
Назовите организмы, которые имеют островной тип распределения популяций. а) среди водных…
Биология, опубликовано 05.12.2018
Вариант 1
А. Выберите один правильный ответ
1. Главный признак живого организма:
а) изменение формы,
б) изменение размера,
в) обмен веществ,
г) пассивное передвижение
А 2. Все живые организмы в отличие от неживых:
а) размножаются, растут, развиваются,
б) существуют независимо от среды,
в) изменяют форму,
г) изменяются под воздействием среды.
А 3. В предложенном перечне найди название систематической категории:
а) вид,
б) собака,
в) роза,
г) человек.
А 4. Группа особей, скрещивающихся между собой и дающих плодовитое потомство – это:
а) вид,
б) род,
в) тип,
г) отряд
А 5. Главная задача систематики – это изучение:
а) этапов эволюции,
б) отношений организмов и окружающей среды,
в) приспособленности организмов к условиям обитания,
г) рганизмов и объединение их в группы на основе родства.
Вариант 2
1) питание
2) рост
3) обмен веществ
4) наследственность.
Б 2. Преемственность поколений обеспечивается за счет
1) развития
2) роста
3) размножения
4) обмена веществ.
Б 3. Удвоение ДНК происходит на уровне организации жизни
1) клеточном
2) молекулярном
3)органо-тканевом
4) организменном
Б 4. Круговорот воды в природе наблюдается на уровне организации жизни
1) популяционно-видовом
2) биосферном
3) экосистемном
4) организменном.
Б 5. Цветение черемухи обыкновенной наблюдается на уровне организации жизни
1) клеточном
2) молекулярном
3)органо-тканевом
4) организменном
Б 6. Деление ядра – это пример проявления жизни на уровне
1) клеточном
2) молекулярном
3)органо-тканевом
4) организменном..
Б 7. Динамика численности уссурийского тигра – это пример на уровне
1) популяционно-видовом
2) биосферном
4) организменном.
Б 8. На каком уровне организации живого происходят генные мутации?
1) организменном
2) клеточном
3) видовом
4) молекулярном
Б 9. Способность организма отвечать на воздействия окружающей среды называют:
1) воспроизведением
2) эволюцией
3) раздражимостью
4) нормой реакции
Б 10. Живое от неживого отличается способностью
1) изменять свойства объекта под воздействием среды
2) участвовать в круговороте веществ
3) воспроизводить себе подобных
4) изменять размеры объекта под воздействием среды
Б 11. Клеточное строение — важный признак живого — характерен для
1) бактериофагов
2) вирусов
3) кристаллов
4) бактерий.
Б 12. Для изучения строения клетки необходимо:
1) рассмотреть под микроскопом орган растения, например, лист;
2) приготовить тонкий срез какого либо растения и рассмотреть его под микроскопом;
3) выделить из органа одну клетку и рассмотреть ее под микроскопом;
uchiru.net
Формирование островных популяций животных
Формирование островных популяций животных

По данным И.М. Кержнера (1979), представители материковых фаун Nabidae заселяют в основном большие острова, такие, как Новая Зеландия и Новая Каледония, или обширные архипелаги, соседствующие с материками.
Полинезийскую часть Океании заселяют в основном виды двух таксонов — родовых групп Nabis sensu lato и Tropiconabis, широко радиирующих и дающих множество эндемиков на таких островных системах, как Гавайские, Маркизские, Галапагос. Эти клопы особенно склонны к формированию островных популяций. Виды Tropiconabis широко радиируют на разнообразных архипелагах, обитают на многих сверхизолированных островах восточной части Океании как в тропических, так и в субтропических поясах. Именно представитель этого рода (T. nigrolineatiis Dist.) нами обнаружен на атолле Дейнджер и вулкане Рауль.
Наиболее обычные полужесткокрылые на самых мелких и удаленных островах — виды родов Remauclieriana и Pachybrachius сем. Lygaeidae. Мы встретили их на всех обследованных островах, в том числе как единственных представителей отряда на атолле Суворова, где обычен трансокеанский Remauclieriana nigriceps Dali.
Много примеров повышенной склонности к формированию островных популяций среди чешуекрылых. Так, по всей экваториальной и тропической Океании, включая острова Кука и Общества, распространены два сборных вида, которые, вероятно, следует считать надвидами. Это Hypolimnas bolina L. и Precis villida F., являющийся представителем рода, замещающего в Ориентальной области нашу Vanessa. Эти бабочки наиболее обычны в восточной части Океании, где фауна дневных чешуекрылых сильно обеднена. Мы их встречали в основном на самых удаленных островах, на атоллах Суворова и Дейнджер, на вулкане Ниуафооу, на которых из дневных чешуекрылых встречаются еще единичные виды голубянок. Эти таксоны образуют множество географических рас, вариететов, а на некоторых островах — симпатрические формы (Gross, 1975). Так, на атолле Дейнджер обычны две формы Н. bolina: типичная темная и светлая, точнее, рыжая (Н. b. alcmene Cv.).
