эпидермисе пигментированной, но не витилигинозной кожи (исходное увеличение ×20). (C) Офтальмологическое обследование 5-го глаза пациента с ДМФ5 с помощью щелевой лампы, через 2 недели после лечения, демонстрирующее мутный клеточный инфильтрат передней камеры (вверху) и (внизу) с индуцированным расширением радужной оболочки через 6 месяцев после лечения TCR и введения стероидных глазных капель, демонстрирующее задние синехии (бессимптомный).(D) Клетки, присутствующие в глазной жидкости на панели C (вверху), проанализированы непосредственно с помощью окрашивания и проточной цитометрии. (E) Аудиологическое обследование пациента 2 с DMF5 до лечения TCR, через 13 дней после лечения, показывающее потерю слуха, и через 11 месяцев после лечения, показывающее восстановление слуха после внутритимпанального лечения стероидами.
Поскольку в глазах присутствуют меланоцитарные клетки, экспрессирующие MART-1 и gp100, все пациенты прошли офтальмологическое обследование до и после лечения. Ни у одного из пациентов в предыдущем испытании DMF4 TCR не развился увеит (клеточный инфильтрат в глазу). Напротив, у 11 из 20 пациентов (55%), получавших более реактивные клетки TCR DMF5, и у 4 из 16 (25%), получавших клетки TCR gp100(154), развился передний увеит, 2 из которых были бессимптомными, а 13 потребовали временного введения. стероидных глазных капель (таблицы 1, 2; рис. 6С). У двух пациентов развились бессимптомные синехии радужной оболочки (рис. 6С). У всех пациентов офтальмологические данные вернулись к норме.Взятие проб жидкости передней камеры глаза у одного пациента с увеитом, который получил клетки, трансдуцированные DMF5, выявило преобладание MART-1-тетрамер-позитивных CD3 + Т-клеток с помощью анализа FACS (рис. 6D), что указывает на трафик генно-модифицированных клеток. в глаз.
Ни у одного из пациентов в предыдущем испытании DMF4 TCR не развился увеит (клеточный инфильтрат в глазу). Напротив, у 11 из 20 пациентов (55%), получавших более реактивные клетки TCR DMF5, и у 4 из 16 (25%), получавших клетки TCR gp100(154), развился передний увеит, 2 из которых были бессимптомными, а 13 потребовали временного введения. стероидных глазных капель (таблицы 1, 2; рис. 6С). У двух пациентов развились бессимптомные синехии радужной оболочки (рис. 6С). У всех пациентов офтальмологические данные вернулись к норме.Взятие проб жидкости передней камеры глаза у одного пациента с увеитом, который получил клетки, трансдуцированные DMF5, выявило преобладание MART-1-тетрамер-позитивных CD3 + Т-клеток с помощью анализа FACS (рис. 6D), что указывает на трафик генно-модифицированных клеток. в глаз.
Меланоцитарные клетки также присутствуют в сосудистых полосках внутреннего уха. Хотя ни у одного из пациентов в предыдущем испытании DMF4 не было потери слуха, аудиометрические исследования выявили признаки потери слуха у 10 из 20 пациентов с DMF5, начавшихся примерно через 1 неделю после лечения (таблица 1; рисунок 6E), 7 из которых получали интратимпанальные инъекции стероидов. .У всех пациентов наступило улучшение, а потеря слуха 3-й степени разрешилась или улучшилась до 1-й или 2-й степени, за исключением 2 пациентов, которые умерли от прогрессирующей метастатической меланомы до повторного тестирования. У пяти из 16 пациентов с gp100(154) развилась легкая потеря слуха (таблица 2), и только одному из них потребовалось лечение, и у него сохраняются легкие остаточные изменения. Девять из 36 пациентов испытали головокружение, связанное с внутренним ухом, которое ответило на лечение. Эта совокупность симптомов аналогична симптомам, наблюдаемым у пациентов с болезнью Фогта-Коянаги-Харада, 20 , которая, как считается, возникает в результате аутоиммунного разрушения меланоцитов, расположенных в глазу, внутреннем ухе, коже и волосах.Никаких побочных аутоиммунных эффектов не наблюдалось ни у одного пациента.
.У всех пациентов наступило улучшение, а потеря слуха 3-й степени разрешилась или улучшилась до 1-й или 2-й степени, за исключением 2 пациентов, которые умерли от прогрессирующей метастатической меланомы до повторного тестирования. У пяти из 16 пациентов с gp100(154) развилась легкая потеря слуха (таблица 2), и только одному из них потребовалось лечение, и у него сохраняются легкие остаточные изменения. Девять из 36 пациентов испытали головокружение, связанное с внутренним ухом, которое ответило на лечение. Эта совокупность симптомов аналогична симптомам, наблюдаемым у пациентов с болезнью Фогта-Коянаги-Харада, 20 , которая, как считается, возникает в результате аутоиммунного разрушения меланоцитов, расположенных в глазу, внутреннем ухе, коже и волосах.Никаких побочных аутоиммунных эффектов не наблюдалось ни у одного пациента.
TCR DMF4 и DMF5 распознают один и тот же эпитоп MART-1:27-35 и были получены от одного и того же пациента с меланомой. 15 Появляется улучшенная способность трансдуцированных клеток, экспрессирующих TCR DMF5 или gp100(154), распознавать антигены-мишени в коже, глазах и ухе и сохраняться у пациентов на уровне до 80% циркулирующих периферических Т-лимфоцитов в течение нескольких месяцев результат более высокой реактивности, обеспечиваемой этими улучшенными конструкциями TCR, по сравнению с предшествующим TCR DMF4.
15 Появляется улучшенная способность трансдуцированных клеток, экспрессирующих TCR DMF5 или gp100(154), распознавать антигены-мишени в коже, глазах и ухе и сохраняться у пациентов на уровне до 80% циркулирующих периферических Т-лимфоцитов в течение нескольких месяцев результат более высокой реактивности, обеспечиваемой этими улучшенными конструкциями TCR, по сравнению с предшествующим TCR DMF4.
У одного пациента, получившего клетки, трансдуцированные DMF5, была проведена серия последовательных биопсий подкожных метастатических поражений до и после лечения. Начиная примерно через 5 дней после лечения, биопсии выявили нарастающую инфильтрацию CD8 + Т-лимфоцитов по всей опухоли, что совпало с прогрессирующим некрозом и частичной регрессией его опухолей (рис. 7А-В).Рост лимфоцитов из резецированных поражений на 9, 15 и 26 дни выявил более 97% MART-1-тетрамер-позитивных лимфоцитов (рис.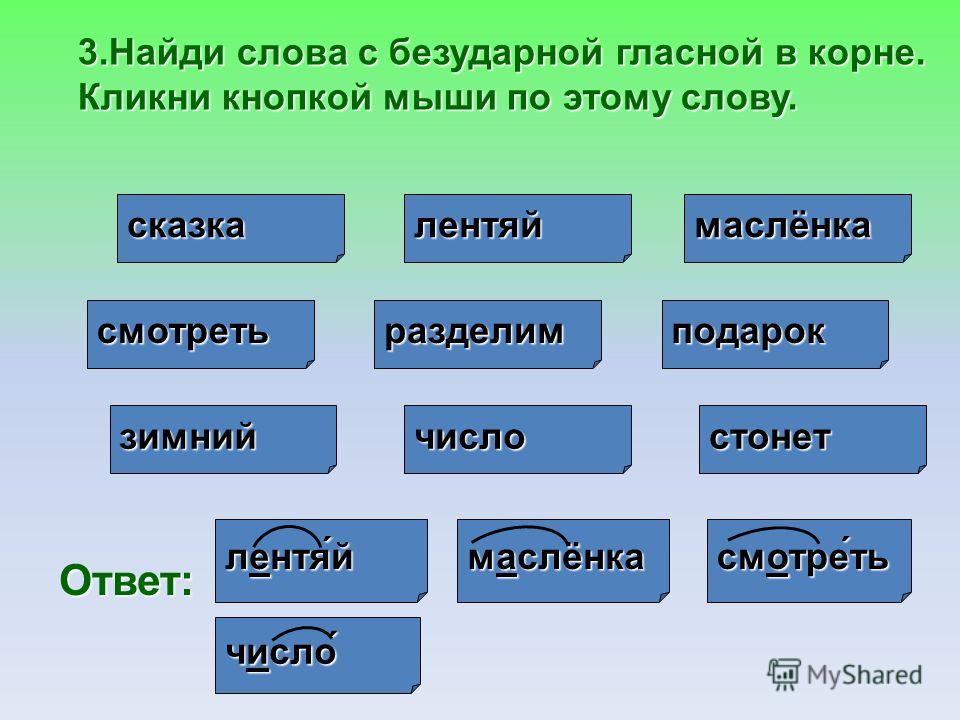 7C). Хотя изначально у большинства пациентов заболевание регрессировало, через 2 месяца развились многочисленные новые поражения; и как таковой, этот пациент не ответил.
7C). Хотя изначально у большинства пациентов заболевание регрессировало, через 2 месяца развились многочисленные новые поражения; и как таковой, этот пациент не ответил.
Рисунок 7
Перенесенные высокореактивные клетки перемещаются в опухоли меланомы и разрушают их у пациентов .(A) Последовательные биопсии подкожных опухолей у пациента 4 с ДМФ5 до (d0) и после лечения, окрашенные гематоксилином и эозином (слева) или анти-CD8 (справа). Исходное увеличение ×20. (B) Бедро 4 пациента с ДМФ5, покрытое множественными очагами подкожной меланомы до лечения и после частичной регрессии опухоли через 5 недель после лечения. (C) Проточная цитометрия TIL, выращенных из подкожной опухоли пациента 4 с DMF5, резецированной через 4 недели после лечения. (DF) До лечения и после лечения КТ легких (D) пациента 2 с DMF5, (E) головного мозга пациента 8 с DMF5 (вверху) и подмышечной впадины (внизу) и (F) gp100(154) легких пациента 14 (вверху) и печень (внизу). Стрелки указывают расположение метастазов меланомы.
Стрелки указывают расположение метастазов меланомы.
Рисунок 7
Перенесенные высокореактивные клетки перемещаются в опухоли меланомы и разрушают их у пациентов . (A) Последовательные биопсии подкожных опухолей у пациента 4 с ДМФ5 до (d0) и после лечения, окрашенные гематоксилином и эозином (слева) или анти-CD8 (справа). Исходное увеличение ×20. (B) Бедро 4 пациента с ДМФ5, покрытое множественными очагами подкожной меланомы до лечения и после частичной регрессии опухоли через 5 недель после лечения.(C) Проточная цитометрия TIL, выращенных из подкожной опухоли пациента 4 с DMF5, резецированной через 4 недели после лечения. (DF) До лечения и после лечения КТ легких (D) пациента 2 с DMF5, (E) головного мозга пациента 8 с DMF5 (вверху) и подмышечной впадины (внизу) и (F) gp100(154) легких пациента 14 (вверху) и печень (внизу). Стрелки указывают расположение метастазов меланомы.
Шесть из 20 пациентов (30%), получавших TCR DMF5, и 3 из 16 (19%), получавших TCR gp100(154), испытали объективный противоопухолевый ответ, как определено критериями RECIST 17 (таблицы 1–2). Опухоли регрессировали во многих органах, включая головной мозг, легкие, печень, лимфатические узлы и подкожные участки (рис. 7D-F). Случаев смерти, связанных с лечением, не было, и все пациенты выздоровели после лечения.
Опухоли регрессировали во многих органах, включая головной мозг, легкие, печень, лимфатические узлы и подкожные участки (рис. 7D-F). Случаев смерти, связанных с лечением, не было, и все пациенты выздоровели после лечения.
При анализе 36 пациентов не было обнаружено корреляции между количеством введенных клеток и вероятностью клинического ответа, при этом некоторые пациенты, ответившие на лечение, получили в логарифмическом отношении меньше клеток, чем другие (таблица 1).Также не было корреляции между клиническим ответом и продолжительностью выращивания клеток ex vivo, а также от того, получали ли они одну или две стимуляции ОКТ-3. Имелась корреляция с клиническим ответом и стойкостью введенных клеток через один месяц, что оценивалось с помощью анализа ELISPOT как специфического высвобождения IFN-γ, так и IL-2 (оба P = 0,02; фигура 3B). Хотя у всех ответивших пациентов через 1 месяц в крови сохранялось более или равно 10% тетрамер-позитивных клеток, этот параметр не коррелировал с ответом ( P = .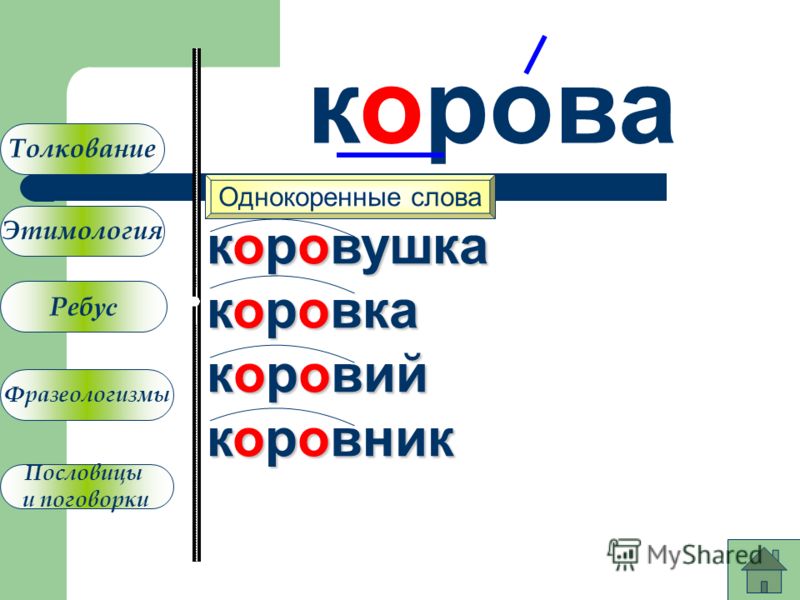 4; Рисунок 3Б).
4; Рисунок 3Б).
Способность генетически модифицированных нормальных периферических лимфоцитов, несущих высокоактивный TCR, разрушать изолированные антигенэкспрессирующие клетки, такие как отдельные меланоциты в ухе, базальном слое эпидермиса и в иммунопривилегированных участках, таких как глаза, а также метастазы меланомы во внутренних органах и головном мозге, имеет важное значение для развития иммунотерапии рака.Этот подход позволяет избежать необходимости идентификации и выделения противоопухолевых эффекторных клеток у каждого пациента. Эти генно-модифицированные клетки нацелены только на специфические общие антигены дифференцировки, и, таким образом, эти результаты подтверждают использование антигенов дифференцировки в качестве мишеней для иммунотерапии рака. Интересно, что клетки, экспрессирующие мышиный gp100(154) TCR, сохранялись на тех же уровнях, что и клетки, экспрессирующие человеческий DMF5 TCR, а также опосредовали регрессию рака, предполагая, что иммунная реакция против мышиных последовательностей TCR не может быть ограничивающим фактором в Генная терапия TCR у человека. Возможность использовать HLA-трансгенных мышей для повышения TCR против раковых антигенов человека представляет собой ценный подход, позволяющий обойти толерантность пациентов к собственным антигенам и, таким образом, сделать возможным создание высокореактивных противоопухолевых TCR. 21
Возможность использовать HLA-трансгенных мышей для повышения TCR против раковых антигенов человека представляет собой ценный подход, позволяющий обойти толерантность пациентов к собственным антигенам и, таким образом, сделать возможным создание высокореактивных противоопухолевых TCR. 21
Удивительно, что процент PBL, окрашивающихся положительно на тетрамер, всегда был выше, чем доля клеток, проявляющих противоопухолевую активность при окрашивании ELISPOT или внутриклеточным цитокином (таблицы 1–2; рисунок 3).Частично это различие может быть объяснено включением всех мононуклеарных клеток в функциональные анализы PBL пациентов по сравнению со способностью гейтировать популяцию CD3 + Т-лимфоцитов с помощью проточной цитометрии. Однако эта разница невелика по сравнению с логарифмами различий между тетрамер-позитивными клетками по данным FACS (приближается к 100% в образцах после лечения и до 80% в PBL после лечения) и в результатах ELISPOT (обычно <1% клеток). Также возможно, что, хотя окрашивание тетрамером идентифицирует все лимфоциты, экспрессирующие опухоле-реактивный TCR, не все клетки TCR + обладают одинаковой противоопухолевой функцией.Поскольку Т-лимфоциты могут иметь противоопухолевый эффекторный фенотип, они также могут быть анергическими или иметь супрессивный фенотип, например регуляторные Т-клетки. В случае этих последних фенотипов противоопухолевый функциональный ответ может быть снижен или отменен. Другим возможным объяснением является различная чувствительность прямого окрашивания тетрамером для проточной цитометрии по сравнению с ELISPOT или окрашиванием внутриклеточных цитокинов.
