
Можно ли ездить в лес за валежником, как в старину, ничего не опасаясь? — Важное — Пресс-центр — Главная — Официальный сайт Туринского городского округа
1 февраля 2019
Можно ли ездить в лес за валежником, как в старину, ничего не опасаясь?
Интервью.
Можно ли ездить в лес за #валежником, как в старину, ничего не опасаясь. За разъяснениями по этому вопросу мы обратились к директору ГКУ СО «Туринское лесничество» Андрею Сергеевичу Сафронову.
Вопрос. Андрей Сергеевич, какие изменения по заготовке валежника произошли с 1 января 2019 года, и что изменится для жителей нашего округа?
Ответ: В соответствии с Федеральным Законом от 18 апреля 2018 года № 77-ФЗ «О внесении изменения в статью 32 Лесного кодекса Российской Федерации» валежник отнесен к недревесным лесным ресурсам. Порядок заготовки гражданами валежника для собственных нужд в лесах, расположенных на территории Свердловской области утвержден Законом Свердловской области от 24 сентября 2007 года № 94-ОЗ «О порядке и нормативах заготовки гражданами недревесных лесных ресурсов для собственных нужд в лесах, расположенных на территории Свердловской области».
Вопрос: В каких лесах может осуществляться заготовка валежника жителями нашего округа?
Ответ: Заготовка гражданами валежника, а именно лежащих на поверхности земли остатков стволов деревьев, сучьев, не являющихся порубочными остатками в местах проведения лесосечных работ, и (или) образовавшихся вследствие естественного отмирания деревьев, при их повреждении вредными организмами, буреломе, снеговале, для собственных нужд может осуществляться в лесах любого целевого назначения, расположенных на территории Свердловской области.
Вопрос: Андрей Сергеевич, расскажите о правилах заготовки валежника?
Ответ: Заготовка валежника осуществляется в течение всего года с применением любого ручного инструмента – топор, бензопила и т.д., способами без нанесения ущерба лесным насаждениям, подросту, несомкнувшимся лесным культурам.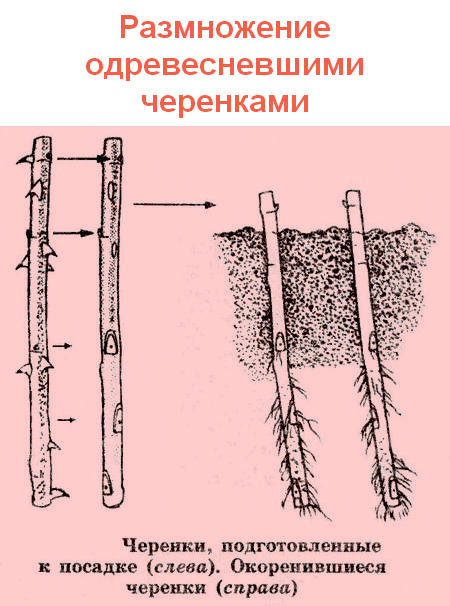
В иных случаях уборка сухостойных деревьев, бурелома, ветровала осуществляется при проведении санитарно-оздоровительных мероприятий. Эти мероприятия на территории лесного фонда планируются по результатам лесопатологических обследований, организуемых и проводимых лесничествами в установленном действующим законодательством порядке. При этом действие Закона Свердловской области от 24 сентября 2007 года № 94-ОЗ на выполнение данных мероприятий не распространяется.
Интервью подготовила ТАТЬЯНА Рудницкая, корреспондент ТТРК «Вектор»
Назад к списку
Саженцы и черенки винограда — что необходимо об этом знать
Красохина С.И., ведущий научный сотрудник лаборатории селекции винограда ВНИИВиВ им. Я.И. Потапенко – филиал ФГБНУ ФРАНЦ, канд. с.-х. наук
Фото Красохиной С.И.
Что такое саженец винограда?
Саженец винограда – это растение возрастом одного года или двух лет, а также вегетирующее в горшке.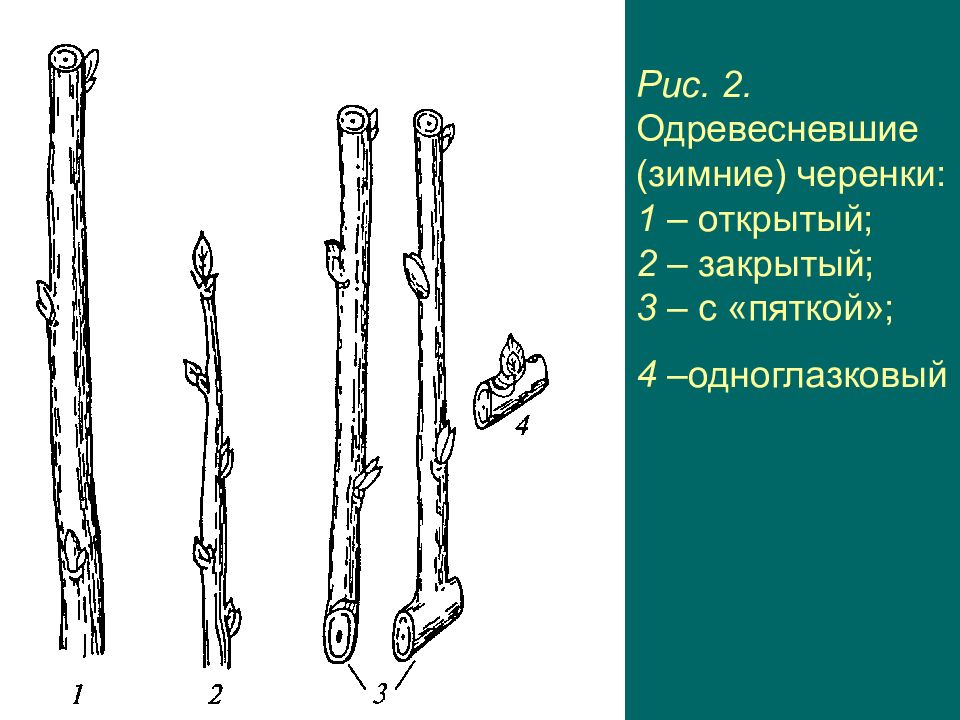 Саженец винограда имеет корни и прирост (и в этом его главное отличие от черенка) и используется для посадки на постоянное место.
Саженец винограда имеет корни и прирост (и в этом его главное отличие от черенка) и используется для посадки на постоянное место.
На фото: одревесневший корнесобственный саженец винограда, фото Красохиной С.И.
Черенок винограда – это кусочек однолетнего прироста виноградной лозы. Черенки бывают одревесневшие (вызревшие), их срезают с куста осенью после листопада или заморозка, и зеленые черенки – их срезают в середине вегетационного сезона (обычно июнь-июль) для последующей прививки.
На фото: черенки винограда, фото Красохиной С.И.
Чем отличается саженец от сеянца?
Сеянец – это растение, которое вырастили из семени винограда. При таком методе размножения у винограда не происходит наследование характеристик исходного сорта.
Внимание! В виноградарстве сеянцы используются только в селекции при выведении новых сортов. Если вы где-то (обычно этим грешит Алиэкспресс) купили семена винограда, клюнув на картинки и дешевизну семян, знайте – из них никогда не вырастет тот сорт, что на картинке. Из них вообще никто не знает и даже не может предположить, что именно вырастет, но результат вам вряд ли понравится.
На фото: сеянцы винограда, выращенные из гибридных семян в научных целях в лаборатории селекции винограда Всероссийского НИИ виноградарства и виноделия им. Я.И. Потапенко. Фото Красохиной С.И.
Какие бывают саженцы винограда?
Саженцы винограда бывают одревесневшие и вегетирующие.
Одревесневший однолетний саженец – это саженец, полученный методом посадки черенка в школку весной и выкопанный из школки осенью. У одревесневшего саженца обычно открытая корневая система (ОКС).
Одревесневший двухлетний саженец – это саженец, посаженный черенком в школку весной этого года, а выкопанный из школки осенью следующего года. Либо, как вариант, непроданные в этом сезоне саженцы коротко обрезают (прирост на 1-2 почки , корни на 1-2 мм) и высаживают в школку как черенки.
Вегетирующий саженец – это саженец, находящийся в периоде своей активной жизнедеятельности (вегетации). У него есть зеленые листья, зеленые побеги и обязательно закрытая корневая система (ЗКС). Назначение вегетирующего саженца – посадка на постоянное место или в школку на доращивание.
На фото: вегетирующий саженец винограда, фото Красохиной С.И.
В зависимости от способа получения однолетние/двухлетние и вегетирующие саженцы бывают корнесобственные и привитые.
Корнесобственные саженцы –это саженцы, выращенные из черенка этого же сорта. То есть с куста взяли черенок, укоренили его в школке или горшке и получили корнесобственный саженец (у него и корни, и прирост свои родные). Корнесобственные саженцы выращивают из длинных и укороченных черенков.
Корнесобственные саженцы выращивают из длинных и укороченных черенков.
Привитые саженцы – это саженцы, состоящие из двух частей: привоя и подвоя. Верх у них сортовой (привой), а корни принадлежат подвойному сорту (подвой). В качестве подвоя при производстве привитых саженцев винограда используют устойчивые к филлоксере американские виды (Рипариа, Рупестрис, Берландиери и др.)
привитой саженец винограда, © Фото Красохиной С.И.
привитой саженец винограда — место прививки, © Фото Красохиной С.И.
На фото: привитые саженцы винограда, фото Красохиной С.И.
Что лучше – привитые саженцы или корнесобственные?
Это философский вопрос. Есть у каждого из этих двух видов саженцев свои достоинства и недостатки.
В местах, где почва заражена филлоксерой, корнесобственные саженцы растут слабо, угнетаются и часто вскоре погибают. Поэтому там целесообразно сажать привитые саженцы.
Если почва не заражена, то вполне можно выращивать корнесобственные саженцы. Их главное преимущество состоит в том, что если вследствие каких-то причин надземная часть корнесобственного куста погибнет, то от корня (подземного корнештамба) вырастут побеги того же самого сорта.
Их главное преимущество состоит в том, что если вследствие каких-то причин надземная часть корнесобственного куста погибнет, то от корня (подземного корнештамба) вырастут побеги того же самого сорта.
Какие документы регулируют качество саженцев и черенков?
Качество посадочного материала регулирует ГОСТ. В РФ есть отдельный ГОСТ на черенки (ГОСТ Р 53050-2008) и на саженцы винограда (ГОСТ 31783-2012). Эти документы предназначены для промышленности и производителей посадочного материала. При покупке черенков и саженцев для себя никто не будет копаться в официальных документах.
Я предлагаю для простоты восприятия при покупке саженцев или черенков воспользоваться вот этими таблицами, которые я постаралась максимально перевести на понятный язык, не сильно нарушая требования документов.
Технические требования к посадочному материалу винограда
Одревесневшие черенки
Показатели |
Признаки |
Внешний вид |
|
Почки (глазки) |
Нормально сформированные, не распустившиеся |
Кора |
Кора не должна иметь повреждения и быть измочаленной |
Срез |
На срезе — интенсивно зеленый цвет, без темных пятен некроза |
Сердцевина |
|
Размеры черенков для прививки и посадки |
|
Длина |
|
Толщина |
|


Зеленые черенки для прививки не должны быть травянистыми. Диаметр — чтобы он подходил (по возможности) к диаметру подготовленного у вас под прививку подвоя, и чтобы было комфортно делать прививку (не слишком тонкий).
На фото: зеленые черенки винограда, подготовленный под прививку методом окулировки в приклад, фото Красохиной С.И.
Если вы купили черенки винограда осенью и вам надо их хранить до весны, то каким способом это лучше сделать, можно почитать здесь: Как сохранить черенки винограда в зимнее время.
Если вы купили саженцы винограда осенью и вам надо их сохранить до весны, то каким способом это лучше сделать, можно почитать здесь: Как сохранить саженцы винограда в зимнее время
Саженцы корнесобственные
Показатели |
Признаки |
Внешний вид |
|

Как должны выглядеть и каким требованиям отвечать корнесобственные саженцы винограда можно узнать из ролика на нашем Ютуб-канале:
Саженцы привитые
Место спайки (там, где была сделана прививка и привой срастается с подвоем) |
Срастание круговое, полное, спайка прочная на изгиб, не должно быть видно щели между привоем и подвоем («зевоты») |
Внешний вид прироста (привоя) |
|
Почки (глазки) |
По требованиям ГОСТ — не менее 4 живых глазков, не распустившихся, хорошо сформированных, но многолетняя практика и стандарты других стран говорят о том, что вполне достаточно и 2 глазков. |
Подвойная часть |
|

Как выглядят привитые саженцы винограда, чем отличаются от корнесобственных и каким требованиям они должны отвечать:
Корни у всех саженцев (и корнесобственных, и привитых):
- На внешний вид: живые, упругие, лучше, если корни расположены равномерно по окружности пятки
- На срезе — сочные, бело-желтоватого или бело-сероватого («мучнистого») цвета
- Количество развитых корней — не менее 3
- Длина корней – не менее 12 см
- Толщина корней – не менее 0, 2 см у основания корня
корни корнесобственных саженцев винограда, © Фото Красохиной С.
 И.
И.корни привитых саженцев винограда, © Фото Красохиной С.И.
На фото: равномерное расположение корней у корнесобственных (а) и привитых (б) саженцев винограда, фото Красохиной С.И.
Помните! Корни и прирост винограда всегда взаимосвязаны между собой.
Если нет возможности увидеть корни (например, при покупке саженца с ЗКС), то об их состоянии можно косвенно судить по виду прироста: прирост тонкий, слабый – значит, и корни тонкие и слабые. И наоборот, если прирост мощный и толстенький, можно смело предполагать наличие хорошо развитых корней.
Саженцы вегетирующие
Внешний вид |
|
Корни |
|

Чем руководствоваться при покупке саженцев и черенков?
Руководствуйтесь данной таблицей и еще несколькими советами:
- Качественный саженец вам должен понравиться.
 Он должен выглядеть крепким, здоровым и достаточно мощным, иметь несколько развитых корней от светло-коричневого до темно-коричневого цвета, не черного. Прирост должен быть эластичный, не сухой.
Он должен выглядеть крепким, здоровым и достаточно мощным, иметь несколько развитых корней от светло-коричневого до темно-коричневого цвета, не черного. Прирост должен быть эластичный, не сухой. - При покупке саженца с ОКС не стесняйтесь попросить продавца сделать срезы верхней части саженца (срез должен быть зеленым и сочным) и немного – корней (срез должен быть светлым).
- При покупке саженца с ЗКС (вегетирующего) обращайте внимание на его внешний вид и силу роста. Листья должны быть здоровыми, и саженец не должен выглядеть как замухрышка.
- При покупке саженцев через интернет у частных лиц обращайте особое внимание на репутацию продавца. Почитайте реальные отзывы о продавце на специализированных форумах или в соцсетях. Это некоторым образом поможет вам избежать не только посадочного материала ненадлежащего качества, но и пересортицы, так как уважающий себя и берегущий свою репутацию продавец следит за чистосортностью.
- При покупке саженцев в специализированных магазинах смотрите не на красивые коробочки с картинкой, а на качество и условия хранения саженцев (таблица вам в помощь).
- Прежде, чем покупать или делать заказ на саженцы понравившегося сорта, не поленитесь – запомните название этого сорта и почитайте о нем на специализированных форумах по винограду. Например, здесь форум Виноград инфо. Ничего не пишут про такой сорт? Задайте про него свой вопрос на форуме. Никто ничего не знает или толком не может рассказать о таком сорте? Это повод призадуматься.
- Также совсем нелишним будет прочесть отзывы о понравившемся Интернет-магазине (если вы решили сделать покупку онлайн) во всех доступных вам источниках (форумы, соцсети, специализированные сайты, слова знакомых или бывалых и т.п.). Интернет-магазины посадочного материала часто грешат несоответствующими реальности описаниями сортов винограда, искусственными крашеными картинками и даже наличием несуществующих сортов.
Внимание! Не лишним также будет помнить, что согласно постановлению Правительства РФ №55 от 19.01.1998 г. (п.13, дополнение от 20.10.1998 г. №1222) проданные растения не подлежат обмену или возврату.
Несколько практических советов при покупке саженцев винограда — статья была написана мной в те недавние времена, когда Интернет-магазинов и специализированных магазинов по продаже растений еще не существовало, но многие советы по-прежнему весьма актуальны и сегодня.
ЧЕРЕНКИ В БАНКЕ | Наука и жизнь
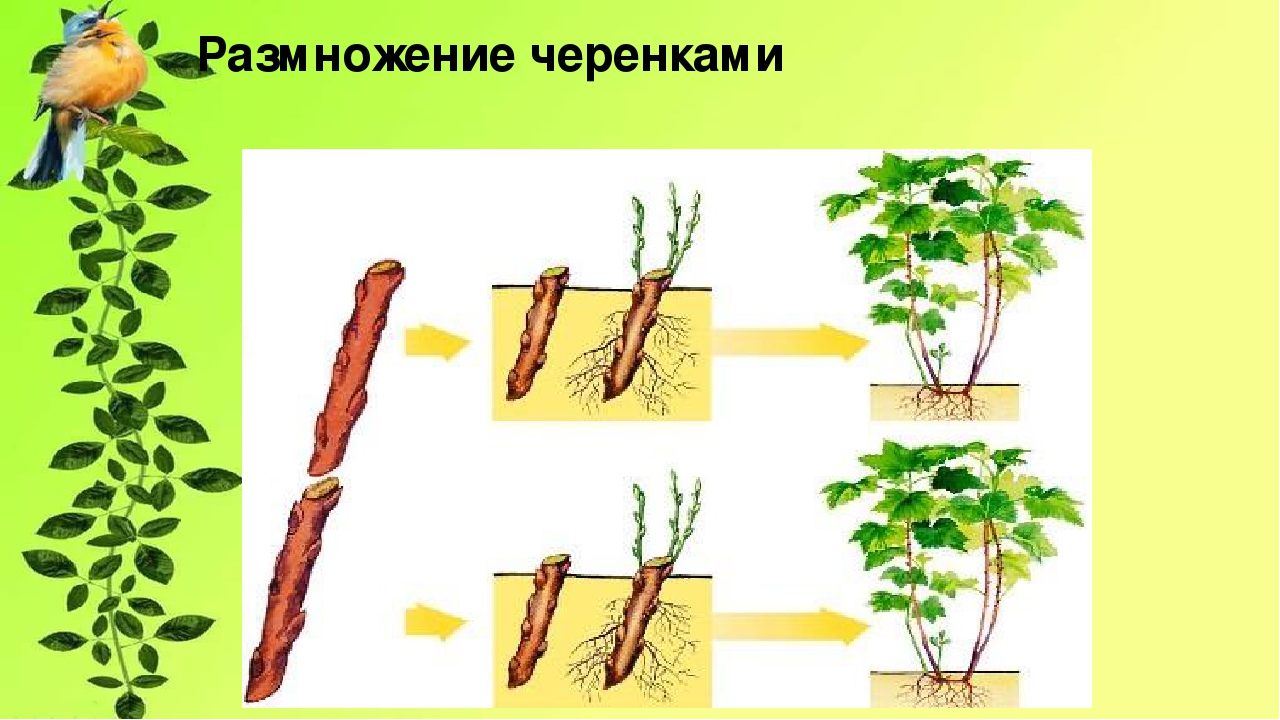
Выращивание растений из черенков — самый популярный способ вегетативного размножения.
Черенки, состоящие из почки с листом, можно нарезать из любых побегов — зеленых, полуодревесневших и одревесневших.
‹
›
Казалось бы, что еще можно написать в дополнение к многочисленным руководствам по укоренению черенков? А вот когда начинаешь сам заниматься черенкованием, обнаруживается масса мелочей, которые решающим образом влияют на результат.
Самый простой способ укоренения — в банке с водой. При определенных условиях им можно пользоваться даже для растений, которые считаются трудно укореняющимися. Но при всей простоте этого метода есть много мелочей, от которых зависит успех.
