
Цитоплазма бактерий — Справочник химика 21
Цитоплазма бактерий. Все содержимое клетки, ограниченное клеточной стенкой, называется протопластом. Протопласт состоит пз цитоплазматической мембраны и живого вещества клетки — цитоплазмы, или протоплазмы. Цитоплазма бактерий является бесцветной, прозрачной, слегка вязкой. [c.249]
Плазмиды наиболее часто используют в качестве векторов. Плазмиды -небольшие кольцевые двухцепочечные ДНК из цитоплазмы бактерий, они могут содержать от 2 до 100 тыс. пар оснований. Каждая плазмида имеет гены, которые могут реплицироваться, транскрибироваться, транслироваться независимо от хромосомных генов, но одновременно с ними. Плазмиды можно перемещать из одной клетки в другую их можно встраивать в другие гены, которые затем переносятся вместе с плазмидой и становятся частью генома клетки-хозяина. [c.61]
Рибосомы находятся в цитоплазме клеток.
Достигнув определенных размеров, диктуемых соотношением объемов ядра и цитоплазмы, бактерии переходят к бесполому размножению путем простого деления, т. е. путем деления на две идентичные дочерние клетки (рис. 2.11). Клеточному делению предшествует репликация [c.27]
Не промывая, препараты подсушивают фильтровальной бумагой и докрашивают 0,25%-ным раствором светлого зеленого или хризоидина. Затем промывают водой и высушивают. На препарате К цитоплазма бактерий окрашена в желто-коричневый цвет, включения волютина — в вишнево-красный.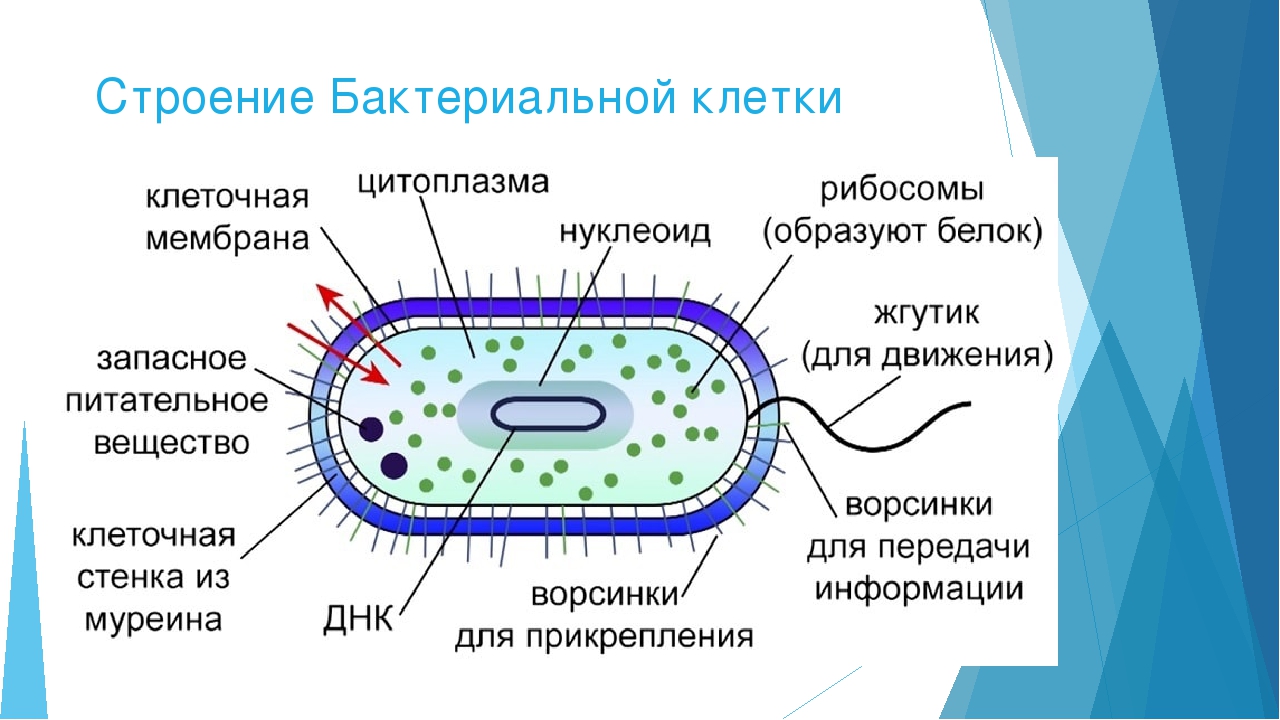
Ртутьредуктаза представляет собой флавопротеин, локализованный в цитоплазме бактерий. [c.460]
Цитоплазма бактерий окрашивается в желтый, а волютин — в темно-синий, почти черный цвет. [c.51]
В цитоплазме бактерий не обнаружены эндоплазматическая сеть и митохондрии, ио имеются рибосомы 703. У гетеротрофных бактерий есть мезосомы — мембранные структуры — производные цитоплазматической мембраны. Нуклеоид в отличие от ядра высших организмов не окружен ядерной мембраной и не содержит ядрышка, он может быть разветвленной формы, имеет одну хромосому, состоящую из кольцевой ДНК, не связанной с гистонами. У бактерий обнаружены дополнительные генетические элементы, содержащие ДНК в виде плазмид, которые могут быть автономны в цитоплазме или включены в хромосомы. Эта особенность плазмид используется в генной инженерии.
Матрикс митохондрии Или цитоплазма бактерии [c.
 78]
78]А хН-зависимая аккумуляция 5 в отсеке, заряженном отрицательно (например, в матриксе митохондрии или в цитоплазме бактерии), описывается уравнением (34) [c.146]
В клетке бактерий обособленное ядро отсутствует, хотя ядерные белки (нук-леопротеиды) находятся в цитоплазме бактерий в большом количестве. [c.20]
Бактерии настолько малы, что находятся на грани разрешения обычного светового микроскопа. Их линейные размеры достигают всего лишь порядка 1 мкм. Поэтому в течение долгого времени было трудно при непосредственном визуальном наблюдении получить информацию об их внутренней структуре. Однако с появлением электронного микроскопа оказалось возможным выявить детальное строение бактериальной клетки, как это можно видеть на приведенной электронной микрофотографии (фиг. 21). Следует отметить, что увеличение на этой микрофотографии в пять раз больше, чем на предыдущей микрофотографии (фиг. 20). Следовательно, размер всей бактериальной клетки не превышает размера митохондрий, находящихся в цитоплазме клеток эукариотов.
Тесная связь между функциями ядра и цитоплазматическими эндосимбионтами была продемонстрирована в конце 60-х годов у Amoeba proteus К.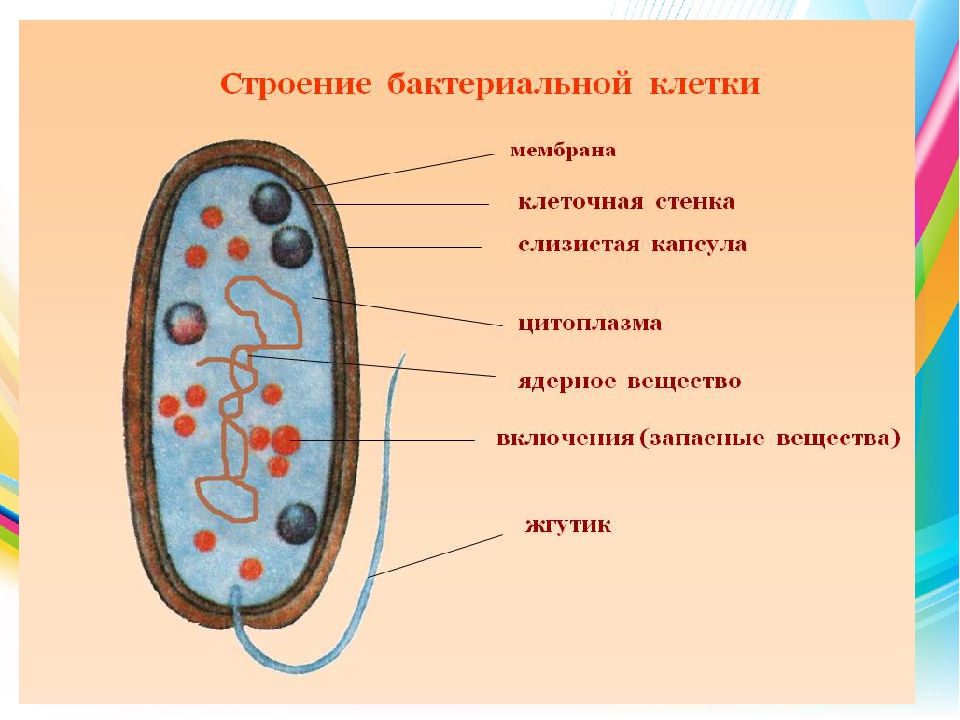 Джеоном. Культура А. proteus была случайно заражена бактериями, которые проникли в цитоплазму простейшего и размножались там до численности 150 тыс. шт. на клетку. Большинство амеб погибло, однако часть из них выжила и активно делилась в присутствии инфицировавшей их цитоплазму бактерии. Число бактериальных клеток в амебе достигало теперь примерно 50 ООО. [c.249]
Джеоном. Культура А. proteus была случайно заражена бактериями, которые проникли в цитоплазму простейшего и размножались там до численности 150 тыс. шт. на клетку. Большинство амеб погибло, однако часть из них выжила и активно делилась в присутствии инфицировавшей их цитоплазму бактерии. Число бактериальных клеток в амебе достигало теперь примерно 50 ООО. [c.249]
Трехмерная структура и расположение в мембране. Я+ — АТФ-синтазный комплекс так велик, что выдается в воду на довольно большое расстояние с одной стороны мембраны. Выступающая часть, которая представляет собой фактор Fi, обращена в цитоплазму бактерий, матрикс митохондрии или строму хлоропласта. [c.132]
Давайте обратимся к механизму транспорта К+ и Ыа+ через бактериальную мембрану. Известно, что между цитоплазмой бактерии и внешней средой существует разность электрических потенциалов, поддерживаемая работой белков-генераторов в бактериальной мембране. Откачивая протоны изнутри клетки наружу, белки-генераторы тем самым заряжают внутренность бактерии отрицательно. В этих условиях накопление ионов К+ внутри клетки могло бы происходить просто за счет электрофореза — движения Боложительно заряженного иона калия в отрицательно заряженную цитоплазму бактерии.
В этих условиях накопление ионов К+ внутри клетки могло бы происходить просто за счет электрофореза — движения Боложительно заряженного иона калия в отрицательно заряженную цитоплазму бактерии.
Цитоплазматическая мембрана является трехслойной структурой и окружает наружную часть цитоплазмы бактерий. По структуре она похожа на цитоплазматическую мембрану клеток животных состоит из двойного слоя липидов, главным образом фосфолипидов со встроенными поверхностными и интефальны-ми белками, как бы пронизываюш.ими насквозь структуру мембраны. Некоторые из них являются пермеазами, участвующими в транспорте веществ. Цитоплазматическая мембрана является динамической структурой с подвижными компонентами, поэтому ее представляют как мобильную, текучую структуру. Она участвует в регуляции осмотического давления, транспорте веществ и энергетическом метаболизме клетки (за счет ферментов цепи переноса электронов, АТФ-азы и др.). [c.25]
Цитоплазма бактерий занимает основной объем клетки и состоит из растворимых белков.
Генетика микроорганизмов как учение о наследственности и изменчивости имеет характерные особенности, соответствующие их сфоению и биологии. Наиболее изучена генетика бактерий, характерными чертами которых являются малые размеры и большая скорость размножения бактериальной клетки, что позволяет проследить генетические изменения в течение небольшого промежутка времени на большом числе популяций.
Ультраструктура бактериальной клетки
Бактериальная клетка, несмотря на внешнюю простоту строения, представляет собой весьма сложный организм, для которого характерны процессы, свойственные всем живым существам.
Ультраструктуру бактерий удалось детально изучить после создания электронных микроскопов с большой разрешающей способностью, разработки техники получения ультратонких срезов клеток, появления фазово-контрастной микроскопии, усовершенствования методов микрохимических анализов. Разнообразные методы исследований дали возможность определить различные поверхностные и внутренние структуры у бактерий, (рис. 5).
К внешним структурам обычно относят капсулы, жгутики, фимбрии и пили, а также клеточную стейку, под которой расположена цитоплазматическая мембрана. Внутреннее содержимое бактерий представлено цитоплазмой, в которой находятся нуклеоид, рибосомы и мембранные структуры, а также разнообразные включения. Бациллы и некоторые другие бактерии образуют споры.
Капсула. Большинство бактерий окружены слоем вещества, расположенного поверх клеточной стенки. Это слизистое образование — капсула (рис. 6). Бывают макрокапсулы (толщина слоя 0,2 мкм), микрокапсулы (менее 0,2 мкм), слизистый слой и растворимая слизь.
По химическому составу капсулы бактерий можно разделить на два типа. Одни представлены полисахаридами, другие — полипептидами. Однако встречаются капсулы, состоящие из липидов, (у туберкулезных бактерий), гетерополисахаридов и других веществ. Капсулы содержат до 98% воды. Поэтому они создают дополнительный осмотический барьер, а также защищают клетку от механических повреждений и высыхания. Капсулы защищают клетки и от других неблагоприятных воздействий окружающей среды.
Замечено, что бактерии, имеющие капсулы, могут жить в такой среде, в которой рост бактерий без капсул ограничен.
Жгутики. Существуют два типа подвижных бактерий: скользящие и плавающие. Скольжение наблюдается у миксобактерий. и серных бактерий. Эти организмы могут совершать скольжение по поверхности в результате волнообразных сокращений, вызывающих периодическое изменение формы клетки.