Еще интересные статьи по теме:
www.zoofirma.ru
Ареал островной
Ареал и численность особей в популяции определяется географическими, физическими и экологическими условиями их обитания. Как и в случае видов, распространение популяций является неравномерным. Поэтому различают «сгущенную» и «островную» формы распределения популяций. В случае «сгущенной» формы распространения популяций их ареал характеризуется наличием особенно плотно заселенных зон. Например, «сгущенное» распределение характерно для популяций берез на русской равнине, где они формируют так называемые высокоплотные на единицу площади чистые березняки наряду с уменьшенным количеством растений в других местах ареала. Напротив, «островное» распределение заключается в очаговом распределении берез в Западной Сибири.[ …]
Агатис может бьггь назван островным родом, так как его ареал, заходя лишь на края двух континентов (на полуостров Малакка в Юго-Восточной Азии и в штат Квинсленд в Австралии), охватывает преимущественно острова (Филиппины, Суматра, Калимантан, Сулавеси, Новая Гвинея, Соломоновы, Санта-Крус, Новые Гебриды, Новая Каледония, Фиджи и остров Северный в Новой Зеландии).[ …]
Обширный ареал в умеренных и северных широтах Евразии от 3. Европы до Камчатки и Сахалина, на юг — до горных лесов Ц. Азии, на север — до севера тайги. На большей части территории нашего региона одни из самых многочисленных лесных птиц, распространены от островных и пойменных лесов в степной зоне почти до лесотундры.[ …]
Таких примеров расширения ареала насекомых человеком и островного распределения популяций энтомологическая наука знает йчень много.[ …]
Зимой живут оседло даже в самых северных островных лесах в тундровой зоне. Молодые осенью и в начале зимы широко перемещаются. Возможно, кочует и часть старых птиц, но за пределы гнездового ареала выходят редко.[ …]
Особи любого вида живых организмов распределены в пределах ареала неравномерно. Наблюдения показывают, что существует «островная» форма распределения групп особей и форма «сгущений». Центры плотности» населения каждого вида и являются, как правило, популяциями. Популяция — первая надорганизменная структурная единица вида2.[ …]
Африки, С. Америки. В нашем регионе — всюду до самых северных островных лесов в тундровой зоне. В целом довольно редкая птица, кое-где обычен. Залетают в южную тундру. В пределах гнездового ареала встречаются круглый год.[ …]
Вся лесная зона Евразии и С. Америки. По пойменным и островным лесам гнездовой ареал заходит в степи и на юг тундровой зоны. Нигде не бывает многочисленным, но местами обычен. Вне гнездового ареала — в арктических морях, в тундре и степи — встречаются летующие линяющие гоголи, в основном самцы.[ …]
С. Африка, на восток — до Средней Сибири. В нашем регионе — от пойменных и островных лесов в степи до северной тайги и лесотундры. Одна из самых обычных или многочисленных птиц на большей части ареала. Перелетные птицы, но на юге региона иногда встречаются и зимой.[ …]
Этот юго-восточноевропейскнй вид на территории СССР имеет крайнюю восточную границу ареала.[ …]
Образ жизни. Прилетают поздно, в степные районы — около середины — конца апреля, на север ареала — в конце мая. Населяют разреженные леса, островные и пойменные леса, опушки и парки. Сплошных лесов избегают, явно предпочитают лиственные породы. Петь самцы начинают спустя несколько дней после прилета, подыскав подходящее дупло. Гнездятся в дятловых дуплах и естественных пустотах древесных стволов и толстых ветвей, охотно поселяются в дуплянках и скворечниках. Могут занимать пустоты в строениях, находили гнезда даже в норах в обрывистых берегах и склонах степных балок.[ …]
Большая часть Азии и Сибири. Северные пределы гнездования — лесотундра и островные леса в южной тундре. Залетают в безлесную тундру. В нашем регионе наиболее обычны и даже многочисленны в северной и частично средней тайге на востоке 3. Си бири. На западе ареал доходит до Урала, возможно — до Печоры. Распространение очень неравномерное и очень изменчивое.[ …]
В основном Евразия от Британских о-вов и Испании до Японии и Ю. Китая. В нашем регионе — от островных лесов в степи и лесостепи примерно до юга северной тайги. Немногочисленна или редка, местами на юге лесной зоны обычна. Залетает далеко на юг в степи и на север до лесотундры. На большей части гнездового ареала встречается круглый год.[ …]
Лесной север Евразии и С. Америки, преимущественно северная и средняя тайга. В нашем регионе — почти вся лесная зона и севернее, до островных лесов в тундровой зоне. В гнездовое время на большей части ареала редки, в северной и средней тайге бывают обычны. Зимуют на всей территории региона, кроме тундровой зоны, куда изредка залетают, обычно в теплое время года.[ …]
Группа закономерностей, характерных для пространственных изменений особей в популяциях, сменяющих друг друга при движении с севера на юг (или наоборот) и живущих в различных условиях островной изоляции, наиболее известна. Кажется, еще Фридрих I Барбаросса (1125— 1190) знал правило измельчания размеров тела теплокровных животных при движении с севера на юг, названное затем правилом К■ Бергмана (1847): у теплокровных животных, подверженных географической изменчивости, размеры тела особей статистически (в среднем) больше у популяций, живущих в более холодных частях ареала вида. Правилу Бергмана подчиняются около 50% видов теплокровных животных, подавляющая часть которых (до 90%) птицы. Механизм действия правила Бергмана очевиден — укрупнение размеров особей термодинамически позитивно меняет отношение объема и площади тела, снижает уровень основного обмена. В то же самое время правило не может действовать в тех случаях, когда наблюдается недостаток кормов или колеблется их доступность: при увеличении размеров тела общая масса потребляемого корма возрастает1. Очевидна связь правила Бергмана с правилом поверхностей (разд. 3.4.2).[ …]
Меньше всего ясности с распространением в 3. Сибири. Из-за трудности обнаружения гнезд для большинства районов факт гнездования не доказан. Кочующие птицы встречаются как к югу, так и к северу от гнездового ареала. Область зимнего обитания располагается в основном на той же территории, что и гнездовой ареал, но в целом несколько южнее, так что на большей части северной тайги зимой корольков нет, но они появляются в южных лесах, вплоть до островных боров и сосновых посадок в степной зоне.[ …]
Растения, будь то на индивидуальном, популяционном или видовом уровне организации, можно рассматривать как острова-хозяева для растительноядных насекомых и паразитов растений, а поэтому к ним вполне приложима теория островной биогеографии. Такой подход был использован в нескольких исследованиях для объяснения видового разнообразия насекомых и грибов [124, 216—218]. Практически во всех случаях при этом была обнаружена зависимость числа видов от площади или от ареала: большие скопления растения-хозяина или растения с очень обширным географическим ареалом обеспечивают существование большего числа видов растительноядных насекомых и паразитов, чем мелкие по площади насаждения или растения с небольшим ареалом. Значения 1 для них бывают выше, чем для настоящих островов (см. [265]). Некоторые примеры приведены на рис. 7.3. Все еще не прекращаются споры о том, правомерно ли рассматривать растение-хозяин как остров, и о некоторых данных, приводимых в качестве доказательства существования для него зависимости число видов — площадь (см. возражения [219, 314] и ответы [220, 221, 315]). Однако эта эмпирически установленная зависимость распространена очень широко и обнаружена в самых различных условиях, поэтому отбрасывать ее на том основании, что несколько из приводимых примеров кажутся сомнительными, неправомерно.[ …]