Например, не все растения выносят смену воды. Во избежание возможной неудачи лучше не менять воду, а только доливать ее при испарении. Видимо, в ней накапливаются какие-то необходимые растению продукты обмена. Мне неоднократно приходилось наблюдать гибель уже давших корни черенков пассифлоры голубой после смены воды, даже если вода была хорошо отстоявшаяся, не содержавшая никаких вредных примесей.
Не менее важно и количество черенков в банке. Жимолость не желает укореняться, если на 200 мл воды приходится более трех черенков, а вот по одному в баночке даже меньшего объема те же черенки прекрасно дают корни.
Выращивание растений из черенков — самый популярный способ вегетативного размножения. Слева — узловой черенок. Нижний его срез находится непосредственно под узлом или почкой. Так обычно режут зеленые невызревшие стебли, поскольку расположенные в этом месте ткани более устойчивы к грибным заболеваниям. Срез в середине междоузлия (справа) делают, как правило, при нарезке черенков из более вызревших одревесневших побегов.
Интересно, что способность растения давать укореняющиеся черенки зависит от питания материнского растения. Если оно в избытке получало азот, корни на черенках образуются очень плохо. Трудно образуются корни и у растений, которые «голодали». Для максимально активного образования корней материнское растение должно быть полностью обеспечено калием, фосфором и немного недополучать азот.
Важно и то, с каких побегов взяты черенки. Жирующие, мощно растущие вверх побеги дают черенки плохого качества. Так что не гонитесь за мощью и размером, гораздо лучше укореняются черенки с боковых, не очень сильно растущих веток. Но любопытно, что у некоторых хвойных место взятия черенка влияет на форму будущего растения. Из черенков с боковых горизонтальных веток тиса остроконечного получаются растрепанные или даже почти стелющиеся растения, а из вертикально росших черенков — вертикальные. Похожие явления замечены и при черенковании елей.
Похожие явления замечены и при черенковании елей.
Для легко укореняемых растений возраст материнского растения роли не играет, но для тех растений, которые черенкуются трудно, это очень важно. Чем старше «родитель», тем, в большинстве случаев, хуже укореняются его черенки. В молодом возрасте черенкуются даже такие растения, которые черенковать не принято, например яблони и груши, у которых при обработке препаратом гетероауксин (стимулятором корнеобразования) легко укореняются верхушечные черенки, взятые с однолетних сеянцев. Но легкость образования корней у черенков, заготовленных из однолетних веток, снижается с увеличением возраста дерева, даже при обработке стимуляторами. Более того, некоторые растения с возрастом вовсе теряют способность к вегетативному размножению. Так произошло в нашем саду с гортензией черешчатой, которая великолепно давала отводки 20 лет назад, а сейчас не укореняются даже прикопанные ветки куста.
Черенки, состоящие из почки с листом, можно нарезать из любых побегов — зеленых, полуодревесневших и одревесневших. Верхний срез делают как можно ближе к почке, по возможности, не оставляя пенька. Нижний — в 2,5-4 см от верхнего. Каждый черенок должен состоять из одного листа (другой лист удаляют), находящейся в его пазухе почки и очень короткого отрезка стебля.
Верхний срез делают как можно ближе к почке, по возможности, не оставляя пенька. Нижний — в 2,5-4 см от верхнего. Каждый черенок должен состоять из одного листа (другой лист удаляют), находящейся в его пазухе почки и очень короткого отрезка стебля.
Небезразлично и то, из какой части побега взят черенок. Часто для черенкования берут очень длинные побеги, из которых нарезают секатором сразу большое количество черенков. Чем ниже расположен отрезок стебля, из которого взят черенок, тем лучше у большинства растений он укореняется. Однажды мне пришлось брать черенки с одной и той же ветки пассифлоры. Несмотря на то, что вся ветка была одинаково одревесневшей, укоренился только последний черенок, взятый с самого ее основания. То же самое можно наблюдать у такого часто черенкуемого растения, как роза.
Большое влияние на укоренение черенков оказывает свет. Черенок, у которого нет листьев, лучше дает корни в темноте. Для укоренения черенка, у которого остался хоть кусочек листа, необходим свет. Но даже при свете в непрозрачной посуде корни образуются гораздо лучше, чем в светлой банке. Считается, что черенок, заготовленный из ветки в период, когда листьев нет, уже содержит некоторое количество гетероауксина, который, по-видимому, разлагается на свету. Если же листья есть, именно они вырабатывают стимулятор корнеобразования гетероауксин. Можно было бы оставлять на черенках все листья, но, находясь в воде, а тем более при укоренении в субстрате, когда поступление воды ограничено, лишние листья высушивают черенки и они могут погибнуть.
Но даже при свете в непрозрачной посуде корни образуются гораздо лучше, чем в светлой банке. Считается, что черенок, заготовленный из ветки в период, когда листьев нет, уже содержит некоторое количество гетероауксина, который, по-видимому, разлагается на свету. Если же листья есть, именно они вырабатывают стимулятор корнеобразования гетероауксин. Можно было бы оставлять на черенках все листья, но, находясь в воде, а тем более при укоренении в субстрате, когда поступление воды ограничено, лишние листья высушивают черенки и они могут погибнуть.
Важен и уровень воды в банке. Если воды слишком много, вблизи дна кислорода недостаточно, а это может вызвать загнивание нижней части черенков: ведь корни образуются только на границе воды и воздуха. Опыты англичан показали, что корни появляются гораздо быстрее и по всей длине черенков, находящихся в банке, в том случае, если проводилась аэрация воды.
И еще одно интересное наблюдение. Многие растения лучше размножаются в период покоя. Высаженные осенью в грунт черенки черной смородины укореняются гораздо лучше, чем весенние, которые почти полностью погибают. Хорошо размножается зимними черенками облепиха. Срезанные в конце января — феврале веточки облепихи дают корни уже через одну-две недели в банке с обычной водой или с водой, в которую добавлен мед. Столовую ложку меда разводят в стакане воды и ставят в раствор черенки на 6-12 часов. Основная ошибка садоводов при зимнем черенковании облепихи — слишком поздняя ее пересадка в почву. Корни на черенках растут очень быстро, они становятся слишком длинными и при перенесении в сад обламываются.
Отличные корни в обыкновенных вазах с водой дали срезанные в феврале и марте для букета длинные ветки тиса ягодного и туи западной (около 60 см длиной и около сантиметра в диаметре). В мае — июне все они с хорошо развитыми корнями были высажены в грунт.
И еще один любопытный метод черенкования, распространенный в последнее время среди садоводов-любителей.
Берут крупный клубень картофеля и тщательно удаляют из него все глазки. Черенки втыкают прямо в клубень, закапывают его в землю и накрывают стеклянной банкой для создания парникового эффекта. При регулярном поливе черенки быстро дают корни, а посаженные в саду молодые растения хорошо развиваются. Так удается укоренить даже плохо черенкующиеся растения. Метод вполне обоснован, поскольку вместо воды черенки получают сразу большое количество готовых питательных веществ, в частности углеводов, в том числе крахмала.
Черенки втыкают прямо в клубень, закапывают его в землю и накрывают стеклянной банкой для создания парникового эффекта. При регулярном поливе черенки быстро дают корни, а посаженные в саду молодые растения хорошо развиваются. Так удается укоренить даже плохо черенкующиеся растения. Метод вполне обоснован, поскольку вместо воды черенки получают сразу большое количество готовых питательных веществ, в частности углеводов, в том числе крахмала.
Возник этот метод случайно, и можно даже предположить, каким образом. Давно известно, что сохранять черенки для прививок в домашних условиях лучше всего в холодильнике. Черенки втыкают в обычные клубни картофеля, заворачивают в бумагу и помещают в полиэтиленовый пакет с дырками. Вероятно, кто-то из садоводов оставил слишком надолго черенки в клубне, и они дали корни.
Таким же образом можно хранить и некоторые цветы. В частности, мне удалось сохранить свежими до 4-5 марта зеленые бутоны тюльпанов, купленные в начале февраля. Поставленные в воду, они через три-четыре дня приобрели нормальную окраску и прекрасно распустились.
В прошлом году уже почти осенью быстро укоренились в картофеле черенки роз.
Cпособ размножения тополя белого пирамидального (мужская форма) в культуре изолированных тканей растений для озеленения населенных пунктов Иркутской области
Тополь является наиболее пригодной древесной породой для озеленения населенных пунктов, так как он быстро растет, образует большую листовую массу, эффективно защищающую воздух от пыли и вредоносных газов. Одним из недостатков тополя является образование пушистых семян у женских растений, что заставляет иногда отказываться от его использования для озеленения. Среди большого числа видов тополя особый интерес для озеленения представляет тополь белый (Populus alba L.), имеющий высоко декоративную листву, благодаря которой его называют тополем серебристым. Кроме того, он устойчив к листовой ржавчине, из-за которой наиболее распространенный в городах Иркутской области тополь бальзамический рано сбрасывает листву и перестает выполнять защитную функцию уже в августе-сентябре.
В Центральном Сибирском Ботаническом Саду СО РАН (Новосибирск) отселектирована форма, названная «Памяти Вавилова». Это мужские особи, не дающие пуха, с узко пирамидальной кроной, с листьями, имеющими темно-зеленую поверхность и бело-войлочную нижнюю сторону. Это придает таким деревьям высокую декоративность. Кроме того, листва долго сохраняется в осенний период, так как устойчива к ржавчине. Эта форма рекомендована для озеленения Иркутска (Сизых и др., 2012). Примером успешного использования тополя белого «Памяти Вавилова» для озеленения является его посадка по периметру комплекса «Микрохирургия глаза» в Иркутске (рисунок 1).
Препятствием для широкого применения этой формы тополя является трудности его размножения, так как зеленые и одревесневшие черенки, взятые из кроны взрослого дерева тополя белого, практически не укореняются (Бакулин, 2012), что наблюдалось и в наших опытах.
Для преодоления слабой укореняемости этого тополя применен биотехнологический метод – клональное микроразмножение. Пробудившиеся почки на ветке, взятой от одного из деревьев, произрастающих на территории «Микрохирургии глаза» были введены в изолированную культуру и размножаются постоянно в пробирках с 2011 года (рис. 2). При их высадке в контейнеры с грунтом они образуют рассаду (рис. 3). Эта рассада используется для выращивания саженцев, которые затем высаживаются на постоянное место (рис. 4).
В 2014 году получено около 150 растений, которые выращиваются на территории института для получения саженцев.
Цитированная литература:
Бакулин В. Т. – Тополь белый в Западной Сибири. Нсб, академическое изд-во «Гео», 2012.
Сизых С. В. и др. – Растения для озеленения Иркутска. – Иркутск, ООО «Байкал-Инновация», 2012.
Инновационные аспекты предложения: Разработан способ массового размножения тополя белого с использованием клонального микроразмножения.
Главные преимущества предложения: Массовое размножение тополя белого с использованием зеленых и одревесневших черенков невозможно, а предложенный способ снимает это ограничение.
Технологические ключевые слова: Размножение древесных растений, озеленение населенных пунктов
Тип требующегося сотрудничества:
1) подача заявок на производство саженцев в количестве 100-200 штук,
2) при большей потребности организация совместного предприятия, в котором Малое инновационное предприятие «Центр инновационных технологий СИФИБР» (ООО «ЦИТ СИФИБР») будет обеспечивать исходный материал для получения саженцев (пробирочные растения), а второе учреждение будет их использовать для выращивания саженцев.
Текущая стадия развития: Первоначальная технология разработана. Планируется дальнейшее ее совершенствование.
Права интеллектуальной собственности: Оформляется ноу-хау.
Контактная информация:
СИФИБР СО РАН, г.Иркутск
Адрес: 664033, г. Иркутск, ул. Лермонтова, 132, а/я 1243
Гамбург Ким Захарович
Лаборатория физиологической генетики
Тел.: (3952) 42-63-03
факс: (3952) 51-07-54
e-mail: Этот адрес электронной почты защищён от спам-ботов. У вас должен быть включен JavaScript для просмотра.
.
Продуктивная способность маточных насаждений смородины при выращивании саженцев из одревесневших черенков
В. Жидехина,
О. С. Родюкова,
ГНУ ВНИИС им. И.В. Мичурина
В статье представлены результаты исследований по оценке пригодности сортов смородины черной и красной к размножению одревесневшими черенками.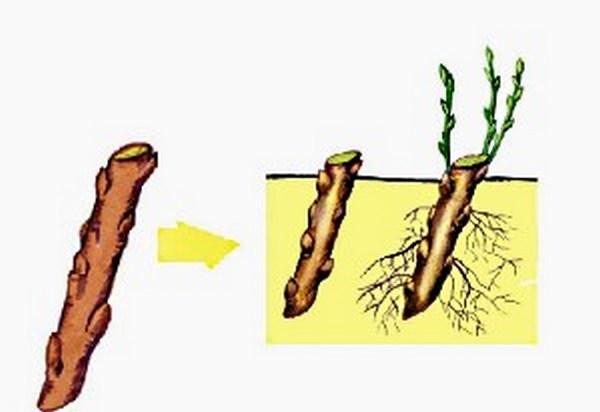 Установлено, что максимальным выходом с 1 га одревесневших черенков (свыше 250 тыс. шт.) и саженцев (более 70 тыс. шт.) характеризуются сорта смородины черной — Кармелита, Тамерлан, Чернавка, Шалунья. Наибольший выход с 1 га черенков (свыше 200 тыс. шт.) и саженцев (более 80 тыс. шт.) на смородине красной получен у сортов Голландская розовая и Jonker van Tets.
Установлено, что максимальным выходом с 1 га одревесневших черенков (свыше 250 тыс. шт.) и саженцев (более 70 тыс. шт.) характеризуются сорта смородины черной — Кармелита, Тамерлан, Чернавка, Шалунья. Наибольший выход с 1 га черенков (свыше 200 тыс. шт.) и саженцев (более 80 тыс. шт.) на смородине красной получен у сортов Голландская розовая и Jonker van Tets.
Продуктивная способность маточных насаждений смородины при выращивании саженцев из одревесневших черенков
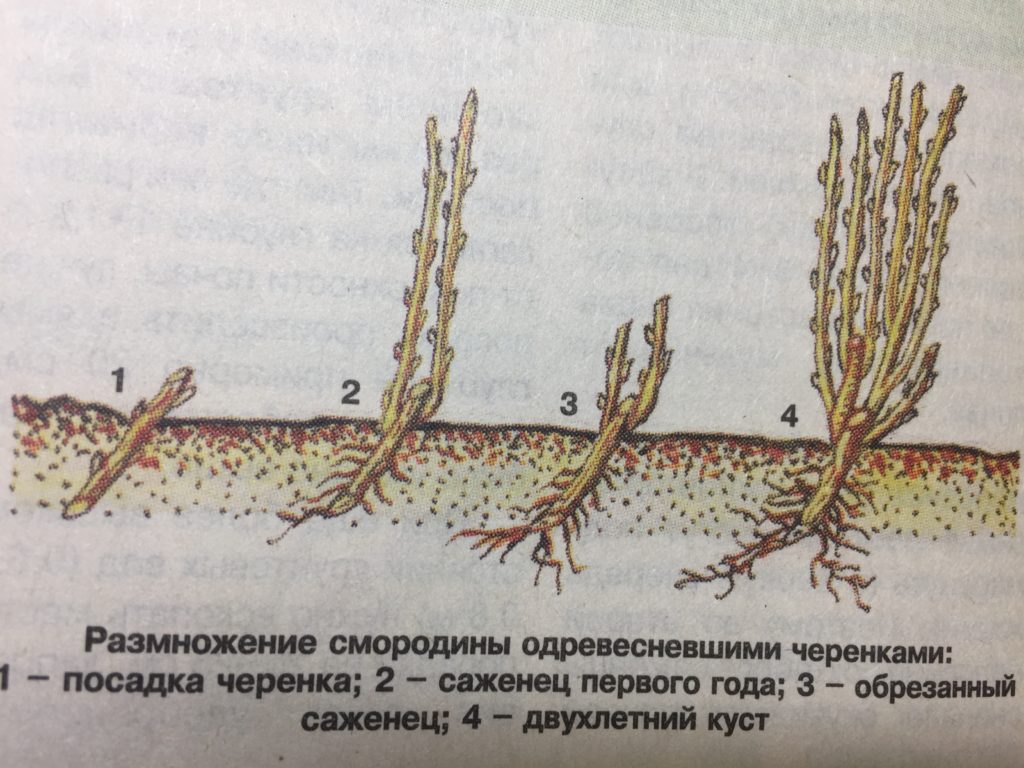
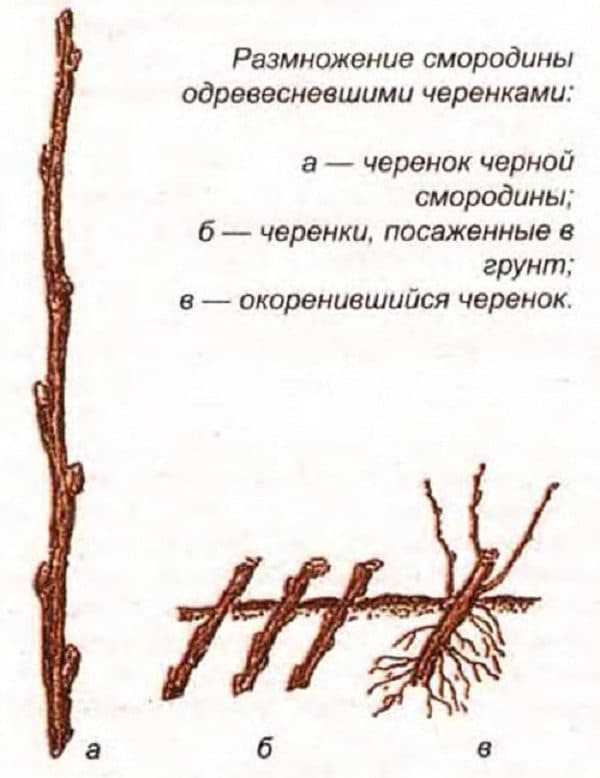
Смородина обладает большой способностью к вегетативному размножению. Основным способом получения саженцев у нее является укоренение одревесневших черенков. Технология предусматривает создание и эксплуатацию маточников, укоренение и доращивание саженцев. Это наиболее доступный и дешевый способ, позволяющий механизировать большинство технологических процессов (Баранова, 1970; Распопова, 1984). Приживаемость черенков смородины черной составляет 8-77 % (Соловьева, 2008), красной — 40-87 % (Ильин, 1981; Соловьева, 2008).
В ГНУ ВНИИС имени И. В. Мичурина Россельхозакадемии были заложены опыты по оценке новых сортов смородины черной селекции института и перспективных — смородины красной на пригодность их к размножению одревесневшими черенками. Маточник был заложен осенью 2004 года двухлетними стандартными саженцами по схеме 3 х 0,7 м. В качестве объектов исследований были использованы сорт; Диво Звягиной, Кармелита, Маленький принц, Тамерлан, Чернавка, Шалунья (смородина черная) и Виксне, Голландская розовая, Jonker van Tets (смородина красная). Уходные работы на маточнике выполняли согласно агротехническим указаниям для Тамбовской области.
Заготовку одревесневших черенков начали проводить с первого года эксплуатации маточника. В первый год выход черенков с 1 растения колебался от 14 (Шалунья) до 27 шт. (Диво Звягиной) у смородины черной (рис. 1) и составил в среднем 8 шт. у всех сортов смородины красной (рис. 2).
Средний выход одревесневшиих черенков с 1 растения смородины черной на традиционном маточникеОценка сортовых особенностей у смородины показала, что максимальное количество одревесневших черенков с одного растения было получено на четвертый год по сортам — Тамерлан (82 шт. ), Кармелита (74), Диво Звягиной (65) у смородины черной и Jonker van Tets (71), Виксне (59 шт.), у смородины красной. На пятый год наиболее продуктивными были сорта Шалунья (103), Чернавка (83), Маленький принц (50) (смородина черная) и Голландская розовая (61 шт.) (смородина красная).