Плавающие палочковидные бактерии передвигаются с помощью особых нитевидных придатков — жгутиков.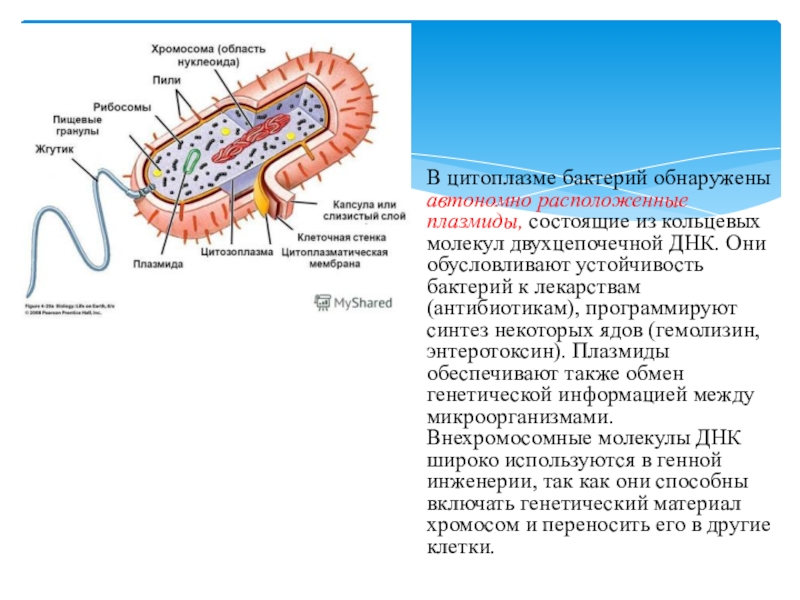 За счет жгутиков передвигается большинство спирилл. Кокки, за исключением отдельных видов, не имеют жгутиков.
За счет жгутиков передвигается большинство спирилл. Кокки, за исключением отдельных видов, не имеют жгутиков.
Бактерия с одним жгутиком называется монотрихом; бактерия с пучком жгутиков на одном конце клетки — лофотрихом; на обоих концах — амфитрихом; бактерия со жгутиками, расположенными по всей поверхности клетки, называется перитрихом (рис. 7).
Число жгутиков различно у разных видов бактерий. Например, спириллы (Spirillum) имеют от 5 до 30 жгутиков, вибрионы (Vibrio) — 1 или 2—3 жгутика на полюсе клетки, а у палочковидных бактерий Proteus vulgaris и Clostridium tetani обнаружено от 50 до 100 жгутиков. Толщина жгутиков колеблется от 10 до 20 нм, длина — от 3 до 15 .мкм, причем у одной и той же бактериальной клетки длина может изменяться в зависимости от состояния культуры и факторов внешней среды. В химическом отношении жгутики представляют собой белок флагеллин.
Белковые молекулы, из которых состоят жгутики, собраны в спиральные цепи, закругленные вокруг полой сердцевины.
Жгутики хорошо видны в электронном микроскопе, для наблюдения через оптический микроскоп требуется их специальная обработка. Жгутики не относятся к жизненно важным структурам бактериальной клетки. Так, бактерии, обладающие жгутиками, можно вырастить в условиях, при которых эти структуры у них не развиваются. У подвижных бактерий наблюдаются «фазовые вариации», то есть в течение одной фазы развития жгутики имеются, в другой — отсутствуют. Жгутики можно разрушить, а клетка останется жизнеспособной.
Жгутики прикрепляются к особой структуре — базальному тельцу, расположенному под цитоплазматической мембраной. Движение жгутику сообщает через его основание жгутиковый «мотор» — базальное тельце, состоящее из центрального стержня, вставленного в систему колец, которые вращаются относительно друг друга.
Бактериальные клетки со жгутиками двигаются со скоростью, которая зависит от особенностей их аппарата движения и свойств среды – вязкости, температуры рН, осмотического давления и др. Большинство бактерий за секунду проходят расстояние, равное размерам их клетки.
Большинство бактерий за секунду проходят расстояние, равное размерам их клетки.
Однако некоторые бактерии при благоприятных условиях за то же время могут передвигаться на расстояние, превышающее размеры клетки в 50 раз и более.
Бактерии передвигаются беспорядочно, но способны и к направленным движениям, так называемым таксисам. В зависимости от внешних факторов, под воздействием которых происходит движение, различают хемотаксис, обусловленный разницей в концентрации химических веществ в среде, аэротаксис, связанный с разницей в содержании кислорода, и, наконец, фототаксис, когда условием направленного движения бактерий является различие в интенсивности освещения.
Фимбрии и пили. Кроме жгутиков, клетки бактерий могут иметь длинные, тонкие, прямые нити — фимбрии. Фимбрии значительно короче и тоньше жгутиков, но более многочисленны. Обнаружены они как у подвижных, так и у неподвижных организмов. Размеры фимбрий 0,3—4 мкм в длину и 5—10 нм в ширину.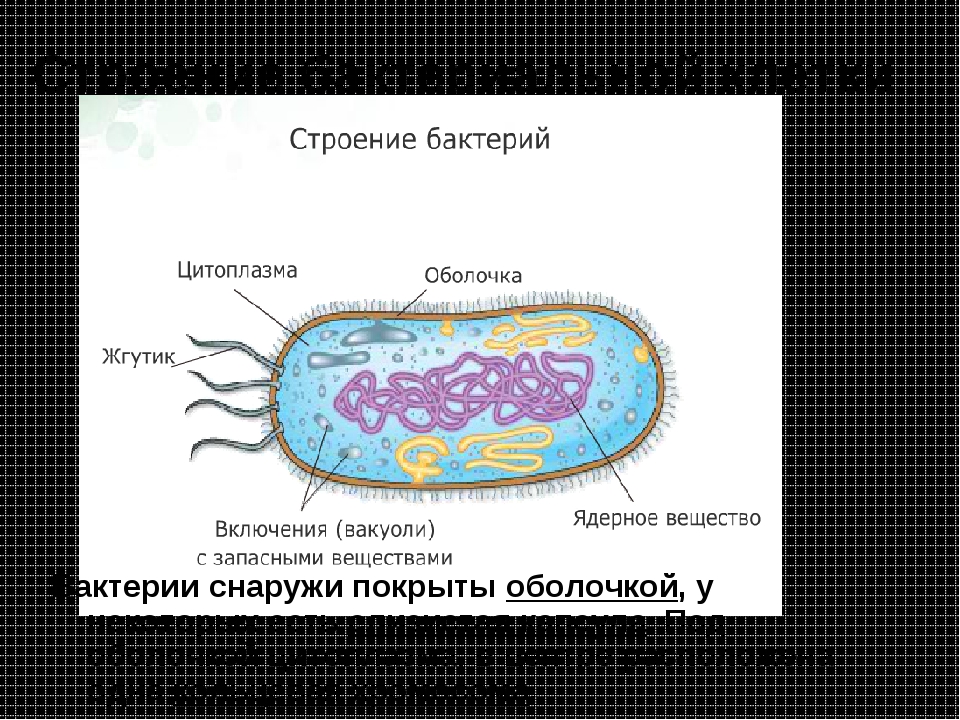 Число их достигает 100—200 и более (до нескольких тысяч) на одну бактериальную клетку.
Число их достигает 100—200 и более (до нескольких тысяч) на одну бактериальную клетку.
Фимбрии состоят из белка — пилина.
В настоящее время известно несколько типов фимбрий, которые отличаются своими функциями. Наиболее изучены функции фимбрий первого и второго типов. Фимбрии первого типа имеют многие бактерии,
Что дало возможность назвать их «фимбриями общего типа». Наличие фимбрий первого типа помогает бактериальной клетке прилипать к другим клеткам или инертному субстрату, или способствовать образованию пленок на поверхности жидкостей, и поэтому считают, что фимбрии этого типа — органы прикрепления (рис. 8).
Большой интерес представляют фимбрии второго типа, так называемые половые фимбрии, или пили, имеющие внутри канал, через который передается генетический материал от одной клетки к другой при конъюгации бактерий. Пили также могут служить для прикрепления патогенных бактерий к тканям животных и человека.
Клеточная стенка — один из главных элементов структуры бактериальной клетки. Клеточная стенка обладает определенной ригидностью (жесткостью), но вместе с тем эластичностью и может изгибаться. Ее можно разрушить ультразвуком, ферментом лизоцимом и другими способами. В результате разрушения клеточной стенки содержимое клетки — цитоплазма с ее включениями, окруженная цитоплазматической мембраной, приобретает шаровидную форму. Такая округлившаяся клетка, образовавшаяся после удаления клеточной стенки у бактерии, называется протопластом (сферопластом). Отсюда следует, что стенка придает ‘бактериальной клетке определенную форму.
Клеточная стенка обладает определенной ригидностью (жесткостью), но вместе с тем эластичностью и может изгибаться. Ее можно разрушить ультразвуком, ферментом лизоцимом и другими способами. В результате разрушения клеточной стенки содержимое клетки — цитоплазма с ее включениями, окруженная цитоплазматической мембраной, приобретает шаровидную форму. Такая округлившаяся клетка, образовавшаяся после удаления клеточной стенки у бактерии, называется протопластом (сферопластом). Отсюда следует, что стенка придает ‘бактериальной клетке определенную форму.
Клеточная стенка имеет и другие функции. Она защищает внутреннее содержимое клетки от действия механических и осмотических сил внешней среды, ей принадлежит важная роль в регуляции роста и деления бактерий и распределении генетического материала.
Толщина клеточной стенки колеблется от 10 до 80 нм и составляет около 20% сухого вещества бактериальной клетки. Клеточная стенка относительно проницаема для крупных молекул. Она связана с цитоплазматической мембраной соединительными тяжами — «мостиками».
Считают, что клеточная стенка ответственна за окрашивание бактерий по Граму. Способность (или, наоборот, неспособность) окрашиваться по Грамму связана с различием в химическом составе клеточных стенок бактерий.
Главным структурным компонентом клеточных стенок большиства исследованных бактерий является пептидогликан (муреин), представляющий собой гетерополимер, который построен из чередующихся остатков N-ацетил-N-глюкозамина и N-ацетилмурамовой кислоты, соединенных в-1,4-связями. Полисахаридные цепи связаны между собой короткими пептидными мостиками. Пептидогликан придает клеточной стенке ригидные свойства, благодаря чему бактериальная клетка способна сохранять свою форму.
У грамположительных бактерий клеточная стенка состоит главным образом из многослойного пептидогликана, в который сложным образом вплетены такие сопутствующие компоненты, как белки, полисахариды, а также так называемые тейхоевые кислоты (полимеры рибитфосфорной и глицеринфосфорной кислот).
У грамотрицательных бактерий пептидогликан однослойный. Сверху пептидогликана располагается структура, получившая название наружной мембраны. Она имеет мозаичное строение и состоит из фосфолипидов, липопротеидов, белков и сложного липополисахарида (ЛПС).
Сверху пептидогликана располагается структура, получившая название наружной мембраны. Она имеет мозаичное строение и состоит из фосфолипидов, липопротеидов, белков и сложного липополисахарида (ЛПС).
Содержание пептидогликана в клеточных стенках колеблется от 50 до 90% у грамположительных бактерий и от 1 до 10% у грамотрицательных.
Таким образом, неодинаковое отношение бактерий к окраске по Граму может быть объяснено различием в количестве пептидогликана и его локализацией в клеточной стенке.
В настоящее время выявлено, что поверхность клеточной стенки некоторых палочковидных форм бактерий покрыта выростами, шипами или буграми.
Клеточной стенки нет у микоплазм, а также у L-форм бактерий. Наименование L-форм (от названия Института имени Листера (Великобритания), где впервые изучены эти формы) получили бактерии, полностью или частично лишенные клеточной стенки (под влиянием антибиотиков или спонтанно без видимой причины) и сохранившие способность к размножению. Для L-форм разных бактерий характерно образование крупных и мелких шаровидных клеток. L-формы описаны у многих болезнетворных и сапрофитных бактерий.
Для L-форм разных бактерий характерно образование крупных и мелких шаровидных клеток. L-формы описаны у многих болезнетворных и сапрофитных бактерий.
Цитоплазматическая мембрана (ЦПМ). К клеточной стенке бактериальной клетки тесно прилегает внешний слой цитоплазмы — цитоплазматическая мембрана, состоящая обычно из двойного слоя липидов, каждая из поверхностей которого покрыта мономолекулярным слоем белка. Мембрана составляет около 8—15% сухого вещества клетки и содержит до 70—90% липидов клетки. Общая толщина мембраны равняется приблизительно 9 нм.
Цитоплазматическая мембрана играет роль осмотического барьера, контролирующего транспорт веществ в бактериальную клетку и из нее. Нередко мембрана дает внутрицитоплазматические впячивания (инвагинации), приводящие к образованию особых телец—мезосом.
Цитоплазматическая мембрана и мезосомы выполняют функции, свойственные мембранам и митохондриям высших организмов, в которых или на которых локализованы ферментные системы-поставщики энергии. В отличие от митохондрий в цитоплазматической мембране и мезосомах бактерий наряду с дыхательными системами ферментов и механизмом регуляции проницаемости располагаются специфичные ферментные системы, участвующие в таких процессах, как азотфиксация, хемосинтез и др.
В отличие от митохондрий в цитоплазматической мембране и мезосомах бактерий наряду с дыхательными системами ферментов и механизмом регуляции проницаемости располагаются специфичные ферментные системы, участвующие в таких процессах, как азотфиксация, хемосинтез и др.
С цитоплазматической мембраной, мезосомами и близкими структурами бактерий связаны также многие другие функции — биосинтез клеточной стенки и капсулы, выделение экзоферментов, деление и спорообразование и т. д.
Цитоплазма. Под цитоплазматической мембраной у бактерий находится цитоплазма. Это коллоидная система, состоящая из воды, белков, жиров, углеводов, минеральных соединений и других веществ, соотношение которых варьирует в зависимости от вида бактерий и их возраста. Цитоплазма бактерий имеет различные структурные элементы — внутрицитоплазматические мембраны, генетический аппарат, рибосомы и включения, остальная часть ее представлена цитозолем.
Цитозоль — это фракция цитоплазмы, которая имеет гомогенную консистенцию и состоит главным образом из белковых макромолекул (растворимых РНК, ферментных белков, продуктов и субстратов различных реакций) и служит поддерживающей средой для клеточных гранул. Изучение структуры цитоплазмы выявило ее мелкогранулярный характер, она представлена цитоплазматическими гранулами диаметром 10—20 нм. Многие из этих гранул являются рибосомами — частицами, состоящими из РНК (60%) и белка (40%). Каждая бактерия содержит от 5000 до 50 000 рибосом, которые служат центрами синтеза белков. Рибосомы совместно с молекулами информационной и транспортных РНК участвуют в синтезе белка в форме не изолированных частиц, а их агрегатов, называемых полирибосомами, или полисомами.