Также возможно, что, хотя окрашивание тетрамером идентифицирует все лимфоциты, экспрессирующие опухоле-реактивный TCR, не все клетки TCR + обладают одинаковой противоопухолевой функцией.Поскольку Т-лимфоциты могут иметь противоопухолевый эффекторный фенотип, они также могут быть анергическими или иметь супрессивный фенотип, например регуляторные Т-клетки. В случае этих последних фенотипов противоопухолевый функциональный ответ может быть снижен или отменен. Другим возможным объяснением является различная чувствительность прямого окрашивания тетрамером для проточной цитометрии по сравнению с ELISPOT или окрашиванием внутриклеточных цитокинов.
Распознавание нормальных покоящихся клеток, экспрессирующих целевой раковый антиген, поднимает очевидные вопросы, касающиеся токсичности этого подхода генной терапии.Эти результаты совпадают с нашими выводами на модели мышиной меланомы, которая показала связь глазной токсичности и противоопухолевой активности с использованием gp100 в качестве антигена-мишени, 22 , хотя клинически местное применение стероидов ослабляло нормальную тканевую токсичность у пациентов, получавших эти меланоцит-специфические TCR. Эти данные подчеркивают важность целевого опухолевого антигена. Может быть очень эффективным нацеливание антигенов дифференцировки на рак, возникающий в второстепенных органах, таких как простата, яичники, молочная железа и щитовидная железа.Другие виды рака имеют более высокую экспрессию антигенов дифференцировки, таких как CEA и Muc-1, чем в нормальных тканях, и это может создать терапевтическое окно, которое можно использовать. Класс антигенов рака яичек, не экспрессируемых в нормальных тканях взрослого человека, может быть идеальной мишенью для этого подхода. В любом случае необходимо тщательно взвесить риски и преимущества инфузии высокореактивных аутоантиген-специфических TCR.
Эти данные подчеркивают важность целевого опухолевого антигена. Может быть очень эффективным нацеливание антигенов дифференцировки на рак, возникающий в второстепенных органах, таких как простата, яичники, молочная железа и щитовидная железа.Другие виды рака имеют более высокую экспрессию антигенов дифференцировки, таких как CEA и Muc-1, чем в нормальных тканях, и это может создать терапевтическое окно, которое можно использовать. Класс антигенов рака яичек, не экспрессируемых в нормальных тканях взрослого человека, может быть идеальной мишенью для этого подхода. В любом случае необходимо тщательно взвесить риски и преимущества инфузии высокореактивных аутоантиген-специфических TCR.
Хотя низкие цифры в этих клинических испытаниях не позволяют сделать статистические выводы, частота объективного ответа на рак 30% и 19%, наблюдаемая при использовании TCR DMF5 и gp100(154), соответственно, была ниже, чем частота ответа от 51% до 72%, наблюдаемая при использование аутологичных TIL, отобранных и выращенных индивидуально из резецированной опухоли каждого пациента. 7,8,11 Передний увеит и ототоксичность наблюдались у 6 из 93 (6,5%) и только у одного из 93 (1,1%) пациентов с меланомой, получавших TIL, соответственно, в отличие от 41,7% случаев обоих видов токсичности, наблюдаемых в текущее исследование ( P < 0,001), иногда при отсутствии регрессии опухоли, подразумевая, что общие антигены меланомы/меланоцитов могут не быть преобладающими мишенями терапевтических TIL. Хотя многие TIL распознают антигены MART-1 и gp100, клоны Т-клеток были выделены из многих клинически эффективных TIL, которые также распознают мутированные или неидентифицированные антигены, и они также могут быть ответственны за противоопухолевые эффекты переноса TIL.В качестве альтернативы, это открытие может объясняться различиями в самонаводящихся молекулах или другими фенотипическими различиями между кровью и лимфоцитами, происходящими из опухоли.
7,8,11 Передний увеит и ототоксичность наблюдались у 6 из 93 (6,5%) и только у одного из 93 (1,1%) пациентов с меланомой, получавших TIL, соответственно, в отличие от 41,7% случаев обоих видов токсичности, наблюдаемых в текущее исследование ( P < 0,001), иногда при отсутствии регрессии опухоли, подразумевая, что общие антигены меланомы/меланоцитов могут не быть преобладающими мишенями терапевтических TIL. Хотя многие TIL распознают антигены MART-1 и gp100, клоны Т-клеток были выделены из многих клинически эффективных TIL, которые также распознают мутированные или неидентифицированные антигены, и они также могут быть ответственны за противоопухолевые эффекты переноса TIL.В качестве альтернативы, это открытие может объясняться различиями в самонаводящихся молекулах или другими фенотипическими различиями между кровью и лимфоцитами, происходящими из опухоли.
Несколько пациентов проявляли токсичность меланоцитов без отторжения опухоли. Лечение пациентов с низкой (DMF4) и высокой (DMF5) авидностью TCR, распознающими один и тот же антиген MART-1, приводило к 13% (4 из 34) и 30% (6 из 20) объективных ответов опухоли, соответственно, что позволяет предположить, что повышенная авидность TCR может улучшить отторжение опухоли.Однако из-за небольшого количества образцов возможно, что авидность TCR in vivo не является основным фактором, определяющим отторжение опухоли. Вероятно, необходимо учитывать такие факторы, как гомогенность лимфоцитов к нормальной ткани по сравнению с опухолевой, гетерогенность экспрессии антигена опухолью, истощение Т-клеток, подавление в микроокружении опухоли или потребность в поливалентных популяциях Т-клеток. для улучшения отторжения опухоли у пациентов. Модели опухолей мышей предсказывают, что усиление лимфодеплеции перед переносом клеток и введение вакцины для стимуляции перенесенных клеток in vivo может улучшить противоопухолевую эффективность этого подхода, 10,23 , и в настоящее время мы проводим клиническое испытание, сочетающее DMF5 и gp100 ( 154) TCR, с усилением иммунодефицита пациента, с последующим введением пептидной вакцины.
Лечение пациентов с низкой (DMF4) и высокой (DMF5) авидностью TCR, распознающими один и тот же антиген MART-1, приводило к 13% (4 из 34) и 30% (6 из 20) объективных ответов опухоли, соответственно, что позволяет предположить, что повышенная авидность TCR может улучшить отторжение опухоли.Однако из-за небольшого количества образцов возможно, что авидность TCR in vivo не является основным фактором, определяющим отторжение опухоли. Вероятно, необходимо учитывать такие факторы, как гомогенность лимфоцитов к нормальной ткани по сравнению с опухолевой, гетерогенность экспрессии антигена опухолью, истощение Т-клеток, подавление в микроокружении опухоли или потребность в поливалентных популяциях Т-клеток. для улучшения отторжения опухоли у пациентов. Модели опухолей мышей предсказывают, что усиление лимфодеплеции перед переносом клеток и введение вакцины для стимуляции перенесенных клеток in vivo может улучшить противоопухолевую эффективность этого подхода, 10,23 , и в настоящее время мы проводим клиническое испытание, сочетающее DMF5 и gp100 ( 154) TCR, с усилением иммунодефицита пациента, с последующим введением пептидной вакцины.
Также было показано, что адоптивная клеточная терапия с использованием геномодифицированных лимфоцитов эффективна при лечении злокачественных новообразований, отличных от меланомы. В доклинических моделях лимфоциты были генетически модифицированы для экспрессии костимулирующих молекул, предназначенных для повышения противоопухолевой активности и выживаемости Т-клеток, таких как лиганд CD80 или 4-1BB, или для устранения эффектов ингибирующих сигналов. 24 Генетически модифицированные лимфоциты человека, экспрессирующие химерные антигенные рецепторы (CAR), 25 , сочетающие распознавание антигена антитела с сигнальными мотивами Т-клеток, были способны уничтожать В-клеточную лимфому человека путем нацеливания на CD19, 26 и человеческий опухоли предстательной железы путем нацеливания на ErbB2/Her2 27 в моделях ксеногенных мышей SCID.Использование Т-клеток человека, специфичных к вирусу Эпштейна-Барр, трансдуцированных CAR, распознающими антиген CD30, специфичный для лимфомы Ходжкина, также оказалось эффективным в ксеногенной мышиной модели. 28 Клинически специфичные к вирусу Эпштейна-Барра клетки, экспрессирующие CAR, которые распознают опухолеспецифический диасиалоганглиозид GD2 нейробластомы, недавно также успешно использовались для лечения пациентов с нейробластомой. 29
28 Клинически специфичные к вирусу Эпштейна-Барра клетки, экспрессирующие CAR, которые распознают опухолеспецифический диасиалоганглиозид GD2 нейробластомы, недавно также успешно использовались для лечения пациентов с нейробластомой. 29
В попытке расширить эту терапию TCR для лечения рака, отличного от меланомы, мы клонировали гены, кодирующие TCR с высокой авидностью, которые распознают различные антигенные эпитопы, такие как NY-ESO-1:157-165, 30 p53:264-272, 21,31 и CEA:691-699, 32 экспрессируются при многих распространенных эпителиальных раковых заболеваниях. 9 Для нацеливания на опухоли способом, не ограниченным главным комплексом гистосовместимости, мы также сконструировали лимфоциты для экспрессии химерных рецепторов, включающих сайт связывания антигена анти-Her2 или анти-CD19 mAb, 26 , которые могут нацеливаться на опухолевые клетки, сверхэкспрессирующие эти детерминанты на клеточной поверхности.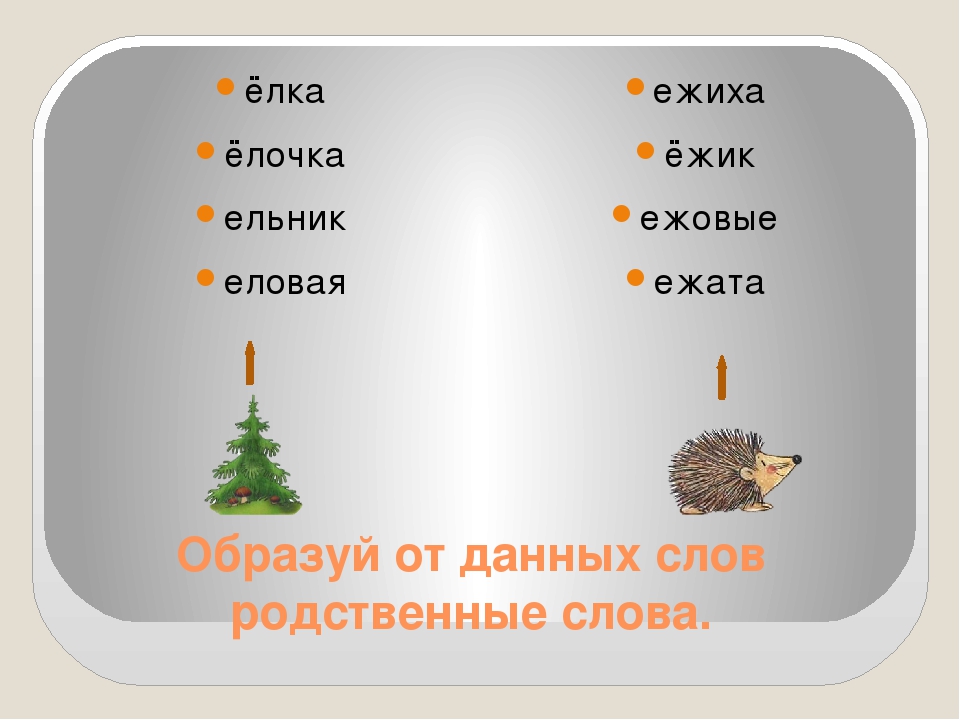
При применении этого подхода необходимо учитывать возможную токсичность, возникающую в результате экспрессии опухолеассоциированных антигенов на нормальных тканях.Наши результаты, однако, подтверждают гипотезу о том, что введение онкологическим больным Т-клеток, трансдуцированных высокореактивными человеческими или мышиными TCR, может опосредовать in vivo разрушение тканей, которые экспрессируют антиген-мишень, и предполагают, что терапия переносом клеток может быть ценным дополнением. для лечения пациентов с метастатическим раком.
Затраты на публикацию этой статьи были частично покрыты за счет оплаты страницы.Поэтому и исключительно для того, чтобы указать на этот факт, эта статья настоящим помечена как «реклама» в соответствии с разделом 1734 18 USC.
Предоставлено: L. A.J. подготовил клетки для лечения пациентов DMF5, провел лабораторные эксперименты и анализ и написал статью; ОЗУ. готовили клетки для лечения пациентов gp100(154) и проводили эксперименты; M.E.D., A.P.C. и J.R.W. подготовленные лечебные ячейки пациента; С.Л.С. проведенные эксперименты; Л.К. и Н.П.Р. идентифицировали и выделили gp100(154) TCR; J.C.Y., M.S.H., U.S.K., R.E.R., R.M.S., J.L.D., A.M., R.T.R. и S.A.R. были врачи и Д.А.Н. была медсестрой-исследователем по клиническим протоколам; ОЗУ. сконструировали ретровирусные векторы TCR; К.-К.Р.Л. проведено иммуногистологическое окрашивание биоптатов пациентов; Р.Дж.Б. проводила офтальмологические осмотры пациентов; H.K., C.C.B., S.F.R. и C.V.W. провела слуховые оценки пациентов; К.М.Л. оказал поддержку в подготовке клинического протокола и отчетности; С.А.Р. руководил клиническими испытаниями и участвовал в анализе данных и написании статьи; Р.Дж.Б. предоставил информацию об офтальмологическом обследовании пациентов; H.K., C.C.B., S.F.R.