), Кармелита (74), Диво Звягиной (65) у смородины черной и Jonker van Tets (71), Виксне (59 шт.), у смородины красной. На пятый год наиболее продуктивными были сорта Шалунья (103), Чернавка (83), Маленький принц (50) (смородина черная) и Голландская розовая (61 шт.) (смородина красная).
За время товарной эксплуатации маточника смородины среднегодовой выход одревесневших черенков свыше 250 тыс. шт. с 1 га получен на сортах Шалунья (302), Тамерлан (282), Чернавка (276), Кармелита (260) (смородина черная) и свыше 200 тыс. шт./1 га — Голландская розовая (250), Jonker van Tets (238) (смородина красная).
Продуктивность маточных насаждений зависит не только от биологических особенностей сорта, но и от условий вегетационного периода. Оценка метеорологических условий в период эксплуатации маточных насаждений показала, что наиболее напряженные климатические условия для роста и развития растений смородины сложились в 2007 и 2010 годах. В мае-июне 2007 г., при отсутствии осадков, температура воздуха поднималась до + 40°С. А в течение всего вегетационного периода 2010 г среднесуточная температура воздуха на 10-15°С превышала среднемноголетние значения. В третьей декаде июля — первой декаде августа среднесуточная температура достигала +42…+44°С. При этом высокая температура воздуха сочеталась с критической воздушной и почвенной засухой. В условиях отсутствия орошения, при значительном распространении почкового клеща в 2010 г., у сорта Маленький принц отмечен минимальный выход одревесневших черенков (40 шт. с 1 растения), а в 2011 г. черенков не получено совсем, т. к. многие маточные растения погибли во время зимы (рис. 3, 4). В целом оценка репродуктивной способности маточных насаждений смородины чёрной и красной выявила их высокий адаптивный потенциал, однако для получения стабильно высокой продуктивности по годам необходимо соблюдение всех агротехнических требований.
В мае-июне 2007 г., при отсутствии осадков, температура воздуха поднималась до + 40°С. А в течение всего вегетационного периода 2010 г среднесуточная температура воздуха на 10-15°С превышала среднемноголетние значения. В третьей декаде июля — первой декаде августа среднесуточная температура достигала +42…+44°С. При этом высокая температура воздуха сочеталась с критической воздушной и почвенной засухой. В условиях отсутствия орошения, при значительном распространении почкового клеща в 2010 г., у сорта Маленький принц отмечен минимальный выход одревесневших черенков (40 шт. с 1 растения), а в 2011 г. черенков не получено совсем, т. к. многие маточные растения погибли во время зимы (рис. 3, 4). В целом оценка репродуктивной способности маточных насаждений смородины чёрной и красной выявила их высокий адаптивный потенциал, однако для получения стабильно высокой продуктивности по годам необходимо соблюдение всех агротехнических требований.
Выход стандартных саженцев во многом определяется качеством одревесневших черенков, биологическими особенностями сортов, уровнем плодородия используемых по школку почв и соблюдением необходимых агротехнических условий. Максимальным выходом саженцев (более 70 тыс. шт./1 га) характеризовались сорта смородины черной Тамерлан (116), Кармелита (107), Шалунья (104), Чернавка (89), Маенький принц (79), при этом выход стандартных саженцев составил соответственно 95, 99, 96, 72, 64 тыс. шт. Высокий выход саженцев получен у сортов смородины красной: Jonker van Tets (91 тыс. шт./1 га) и Голландская розовая (85), при этом выход стандартных саженцев у них составил 71 и 74 тыс. шт./1 га.
Максимальным выходом саженцев (более 70 тыс. шт./1 га) характеризовались сорта смородины черной Тамерлан (116), Кармелита (107), Шалунья (104), Чернавка (89), Маенький принц (79), при этом выход стандартных саженцев составил соответственно 95, 99, 96, 72, 64 тыс. шт. Высокий выход саженцев получен у сортов смородины красной: Jonker van Tets (91 тыс. шт./1 га) и Голландская розовая (85), при этом выход стандартных саженцев у них составил 71 и 74 тыс. шт./1 га.
Таким образом, новые сорта смородины черной селекции ВНИИС имени И. В. Мичурина и перспективные для условий ЦЧР сорта смородины красной пригодны для размножения их одревесневшими черенками. От качества посадочного материала во многом зависят повышение урожайности товарных плантаций и снижение производственных затрат в первые годы после посадки. Закладка насаждений сортами смородины черной Кармелита, Маленький принц, Тамерлан, Чернавка, Шалунья и смородины красной — Jonker van Tets, Голландская розовая обеспечит получение высокого экономического эффекта, как при выращивании посадочного материала, так и при получении ягодной продукции.
Литература
- Баранова О. А. Размножение черной смородины // Горно- Алтайское садоводство: мат. научно-техн. конф. Алтайской опытной станции садоводства им. М. А. Лисавенко. Барнаул, 1970. С. 22-23.
- Ильин В. С. Выращивание красной смородины в Челябинской области: метод, рекомендации. Новосибирск, 1981. 118 с.
- Распопова Г. И. Рекомендации по размножению смородины комбинированными черенками. М.: Колос, 1984. 17 с.
- Соловьева А. Е. Научные основы питомниководства ягодных культур. Новосибирск, 2008. 280 с.
30 января в 11:00 пройдет пресс-тур «Деревья-экзоты Бирюлевского дендропарка»
На сегодняшний день дендрологическая коллекция Бирюлевского дендропарка насчитывает — 205 видов растений, в том числе таких экзотических как: сосна кедровая сибирская, сосна Веймутова, сосна румелийская, пихта сибирская, пихта одноцветная, лжетсуга Мензиса, береза бумажная, бархат амурский, робиния лжеакация, аралия маньчжурская, скумпия кожевенная, виноград прибрежный, виноград амурский, можжевельник скальный, можжевельник китайский, можжевельник казацкий, сосна горная, кипарисовик горохоплодный, дрок красильный, магония падуболистная, леспедеция двуцветная. Эти растения — интродуценты, естественный ареал произрастания — Дальний Восток, юг России, Китай, Япония, Канада, Западная Европа, Северная Америка, Кавказ.
Эти растения — интродуценты, естественный ареал произрастания — Дальний Восток, юг России, Китай, Япония, Канада, Западная Европа, Северная Америка, Кавказ.
Дрок красильный (Genīsta tinctōria) – интересен тем, что в зимний период сохраняет зеленый цвет листьев и стеблей, снег служит покрывалом и не дает листочкам пересыхать от морозов.
Кустарник магония падуболистная (Mahōnia aquifōlium) семейства барбарисовые имеет красивые блестящие сложные непарноперистые листья, которые экзотично выглядывают из-под снега.
Сосны кедровая, румелийская и Веймутова отлично адаптировались к зимним условиям Москвы и в настоящее время не испытывают никакого стресса от переменчивой московской погоды.
Виноград прибрежный и виноград амурский могут обмерзать в сильные морозы. Сохраняется только та часть растения, которая находится под снегом. Эти растения сотрудники максимально присыпают снегом. Весной от сохранившейся части идет отрастание и восстановление лианы.
Труднее всего в наших условиях приходится леспедеции двуцветной(lespedeza bicolor)семейства бобовые. В природных условиях встречается на юге Дальнего Востока, в северном Китае, в Японии и Корее. Это теплолюбивое растение с тонкими изящными веточками. Укрытие на зиму не спасет растение и в сильные продолжительные морозы обмерзает до одревесневшей части, но за летний период быстро отрастает и цветет на побегах этого года.
Можжевельник скальный, можжевельник китайский, можжевельник казацкий, сосна горная, кипарисовик горохоплодный отлично зимуют в условиях Москвы. Единственное что может испортить их декоративный вид – это обильные снегопады, во время которых снег налипает на ветви и может даже поломать их. Поэтому после снегопадов сотрудники Мосприроды обходят участок с данными видами растений и освобождают их от налипшего снега.
В ходе пресс-тура специалисты Мосприроды покажут эти необычные для нашей местности виды, расскажут, как деревья переживают морозную московскую зиму и какие меры принимают сотрудники для сохранения растений.
Адрес: ул. Липецкая, вл.5а
Аккредитация СМИ: +7 (495) 691-28-90, [email protected]
Пресс-служба Департамента природопользования и охраны окружающей среды Москвы.
Лигнификация — обзор | ScienceDirect Topics
3.18.7.1 Ранние исследования дегидрирования монолигнолов
in vitroВпервые участие монолигнолов в лигнификации можно проследить до исследований Питера Класона, 509,510 , который пришел к выводу 19 конифериловый спирт ( 2 ) и оксиконифериловый спирт. 511 Позже, в 1925 году, Кюршнер предположил, что лигнин был получен в результате полимеризации кониферина ( 73 ). 512 513
Однако в 1932 г. Холгар Эрдтман получил результат, оказавшийся крайне важным для биохимии лигнина. 508,514 К тому времени уже было официально известно, что окислители, такие как FeCl 3 , удаляют атом водорода из фенола с образованием свободного радикала, который может подвергаться радикальному связыванию с образованием бифенильного соединения или дифенилового эфира. Действительно, изоэвгенол ( 105 ) ранее окислялся либо FeCl 3 , либо грибной оксидазой, а Cousin and Hérissey 515 получили продукт, которому была приписана бифенильная структура ( 124 ). (Рисунок 42). 516 С другой стороны, фенол с боковой цепью в положении o или p , несущий двойную связь, конъюгированную с ароматическим кольцом, как предполагал Эрдтман, подвергается связыванию по 8-й положение с образованием 8-8′-связанного димера дегидрогваяретовой кислоты ( 125 ). Эксперименты Эрдтмана впоследствии опровергли обе структуры, вместо которых был получен рацемический (±)-фенилкумаран, дегидродиизоэвгенол ( 126 ), который возникает в результате сочетания радикалов 8–5′, как показано. 508,514 Это привело Erdtman 508,514 к предположению, что лигнины образуются путем дегидрирования фенольных предшественников, несущих сопряженные 7,8-двойные связи, таких как конифериловый спирт ( 2 ).
Действительно, изоэвгенол ( 105 ) ранее окислялся либо FeCl 3 , либо грибной оксидазой, а Cousin and Hérissey 515 получили продукт, которому была приписана бифенильная структура ( 124 ). (Рисунок 42). 516 С другой стороны, фенол с боковой цепью в положении o или p , несущий двойную связь, конъюгированную с ароматическим кольцом, как предполагал Эрдтман, подвергается связыванию по 8-й положение с образованием 8-8′-связанного димера дегидрогваяретовой кислоты ( 125 ). Эксперименты Эрдтмана впоследствии опровергли обе структуры, вместо которых был получен рацемический (±)-фенилкумаран, дегидродиизоэвгенол ( 126 ), который возникает в результате сочетания радикалов 8–5′, как показано. 508,514 Это привело Erdtman 508,514 к предположению, что лигнины образуются путем дегидрирования фенольных предшественников, несущих сопряженные 7,8-двойные связи, таких как конифериловый спирт ( 2 ). Структура дегидродиизоэвгенола ( 126 ) была подтверждена почти десять лет спустя Фройденбергом и Рихтценхайном. 517 Следует также отметить, что Фройденберг и Дюрр 518 (ошибочно) предположили, что структуры фенилкумарана могут быть получены в результате перестройки предполагаемых первичных цепей лигнина, связанных линейным образом через 9– O –4 ′ облигации.
Структура дегидродиизоэвгенола ( 126 ) была подтверждена почти десять лет спустя Фройденбергом и Рихтценхайном. 517 Следует также отметить, что Фройденберг и Дюрр 518 (ошибочно) предположили, что структуры фенилкумарана могут быть получены в результате перестройки предполагаемых первичных цепей лигнина, связанных линейным образом через 9– O –4 ′ облигации.
Рисунок 42. Дегидрирующее связывание изоэвгенола ( 105 ). (a) Продукты, изначально предусмотренные Cousin and Hérissey (соединение ( 124 )) и Erdtman (соединение ( 125 )). (b) Открытие Эрдтманом образования дегидродиизоэвгенола ( 126 ) привело к теории дегидрирования для полимеризации монолигнола.
Во второй половине 1930-х годов было обнаружено, что окисление кониферилового спирта ( 2 ) FeCl 3 приводит к образованию аморфного продукта 519 , который дает, inter alia , verapia кислоты ( 127 ) при обработке горячей щелочью с последующим метилированием и перманганатным окислением. Поскольку одни и те же продукты были получены из дегидродиизоэвгенола ( 126 ) и из лигнинов, эти результаты, как правило, подтверждают идею о том, что лигнины могут быть образованы дегидрированием кониферилового спирта ( 2 ) и родственных соединений, как предположил Эрдтман.
Поскольку одни и те же продукты были получены из дегидродиизоэвгенола ( 126 ) и из лигнинов, эти результаты, как правило, подтверждают идею о том, что лигнины могут быть образованы дегидрированием кониферилового спирта ( 2 ) и родственных соединений, как предположил Эрдтман.
В 1940-х и 1950-х годах были предприняты значительные усилия для изучения результатов реакций связывания монолигнолов in vitro с использованием различных неочищенных препаратов окислительных ферментов, полученных из организмов, некоторые из которых даже не биосинтезировали лигнины.В любом случае, дегидрирование кониферилового спирта ( 2 ), например, O 2 , катализируемое препаратом грибной оксидазы 520 или лакказой, присутствующей в неочищенном грибном ферменте, 521,52 с H 2 O 2 в присутствии пероксидазы 523 давал дегидрирующий полимеризат, имеющий некоторое сходство с лигнином, экстрагируемым водным 90% диоксаном из муки хвойных пород (см. Таблицу 20).Среди образовавшихся исходных димерных, тримерных и олигомерных продуктов удалось предположить структуры около 30 различных соединений. Помимо неохарактеризованного (предположительно) полимерного материала (45%), основными продуктами были (±)-дегидродиконифериловые спирты ( 68 ) (26%), (±)-пинорезинолы ( 128 ) (13%) и ( ±)-гваяцилглицерин 8– O –4’ эфиры кониферилового спирта ( 94 ) (9%). 524 Впоследствии сообщалось также о небольших количествах тримера ( 129 ), тетрамера ( 130 ) и пентамера ( 131 ); 525,526 На рис. 43 показаны задействованные процессы радикально-радикального сочетания и последующие реакции, дающие высшие олигомеры.Как видно, соединения ( 129 )–( 131 ) включают трифункциональную точку разветвления, которая возникает в результате нуклеофильного присоединения фенольной гидроксильной группы либо к монолигнолу (например, ( 2 ), либо к 8–5′ связанный дилигнол ( 94 ) с C-7 промежуточного метиленхинона, образованного из 8- O -4′-сочетания между двумя радикалами кониферилового спирта) (рис.
Таблицу 20).Среди образовавшихся исходных димерных, тримерных и олигомерных продуктов удалось предположить структуры около 30 различных соединений. Помимо неохарактеризованного (предположительно) полимерного материала (45%), основными продуктами были (±)-дегидродиконифериловые спирты ( 68 ) (26%), (±)-пинорезинолы ( 128 ) (13%) и ( ±)-гваяцилглицерин 8– O –4’ эфиры кониферилового спирта ( 94 ) (9%). 524 Впоследствии сообщалось также о небольших количествах тримера ( 129 ), тетрамера ( 130 ) и пентамера ( 131 ); 525,526 На рис. 43 показаны задействованные процессы радикально-радикального сочетания и последующие реакции, дающие высшие олигомеры.Как видно, соединения ( 129 )–( 131 ) включают трифункциональную точку разветвления, которая возникает в результате нуклеофильного присоединения фенольной гидроксильной группы либо к монолигнолу (например, ( 2 ), либо к 8–5′ связанный дилигнол ( 94 ) с C-7 промежуточного метиленхинона, образованного из 8- O -4′-сочетания между двумя радикалами кониферилового спирта) (рис. 43(e)). Однако следует отметить, что в изолированных образцах лигнина не было возможности обнаружить в таких количествах присутствие связей 7- O -4′ ациклического бензиларилового эфира, которые вносят столь заметный вклад в скелет этих олиголигнолов.Это может быть связано с относительно жесткими условиями, необходимыми для солюбилизации лигнина, что может привести к разрыву таких лабильных связей.
43(e)). Однако следует отметить, что в изолированных образцах лигнина не было возможности обнаружить в таких количествах присутствие связей 7- O -4′ ациклического бензиларилового эфира, которые вносят столь заметный вклад в скелет этих олиголигнолов.Это может быть связано с относительно жесткими условиями, необходимыми для солюбилизации лигнина, что может привести к разрыву таких лабильных связей.
Рисунок 43. Одноэлектронное окисление кониферилового спирта ( 2 ) in vitro . (а) Гибридные формы резонанса (свободнорадикальный) резонанс; (б)–(г): 8–5′, 8–8′ и 8– O –4′ способы сочетания либо с внутримолекулярной циклизацией, либо с нуклеофильным присоединением H 2 O при С-7; (e) нуклеофильная атака промежуточного соединения 8- O -4′ хинонметида с различными анионами феноксида с образованием олиголигнолов ( 129 )–( 131 ).
Со временем обнаружилась довольно неприятная изменчивость в выходах, о которых сообщил Фройденберг для основных дилигнолов ( 68 ), ( 94 ) и ( 128 ), образующихся в результате дегидрирования in vitro кониферила. спирт ( 2 ). 527 528 527 528 Действительно, результаты были заметно пересмотрены 495 без идентифицируемого экспериментального подтверждения 528 таким образом, чтобы способствовать пропорции 8- O -4 ‘связанных димеров ( 94 ) образуется относительно двух других дилигнолов ( 68 ) и ( 128 ).Например, в 1957 г. 8- O -4′ связанные дилигнолы были названы 529 на основании обзоров Фройденберга 494,530,531 как преобладающие продукты (~70%), образующиеся в результате дегидратации. кониферилового спирта ( 2 ) in vitro , тогда как все предыдущие экспериментальные данные подтверждали, что 8-5′-связанный дилигнол ( 68 ) фактически преобладал. 524
спирт ( 2 ). 527 528 527 528 Действительно, результаты были заметно пересмотрены 495 без идентифицируемого экспериментального подтверждения 528 таким образом, чтобы способствовать пропорции 8- O -4 ‘связанных димеров ( 94 ) образуется относительно двух других дилигнолов ( 68 ) и ( 128 ).Например, в 1957 г. 8- O -4′ связанные дилигнолы были названы 529 на основании обзоров Фройденберга 494,530,531 как преобладающие продукты (~70%), образующиеся в результате дегидратации. кониферилового спирта ( 2 ) in vitro , тогда как все предыдущие экспериментальные данные подтверждали, что 8-5′-связанный дилигнол ( 68 ) фактически преобладал. 524
Изменения в распределении продуктов, которые, как утверждается, были получены in vitro Фройденбергом, похоже, полностью повторяют или отражают результаты, полученные другими исследователями при анализе реальных одревесневших тканей. Таким образом, аналитические исследования разложения начали показывать, что связи 8- O -4′ составляют около 50% от общего количества лигнинов хвойной древесины, эти выводы без поддержки каких-либо новых данных.
Таким образом, аналитические исследования разложения начали показывать, что связи 8- O -4′ составляют около 50% от общего количества лигнинов хвойной древесины, эти выводы без поддержки каких-либо новых данных.
Однако нет внутреннего противоречия между этими выводами и наблюдением, что связи 8- O -4′ присутствуют менее чем в 20% дилигнолов, дегидрированно образованных из кониферилового спирта ( 2 ) in vitro .Сборка макромолекулярного лигнина в клеточных стенках растений, как полагают, в первую очередь включает дегидрогенативное связывание между монолигнолами и остатками мономеров на растущих биополимерных цепях, 495 , где по крайней мере одно положение (обычно С-8) уже ковалентно связано с другим звеном. То, что происходит при таких обстоятельствах, отражено в результате дегидрогенеративной связи между спиртом Coniferyl ( 2 ) и димерами Dehydrodiconiferyl Алкоголь ( 68 ), 532 EPI -PinOSINOLOL ( 132 ) 533 и гваяцилглицерол 8- O -4′ эфир кониферилового спирта ( 94 ), 532 соответственно, где 8- O -4′ связи были обнаружены Фройденбергом и коллегами преобладать во всех трех случаях. Таким образом, настойчивость Фройденберга в конце 1950-х годов, 2 391 533 на неразличимости лигниновых биополимеров и монолигнолдегидрополимеризатов, произведенных in vitro , больше не диктовала, что 8– O –4’ связаны с дилигнолами. наиболее часто среди дегидродимеров образуются из кониферилового спирта ( 2 ). В самом деле, это могло объяснить несколько более умеренное утверждение, сделанное Фройденбергом в 1968 г. 528 , что гваяцилглицерин 8– O –4’ эфир кониферилового спирта ( 94 ) , вероятно, превосходит выход пинорезинола (). 128 ) и дегидродиконифериловый спирт ( 68 ).