Изучение структуры цитоплазмы выявило ее мелкогранулярный характер, она представлена цитоплазматическими гранулами диаметром 10—20 нм. Многие из этих гранул являются рибосомами — частицами, состоящими из РНК (60%) и белка (40%). Каждая бактерия содержит от 5000 до 50 000 рибосом, которые служат центрами синтеза белков. Рибосомы совместно с молекулами информационной и транспортных РНК участвуют в синтезе белка в форме не изолированных частиц, а их агрегатов, называемых полирибосомами, или полисомами.
В цитоплазме цианобактерий имеются так называемые Тилакоиды (или Фикобилисомы) — мембранные фотосинтезирующие структуры, содержащие хлорофилл и каротиноиды, при помощи которых осуществляется фотосинтез. У пурпурных серобактерий фотосинтезирующие пигменты (бактериохлорофилл и каротиноиды) локализованы в хроматофорах, которые составляют от 40 до 50% массы клетки. Тилакоиды в основном состоят из белков и липидов. Предполагают, что тилакоиды связаны с цитоплазматической или внутренними мембранами.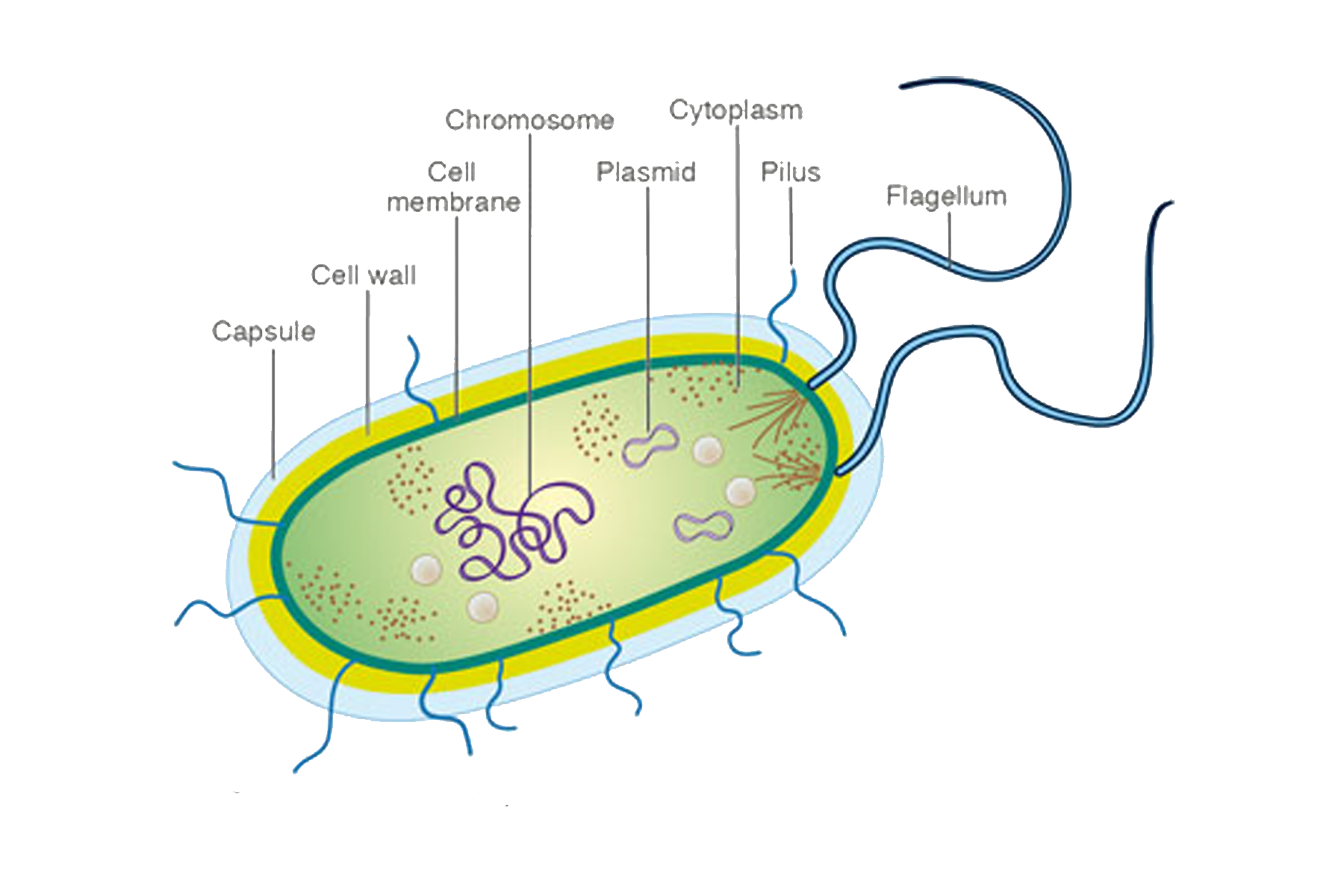 У зеленых бактерий пигменты, участвующие в фотосинтезе, содержатся в мембранных структурах, называемых хлоросомами. Клетки ряда водных бактерий содержат наполненные газом структуры — газовые вакуоли (аэросомы). Некоторые бактерии имеют структуры, называемые полиэдральными телами (в виде многоугольника), или карбоксисомами, где осуществляется процесс связывания С02 (у автотрофных бактерий).
У зеленых бактерий пигменты, участвующие в фотосинтезе, содержатся в мембранных структурах, называемых хлоросомами. Клетки ряда водных бактерий содержат наполненные газом структуры — газовые вакуоли (аэросомы). Некоторые бактерии имеют структуры, называемые полиэдральными телами (в виде многоугольника), или карбоксисомами, где осуществляется процесс связывания С02 (у автотрофных бактерий).
Включения. В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Их присутствие нельзя рассматривать как постоянный признак микроорганизма, обычно они в значительной степени связаны с физическими и химическими условиями среды обитания.
Многие внутрицитоплазматические гранулы состоят из соединений, которые служат для микроорганизмов источником энергии и углерода. Такие соединения обычно образуются, когда микроорганизм снабжается достаточным количеством питательных веществ, и используются, когда он попадает в неблагоприятные в отношении питания условия. В качестве резервных питательных веществ в клетках бактерий могут накапливаться вещества, состоящие из углеводов — гранулы гликогена (крахмала) или гранулезы (близкого к крахмалу полисахарида). При недостаточном поступлении углеродсодержащих веществ в питательную среду гранулы гликогена или гранулезы постепенно исчезают из клеток бактерий.
В качестве резервных питательных веществ в клетках бактерий могут накапливаться вещества, состоящие из углеводов — гранулы гликогена (крахмала) или гранулезы (близкого к крахмалу полисахарида). При недостаточном поступлении углеродсодержащих веществ в питательную среду гранулы гликогена или гранулезы постепенно исчезают из клеток бактерий.
Большая часть бактерий в качестве резервного вещества синтезирует полимер из в — оксимасляной кислоты (поли — в — оксимасляная кислота). У некоторых видов бактерий в клетках накапливаются гранулы жира и волютина. Волютиновые гранулы, называемые еще метахроматическими гранулами, состоят преимущественно из полифосфатов и служат запасным источником фосфора. Волютин обнаруживается в виде крупных, хорошо видимых гранул, образующихся в больших количествах на средах, богатых глицерином или углеводами. В клетках серных бактерий в качестве включений встречается сера, которая образуется в результате окисления сероводорода и видна в виде расположенных непосредственно в цитоплазме блестящих полужидких капелек. Включения серы для аэробных тионовых бактерий, окисляющих сероводород, служат источником энергии. Некоторые серные бактерии наряду с капельками серы имеют зернышки аморфного карбоната кальция, роль которого пока не выяснена.
Включения серы для аэробных тионовых бактерий, окисляющих сероводород, служат источником энергии. Некоторые серные бактерии наряду с капельками серы имеют зернышки аморфного карбоната кальция, роль которого пока не выяснена.
В цитоплазматическом матриксе содержатся также растворимые белки, различные ферменты, РНК, пигменты и низкомолекулярные соединения — углеводы, аминокислоты и нуклеотиды. Наличие в цитоплазме низкомолекулярных соединений обусловливает разность в осмотическом давлении клеточного содержимого и внешней среды. Величина внутриклеточного осмотического давления значительно изменяется у разных микроорганизмов.
Нуклеоид. В цитоплазме бактериальных клеток расположен эквивалент ядра, называемый нуклеоидом. Нуклеоид бактериальной клетки находится в ее центральной части. Предполагают, что в зависимости от стадии развития клетки нуклеоид может быть либо дискретным (прерывистым, состоящим из отдельных форменных структур), либо в виде хроматиновой сети — ядерного вещества, дисперсно распределенного в цитоплазме.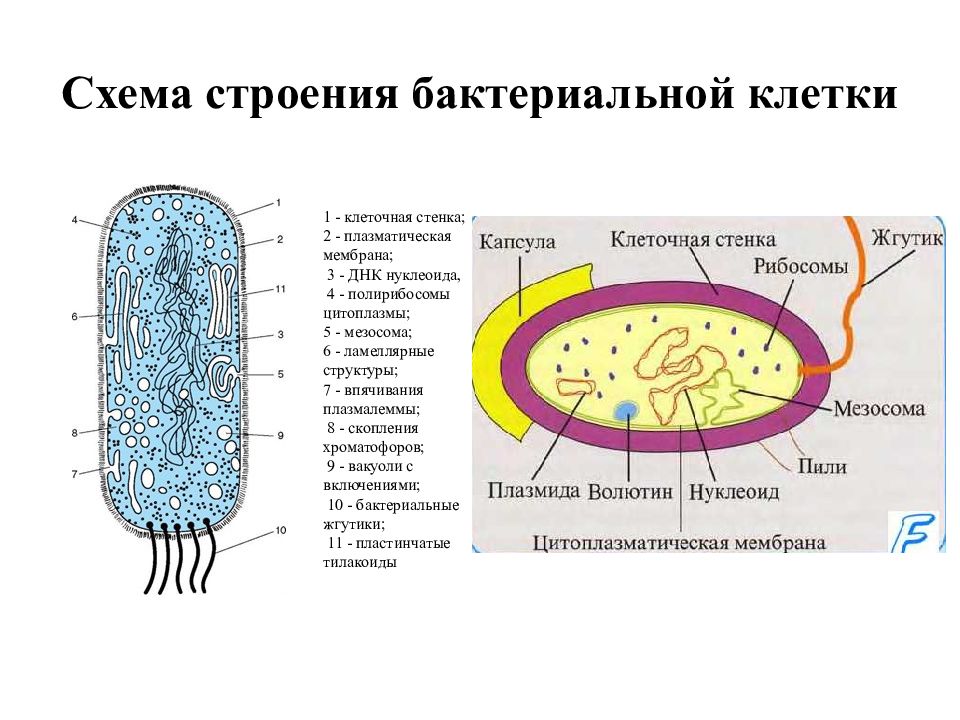
Ядерные структуры, присутствующие в цитоплазме бактерий в дискретном состоянии, имеют палочковидную форму (рис. 9).
Нуклеоиды бактерий содержат ДНК с молекулярной массой 2—3-109. Установлено, что бактериальная ДНК имеет форму свернутой в кольцо нити длиной 1,1 — 1,4 мм, называемой также бактериальной хромосомой и генофорой.
В покоящейся бактериальной клетке обычно содержится один нуклеоид; клетки, находящиеся в фазе, предшествующей делению, имеют два нуклеоида; в фазе логарифмического роста — размножения — до четырех и более нуклеоидов. В условиях, которые отрицательно сказываются на росте бактерий, могут возникать нитевидные многоядерные клетки. Это объясняется нарушением синхронизации между скоростью роста клетки и скоростью деления клетки и нуклеоида.
Нуклеоид бактерий — основной носитель информации о свойствах клетки и основной фактор передачи этих свойств потомству.
Кроме нуклеоида, в цитоплазме бактериальной клетки могут — находиться в сотни раз более короткие нити ДНК — так называемые внехромосомные факторы наследственности, получившие название плазмид. Как выяснено, плазмиды необязательно имеются у бактерий, но они придают организму дополнительные, полезные для него свойства, в частности связанные с размножением, устойчивостью к лекарственным препаратам, болезнетворностью и др.
Как выяснено, плазмиды необязательно имеются у бактерий, но они придают организму дополнительные, полезные для него свойства, в частности связанные с размножением, устойчивостью к лекарственным препаратам, болезнетворностью и др.
Споры и спорообразование. Бактерии рода Bacillus, Clostridium и Desulfotomaculum, так же как и отдельные кокки и спириллы, способны образовывать споры (эндоспоры) — тельца сферической или эллиптической формы, устойчивые к воздействию неблагоприятных факторов (рис.10).
Споры преломляют свет и четко видны в световом микроскопе. Как правило, внутри бактериальной клетки образуется только одна спора. Однако в последнее время у отдельных видов Clostridium обнаружены клетки с двумя и более спорами. Обычно спорообразование начинается, когда бактерии испытывают недостаток питательных веществ или когда в среде в большом количестве накапливаются продукты обмена веществ бактерий. Поэтому споры можно рассматривать как приспособление организма для выживания в неблагоприятных условиях среды.
Формирование спор зависит от условий роста. Споры могут оставаться живыми в условиях, когда вегетативные клетки, то есть клетки, не образовавшие споры, погибают. Большинство спор хорошо переносит высушивание, многие споры нельзя убить даже кипячением в течение нескольких часов. Для их уничтожения требуется температура пара 120° С при давлении его 1 атм. (1,01* 105 Па). При этих условиях споры погибают через 20 мин. В сухом состоянии они погибают лишь при сильном нагревании (до 150—160°С) в течение нескольких часов. Споры отдельных видов бактерий отличаются особенной термоустойчивостью.