В нашем регионе редкая и очень редкая птица с неясными границами распространения. Относительно обычен лишь кое-где в горных лесах Урала. Возможно, на больших территориях в 3. Сибири вид совсем отсутствует, а распространение на Урале имеет островной характер, без связи с остальным ареалом в Средней и В. Сибири. На пролете встречается к югу от гнездового ареала, в т.ч. в лесостепи и степи. Залетает до 3. Европы.[ …]
Преимущественно лесная зона Евразии от Атлантики до Тихого океана. В нашем регионе — от лесостепи до северной тайги относительно обычный вид, но локальная плотность не бывает высокой и очень изменчива год от года. Иногда гнездятся в степных островных борах. Менее всего известно о границах ареала на территории Сибири. Нерегулярно-перелетный и кочующий вид, в средней и южной тайге в некоторые годы зимует, в лесостепи и степи — чаще.[ …]
Литогенная основа и типы ландшафта. Подчиненная роль литогенной основы в формировании типов ландшафта вытекает из сущности последних как системы комплексов, формирующихся в условиях определенного соотношения тепла и влаги. Но, будучи климатогенными по своей природе, типы ландшафта в своем географическом распространении обнаруживают замечательную связь с литогенной основой. Орография — один из элементов литогенной основы — определяет прерывистый ареал типов ландшафта — от крупных контуров в виде природных зон до мелких островных пятен, вкрапленных в другие зоны или горные страны.[ …]
За последние 20 лет соотношение грызунов в портах и некоторых центральных областях России (например, Тульская обл.) заметно изменилось, а взаимоотношения крыс двух родственных конкурирующих видов дополнились новыми. Морфологическое и репродуктивное разобщение черных крыс России, особенно выраженное между судовыми и прибалтийскими группировками, почти во всех современных портах поддерживается своеобразным барьером, создаваемым портовыми пасюками. Локализация европейских популяций одного вида усугубляет изолированность популяций другого в Прибалтике и в Центре России (Тульская обл.). Влияя на видовое разнообразие, увеличивая напряженность межвидовых отношений, изолируя и расчленяя некоторые поселения черной крысы, пасюк создает новые условия для дифференциации и, вероятно, эволюции черной крысы, и без того весьма изменчивой даже в пределах одного цитогенетического типа. У черной крысы, будто бы повсеместно вытесняемой пасюком, не заметно сужения пищевой ниши. Взаимоотношения этих видов не соответствуют тем, что схематически изображены на рис. 125 книги Н. В. Тимофеева-Ресовского с соавторами (1969), а больше соответствуют гипотезе компрессии (MacArthur, Wilson, 1967, по: Пианка, 1981), согласно которой по мере проникновения в сообщество все большего числа видов ниши (места обитания) сужаются, а пищевые ниши (спектр кормов) не меняются или даже расширяются. Из приведенных выше расчетов следует, что относительное обилие пасюков в городах Восточной Сибири и Дальнего Востока положительно коррелирует с обилием, численностью и биомассой черных крыс. Несмотря на молодость городов, это может быть связано с давностью обитания здесь крыс. Такие процессы заслуживают пристального внимания. Последствия изоляции городских островных местообитаний черных и серых крыс, разобщенности крыс отдельных свинокомплексов, мясокомбинатов, зерноскладов, птицефабрик и пивных заводов хорошо заметны по черепным и эк-стерьерным признакам этих грызунов (Вигоров, 1992). Морфологическое разнообразие пасюков Среднего Урала и Башкирии, т. е. на относительно недавно ими освоенной (100-400 лет) периферии ареала, выражено столь же явно, как и цитогенетическое разнообразие, и, вероятно, вызвано генетическим своеобразием поселений вследствие разной истории заселения Урала крысами. Комплексами экстерьерных, черепных и поведенческих признаков черные крысы Центра и Северо-Запада европейской части России отличаются от крыс флотов и юга Европы не меньше, чем 38- и 42-хромосомные формы крыс друг от друга, а крысы центральных областей (города Данков, Ельня и др.) различаются не меньше, чем сухопутные от корабельных. Морфологическое и поведенческое разнообразие черных крыс России хорошо заметно в пределах одной хромосомной расы, хотя морфологическое — менее резкое, чем у пасюка, и пока не вызвало расхождения колоний до уровня подвидов.[ …]
ru-ecology.info
Пространственное распределение особей в популяции
⇐ ПредыдущаяСтр 8 из 11Следующая ⇒
Пространственное распределение— это распределение особей на территории, занимаемой популяцией. Оно зависит как от разнообразия внешних условий, так и биологических особенностей организмов, в частности их подвижности и степени агрегированности.
Различают три типа распределения особей внутри популяции (см. рисунок 22).
1) равномерное распределение в природе встречается редко. Чаще оно связано с острой конкуренцией между особями популяции. Такой тип распределения отмечают у хищных рыб и у колюшек с их территориальным инстинктом и сугубо индивидуальным характером.
2) случайное распределение имеет место только в однородной среде, например, в водной. Такое распределение на первых порах имеет и тля на поле.
3) групповое распределение встречается наиболее часто, при этом оно может быть и случайным. Групповое распределение обеспечивает более высокую устойчивость популяции по отношению к неблагоприятным условиям среды.
Знание типа распределения имеет большое значение при оценке плотности популяции.
Рисунок 22. Типы распределения особей в популяции
A — равномерное распределение, Б — случайное распределение, B — групповое распределение
Наиболее широко встречается групповое (или агрегированное) распределение организмов. Оно характерно для многих растений.
Групповое распределение многих растений связано с характером распространения их семян. Если семена тяжелые, трудно распространяемые на большие расстояния, то они остаются вблизи родительской особи. Тогда возникают скопления. Наблюдать такое распределение можно у кедра, дуба, тысячелистника, зверобоя. В противоположность им семена тополя, злаков, одуванчика легко распространяются на значительные расстояния.
Образование групп — важный фактор повышения жизнеспособности организмов. Группа деревьев лучше противостоит ветру, эффективнее снижает потери воды. Стае птиц легче защитить гнезда от разорения, легче отогнать хищника. Прайд львов или стая волков легче добывают пищу, чем это может сделать одна особь.
Образование скоплений возможно в результате влияния других организмов. Некоторые растения образуют скопления в местах, менее доступных для животных, их поедающих. Если в результате выпаса скота какой-то вид растения активно выедается, то он может сохраниться где-нибудь среди кустарников, в овраге и других малодоступных для скота местах. Это же может происходить и в результате влияния сенокоса.
Одна из существенных черт внутрипопуляционных пространственных группировок — относительная недолговечность: их существование ограничено обычно несколькими поколениями; они многократно возникают и распадаются в процессе существования популяции.
Рождаемость
Рождаемость – это способность популяций к увеличению численности. В живых организмах заложена огромная возможность к размножению: в популяции имеется тенденция к образованию теоретически максимально возможного количества новых особей (правило максимальной рождаемости). Это достигается в идеальных условиях среды, когда отсутствуют лимитирующие факторы и размножение ограничено лишь физиологическими особенностями вида (т.н. биотический потенциал).