Таким образом, настойчивость Фройденберга в конце 1950-х годов, 2 391 533 на неразличимости лигниновых биополимеров и монолигнолдегидрополимеризатов, произведенных in vitro , больше не диктовала, что 8– O –4’ связаны с дилигнолами. наиболее часто среди дегидродимеров образуются из кониферилового спирта ( 2 ). В самом деле, это могло объяснить несколько более умеренное утверждение, сделанное Фройденбергом в 1968 г. 528 , что гваяцилглицерин 8– O –4’ эфир кониферилового спирта ( 94 ) , вероятно, превосходит выход пинорезинола (). 128 ) и дегидродиконифериловый спирт ( 68 ).
Лигнификация клеточных стенок растений: влияние генетических манипуляций
Лигнин является вторым наиболее распространенным полимером, обнаруженным в
природа после целлюлозы. Среди многих ролей, которые лигнин играет в растении
рост и развитие обеспечивают структурную поддержку земли. растений и в качестве механического барьера в борьбе с вредителями. Хотя жизненно важно
на пригодность растений лигнин отрицательно влияет на переработку бумажной массы и
корм для скота. Большая часть стоимости, сложности и загрязнения
связанный с процессом варки целлюлозы, происходит от удаления лигнина
из дерева для изготовления высококачественной бумаги.Точно так же количество
усваиваемая энергия, доступная жвачным животным в результате ферментации
полисахариды клеточной стенки ограничены лигнином, что делает большую часть
целлюлоза и гемицеллюлоза, недоступные микроорганизмам рубца.
Из-за значительных экономических выгод, которые могут быть достигнуты,
значительные исследовательские усилия были направлены на снижение
количество лигнина или изменение структуры лигнина для облегчения получения целлюлозы
деревьев и улучшения усвояемости кормовых культур. Как результат,
трансгенные эксперименты по оценке генов на многих этапах
пути биосинтеза лигнина предоставили важные сведения (1–3).
растений и в качестве механического барьера в борьбе с вредителями. Хотя жизненно важно
на пригодность растений лигнин отрицательно влияет на переработку бумажной массы и
корм для скота. Большая часть стоимости, сложности и загрязнения
связанный с процессом варки целлюлозы, происходит от удаления лигнина
из дерева для изготовления высококачественной бумаги.Точно так же количество
усваиваемая энергия, доступная жвачным животным в результате ферментации
полисахариды клеточной стенки ограничены лигнином, что делает большую часть
целлюлоза и гемицеллюлоза, недоступные микроорганизмам рубца.
Из-за значительных экономических выгод, которые могут быть достигнуты,
значительные исследовательские усилия были направлены на снижение
количество лигнина или изменение структуры лигнина для облегчения получения целлюлозы
деревьев и улучшения усвояемости кормовых культур. Как результат,
трансгенные эксперименты по оценке генов на многих этапах
пути биосинтеза лигнина предоставили важные сведения (1–3).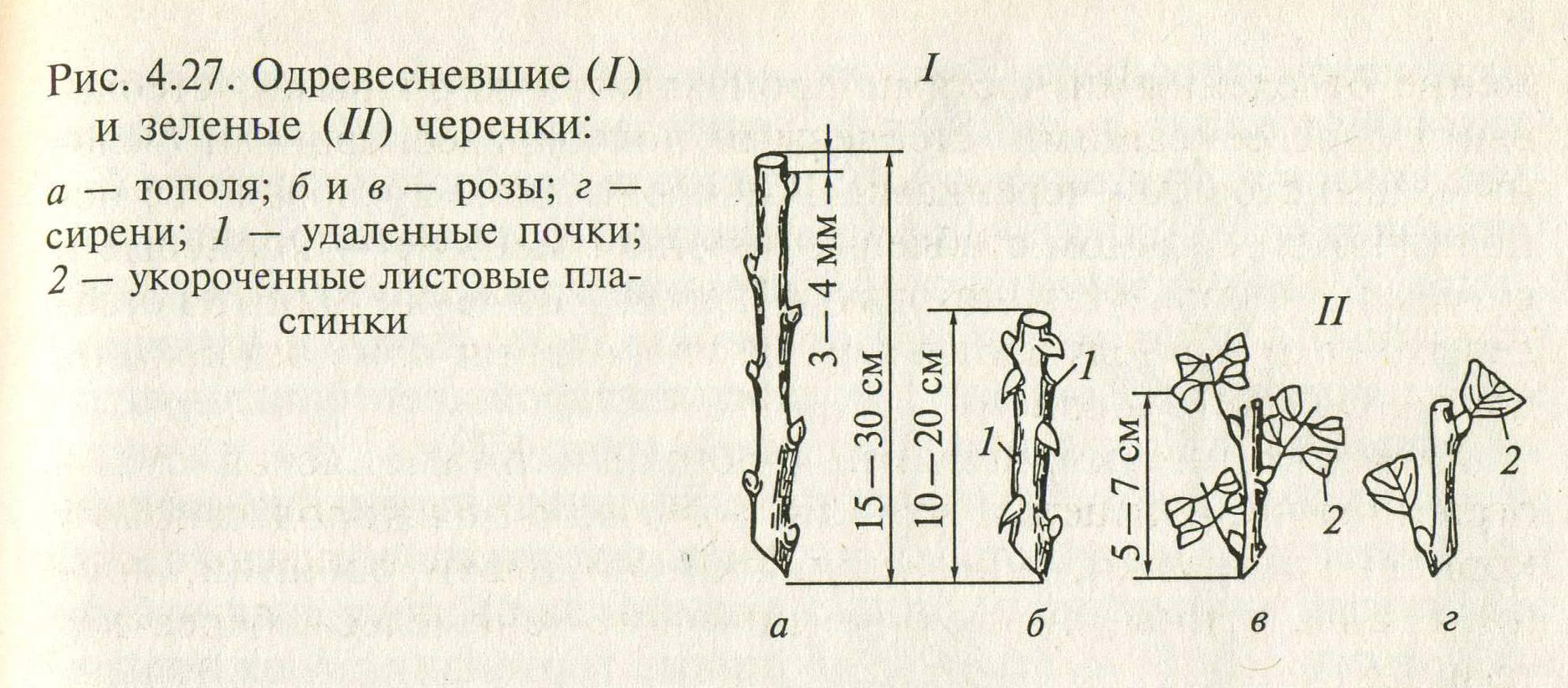
В этом выпуске Proceedings , Ralph et al. (4) сообщение о ряде новых структур, включенных в лигнины
трансгенные растения табака, подавляемые гомологичным антисмысловым
генетические конструкции циннамоил-КоА-редуктазы (CCR) и циннамил
алкогольдегидрогеназа (CAD). Ферментные продукты этих генов
отвечает за катализ двух последних стадий биосинтеза
предшественники лигнина (CCR превращает производные гидроксициннамоил-КоА в
гидроксикоричные альдегиды и CAD восстанавливают эти альдегиды до
гидроксикоричные спирты) перед экспортом монолигнолов в клетку
стенки, где происходит полимеризация.Использование антисмысловой CCR трансгенной
табачная линия, Ralph и др. (4) продемонстрировали, что
Ранее было показано, что тираминовые ферулаты присутствуют в клеточных стенках
обычный табак (5) фактически включены в полимер лигнина.
Ядерно-магнитно-резонансный анализ был использован для подтверждения подлинности
наличие ферулоильных 4-O и 5-положительных связей тирамина
ферулирует в лигнин. Увеличение сирингил-гваяцила (S/G)
соотношение лигнина также было обнаружено, и это изменение произошло в результате снижения
гваяцильные звенья в лигнине трансгенного табака CCR (4).В отличие
к антисмысловому трансгенному табаку CCR, к антисмысловому трансгенному CAD
линия накапливала альдегидные структуры в лигнине, а не в тирамине
ферулирует. В дополнение к увеличению содержания коричного альдегида и бензальдегида,
новая группа альдегидных структур, о которой ранее не сообщалось в растениях
лигнины оказались преобладающими компонентами лигнина. Ральф и др. . (4) использовали анализ ЯМР, чтобы продемонстрировать, что эти новые
альдегиды получают из кониферальдегида и синапальдегида.С этим увеличением содержания альдегидных компонентов связано снижение
структуры, производные кониферилового и синапилового спиртов, обнаруживаемые в норме.
лигнин табака. Нарушение синтеза сирингиллигнина
по антисмысловому трансгену CAD, по-видимому, больше, чем по гваяцилу.
лигнина, поскольку в
трансгенный табак.
Увеличение сирингил-гваяцила (S/G)
соотношение лигнина также было обнаружено, и это изменение произошло в результате снижения
гваяцильные звенья в лигнине трансгенного табака CCR (4).В отличие
к антисмысловому трансгенному табаку CCR, к антисмысловому трансгенному CAD
линия накапливала альдегидные структуры в лигнине, а не в тирамине
ферулирует. В дополнение к увеличению содержания коричного альдегида и бензальдегида,
новая группа альдегидных структур, о которой ранее не сообщалось в растениях
лигнины оказались преобладающими компонентами лигнина. Ральф и др. . (4) использовали анализ ЯМР, чтобы продемонстрировать, что эти новые
альдегиды получают из кониферальдегида и синапальдегида.С этим увеличением содержания альдегидных компонентов связано снижение
структуры, производные кониферилового и синапилового спиртов, обнаруживаемые в норме.
лигнин табака. Нарушение синтеза сирингиллигнина
по антисмысловому трансгену CAD, по-видимому, больше, чем по гваяцилу.
лигнина, поскольку в
трансгенный табак.
Как указано Ральфом и др. . (4), эти необычные лигнин
структуры являются еще одним свидетельством кажущейся пластичности лигнина
образования в растениях, где могут быть многочисленные фенольные предшественники
встраивается в полимер.Аналогичные альдегидные структуры
наблюдаемое для трансгенного табака с пониженной регуляцией CAD, будет
ожидается у мутанта кукурузы brown midrib-1, который недавно был показан
быть мутацией CAD (6). Ральф и др. (4) отметил, что
природный CAD-дефицитный мутант у сосны обыкновенной (7) также накапливал
альдегидные структуры, хотя и не в такой степени, как CAD табака
трансгенный. Но мутант CAD из сосны также обладал дополнительным необычным свойством.
компонент лигнина, полученный из дигидрокониферилового спирта, который не был
наблюдается в трансгенном табаке CAD.Эти наблюдения демонстрируют
насколько гибки растения в адаптации к метаболическим изменениям, связанным с
биосинтез лигнина. Растения, по-видимому, адаптировали несколько стратегий
чтобы обойти генетические и метаболические препятствия для производства лигнина. В
Помимо использования различных предшественников монолигнола, растения также, по-видимому,
обладают метаболической сеткой для биосинтеза монолигнола (8). в
метаболическая сетка, сходные реакции осуществляются одним и тем же ферментом или
изоферменты с различной субстратной специфичностью (9, 10).Эти ферменты
кодируются мультигенными семействами и по-разному экспрессируются
во время развития и в ответ на стресс окружающей среды. То
гибкость метаболической сетки дает лигнины, которые различаются по
состав и строение по типам клеток и тканей, стадиям
развития и в ответ на экологические стимулы (9). Должно
Следует отметить, что, хотя существуют некоторые разногласия относительно того, является ли
необычные структуры лигнина, как сообщает Ralph et al. (4, 7)
фактически включены в полимер лигнина клеточной стенки
(11), текущие данные убедительно подтверждают такой вывод.
В
Помимо использования различных предшественников монолигнола, растения также, по-видимому,
обладают метаболической сеткой для биосинтеза монолигнола (8). в
метаболическая сетка, сходные реакции осуществляются одним и тем же ферментом или
изоферменты с различной субстратной специфичностью (9, 10).Эти ферменты
кодируются мультигенными семействами и по-разному экспрессируются
во время развития и в ответ на стресс окружающей среды. То
гибкость метаболической сетки дает лигнины, которые различаются по
состав и строение по типам клеток и тканей, стадиям
развития и в ответ на экологические стимулы (9). Должно
Следует отметить, что, хотя существуют некоторые разногласия относительно того, является ли
необычные структуры лигнина, как сообщает Ralph et al. (4, 7)
фактически включены в полимер лигнина клеточной стенки
(11), текущие данные убедительно подтверждают такой вывод.
Еще одно важное наблюдение, сделанное Ralph et al. (4) есть
что обычный табак, по-видимому, производит лигнин «объемного» типа, где
монолигнолы реагируют с образованием димеров перед включением в полимер. Это первое сообщение о растительном лигнине, содержащем высокое
пропорции концевых групп α-эфиров и циннамоилового спирта, обнаруженные в
синтетические лигнины, вызванные объемной полимеризацией, а не низким
концентрации этих структур в лигнинах, полученных из типичных
Обычно считается, что в растениях происходит процесс сложения поперек.Данный
эта необычная схема биосинтеза лигнина в табаке будет
интересно узнать, проявляют ли другие виды растений типичные
поперечная модель полимеризации лигнина откладывает необычные структуры лигнина
аналогичны наблюдаемым в табаке в ответ на генетические
модификации.
Это первое сообщение о растительном лигнине, содержащем высокое
пропорции концевых групп α-эфиров и циннамоилового спирта, обнаруженные в
синтетические лигнины, вызванные объемной полимеризацией, а не низким
концентрации этих структур в лигнинах, полученных из типичных
Обычно считается, что в растениях происходит процесс сложения поперек.Данный
эта необычная схема биосинтеза лигнина в табаке будет
интересно узнать, проявляют ли другие виды растений типичные
поперечная модель полимеризации лигнина откладывает необычные структуры лигнина
аналогичны наблюдаемым в табаке в ответ на генетические
модификации.
Хотя можно было ожидать, что подавление ферментов в
путь биосинтеза лигнина должен снижать концентрацию лигнина
вызвано сокращением поставок предшественников, исследованиями с природными мутантами и
трансгенных растений показало, что результаты такого подавления
сильно зависят от вовлеченного фермента.Снижение активности CCR
Сообщалось о снижении концентрации лигнина в табаке и Арабидопсис (4, 12, 13). В противоречивом отчете Кадзита и др. (14) обнаружили, что концентрация лигнина не снижается
в трансгенном табаке с подавленным смыслом и антисмысловым CCR. Эта учеба
(14) также был необычен тем, что сообщал о сниженном соотношении S/G лигнина.
в то время как другие отчеты о трансгенных CCR со сниженной регуляцией обнаружили
повышенное соотношение S/G (4, 12, 13). В отличие от ситуации с
трансгенных CCR, все отчеты о трансгенных CAD с подавленной регуляцией
не обнаружили снижения отложений лигнина и последовательного снижения
Соотношение S/G лигнин (15-18).Очевидно, нам еще многому предстоит научиться
о контрольных точках пути биосинтеза лигнина.
В противоречивом отчете Кадзита и др. (14) обнаружили, что концентрация лигнина не снижается
в трансгенном табаке с подавленным смыслом и антисмысловым CCR. Эта учеба
(14) также был необычен тем, что сообщал о сниженном соотношении S/G лигнина.
в то время как другие отчеты о трансгенных CCR со сниженной регуляцией обнаружили
повышенное соотношение S/G (4, 12, 13). В отличие от ситуации с
трансгенных CCR, все отчеты о трансгенных CAD с подавленной регуляцией
не обнаружили снижения отложений лигнина и последовательного снижения
Соотношение S/G лигнин (15-18).Очевидно, нам еще многому предстоит научиться
о контрольных точках пути биосинтеза лигнина.
Было отмечено Ralph et al. (4) что хотя антисмысл
Трансформант CAD имел нормальный внешний вид и рост, за исключением
красно-коричневый цвет ксилемы, напоминающий мутанты коричневой средней жилки у кукурузы и
sorghum, антисмысловой CCR-трансформант имел измененный цвет ксилемы
(оранжево-коричневые) и сниженный рост с аномальной морфологией листьев и
разрушенные сосуды. Это явление аномального роста растений и
развитие некоторых трансгенных растений с измененным биосинтезом лигнина
сообщалось ранее.Аномальный рост, по-видимому, ограничен
к случаям, когда концентрация лигнина была заметно снижена
трансген (4, 12, 13, 19, 20). Аномалия, вероятно, связана
с ослабленной сосудистой системой, вызванной коллапсом сосудов (13). В
Напротив, даже очень резкие изменения в составе лигнина, такие как
полное отсутствие лигнина сирингильного типа, вызванное подавлением
ген ферулат-5-гидроксилазы (F5H) у арабидопсиса мутант fah2 (21), не изменили рост и развитие при
концентрация лигнина оставалась нормальной.Этот вывод говорит о том, что
количество лигнина может играть более важную роль, чем состав лигнина
и структура общего роста. Улучшение понимания того, насколько
потребность растений в лигнине для нормального роста и развития будет иметь большое
важное значение для успешного управления биосинтезом лигнина.
Достигнут значительный прогресс в модификации лигнина
биосинтез посредством биотехнологии; тем не менее, критический вопрос
принесут ли модификации желаемую практическую пользу. В области улучшения переваримости клеточных стенок кормовых культур
результаты были смешанными. Бернар Вайль и др. (22, 23)
сообщили об улучшении усвояемости клеточной стенки O -метилтрансфераза (OMT) и CAD трансгенные растения табака.
Повышенная усвояемость была приписана измененному лигнину.
состава (уменьшенное соотношение S/G), так как концентрация лигнина не
изменились у этих трансгеников. Напротив, Sewalt et al. (24) объяснили улучшенную усвояемость клеточной стенки
фенилаланин-аммиак-лиаза и трансгены табака ОМТ к наблюдаемым
снижение концентрации лигнина, а не измененный лигнин
состав этих трансформантов.Наконец, недавняя работа в нашей
лаборатория с мутантом F5H Arabidopsis не обнаружена
изменение усвояемости клеточной стенки, даже если состав лигнина был
резко перешел от смешанного сирингил/гваяциллигнина к
сирингил-свободный лигнин этой мутацией (25). Предстоит дальнейшая работа
сделано, чтобы определить, какие типы модификаций лигнина улучшат
усвояемость клеточной стенки и как добиться таких изменений с помощью
биотехнология.
 Однако данные мутантов по коричневой средней жилке явно
указывают на то, что изменение лигнификации может привести к кормовым культурам с
улучшенная усвояемость клеточной стенки.
Однако данные мутантов по коричневой средней жилке явно
указывают на то, что изменение лигнификации может привести к кормовым культурам с
улучшенная усвояемость клеточной стенки.
Также имеется ограниченная информация о влиянии генетического
манипуляции с лигнификацией влияют на процесс варки целлюлозы. Халпин и др. (26) сообщили, что трансгенные
табак требовал меньше отбеливания для производства целлюлозы. Вскоре после этого
Баухер и др. (16) сообщили, что подавление ИБС
трансгенный тополь демонстрировал пониженное число Каппа во время варки целлюлозы,
что указывает на более эффективное удаление лигнина, как у 3-месячных проростков
и через 2 года роста.Недавно было обнаружено, что подавление O -трансформанты метилтрансферазы тополя имели более высокую каппа
номера для производства целлюлозы (W. Boerjan, личное сообщение) с указанием
что восстановленный лигнин, богатый сирингилом, дает структуру лигнина менее
поддается экстракции, чем богатый альдегидами лигнин CAD
трансформанты.