В процессе образования спор осуществляется синтез особого соединения — дипиколиновой кислоты (пиридин-2,6-дикарбоновая кислота), обычно отсутствующей у вегетативных клеток бактерий. Дипиколиновая кислота может составлять 10—15% массы сухой споры. Показано, что это вещество накапливается в центральной части споры — ее цитоплазме, образуя с ионами кальция комплекс, который вместе с повышенным содержанием других катионов (магния, марганца и калия)
Обеспечивает пребывание спор в состоянии покоя и их термоустойчивость.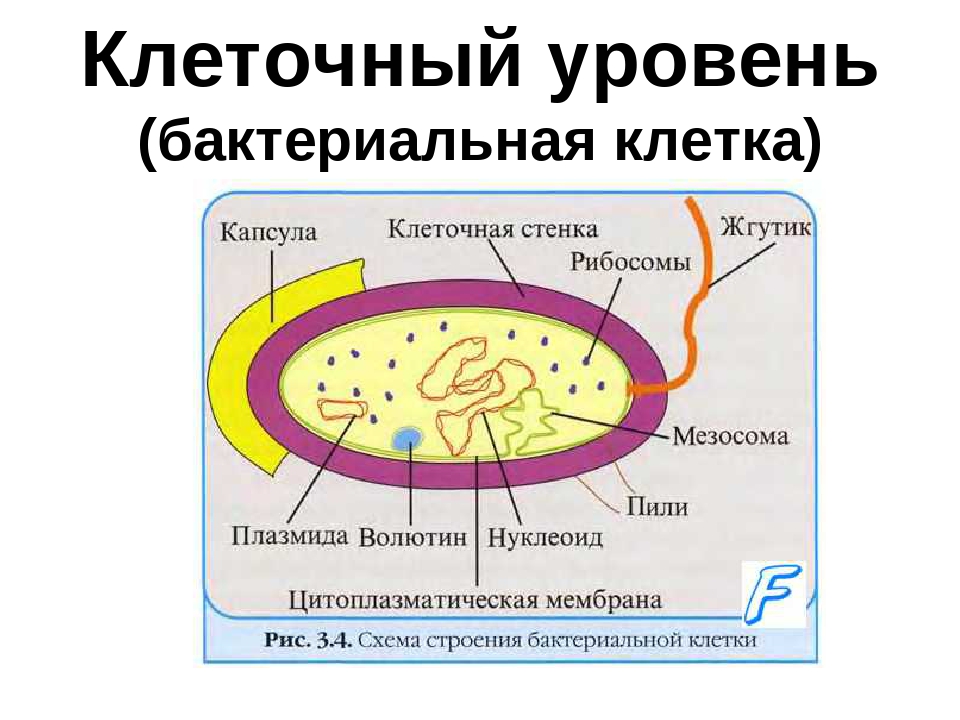
Общая схема спорообразования может быть представлена в следующем виде. В результате неравномерного деления бактериальной клетки, сопровождающегося впячиванием цитоплазматической мембраны, наблюдается обособление части нуклеоида с небольшой частью цитоплазмы. Образовавшаяся проспора затем покрывается цитоплазматической мембраной бактериальной клетки.
Таким образом, внутри клетки бактерии возникает новая клетка — проспора, окруженная двумя мембранами. Затем между мембранами образуется кортикальный слой, или кортекс, состоящий из особых молекул пептидогликана.
Дальнейшее развитие споры заключается в образовании нескольких слоев споровых покровов и ее созревании. Споровые покровы формируются в основном из вновь синтезированных особых белков, а также липидов и гликолипидов. Электронномикроскопическое изучение ультратонких срезов спор многих бактерий показало, что поверх покровов споры образуется еще одна структура — экзоспориум, часто состоящий из ряда слоев и имеющий подчас разнообразную «лепную» форму. Диаметр споры приблизительно равен диаметру клетки, в которой она образовалась, или несколько превышает его. У некоторых бактерий спора формируется на конце клетки, которая при этом несколько расширяется, приобретая вид барабанной палочки. У других бактерий спора образуется в центре клетки, и последняя либо не меняет формы (род Bacillus), либо расширяется в середине, принимая вид веретена (род Clostridium).
Диаметр споры приблизительно равен диаметру клетки, в которой она образовалась, или несколько превышает его. У некоторых бактерий спора формируется на конце клетки, которая при этом несколько расширяется, приобретая вид барабанной палочки. У других бактерий спора образуется в центре клетки, и последняя либо не меняет формы (род Bacillus), либо расширяется в середине, принимая вид веретена (род Clostridium).
После созревания споры клеточная стенка вегетативной части клетки разрушается, и спора выходит в окружающую среду. При попадании в благоприятные условия спора начинает прорастать.
Прорастанию предшествует поглощение спорой воды и последующее набухание. Затем оболочка под влиянием давления, вызванного ростом, разрывается, возникает ростовая трубка. В дальнейшем происходит удлинение освободившегося бактериального организма и, наконец, деление уже удлиненной клетки.
Споры бактерий могут длительное время (десятки, сотни и даже тысячи лет) существовать в покоящемся состоянии.
Имеются микроорганизмы, образующие относительно устойчивые к неблагоприятным условиям среды (температура, кислотность, аэрация и др.) покоящиеся клетки — цисты, не являющиеся спорами. Например, азотобактер образует цисты, устойчивые к высушиванию и теплу.
Известны и другие группы покоящихся клеток (миксоспоры миксобактерий, акинеты цианобактерий, эндоспоры актиномицетов и др.).
Организация и функция структур бактериальной клетки
Более 80 лет назад ученые заметили, что бактерия устойчива к растворам кислот и щелочей.
Было высказано предположение о существовании наружного слоя, который защищает бактериальную клетку от вредных внешних воздействий. Эта «защитная одежда» клеток должна быть достаточно прочной, так как клетка не теряет своей формы даже после смерти. Сейчас удалось не только совершенно точно доказать существование у бактерий клеточной стенки, но и изолировать ее от всей остальной части клетки и даже измерить ее толщину.
Если обработать живые бактерии щелочью, некоторыми ферментами, поместить бактериальную клетку б струю горячей воды или короткое время встряхивать в смеси с мелким песком (3 тысячи встряхиваний в минуту), то клетка лопнет, и содержимое ее вытечет наружу. Отмыв его и поместив в электронный микроскоп, вы увидите пустой «мешок» — это и есть клеточная стенка.
Отмыв его и поместив в электронный микроскоп, вы увидите пустой «мешок» — это и есть клеточная стенка.
Вес ее составляет 20% всей клетки. По новейшим данным, она имеет толщину, равную 200—600 ангстремам, и состоит из трех концентрических слоев. Наружный — липопротеиновый — содержит большое количество жиров, средний — липополисахаридный и, наконец, внутренний — ригидный слой, благодаря которому клетка и сохраняет свою форму. Этот слой состоит из белка и жесткого вещества мукоида. Тонкий химический и физико-химический анализы показали, что белковые молекулы стенки расположены на расстоянии 80 ангстрем друг от друга и сами имеют диаметр, равный 50 ангстрем. Между белковыми молекулами расположены липиды; они играют как бы цементирующую роль.
Такая сложная многослойная структура стенки не случайна. Внутри бактериальной клетки из-за присутствия солей органических соединений и воды создается высокое внутреннее давление, которому и противостоит упругая наружная стенка.
Среда, окружающая бактериальную клетку, таит в себе много вредоносных факторов.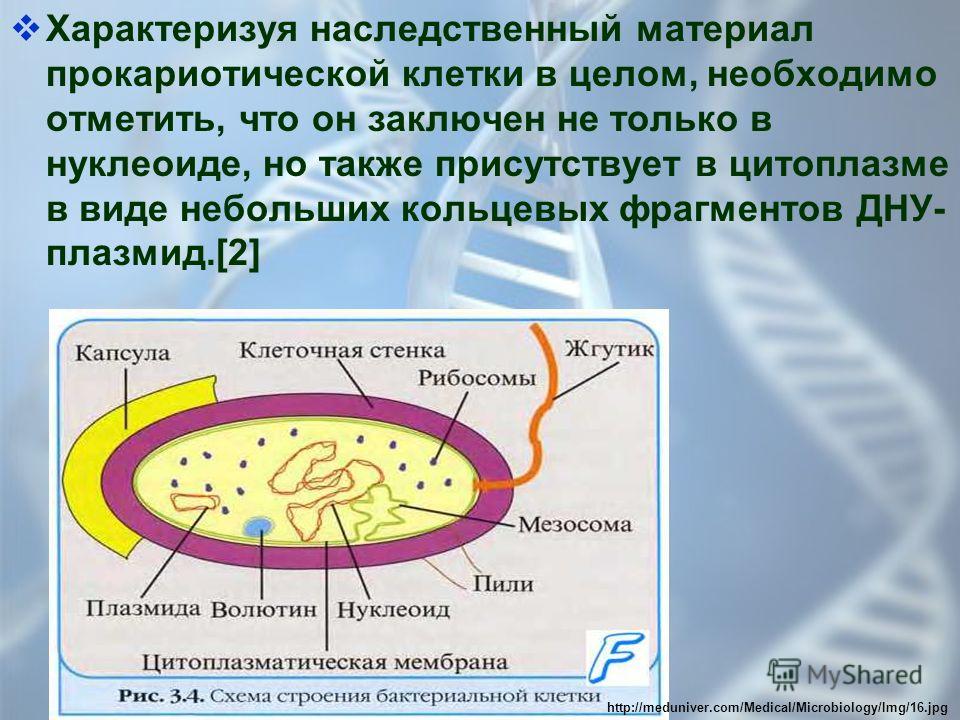 Поэтому, кроме стенки, многие бактерии окружены еще одним защитным слоем — слизистой капсулой. Толщина капсулы у некоторых бактерий вдвое превышает толщину самой клетки (пневмококк). Образование капсулы придает бактерии большую устойчивость к губительным воздействиям внешней среды. Таким образом, клеточная стенка и капсула служат бактерии надежной защитой от разнообразных вредных воздействий: солнечных лучей, давления и др.
Поэтому, кроме стенки, многие бактерии окружены еще одним защитным слоем — слизистой капсулой. Толщина капсулы у некоторых бактерий вдвое превышает толщину самой клетки (пневмококк). Образование капсулы придает бактерии большую устойчивость к губительным воздействиям внешней среды. Таким образом, клеточная стенка и капсула служат бактерии надежной защитой от разнообразных вредных воздействий: солнечных лучей, давления и др.
Что же будет с бактериальной клеткой после удаления клеточкой стенки? Оказывается, если осторожно «раздеть» клетку, имеющую форму палочки, а это можно сделать с помощью лизоцима (фермент, который растворяет клеточную стенку), — она превратится в шар. Такие шарообразные бактерии, лишенные клеточной стенки, называют протопластами.
Протопласты — очень хрупкие образования. Они сохраняют свою форму лишь в концентрированных солевых растворах. В ином случае протопласты лопаются, и их содержимое выливается наружу. Если его удалить, то можно увидеть «тени» — цитоплазматическую мембрану, которая и удерживает цитоплазму после удаления клеточной стенки.
Цитоплазматическая мембрана имеет толщину около 50 ангстрем и составляет, так же как и стенка, около 20% веса клетки. Химический анализ изолированных и отмытых «теней» показал, что они состоят из липопротеинов.
Цитоплазматическая мембрана играет очень важную роль в жизни бактериальной клетки; она полупроницаема, то есть через нее проходят одни вещества и задерживаются другие. Именно она регулирует обмен веществ между бактериальной клеткой и внешней средой. Этот процесс заключается в поступлении в клетку из среды питательных веществ и выходе из клетки продуктов обмена.
Еще сравнительно недавно казалось, что проникновение веществ в бактерии зависит лишь от величины пор в мембране, растворимости проходящего вещества и электрических зарядов мембраны и вещества. Сейчас в цитоплазматической мембране установлено существование специальных ферментов, обеспечивающих перенос различных веществ из среды в клетку. Эти ферменты получили название пермеаз.
Таким образом, цитоплазматическая мембрана — сложный химический и физико-химический барьер клеточной проницаемости.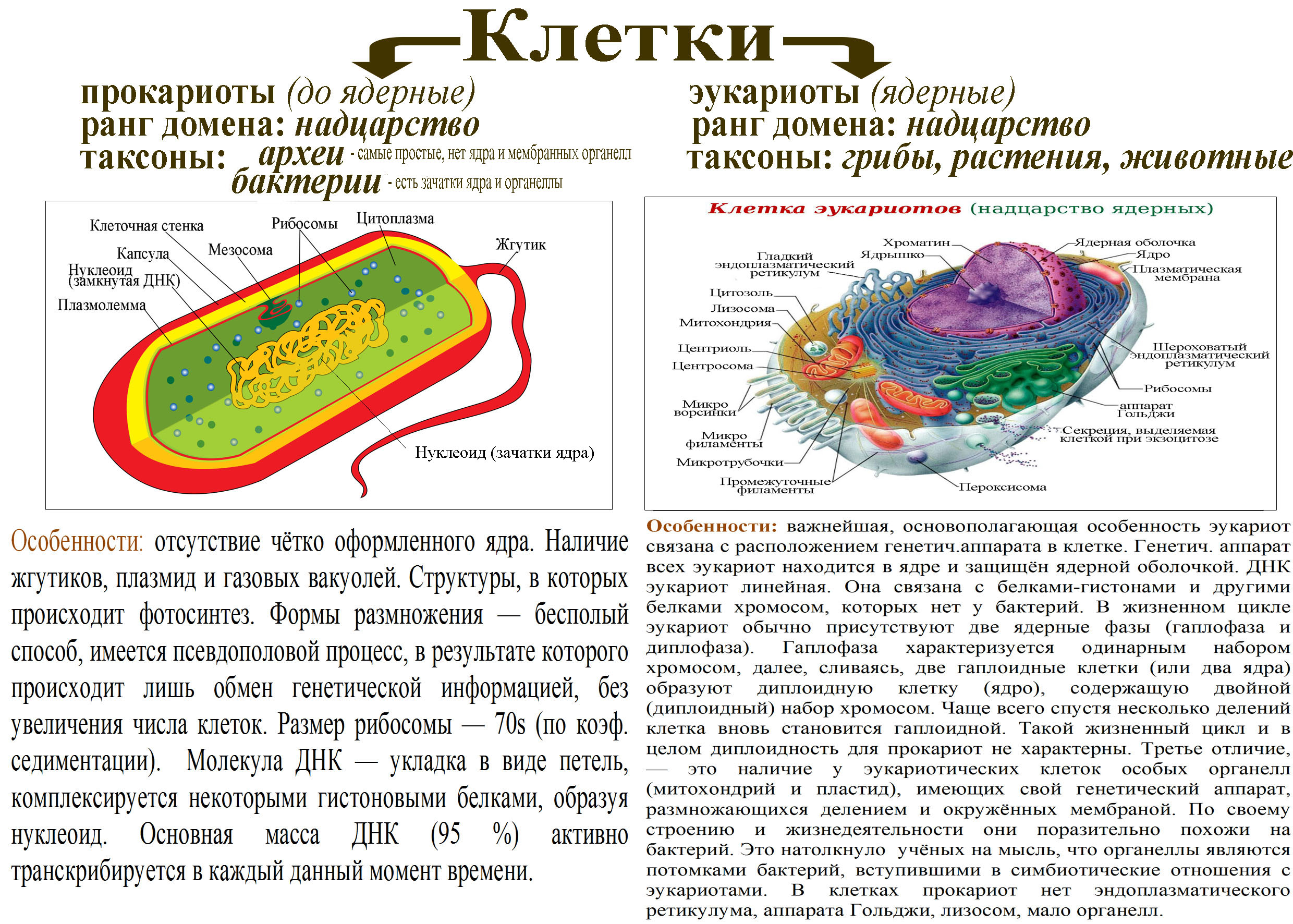 Изучение ее структуры и функции очень важно для понимания механизма действия лекарственных веществ на клетку и для расшифровки механизмов обмена самой бактериальной клетки.