A.J. подготовил клетки для лечения пациентов DMF5, провел лабораторные эксперименты и анализ и написал статью; ОЗУ. готовили клетки для лечения пациентов gp100(154) и проводили эксперименты; M.E.D., A.P.C. и J.R.W. подготовленные лечебные ячейки пациента; С.Л.С. проведенные эксперименты; Л.К. и Н.П.Р. идентифицировали и выделили gp100(154) TCR; J.C.Y., M.S.H., U.S.K., R.E.R., R.M.S., J.L.D., A.M., R.T.R. и S.A.R. были врачи и Д.А.Н. была медсестрой-исследователем по клиническим протоколам; ОЗУ. сконструировали ретровирусные векторы TCR; К.-К.Р.Л. проведено иммуногистологическое окрашивание биоптатов пациентов; Р.Дж.Б. проводила офтальмологические осмотры пациентов; H.K., C.C.B., S.F.R. и C.V.W. провела слуховые оценки пациентов; К.М.Л. оказал поддержку в подготовке клинического протокола и отчетности; С.А.Р. руководил клиническими испытаниями и участвовал в анализе данных и написании статьи; Р.Дж.Б. предоставил информацию об офтальмологическом обследовании пациентов; H.K., C.C.B., S.F.R. и C.V.W. предоставил информацию об отоларингологическом обследовании пациентов; и Л.А.Дж. и С.А.Р. проанализированы данные, к которым имеют доступ все авторы.
и C.V.W. предоставил информацию об отоларингологическом обследовании пациентов; и Л.А.Дж. и С.А.Р. проанализированы данные, к которым имеют доступ все авторы.
Раскрытие информации о конфликте интересов: авторы заявляют об отсутствии конкурирующих финансовых интересов.
Текущий адрес L.A.J. Отделение нейрохирургии, отделение хирургии, Центр опухолей головного мозга имени Престона Роберта Тиша, Медицинский центр Университета Дьюка, Дарем, Северная Каролина.
Переписка: Стивен А. Розенберг, отделение хирургии, Национальный институт рака, здание 10CRC, Центр клинических исследований Хэтфилда, Бетесда, Мэриленд, 20892; электронная почта SAR@nih.правительство
итальянских родственных и заимствованных слов: 17 правил итальянизации английских слов, которые вы уже знаете
Начинающий итальянский? Вместо того, чтобы нырять прямо в глубь огромного пула итальянских слов, сначала мы собираемся окунуться в них. В этом руководстве вы выучите несколько итальянских слов, которые не покажутся вам совершенно иностранными. потому что они точно такие же на английском языке. Ура!
В этом руководстве вы выучите несколько итальянских слов, которые не покажутся вам совершенно иностранными. потому что они точно такие же на английском языке. Ура!
Затем мы научимся применять 17 простых правил, чтобы вы могли начать итальянизировать английских слов, которые вы уже знаете, включая существительные, глаголы и наречия.Изучение этих правил пригодится. С помощью этих правил вы сможете разблокировать тонну итальянского словарного запаса без особых усилий.
Пронти? Готов? Коминциамо! Начнем!
Английские заимствования на итальянском языке
Хотя итальянцы очень гордятся своим языком, они позволили проникнуть в него некоторым английским словам. Как правило, они произносятся так же, как и в английском, за некоторыми исключениями.Вы услышите такие слова, как гаджеты , бег трусцой , чувство и шок . Вы даже услышите, как они используют слово , хорошо, . С тех пор как компьютеры вошли в нашу жизнь, итальянцы даже говорят cliccare sul mous e , что означает «щелкнуть мышью». Наконец, есть lo zapping , что означает переключение телеканалов с помощью пульта дистанционного управления.
Вы даже услышите, как они используют слово , хорошо, . С тех пор как компьютеры вошли в нашу жизнь, итальянцы даже говорят cliccare sul mous e , что означает «щелкнуть мышью». Наконец, есть lo zapping , что означает переключение телеканалов с помощью пульта дистанционного управления.
Рассмотрим следующий список слов. Каждый показан с соответствующим итальянским определенным артиклем или более известным как слово «the», о котором мы узнаем больше позже.
- радио
- л’автобус
- антенна
- отель
- район
- идея
- Гамбургер
- иль коктейль
- иль джаз
- иль кинотеатр
- ил компьютер
- иль бар
- иль блюз
- иль пленка
- иль рок-н-ролл
- иль выходные
- иль кемпинг
- я джинсы
- вот сноб
- шок
- пойти за покупками
- спорт
- вот шампунь
- ло зоопарк
Это лишь некоторые из множества английских слов, вошедших в итальянский язык. Вы можете увидеть мое полное руководство по английским словам на итальянском языке здесь . Точно так же многие итальянские слова используются и в англоязычных странах! В том числе:
Вы можете увидеть мое полное руководство по английским словам на итальянском языке здесь . Точно так же многие итальянские слова используются и в англоязычных странах! В том числе:
- пицца
- макароны
- спагетти гли
- тортеллини
- моцарелла
- эспрессо
- Капучино
- il panino – бутерброд / i panini – бутерброды
- i biscotti – печенье / il biscotti – печенье
- тирамису
Кстати, знаете ли вы, что тирамису буквально означает «подтяни меня»? Это относится к тому факту, что этот сладкий десерт сделан из крепкого итальянского кофе эспрессо, который эффективно дает вам заряд энергии или «поднимает настроение».
Итальянские родственные слова
Помимо слов, которые проникли в язык напрямую, итальянский и английский языки имеют много родственных слов. Родственное слово — это слово в одном языке, которое имеет то же происхождение, что и слово в другом языке, и может звучать похоже. Вот несколько примеров:
Родственное слово — это слово в одном языке, которое имеет то же происхождение, что и слово в другом языке, и может звучать похоже. Вот несколько примеров:
- аэропорт – аэропорт
- внимание – внимание
- связь – связь
- важное – важное
- невероятно – невероятно
К счастью, есть несколько правил, которые можно применить к итальянским словам, которые вы уже знаете.Вот несколько советов, которые помогут вам распознавать и использовать родственные итальянские слова.
Итальянизация существительных
Теперь, когда мы начинаем изучать грамматику, давайте освежим в памяти некоторые грамматические термины. Не напрягайтесь. Мы собираемся сделать это очень простым.
Не знаете, что такое существительное? Чтобы определить существительное, задайте себе этот простой вопрос. Могу ли я поставить «the», «a» или «an» перед словом? Если ответ да, то вы нашли себе существительное!
Например, слово пицца является существительным, потому что когда мы говорим « пицца» или « пицца» это имеет смысл. Однако говорить « восхитительный» или « восхитительный» сам по себе не имеет смысла.
Однако говорить « восхитительный» или « восхитительный» сам по себе не имеет смысла.
Имея это в виду, вот несколько правил, которые помогут увеличить словарный запас существительных.
Правило 1: английские существительные, оканчивающиеся на -y, становятся -ia в итальянском
Вот несколько примеров:
- биология – биология
- философия – философия
- социология – социология
- анатомия – анатомия
Вот несколько примеров:
- музыка – музыка
- политика – политика
- республика – ла республика
- математика – математика
Правило 3: Существительные в английском языке, оканчивающиеся на -ty, становятся на -tà в итальянском
Вот несколько примеров:
- город – ла-Сити
- удостоверение личности – удостоверение личности
- общество – ла общество
- университет – университет
Правило 4: Существительные в английском языке, оканчивающиеся на -ce, становятся -za в итальянском
Вот несколько примеров:
- важность – l’importanza
- элегантность – элегантность
- насилие – насилие
- терпения – la pazienza
Правило 5: Существительные в английском языке, оканчивающиеся на -tion, становятся -zione в итальянском
Вот несколько примеров:
- нация – ла национе
- внимание – внимание
- образование – образование
- ситуация – ситуация
Правило 6: Существительные в английском языке, оканчивающиеся на -o(u)r, становятся на -ore в итальянском
Вот несколько примеров:
- актер – актер
- врач – иль дотторе
- профессор – иль профессор
- мотор – иль мотор
Правило 7: Существительные в английском языке, оканчивающиеся на -ary, становятся -ario в итальянском
Вот несколько примеров:
- секретарь – il segretario
- словарный запас – словарный запас зарплата
- – иль зарплата
- функционер – il funzionario
Правило 8: Существительные в английском языке, оканчивающиеся на -ist, становятся в итальянском языке -ista
Вот несколько примеров:
- художник – художник
- скрипач – скрипач
- пианист – пианист
- оптимист – оптимист
Итальянизация прилагательных
Ранее мы говорили о существительных в нашем примере пицца . Давайте сделаем еще один шаг в этом примере и добавим к нему прилагательное.
Давайте сделаем еще один шаг в этом примере и добавим к нему прилагательное.
Прилагательные можно определить, поставив перед словом «есть». Например, «пицца вкусная» или «кофе горячая».
Вот несколько правил, которые помогут вам расширить словарный запас прилагательных.
Правило 9: Прилагательные в английском языке, оканчивающиеся на -al, превращаются в -ale в итальянском
Вот несколько примеров:
- специальный – специальный
- личных – личных
- оригинал – оригинал
- сентиментальный – сентиментальный
Правило 10: Прилагательные в английском языке, оканчивающиеся на -ect, становятся на -etto в итальянском
Вот несколько примеров:
- совершенный – совершенный
- правильно – правильно
- прямостоячий – эретто
- прямой – прямой
Правило 11: Прилагательные в английском языке, оканчивающиеся на -ical, становятся на -ico в итальянском
Вот несколько примеров:
- типичный – типико
- классический – классический
- политик – политик
- географический – географический
Правило 12: Прилагательные в английском языке, оканчивающиеся на -ous, становятся в итальянском языке -oso
Вот несколько примеров:
- щедрый – щедрый
- любопытных – любопытных
- нервный – нервный
- амбициозный – амбициозный
Правило 13: Прилагательные в английском языке, оканчивающиеся на -able или -ible, становятся -abile и -ibile в итальянском
Вот несколько примеров:
- возможно – возможно
- видимых – видимых
- ужасный – ужасный
- ответственный – ответственный
- очаровательный – очаровательный
Итальянизация глаголов
И последнее, но не менее важное: у нас есть все важные глаголы.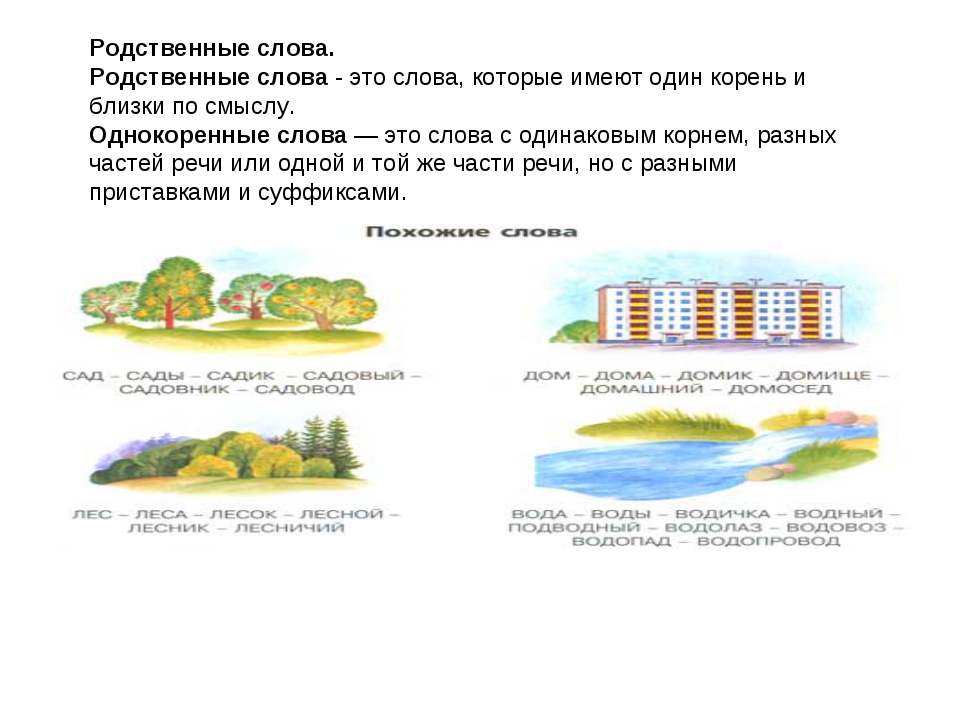 Глаголы составляют основу любого языка. Один только этот термин может вызвать страх, но, как и раньше, мы будем делать все очень просто.
Глаголы составляют основу любого языка. Один только этот термин может вызвать страх, но, как и раньше, мы будем делать все очень просто.
Глагол можно определить, добавив перед словом от до . Например, от до делать, от до есть, от до готовить, от до путешествовать . Это все глаголы. Это так просто! Важно, чтобы вы узнавали глаголы, так как они помогут вам легко строить предложения.
Стоит отметить, что в итальянском языке часть глагола «to» уже включена в состав слова и определяется тремя последними буквами.Есть только три разных окончания глагола, это:
Например, глагол для съесть имеет вид mangiare , поэтому часть «to» — это окончание -are . Вы сможете определить эти окончания в следующих правилах.
Правило 14: Глаголы в английском языке, оканчивающиеся на -cate, в итальянском превращаются в -care.
Вот несколько примеров:
- для обучения – educare
- для указания – indicare
- усложнить – усложнить
- подразумевать, подразумевать – подразумевать
Правило 15: Глаголы в английском языке, оканчивающиеся на -ize(-ise) или -yze(-yse), становятся на -izzare в итальянском
Вот несколько примеров:
- для организации – organizzare
- сочувствовать – сочувствовать
- для анализа – для анализа
- для минимизации – для минимизации
Правило 16: Глаголы в английском языке, оканчивающиеся на -ish, становятся -ire в итальянском
Вот несколько примеров:
- до конца – до конца
- отменить – отменить
- для украшения – abbellire
- установить – стабильный
Загрузите руководство по 24 наиболее употребительным итальянским глаголам и и узнайте, как их спрягать, выполнив 3 простых шага.
Итальянизация наречий
Любезно , медленно , здесь , часто и очень — все это примеры наречий. Наречия изменяют глаголы, прилагательные или другие наречия. Под изменить мы подразумеваем добавление или изменение значения слова.
В следующих примерах наречие выделено жирным шрифтом , а глагол, который оно изменяет, выделено курсивом .
- Джон говорит громко .(Как говорит Джон?)
- Потом она выкурила сигарету. (Когда она курила?)
- Мэри живет локально . (Где живет Мэри?)
Но наречия также могут изменять прилагательные. Посмотрите на эти примеры:
Изменить прилагательное:
- Он действительно красивый .
 (Насколько он красив?)
(Насколько он красив?) - Это было чрезвычайно любезно с вашей стороны.
Изменить другое наречие:
- Она водит невероятно медленно .(Как медленно она ездит?)
- Он водит чрезвычайно быстро .
Теперь, когда мы знаем, что такое наречие, давайте рассмотрим следующий набор правил.
Правило 17: Наречия в английском языке, оканчивающиеся на
-ably и -ily , становятся — abilmente и -ibilmente соответственно, в итальянскомВот несколько примеров:
- вероятно – вероятно
- возможно – возможно
По мере того, как вы пополняете свой словарный запас и еще не знаете слово для чего-либо, вы можете итальянизировать английское слово, используя эти 17 правил.Конечно, есть некоторые исключения из этих правил, но вы будете удивлены, как часто они применяются! Как и во всем, вы учитесь на собственном опыте. Какое правило/какие правила удивили вас больше всего? Позвольте мне знать в комментариях ниже.
Какое правило/какие правила удивили вас больше всего? Позвольте мне знать в комментариях ниже.
Понравился этот урок? Вы новичок или уже изучаете итальянский? Предстоит поездка или вы хотите пообщаться со своим итальянским партнером или родственниками на итальянском языке? Учи итальянский по моей уникальной методике 80/20
Открыта регистрация для присоединиться к Intrepid Italian, к моей новой серии онлайн-видеокурсов, в которых используется мой уникальный метод 80/20.Вы пройдете путь от застенчивого, сбитого с толку новичка до опытного и уверенного в себе оратора среднего уровня, а я буду вашим верным проводником.