Ясно, что лигнификацию можно значительно изменить за счет биотехнологии, хотя фенотип полученных растений не всегда предсказуем из-за сложной природы биосинтетического путь лигнификации.Однако такие генетические манипуляции повысили нашу осведомленность о тонкостях и сложности лигнина синтез. Ральф и др. (4) теперь увеличили наши знания о типах предшественников растений, которые могут входить в состав лигнина в ответ на генетические манипуляции. С увеличением понимания лигнификации представляется разумным ожидать, что полезные технологии появится для улучшения производства бумажной массы и использования кормов волокнистых культур в недалеком будущем.
Сноски
↵‡ Кому следует направлять запросы на перепечатку по адресу: U.S. Департамент сельского хозяйства и Служба сельскохозяйственных исследований, 411 Borlaug Hall, 1991 Upper Buford Circle, St. Paul, MN 55108. jungx002{at}maroon.tc.umn.edu.
Дополнение к этому Комментарию начинается на странице 12803.
- Copyright © 1998, Национальная академия наук
Транскрипционный ландшафт сильно одревесневших стеблей тополя при одноклеточном разрешении | Геномная биология
Лукас В.Дж., Грувер А., Лихтенбергер Р., Фурута К., Ядав С.Р., Хелариутта Ю. и др. Сосудистая система растений: эволюция, развитие и функции. J Integr Plant Biol. 2013;55(4):294–388. https://doi.org/10.1111/jipb.12041.
КАС Статья пабмед Google Scholar
Miyashima S, Sebastian J, Lee JY, Helariutta Y. Функция стволовых клеток во время развития сосудов растений. EMBO J. 2013;32(2):178–93. https://doi.org/10.1038/emboj.2012.301.
КАС Статья пабмед Google Scholar
De Rybel B, Mähönen AP, Helariutta Y, Weijers D. Развитие сосудов растений: от ранней спецификации до дифференциации. Nat Rev Mol Cell Biol. 2016;17(1):30–40. https://doi.org/10.1038/nrm.2015.6.
КАС Статья пабмед Google Scholar
Бар-Он Ю.М., Филлипс Р., Майло Р.Распределение биомассы на Земле. ПНАС. 2018;115(25):6506–11. https://doi.org/10.1073/pnas.1711842115.
КАС Статья пабмед ПабМед Центральный Google Scholar
Миентс М.Дж., Ватанабэ Ю., Сэмюэлс А.Л. Клеточная биология биосинтеза вторичной клеточной стенки. Энн Бот. 2018;121(6):1107–25. https://doi.org/10.1093/aob/mcy005.
КАС Статья пабмед ПабМед Центральный Google Scholar
Schuetz M, Smith R, Ellis B. Спецификация тканей ксилемы, формирование паттерна и механизмы дифференцировки. J Опытный бот. 2013;64(1):11–31. https://doi.org/10.1093/jxb/ers287.
КАС Статья пабмед Google Scholar

Ван Х-З, Диксон Р.А. Выключатели для вторичного биосинтеза клеточной стенки. Мол завод. 2012;5(2):297–303. https://doi.org/10.1093/mp/ssr098.
КАС Статья пабмед Google Scholar
Чжун Р., Маккарти Р.Л., Хагигат М., Е З-Х. Главные переключатели MYB тополя связываются с сайтом SMRE и активируют программу биосинтеза вторичной стенки во время формирования древесины. ПЛОС Один. 2013;8(7):e69219. https://doi.org/10.1371/journal.pone.0069219.
КАС Статья пабмед ПабМед Центральный Google Scholar
Zhang J, Elo A, Helariutta Y. Arabidopsis как модель образования древесины. Курр Опин Биотехнолог.2011;22(2):293–9. https://doi.org/10.1016/j.copbio.2010.11.008.
Чжан Дж., Се М., Тускан Г.А., Мучеро В., Чен Дж.-Г. Последние достижения в транскрипционной регуляции биосинтеза вторичной клеточной стенки у древесных растений. Фронт завод науч. 2018;9:1535. https://doi.org/10.3389/fpls.2018.01535.
Фронт завод науч. 2018;9:1535. https://doi.org/10.3389/fpls.2018.01535.
Артикул пабмед ПабМед Центральный Google Scholar
Zhao C, He Y, Yu Y, Zhou M, Zhao L, Xia X, et al.Транскриптомный анализ сезонной экспрессии генов и регуляции во время развития ксилемы у гибридного тополя Shanxin ( Populus davidiana × Populus bolleana ). Леса. 2021;12(4):451. https://doi.org/10.3390/f12040451.
Артикул Google Scholar
Аренд М., Фромм Дж. Сезонные изменения реакции развития клеток древесины тополя на засуху. Физиол дерева. 2007;27(7):985–92. https://doi.org/10.1093/treephys/27.7.985.
Артикул пабмед Google Scholar
Камарго Э.Л., Плоет Р., Кассан-Ванг Х., Муне Ф., Грима-Петтенати Дж. Копание в древесине: новый взгляд на регуляцию образования древесины у древесных пород. Рекламный бот Res. 2019;89:201–33. https://doi.org/10.1016/bs.abr.2018.11.007.
Рекламный бот Res. 2019;89:201–33. https://doi.org/10.1016/bs.abr.2018.11.007.
КАС Статья Google Scholar
Shi R, Wang JP, Lin Y-C, Li Q, Sun Y-H, Chen H, et al.Тканевые и клеточные коэкспрессивные сети факторов транскрипции и генов компонентов древесины у Populus trichocarpa . Планта. 2017;245(5):927–38. https://doi.org/10.1007/s00425-016-2640-1.
КАС Статья пабмед Google Scholar
Sundell D, Street NR, Kumar M, Mellerowicz EJ, Kucukoglu M, Johnsson C, et al. AspWood: профили транскриптома с высоким пространственным разрешением выявляют нехарактерную модульность формирования древесины у Populus tremula .Растительная клетка. 2017;29(7):1585–604. https://doi.org/10.1105/tpc.17.00153.
КАС Статья пабмед ПабМед Центральный Google Scholar

Chen HC, Song J, Wang JP, Lin Y-C, Ducoste J, Shuford CM, et al. Системная биология биосинтеза лигнина у Populus trichocarpa : гетеромерная 4-кумаровая кислота: образование, регуляция и численное моделирование белковых комплексов кофермента А-лигазы. Растительная клетка. 2014;26(3):876–93.https://doi.org/10.1105/tpc.113.119685.
КАС Статья пабмед ПабМед Центральный Google Scholar
Goué N, Lesage-Descauses MC, Mellerowicz EJ, Magel E, Label P, Sundberg B. Микрогеномный анализ выявил характерные для типа клеток паттерны экспрессии генов между лучевыми и веретенообразными инициалями в камбиальной меристеме Populus . Новый Фитол. 2008;180(1):45–56. https://doi.org/10.1111/j.1469-8137.2008.02556.х.
КАС Статья пабмед Google Scholar
Ислам С., Зейсель А., Джуст С., Ла Манно Г., Заджак П., Каспер М. и др. Количественная последовательность одноклеточной РНК с уникальными молекулярными идентификаторами. Нат Методы. 2014;11(2):163–6. https://doi.org/10.1038/nmeth.2772.
Нат Методы. 2014;11(2):163–6. https://doi.org/10.1038/nmeth.2772.
КАС Статья пабмед Google Scholar
Macosko EZ, Basu A, Satija R, Nemesh J, Shekhar K, Goldman M, et al.Высокопараллельное профилирование полногеномной экспрессии отдельных клеток с использованием нанолитровых капель. Клетка. 2015;161(5):1202–14. https://doi.org/10.1016/j.cell.2015.05.002.
КАС Статья пабмед ПабМед Центральный Google Scholar
Seyfferth C, Renema J, Wendrich JR, Eekhout T, Seurinck R, Vandamme N, et al. Достижения и возможности транскриптомики одиночных клеток для исследований растений. Annu Rev Plant Biol. 2021;72(1):847–66.https://doi.org/10.1146/annurev-arplant-081720-010120.
КАС Статья пабмед Google Scholar
Shaw R, Tian X, Xu J. Анализ транскриптома одной клетки у растений: достижения и проблемы. Мол завод. 2020;14(1):115–26. https://doi.org/10.1016/j.molp.2020.10.012.
Мол завод. 2020;14(1):115–26. https://doi.org/10.1016/j.molp.2020.10.012.
КАС Статья пабмед Google Scholar
Liu Q, Liang Z, Feng D, Jiang S, Wang Y, Du Z и др.Транскрипционный ландшафт корней риса при одноклеточном разрешении. Мол завод. 2021;14(3):384–94. https://doi.org/10.1016/j.molp.2020.12.014.
КАС Статья пабмед Google Scholar
Чжан Т-К, Сюй З-Г, Шан Г-Д, Ван Дж-В. Секвенирование одноклеточной РНК описывает ландшафт развития корня Arabidopsis . Мол завод. 2019;12(5):648–60. https://doi.org/10.1016/j.molp.2019.04.004.
КАС Статья пабмед Google Scholar
Саттерли Дж.В., Стрэйбл Дж., Скэнлон М.Дж. Организация и дифференцировка стволовых клеток растений при разрешении одной клетки. ПНАС. 2020;117(52):33689–99. https://doi.org/10.1073/pnas. 2018788117.
2018788117.
КАС Статья пабмед ПабМед Центральный Google Scholar
Tian C, Du Q, Xu M, Du F, Jiao Y. Одноядерная РНК-секвенция разрешает пространственно-временные траектории развития в верхушке побега томата. БиоРксив. 2020.
Чжан Т.К., Чен Ю., Ван Дж.В. Одноклеточный анализ верхушки вегетативного побега Arabidopsis . Ячейка Дев. 2021;56(7):1056–74. https://doi.org/10.1016/j.devcel.2021.02.021.
КАС Статья пабмед Google Scholar
Kim J-Y, Symeonidi E, Pang TY, Denyer T, Weidauer D, Bezrutczyk M, et al. Различная идентичность клеток флоэмы листа, выявленная с помощью транскриптомики отдельных клеток. Растительная клетка. 2021;33(3):511–30.https://doi.org/10.1093/plcell/koaa060.
Артикул пабмед ПабМед Центральный Google Scholar

Xu X, Crow M, Rice BR, Li F, Harris B, Liu L, et al. Секвенирование одноклеточной РНК развивающихся початков кукурузы облегчает функциональный анализ и обнаружение генов-кандидатов признаков. Ячейка Дев. 2021;56(4):557–68. https://doi.org/10.1016/j.devcel.2020.12.015.
КАС Статья пабмед Google Scholar
Нелмс Б., Уолбот В. Определение программы развития, ведущей к мейозу у кукурузы. Наука. 2019;364(6435):52–6. https://doi.org/10.1126/science.aav6428.
КАС Статья пабмед Google Scholar
Shulse CN, Cole BJ, Ciobanu D, Lin J, Yoshinaga Y, Gouran M, et al. Высокопроизводительное профилирование одноклеточного транскриптома типов растительных клеток. Сотовые отчеты. 2019;27(7):2241–7. https://doi.org/10.1016/j.celrep.2019.04.054.
КАС Статья пабмед Google Scholar

Denyer T, Ma X, Klesen S, Scacchi E, Nieselt K, Timmermans MC. Пространственно-временные траектории развития в корне Arabidopsis , выявленные с помощью высокопроизводительного секвенирования одноклеточной РНК. Ячейка Дев. 2019;48(6):840–52. https://doi.org/10.1016/j.devcel.2019.02.022.
КАС Статья пабмед Google Scholar
Ryu KH, Huang L, Kang HM, Schiefelbein J. Секвенирование одноклеточной РНК разрешает молекулярные отношения между отдельными растительными клетками. Завод Физиол. 2019;179(4):1444–56. https://doi.org/10.1104/pp.18.01482.
КАС Статья пабмед ПабМед Центральный Google Scholar
Liu Z, Zhou Y, Guo J, Li J, Tian Z, Zhu Z, et al. Глобальное динамическое молекулярное профилирование развития клеток устьичной линии с помощью секвенирования одноклеточной РНК.Мол завод. 2020;13(8):1178–93. https://doi.org/10. 1016/j.molp.2020.06.010.
1016/j.molp.2020.06.010.
КАС Статья пабмед Google Scholar
Lopez-Anido CB, Vaten A, Smoot NK, Sharma N, Guo V, Gong Y, et al. Одноклеточное разрешение траекторий клонов устьичной линии Arabidopsis и развивающегося листа. Ячейка Дев. 2021;56(7):1043–55. https://doi.org/10.1016/j.devcel.2021.03.014.
КАС Статья пабмед Google Scholar
Лю Х., Ху Д., Ду П., Ван Л., Лян Х., Ли Х. и др. Одноклеточная РНК-секвенция описывает ландшафт транскриптома и идентифицирует критические факторы транскрипции в листовой пластинке аллотетраплоидного арахиса ( Arachis hypogaea L.). Plant Biotechnol J. 2021;19(11):2261–76.
КАС Статья Google Scholar
Gala HP, Lanctot A, Jean-Baptiste K, Guiziou S, Chu JC, Zemke JE, et al. Вид транскриптома одной клетки во время инициации латерального корня у Arabidopsis thaliana . Растительная клетка. 2021;33(7):2197–220. https://doi.org/10.1093/plcell/koab101.
Растительная клетка. 2021;33(7):2197–220. https://doi.org/10.1093/plcell/koab101.
Артикул пабмед Google Scholar
Li H, Dai X, Huang X, Xu M, Wang Q, Yan X и др. Секвенирование РНК одиночных клеток выявило клеточный атлас ксилемы с высоким разрешением у Populus . J Integr Plant Biol. 2021;00(11):1–16. https://doi.org/10.1111/jipb.13159.
Артикул Google Scholar
Lin Y-C, Li W, Chen H, Li Q, Sun Y-H, Shi R и другие. Простая система протопластов ксилемы с улучшенной пропускной способностью для изучения образования древесины. Нат Проток. 2014;9(9):2194–205. https://doi.org/10.1038/nprot.2014.147.
КАС Статья пабмед Google Scholar
Wu AM, Hörnblad E, Voxeur A, Gerber L, Rihouey C, Lerouge P, et al. Анализ пар генов гликозилтрансфераз Arabidopsis IRX9/IRX9-L и IRX14/IRX14-L показывает критический вклад в биосинтез глюкуроноксилана гемицеллюлозы. Завод Физиол. 2010;153(2):542–54. https://doi.org/10.1104/pp.110.154971.
Завод Физиол. 2010;153(2):542–54. https://doi.org/10.1104/pp.110.154971.
КАС Статья пабмед ПабМед Центральный Google Scholar
Lee C, O’Neill MA, Tsumuraya Y, Darvill AG, Ye Z-H. Мутант с неправильной ксилемой9 лишен активности ксилан-ксилозилтрансферазы. Физиология клеток растений. 2007;48(11):1624–34. https://doi.org/10.1093/pcp/pcm135.
КАС Статья пабмед Google Scholar
Lasserre E, Jobet E, Llauro C, Delseny M. AtERF38 ( At2g35700 ), ген транскрипционного фактора семейства AP2/ERF из Arabidopsis thaliana , экспрессируется в определенных типах клеток корней, стеблей и семян, подвергающихся суберизация. Завод Физиол Биохим. 2008;46(12):1051–61. https://doi.org/10.1016/j.plaphy.2008.07.003.
Берте С., Демон-Коле Н., Полле Б., Бидзинский П., Сезар Л., Ле Брис П. и др. Разрушение LACCASE4 и 17 приводит к тканеспецифическим изменениям лигнификации стеблей Arabidopsis thaliana . Растительная клетка. 2011;23(3):1124–37. https://doi.org/10.1105/tpc.110.082792.
Растительная клетка. 2011;23(3):1124–37. https://doi.org/10.1105/tpc.110.082792.
КАС Статья пабмед ПабМед Центральный Google Scholar
Li E, Bhargava A, Qiang W, Friedmann MC, Forneris N, Savidge RA, et al. Ген класса II KNOX KNAT7 отрицательно регулирует образование вторичной стенки у Arabidopsis и функционально консервативен у Populus . Новый Фитол. 2012;194(1):102–15.https://doi.org/10.1111/j.1469-8137.2011.04016.x.
КАС Статья пабмед Google Scholar
Tan TT, Endo H, Sano R, Kurata T, Yamaguchi M, Ohtani M, et al. Факторы транскрипции VND1-VND3 способствуют формированию сосудов ксилемы семядолей. Завод Физиол. 2018;176(1):773–89. https://doi.org/10.1104/pp.17.00461.
КАС Статья пабмед Google Scholar
Feuillet C, Lauvergeat V, Deswarte C, Pilate G, Boudet A, Grima-Pettenati J. Ткане- и клеточно-специфическая экспрессия промотора коричной алкогольдегидрогеназы в трансгенных растениях тополя. Завод Мол Биол. 1995;27(4):651–67. https://doi.org/10.1007/BF00020220.
Ткане- и клеточно-специфическая экспрессия промотора коричной алкогольдегидрогеназы в трансгенных растениях тополя. Завод Мол Биол. 1995;27(4):651–67. https://doi.org/10.1007/BF00020220.
КАС Статья пабмед Google Scholar
Bonke M, Thitamadee S, Mähönen AP, Hauser M-T, Helariutta Y. APL регулирует идентичность сосудистой ткани у Arabidopsis .Природа. 2003; 426 (6963): 181–6. https://doi.org/10.1038/nature02100.
КАС Статья пабмед Google Scholar
Dinant S, Clark AM, Zhu Y, Vilaine F, Palauqui J-C, Kusiak C, et al. Разнообразие надсемейства лектинов флоэмы (белок флоэмы 2) у покрытосеменных растений. Завод Физиол. 2003;131(1):114–28. https://doi.org/10.1104/pp.013086.
КАС Статья пабмед ПабМед Центральный Google Scholar
Sauer N, Stolz J. SUC1 и SUC2: два переносчика сахарозы из Arabidopsis thaliana ; экспрессия и характеристика в пекарских дрожжах и идентификация меченого гистидином белка. Плант Дж. 1994;6(1):67–77. https://doi.org/10.1046/j.1365-313X.1994.6010067.x.
Плант Дж. 1994;6(1):67–77. https://doi.org/10.1046/j.1365-313X.1994.6010067.x.
КАС Статья пабмед Google Scholar
Pelissier HC, Peters WS, Collier R. Bel AJv, Knoblauch M: Маркировка GFP белков окклюзии ситовидных элементов (SEO) приводит к появлению зеленых флуоресцентных форизом.Физиология клеток растений. 2008;49(11):1699–710. https://doi.org/10.1093/pcp/pcn141.
КАС Статья пабмед ПабМед Центральный Google Scholar
Chardon F, Bedu M, Calenge F, Klemens PA, Spinner L, Clement G, et al. Содержание фруктозы в листьях контролируется вакуолярным переносчиком SWEET17 в Arabidopsis . Карр Биол. 2013;23(8):697–702. https://doi.org/10.1016/j.cub.2013.03.021.
КАС Статья пабмед Google Scholar
Long Y, Goedhart J, Schneijderberg M, Terpstra I, Shimotohno A, Bouchet BP, et al. SCARECROW-LIKE 23 и SCARECROW совместно определяют судьбу энтодермальных клеток, но отчетливо контролируют движение SHORT-ROOT. Плант Дж. 2015;84(4):773–84. https://doi.org/10.1111/tpj.13038.
SCARECROW-LIKE 23 и SCARECROW совместно определяют судьбу энтодермальных клеток, но отчетливо контролируют движение SHORT-ROOT. Плант Дж. 2015;84(4):773–84. https://doi.org/10.1111/tpj.13038.
КАС Статья пабмед Google Scholar
Hassan H, Scheres B, Blilou I. JACKDAW контролирует формирование эпидермального паттерна в корневой меристеме Arabidopsis посредством неклеточно-автономного механизма.Разработка. 2010;137(9):1523–1529. https://doi.org/10.1242/dev.048777.
КАС Статья пабмед Google Scholar
David LC, Berquin P, Kanno Y, Seo M, Daniel-Vedele F, Ferrario-Méry S. Доступность N модулирует роль NPF3. 1, транспортер гиббереллина, в GA-опосредованных фенотипах у Arabidopsis . Планта. 2016;244(6):1315–28. https://doi.org/10.1007/s00425-016-2588-1.