Изучение ее структуры и функции очень важно для понимания механизма действия лекарственных веществ на клетку и для расшифровки механизмов обмена самой бактериальной клетки.
Если мы снимем с бактериальной клетки ее стенку и поместим образовавшийся протопласт в среду с низкой концентрацией солей, то в силу физико-химических законов вода начнет проникать внутрь протопласта. В результате резкого увеличения внутреннего давления цитоплазматическая мембрана разорвется, и содержимое протопласта выйдет наружу. Этот способ и был использован для выделения внутреннего содержимого — цитоплазмы бактерий, изучения ее свойств и составных частей.
Было найдено, что цитоплазма представляет собой прозрачную сиропообразную жидкость, вязкость которой в 8 тысяч раз превышает вязкость воды.
Уже давно в оптическом микроскопе при различных методах окраски в цитоплазме бактерий наблюдали довольно крупные гранулы. Неоднородность цитоплазмы была подтверждена при исследовании бактерий в фазовоконтрастном и люминесцентном микроскопах. Наиболее обстоятельно состав цитоплазмы был изучен методом дифференциального центрифугирования и в ультратонких срезах под электронным микроскопом.
Наиболее обстоятельно состав цитоплазмы был изучен методом дифференциального центрифугирования и в ультратонких срезах под электронным микроскопом.
Основная масса цитоплазмы состоит из очень мелких зерен диаметром 100—200 ангстрем — рибосом, где происходит синтез различных белков. Часть этих белков микроб использует как строительный материал для создания капсулы, стенки, цитоплазматической мембраны, самой цитоплазмы. Другая их часть — ферменты, которые являются катализаторами различных химических реакций.
Бактериальная клетка содержит огромное количество ферментов. Они либо прочно связаны с клеточной структурой, либо находятся в свободном состоянии. Некоторые из этих свободных ферментов выделяются из клетки наружу и разлагают сложные органические соединения на более простые вещества. Эти вещества затем могут поступать внутрь клетки и только в таком виде использоваться как питательные. Способность бактерий выделять наружу ферменты используется для промышленного получения этих ценных химических соединений.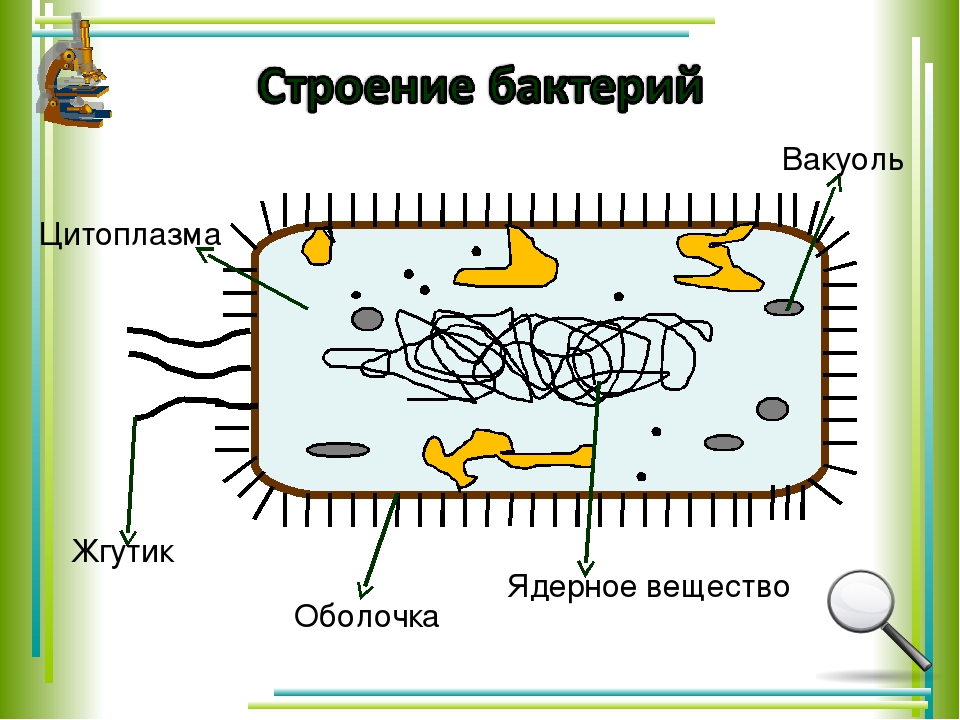
У некоторых болезнетворных бактерий ферменты служат мощным оружием в борьбе с организмом человека. 30 лет назад из шарообразных бактерий — стрептококков (возбудителей ангины, сепсиса, гнойничковых поражений кожи) был выделен фермент, названный гиалуронидазой. Этот фермент разрушает гиалуроновую кислоту, входящую в состав живой ткани. Благодаря его действию бактерии могут быстро продвигаться в глубь ткани, захватывая все новые участки организма. За это свойство фермент гиалуронидаза получил название «фактор распространения».
Позднее ученые выделили гиалуронидазу из пневмококков, стафилококков, палочки дифтерии и газовой гангрены.
Различные бактерии отличаются друг от друга внутренним содержимым цитоплазмы. Так, у некоторых из них уже при обработке анилиновыми красителями в цитоплазме выявлялись крупные, резко очерченные глыбки диаметром около 0,6 микрона. Это зерна волютина — вещества, чрезвычайно богатого полифосфатами. В зернах волютина происходят интенсивные окислительные процессы.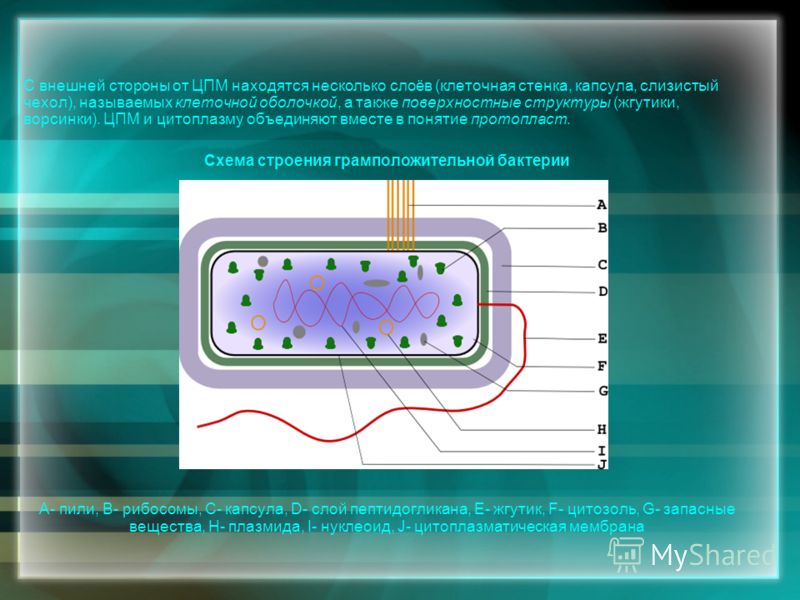 В палочке дифтерии зерна волютина настолько велики и располагаются так характерно (на обоих концах палочки), что по ним определяют эту бактерию под микроскопом.
В палочке дифтерии зерна волютина настолько велики и располагаются так характерно (на обоих концах палочки), что по ним определяют эту бактерию под микроскопом.
В цитоплазме бактериальных клеток могут накапливаться различные питательные «вещества, которые в неблагоприятных условиях будут постепенно расходоваться. Эти своеобразные кладовые продуктов, или, как их называют, вакуоли, содержат жиры, зерна гликогена, крахмала, капли серы и др.
Некоторые бактерии содержат в своей цитоплазме разнообразные красящие вещества — пигменты.
Однажды на прибрежных скалах Гренландии моряки с изумлением обнаружили… красный снег. Советские ученые нашли на Новой Земле… зеленый снег! И в том и в другом случае в этой неожиданной раскраске снега были повинны бактерии: пигмент служил им защитой от действия солнечных лучей.
Энергия, необходимая для поддержания жизнедеятельности бактерий, образуется «в клетке в результате дыхания. По типу дыхания все бактерии делятся на две основные группы: 1) аэробные — развивающиеся только в присутствии кислорода и 2) анаэробные — развивающиеся только в бескислородной среде. Дыхание представителей той и другой группы осуществляется в виде сложных окислительно-восстановительных реакций. У первой группы в них принимает участие атмосферный кислород, у второй роль кислорода выполняют различные органические вещества. Процесс дыхания сопровождается выделением тепла. Все вы знаете, что влажное сено, навоз, торф и т. д. «самонагреваются». Виновники этого — бактерии.
Дыхание представителей той и другой группы осуществляется в виде сложных окислительно-восстановительных реакций. У первой группы в них принимает участие атмосферный кислород, у второй роль кислорода выполняют различные органические вещества. Процесс дыхания сопровождается выделением тепла. Все вы знаете, что влажное сено, навоз, торф и т. д. «самонагреваются». Виновники этого — бактерии.
У некоторых бактерий окислительные процессы в клетке сопровождаются выделением световой энергии. Свечение гнилого дерева, пищевых продуктов (мясо, рыба), морской воды — все это результат интенсивного окисления особых фотогенных веществ в теле бактериальной клетки. Более 60 лет назад светящихся бактерий предложили использовать как «безопасные лампы» в пороховых погребах и везде, где можно опасаться взрыва. А свечение моря рыбаки Северного Приморья используют, определяя местонахождение косяков рыбы.
У многих видов бактерий при неблагоприятных условиях жизни — обеднение среды питательными веществами, нарушении режима влажности, старении клеток — внутри цитоплазмы возникает участок уплотнения, вокруг которого, тоже из цитоплазмы, образуется двуслойная оболочка, плохо проницаемая для воды и других веществ. Этот процесс называют спорообразованием. Спора служит бактерии для сохранения вида. Споры очень устойчивы: температура 100° их не убивает. Для этого необходим пар при температуре 120°. В таких условиях их гибель наступает только через 20 минут. Споры могут сохраняться многие годы, и только попав в благоприятные условия дают начало вегетативной форме бактерии (прорастают).
Этот процесс называют спорообразованием. Спора служит бактерии для сохранения вида. Споры очень устойчивы: температура 100° их не убивает. Для этого необходим пар при температуре 120°. В таких условиях их гибель наступает только через 20 минут. Споры могут сохраняться многие годы, и только попав в благоприятные условия дают начало вегетативной форме бактерии (прорастают).
Процесс прорастания длится 4—5 часов. Он начинается с набухания споры и усиления биохимической активности внутри нее. Затем через отверстие в оболочке появляется проросток, который вытягивается в палочку. Оболочка споры какое-то время сохраняется на конце палочки, а затем отделяется от нее. Спорообразование имеет важное практическое значение и всегда учитывается при стерилизации, например, перевязочного материала в больницах: стерилизация проводится в автоклавах паром под давлением — в режиме, при котором бактерии и их споры безусловно погибают.
Уже в момент открытия бактерий было замечено, что многие из них подвижны.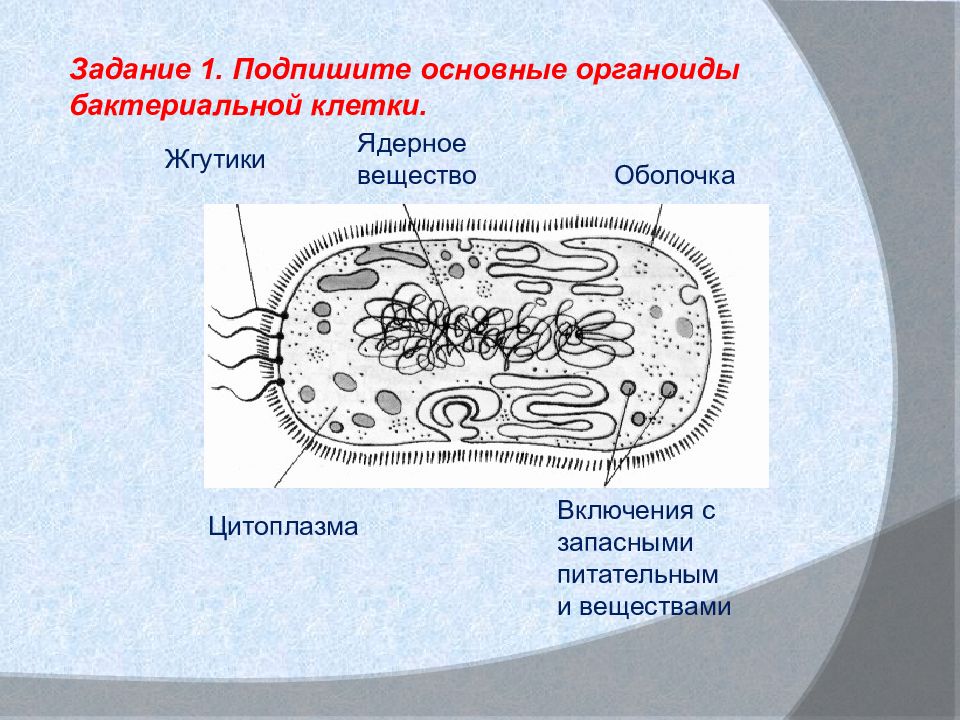 Около 100 лет назад обнаружили, что у некоторых бактерий есть жгутики — очень тонкие нити, отходящие от тела бактериальной клетки. Однако изучение их в оптическом микроскопе оказалось весьма затруднительным, так как жгутики были настолько тонкими, что различались лишь после предварительной специальной обработки. Нечего было и думать об их изучении. Ценные сведения о строении жгутиков удалось получить только с помощью электронного микроскопа.