Наконец-то вы сможете связаться со своим итальянским партнером, поговорить с родственниками и насладиться подлинными впечатлениями от путешествий по Италии, о которых вы всегда мечтали, и многое другое.
Как носитель английского языка, выучивший итальянский во взрослом возрасте, я знаю, каково это чувствовать себя безнадежным и неуверенным в себе, чтобы говорить. Я знаю, каково это начинать с нуля и даже вернуться к абсолютным основам и узнать, что такое глагол!
Я знаю, каково это начинать с нуля и даже вернуться к абсолютным основам и узнать, что такое глагол!
Intrepid Italian был создан с мыслью о ВАС.Я использую свои практические знания английского языка, чтобы помочь вам войти в «итальянское мышление», чтобы вы могли избежать распространенных ловушек и ошибок, которые допускают носители английского языка — потому что я тоже когда-то их делал! Я разбиваю все таким образом, чтобы это «щелкало» и имело смысл.
Независимо от вашего уровня, для вас есть курс итальянского языка Intrepid, включающий:
Вы можете присоединиться к 1, 2 или всем 3 курсам, решать только вам. Самое приятное то, что у вас есть пожизненный доступ, поэтому вы можете учиться в любое время, в любом месте и на любом устройстве.
В качестве вашего проводника я проведу вас через каждый урок, шаг за шагом, используя мой уникальный метод 80/20. Мой подход отличается от традиционных методов, потому что я с самого начала учу вас самым важным 20% языка, чтобы вы сразу могли начать говорить.
Каждый курс включает в себя видеоуроки, аудиоупражнения, загружаемые рабочие листы, бонусные руководства, частное сообщество поддержки и пожизненный доступ — все это предназначено для упрощения обучения и развлечения.
Он даже поставляется с моим знаменитым « Празднуйте с гарантией Spritz» .После 30 дней использования Intrepid Italian, если вы не хотите отмечать свои новообретенные навыки итальянского с помощью Aperol Spritz, вам не нужно платить ни копейки! Ваше здоровье! 🥂
Присоединяйтесь к Intrepid Italian здесь и начните учить уже сегодня!
Ci vediamo lì! (До встречи!)
Учить итальянский? Ознакомьтесь с этими справочниками по итальянскому языку
Нравится? Закрепите это на потом!
Вам слово!
Было ли это руководство полезным для вас? Есть вопрос? Дайте мне знать, используя раздел комментариев ниже, или присоединяйтесь ко мне в социальных сетях @intrepidguide или @intrepiditalian , чтобы начать разговор.
Спасибо за прочтение. Надеюсь, вам понравился этот пост.
Нравится то, что вы видите? Подпишитесь, используя форму ниже, чтобы все мои сообщения доставлялись прямо на вашу электронную почту.
Концентрация CD8+ Т-клеток определяет их эффективность в уничтожении родственных антиген-экспрессирующих сингенных клеток млекопитающих in vitro и в тканях мыши | Журнал экспериментальной медицины
Представленные здесь результаты имеют очевидное значение для клеточной иммунотерапии меланомы и, возможно, других опухолей.Клеточная иммунотерапия приводит к объективной регрессии меланомы более чем у 50% пациентов, но излечивает очень небольшой процент из них (Dudley et al., 2002; Dudley and Rosenberg, 2003). В этой статье предлагается по крайней мере одна причина такого разрыва между реакцией и излечением: это неспособность доставлять и поддерживать достаточную внутриопухолевую концентрацию специфичных к опухолевому антигену CD8 + Т-клеток/мл опухоли в течение периода времени, достаточного для уничтожения 100 % клоногенных опухолевых клеток (рис. 5 и 7). Исследования, о которых сообщается в этой статье, предоставляют количественную информацию об этих параметрах.Они показывают, что концентрация ≥10 7 активированных клеток OT-1/мл необходима для уничтожения 100% клеток SIINFEKL-B16/мл коллагеново-фибринового геля за 7 дней (рис. 5). В этих гелях цитолиз клеток-мишеней зависит только от концентрации Т-клеток CD8 + (рис. 2, 5 и 7). Соответственно, если предположить, что меланомы содержат 3 × 10 8 клеток меланомы/мл или/г опухоли (Stephens and Peacock, 1978), и что антиген-специфические CD8 + Т-клетки меланомы убивают клетки меланомы in vivo с той же скоростью, что и клетки ОТ-1, использованные в экспериментах, показанных на рис.5, доставка и поддержание ≥10 7 меланомных антиген-специфических CD8 + Т-клеток/г меланомы в течение ≥8 дней должно привести к эрадикации меланомы. Однако наш анализ результатов Petersen et al. (2006; рис. 7 и таблица S4) показывает, что клетки OT-1 убивают клетки ova-B16 примерно в девять раз ниже ( k ) и имеют примерно в восемь раз более высокий CTC в опухолях in vivo, чем в коллагеново-фибриновых гелях.
5 и 7). Исследования, о которых сообщается в этой статье, предоставляют количественную информацию об этих параметрах.Они показывают, что концентрация ≥10 7 активированных клеток OT-1/мл необходима для уничтожения 100% клеток SIINFEKL-B16/мл коллагеново-фибринового геля за 7 дней (рис. 5). В этих гелях цитолиз клеток-мишеней зависит только от концентрации Т-клеток CD8 + (рис. 2, 5 и 7). Соответственно, если предположить, что меланомы содержат 3 × 10 8 клеток меланомы/мл или/г опухоли (Stephens and Peacock, 1978), и что антиген-специфические CD8 + Т-клетки меланомы убивают клетки меланомы in vivo с той же скоростью, что и клетки ОТ-1, использованные в экспериментах, показанных на рис.5, доставка и поддержание ≥10 7 меланомных антиген-специфических CD8 + Т-клеток/г меланомы в течение ≥8 дней должно привести к эрадикации меланомы. Однако наш анализ результатов Petersen et al. (2006; рис. 7 и таблица S4) показывает, что клетки OT-1 убивают клетки ova-B16 примерно в девять раз ниже ( k ) и имеют примерно в восемь раз более высокий CTC в опухолях in vivo, чем в коллагеново-фибриновых гелях. В этих условиях для эрадикации опухоли потребуется доставка и поддержание ≥10 8 меланомных антиген-специфических CD8 + Т-клеток/мл или/г меланомы в течение ≥8 дней.Это очень высокий порог, который не сулит успеха в клеточной иммунотерапии рака. Тем не менее, в небольшом числе случаев было показано, что клеточная иммунотерапия является излечивающей (Dudley et al., 2002), предполагая, что in vivo существуют другие факторы, которые при определенных обстоятельствах способствуют эффективности антиген-специфических CD8 + Т-клетки. Мы опишем четыре из этих факторов в следующем параграфе.
В этих условиях для эрадикации опухоли потребуется доставка и поддержание ≥10 8 меланомных антиген-специфических CD8 + Т-клеток/мл или/г меланомы в течение ≥8 дней.Это очень высокий порог, который не сулит успеха в клеточной иммунотерапии рака. Тем не менее, в небольшом числе случаев было показано, что клеточная иммунотерапия является излечивающей (Dudley et al., 2002), предполагая, что in vivo существуют другие факторы, которые при определенных обстоятельствах способствуют эффективности антиген-специфических CD8 + Т-клетки. Мы опишем четыре из этих факторов в следующем параграфе.
Во-первых, наши расчеты относительно концентрации специфичных к опухолевому антигену CD8 + Т-клеток, которые должны накапливаться в ложе опухоли для эрадикации опухоли, предполагают, что эти CD8 + Т-клетки являются единственными цитолитическими эффекторами.Однако присутствие активированных опухолевых антиген-специфических CD8 + Т-клеток в ложе опухоли может способствовать накоплению и уничтожению опухолевых клеток другими цитолитическими эффекторными клетками (например, NK-клетками и активированными макрофагами). В этих условиях CD8 + Т-клетки могут быть только одной из нескольких цитолитически активных эффекторных клеток, работающих в опухоли. Следовательно, наши расчеты могут недооценивать общую цитоцидную активность всех эффекторных лейкоцитов, которые собираются в опухоли в ответ на сигналы, инициированные опухолевыми антиген-специфическими CD8 + Т-клетками и/или цитолизом опухолевых клеток.Во-вторых, наши расчеты предполагают, что уничтожение опухолевых клеток клонами CD8 + Т-клеток, TCR которых распознают разные опухолевые антигены, является просто аддитивным. Возможно, он мультипликативен. Если да, то наличие внутри опухоли 3 × 10 6 CD8 + Т-клеток по сравнению с предполагаемым опухолевым антигеном А, 3 × 10 6 CD8 + Т-клеток по сравнению с предполагаемым опухолевым антигеном В и 3 × 10 6 CD8 + Т-клетки по сравнению с предполагаемым опухолевым антигеном С могут привести к 27-кратному увеличению гибели опухолевых клеток.
В этих условиях CD8 + Т-клетки могут быть только одной из нескольких цитолитически активных эффекторных клеток, работающих в опухоли. Следовательно, наши расчеты могут недооценивать общую цитоцидную активность всех эффекторных лейкоцитов, которые собираются в опухоли в ответ на сигналы, инициированные опухолевыми антиген-специфическими CD8 + Т-клетками и/или цитолизом опухолевых клеток.Во-вторых, наши расчеты предполагают, что уничтожение опухолевых клеток клонами CD8 + Т-клеток, TCR которых распознают разные опухолевые антигены, является просто аддитивным. Возможно, он мультипликативен. Если да, то наличие внутри опухоли 3 × 10 6 CD8 + Т-клеток по сравнению с предполагаемым опухолевым антигеном А, 3 × 10 6 CD8 + Т-клеток по сравнению с предполагаемым опухолевым антигеном В и 3 × 10 6 CD8 + Т-клетки по сравнению с предполагаемым опухолевым антигеном С могут привести к 27-кратному увеличению гибели опухолевых клеток. В-третьих, наши расчеты предполагают, что все Т-клетки CD8 + убивают с той же эффективностью, что и клетки ОТ-1. CD8 + Т-клетки сильно различаются по своей эффективности уничтожения (Stuge, et al., 2004). Соответственно, возможно получить популяции Т-клеток CD8 + , которые убивают с более высокой скоростью, чем наблюдаемая для клеток ОТ-1. Действительно, мы определили протоколы активации клеток OT-1, которые более чем удваивают свою эффективность в уничтожении клеток SIINFEKL-B16 в коллагеново-фибриновых гелях (неопубликованные данные).В-четвертых, мы предположили, что опухолевая сосудистая сеть всех меланом одинаково эффективна в доставке CD8 + Т-клеток в паренхиму опухоли. Буканович и др. (2008) показали, что эффективность этого процесса можно повысить более чем в 10 раз. По всем изложенным здесь причинам важно определить эффективность проникновения ex vivo размноженных CD8 + Т-клеток в ложе опухоли, время их пребывания в ложе опухоли и их цитолитическую активность как в идеальных условиях (т.
В-третьих, наши расчеты предполагают, что все Т-клетки CD8 + убивают с той же эффективностью, что и клетки ОТ-1. CD8 + Т-клетки сильно различаются по своей эффективности уничтожения (Stuge, et al., 2004). Соответственно, возможно получить популяции Т-клеток CD8 + , которые убивают с более высокой скоростью, чем наблюдаемая для клеток ОТ-1. Действительно, мы определили протоколы активации клеток OT-1, которые более чем удваивают свою эффективность в уничтожении клеток SIINFEKL-B16 в коллагеново-фибриновых гелях (неопубликованные данные).В-четвертых, мы предположили, что опухолевая сосудистая сеть всех меланом одинаково эффективна в доставке CD8 + Т-клеток в паренхиму опухоли. Буканович и др. (2008) показали, что эффективность этого процесса можно повысить более чем в 10 раз. По всем изложенным здесь причинам важно определить эффективность проникновения ex vivo размноженных CD8 + Т-клеток в ложе опухоли, время их пребывания в ложе опухоли и их цитолитическую активность как в идеальных условиях (т. g., коллагеново-фибриновые гели) и в условиях in vivo.
g., коллагеново-фибриновые гели) и в условиях in vivo.
Исследования, о которых сообщается в этой статье, определяют три параметра, которые представляются особенно важными для понимания терапевтического потенциала клеточной иммунотерапии. Во-первых, это концентрация всех цитолитических эффекторных клеток, доставленных в опухоль и активных внутри нее. Во-вторых, эффективность уничтожения ( k ) всех эффекторных клеток по отдельности и в комбинации в ложе опухоли.В-третьих, это продолжительность времени, в течение которого каждая из этих эффекторных клеток остается цитолитически активной в ложе опухоли. Информация об этих параметрах позволит исследователям оценить, является ли высокая частота объективной регрессии опухоли, но низкая частота излечения, наблюдаемая у пациентов с меланомой, получавших ex vivo-активированные CD8 + Т-клетки, следствием доставки недостаточного количества опухолевых антиген-специфических CD8 + Т-клеток к опухолям для повышения внутриопухолевой концентрации в 30 раз выше ЦОК (рис. 5), или вредного воздействия окружения опухоли на жизнеспособность и/или эффекторные функции CD8 + Т-клеток. Текущие исследования показывают, что система коллаген-фибриновый гель, описанная в этой статье, также полезна для оценки уничтожения клонированными опухолевыми антиген-специфичными CD8 + Т-клетками человека клеток меланомы человека, экспрессирующих родственный антиген (неопубликованные данные).
5), или вредного воздействия окружения опухоли на жизнеспособность и/или эффекторные функции CD8 + Т-клеток. Текущие исследования показывают, что система коллаген-фибриновый гель, описанная в этой статье, также полезна для оценки уничтожения клонированными опухолевыми антиген-специфичными CD8 + Т-клетками человека клеток меланомы человека, экспрессирующих родственный антиген (неопубликованные данные).
| [Другие продукты] Нуклеотид GenBank NCBI № | [Другие продукты] Основной регистрационный номер UniProt № | [Другие продукты] Связанный с UniProt номер доступа Официальное полное имя NCBI родственный белок теплового шока 80 Информация о белках NCBI родственный белок теплового шока 80 Название белка UniProt Родственный белок теплового шока 80 Комментарии UniProt для HSC80 Молекулярный шаперон, который способствует созреванию, поддержанию структуры и правильной регуляции специфических белков-мишеней, участвующих, например, в контроле клеточного цикла и передаче сигналов.Проходит функциональный цикл, связанный с его АТФазной активностью. Этот цикл, вероятно, индуцирует конформационные изменения в клиентских белках, тем самым вызывая их активацию. Динамически взаимодействует с различными ко-шаперонами, которые модулируют распознавание субстрата, АТФазный цикл и функцию шаперона (4). Меры предосторожности Все продукты MyBioSource предназначены для научных лабораторных исследований и не предназначены для диагностики, терапии, профилактики или использования in vivo. Покупая, вы прямо заявляете и гарантируете MyBioSource, что будете должным образом тестировать и использовать любые Продукты, приобретенные у MyBioSource, в соответствии с отраслевыми стандартами.MyBioSource и его авторизованные дистрибьюторы оставляют за собой право отказать в обработке любого заказа, если мы обоснованно считаем, что предполагаемое использование выходит за рамки наших приемлемых правил. Отказ от ответственности Несмотря на то, что были предприняты все усилия для обеспечения точности информации, представленной в этом листе данных, MyBioSource не несет ответственности за какие-либо упущения или ошибки, содержащиеся в нем. MyBioSource оставляет за собой право вносить изменения в данное техническое описание в любое время без предварительного уведомления.Клиент обязан сообщить MyBioSource о проблемах с производительностью продукта в течение 30 дней с момента получения продукта. Пожалуйста, посетите нашу страницу «Условия и положения» для получения дополнительной информации. Пути, связанные с антителом против HSC80Органы/ткани, связанные с антителом против HSC80 |
Комплекс нового ингибитора протеаз, гомолога овостатина, с родственными ему протеазами в просветной жидкости матки неполовозрелых мышей глутатион (GSH), тритон X-100, желатин, коллаген плаценты человека IV типа, коллаген I типа бычьей кожи, мышиный альбумин, 1-10-фенантролин, йодацетамид, пепстатин, безамидин, химостатин и лейпептин были приобретены у Sigma-Aldrich (St. .Луис, Миссури, США). Sephacryl S-400, DEAE-Sepharose, глутатион-сефароза 4B, CNBr-активированная Sepharose 4B, мембрана из поливинилидендифторида (PVDF) и pGEX-4T-1 были заказаны в GE Healthcare Life Sciences (Piscataway, NJ, USA). Рестриктазы
Bam HI и Eco RI были заказаны в New England BioLabs, Inc. (Ипсвич, Массачусетс, США). Антитело против мышиного CLCA3 было приобретено у Abcam (Кембридж, Великобритания).Заявление об этике
Самки новозеландских белых кроликов были приобретены в Научно-исследовательском институте животноводства Совета по сельскому хозяйству (Тайнань, Тайвань).Процедуры ухода, содержания и экспериментов с кроликами и мышами были рассмотрены и одобрены Комитетом Института по уходу и использованию животных (IACUC) Университета Фуинь. Животных содержали в соответствии с институциональными рекомендациями по уходу и использованию экспериментальных животных.