Биотический потенциал —максимально возможная удельная скорость роста, которая может быть достигнута в стационарном состоянии при стабильном возрастном распределении в популяции.
В экологии различают:
— максимальную (физиологическую) рождаемость — теоретический максимум скорости рождения новых особей при отсутствии внешних факторов, сдерживающих процессы размножения;
— экологическую (реализованную) рождаемость — увеличение численности популяции при фактических условиях среды.
Различия между этими типами рождаемости видны на следующем примере. В одном из опытов самки мучного хрущака отложили 12 000 яиц, что характеризует максимальную рождаемость. Однако из этих яиц вывелись только 773 личинки (6 %). Это и была реализованная рождаемость.
Рождаемость обычно выражается либо как:
абсолютная — это скорость, определяемая путем деления числа вновь появившихся особей на время;
либо как удельная — число появившихся потомков в единицу времени на 1 особь в популяции.
Например, популяция простейших, состоявшая из 1000 особей, через 1 ч составила 3000 особей. Тогда абсолютная рождаемость здесь равна 2000 особей/ч, а удельная — 2 особям.
Смертность
Смертность — количество особей, погибших в популяции за определенный период.
Смертность изменяется в зависимости от условий среды, возраста и состояния популяции. Она выражается чаще в виде относительной величины — доли особей (от исходного количества), погибших за определенное время.
Смертность может быть минимальной и экологической. Минимальная смертность представляет собой гибель особей в идеальных для популяции условиях существования и является величиной постоянной для данного вида. Экологическая смертность – гибель особей в реальных условиях существования.
Различают следующие типы смертности:
1) равномерная смертность во всех возрастах,
2) повышенная смертность особей на ранних стадиях развития,
3) преимущественная гибель взрослых, прежде всего старых особей.
Перечисленным типам смертности соответствуют кривые выживания на рисунке 23.
Равномерная смертность во всех возрастах (тип гидры) характерна для некоторых ящериц, бородавочника, многих птиц, рыб, многолетних травянистых растений. Повышенная гибель взрослых особей (тип дрозофилы) характерная для некоторых насекомых, пресмыкающихся, африканских копытных и человека. Повышенная гибель особей на ранних стадиях развития (тип устрицы) наблюдается у многих морских беспозвоночных, рыб, лягушек, большинства насекомых и многих растений.
Реально встречающиеся кривые выживания представляют чаще всего комбинацию перечисленных трех типов.
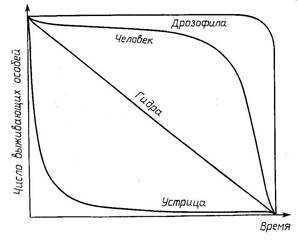
Рисунок 23. Различные типы кривых выживания
Типы роста популяции
Разница между рождаемостью и смертностью характеризует скорость изменения численности популяции.
Если рождаемость равна смертности, а притока особей со стороны нет, то численность популяции не изменится. В этом случае формируется стабильная популяция. Однако фактически такого в природе не наблюдается. Если рождаемость выше смертности, то популяция растет. Наконец, если смертность превышает рождаемость, то численность популяции будет снижаться.
Если удельная скорость изменения численности популяции постоянна, то рост популяции должен идти по экспоненте. Скорость увеличения такой популяции зависит от ее численности. Чем больше численность особей в популяции, тем быстрее она увеличивается.
Экспоненциальный рост популяций иногда наблюдается в природе. Например, это «цветение» воды в результате бурного развития фитопланктона, вспышка массового размножения некоторых вредителей, рост бактерий в свежей культуре. Однако это происходит непродолжительное время.
Рост популяции в реальных экологических условиях описывается кривыми S-образной формы. Сначала рост популяции невелик. Затем он нарастает, но через некоторое время замедляется (см. рисунок 24). Такой тип роста описывается уравнением П.-Ф. Ферхюльста и соответствует на рисунке логистической кривой роста.
Константы r и К из уравнения П.-Ф. Ферхюльста дали название двум типам жизненных стратегий организмов, направленных на повышение вероятностей выжить в тех или иных условиях.
При r-стратегии отбор направлен на повышение скорости роста популяции в начальный период увеличения ее численности, т. е. когда плотность мала и слабо выражено тормозящее действие конкуренции. Этот отбор направлен на высокую плодовитость, быстрое достижение половозрелости, короткий жизненный цикл, способность быстро распространяться в новых местообитаниях, переживать неблагоприятные условия в состоянии покоящихся стадий. r-Виды получают преимущество на ранних стадиях развития экосистем, при заселении новых мест обитания, в молодых, небогатых видами сообществах.
При К-стратегии отбор направлен на повышение выживаемости в условиях стабилизировавшейся численности, при сильном влиянии конкуренции. Отбор направлен на повышение конкурентоспособности, защищенности от хищников, вероятности выживания каждого потомка, на развитие более совершенных внутрипопуляционных механизмов регуляции численности. К-виды — это виды зрелых сообществ.
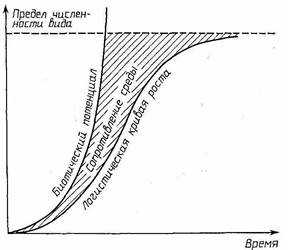
Рисунок 24. Теоретические кривые роста популяции
На основании жизненных стратегий виды растений подразделяются на три группы.
— Виоленты — подавляют конкурентов за счет более интенсивного роста и более полного использования территории. Имеют мощную корневую систему и хорошо развитую надземную часть. К ним относятся деревья, тростники, сфагновые мхи, доминирующие на заболоченных территориях.
— Патиенты — способны выживать в неблагоприятной среде, где многие виды жить не могут: в условиях недостатка влаги, освещенности, бедности почв элементами питания. К ним относятся многие сухо- и тенелюбивые виды.
— Эксплеренты — быстро размножающиеся и быстро расселяющиеся виды. У многих из них семена длительно сохраняют всхожесть. Виды эксплеренты в ходе развития сообщества вытесняются виолентами. Среди эксплерентов широко представлены виды вырубок и гарей (иван-чай, осина).