Около 100 лет назад обнаружили, что у некоторых бактерий есть жгутики — очень тонкие нити, отходящие от тела бактериальной клетки. Однако изучение их в оптическом микроскопе оказалось весьма затруднительным, так как жгутики были настолько тонкими, что различались лишь после предварительной специальной обработки. Нечего было и думать об их изучении. Ценные сведения о строении жгутиков удалось получить только с помощью электронного микроскопа.
Оказалось, что различные бактерии имеют неодинаковое количество жгутиков, расположенных в разных частях клеточной поверхности. Встречаются клетки, у которых от одного из концов отходит от 1 до 100 жгутиков («косы»). У некоторых бактерий жгутики расположены по всей поверхности.
Жгутики имеют вид цилиндрической спирали и составляют часть живой клетки, хотя, если их удалить, это не ведет к гибели бактерии. Длина жгутиков достигает 75 микрон и зависит от возраста бактерии и условий ее существования. При более детальном исследовании жгутиков было обнаружено, что каждый жгутик состоит из 3—4 (или более) отдельных нитей, свернутых спиралеобразно друг с другом.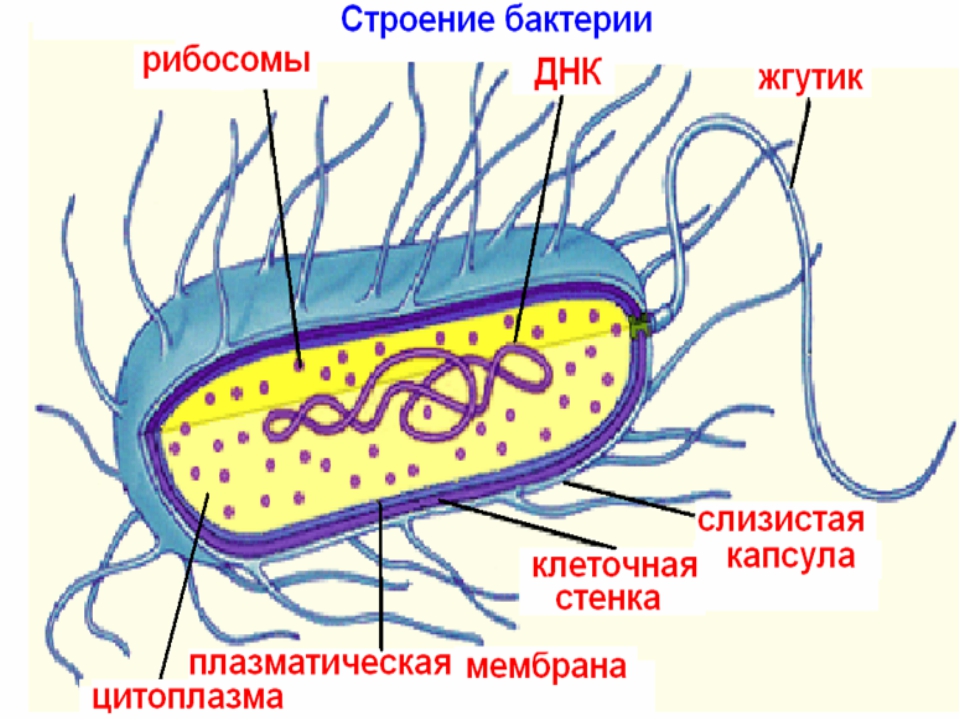 Средняя толщина жгутика составляет 15 миллимикрон.
Средняя толщина жгутика составляет 15 миллимикрон.
Жгутик начинается в теле клетки от блефаропласта — зернышка, расположенного в цитоплазме недалеко от клеточной стенки. Блефаропласт регулирует активность жгутика, который, проходя сквозь клеточную стенку, прикрепляется также и к ней. Если стенку удалить, то жгутик останется, но потеряет способность к сокращению.
Движение бактерии обусловлено спиралеобразными движениями жгутика. Скорость движения безжгутиковых бактерий около 5 микрон в секунду, а жгутиковых — более 50 микрон в секунду. Это огромная скорость: ведь бактерия длиной в 3 микрона за 1 секунду покрывает расстояние, в 16 раз превышающее длину ее тела! За это же время человек (в лучшем случае) способен преодолеть путь, равный не более пятикратной длине своего тела. Направление, движение и повороты зависят от угла, который образуется между телом бактерии и ее жгутиком, располагающимся чаще всего сзади бактерии. Иногда, впрочем, он находится впереди и действует наподобие пропеллера. Перемещаясь под влиянием жгутика вперед, бактериальная клетка одновременно совершает воронкообразные движения.
Перемещаясь под влиянием жгутика вперед, бактериальная клетка одновременно совершает воронкообразные движения.
Подвижность бактерий имеет большое практическое значение и используется при диагностике инфекционных заболеваний. Так, различают очень сходные по форме и размерам палочки дизентерии и брюшного тифа: первые неподвижны, вторые весьма активны.
Схема строения бактериальной клетки
При исследовании бактериальных клеток в электронном микроскопе было обнаружено, что поверхность их тела покрыта короткими и чрезвычайно тонкими ворсинками. Эти ворсинки найдены у подвижных и у неподвижных бактерий. Следовательно, они не имеют отношения к подвижности бактерий. Позднее было показано, что ворсинки — это органы прикрепления бактериальной клетки к поверхности тканей человека, животных и растений.
Долгое время вопрос о существовании ядра у бактерий был предметом горячих споров. И это понятно. Именно в ядре происходят такие жизненно важные процессы, от которых зависят рост и развитие клетки, в ядре начинается процесс деления, и, наконец, в ядре каждой живой клетки — животной или растительной — заложен материал, в котором зашифрованы все наследственные признаки.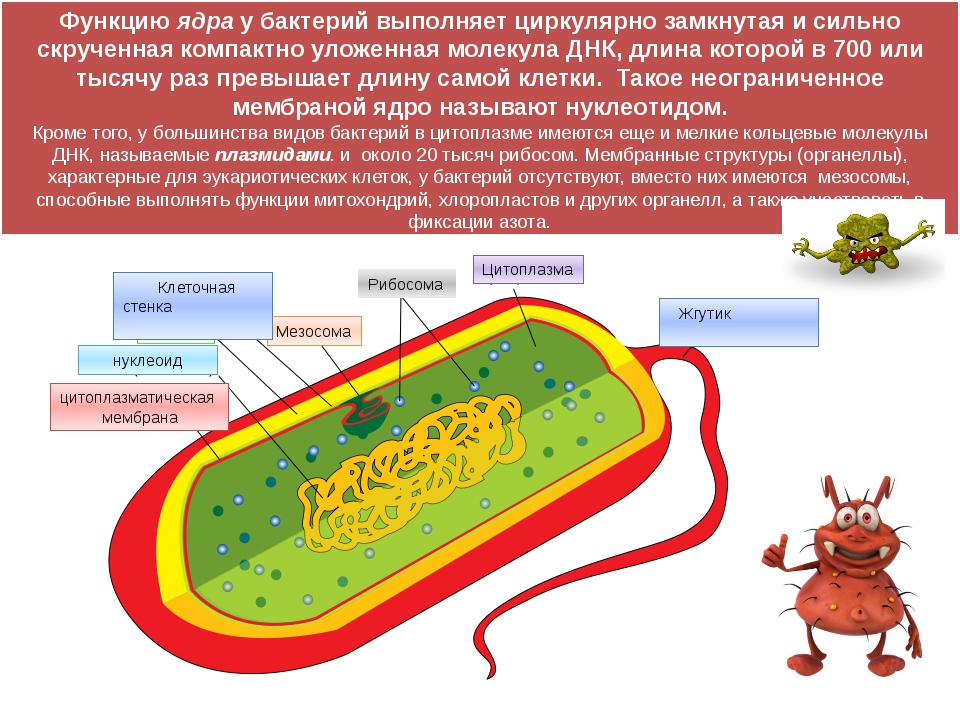
Высказывалось предположение, что бактерии вовсе «не имеют ядер, так как четко очерченного ядра, какое мы ясно видим в клетках животных или растений, в бактериальных клетках, при их изучении в оптическом микроскопе, найти не удавалось. Отсутствие внутри бактерии ясно выраженного ядра позволило высказать и прямо противоположную точку зрения — сама бактериальная клетка представляет собой голое ядро, лишенное цитоплазмы. В дальнейшем разработка и применение новых методов исследования (специальная окраска, изготовление ультратонких срезов и др.) позволили накопить факты, решившие эту проблему.
Ядра всех животных и растительных клеток содержат особое вещество — хроматин, который состоит из дезоксирибонуклеиновой кислоты (ДНК). В состав ДНК входит, в частности, дезоксирибоза — сахар, который есть только в составе ДНК. Следовательно, если удастся найти в каких-нибудь структурах дезоксирибозу, значит, в них находится ДНК — характерное ядерное вещество. Так и было сделано. При окраске бактерий на дезоксирибозу (реакция Фельгена) было обнаружено характерное фиолетовое окрашивание нескольких участков цитоплазмы.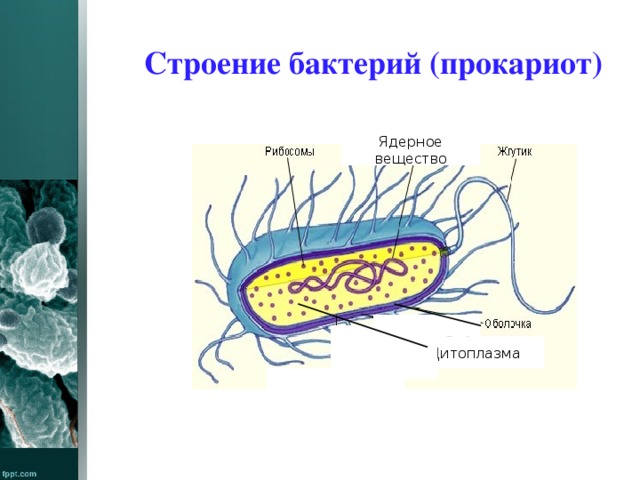 Был сделан вывод о присутствии внутри бактериальной клетки ядерного вещества.
Был сделан вывод о присутствии внутри бактериальной клетки ядерного вещества.
Позднее было показано, что эти прокрашивающиеся, по Фельгену, участки начинают делиться раньше остальных частей бактерий. Это тоже характерно для ядерных структур. С помощью гонких химических исследований удалось определить, что вся ДНК бактерий находится в ядре, так как ДНК изолированных ядер оказалась равной ДНК, выделенной из целой клетки. Наконец, были получены ультратонкие срезы бактериальных клеток, и на них достаточно ясно было видно ядро.
Выявить бактериальное ядро было трудно. Дело в том, что ядро бактерии действительно отличается от ядер животных и растительных клеток. Оно не имеет оболочки и поэтому легко может менять свою форму даже на различных этапах жизни одной и той же клетки.
Отличие ядерного аппарата бактерии проявилось и в другом. В ядре каждой животной или растительной клетки из хроматина образованы особые тяжи — хромосомы. В каждой клетке любого организма содержится строго определенное количество хромосом.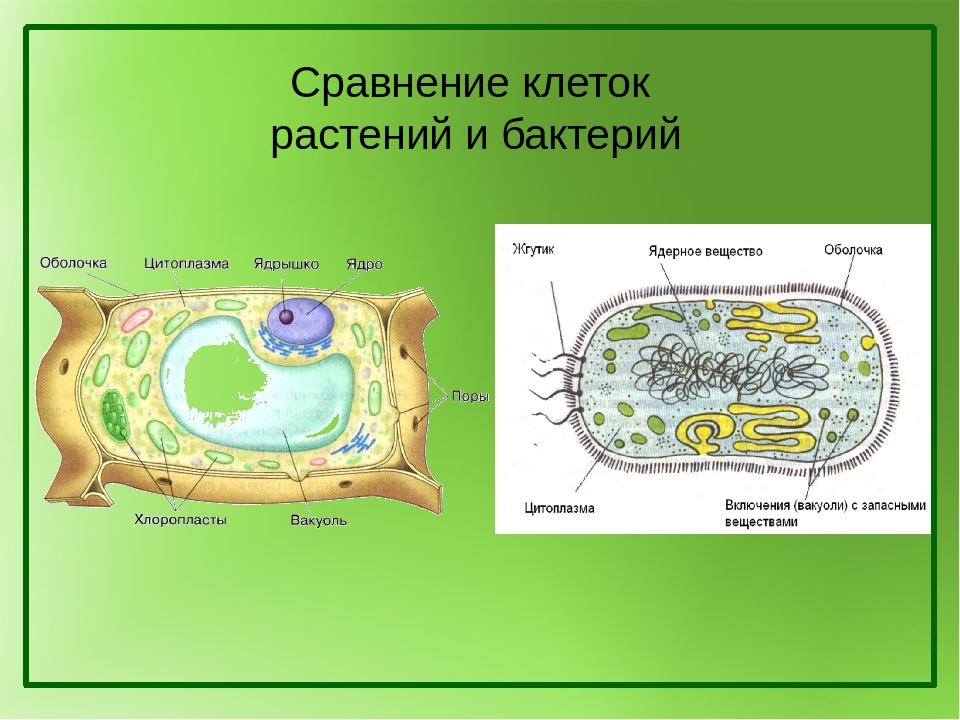 Так, у человека их 46. Следовательно, каждая клетка каждого человека содержит по 46 хромосом. Разные виды животных различаются по числу хромосом.
Так, у человека их 46. Следовательно, каждая клетка каждого человека содержит по 46 хромосом. Разные виды животных различаются по числу хромосом.
У одного вида круглых червей в каждой клетке насчитывается лишь по две хромосомы, в то время как у радиолярии — микроскопического морского животного — их около 1600. Хромосомы всегда парны, то есть в клетке всегда имеется по две хромосомы каждого типа. Таким образом, у человека их 23 пары. Каждая пара отличается от других пар по длине, форме и наличию утолщений или перетяжек.