Подготовка неполовозрелых мышей ULF
Неполовозрелым самкам мышей ICR (возраст 21 день) вводили подкожно суточную дозу DES (1 мкг), растворенного в кукурузном масле, соответственно, в течение четырех дней подряд.Мышей забивали на 25-й день возраста. Затем из просвета матки мышей собирали УНЧ. Раствор УНЧ, содержащий 2 мМ ЭДТА, центрифугировали при 10000× г в течение 2 мин для удаления нерастворимых компонентов, а затем хранили при –70°С до дальнейшего анализа.
Зимография субстрата
Обозначенные образцы смешивали с невосстанавливающим буфером для загрузки образцов, а затем подвергали SDS-PAGE анализу с 12% полиакриламидными гелями, содержащими желатин (2 мг/мл), человеческий плацентарный коллаген IV типа (1 мг/мл ), или коллаген I типа бычьей кожи (1 мг/мл).После электрофореза срезы геля промывали буфером (0,1 М Трис-HCl, рН 7,4, содержащий 2,5% Тритона Х-100) два раза при комнатной температуре по 18 минут, а затем инкубировали в 0,1 М Трис-HCl (рН 7,4). при 37 °С. После инкубации в течение ночи гели окрашивали кумасси бриллиантовым синим в течение 20 минут с последующим обесцвечиванием в 10% растворе уксусной кислоты. Очищенные гели сканировали фотосканером HP Scanjet G3010. Желатинолитическую активность определяли по светлым зонам, указывающим на деградацию субстрата, и количественно определяли путем измерения интенсивности площади указанной полосы за вычетом идентичной фоновой интенсивности той же дорожки с использованием программного обеспечения ImageJ.
Выделение белков
1 мл УЛФ фракционировали методом гель-фильтрационной хроматографии на колонке с сефакрилом S-400 (15 мм × 100 см), проявляли буфером PBS, содержащим 2 мМ ЭДТА, со скоростью потока 0,2 мл/мин на холоду. номер. Каждую фракцию собирали по 3,2 мл. Аликвоты (0,3 мл) указанных фракций диализовали против воды, лиофилизировали и наносили на желатиновую зимографию для определения активности. Объединенные фракции (140 мл), содержащие желатинолитический фермент 26 кДа, собранные при хроматографии на Сефакриле S-400, диализовали, лиофилизировали, затем растворяли в 20 мл уравновешенного буфера А (5 мМ фосфатный буфер, содержащий 2 мМ ЭДТА при рН 7.4) и загружали в колонку с ДЭАЭ-Сефарозой (15 мм × 2 см). После промывки десятью объемами буфера А колонку обрабатывали линейным градиентом от 75 мл буфера А до 75 мл буфера А плюс 0,4 М NaCl. Каждая фракция собирала 3,6 мл. 0,25 мл, взятые из указанных фракций, диализовали, лиофилизировали, а затем подвергали желатиновой зимографии для определения активности. Фракции, содержащие желатинолитический фермент 26 кДа, объединяли, подвергали диализу против воды для удаления избытка реагентов, лиофилизировали и использовали в качестве антигена для получения кроличьих антител против mOH.
Масс-спектрометрия
Очищенный желатинолитический ферментный комплекс анализировали с помощью SDS-PAGE, а затем окрашивали кумасси бриллиантовым синим. Назначенные полосы вырезали из геля, расщепляли трипсином и анализировали жидкостной хроматографией-ионизацией электрораспылением в сочетании с тандемной масс-спектрометрией (ЖХ-МС/МС) соответственно.
Экспрессия и очистка слитого белка GST-mOH[870–940]
кДНК частичной последовательности mOH (остатки 870–940 аминокислот) амплифицировали из маток неполовозрелых мышей, получавших DES, с помощью ОТ-ПЦР с использованием праймеров ( 5′-CTC GGA TCC ACT GTA GTG GCT ACA TCC-3′ и 5′-CTC GAA TTC TCA ACC TTC TAC TAC ATT GCT -3′).Полученный продукт расщепляли с помощью Bam HI/ Eco RI и субклонировали в pGEX 4T-1 с получением вектора pGEX 4T-1-mOH [870–940]. штаммов E. coli BL21 (DE 3), содержащих вектор pGEX-4T-1-mOH[870–940], выращивали в 1 л бульона Луриа-Бертани (LB), содержащего ампициллин (100 мкг/мл), при 37 °C. Пока поглощение при 600 нм не достигло приблизительно 0,4, для индукции экспрессии белка добавляли изопропил-β-D-1-тиогалактопиранозид (конечная концентрация 0,4 мМ). Через четыре часа бактерии собирали центрифугированием при 6000 × г в течение 15 минут.Клетки в осадке ресуспендировали в буфере (PBS, содержащем 1 мкг/мл лизоцима, 2 мМ EDTA и 1 мМ PMSF), инкубировали в течение 1 часа при 37 °C, а затем разрывали тремя циклами замораживания и оттаивания. Лизат обрабатывали ДНКазой I (1 мкг/мл) в течение 30 мин при 37°С с последующим центрифугированием при 6000× г при 4°С в течение 30 мин. Супернатант загружали в колонку, заполненную смолой Glutathione Sepharose 4B. Несвязавшиеся белки удаляли промыванием PBS. Слитый белок GST-mOH[870–940] элюировали PBS, содержащим 10 мМ GSH.Элюат подвергали диализу против воды для удаления избытка реагентов, лиофилизировали и хранили для получения антитела mOH[870–940]. Антитело GST-mOH[870–940] распознает только восстановленную форму mOH.
Получение очищенного mOH-антитела (POHA)
ULF очищали гель-фильтрацией с использованием колонки Sephacryl S-400 (15 мм × 100 см), разработанной PBS. Каждая фракция собирала 3,2 мл. Основные белки mCLCA3, mOH и C3 в УНЧ элюировали и собирали во фракциях 26–30, 32–35 и 38–40 соответственно.Фракции, содержащие mCLAC3, mOH и C3, объединяли соответственно, подвергали диализу против дистиллированной воды и лиофилизировали. МОН, очищенный от УНЧ (15 мл) с использованием хроматографов Sephacryl S-400 и DEAE-Sepharose, конъюгировали с 1,0 мл гранул CNBr-активированной сефарозы в соответствии с инструкциями производителя. Этот очищенный mOH также содержит C3, альбумин и mCLCA3. Таким образом, анти-С3, анти-альбумин и анти-mCLAC3 антитела в антителах, выращенных с помощью mOH, удаляли путем абсорбции гранулами сефарозы, конъюгированными с С3, альбумином и mCLCA3.По одному миллиграмму С3, mCLCA3 и мышиного альбумина конъюгировали с 1,5 мл гранул сефарозы, активированной CNBr, в соответствии с инструкциями производителя. Кроличье антитело против mOH (0,5 мл) применяли для аффинной хроматографии на колонке, заполненной шариками, конъюгированными с mOH. После нескольких промывок антитело элюировали 0,1 М раствором глицина (рН 3) и немедленно нейтрализовали добавлением 1 М трис-HCl (рН 8). Элюированные пулы (6,0 мл) затем инкубировали с абсорбированными гранулами в течение 1 часа.6,0 мл очищенного антитела mOH (POHA) собирали из несвязанного раствора для элюирования и затем хранили при -70 °C. POHA может распознавать нативную форму mOH в нашем анализе.
Вестерн-блоттинг
Обозначенные образцы разделяли с помощью SDS-PAGE с полиакриламидным гелем с линейным градиентом (4–15%), с последующим электроблоттингом на мембране PVDF, зондировали POHA (250 мкл), мОН [870–940 ] антисыворотка (разведение 1:2000) или антитело mCLCA 3 (разведение 1:2000).
Анализ совместного фракционирования
1 мл ULF фракционировали с помощью хроматографии на сефакриле S-400, как описано выше.Аликвоты (0,30 мл) указанных фракций (21–47) из гель-фильтрационной хроматографии разделяли с помощью SDS-PAGE, а затем применяли для вестерн-блоттинга, определяли невосстановленный mOH с использованием POHA или восстановленный mCLCA3 с использованием кроличьих антител mCLCA 3. Кроме того, идентичные образцы подвергали желатиновой зимографии для обнаружения желатинолитических ферментов.
Ко-иммунопреципитационный анализ
Аликвоту (60 мкл) гранул протеина А-Сефарозы инкубировали либо с 5 мкг преиммунного антитела (PIA), либо с 5 мкл POHA, вращали при комнатной температуре в течение одного часа, а затем промывали 0.1 мл PBS три раза. Ко всем компонентам добавляли 30 мкл УНЧ и затем вращали при комнатной температуре в течение одного часа. Гранулы трижды промывали 0,1 мл PBS с последующим центрифугированием. Осажденные белки повторно суспендировали с двумя объемами буфера для загрузки геля SDS (30 мкл). Затем аликвоты (6 мкл) иммунопреципитированных белков подвергали желатиновой зимографии.
Анализ ингибирования
Аликвоту (0,3 мл) комплекса желатинолитических ферментов, выделенного из хроматографии Sephacryl S-400, анализировали методом зимографии желатина в присутствии различных белковых ингибиторов, включая 1–10-фенантролин (1 мМ), йодацетамид (0 .1 мМ), пепстатин (20 мкМ), безамидин (2 мМ), химостатин (100 мкМ) или лейпептин (100 мкМ).
Определение оптимального рН для активности желатинолитических ферментов
Частично очищенный комплекс желатинолитических ферментов, полученный хроматографией на Сефакриле S-400, подвергали желатинзимографии. Каждую дорожку геля вырезали и промывали буфером (0,1 М Трис-HCl, рН 7,4, содержащий 2,5% Тритон Х-100) два раза при комнатной температуре в течение 18 минут, а затем инкубировали в буферах с различными значениями рН соответственно.Буферы включали 0,1 М цитрата (рН 5,0 и 6,0), 0,1 М Трис (7,4 и 8,0) или 0,1 М глицина (рН 9,0, 10,0 и 10,5). После инкубации при 37°С в течение ночи нарезанные гели окрашивали кумасси бриллиантовым синим, а затем обесцвечивали 10% уксусной кислотой.
Анализ деградации коллагена типа I
6 мкг комплекса mOH-протеазы после очистки разделяли в 12% полиакриламидном геле с SDS в невосстанавливающих условиях. После электрофореза гель промывали буфером (0.1M Tris-HCl, pH 7,4, содержащий 2,5% Triton X-100) при комнатной температуре в течение 15 минут три раза для удаления SDS. Белковые полосы с кажущейся молекулярной массой 26 и 23/22 кДа соответственно вырезали, измельчали и вращательно инкубировали со 100 мкл коллагена I типа (1 мг/мл). После инкубации в течение ночи при 37 °C аликвоты (8 мкл) супернатанта подвергали анализу с помощью SDS-PAGE с использованием полиакриламидного геля с линейным градиентом (4–15%).
Обратная транскрипция – полимеразная цепная реакция (ОТ-ПЦР)
ОТ-ПЦР использовали для определения распределения в тканях транскриптов мРНК mOH у мышей.Панель мультитканевой мРНК мыши была получена от Clontech Laboratories, Inc. (Маунтин-Вью, Калифорния, США). Реакцию обратной транскрипции проводили по инструкции производителя (Invitrogen, Гранд-Айленд, Нью-Йорк, США). Полученный продукт служил матрицей для ПЦР-амплификации транскрипта mOH с помощью праймеров (5′-GCT ACA CTG GAG CTC GTG AAA GC-3′ и 5′-GGT GCT TGA TCA GGC TCT TGG TAG-3′). GAPDH амплифицировали параллельно с праймерами (5′-ACC ACA GTC CAT GCC ATC AC-3′ и 5′-TCC ACC ACC CTG TTG CTG TA-3′) и служили внутренним контролем.
MP06-01 ИДЕНТИФИКАЦИЯ РОДСТВЕННЫХ ПРОКСИМАЛЬНЫХ ТИПОВ КЛЕТОК ПРОСТАТЫ МЫШИ И ЧЕЛОВЕКА И ИХ ОБОГАЩЕНИЕ ПРИ ДОБРОКАЧЕСТВЕННОЙ ГИПЕРПЛАЗИИ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ ЧЕЛОВЕКА
ВВЕДЕНИЕ И ЦЕЛЬ:
Доброкачественная гиперплазия предстательной железы (ДГПЖ) широко распространена у стареющих мужчин и представляет значительную бремя здравоохранения, связанное с лечением симптомов нижних мочевыводящих путей (СНМП). ДГПЖ/СНМП по-прежнему трудно поддаются лечению из-за фенотипической гетерогенности, в результате чего при необходимости хирургического вмешательства.Типы клеток, вызывающие рост ДГПЖ: Неизвестный. Анатомические исследования показали, что переходная зона, расположенная вблизи уретра является местом роста ДГПЖ. Ранее мы идентифицировали два новых типа клеток, булавой и бугорком, которые обогащаются в предстательной уретре и проксимальных протоках. Здесь, мы идентифицируем новый проксимальный фибробласт у человека, а также родственный бугорковый эпителий и проксимальные фибробласты у мышей. Мы оцениваем вклад каждого проксимального тип клеток при ДГПЖ человека, установив новую парадигму эпителия уретры и проксимальных фибробласты в дискретных фенотипах ДГПЖ.
МЕТОДЫ:
Мы использовали беспристрастный подход с помощью секвенирования РНК отдельных клеток (scRNA-seq) для идентификации родственные типы клеток предстательной железы мыши и человека и разработать новую проточную цитометрию и IHC панели антител для очистки и локализации каждого типа клеток. Частота проксимальных фибробласты и уретральный эпителий при ДГПЖ по сравнению с нормальной простатой человека оценивали с помощью scRNA-seq, проточная цитометрия и иммунофлуоресценция.