Рекомендуемые страницы:
lektsia.com
Модель островных популяций — Справочник химика 21
Химия и химическая технология
Статьи Рисунки Таблицы О сайте English Модели с изолированными группами ( островами ) и с однородным непрерывным расселением — это два крайних случая. В действительности могут существовать все градации промежуточных типов распределения и соответствующие им типы дифференциации могут иметь смешанную природу. По-видимому, когда рассматривают различие между сравнительно большими площадями, более применима модель дифференциации островного типа, а когда рассматривают вариацию внутри одной такой площади, более адекватна модель дифференциации непрерывного типа. Как мы отмечали раньше, при математическом рассмотрении структуры популяции приходится прибегать к простым моделям. Именно по этой причине для решения вопроса изоляции расстоянием мы предложили непрерывное и однородное распределения особей на площади или вдоль узкого пояса. Любой биолог-полевик знает, что, как правило, фактическое распределение любых видов бывает каким угодно, только не однородным. Усложняющим фактором является изменение плотности популяций от участка к участку. При этом размер [c.478]Модель островных популяций. Рассмотрим вымышленный пример, представляющий собой крайний случай. Предположим, что в Тихом океане существует 160 изолированных островов, на каждом из которых поселилась одна супружеская пара. Все эти 320 поселенцев имеют группу крови MN. У каждой пары родились сын и дочь, которые, вступив в инцестный брак, становятся предками островной популяции. Ни один из генотипов системы МК не является селективно благоприятным или невыгодным. Через несколько сотен лет мы возвращаемся на острова, чтобы определить группы крови МК в этих популяциях. Что же мы обнаружим [c.367]
Рассмотрим модель подразделенности популяции па песколько уровней иерархии. На нулевом уровне все элементарные единицы подразделения представляют собой субпопуляции со случайным скреш ивапием и равными между собой размерами, которые постоянны во времени. Этн субпопуляции группируются в единицы первого уровня, так сказать, районы , состояш ие из одинакового количества субнонуляций. Внутри единицы первого уровня между субпопуляциями осуш ествляются миграции по описанной в 1 островной схеме, т. е. здесь панмиксия нарушена. Далее группы первого уровня образуют равные по размеру единицы второго уровня ( области ), также соединенные внутри себя миграциями между районами по островной схеме, и т. д. Последней единицей высшего уровня иерархии будет вся подразделенная популяция. [c.471]
Совместное влияние отбора и генного дрейфа приводит в конце концов к гомозиготности популяции. Фактором, позволяющим сохранить ее полиморфность и обеспечивающим существование стационарного распределения вероятности, является миграция, заносящая аллели разных типов. В островной модели предполагается, что интенсивность обмена мигрантами между ионуляцией ( островом ) и внешним источником (с постоянными частотами аллелей q и 1 — ) равна константе т. Для случая двух аллелей снос нз-за миграций (1.3) удобно переписать [c.411]
Смотреть страницы где упоминается термин Модель островных популяций: [c.395] [c.479] [c.468] [c.475]
Генетика человека Т.3 (1990) — [ c.0 ]
© 2019 chem21.info Реклама на сайте
www.chem21.info
Сайт учителей биологии МБОУ Лицей № 2 города Воронежа
Популяции
В жизни любого живого существа большую роль играют отношения с другими представителями собственного вида. Отношения эти осуществляются в популяциях. “Популюс” по-латыни – “народ”, и в точном переводе слово “популяция” означает “население вида на какой-либо территории”.
Любой вид – это большая популяция и, в свою очередь, состоит из более мелких популяций, так как вид неоднородно распределен в пространстве.
Взаимосвязь отдельных популяций
В пределах занимаемой видом общей территории – ареала – встречаются места как более пригодные, так и мало подходящие для жизни. Поэтому возникает некоторая отграниченность одной популяции от другой. Соседние популяции сообщаются друг с другом в процессах расселения, переноса семян и зачатков, сезонных миграций. У некоторых видов такая связь соседних популяций постоянная, у других – эпизодическая. Например, у озерных окуней – островной тип распределения, когда одно пригодное для жизни пространство (озеро) резко отделено от другого, как острова в море. У них связь популяций возникает только изредка, в половодья или наводнения. У семиточечной божьей коровки, свободно перелетающей из одного местообитания в другое, границы между отдельными популяциями сильно размыты.
Наземные улитки из рода цепея всю жизнь проводят на участке в несколько десятков квадратных метров. Для них труднопреодолимы сухие пространства даже небольшой протяженности. Вид распадается на множество мелких популяций, которые развиваются только в достаточно влажных местообитаниях. Волки, лоси, северные олени – подвижные крупные животные, активно передвигающиеся по территории с разнородными условиями. В пределах таких видов вычленяется меньшее число популяций, каждая из которых обычно связана с обширным пространством между какими-либо географическими границами.
У перелетных птиц и других мигрирующих животных популяции выделяются по местам их размножения. На зимовках могут скапливаться представители различных популяций.
Скорость расселения вида можно характеризовать средним расстоянием между местом рождения и местом размножения большинства членов популяции. Для ежа обыкновенного оно составляет в разных условиях от 200 до 1000 м, для крота – 170 м, зайца беляка – 400 м, зайца-русака – 3 км, белки обыкновенной – 7 км, соболя – 19 км. Некоторые перелетные птицы могут размножаться за 1–1,5 тыс. км от места рождения.
У растений распространяются семена, пыльца, споры.
За счет постоянно протекающего расселения молодых, а у некоторых видов – взрослых особей осуществляется связь различных популяций и освоение новых территорий.
Условия жизни видов в разных частях ареала могут сильно различаться. Например, белки в лесах Белоруссии и Дальнего Востока живут в условиях разного климата, в разных типах леса, в окружении разных врагов и т. д. Каждая популяция оказывается приспособленной к своим условиям. Поэтому любой широко распространенный вид экологически неоднороден.
Зайцы-беляки, обитающие в районе Верхоянска, отличаются от своих собратьев из популяций, населяющих районы Средней Волги, целым рядом особенностей. Например, у них разный состав пищи: южные грызут преимущественно лиственные породы, северные – больше хвойные, ягодные кустарнички и травы. Из-за более грубых кормов у них вдвое больше длина слепой кишки. В северных популяциях сокращен период размножения, самки дают только один приплод за лето, а приволжские зайцы – два-три. Возрастной состав популяций разный.
Взаимоотношения особей внутри популяции
Члены одной и той же популяции – это соседи, вынужденные сообща осваивать занимаемую территорию и размножаться на ней, поэтому у них много общего в приспособительных особенностях и возникает разная степень взаимодействий. По характеру этих взаимодействий популяции разных видов чрезвычайно различны. У некоторых видов все особи живут поодиночке, независимо друг от друга, лишь изредка встречаясь для размножения (например, жуки-жужелицы или некоторые пауки). Другие образуют в неблагоприятные периоды скопления, вместе переживают зиму или засуху в подходящих условиях (божьи коровки). У третьих создаются временные или постоянные семьи, объединяющие родителей и потомство.