В отличие от этого в ядре бактерии содержится только одна хромосома. Согласно новейшим представлениям, ДНК бактериальной хромосомы имеет вид замкнутой зигзагообразной нити. Между отдельными отрезками ДНК располагаются белковые молекулы. Такое строение придает всей структуре компактность и вместе с тем подвижность.
Теперь мы с вами узнали, что бактериальная клетка представляет собой сложный живой организм. Оказывается, его микроскопически малые размеры сочетаются с огромной скоростью размножения.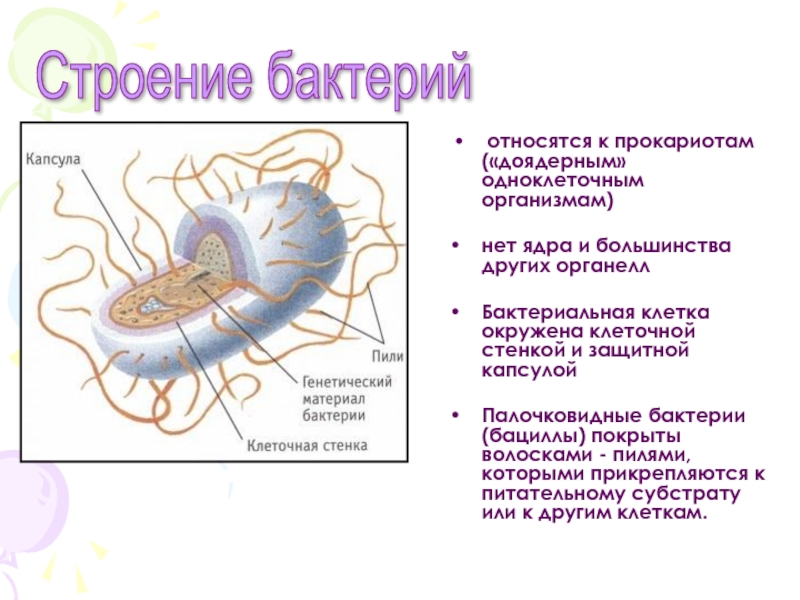 Бактериальная клетка делится каждые 30 минут. За 24 часа у бактерий сменяется столько же поколений, сколько у человека за 5 тысяч лет!
Бактериальная клетка делится каждые 30 минут. За 24 часа у бактерий сменяется столько же поколений, сколько у человека за 5 тысяч лет!
Размножаются бактерии делением. Достигнув определенной величины, клетка начинает изменяться. Прежде всего, внутри нее увеличивается, а затем и разделяется ядерное вещество. Затем посередине бактерии в цитоплазматической мембране образуется утолщение, которое увеличивается и постепенно превращается в перегородку, делящую бактерию на две половины. Образование перегородки сопровождается появлением перетяжки в клеточной стенке как раз напротив места образования перегородки. Таким образом, одна клетка разделилась на две дочерних бактериальных клетки.
В последние годы было обнаружено, что бактерии способны передавать генетический материал от одной клетки к другой, что напоминает процесс, сходный с половым.
В 1947 году американские ученые Ледерберг и Татум поставили простой опыт. Они взяли два сорта бактерий (два штамма). У одного из них не хватало признаков А, Б и В, а признаки Г и Д были.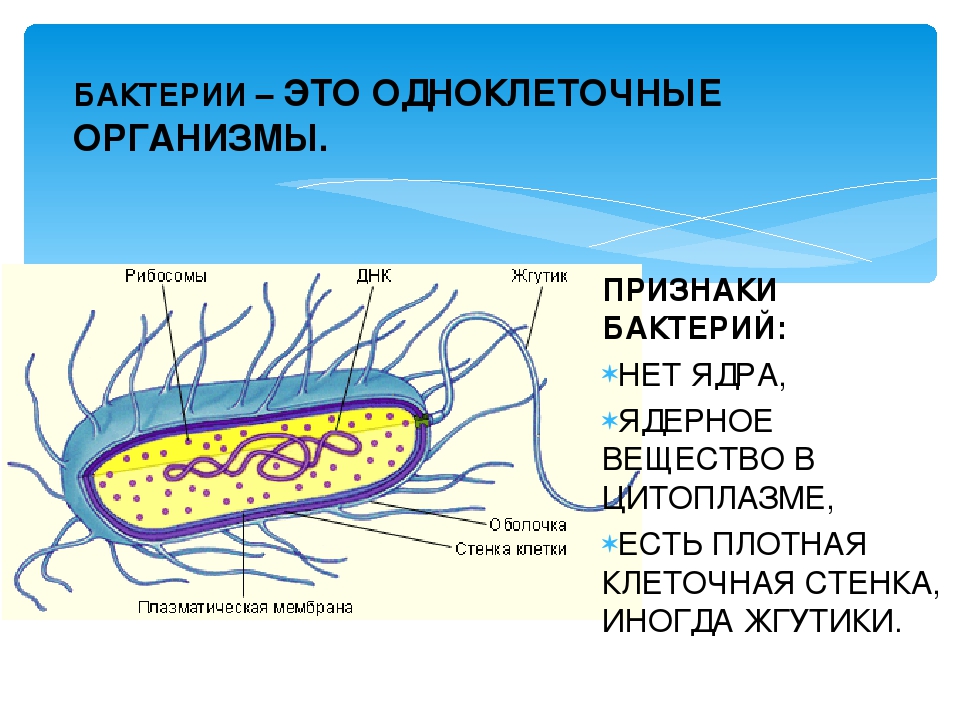 А у другого наоборот — были признаки А, Б и В, но не хватало признаков Г и Д. Из-за этих дефектов каждый из штаммов в отдельности не мог расти на питательной среде. Но стоило их смешать, как начинался рост. Тогда ученые убили один из штаммов и после этого смешали их. Роста не было. Значит, решили они, для роста клеток оба штамма должны быть живыми. Поставили еще опыт. Оба штамма поместили в один сосуд, но разделили тонким фильтром. Он пропускал жидкую среду, а бактерии задерживал. Роста опять не было. Следовательно, для роста необходим контакт.
А у другого наоборот — были признаки А, Б и В, но не хватало признаков Г и Д. Из-за этих дефектов каждый из штаммов в отдельности не мог расти на питательной среде. Но стоило их смешать, как начинался рост. Тогда ученые убили один из штаммов и после этого смешали их. Роста не было. Значит, решили они, для роста клеток оба штамма должны быть живыми. Поставили еще опыт. Оба штамма поместили в один сосуд, но разделили тонким фильтром. Он пропускал жидкую среду, а бактерии задерживал. Роста опять не было. Следовательно, для роста необходим контакт.
Ученые предположили, что оба дефектных штамма при смешивании обмениваются генами, которые отвечают за все имеющиеся признаки данного организма. Иначе говоря, при объединении А—+Б—+В—+Г++Д++А++Б++В++Г—+Д— у каких-нибудь бактерий окажется полный набор всех пяти генов, ответственных за признаки А, Б, В, Г и Д, и такие бактерии смогут расти на питательной среде.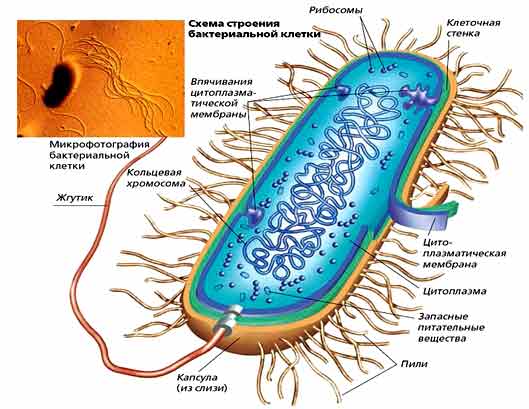
Позднее англичанин Хейс показал, что действительно обмен наследственным материалом происходит не случайно, а в строго определенном направлении. Получалось что-то похожее на половой процесс, так как передающие бактерии напоминали мужской тип, а воспринимающие — женский.
Несколько лет назад эти предположения были блестяще подтверждены французскими микробиологами Жакобом и Вольманом. Они увидели в электронном микроскопе, что «мужские» и «женские» клетки соединяются между собой цитоплазматическими мостиками. Если такие соединенные бактерии осторожным встряхиванием разъединять через 1, 10, 15, 20, 30 и т. д. минут после объединения, то можно обнаружить, что через различные интервалы времени от «мужских» к «женским» передаются различные признаки: последовательно друг за другом. Это было убедительным доказательством передачи наследственного материала (участков хромосом) от одной бактерии к другой.
После передачи материала хромосом от мужской клетки в женскую «оплодотворенная» бактерия делится, как описано выше, и передает по наследству новые признаки.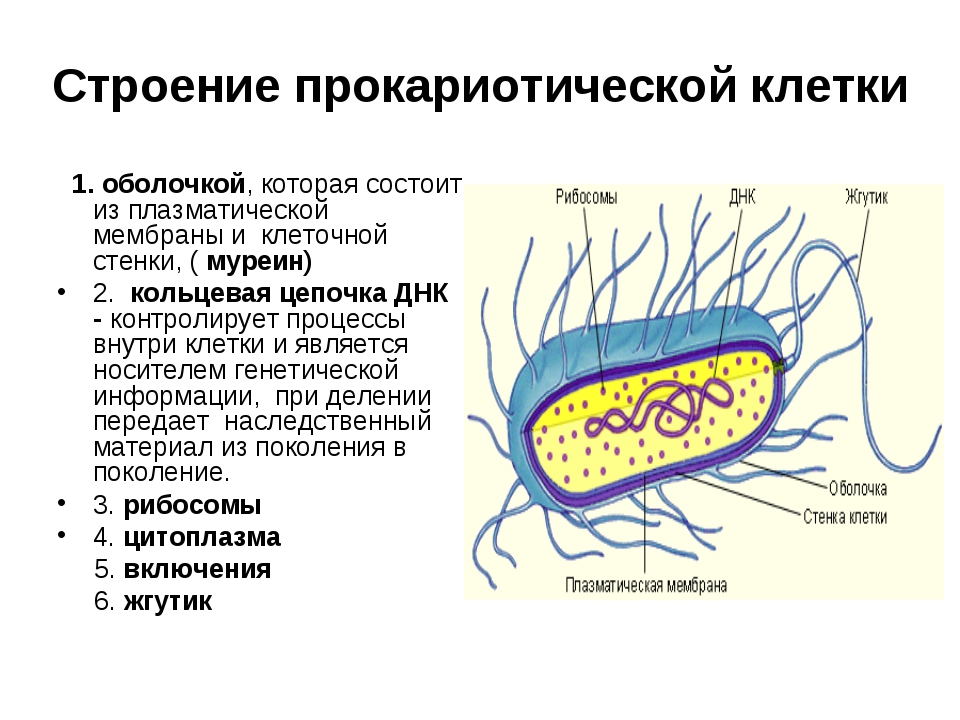 Открытие «полового» процесса у бактерий открывает перед учеными широкие горизонты новых теоретических исследований, которые позволят познать способы выведения новых полезных видов бактерий для промышленности и сельского хозяйства, позволят изменять свойства болезнетворных бактерий и др.
Открытие «полового» процесса у бактерий открывает перед учеными широкие горизонты новых теоретических исследований, которые позволят познать способы выведения новых полезных видов бактерий для промышленности и сельского хозяйства, позволят изменять свойства болезнетворных бактерий и др.
Чтобы представить себе, как велика скорость размножения бактерий, достаточно сказать, что через 5 дней потомство одной бактериальной клетки могло бы заполнить пространство, занимаемое всеми морями и океанами!
Почему же этого не происходит? Дело в том, что высокая скорость размножения бактерий сопровождается их быстрой гибелью. В этом можно легко убедиться, если проследить за развитием бактерий в мясном бульоне.
Регулярно определяя количество бактерий, мы увидим, что в первые часы после внесения бактерий в среду их количество не будет увеличиваться. Это объясняется тем, что вначале бактериальные клетки приспосабливаются к новым условиям существования и число вновь образующихся клеток будет равно числу отмирающих. Затем наступит фаза бурного размножения, когда за одно и то же время будет делиться гораздо больше клеток, чем отмирать. Поэтому общее количество их будет увеличиваться.
Затем наступит фаза бурного размножения, когда за одно и то же время будет делиться гораздо больше клеток, чем отмирать. Поэтому общее количество их будет увеличиваться.
Но постепенное расходование питательных веществ и накопление вредных продуктов обмена начнут замедлять скорость клеточного деления. Скорость деления сравняется со скоростью отмирания (опять прекратится увеличение общего количества бактерий), а затем снизится еще более. Отмирание клеток начнет преобладать — абсолютное количество бактерий будет уменьшаться.
Так происходит не только в пробирке на столах ученых, но и в естественных условиях.
В организме больного бактерии, приспособившись к новым условиям, начинают бурно размножаться. Но вскоре, под воздействием повышенной температуры тела больного, клеток крови, накапливающихся антител «и других защитных факторов начинает отмирать все больше бактерий.
Если они попадают во внешнюю среду, то здесь их гибель ускоряют солнечные лучи, высыхание, отсутствие питательных веществ и др.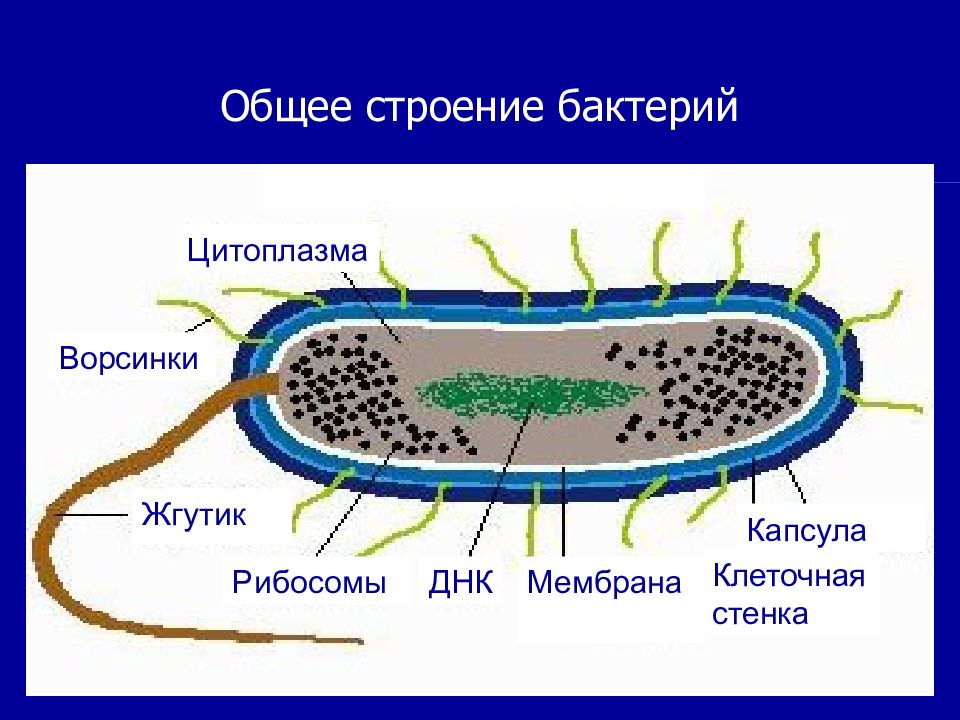 Диалектическое единство двух противоположных процессов — рождения новых бактериальных клеток и отмирания старых — ограничивает распространение бактерий, имеющих столь бурный темп роста.