КАС Статья пабмед Google Scholar

Соццани Р., Куи Х., Морено-Рисуэно М., Буш В., Ван Норман Дж., Верну Т. и другие. Пространственно-временная регуляция генов клеточного цикла с помощью SHORTROOT связывает формирование паттерна и рост. Природа. 2010;466(7302):128–32. https://doi.org/10.1038/nature09143.
КАС Статья пабмед ПабМед Центральный Google Scholar
Гельднер Н. Энтодерма. Annu Rev Plant Biol. 2013;64(1):531–58. https://doi.org/10.1146/annurev-arplant-050312-120050.
КАС Статья пабмед Google Scholar
Molina I, Li-Beisson Y, Beisson F, Ohlrogge JB, Pollard M. Идентификация Arabidopsis ферулоил-кофермента трансферазы A, необходимой для синтеза суберина. Завод Физиол. 2009;151(3):1317–28. https://doi.org/10.1104/pp.109.144907.
КАС Статья пабмед ПабМед Центральный Google Scholar

Naseer S, Lee Y, Lapierre C, Franke R, Nawrath C, Geldner N. Диффузионный барьер из полоски Casparian в Arabidopsis изготовлен из полимера лигнина без суберина. ПНАС. 2012;109(25):10101–6. https://doi.org/10.1073/pnas.1205726109.
Артикул пабмед ПабМед Центральный Google Scholar
Joubès J, Raffaele S, Bourdenx B, Garcia C, Laroche-Traineau J, Moreau P, et al. Семейство генов элонгазы ЖКОДЦ в Arabidopsis thaliana : филогенетический анализ, трехмерное моделирование и профилирование экспрессии.Завод Мол Биол. 2008;67(5):547–66. https://doi.org/10.1007/s11103-008-9339-z.
КАС Статья пабмед Google Scholar
Лю С., Сонг Т., Косма Д.К., Парсонс Э.П., Роуленд О., Дженкс М.А. Arabidopsis CER8 кодирует длинноцепочечную ацил-коасинтетазу 1 (LACS1), функции которой перекрываются с LACS2 в синтезе растительного воска и кутина. Плант Дж. 2009; 59 (4): 553–64. https://doi.org/10.1111/j.1365-313X.2009.03892.x.
Плант Дж. 2009; 59 (4): 553–64. https://doi.org/10.1111/j.1365-313X.2009.03892.x.
Masucci JD, Rerie WG, Foreman DR, Zhang M, Galway ME, Marks MD, et al. Ген гомеобокса GLABRA2 необходим для зависимой от положения дифференцировки клеток в эпидермисе корней Arabidopsis thaliana . Разработка. 1996;122(4):1253–60. https://doi.org/10.1242/dev.122.4.1253.
КАС Статья пабмед Google Scholar
Фишер У., Кукукоглу М., Хелариутта Ю., Бхалерао Р.П. Динамика активности камбиальных стволовых клеток.Annu Rev Plant Biol. 2019;70(1):293–319. https://doi.org/10.1146/annurev-arplant-050718-100402.
КАС Статья пабмед Google Scholar
Kucukoglu M, Chaabouni S, Zheng B, Mähönen AP, Helariutta Y, Nilsson O. Пептид, кодирующий Populus CLV3/ESR-RELATED 47 ( PttCLE47 ) способствует камбиальному развитию и вторичному xx . Новый Фитол. 2020;226(1):75–85. https://дои.орг/10.1111/nph.16331.
Новый Фитол. 2020;226(1):75–85. https://дои.орг/10.1111/nph.16331.
КАС Статья пабмед Google Scholar
Zhu Y, Song D, Xu P, Sun J, Li L. Ген A HD-ZIP III , PtrHB4 , необходим для развития межпучкового камбия у Populus . Plant Biotechnol J. 2018;16(3):808–17. https://doi.org/10.1111/pbi.12830.
КАС Статья пабмед Google Scholar
Zhu Y, Song D, Sun J, Wang X, Li L. PtrHB7 , ген HD-Zip класса III, играет критическую роль в регуляции дифференцировки сосудистого камбия у Populus . Мол завод. 2013;6(4):1331–43. https://doi.org/10.1093/mp/sss164.
Бреда А.С., Хазак О., Хардтке К.С. Активность OCTOPUS в протофлоэме корня определяется зарядом фосфозита, а не локализацией в направлении побега. ПНАС. 2017;114(28):E5721–30. https://doi.org/10.1073/pnas.1703258114.
КАС Статья пабмед ПабМед Центральный Google Scholar

Wallner ES, Tonn N, Shi D, Jouannet V, Greb T. SUPPRESSOR OF MAX2 1-LIKE 5 способствует образованию вторичной флоэмы во время радиального роста стебля. Плант Дж. 2020;102(5):903–15. https://doi.org/10.1111/tpj.14670.
Gu F, Bringmann M, Combs JR, Yang J, Bergmann DC, Nielsen E. Arabidopsis CSLD5 действует при формировании клеточных пластинок в зависимости от клеточного цикла. Растительная клетка. 2016;28(7):1722–37. https://doi.org/10.1105/tpc.16.00203.
Мюллер С., Фукс Э., Овецка М., Высоцкая-Диллер Дж., Бенфей П.Н., Хаузер М-Т. Два новых локуса, PLEIADE и HYADE , предполагают органоспецифическую регуляцию цитокинеза у Arabidopsis . Завод Физиол. 2002;130(1):312–24. https://doi.org/10.1104/pp.004416.
КАС Статья пабмед ПабМед Центральный Google Scholar
Strompen G, El Kasmi F, Richter S, Lukowitz W, Assaad FF, Jürgens G, et al. Ген Arabidopsis HINKEL кодирует родственный кинезину белок, участвующий в цитокинезе, и его экспрессия зависит от клеточного цикла. Карр Биол. 2002;12(2):153–8. https://doi.org/10.1016/S0960-9822(01)00655-8.
Ген Arabidopsis HINKEL кодирует родственный кинезину белок, участвующий в цитокинезе, и его экспрессия зависит от клеточного цикла. Карр Биол. 2002;12(2):153–8. https://doi.org/10.1016/S0960-9822(01)00655-8.
КАС Статья пабмед Google Scholar
Boruc J, Mylle E, Duda M, De Clercq R, Rombauts S, Geelen D, et al. Систематическая локализация основных белков клеточного цикла Arabidopsis выявляет новые комплексы клеточного деления.Завод Физиол. 2010;152(2):553–65. https://doi.org/10.1104/pp.109.148643.
КАС Статья пабмед ПабМед Центральный Google Scholar
Boudolf V, Rombauts S, Naudts M, Inzé D, De Veylder L. Идентификация новых циклинзависимых киназ, взаимодействующих с белком CKS1 Arabidopsis . J Опытный бот. 2001; 52 (359): 1381–2. https://doi.org/10.1093/jexbot/52.359.1381.
КАС Статья пабмед Google Scholar
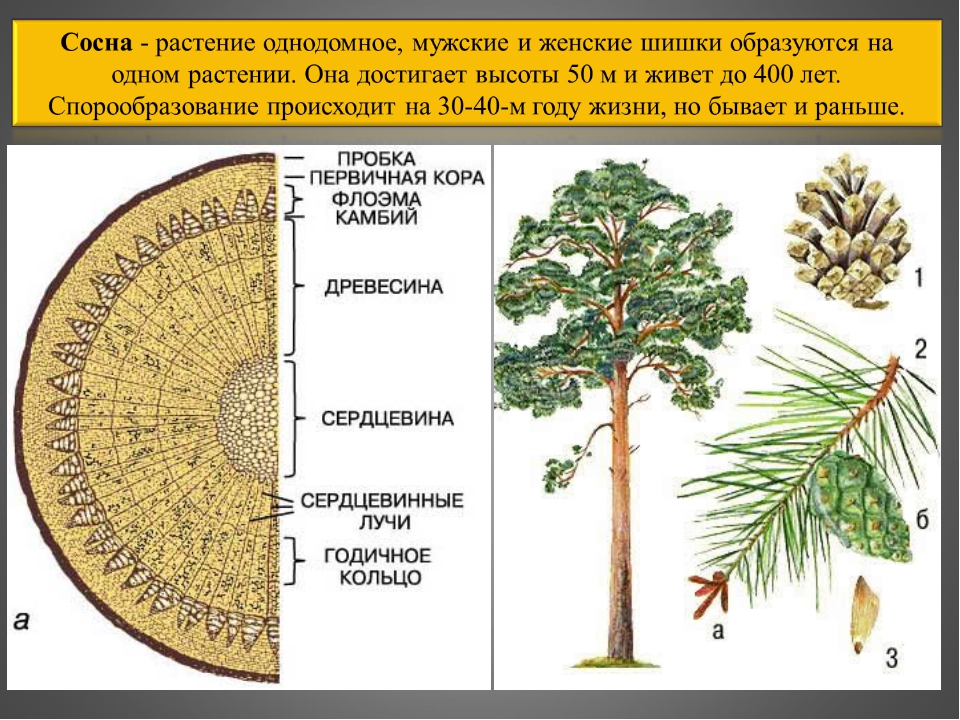
Qiu X, Mao Q, Tang Y, Wang L, Chawla R, Pliner HA и др. Обратное вложение графа разрешает сложные траектории с одной ячейкой. Нат Методы. 2017;14(10):979–82. https://doi.org/10.1038/nmeth.4402.
КАС Статья пабмед ПабМед Центральный Google Scholar
Kleine-Vehn J, Dhonukshe P, Swarup R, Bennett M, Friml J. Субклеточный перенос переносчика притока ауксина Arabidopsis AUX1 использует новый путь, отличный от PIN1.Растительная клетка. 2006;18(11):3171–81. https://doi.org/10.1105/tpc.106.042770.
КАС Статья пабмед ПабМед Центральный Google Scholar
Liu L, Liu C, Hou X, Xi W, Shen L, Tao Z, et al. FTIP1 является важным регулятором, необходимым для транспорта флоригена. PLoS биол. 2012;10(4):e1001313. https://doi.org/10.1371/journal.pbio.1001313.
КАС Статья пабмед ПабМед Центральный Google Scholar

Яо Р., Мин З., Ян Л., Ли С., Ван Ф., Ма С. и др. DWARF14 — неканонический гормональный рецептор стриголактона. Природа. 2016; 536(7617):469–73. https://doi.org/10.1038/nature19073.
КАС Статья пабмед Google Scholar
Ван Л., Ван Б., Ю. Х., Го Х., Линь Т., Коу Л. и др. Транскрипционная регуляция передачи сигналов стриголактона у Arabidopsis . Природа. 2020;583(7815):277–81. https://doi.org/10.1038/с41586-020-2382-х.
КАС Статья пабмед Google Scholar
Barratt DP, Kölling K, Graf A, Pike M, Calder G, Findlay K, et al. Синтаза каллозы GSL7 необходима для нормального транспорта флоэмы и роста соцветий у Arabidopsis . Завод Физиол. 2011;155(1):328–41. https://doi.org/10.1104/pp.110.166330.
КАС Статья пабмед Google Scholar
Шёлунд РД. Элемент сита флоэмы: через него протекает река. Растительная клетка. 1997;9(7):1137–46. https://doi.org/10.1105/tpc.9.7.1137.
Элемент сита флоэмы: через него протекает река. Растительная клетка. 1997;9(7):1137–46. https://doi.org/10.1105/tpc.9.7.1137.
КАС Статья пабмед ПабМед Центральный Google Scholar
Хан Дж.А., Ван К., Шёлунд Р.Д., Шульц А., Томпсон Г.А. Ранний нодулин-подобный белок накапливается в плазматической мембране ситовидного элемента Arabidopsis . Завод Физиол. 2007;143(4):1576–89. https://doi.org/10.1104/стр.106.092296.
КАС Статья пабмед ПабМед Центральный Google Scholar
Song, N. Характеристика раннего нодулинового белка Arabidopsis , специфичного к ситовому элементу. Дипломная работа. Университет штата Оклахома, кафедра биохимии и молекулярной биологии; 2010.
Google Scholar
Деттмер Дж., Урсаче Р., Кампильо А., Мияшима С., Белевич И., О’Реган С. и др.CHOLINE TRANSPORTER-LIKE1 необходим для развития ситовидных пластинок, чтобы обеспечить связь между клетками на большие расстояния. Нац коммун. 2014;5(1):1–11. https://doi.org/10.1038/ncomms5276.
и др.CHOLINE TRANSPORTER-LIKE1 необходим для развития ситовидных пластинок, чтобы обеспечить связь между клетками на большие расстояния. Нац коммун. 2014;5(1):1–11. https://doi.org/10.1038/ncomms5276.
КАС Статья Google Scholar
Milhinhos A, Prestele J, Bollhöner B, Matos A, Vera-Sirera F, Rambla JL, et al. Уровни термоспермина контролируются ауксин-зависимым механизмом петли обратной связи в ксилеме Populus . Плант Дж.2013;75(4):685–98. https://doi.org/10.1111/tpj.12231.
КАС Статья пабмед Google Scholar
Morris H, Jansen S. Паренхима вторичной ксилемы – от классической терминологии к функциональным признакам. Журнал Лава. 2016;37(1):1–15. https://doi.org/10.1163/22941932-20160117.
Артикул Google Scholar
Секки Ф., Пальярани С., Звенецкий М.А.Функциональная роль клеток ксилемной паренхимы и аквапоринов при восстановлении после тяжелого водного стресса. Окружающая среда растительной клетки. 2017;40(6):858–71. https://doi.org/10.1111/pce.12831.
КАС Статья пабмед Google Scholar
Юань Ю, Тэн Кью, Чжун Р, Е З-Х. Белок ESK1, содержащий домен Arabidopsis DUF231, опосредует 2-O- и 3-O-ацетилирование ксилозильных остатков в ксилане. Физиология клеток растений. 2013;54(7):1186–99.https://doi.org/10.1093/pcp/pct070.
КАС Статья пабмед Google Scholar
Цао С., Го М., Ван С., Сюй В., Ши Т., Тонг Г. и др. Полногеномная характеристика семейства генов аспарагиновой протеазы (AP) у Populus trichocarpa и идентификация потенциальных PtAP, участвующих в формировании древесины. BMC Растение Биол. 2019;19(1):1–17. https://doi.org/10.1186/s12870-019-1865-0.
КАС Статья Google Scholar
Song D, Sun J, Li L. Различные роли белков PtrDUF579 в функции Populus и PtrDUF579-1 в пролиферации сосудистого камбия во время вторичного роста. Завод Мол Биол. 2014;85(6):601–12. https://doi.org/10.1007/s11103-014-0206-9.
Различные роли белков PtrDUF579 в функции Populus и PtrDUF579-1 в пролиферации сосудистого камбия во время вторичного роста. Завод Мол Биол. 2014;85(6):601–12. https://doi.org/10.1007/s11103-014-0206-9.
КАС Статья пабмед Google Scholar
Gui J, Lam PY, Tobimatsu Y, Sun J, Huang C, Cao S, et al. Специфическая для волокон регуляция биосинтеза лигнина улучшает качество биомассы Populus .Новый Фитол. 2020;226(4):1074–87. https://doi.org/10.1111/nph.16411.
КАС Статья пабмед ПабМед Центральный Google Scholar
Zhao Y, Song D, Sun J, Li L. Populus эндо-бета-манназа PtrMAN6 играет роль в координации ремоделирования клеточной стенки с подавлением вторичного утолщения стенки посредством генерации сигналов олигосахаридов. Плант Дж. 2013;74(3):473–85. https://doi.org/10.1111/tpj.12137.

Фанк В., Косицуп Б., Чжао С., Бирс EP. Ксилемная пептидаза XCP1 Arabidopsis представляет собой вакуолярный белок трахеарного элемента, который может быть ортологом папаина. Завод Физиол. 2002;128(1):84–94. https://doi.org/10.1104/pp.010514.
КАС Статья пабмед ПабМед Центральный Google Scholar
Ma T, Wang J, Zhou G, Yue Z, Hu Q, Chen Y, et al. Геномный взгляд на адаптацию к соли у пустынного тополя.Нац коммун. 2013;4(1):1–9. https://doi.org/10.1038/ncomms3797.
КАС Статья Google Scholar
Tuskan GA, Difazio S, Jansson S, Bohlmann J, Grigoriev I, Hellsten U, et al. Геном тополя черного, Populus trichocarpa (Torr. & Grey). Наука. 2006;313(5793):1596–604. https://doi.org/10.1126/science.1128691.
КАС Статья пабмед Google Scholar
Блан Г.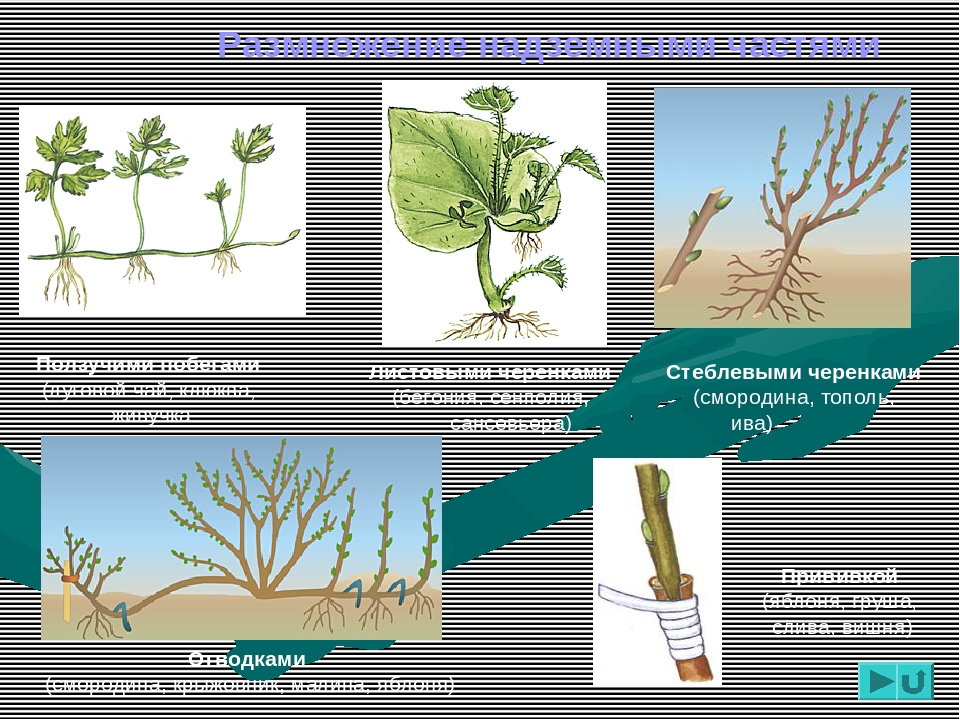 , Вулф К.Х. Функциональная дивергенция дуплицированных генов, образованных полиплоидией в ходе эволюции Arabidopsis . Растительная клетка. 2004;16(7):1679–91. https://doi.org/10.1105/tpc.021410.
, Вулф К.Х. Функциональная дивергенция дуплицированных генов, образованных полиплоидией в ходе эволюции Arabidopsis . Растительная клетка. 2004;16(7):1679–91. https://doi.org/10.1105/tpc.021410.
КАС Статья пабмед ПабМед Центральный Google Scholar
Xu W, Cheng H, Zhu S, Cheng J, Ji H, Zhang B, et al. Функциональное понимание синтаз целлюлозы вторичной клеточной стенки у Populus trichocarpa с помощью индуцированных Cas9 / гРНК нокаутов генов.Новый Фитол. 2021;231(4):1478–95. https://doi.org/10.1111/nph.17338.
КАС Статья пабмед ПабМед Центральный Google Scholar
Abbas M, Peszlen I, Shi R, Kim H, Katahira R, Kafle K, et al. Участие CesA4 , CesA7-A/B и CesA8-A/B в формировании вторичной стенки в древесине Populus trichocarpa . Физиол дерева. 2020;40(1):73–89. https://doi. org/10.1093/treephys/tpz020.
org/10.1093/treephys/tpz020.