Шимпанзе живут в тропических лесах Африки группами до 28 особей и связаны сложными родственными отношениями. Один из самцов, обычно наиболее сильный и опытный, является вожаком, его слушаются все остальные. Положение особи в группе обычно определяется ее полом и возрастом. Устанавливается сложная система отношений соподчинения. Она проявляется в основном в распределении пищи. Шимпанзе – очень шумные, но мирные обезьяны. Проявления агрессии у них сравнительно редки. Мать долго заботится о детеныше, ей помогают его старшие братья и сестры. Взрослые часто проявляют устойчивые дружеские привязанности к кому-нибудь из группы, не обязательно того же положения. Шимпанзе стремится к обществу других. Он может покинуть свою группу и присоединиться к соседней.
Иногда шимпанзе охотятся поодиночке на животных среднего размера: мартышек, молодых антилоп и других. На дележ добычи собираются все, и практически каждый получает свою часть по эстафете от старших к младшим в соответствии со своим положением в группе.
Есть виды, у которых в пределах популяций особи объединяются в крупные группы, – стаи, стада, колонии, где совершают совместные действия (защиту, миграции, добывание пищи). Отношения в популяциях – это внутривидовые взаимодействия. В популяциях встречаются все типы биотических связей, характерные для разных видов, но наиболее распространены два прямо противоположных типа: конкуренция и мутуализм. Например, всем известно, что грачи при прилете сначала устраивают драки из-за пригодных мест для гнезда, а затем сообща защищают свою колонию и совместно кормятся на пашнях. В стаях собак или обезьян более сильные особи имеют первое право на пищу (результат конкурентных отношений), но они же в наибольшей мере защищают всю стаю (взаимовыгодные связи).
Отношения особей внутри популяций могут быть очень разнообразными, от полной нейтральности до тесного семейного и группового образа жизни у разных видов.
Среди рыб глубоководных удильщиков существуют необычайные для позвоночных животных связи между самцами и самками по типу паразит-хозяин. Маленький самец сначала прикрепляется к телу взрослой самки зубами, а затем полностью прирастает к ней. Его челюсти, зубы, глаза и кишечник редуцируются, кровеносные сосуды соединяются с кровеносными сосудами самки, и он превращается в ее придаток. Сохраняются лишь собственные жабры и половая система. Карликовые самцы, паразитирующие на самках, встречаются также у некоторых донных беспозвоночных животных.
Численность, плотность и структура популяций
Главная характеристика любой популяции – это ее численность. Она сразу говорит нам о том, хорошо или плохо чувствует себя вид в данных условиях. Однако не всегда легко получить эту характеристику, так как для этого надо пересчитать всех особей популяции. Поэтому чаще используют другой показатель – плотность популяции. Плотность – это число особей, приходящихся на единицу пространства, которую мы выбираем для учета, например число растений пшеницы или одуванчика на квадратный метр или число рачков-дафний в литре воды из данного водоема. Таким образом, можно количественно сравнивать разные популяции, независимо от общего размера занимаемой ими территории. Соотношение особей разного пола или разного возраста – показатели структуры популяции. Структура – это соотношение частей в любой системе. Популяции можно сравнивать и по распределению особей в пространстве, т. е. по их пространственной структуре, и по другим признакам. Все эти признаки – количественные. Следовательно, популяции характеризуются прежде всего количественными показателями. Ведя наблюдения за отдельными популяциями, мы должны уметь учитывать и рассчитывать, оценивать и прогнозировать их численность.
Основные процессы, происходящие в популяции. Основные процессы, происходящие в популяциях, – это рождение и смерть отдельных особей, их частичное расселение за пределы данной территории или появление вселенцев из других популяций. Эти процессы отражаются в соответствующих характеристиках: рождаемость, смертность, вселение (иммиграция) и выселение (эмиграция).
Указанные характеристики отражают скорость, интенсивность происходящих процессов и включают в себя единицы времени, т. е. это временные показатели. Так, рождаемость – это число особей, родившихся в популяции за месяц, год или десятилетие, смертность – число особей, погибших за это же время. Эти характеристики можно выражать в долях или процентах от общей численности. Например, рождаемость в 20% за год означает, что за это время на каждые 100 особей популяции появилось 20 новых.
Соотношение процессов рождаемости, смертности, вселения и выселения определяет численность конкретных популяций.
Человек в своей конкретной практике имеет дело не с отдельными особями или целыми видами, а именно с популяциями растений, животных и микроорганизмов. Домашние животные также живут по популяционным законам. Поэтому так важно понимание этих законов и экологически грамотное управление численностью популяций.
biolicey2vrn.ru
10. Популяция. Популяционная структура вида. Понятие о генетической целостности популяции. Пространственная структура популяции, типы распределения особей в пространстве.
Популяцией в экологии называют группу особей одного вида, находящихся во взаимодействии между собой и совместно населяющих общую территорию.
Члены одной популяции оказывают друг на друга не меньшее воздействие, чем физические факторы среды или другие обитающие совместно виды организмов. В популяциях проявляются в той или иной степени все формы связей, характерные для межвидовых отношений, но наиболее ярко выражены мутуалистические (взаимно полезные) и конкурентные. Специфические внутривидовые взаимосвязи– это отношения, связанные с воспроизводством: между особями разных полов и между родительским и дочерним поколениями.
При половом размножении обмен генами превращает популяцию в относительно целостную генетическую систему. Если перекрестное оплодотворение отсутствует и преобладает вегетативное, партеногенетическое или другие способы размножения, генетические связи слабее и популяция представляет собой систему клонов, или чистых линий, совместно использующих среду. Такие популяции объединены в основном экологическими связями. Во всех случаях в популяциях действуют законы, позволяющие таким образом использовать ограниченные ресурсы среды, чтобы обеспечить оставление потомства. Достигается это в основном через количественные изменения населения. Популяции многих видов обладают свойствами, позволяющими им регулировать свою численность.
Поддержание оптимальной в данных условиях численности называют гомеостазом популяции. Гомеостатические возможности популяций по-разному выражены у различных видов. Осуществляются они также через взаимоотношения особей.
Таким образом, популяции, как групповые объединения, обладают рядом специфических свойств, которые не присущи каждой отдельно взятой особи.
Основные характеристики популяций:
1) численность– общее количество особей на выделяемой территории;
2) плотность популяции – среднее число особей на единицу площади или объема занимаемого популяцией пространства; плотность популяции можно выражать также через массу членов популяции в единице пространства;
3) рождаемость– число новых особей, появившихся за единицу времени в результате размножения;
4) смертность – показатель, отражающий количество погибших в популяции особей за определенный отрезок времени;
5) прирост популяции– разница между рождаемостью и смертностью; прирост может быть как положительным, так и отрицательным;
6) темп роста – средний прирост за единицу времени.