РЕЗУЛЬТАТОВ:
Бугорчатые и булавовидные клетки устанавливаются рано, и эти клетки распространяются в проксимальные протоки переходной зоны предстательной железы взрослого человека.Проксимальные фибробласты окружают мочеиспускательный канал и проксимальные протоки. У пациентов с ДГПЖ увеличены булавовидные и бугорчатые клетки. внутри железистых узелков по сравнению с нормальной тканью предстательной железы и проксимальными фибробластами увеличиваются в зонах периуретрального фиброза. Данные scRNA-seq и IHC мыши подтверждены наличие родственных проксимальных фибробластов и бугоркового эпителия, но булавовидных клеток не были найдены.
ВЫВОДЫ:
Наши результаты показывают, что булавовидный эпителий уретры и проксимальных протоков образуются до зачатия предстательной железы и обогащены железистыми узелками ДГПЖ, предполагая потенциальное клеточное происхождение нового роста простаты.Мы также выявляем проксимальный фибробласт человека как клеточный источник отложения коллагена в фиброз предстательной железы, создающий новый фенотип для пациентов с СНМП. Мы создали клеточный атлас уретры и проксимального отдела предстательной железы мышей, который позволит для создания специализированных моделей мышей для отслеживания уретральных и проксимальных фибробластов родословная. Типы клеток проксимальных протоков могут стать новыми мишенями для лечение ДГПЖ/СНМП.
Источник финансирования:
R01 ДК115477
Нейропептид S способствует обонятельной функции мышей за счет активации экспрессирующих родственные рецепторы нейронов в обонятельной коре
Abstract
Нейропептид S (НПС) представляет собой недавно обнаруженный нейромодулятор, расположенный в стволе головного мозга и регулирующий различные биологические функции путем избирательной активации рецепторов НПВ (НПСР). Высокий уровень экспрессии мРНК NPSR в обонятельной коре предполагает, что система NPS-NPSR может участвовать в регуляции обонятельной функции.Настоящее исследование было предпринято для изучения влияния интрацеребровентрикулярной (i.c.v.) инъекции НПВ или совместной инъекции антагониста NPSR на обонятельное поведение, потребление пищи и экспрессию c-Fos в обонятельной коре у мышей. Кроме того, двойная иммунофлуоресценция использовалась для идентификации NPS-индуцированных Fos-иммунореактивных (-ir) нейронов, которые также несут NPSR. НПВ (0,1–1 нмоль) в.ц.в. инъекция значительно уменьшила латентный период для поиска закопанной пищи и увеличила обонятельную дифференциацию различных запахов и общее время, затрачиваемое на обнюхивание в задачах обонятельного привыкания / избавления от привычки.НПС больше всего улучшал обонятельную способность в дозе 0,5 нмоль, которую можно было заблокировать совместным введением 40 нмоль антагониста НПСР [D-Val 5 ]НПС. Введение НПВ дозозависимо ингибировало потребление пищи голодающими мышами. Иммуногистохимия ex-vivo c-Fos и NPSR в обонятельной коре показала, что по сравнению с мышами, получавшими носитель, NPS заметно усиливал экспрессию c-Fos в переднем обонятельном ядре (AON), грушевидной коре (Pir), вентральной тени. tecta (VTT), переднее корковое миндалевидное ядро (ACo) и латеральная энторинальная кора (LEnt).Процент нейронов Fos-ir, которые также экспрессируют NPSR, составил 88,5% и 98,1% в AON и Pir соответственно. Настоящие результаты показали, что NPS посредством избирательной активации нейронов, несущих NPSR в обонятельной коре, облегчает обонятельную функцию у мышей.
Образец цитирования: Shao Y-F, Zhao P, Dong C-Y, Li J, Kong X-P, Wang H-L, et al. (2013) Нейропептид S облегчает обонятельную функцию мышей за счет активации нейронов, экспрессирующих родственные рецепторы, в обонятельной коре.ПЛОС ОДИН 8(4): е62089. https://doi.org/10.1371/journal.pone.0062089
Редактор: Пранела Рамешвар, Университет медицины и стоматологии Нью-Джерси, США
Получено: 30 января 2013 г.; Принято: 16 марта 2013 г.; Опубликовано: 16 апреля 2013 г.
Авторское право: © 2013 Shao, et al. Это статья с открытым доступом, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Работа выполнена при поддержке грантов Национального фонда естественных наук Китая (№ 81171254, 81071076 и 30670677), а также гранта Ключевой национальной научно-технической программы «Крупная разработка новых лекарств» Министерства науки и Технология Китая (2009ZX09503-017). Спонсоры не участвовали в разработке исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Нейропептид S (НПС) — недавно обнаруженный нейромодулятор, расположенный в стволе головного мозга. NPS избирательно связывается с высоким сродством к рецепторам, связанным с белками Gs и Gq, ранее идентифицированными как GPR 154 и теперь обозначаемыми как NPSR, вызывая мобилизацию внутриклеточного Ca 2+ и повышая уровни цАМФ [1]. мРНК предшественника NPS у крыс экспрессируется в группе нейронов, расположенных между голубым пятном (LC) и ядром Баррингтона, главным сенсорным ядром тройничного нерва и латеральным парабрахиальным ядром [1].У мышей мРНК предшественника NPS экспрессируется только в ядре Kölliker-Fuse и перикоэррулярной области ствола мозга [2]. Напротив, мРНК NPSR широко распространена в мозге крыс и мышей, в основном в обонятельной коре, коре головного мозга, таламусе, гипоталамусе, миндалевидном теле и субкулуме [1]–[4].
Этот профиль экспрессии мРНК NPSR предполагает участие системы NPS-NPSR в регуляции множества центральных функций. Действительно, активация НПСР при центральном введении НПВ усиливает двигательную и исследовательскую активность, вызывает анксиолитические эффекты у мышей [1], [5], [6] и способствует бодрствованию у крыс [1], [7].NPS также участвует в антиноцицепции [8], [9], выражении и угасании страха [10] и процессах памяти у мышей [11], [12], а также способствует рецидиву поиска кокаина у крыс [13].
СистемаNPS-NPSR предлагается как недавно идентифицированная система регуляции обоняния, участвующая в регуляции обонятельного восприятия и/или интеграции обонятельной или феромонной информации [3], поскольку высокие уровни экспрессии мРНК NPSR были обнаружены во многих областях обонятельной коры. включая переднее обонятельное ядро (AON), грушевидную кору (Pir), tenia tecta (TT), а также переднее корковое миндалевидное ядро (ACo) и латеральную энторинальную кору (LEnt) у мышей [2].Эти области обонятельной коры непосредственно получают синаптический вход от обонятельной луковицы [14], [15] и, по-видимому, играют решающую роль в переводе характеристик вдыхаемых молекул в насыщенные, вызывающие эмоции и память окрашенные восприятия, называемые запахами [15]. ]. Однако неизвестно, как система NPS-NPSR регулирует обонятельное поведение.
Настоящее исследование было разработано для наблюдения за влиянием системы NPS-NPSR на обонятельную функцию у мышей после интрацеребровентрикулярной инъекции (т.в.) инъекции. Обонятельные способности у мышей оценивали с помощью теста зарытой пищи (для оценки способности обнаруживать летучие запахи) и теста обонятельного привыкания/отторжения (для оценки способности обнаруживать и различать одинаковые и разные запахи). Тест на потребление пищи использовали для выяснения взаимосвязи между обонянием и приемом пищи у мышей после i.c.v. администрация НПС. Для дальнейшей идентификации потенциальных нейронных мишеней NPS в обонятельной коре NPS-индуцированные Fos-иммунореактивные (-ir) нейроны были проанализированы с использованием иммуногистохимии ex vivo , а наличие NPSR в этих нейронах было исследовано с помощью двойной иммунофлуоресцентной микроскопии.
Материалы и методы
Животные и хирургическая имплантация
взрослых самца мышей C57BL/6J (возраст 6 недель) были приобретены в Центре экспериментальных животных Университета Ланьчжоу (Ланьчжоу, КНР). Их содержали при температуре окружающей среды (22±1°C) и относительной влажности 50% при автоматически регулируемом цикле свет/темнота 12:12 (свет включен с 8:00 до 20:00, интенсивность освещения ≈ 100 лк). Еда и вода были доступны вволю за исключением периода голодания.Каждое животное использовали только один раз для межгрупповых сравнений в тесте на зарытую пищу, в тесте на привыкание и отвыкание от обоняния и в тесте на потребление пищи. За всеми животными ухаживали, и эксперименты проводились в соответствии с рекомендациями Европейского сообщества по использованию экспериментальных животных (86/609/EEC). Экспериментальный протокол был одобрен Комитетом по этике Университета Ланьчжоу (номер разрешения: SCXK Gan 2009–0004).
Под наркозом хлоралгидратом (350 мг/кг, т.е.р.), мышей помещали в стереотаксический аппарат. Направляющую канюлю из нержавеющей стали (калибр 25) стереотаксически имплантировали над правым боковым желудочком (AP -0,2 мм, ML +1,0 мм, DV -1,4 мм, согласно атласам Paxinos and Franklin, 2001 [16]) для и.к.в. инъекция. Канюля хронически фиксировалась к черепу стоматологическим цементом. В направляющую канюлю вставляли постоянный стилет из нержавеющей стали (размер 32) для предотвращения окклюзии.
Администрации лекарственных средств
NPS (мышь, Ser-Phe-Arg-Asn-Gly-Val-Gly-Ser-Gly-Ala-Lys-Lys-Thr-Ser-Phe-Arg-Arg-Ala-Lys-Gln) и [D-Val 5 ]НПВ (человек, Ser-Phe-Arg-Asn-D-Val-Val-Gly-Thr-Gly-Met-Lys-Lys-Thr-Ser-Phe-Gln-Arg-Ala-Lys-Ser) , подарки от проф.Rui Wang, были синтезированы на кафедре биохимии и молекулярной биологии Школы наук о жизни Ланьчжоуского университета [7], [9], [17]. Свежий НПС (0,1-1 нмоль) и НПС (0,5 нмоль) + [D-Val 5 ]НПС (20 или 40 нмоль) растворяли в 1 мкл физиологического раствора. Препараты и носитель (физиологический раствор) вводили через насаженную направляющую канюлю со скоростью потока 1 мкл/мин в 17:00 дня исследования.
По окончании эксперимента мышам вводили и.ц.в. через направляющую канюлю вводили по 1 мкл красителя метиленового синего и через 5 мин декапитировали под глубокой анестезией хлоралгидратом натрия.Мозги были удалены и заморожены. Макродиссекцию головного мозга использовали для проверки места введения лекарств или носителя. Использовались только данные по животным с дисперсией красителя через желудочек.
Тесты обонятельного поведения
Тест зарытой пищи.
Тест зарытой пищи проводили, как ранее описано Янгом и Кроули [18]. Вкратце, после 7 дней восстановления после операции мышей не кормили в течение 32 часов, начиная с 9:00, с доступной водой.В день тестирования каждая мышь получала i.c.v. инъекции носителя, НПВ или НПВ + [D-Val 5 ]НПВ, а затем помещали в испытательную камеру из плексигласа (46 см Д × 23,5 см Ш × 20 см В), содержащую чистые подстилки глубиной 3 см из свежестерилизованной и дезодорированной древесной щепы. После адаптации к окружающей среде в течение 15 минут мышь удаляли из камеры и гранулы корма для мышей (1,5 г, Beijing keaoxieli feedstuff Co. Ltd.) случайным образом закапывали на 1 см ниже поверхности подстилки. Затем мышь помещали обратно в камеру и измеряли время ожидания закопанной пищи.Латентный период определяли как время от момента помещения мыши в тестовую камеру до момента, когда она открывала и хватала пищу передними лапами и/или зубами [19]. После каждого опыта испытательные камеры промывали дистиллированной водой и высушивали на воздухе. Постельное белье меняли перед каждым испытанием. Животных записывали на видео, оценивали и анализировали исследователи, которые не знали о вводимых препаратах.
Тест на привыкание и отвыкание от обоняния.
Тест на привыкание и отвыкание от обоняния проводили в соответствии с предыдущим описанием Янга и Кроули [18].В день тестирования каждую мышь помещали в тестовую камеру (26 см Д × 12 см Ш × 12 см В), содержащую подстилку из чистой свежей древесной стружки толщиной 1 см, и адаптировали к условиям тестирования, в которых была чистая, Сухой аппликатор с ватным наконечником вводили на глубину 4 см через отверстие в крышке на 15 мин после внутривенного введения. инъекции наркотиков или транспортного средства. Обнюхивание определяли, когда мышь ориентировалась на кончик замененного аппликатора, впитанного дистиллированной водой, экстрактом миндаля (разведение 1:100, Supercook, Лидс, Великобритания) или экстрактом ванили (разведение 1:100, Supercook, Лидс, Великобритания). ) носом в пределах 2 см или ближе к кончику.Обнюхивание регистрировали в течение 2 минут в каждом испытании. Последовательность замены аппликатора: вода, вода, вода, миндаль, миндаль, миндаль, ваниль, ваниль и ваниль с интервалом в 1 мин. Привыкание определяется как прогрессивное уменьшение времени обонятельного обнюхивания в сторону повторного предъявления одного и того же запахового стимула и используется для оценки того, может ли животное различать один и тот же запах. Отказ от привычек определяется восстановлением обнюхивания при появлении нового запаха и используется для оценки того, может ли животное обнаружить другой запах [20]–[22].Испытательные камеры промывали дистиллированной водой и меняли подстилку перед каждым испытанием. Животных записывали на видео, и экспериментатор, не имевший отношения к лечению, анализировал поведение животных.
Тест приема пищи
Хорошо известно, что обоняние тесно связано с приемом пищи у млекопитающих, особенно у грызунов [23], [24]. Чтобы определить, влияет ли изменение обоняния после введения НПВ на прием пищи, количество потребляемой пищи было соответственно измерено как 0.5, 1, 2, 4 и 24 часа после центрального введения носителя (n = 10) или NPS (0,5 или 1 нмоль, n = 9 в каждой группе) у мышей, голодавших в течение 32 часов.
Иммуногистохимия
Подготовка ткани.
Через полтора часа после введения НПВ (0,5 нмоль, n = 4) или носителя 1 мкл (n = 5) в.ц.в. введения животных анестезировали избыточной дозой хлоралгидрата (400 мг/кг) и перфузировали через восходящую аорту 30 мл физиологического раствора, содержащего гепарин (1 ЕД/мл), а затем 4% параформальдегида в 0.1 М фосфатный буфер (ФБ). Мозг извлекали, затем фиксировали в том же фиксаторе в течение ночи и погружали в 30% раствор сахарозы в 0,1 М ФБ при 4°C на 36 ч и делали корональные срезы (30 мкм) на криостате (CM1900, Leica Micro-systems, Heidelberg). , Германия) при -20°C и срезы собирали в 0,01 М натрий-фосфатный буфер (PBS).
Фос иммуногистохимия.