КАС Статья пабмед Google Scholar
Farmer A, Thibivilliers S, Ryu KH, Schiefelbein J, Libault M. Секвенирование одноядерной РНК и ATAC выявило влияние доступности хроматина на экспрессию генов в корнях Arabidopsis на уровне одной клетки. Мол завод. 2021;14(3):372–83. https://doi.org/10.1016/j.molp.2021.01.001.
КАС Статья пабмед Google Scholar
Ding J, Adiconis X, Simmons SK, Kowalczyk MS, Hession CC, Marjanovic ND, et al. Систематическое сравнение одноклеточных и одноядерных методов секвенирования РНК. Нац биотехнолог. 2020;38(6):737–46. https://doi.org/10.1038/s41587-020-0465-8.
КАС Статья пабмед ПабМед Центральный Google Scholar
Grindberg RV, Yee-Greenbaum JL, McConnell MJ, Novotny M, O’Shaughnessy AL, Lambert GM, et al. Секвенирование РНК из отдельных ядер.ПНАС. 2013;110(49):19802–7. https://doi.org/10.1073/pnas.1319700110.
Секвенирование РНК из отдельных ядер.ПНАС. 2013;110(49):19802–7. https://doi.org/10.1073/pnas.1319700110.
КАС Статья пабмед ПабМед Центральный Google Scholar
Zhang L, Zhao J, Bi H, Yang X, Zhang Z, Su Y, et al. Биоинформационный анализ организации хроматина и предвзятой экспрессии дуплицированных генов между двумя тополями с общей полногеномной дупликацией. Хортик Рез. 2021;8(1):1–12. https://doi.org/10.1038/s41438-021-00494-2.
КАС Статья Google Scholar
Сатия Р., Фаррелл Дж. А., Геннерт Д., Шир А. Ф., Регев А. Пространственная реконструкция данных экспрессии генов одноклеточных. Нац биотехнолог. 2015;33(5):495–502. https://doi.org/10.1038/nbt.3192.
КАС Статья пабмед ПабМед Центральный Google Scholar
McGinnis CS, Murrow LM, Gartner ZJ. DoubletFinder: обнаружение дублетов в данных секвенирования одноклеточной РНК с использованием искусственных ближайших соседей. Клеточные системы. 2019;8(4):329–37 e324.
Клеточные системы. 2019;8(4):329–37 e324.
КАС Статья Google Scholar
Батлер А., Хоффман П., Смиберт П., Папалекси Э., Сатия Р. Интеграция транскриптомных данных отдельных клеток в различных условиях, технологиях и видах. Нац биотехнолог. 2018;36(5):411–20. https://doi.org/10.1038/nbt.4096.
КАС Статья пабмед ПабМед Центральный Google Scholar
Хагверди Л., Лун А.Т., Морган М.Д., Мариони Дж.К.Пакетные эффекты в данных секвенирования одноклеточной РНК исправляются путем сопоставления взаимных ближайших соседей. Нац биотехнолог. 2018;36(5):421–7. https://doi.org/10.1038/nbt.4091.
КАС Статья пабмед ПабМед Центральный Google Scholar
Gutierrez, C. Цикл деления клеток Arabidopsis . Arabidopsis Book, 2009;e0120, 7. https://doi.org/10. 1199/tab.0120.
1199/tab.0120.
Ядав Р.К., Гирке Т., Пасала С., Се М., Редди Г.В.Карта экспрессии генов ниши стволовых клеток апикальной меристемы побега Arabidopsis . ПНАС. 2009;106(12):4941–6. https://doi.org/10.1073/pnas.03106.
Артикул пабмед ПабМед Центральный Google Scholar
Виллани А.С., Сатия Р., Рейнольдс Г., Саркизова С., Шекхар К., Флетчер Дж. и др. Одноклеточная РНК-секвенция выявляет новые типы дендритных клеток крови человека, моноцитов и предшественников. Наука. 2017;356(6335):eaah5573.
Артикул Google Scholar
Ван дер Маатен Л., Хинтон Г. Визуализация данных с использованием t-SNE. Дж. Мах Узнать Рез. 2008;9(11):2579–605.
Google Scholar
Becht E, McInnes L, Healy J, Dutertre C-A, Kwok IW, Ng LG, et al. Уменьшение размерности для визуализации данных одной ячейки с использованием UMAP. Нац биотехнолог. 2019;37(1):38–44. https://doi.org/10.1038/nbt.4314.
Нац биотехнолог. 2019;37(1):38–44. https://doi.org/10.1038/nbt.4314.
КАС Статья Google Scholar
Ким Д., Лангмид Б., Зальцберг С.Л. HISAT: быстродействующий выравниватель с низкими требованиями к памяти. Нат Методы. 2015;12(4):357–60. https://doi.org/10.1038/nmeth.3317.
КАС Статья пабмед ПабМед Центральный Google Scholar
Pertea M, Pertea GM, Antonescu CM, Chang T-C, Mendell JT, Salzberg SL.StringTie обеспечивает улучшенную реконструкцию транскриптома из чтений РНК-seq. Нац биотехнолог. 2015;33(3):290–5. https://doi.org/10.1038/nbt.3122.
КАС Статья пабмед ПабМед Центральный Google Scholar
Wang Y, Tang H, DeBarry JD, Tan X, Li J, Wang X, Lee TH, Jin H, Marler B, Guo H. MCScanX: набор инструментов для обнаружения и эволюционного анализа синтении и коллинеарности генов . Нуклеиновые Кислоты Res. 2012;40(7):e49-e49.https://doi.org/10.1093/nar/gkr1293.
Нуклеиновые Кислоты Res. 2012;40(7):e49-e49.https://doi.org/10.1093/nar/gkr1293.
Ли Г.С., Мэн З., Конг Х.З., Чен З.Д., Тейссен Г., Лу А.М. Характеристика цветочных гомеозисных генов-кандидатов класса A, B и E из базального покрытосеменного без околоцветника Chloranthus spicatus (Chloranthaceae). Дев Гены Эвол. 2005;215(9):437–49. https://doi.org/10.1007/s00427-005-0002-2.
КАС Статья пабмед Google Scholar
Heinz S, Benner C, Spann N, Bertolino E, Lin YC, Laslo P, et al.Простые комбинации транскрипционных факторов, определяющих происхождение, задают цис-регуляторные элементы, необходимые для идентичности макрофагов и В-клеток. Мол Ячейка. 2010;38(4):576–89. https://doi.org/10.1016/j.molcel.2010.05.004.
КАС Статья пабмед ПабМед Центральный Google Scholar
Kohl M, Wiese S, Warscheid B. Cytoscape: программное обеспечение для визуализации и анализа биологических сетей. Методы Мол Биол. 2011; 696: 291–303.https://doi.org/10.1007/978-1-60761-987-1_18.
Методы Мол Биол. 2011; 696: 291–303.https://doi.org/10.1007/978-1-60761-987-1_18.
КАС Статья пабмед Google Scholar
Ян З. PAML: пакет программ для филогенетического анализа методом максимального правдоподобия. Биоинформатика. 1997;13(5):555–6. https://doi.org/10.1093/bioinformatics/13.5.555.
КАС Статья Google Scholar
Chen Y, Tong S, Jiang Y, Ai F, Feng Y, Zhang J, Gong J, Qin J, Zhang Y, Zhu Y, Liu J, Ma T: Транскрипционный ландшафт сильно одревесневших стеблей тополя в одноклеточное разрешение.НГДК. БиоПроект. https://ngdc.cncb.ac.cn/bioproject/browse/PRJCA005543 (2021 г.).
Лигнификация вторичных клеточных стенок трахеид ели, связанная с продольной твердостью и модулем упругости, с использованием наноиндентирования
Цитируется по
1. Стратегии защиты семян мозгового эндокарпа Elaeocarpus ganitrus: Gradient motif 2. Локальные вариации содержания углеводов и матричного лигнина в бамбуковых стеблях механической сортировки
Локальные вариации содержания углеводов и матричного лигнина в бамбуковых стеблях механической сортировки
3. Сравнительные исследования механических свойств и микроструктуры внешней и сердцевинной древесины Pinus radiata D. Don
4. Поперечные и продольные константы упругости волокон целлюлозы в бумажных листах
5. Влияние генотипа и почвенного субстрата на качество древесины тополя, выращенного на рекультивированном участке добычи бурого угля
6. Атипичное одревеснение в кожевенной древесине восточной ( Дирка болотная )
7. Механическая характеристика клеточных стенок Pinus massoniana, зараженных грибами синей окраски, с использованием наноиндентирования in situ обзор
10. Анатомия пальмы ветряной мельницы (Trachycarpus Fortunei) и возможности ее применения
11. Наноиндентирование и характеристика воспламеняемости пяти видов биомассы рисовой шелухи для применения в биокомпозитах
12. 215 1 Мезокарпий бразильского ореха (
Бертолетия превосходная
) в качестве вдохновения для создания новых ударопрочных материалов
215 1 Мезокарпий бразильского ореха (
Бертолетия превосходная
) в качестве вдохновения для создания новых ударопрочных материалов
13. Наномеханические и топохимические изменения в древесине вяза древних деревянных конструкций в связи с естественным старением свойства наномеханических клеток
15. К разработке высокоэффективных композитов растительного волокна
16. Изменения в бамбуковом волокне, подвергнутом различным химическим обработкам и сушке вымораживанием, измеренные с помощью наноиндентирования , XRD и мокрая химия
18. Характеристика влагозависимых механических свойств органических материалов: контролируемое влажностью статическое и динамическое наноиндентирование клеточных стенок древесины
20. Оценка эффектов сжатия в сочетании с термической обработкой путем наноиндентирования (НИ) клеточных стенок тополя
21. Наноиндентирование клеточных стенок древесины: влияние подготовки образца и протокола индентирования
22.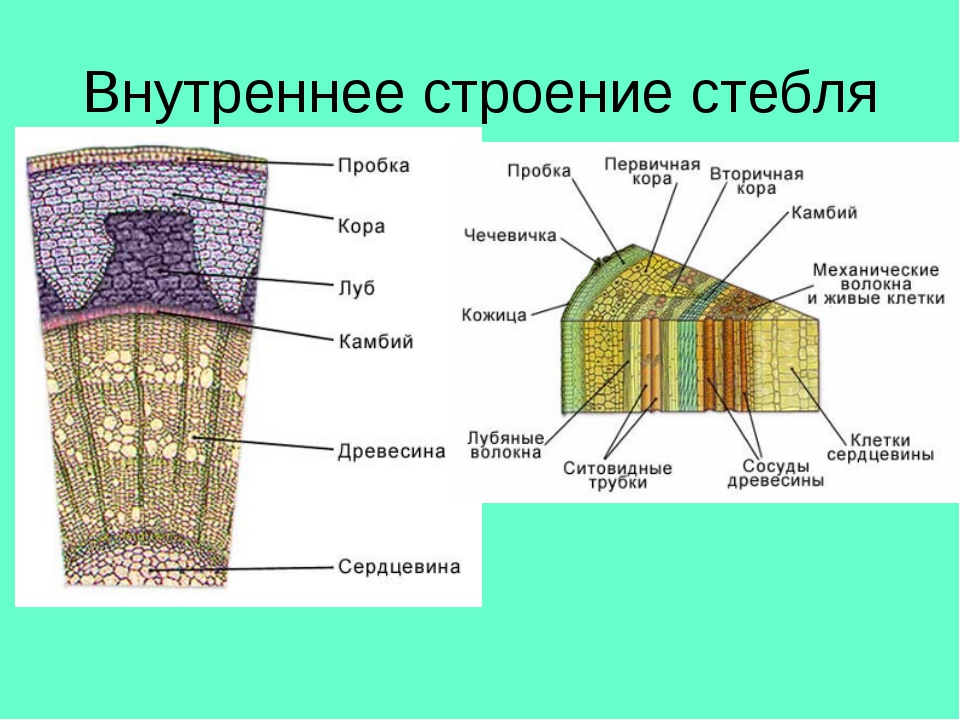 Древесные волокна в качестве армирующих материалов в композиты из натуральных волокон: структура, свойства, обработка и применение
Древесные волокна в качестве армирующих материалов в композиты из натуральных волокон: структура, свойства, обработка и применение
23. Сравнение твердости клеточной стенки при наноиндентировании и твердости древесины по Бринеллю сосны обыкновенной (Pinusbankiana Lamb.
24. Изучение нано- и макромасштабных структурно-механических взаимосвязей археологического дуба Новый метод подготовки образцов, позволяющий избежать влияния заливочной среды во время наноиндентирования
27. Экспериментальная микромеханическая характеристика стенок клеток древесины
28. Характеристика поверхности
29. Eucalyptus nitens: наномеханические свойства волокон коры и древесины
30. Структура клеточной стенки и формирование созревающих волокон бамбука мозо ( Phyllostachys pubescens ) повысить устойчивость к продольному изгибу
31. Влияние снижения содержания углеводов на свойства древесных прядей
32. Связь между модулем вдавливания, углом микрофибрилл и упругими свойствами клеточных стенок древесины
Связь между модулем вдавливания, углом микрофибрилл и упругими свойствами клеточных стенок древесины
33. Уменьшение жесткости и прочности древесины и изменение формы ствола у молодых антисмысловых 4CL трансгенные тополя с пониженным содержанием лигнина
34. Влияние Physisporinus vitreus на свойства древесины ели европейской. Часть 1: Аспекты делигнификации и поверхностной твердости
35. Механическая характеристика одиночных бамбуковых волокон методом наноиндентирования и микрорастяжения
36. Ультраструктура и механические свойства древесины тополя с пониженным содержанием лигнина, Циннамат-4-гидроксилаза
37. Наноразмерная характеристика натуральных волокон и их композитов с помощью контактно-резонансной силовой микроскопии
38. Инструменты для характеристики биомассы в нанометровом масштабе
39. Экспериментальный метод для учета структурной податливости в измерениях наноиндентирования
40.
41. Механические свойства пиролизованной древесины: исследование наноиндентирования
42. Влияние пропитки и полимеризации метакрилатов на месте на твердость древесины сахарного клена
43. Пилотное исследование механических свойств пленок красок для деревянных полов методом наноиндентирования in situ
44. Модуль жесткости в Норвегии ели в зависимости от годичных колец
45. Механические свойства клеточных стенок древесины ели методом наноиндентирования
46. Значение модуля упругости клеточных стенок древесины, полученное из измерений наноиндентирования
47. Прочность на осевое сжатие ели европейской в зависимости от структурной изменчивости и содержания лигнина
Лигнификация в трансгенных растениях с дефицитом 4-кумарат-3-гидроксилазы (C3H) или ассоциированной гидроксициннамоилтрансферазы (HCT)
Лигнификация в трансгенных растениях с дефицитом 4-кумарат-3-гидроксилазы (C3H), или ассоциированная гидроксициннамоилтрансфераза (HCT) | Поиск по дереву Перейти к основному содержанию . gov означает, что это официально.
gov означает, что это официально.
Веб-сайты федерального правительства часто заканчиваются на .gov или .mil. Прежде чем делиться конфиденциальной информацией, убедитесь, что вы находитесь на сайте федерального правительства.
Сайт защищен.
https:// гарантирует, что вы подключаетесь к официальному веб-сайту и что любая предоставленная вами информация шифруется и передается безопасно.
Автор(ы):
Джон Ральф
Такуя Акияма
Хун Ким
Фачуан Лу
Клинт Чаппл
Рамеш Б. Наир
Наир
Армин Вагнер
Фан Чен
М.С. Шриниваса Редди
Ричард Диксон
Хизер Д. Коулман
Шон Д. Мэнсфилд
Тип публикации:
Разное Публикация
Первичная(ые) станция(и):
Лаборатория лесных товаров
Источник:
Polyphenols Communications 2006: XXIII Международная конференция по полифенолам, Виннипег, Манитоба, Канада, 22-25 августа 2006 г. [Виннипег]: Университет Манитобы, [2006]: ISBN: 0978158903: 9780978158903: [2] страницы.
[Виннипег]: Университет Манитобы, [2006]: ISBN: 0978158903: 9780978158903: [2] страницы.
Описание
Понижающая регуляция гена, кодирующего 4-кумарат-3-гидроксилазу (C3H), у покрытосеменных значительно, но предсказуемо увеличила долю p-гидроксифенильных (P) звеньев по сравнению с обычно преобладающими сирингильными (S) и гваяциловыми (G) звеньями. Уровни стебля люцерны до ~ 65% P (от уровней дикого типа (WT) ~ 1%), возникающие в результате подавления C3H, были измерены с помощью традиционных анализов деградации, а также коррелятивных методов 2D 13C-1H ЯМР.Такие уровни помещают эти трансгенные растения далеко за пределы состава P:G:S нормальных растений. ЯМР также выявил структурные различия в распределении межзвенных связей, которое характеризует полимер лигнина. Менее сильно повышенные уровни P были также обнаружены в культурах тополя с подавлением C3H и в культурах трахеид P.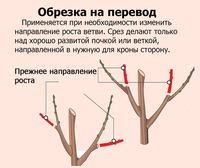 radiata с подавлением HCT, но с различными структурными эффектами. Композиционные и структурные изменения остаются в соответствии с существующей теорией лигнификации, основанной на комбинаторных реакциях радикального сочетания под простым химическим контролем.Такие структурные различия составляют основу для объяснения различий в усвояемости и производительности варки C3H/HCT-дефицитных растений.
radiata с подавлением HCT, но с различными структурными эффектами. Композиционные и структурные изменения остаются в соответствии с существующей теорией лигнификации, основанной на комбинаторных реакциях радикального сочетания под простым химическим контролем.Такие структурные различия составляют основу для объяснения различий в усвояемости и производительности варки C3H/HCT-дефицитных растений.
Цитата
Ральф, Джон; Акияма, Такуя; Ким, Хун; Лу, Фачуан; Ральф, Салли А.; Чаппл, Клинт; Наир, Рамеш Б.; Вагнер, Армин; Чен, Фанг; Редди, М.С. Шриниваса; Диксон, Ричард А; Коулман, Хизер Д.; Мэнсфилд, Шон Д. 2006. Лигнификация трансгенных растений с дефицитом 4-кумарат-3-гидроксилазы (C3H) или ассоциированной гидроксициннамоилтрансферазы (HCT).Коммуникации полифенолов 2006: XXIII Международная конференция по полифенолам, Виннипег, Манитоба, Канада, 22–25 августа 2006 г. [Виннипег]: Университет Манитобы, [2006]: ISBN: 0978158903: 9780978158903: [2] страницы.
Примечания к публикации
- Мы рекомендуем вам также распечатать эту страницу и прикрепить ее к распечатке статьи, чтобы сохранить полную информацию о цитировании.
- Эта статья была написана и подготовлена служащими правительства США в официальное время и поэтому находится в открытом доступе.
https://www.fs.usda.gov/treesearch/pubs/27092
Plantae | Лигнифицирующие и окислительные ферменты: локализация, локализация, локализация!
Имя Марк Сомссич
Филиал Мельбурнского университета
ОРЦИД 0000-0001-5092-6168
Лигнификация клеточных стенок — ключевой процесс в жизни растений. Лигнин представляет собой биополимер, который придает дополнительную стабильность вторичным клеточным стенкам (SCW), позволяет сосудистым клеткам транспортировать воду и защищает растение от патогенной атаки (Tobimatsu and Schuetz, 2019). Он производится из монолигнолов, которые синтезируются в цитозоле и впоследствии секретируются в апопласт, где они сшиваются за счет радикально-радикальной связи. Для этой стадии полимеризации необходимы окислительные ферменты, лакказы (LAC) и пероксидазы (PRX), а также активные формы кислорода (ROS), такие как H 2 O 2 .Полимеризованный лигнин включается в SCW, где он сшивает различные полисахариды клеточной стенки. Однако только некоторые типы клеток имеют лигнифицированные СКВ, и даже в этих специализированных клетках могут быть локальные различия в лигнификации между стенками. Таким образом, лигнификация представляется жестко направленным и точно локализованным событием. Из недавней работы выяснилось, что это точное нацеливание частично достигается за счет специфической локализации окислительных ферментов. В связи с этим лаборатория А.Lacey Samuels ранее описала локализацию LAC4 и PRX64 либо во всей SCW (LAC4), либо только в средней пластинке и углах клеток (PRX64) (Schuetz et al.