Популяции свойственна определенная организация. Распределение особей по территории, соотношения групп по полу, возрасту, морфологическим, физиологическим, поведенческим и генетическим особенностям отражают структуру популяции. Она формируется, с одной стороны, на основе общих биологических свойств вида, а с другой – под влиянием абиотических факторов среды и популяций других видов. Структура популяций имеет, следовательно, приспособительный характер. Разные популяции одного вида обладают как сходными особенностями структуры, так и отличительными, характеризующими специфику экологических условий в местах их обитания.
8.2. Популяционная структура вида
Каждый вид, занимая определенную территорию (ареал), представлен на ней системой популяций. Чем сложнее расчленена территория, занимаемая видом, тем больше возможностей для обособления отдельных популяций. Однако в не меньшей степени популяционную структуру вида определяют его биологические особенности, такие, как подвижность составляющих его особей, степень их привязанности к территории, способность преодолевать естественные преграды.
8.2.1. Степень обособленности популяций
Если члены вида постоянно перемещаются и перемешиваются на обширных пространствах, такой вид характеризуется небольшим числом крупных популяций. Большими миграционными способностями отличаются, например, северные олени и песцы. Результаты мечения показывают, что песцы перемещаются за сезон на сотни, а иногда более чем на тысячу километров от мест размножения. Северные олени совершают регулярные сезонные кочевки также в масштабах сотен километров. Границы между популяциями таких видов проходят обычно по крупным географическим преградам: широким рекам, проливам, горным хребтам и т. п. В некоторых случаях подвижный вид при относительно небольшом ареале может быть представлен одной-единственной популяцией, например кавказский тур, стада которого постоянно кочуют по двум основным хребтам этого горного массива.
При слабо развитых способностях к перемещению в составе вида формируется множество мелких популяций, отражающих мозаичность ландшафта. У растений и малоподвижных животных число популяций находится в прямой зависимости от степени разнородности среды. Например, в горных районах территориальная дифференцировка таких видов всегда более сложна, чем на ровных открытых пространствах. Примером вида, у которого множественность популяций определяется не столько дифференцировкой среды, сколько особенностями поведения, является бурый медведь. Медведи отличаются большой привязанностью к местам своего обитания, поэтому в пределах обширного ареала представлены множеством относительно мелких группировок, отличающихся друг от друга по ряду свойств.
Степень обособленности соседних популяций вида очень различна. В некоторых случаях они резко разделены территорией, непригодной для обитания, и четко локализованы в пространстве, например популяции окуня и линя в изолированных друг от друга озерах или популяции пластинчатозубой крысы, белоусой славки, индийской камышевки и других видов в оазисах и долинах рек среди пустынь.
Противоположный вариант – сплошное заселение видом обширных территорий. Такой характер распространения свойствен, например, малым сусликам в сухих степях и полупустынях. В этих ландшафтах плотность их населения повсеместно высока. Отдельные непригодные для жизни участки легко преодолеваются при расселении молодняка, а в благоприятные годы на них возникают временные поселения. Здесь вычленить границы между популяциями можно лишь условно, между областями с разной плотностью населения.
Пример сплошного распространения вида – семиточечная божья коровка.
В пределах одного и того же вида могут быть популяции как с хорошо различимыми, так и со смазанными границами (рис. 95).
Между популяциями осуществляется обмен отдельными особями, который может быть либо достаточно регулярным, либо эпизодическим. При сезонных кочевках ворон, например, часть молодых птиц ежегодно остается на местах зимовок, образуя пары с представителями оседлого населения. Связь между населением отдельных видов рыб в озерах осуществляется значительно реже, например в годы с особо сильными паводками, когда обособленные водоемы соединяются в единую водную систему.
Связи между популяциями поддерживают вид как единое целое. Слишком длительная и полная изоляция популяций приводит обычно к образованию новых видов.
Различия между отдельными популяциями выражены в разной степени. Они могут затрагивать не только их групповые характеристики, но и качественные особенности физиологии, морфологии и поведения отдельных особей (рис. 96). Эти различия создаются в основном под влиянием естественного отбора, приспосабливающего каждую популяцию к конкретным условиям ее существования.
Зайцы-беляки из разных частей ареала различаются характером окраски, размерами, строением пищеварительной системы. Например, длина слепой кишки у зайцев полуострова Ямал в 2 раза больше, чем у представителей из лесостепного Урала. Это связано с характером питания, разной долей грубых кормов в составе рациона.
8.3.3. Пространственная структура популяций растений и животных
Типы распределения особей в пространстве. Занимаемое популяцией пространство предоставляет ей средства к жизни. Каждая территория может прокормить лишь определенное число особей. Естественно, что полнота использования имеющихся ресурсов зависит не только от общей численности популяции, но и от размещения особей в пространстве. Это наглядно проявляется у растений, площадь питания которых не может быть меньше некоторой предельной величины. Перехватывая корнями питательные вещества и воду, затеняя пространство, выделяя ряд активных веществ, каждое растение распространяет свое влияние на определенную площадь, поэтому оптимальным для популяции является такой интервал между соседними экземплярами, при котором они не влияют отрицательно друг на друга, но при этом не остается и недоиспользованного пространства.
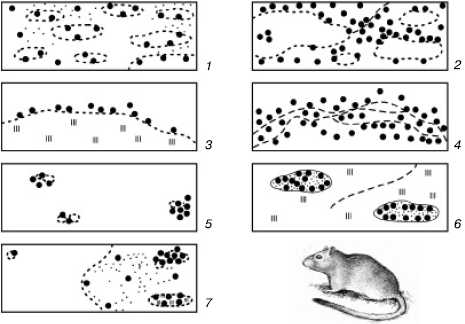
Рис. 104. Основные варианты размещения колоний больших песчанок (по Е. В. Ротшильду, 1966):
1– сплошные равномерные поселения;2– сплошные кружевные поселения;3– узколенточные;4– широколенточные;5– мелкоостровные;6– крупноостровные;7– отдельные скопления колоний
В природе изредка встречается почти равномерное упорядоченное распределение особей на занимаемой территории, например в уплотненных популяциях сидячих морских полихет, в чистых зарослях некоторых растений. Однако чаще всего члены популяции распределяются в пространстве неравномерно (рис. 104), что обусловлено двумя причинами: во-первых, неоднородностью занимаемого пространства, а во-вторых, некоторыми особенностями биологии видов, которые способствуют возникновению скоплений индивидуумов. У растений такая агрегированность возникает, например, при вегетативном размножении, при слабом распространении семян и прорастании их вблизи материнской особи; у животных – при групповом образе жизни семьями, стадами, колониями, при концентрациях для размножения, перезимовки и т. п.