Плавающие срезы промывали 0,01 М раствором PBS (pH 7,4), обрабатывали в течение 30 минут 0,3% H 2 O 2 в PBS и инкубировали в блокирующем растворе (10% бычьей сыворотки в PBS) в течение 1 часа.Затем срезы инкубировали с кроличьим поликлональным антителом против c-Fos (1∶5000, sc-253, Santa Cruz Biotechnology, Санта-Крус, Калифорния, США), разведенным в PBS, содержащем 1% бычьей сыворотки, в течение 48 ч при 4°C на агитатор. После промывания в PBS срезы инкубировали с биотинилированным козьим антикроличьим IgG (1∶1000, AP132B, Millipore, Темекула, Калифорния, США), а затем со стрептавидином, конъюгированным с пероксидазой хрена (1∶2000, SA202, Millipore, Темекула, Калифорния, США). США). Обе инкубации проводили на мешалке при 4°С в течение ночи.После промывания срезы погружали в 0,05 М трис-HCl-буфер, рН 7,6, содержащий 0,05% 3,3′-диаминобензидина (ДАБ), 0,01% H 2 O 2 и 0,6% сульфата аммония никеля для 2– 5 мин при комнатной температуре. Наконец, срезы помещали на покрытые желатином предметные стекла, обрабатывали контрастным окрашиванием нейтральным красным, сушили, обезвоживали и накрывали покровным стеклом с использованием DPX для световой микроскопии.
Двойная иммунофлуоресценция для Fos и NPSR.
Эти срезы инкубировали со смешанным раствором, содержащим кроличьи поликлональные антитела против c-Fos и козьи анти-NPSR (1∶1,000, sc-162893, Santa Cruz Biotechnology, Санта-Крус, Калифорния, США), разведенные в PBS, содержащем 1% бычьего сыворотки в течение 48 ч при 4°С на мешалке после инкубации в 10% бычьей сыворотке в PBS.Специфичность антитела против NPSR была продемонстрирована в предыдущих исследованиях [25], [26]. После нескольких промываний в PBS срезы инкубировали с Alexa Fluor® 488-конъюгированным affinipure ослиным IgG против IgG кролика (1∶200, 711-545-152, Jackson ImmunoResearch Laboratories, Inc., Пенсильвания, США) и CyTM 3-конъюгированным affinipure. ослиный антикозий IgG (1∶200, 705-165-147, Jackson ImmunoResearch Laboratories, Inc., PA, USA) в течение 2 ч при 37°C. Наконец, срезы помещали на предметные стекла, накрывали покровным стеклом, используя 90% глицерин в PBS, и исследовали под флуоресцентным микроскопом.
Анализ данных
Подсчет клеток.
Fos-ir нейроны в AON (Брегма 1,98 мм, рис. 1), Pir (Брегма 0,62 мм, рис. 1), VTT (Брегма 2,34 мм), ACo (Брегма -0,94 мм) и LEnt (Брегма — 3,40 мм) подсчитывали двусторонне для каждого животного, получавшего НПВ или носитель. Рассчитывали среднее значение для двух сторон.
Рис. 1. Схематические рисунки показывают локализацию секций, используемых для подсчета нейронов Fos-ir.
Серые зоны представляют AON (Брегма 1.98 мм) и Пир (Брегма 0,62 мм). Сокращения: АОН — переднее обонятельное ядро; Пир, грушевидная кора.
https://doi.org/10.1371/journal.pone.0062089.g001
Статистический анализ.
Значения были выражены как среднее значение ± стандартная ошибка среднего. Данные теста зарытой пищи, теста потребления пищи и общего времени обнюхивания, затраченного на задачи обонятельного привыкания и отказа от привычки, были проанализированы с использованием однофакторного дисперсионного анализа (ANOVA) и теста наименьшей значимой разницы Фишера (LSD).Данные теста привыкания и отказа от обоняния были проанализированы с использованием внутригруппового дисперсионного анализа с повторными измерениями, а затем с помощью тестов Ньюмана-Кеулса. Количество нейронов Fos-ir между мышами, получавшими NPS и носитель, анализировали с использованием независимого t-критерия Стьюдента. Во всех статистических сравнениях уровень значимости был установлен на уровне p <0,05.
Результаты
Влияние НПВ на обонятельные функции
Тест зарытой пищи.
По сравнению с мышами, получавшими носитель, т.е.резюме. введение 0,1, 0,5 и 1 нмоль НПВ достоверно снижало латентность поиска закопанной пищи с 73,43±11,77 с до 35,74±5,37 ( p <0,001), 12,72±1,34 ( p <0,001) и 24,61±5,04 с. с ( p <0,001) соответственно (рис. 2). Среди трех доз 0,5 нмоль NPS больше всего снижала латентный период ( p <0,001 и p <0,05 по сравнению с носителем и 0,1 нмоль NPS соответственно; рис. 2). Действительно, высокая доза (1 нмоль) НПС незначительно снижала латентность по сравнению с 0.1 нмоль НПС (35,74±5,37 с против 24,61±5,04 с, p = 0,24; рис. 2).
Рисунок 2. Задержка поиска закопанной пищи после i.c.v. инъекции носителя или НПВ мышам.
Значения средние ± SEM (n = 10 мышей в каждой группе). * р <0,05, ** р <0,001. Данные анализировали с помощью однофакторного дисперсионного анализа с последующим применением критерия LSD Фишера.
https://doi.org/10.1371/journal.pone.0062089.g002
Тест на привыкание и отвыкание от обоняния.
На рис. 3A-D обобщены результаты тестов привычного и ненормального обонятельного поведения у мышей, которым интрацеребровентрикулярно вводили носитель или НПВ (0,1, 0,5 или 1 нмоль) на один и тот же и разные запахи соответственно. Центральное введение носителя вызывало привыкание к воде ( p <0,05) и ванили ( p <0,001) и отвыкание от ванили ( p <0,001, рис. 3A). Мыши, которым вводили NPS в дозе 0,1 нмоль, были способны различать миндаль и ваниль как новые запахи, но не привыкали к запаху миндаля (рис.3Б). По сравнению с контролем с носителем, мыши привыкали и выходили из привычки ко всем тестовым запахам после введения 0,5 или 1 нмоль NPS (рис. 3C и D), что указывает на то, что NPS в этих дозах может облегчить мышам различение всех одинаковых и разных тестируемых запахов. Как показано на рис. 3E, NPS в зависимости от дозы увеличивал общее время обнюхивания, затрачиваемое на обонятельные поведенческие задачи привыкания и отказа от привычки.
Рис. 3. Тест на привыкание и отвыкание от обоняния после i.c.v. инъекции носителя или НПВ мышам.
A. Мыши, получавшие носитель, демонстрировали значительное привыкание к воде, отказ от миндаля к ванили и привыкание к ванили. B. NPS при 0,1 нмоль проявлял значительное привыкание к воде, отвыкание от воды к миндалю, отвыкание от миндаля к ванили и привыкание к ванили. C. NPS при 0,5 нмоль проявляли значительное привыкание к воде, отказ от воды к миндалю, привыкание к миндалю, отказ от миндаля к ванили и привыкание к ванили. D. NPS при 1 нмоль демонстрировал значительное привыкание к воде, отказ от воды к миндалю, привыкание к миндалю, отказ от миндаля к ванили и привыкание к ванили. E. НПВ в зависимости от дозы увеличивает общее время вдыхания, затрачиваемое на обонятельные задачи привыкания и избавления от привычки. Значения представляют собой средние значения ± стандартная ошибка среднего (n = 10 мышей в каждой группе). * p <0,05, ** p <0,01, *** p <0,001 для привыкания; # р <0.05, ## p <0,01, ### p <0,001 для отказа от привычки; данные были проанализированы с использованием внутригруппового дисперсионного анализа с повторными измерениями, а затем с помощью тестов Ньюмана-Кеулса. и р <0,01, && р <0,001; данные были проанализированы с помощью однофакторного дисперсионного анализа с последующим применением критерия LSD Фишера.
https://doi.org/10.1371/journal.pone.0062089.g003
Влияние НПВ на обонятельное поведение блокировалось [D-Val
5 ]НПВ.Чтобы определить, блокирует ли антагонист NPSR действие NPS на обонятельные способности, [D-Val 5 ]NPS, селективный антагонист NPSR [27], вводили мышам с 0,5 нмоль NPS (в/в) или без него.
Наши результаты показали, что 40 нмоль [D-Val 5 ]НПС значительно противодействуют эффекту 0,5 нмоль НПВ на латентный период для поиска спрятанной пищи (рис. 4). Однако при введении отдельно 40 нмоль [D-Val 5 ]НПС не влияли на латентный период по сравнению с носителем (рис.4).
Рисунок 4. Задержка поиска закопанной пищи после i.c.v. инъекции носителя, НПВ или [D-Val 5 ]НПВ и НПВ + [D-Val 5 ]НПВ мышам.
Значения средние ± SEM (n = 10 мышей в каждой группе). * р <0,05, ** р <0,001. Данные анализировали с помощью однофакторного дисперсионного анализа с последующим применением критерия LSD Фишера.
https://doi.org/10.1371/journal.pone.0062089.g004
Введение 20 нмоль [D-Val 5 ] НПВ значительно блокировало эффекты 0.5 нмоль NPS на обонятельную дифференцирующую способность (рис. 3C) в сторону воды и миндаля, но не ванили (рис. 5A). Кроме того, 40 нмоль [D-Val 5 ]НПВ полностью ингибировали влияние НПВ на обонятельное дифференцирующее поведение (рис. 5В) и заметно обращали вспять вызванное НПВ увеличение общего времени обнюхивания, затрачиваемого на задачи обонятельного привыкания и отвыкания (рис. 5С). ).
Рис. 5. Тест привыкания и отказа от обоняния у мышей после внутривенного введения. совместная инъекция НПВ и [D-Val 5 ]НПВ.
A. Введение 0,5 нмоль NPS + 20 нмоль [D-Val 5 ] NPS показало значительное привыкание к воде и отказ от привычки почти к ванили. B. Введение 0,5 нмоль NPS + 40 нмоль [D-Val 5 ]NPS вызывало значительное привыкание только к воде. C. Введение 0,5 нмоль NPS + 40 нмоль [D-Val 5 ]NPS значительно блокировало эффект NPS в отношении увеличения общего времени вдыхания в задачах привыкания к обонянию и отказа от привыкания.Значения представляют собой средние значения ± стандартная ошибка среднего (n = 10 мышей в каждой группе). * p <0,001 для привыкания; # p <0,05 для отказа от привычки; данные были проанализированы с использованием внутригруппового дисперсионного анализа с повторными измерениями, а затем с помощью тестов Ньюмана-Кеулса. и p <0,001, данные анализировали с помощью однофакторного дисперсионного анализа с последующим применением критерия LSD Фишера.
https://doi.org/10.1371/journal.pone.0062089.g005
Ингибирующее действие НПВ на потребление пищи
Рис.6A и B суммированы эффекты NPS (i.c.v.) на кумулятивное и рассчитанное по времени потребление пищи через 0,5, 1, 2, 4 и 24 часа после лечения у голодных мышей. В течение первых получаса 0,5 и 1 нмоль НПВ дозозависимо ингибировали потребление пищи по сравнению с внутривенным введением. мышей, получавших носитель ( p <0,01 и p <0,001 соответственно; фиг. 6A и B). По сравнению с носителем, у мышей, получавших NPS, наблюдалось значительное снижение потребления пищи в период 0-0,5 ч в течение 24-часового периода (фиг. 6). Однако в течение второго часа после лечения мыши, которым вводили НПВ, ели значительно больше, чем контрольная группа ( p <0.01; Рис. 6Б).
Рисунок 6. Влияние НПВ на кумулятивное потребление пищи (А) и потребление в определенные периоды времени (В) у голодающих мышей.
Носитель, 0,5 или 1 нмоль НПВ вводили внутривенно. вводили мышам, голодавшим в течение 32 часов. Значения представляют собой средние значения ± стандартная ошибка среднего (n = 10 мышей в группе с носителем, n = 9 мышей в каждой группе NPS). * р <0,05, ** р <0,01, *** р <0,001. Данные анализировали с помощью однофакторного дисперсионного анализа с последующим применением критерия LSD Фишера.
https://дои.org/10.1371/journal.pone.0062089.g006
NPS-индуцированное мечение c-Fos в обонятельной коре
В обонятельной коре центральное введение NPS (0,5 нмоль) индуцировало большое количество Fos-ir нейронов в AON (рис. 7A) и Pir (рис. 7C) и умеренное количество Fos-ir нейронов в VTT , ACo и LEnt (Таблица 1), и цифры были значительно больше, чем при впрыске носителя. В настоящем исследовании мы сосредоточились на областях AON и Pir, поскольку хорошо известно, что они играют ключевую роль в обонятельной функции и регуляции [28].По сравнению с мышами, получавшими носитель, NPS значительно увеличивал количество нейронов Fos-ir в 13,2 раза (2236 ± 199 против 169 ± 15) в AON (рис. 7E) и в 8,6 раз (1113 ± 49 против 169 ± 15). 128±16) в Пире (рис. 7F). Кроме того, NPS также индуцировал увеличение количества Fos-ir нейронов в моторной и соматосенсорной коре, миндалевидном теле, околоводопроводном сером, туберомаммилярном ядре, аркуатном гипоталамическом ядре, перифорникальном ядре и латеральной гипоталамической области (данные не показаны).
Рис. 7.Эффекты i.c.v. инъекция NPS на иммунореактивность Fos в AON и Pir у мыши. На микрофотографиях
AD показаны нейроны Fos-ir (черные) в AON и Pir у мышей, получавших NPS и носитель, соответственно. E, F. Гистограммы показывают количественный анализ количества Fos-ir нейронов в AON и Pir после NPS (n = 4 мыши) и носителя (n = 5 мышей) i.c.v. инъекция. Значения являются средними значениями ± SEM. * р <0,001. Данные анализировали с помощью независимого t-критерия Стьюдента.Бар = 100 мкм. Сокращения: aci, передняя спайка, внутрибульбарная часть; AON, переднее обонятельное ядро; вот, латеральный обонятельный тракт; Пир, грушевидная кора; OV, обонятельный желудочек.
https://doi.org/10.1371/journal.pone.0062089.g007
NPS-индуцированные Fos-ir нейроны в обонятельной коре экспрессируют NPSR
Определить, экспрессируют ли NPS-индуцированные Fos-ir нейроны в AON и Pir NPSR. Было проведено окрашивание Fos-ir в сочетании с окрашиванием NPSR-ir.Как показано на рис. 8, процент нейронов Fos-ir, которые также демонстрируют окрашивание для NPSR, составлял 88,5 ± 1,1% в AON (рис. 8A-C) и 98,1 ± 0,4% в Pir (рис. 8D-F). , соответственно.
Рис. 8. NPS-индуцированные нейроны Fos-ir, несущие NPSR в AON и Pir.
На микрофотографиях показаны нейроны Fos-ir в AON (A) и Pir (D) после NPS i.c.v. введения, NPSR-ir нейронов в AON (B) и Pir (E), и коэкспрессии Fos-ir и NPSR-ir нейронов в AON (C) и Pir (F) соответственно.Стрелки (C и F) показывают коэкспрессию нейронов Fos-ir и NPSR-ir. Бар = 50 мкм.
https://doi.org/10.1371/journal.pone.0062089.g008
Обсуждение
Настоящее исследование впервые продемонстрировало, что i.c.v. введение мышам NPS улучшало обонятельные способности, уменьшая латентный период для поиска закопанной пищи и усиливая обонятельную дифференциацию различных запахов (рис. 2–3). Среди трех доз 0,5 нмоль NPS, вероятно, уже максимально активирует его мозговые мишени, потому что в этой дозе NPS больше всего сокращает латентный период для поиска закопанной пищи и значительно способствует отвыканию и привыканию ко всем тестируемым запахам.Эти результаты показывают, что НПВ могут улучшать способность ощущать летучие запахи, а также обнаруживать и различать различные запахи. Было показано, что несколько классических нейротрансмиттеров, происходящих из области ствола мозга, например, норадренергическое ядро голубого пятна [29]–[32] и серотонинергическое ядро шва [33], модулируют обонятельное поведение. Кроме того, некоторые нейропептиды, такие как пептиды, родственные тахикинину, короткий нейропептид F и FMRFамид, также участвуют в модуляции обонятельной функции [34].Кларк и др. сообщили, что NPS происходит из ядра Kölliker-Fuse и перикоэрулярной области ствола мозга мыши и что мРНК NPSR сильно экспрессируется в обонятельной коре [2]. Взятые вместе, проекции NPS из ядра Kölliker-Fuse и перикоэрулярной области ствола мозга мышей в обонятельную кору могут обеспечивать механистическую основу для регуляции ею обонятельной функции.