Он производится из монолигнолов, которые синтезируются в цитозоле и впоследствии секретируются в апопласт, где они сшиваются за счет радикально-радикальной связи. Для этой стадии полимеризации необходимы окислительные ферменты, лакказы (LAC) и пероксидазы (PRX), а также активные формы кислорода (ROS), такие как H 2 O 2 .Полимеризованный лигнин включается в SCW, где он сшивает различные полисахариды клеточной стенки. Однако только некоторые типы клеток имеют лигнифицированные СКВ, и даже в этих специализированных клетках могут быть локальные различия в лигнификации между стенками. Таким образом, лигнификация представляется жестко направленным и точно локализованным событием. Из недавней работы выяснилось, что это точное нацеливание частично достигается за счет специфической локализации окислительных ферментов. В связи с этим лаборатория А.Lacey Samuels ранее описала локализацию LAC4 и PRX64 либо во всей SCW (LAC4), либо только в средней пластинке и углах клеток (PRX64) (Schuetz et al. , 2014; Yi Chou et al., 2018). Опираясь на эту работу, они теперь намеревались отнести еще несколько ферментов LAC и PRX, участвующих в лигнификации, к их конкретному субдомену SCW, приближая нас к визуализации полимеризации лигнина на субклеточном уровне (Hoffmann et al., 2020). ).
, 2014; Yi Chou et al., 2018). Опираясь на эту работу, они теперь намеревались отнести еще несколько ферментов LAC и PRX, участвующих в лигнификации, к их конкретному субдомену SCW, приближая нас к визуализации полимеризации лигнина на субклеточном уровне (Hoffmann et al., 2020). ).
В своей новой рукописи в этом выпуске журнала Plant Physiology Hoffmann et al.использовать данные об экспрессии для различных генов LAC и PRX , данные о совместной экспрессии этих генов с другими компонентами синтеза SCW и известные фенотипы мутантов, чтобы сузить длинный список членов семейства Arabidopsis thaliana LAC и PRX. до восьми основных кандидатов, которые могут быть центральными ферментами, облегчающими лигнификацию клеток ксилемы и межпучковых волокон в стеблях арабидопсиса. Этот тщательный анализ составляет основу работы, представленной здесь, и авторы включили полезную таблицу, в которой обобщаются идентификаторы генов LAC и PRX, паттерны экспрессии и данные о совместной экспрессии, а также фенотипы стволовых мутантов. Большая часть работы по пониманию роли LAC в полимеризации лигнина была сосредоточена на LAC4 и LAC17, причем LAC4 является наиболее высокоэкспрессируемым LAC в стеблях, а двойные мутанты lac4 lac17 демонстрируют снижение лигнификации стеблей до 40% (Berthet и др., 2011; Холл и Эллис, 2013). Точно так же работа над PRX часто была сосредоточена на PRX64, наиболее сильно выраженном PRX в стеблях (Hall and Ellis, 2013; Yi Chou et al., 2018).
Большая часть работы по пониманию роли LAC в полимеризации лигнина была сосредоточена на LAC4 и LAC17, причем LAC4 является наиболее высокоэкспрессируемым LAC в стеблях, а двойные мутанты lac4 lac17 демонстрируют снижение лигнификации стеблей до 40% (Berthet и др., 2011; Холл и Эллис, 2013). Точно так же работа над PRX часто была сосредоточена на PRX64, наиболее сильно выраженном PRX в стеблях (Hall and Ellis, 2013; Yi Chou et al., 2018).
 LAC10, PRX42 и PRX52 локализуются в не лигнифицированных тканях и, следовательно, могут быть активны в других путях. Интересно, что LAC4 гомогенно распределяется по SCW сосудистых элементов протоксилемы на самых ранних стадиях развития, а затем дополнительно локализуется специфически в углах клеток и средней ламелле волоконных клеток, прежде чем он также показывает гомогенное распределение в SCW этих клеток для остальные стадии развития (рис. 1). Следовательно, LAC4 может играть роль в лигнификации SCW как в элементах сосудов, так и в клетках волокон на разных стадиях развития.
LAC10, PRX42 и PRX52 локализуются в не лигнифицированных тканях и, следовательно, могут быть активны в других путях. Интересно, что LAC4 гомогенно распределяется по SCW сосудистых элементов протоксилемы на самых ранних стадиях развития, а затем дополнительно локализуется специфически в углах клеток и средней ламелле волоконных клеток, прежде чем он также показывает гомогенное распределение в SCW этих клеток для остальные стадии развития (рис. 1). Следовательно, LAC4 может играть роль в лигнификации SCW как в элементах сосудов, так и в клетках волокон на разных стадиях развития. Продемонстрировав эти специфические локализации различных ферментов, авторы переходят к тестированию фактической окислительной активности в этих областях. Инкубация срезов ствола с субстратом окисления PRX TMB (3,3′,5,5′-тетраметилбензидин) и экзогенным H 2 O 2 приводит к окислению TMB во всех клеточных стенках, независимо от лигнификации, что указывает на то, что PRX являются обычно присутствует на всех стенах. Однако когда срезы стебля инкубировали только с TMB, окисление ограничивалось лигнифицированным SCW, перекрывая локализацию различных ферментов LAC и PRX.Это наблюдение демонстрирует, что не только локализация ферментов в этих специфических областях, но и направленное высвобождение эндогенных H 2 O 2 в апопласт в этих доменах способствует специфической картине лигнификации.
Хоффманн и др. идентифицировали ферменты LAC и PRX (а также их комбинации), участвующие в лигнификации SCW на разных стадиях развития и в определенных субдоменах SCW, что поможет уточнить нашу модель лигнификации.Кроме того, они демонстрируют, что не только пространственное распределение различных наборов ферментов и целенаправленная секреция монолигнолов в этих областях, но и локализованное накопление АФК необходимы для достижения сильно локализованной лигнификации. Это согласуется с недавними выводами о том, что лигнификация полоски Каспари также зависит от целенаправленного накопления АФК и локализации окислительных ферментов (Fujita et al. , 2020).
, 2020).
Представленные здесь данные также демонстрируют, что многие члены семейства LAC и PRX не просто дублируют друг друга, но, по-видимому, выполняют специфические функции в специфических доменах и в специфические моменты времени.Будет интересно узнать больше о том, как ферменты направляются к своим специфическим доменам и как они там секвестрируются. В этом отношении особенно интересным кандидатом будет LAC4, который изменяет свою интрамуральную локализацию с углов волокнистых клеток на гомогенное распределение. В целом результаты этой публикации представляют собой еще один шаг к пониманию полимеризации лигнина на субклеточном уровне.
ЦИТИРОВАННАЯ ЛИТЕРАТУРА
Berthet S, Demont-Caulet N, Pollet B, Bidzinski P, Cézard L, Le Bris P, Borrega N, Hervé J, Blondet E, Balzergue S, et al (2011) Нарушение LACCASE4 и 17 результатов в тканях- Специфические изменения лигнификации стеблей Arabidopsis thaliana.Растительная клетка 23 : 1124–1137
Fujita S, Bellis D De, Edel KH, Köster P, Andersen TG, Schmid-siegert E, Tendon VD, Pfister A, Marhavy P, Ursache R, et al (2020) ШЕНГЕНСКИЙ рецепторный модуль стимулирует локализованное производство и лигнификацию АФК в корнях растений. EMBO J 1–18
EMBO J 1–18
Hall H, Ellis B (2013) Программирование транскрипции во время созревания клеточной стенки в расширяющемся стебле арабидопсиса. BMC Plant Biol 13 : 14
Hoffmann N, Benske A, Betz H, Schuetz M, Samuels AL (2020) Лакказы и пероксидазы совместно локализуются в одревесневших вторичных клеточных стенках на протяжении всего развития стебля арабидопсиса.Завод Физиол 183 : https://doi.org/10.1104/pp.20.00473
Schuetz M, Benske A, Smith RA, Watanabe Y, Tobimatsu Y, Ralph J, Demura T, Ellis B, Samuels AL (2014) Лакказы направляют лигнификацию в дискретных доменах вторичной клеточной стенки протоксилемы. Завод Физиол 166 : 798–807
Tobimatsu Y, Schuetz M (2019) Полимеризация лигнина: как растения так хорошо справляются с химией? Curr Opin Biotechnol 56 : 75–81
Yi Chou E, Schuetz M, Hoffmann N, Watanabe Y, Sibout R, Samuels AL (2018)Распределение, подвижность и закрепление связанных с лигнином окислительных ферментов во вторичных клеточных стенках арабидопсиса. J Exp Bot 69 : 1849–1859
J Exp Bot 69 : 1849–1859
Одревеснение в развивающихся стеблях бамбука Sinobambusa tootsik | Journal of Wood Science
Chemicals
Кониферин и сирингин были предоставлены почетным профессором Н. Терашима, Университет Нагоя, Япония. Другие химикаты, за исключением p -кумарилового спирта и p -глюкокумарилового спирта, были приобретены у Wako Pure Chemical Industries (Осака, Япония), Tokyo Chemical Industry Co. (Токио, Япония) или Sigma-Aldrich (Сент-Луис). , Миссури, США).
Синтезы
p — глюкокумарилового спирта и p — кумарилового спирта p -Глюкокумариловый спирт синтезировали, как описано ранее [23]. Синтез p -кумарилового спирта в основном повторял синтез p -глюкокумарилового спирта с некоторыми модификациями. Вкратце, смесь p -гидроксибензальдегида (6,0 г), моноэтилмалоната (8,4 г), пиридина (6 мл) и пиперидина (10 капель) нагревали при 60 °C в течение 24 часов. После подкисления (pH 3) реакционную смесь экстрагировали этилацетатом с последующей экстракцией насыщенным раствором гидрокарбоната натрия и 10%-ным раствором гидросульфита натрия. Органический слой промывали водой и затем концентрировали. Перекристаллизация из смеси с этилацетатом и гексаном давала п этиловый эфир кумаровой кислоты (5,6 г, выход 59%).
После подкисления (pH 3) реакционную смесь экстрагировали этилацетатом с последующей экстракцией насыщенным раствором гидрокарбоната натрия и 10%-ным раствором гидросульфита натрия. Органический слой промывали водой и затем концентрировали. Перекристаллизация из смеси с этилацетатом и гексаном давала п этиловый эфир кумаровой кислоты (5,6 г, выход 59%).
Алюмогидрид лития (0,8 г) помещали в колбу и добавляли абсолютный тетрагидрофуран (50 мл). p — Раствор этилового эфира кумаровой кислоты (1,0 г в 15 мл абсолютного тетрагидрофурана) добавляли по каплям в течение 1 часа при перемешивании, и реакцию поддерживали в течение 2 часов. После подкисления (pH 3) реакционную смесь трижды экстрагировали этилацетатом. Органический слой промывали соляным раствором и затем удаляли с получением p -кумарилового спирта (0,12 г, выход 15%).
Экстракция предшественников лигнина
Растения S. tootsik , используемые в этих экспериментах, выращивались в кампусе Университета Миядзаки. Пробы были собраны в период с мая по сентябрь (см. Таблицу 1) и в ноябре (рис. 1c). При каждом отборе проб собирали три стебля бамбука одинаковой общей высоты. Стебли возрастом более 1 года были отобраны как «зрелые» образцы, хотя их точный возраст неизвестен. Самое низкое междоузлие над землей или укоренившееся междоузлие обозначали как 0, а междоузлия нумеровали снизу вверх стебля. Блоки, содержащие как внешние, так и самые внутренние клетки междоузлия, вырезали из верхней части 2-го, 4-го, 6-го, 8-го и 10-го междоузлий.Блоки замораживали в жидком азоте и хранили при температуре -80 °C до использования в следующих экспериментах.
Пробы были собраны в период с мая по сентябрь (см. Таблицу 1) и в ноябре (рис. 1c). При каждом отборе проб собирали три стебля бамбука одинаковой общей высоты. Стебли возрастом более 1 года были отобраны как «зрелые» образцы, хотя их точный возраст неизвестен. Самое низкое междоузлие над землей или укоренившееся междоузлие обозначали как 0, а междоузлия нумеровали снизу вверх стебля. Блоки, содержащие как внешние, так и самые внутренние клетки междоузлия, вырезали из верхней части 2-го, 4-го, 6-го, 8-го и 10-го междоузлий.Блоки замораживали в жидком азоте и хранили при температуре -80 °C до использования в следующих экспериментах.
Рост в длину Sinobambusa tootsik стеблей. a Общая длина образцов стебля (см. Таблицу 1). Данные представляют собой среднее значение ± стандартное отклонение для трех биологических повторов. b Длина 2-го, 4-го, 6-го, 8-го и 10-го междоузлий стеблей (см. Таблицу 1). Данные представляют собой средние значения трех биологических повторов. c Длина каждого междоузлия зрелых (старше 1 года) стеблей, собранных в ноябре
b Длина 2-го, 4-го, 6-го, 8-го и 10-го междоузлий стеблей (см. Таблицу 1). Данные представляют собой средние значения трех биологических повторов. c Длина каждого междоузлия зрелых (старше 1 года) стеблей, собранных в ноябре
поперечных криосрезов были получены из каждого блока замороженных образцов и помещены в 1,5-мл флакон при температуре -20 °C. Флаконы взвешивали до и после добавления срезов для определения веса свежих срезов. После взвешивания флакона добавляли 1 мл 80% метанола и срезы экстрагировали в течение ночи при 4 °C. Экстракт собирали и определяли предшественники лигнина жидкостной хроматографией-масс-спектрометрией (ЖХ-МС).Срезы обрабатывали ацетоном в течение ночи после экстракции метанолом. После удаления супернатанта срезы высушивали и использовали для анализа на лигнин.
Количественное определение лигнина
Для количественного определения лигнина использовали метод тиогликолевой кислоты (ТГА) [24]. Вкратце, образцы реагировали с 1 мл 3 н. HCl и 0,1 мл ТГА в 2-мл пробирках с завинчивающимися крышками при 80 °C в течение 3 часов. Пробирки центрифугировали при 20000× g в течение 10 мин, осадок промывали 1 мл дистиллированной воды, затем добавляли 1 мл 1 н. NaOH и пробирку встряхивали вертикально при 80 об/мин в течение ночи.После центрифугирования, как описано выше, 1 мл супернатанта смешивали с 0,2 мл концентрированной HCl. Лигнин TGA осаждали при 4 °C в течение 4 ч и собирали центрифугированием, как описано выше. Осадок растворяли в 1 н. растворе NaOH и измеряли поглощение при 280 нм для определения количества производных ТГА по стандартной кривой, полученной с использованием известных концентраций лигнина измельченной древесины бамбука [24].
HCl и 0,1 мл ТГА в 2-мл пробирках с завинчивающимися крышками при 80 °C в течение 3 часов. Пробирки центрифугировали при 20000× g в течение 10 мин, осадок промывали 1 мл дистиллированной воды, затем добавляли 1 мл 1 н. NaOH и пробирку встряхивали вертикально при 80 об/мин в течение ночи.После центрифугирования, как описано выше, 1 мл супернатанта смешивали с 0,2 мл концентрированной HCl. Лигнин TGA осаждали при 4 °C в течение 4 ч и собирали центрифугированием, как описано выше. Осадок растворяли в 1 н. растворе NaOH и измеряли поглощение при 280 нм для определения количества производных ТГА по стандартной кривой, полученной с использованием известных концентраций лигнина измельченной древесины бамбука [24].
Характеристика лигнина
Безэкстрактивные образцы использовали для характеристики лигнина методом микромасштабного тиоацидолиза [25].В качестве внутреннего стандарта использовали тетракозан. Газохроматографический-масс-спектрометрический (ГХ-МС) анализ проводили на приборе GCMS-QP 2010 SE (Shimadzu, Киото, Япония), оснащенном колонкой Inert Cap (60 м × 0,25 мм внутреннего диаметра; GL Sciences, Токио, Япония). Объем вводимой пробы составлял 1 мкл в импульсном режиме без разделения. Инжектор и передаточная линия работали при 250 и 280 °C соответственно. Начальная температура печи составляла 200°C (выдерживалась 1 мин) и повышалась при 4°C min -1 до 280°C и выдерживалась 10 мин.Количественную оценку мономеров p -гидроксифенила (H), гваяцила (G) и сирингила (S) проводили с использованием ионных хроматограмм, реконструированных по их характерным бензильным ионам при мкм / z 239, 269 и 299 соответственно. .
Объем вводимой пробы составлял 1 мкл в импульсном режиме без разделения. Инжектор и передаточная линия работали при 250 и 280 °C соответственно. Начальная температура печи составляла 200°C (выдерживалась 1 мин) и повышалась при 4°C min -1 до 280°C и выдерживалась 10 мин.Количественную оценку мономеров p -гидроксифенила (H), гваяцила (G) и сирингила (S) проводили с использованием ионных хроматограмм, реконструированных по их характерным бензильным ионам при мкм / z 239, 269 и 299 соответственно. .
Анализ ЖХ-МС
Метанольный экстракт, хранившийся при -80 °C, центрифугировали при 20 000× g в течение 20 мин, а затем надосадочную жидкость фильтровали через мембранный фильтр (0,45 мкм) перед анализом ЖХ-МС. Анализ ЖХ-МС проводили с использованием системы УВЭЖХ Dionex Ultimate 3000 (Thermo Scientific, Уолтем, Массачусетс, США), соединенной с масс-спектрометром Q-Exactive (Thermo Fisher, Рокфорд, Иллинойс, США).Аликвоту 2 мкл каждого образца вводили в колонку InertSustain C18 с обращенной фазой (100 мм × 2,1 мм, внутренний диаметр, 2 мкм; GL Sciences, Токио, Япония) при температуре 35 °C. Скорость потока составляла 0,3 мл мин -1 , а программа градиента была следующей: метанол 15% (об./об.) выдерживали в течение 5 минут, увеличивали до 35% метанола за 15 минут, повышали до 80% метанола за 20 минут. , и выдерживали до 28 мин с постоянной 0,1% уксусной кислотой. Масс-спектральные данные собирали при положительной или отрицательной полярности ионов с использованием квадрупольного масс-спектрометра, снабженного нагретым капиллярным интерфейсом электрораспыления.Предшественники лигнина идентифицировали и определяли сравнением со стандартными кривыми, рассчитанными по данным МС для аутентичных соединений. Время удерживания и масса каждого аутентичного соединения (допуск массы = 3 ppm), использованного для идентификации, были следующими: p -кумариловый спирт (10,1 мин, +, m / z 133,065), p -глюкокумарил спирт (4,1 мин, +, м / z 335.110), кониферальдегид (15,5 мин, -, м / z 177.056), конифериловый спирт (12,3 мин, +, м / z 163,075), кониферин (6,2 мин, +, м / z 365,121), синапальдегид (16,4 мин, −, м 365,121), синапальдегид z 207.
Скорость потока составляла 0,3 мл мин -1 , а программа градиента была следующей: метанол 15% (об./об.) выдерживали в течение 5 минут, увеличивали до 35% метанола за 15 минут, повышали до 80% метанола за 20 минут. , и выдерживали до 28 мин с постоянной 0,1% уксусной кислотой. Масс-спектральные данные собирали при положительной или отрицательной полярности ионов с использованием квадрупольного масс-спектрометра, снабженного нагретым капиллярным интерфейсом электрораспыления.Предшественники лигнина идентифицировали и определяли сравнением со стандартными кривыми, рассчитанными по данным МС для аутентичных соединений. Время удерживания и масса каждого аутентичного соединения (допуск массы = 3 ppm), использованного для идентификации, были следующими: p -кумариловый спирт (10,1 мин, +, m / z 133,065), p -глюкокумарил спирт (4,1 мин, +, м / z 335.110), кониферальдегид (15,5 мин, -, м / z 177.056), конифериловый спирт (12,3 мин, +, м / z 163,075), кониферин (6,2 мин, +, м / z 365,121), синапальдегид (16,4 мин, −, м 365,121), синапальдегид z 207.