Неравномерное размещение членов популяции может проявляться в двух крайних вариантах со всевозможными переходами между ними: 1) в резко выраженной мозаичности с незанятым пространством между отдельными скоплениями особей и 2) в распределении случайного, диффузного типа. Примером первого могут служить гнездовья грачей, поселяющихся колониями в рощах или парках, к которым примыкают благоприятные кормовые угодья. Диффузное распределение встречается в природе, если члены популяции относительно независимы друг от друга и обитают в однородной для них среде. Таково, например, размещение мучных хрущаков Tribolium confusum в муке, личинок-поденок в воде ручья, норок пауков-каракуртов на лугах и т. п.
В каждом конкретном случае тип распределения в занимаемом пространстве оказывается приспособительным, т. е. позволяет оптимально использовать имеющиеся ресурсы. Способы, которыми достигается рациональный характер размещения, определяются системой взаимоотношений между членами популяции.
Растения в ценопопуляции чаще всего распределены крайне неравномерно, образуя более или менее изолированные группы, скопления, так называемые микроценопопуляции, субпопуляции илиценопопуляционные локусы. Эти скопления отличаются друг от друга числом особей, плотностью, возрастной структурой, протяженностью. Часто более плотный центр скопления окружен особями, расположенными менее плотно.
Пространственная неоднородность ценопопуляции связана с характером развития скоплений во времени. У люцерны серповидной, например, семена обычно опадают в непосредственной близости от материнского растения, поэтому около обильно плодоносящих средневозрастных генеративных растений образуются скопления молодых (проростков, ювенильных и имматурных). Эти скопления характеризуются большой плотностью. По мере того как особи переходят в следующие возрастные состояния, скопления меняют возрастную структуру, изреживаются. Одновременно происходит приживление новых зачатков в пределах скопления, оно уплотняется, усложняется его структура, расширяется занятая им территория. Если приживление зачатков происходит за пределами скопления, то возникает новое. Скопления могут частично сливаться друг с другом, т. е. достигать более высокого уровня агрегированности.
Таким образом, жизнь ценопопуляции протекает в форме несинхронных возрастных изменений различных локусов, при этом меняется и ее пространственная структура, ибо изменяются конфигурация, протяженность локусов и их местоположение в фитоценозе.
У животных благодаря их подвижности способы упорядочивания территориальных отношений более разнообразны по сравнению с растениями. Даже у сидячих форм имеется ряд приспособлений к рациональному размещению в пространстве. У асцидий и мшанок растущий край колонии, наталкиваясь на колонию другого вида, растет поверх нее и в конечном счете подавляет ее. Но если встречаются колонии одного вида, каждая из них тормозит рост соседней и они начинают распространяться в другом направлении. При полном окружении колониями своего вида вегетативное размножение прекращается, но усиливается образование половых продуктов и подвижных личинок.
Личинки устриц, морских желудей, сидячих полихет, руководствуясь химическими стимулами, оседают обычно в местах, где уже имеются особи того же вида. Для осевшей личинки перед окончательным прикреплением характерно специальное поисковое поведение, позволяющее ей занять определенное место в скоплении. По наблюдениям, циприсовидные личинки морского желудя Balanus balanoides обычно прикрепляются на расстоянии не менее 2,5 мм от молодых и не менее 2 мм от старых особей своего вида. В то же время они свободно оседают в близком соседстве и даже на поверхности представителей других видов.
У высших животных внутрипопуляционное распределение регулируется системой инстинктов. Им свойственно особое территориальное поведение – реакция на местонахождение других членов популяции. Инстинкты, поддерживающие размещение по территории отдельных особей или групп в популяциях, существуют у птиц, млекопитающих, пресмыкающихся, ряда рыб, в меньшей степени – у амфибий. Они выражены также у многих видов беспозвоночных со сложной нервной системой – ряда насекомых, пауков, крабов, осьминогов и др.
По типу использования пространства все подвижные животные делятся на две основные группы: оседлых икочевых.
При оседлом существовании животное в течение всей или большей части жизни использует довольно ограниченный участок среды. Такие животные отличаются инстинктами привязанности к своему участку, а в случае вынужденного переселения – стремлением вернуться на хорошо знакомую территорию. Это «чувство дома» получило в экологии название «хоминг» (от англ. home– дом). Многие виды возвращаются к месту размножения даже после длительных и дальних миграций. Известно, например, что одна и та же пара скворцов из года в год может занимать «свой» скворечник. Хоминг голубей использовали даже в практических целях – для переноса почты.
Оседлый образ жизни имеет значительные биологические преимущества. На хорошо знакомой территории животное свободно ориентируется, тратит меньше времени на поиски корма, кратчайшим путем спасается в известные ему укрытия. Кроме того, многие оседлые виды создают систему запасов, прокладывают тропинки, строят дополнительные гнезда, норы, что помогает им выжить. Например, у белок есть основное гнездо, где выводится молодняк, и несколько дополнительных, в которые зверьки скрываются от непогоды или врагов. Белки создают также серии кладовых, запасая на зиму орехи, семена, грибы. Длительное использование определенной территории помогает более полному ее освоению.
На чужом участке поведение животного меняется. Наблюдения за сусликами показали, например, что такие зверьки отличаются суетливостью, неуверенностью передвижения, часто осматриваются, убежища находят лишь случайно и поэтому чаще гибнут, чем хозяева территории.
Уменьшение вероятности гибели от хищников на знакомой территории доказано и в экспериментах. Например, при выпускании ушастой совы в комнату, где находились хомячки, оказалось, что зверьков, ранее ознакомившихся с обстановкой комнаты, сова ловила в пять раз реже, чем тех, которые попадали в нее впервые.
Однако оседлый образ жизни таит в себе угрозу быстрого истощения ресурсов, если плотность популяции окажется слишком высокой. У оседлых видов выработались такие приспособительные особенности поведения, которые обеспечивают разграничение мест обитания между отдельными особями, семьями или другими внутрипопуляционными группировками. Общая площадь, занимаемая популяцией, оказывается поделена на отдельные индивидуальные или групповые участки, чем достигается упорядоченное использование запасов пищи, естественных укрытий, мест для размножения и т. п.
studfiles.net