Обоняние имеет большое значение для выживания млекопитающих и влияет на различные виды социальной деятельности, включая распознавание, выбор партнера, реакцию страха на запах хищника и потребление пищи, особенно у грызунов [23], [24].В настоящем исследовании, когда 0,5 или 1 нмоль NPS вводили соответственно i.c.v. при введении мышам натощак потребление пищи дозозависимо снижалось в течение первых получаса по сравнению с мышами, получавшими носитель (рис. 6). Эти результаты согласуются с более ранними наблюдениями, согласно которым NPS ингибирует потребление пищи у мышей и крыс [9], [35]–[37]. Механизмы, с помощью которых НПВ регулируют потребление пищи, до сих пор неизвестны. Однако в некоторых отчетах, описанных Fedeli et al. и Пэн и др. предполагают, что паравентрикулярные ядра гипоталамуса, а также НПСР участвуют в аноректическом действии НПВ [9], [36].Эти результаты показывают, что НПВ усиливает обонятельную функцию, но препятствует проглатыванию.
Как показано на рис. 3, NPS в зависимости от дозы увеличивает общее время вдыхания запаха во время задач привыкания и отказа от обоняния. Обычно считается, что фырканье является частью поведения, вызывающего возбуждение [38]. Во время быстроволнового состояния неокортикальной ЭЭГ обонятельные нейроны коры демонстрировали сильные спайковые ответы на адекватные одоранты, тогда как они демонстрировали только слабые ответы в медленноволновом состоянии [39]–[41]. Наша более ранняя работа продемонстрировала, что NPS значительно увеличивает бодрствование, сопровождаемое увеличением высокочастотной активности ЭЭГ (14.5–60 Гц) и значительно уменьшает медленноволновой сон и парадоксальный сон у крыс [7]. Следовательно, обнюхивание и увеличение времени обнюхивания, вероятно, связаны с увеличением возбуждения, передвижения и исследования, вызванных NPS.
Что еще более важно, наше исследование также направлено на выявление потенциальных мишеней, посредством которых NPS облегчает обонятельную функцию, путем изучения нейронов, экспрессирующих Fos, продукт непосредственно раннего гена, который экспрессируется в связи с активацией нейронов [42], [43].Наши результаты показывают, что центральное введение НПС вызывало увеличение количества Fos-ir нейронов в нескольких областях обонятельной коры, включая AON, Pir (рис. 7), VTT, ACo и LEnt (табл. 1). Считается, что это происходит в AON для большей части начальной конвергенции признаков запаха, связанной с ранними стадиями создания объектов запаха [28]. В то время как Пир, самая большая область обонятельной коры, будет выполнять ассоциации более высокого порядка между объектами запаха и гедоникой, контекстом и другими запахами [15].Наше настоящее исследование также было разработано для изучения того, были ли эффекты NPS на регуляцию обонятельной функции избирательно противодействовать антагонисту NPSR, и, кроме того, экспрессировали ли нейроны, активированные NPS, свой родственный рецептор в обонятельной коре. Результаты показывают, что эффекты NPS на регуляцию обонятельной функции блокируются антагонистом NPSR [D-Val 5 ]NPS (рис. 4 и 5), и что подавляющее большинство Fos-ir нейронов активируется АНП в АОН и Пире также содержали НПСР (рис.8С и F). В большом количестве литературы показано, что NPS избирательно связывает NPSR с высокой аффинностью, вызывая биологические действия [1], [9], [10], [12], [44], [45]. Наши результаты убедительно свидетельствуют о том, что NPS способствует обонятельной способности за счет активации нейронов, несущих NPSR, в обонятельной коре.
Таким образом, центральное введение НПВ у мышей усиливает обонятельные функции, уменьшая латентный период для поиска закопанной пищи и усиливая обонятельную дифференциацию одинаковых и разных запахов.Эти эффекты являются рецептор-специфичными, поскольку они могут быть заблокированы его селективным антагонистом NPSR [D-Val 5 ]NPS. Кроме того, центральное введение NPS заметно активирует экспрессию c-Fos в нейронах обонятельной коры, большинство из которых также экспрессируют NPSR, что указывает на то, что NPS облегчает обонятельные функции посредством активации NPSR в нейронах обонятельной коры.
Благодарности
Мы благодарим профессора Руи Ванга за подарки NPS и [D-Val 5 ]NPS и Dr.Shuye Pu за исправление рукописи.
Авторские взносы
Инициатива и разработка экспериментов: YH YS. Выполнены опыты: YS PZ CD JL HW. Проанализированы данные: YH YS CD PZ. Предоставленные реагенты/материалы/инструменты для анализа: YH LD. Написал статью: YH YS XK.
Каталожные номера
- 1. Xu YL, Reinscheid RK, Huitron-Resendiz S, Clark SD, Wang Z, et al. (2004) Нейропептид S: нейропептид, способствующий возбуждению и анксиолитическим эффектам.Нейрон 43: 487–497.
- 2. Кларк С.Д., Дуангдао Д.М., Шульц С., Чжан Л., Лю С. и др. (2011) Анатомическая характеристика системы нейропептида S в мозге мыши с помощью гибридизации in situ и иммуногистохимии. J Comp Neurol 519: 1867–1893.
- 3. Xu YL, Gall CM, Jackson VR, Civelli O, Reinscheid RK (2007)Распределение мРНК рецептора нейропептида S и нейрохимические характеристики нейронов, экспрессирующих нейропептид S, в мозге крысы. J Comp Neurol 500: 84–102.
- 4. Леонард С.К., Ринг Р.Х. (2011)Иммуногистохимическая локализация рецептора нейропептида S в центральной нервной системе крыс. Неврология 172: 153–163.
- 5. Duangdao DM, Clark SD, Okamura N, Reinscheid RK (2009)Поведенческое фенотипирование мышей с нокаутом рецептора нейропептида S. Behav Brain Res 205: 1–9.
- 6. Энквист Дж., Ферверда М., Мадхаван А., Хок Д., Уистлер Дж.Л. (2012) Хронический этанол усиливает действие нейропептидов в базолатеральной миндалевидном теле и проявляет повышенный анксиолитический и антидепрессивный эффект.Нейропсихофармакология 37: 2436–2445.
- 7. Чжао П., Шао Ю.Ф., Чжан М., Фань К., Конг Х.П. и др. (2012) Нейропептид S способствует бодрствованию за счет активации гистаминергических и орексинергических нейронов заднего гипоталамуса. Неврология 207: 218–226.
- 8. Li W, Chang M, Peng YL, Gao YH, Zhang JN и др. (2009) Нейропептид S оказывает антиноцицептивное действие на супраспинальном уровне у мышей. Регул Пепт 156: 90–95.
- 9. Пэн Ю.Л., Чжан Д.Н., Чанг М., Ли В., Хань Р.В. и др.(2010)Влияние центрального нейропептида S на формалиновый тест на мышах. Пептиды 31: 1878–1883.
- 10. Джунглинг К., Зайденбехер Т., Сосулина Л., Лестинг Дж., Сангха С. и др. (2008) Нейропептид S-опосредованный контроль выражения и угасания страха: роль интеркалированных ГАМКергических нейронов в миндалевидном теле. Нейрон 59: 298–310.
- 11. Han RW, Yin XQ, Chang M, Peng YL, Li W и др. (2009) Нейропептид S облегчает пространственную память и смягчает ухудшение пространственной памяти, вызванное антагонистом рецептора N-метил-D-аспартата у мышей.Neurosci Lett 455: 74–77.
- 12. Окамура Н., Гарау С., Дуангдао Д.М., Кларк С.Д., Джунглинг К. и др. (2011) Нейропептид S улучшает память во время фазы консолидации и взаимодействует с норадренергическими системами в головном мозге. Нейропсихофармакология 36: 744–752.
- 13. Каллупи М., Каннелла Н., Экономиду Д., Убальди М., Руджери Б. и др. (2010) Нейропептид S способствует индуцированному сигналом рецидиву поиска кокаина за счет активации гипоталамической гипокретиновой системы.Proc Natl Acad Sci USA 107: 19567–19572.
- 14. Невилл К.Р., Хаберли Л.Б. (2004)Обонятельная кора. В Синаптической организации мозга, 415–454.
- 15. Уилсон Д.А., Салливан Р.М. (2011)Корковая обработка объектов запаха. Нейрон 72: 506–519.
- 16. Paxinos G, Franklin KBJ (2001) Мозг мыши в стереотаксических координатах, 2-е изд. Сан-Диего: Академическая пресса.
- 17. Чанг М., Пэн Ю.Л., Донг С.Л., Хань Р.В., Ли В. и др.(2005)Исследования структуры-активности различных модификаций ноцицептина/орфанина FQ: идентификация сильнодействующих агонистов и антагонистов его рецептора. Регул Пепт 130: 116–122.
- 18. Ян М., Кроули Дж. Н. (2009) Простая поведенческая оценка обоняния мыши. Curr Protoc Neurosci Глава 8: Блок 8.24.
- 19. Nathan BP, Yost J, Litherland MT, Struble RG, Switzer PV (2004)Обонятельная функция у мышей с нокаутом ароЕ. Behav Brain Res 150: 1–7.
- 20.Вудли С.К., Баум М.Дж. (2003)Влияние половых гормонов и пола на пороги влечения к летучим запахам анальных желез у хорьков. Хорм Бехав 44: 110–118.
- 21. Wrenn CC, Kinney JW, Marriott LK, Holmes A, Harris AP, et al. (2004)Обучение и память у мышей, у которых отсутствует подтип GAL-R1 рецептора галанина. Eur J Neurosci 19: 1384–1396.
- 22. Версингер С.Р., Колдуэлл Х.К., Мартинес Л., Голд П., Ху С.Б. и др. (2007) Мыши с нокаутом рецептора вазопрессина 1а имеют тонкий обонятельный дефицит, но нормальную агрессию.Genes Brain Behav 6: 540–551.
- 23. Mandairon N, Poncelet J, Bensafi M, Didier A (2009) Люди и мыши выражают схожие обонятельные предпочтения. PLoS One 4: e4209.
- 24. Санчес-Андраде Г., Кендрик К.М. (2009)Основная обонятельная система и социальное обучение у млекопитающих. Behav Brain Res 200: 323–335.
- 25. Лайтинен Т., Полви А., Ридман П., Венделин Дж., Пулккинен В. и др. (2004) Характеристика общего локуса восприимчивости к признакам, связанным с астмой.Наука 304: 300–304.
- 26. Венделин Дж., Пулккинен В., Рен М., Пирсканен А., Райсанен-Соколовски А. и соавт. (2005) Характеристика GPRA, нового рецептора, связанного с G-белком, связанного с астмой. Am J Respir Cell Mol Biol 33: 262–270.
- 27. Геррини Р., Камарда В., Трапелла С., Кало Дж., Рицци А. и др. (2009)Синтез и биологическая активность аналогов нейропептида S человека, модифицированных в положении 5: идентификация сильнодействующих и чистых антагонистов рецептора нейропептида S.J Med Chem 52: 524–529.
- 28. Haberly LB (2001)Параллельно-распределенная обработка в обонятельной коре: новое понимание морфологического и физиологического анализа нейронных цепей. Химические чувства 26: 551–576.
- 29. Bouret S, Sara SJ (2002)Активация голубоватого пятна модулирует скорость стрельбы и временную организацию вызванных запахом одноклеточных ответов в грушевидной коре крысы. Eur J Neurosci 16: 2371–2382.
- 30. Jiang M, Griff ER, Ennis M, Zimmer LA, Shipley MT (1996)Активация голубого пятна усиливает реакцию митральных клеток обонятельной луковицы на слабый вход обонятельного нерва.J Neurosci 16: 6319–6329.
- 31. Hasselmo ME, Linster C, Patil M, Ma D, Cekic M (1997)Норадренергическое подавление синаптической передачи может влиять на кортикальное отношение сигнал/шум. Дж. Нейрофизиол 77: 3326–3339.
- 32. Smith JJ, Shionoya K, Sullivan RM, Wilson DA (2009)Слуховая стимуляция выводит из строя обонятельные реакции посредством норадренергической корковой модуляции. Нейрал Пласт 2009: 754014.
- 33. Шипли М.Т., Эннис М. (1996)Функциональная организация обонятельной системы.Дж. Нейробиол 30: 123–176.
- 34. Leinwand SG, Chalasani SH (2011)Обонятельные сети: от ощущения к восприятию. Curr Opin Genet Dev 21: 806–811.
- 35. Beck B, Fernette B, Stricker-Krongrad A (2005) Пептид S является новым мощным ингибитором произвольного и быстрого приема пищи у крыс. Biochem Biophys Res Commun 332: 859–865.
- 36. Федели А., Бракони С., Экономиду Д., Каннелла Н., Каллупи М. и др. (2009) Паравентрикулярное ядро гипоталамуса является нейроанатомическим субстратом для ингибирования приема вкусной пищи нейропептидом S.Евр Джей Нейроски. 30: 1594–1602.
- 37. Смит К.Л., Паттерсон М., Дхилло В.С., Патель С.Р., Семджонус Н.М. и др. (2006) Нейропептид S стимулирует гипоталамо-гипофизарно-надпочечниковую ось и подавляет прием пищи. Эндокринология 147: 3510–3518.
- 38. Seelke AM, Blumberg MS (2004)Обнюхивание крысят во время сна и бодрствования. Behav Neurosci 118: 267–273.
- 39. Мураками М., Кашивадани Х., Кирино Ю., Мори К. (2005)Зависящие от состояния сенсорные ворота в обонятельной коре.Нейрон 46: 285–296.
- 40. Барнс Д.К., Чапюи Дж., Чаудхури Д., Уилсон Д.А. (2011)Кондиционирование страха перед запахом изменяет локальные полевые потенциалы грушевидной коры как во время кондиционирования, так и во время сна после кондиционирования. PLoS One 6: e18130.
- 41. Уилсон Д.А. (2010) Единичная активность грушевидной коры во время медленноволнового состояния определяется недавним ощущением запаха. J Neurosci 30: 1760–1765.
- 42. Morgan JI, Curran T (1986) Роль потока ионов в контроле экспрессии c-fos.Природа 322: 552–555.
- 43. Драгунов М., Фаулл Р. (1989)Использование c-fos в качестве метаболического маркера при отслеживании путей нейронов. J Neurosci Methods 29: 261–265.
- 44. Филаферро М., Нови С., Руджери В., Дженедани С., Альбони С. и др. (2012) Нейропептид S стимулирует хемотаксис моноцитов человека посредством активации рецептора NPS. Пептиды 39С: 16–20.
- 45. Руцца С., Рицци А., Камарда В., Пульга А., Марзола Г. и др. (2012) [tBu-D-Gly5] NPS, чистый и мощный антагонист рецептора нейропептида S: исследования in vitro и in vivo.Пептиды 34: 404–411.