Диалектическое единство двух противоположных процессов — рождения новых бактериальных клеток и отмирания старых — ограничивает распространение бактерий, имеющих столь бурный темп роста.
Изучение законов роста и размножения бактерий имеет огромное практическое значение для борьбы с инфекционными болезнями.
Из бактериальных клеток готовят различные вакцины — препараты, содержащие убитые или ослабленные болезнетворные бактерии. Такие бактерии при (введении в организм уже не могут вызвать болезнь, но они способны вызывать образование антител. Бактерии, попавшие в организм, при последующем заражении встретятся со специфическими антителами и будут уничтожены. Человек не заболевает, у него образовался иммунитет — невосприимчивость к инфекционному заболеванию.
Современные представления об организации бактерии, построенные на основе разносторонних исследований методами цитохимии, энзимологии, химического фракционирования и электроноскопического анализа ультратонких срезов, позволяют утверждать, что бактерия обладает структурами, характерными для клеточной организации.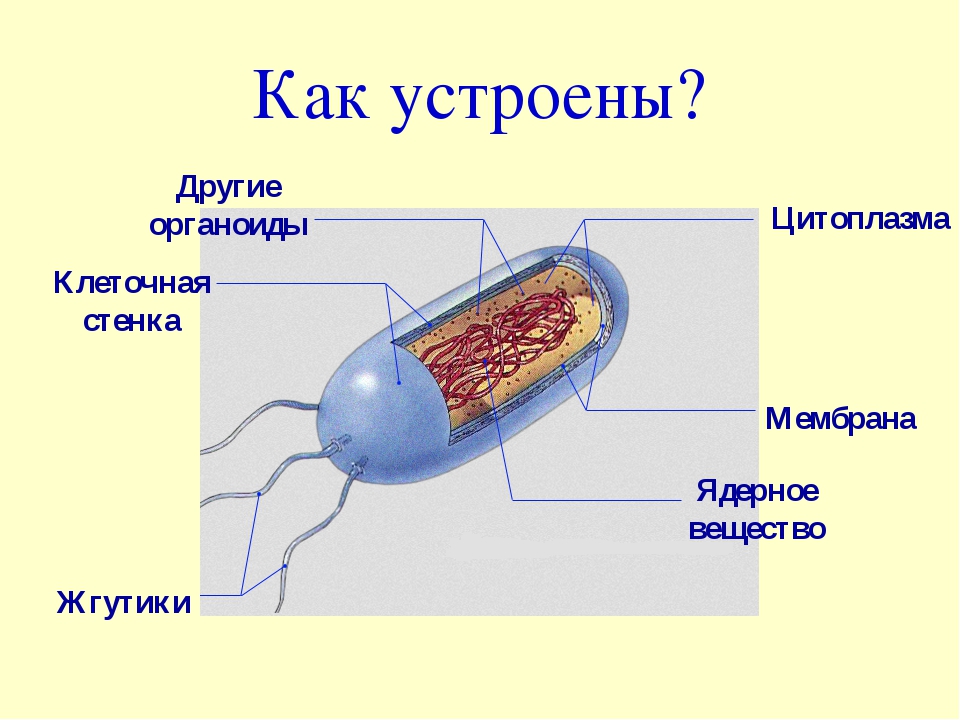 Следовательно, бактериальная клетка принципиально не отличается от клеток животного и растительного происхождения. Этот важный вывод позволяет использовать бактерии в качестве модели для изучения самых общих вопросов биологии и медицины.
Следовательно, бактериальная клетка принципиально не отличается от клеток животного и растительного происхождения. Этот важный вывод позволяет использовать бактерии в качестве модели для изучения самых общих вопросов биологии и медицины.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Бактериальная клетка. Строение и химический состав : Farmf
Бактериальная клетка. Строение и химический состав
Бактериальная клетка имеет сложную внутреннюю организацию. Каждая систематическая группа микроорганизмов имеет свои специфические особенности строения.
Бактериальная клетка. Клеточная стенка.
Клетка бактерий одета плотной оболочкой. Этот поверхностный слой, расположенный снаружи от цитоплазматической мембраны, называют клеточной стенкой. Стенка выполняет защитную и опорную функции, а также придает клетке постоянную, характерную для нее форму (например, форму палочки или кокка) и представляет собой наружный скелет клетки.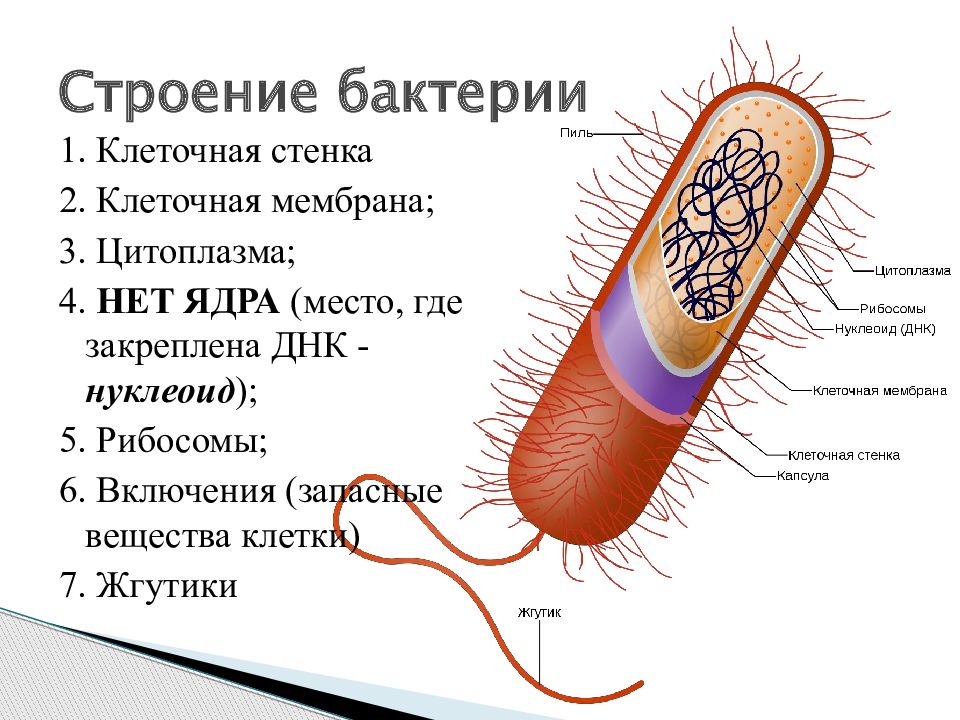
Эта плотная оболочка роднит бактерии с растительными клетками, что отличает их от животных клеток, имеющих мягкие оболочки. Внутри бактериальной клетки осмотическое давление в несколько раз, а иногда и в десятки раз выше, чем во внешней среде. Поэтому клетка быстро разорвалась бы, если бы она не была защищена такой плотной, жесткой структурой, как клеточная стенка.
Толщина клеточной стенки 0,01—0,04 мкм. Она составляет от 10 до 50% сухой массы бактерий. Количество материала, из которого построена клеточная стенка, изменяется в течение роста бактерий и обычно увеличивается с возрастом.
Основным структурным компонентом стенок, основой их жесткой структуры почти у всех исследованных до настоящего времени бактерий является муреин (гликопептид, мукопептид). Это органическое соединение сложного строения, в состав которого входят сахара, несущие азот, аминосахара и 4—5 аминокислот. Причем аминокислоты клеточных стенок имеют необычную форму (D-стереоизомеры), которая в природе редко встречается.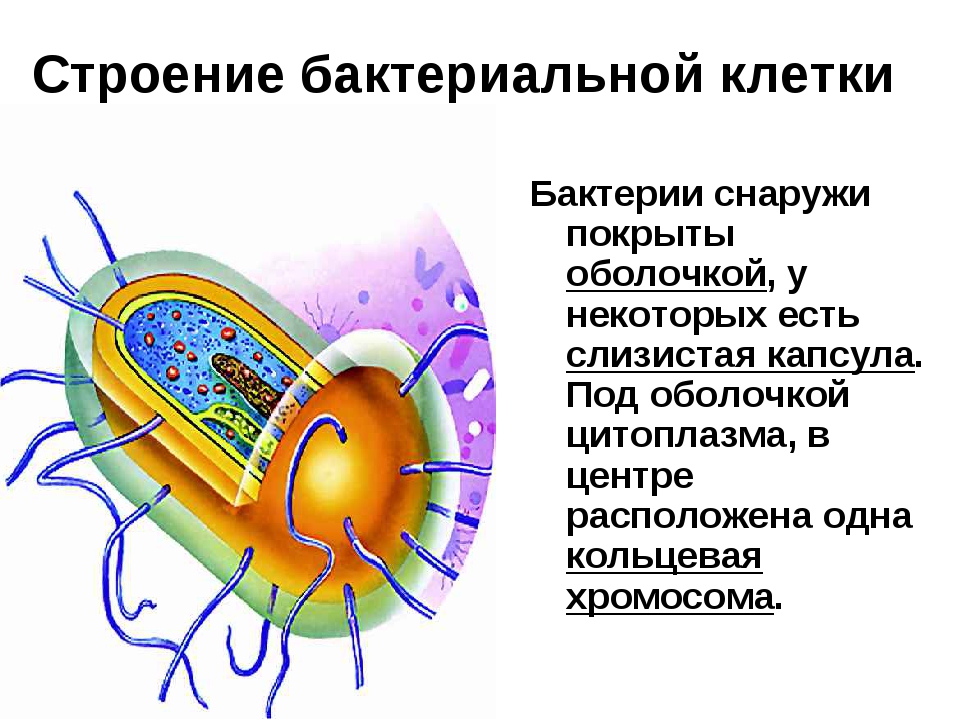
Составные части клеточной стенки, ее компоненты, образуют сложную прочную структуру.
С помощью способа окраски, впервые предложенного в 1884 г. Кристианом Грамом, бактерии могут быть разделены на две группы: грамположительные и грамотрицательные. Грамположительные организмы способны связывать некоторые анилиновые красители, такие, как кристаллический фиолетовый, и после обработки иодом, а затем спиртом (или ацетоном) сохранять комплекс иод-краситель. Те же бактерии, у которых под влиянием этилового спирта этот комплекс разрушается (клетки обесцвечиваются), относятся к грамотрицательным.
Химический состав клеточных стенок грамположительных и грамотрицательных бактерий различен.
У грамположительных бактерий в состав клеточных стенок входят, кроме мукопептидов, полисахариды (сложные, высокомолекулярные сахара), тейхоевые кислоты (сложные по составу и структуре соединения, состоящие из сахаров, спиртов, аминокислот и фосфорной кислоты). Полисахариды и тейхоевые кислоты связаны с каркасом стенок — муреином.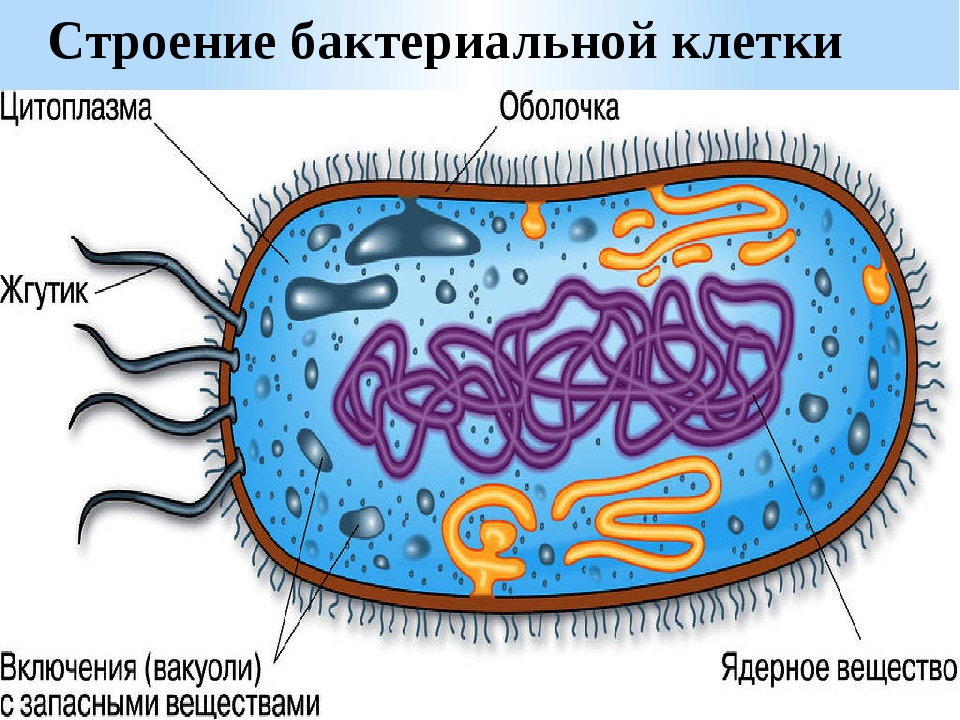 Какую структуру образуют эти составные части клеточной стенки грамположительных бактерий, мы пока еще не знаем. С помощью электронных фотографий тонких срезов (слоистости) в стенках грамположительных бактерий не обнаружено. Вероятно, все эти вещества очень плотно связаны между собой.
Какую структуру образуют эти составные части клеточной стенки грамположительных бактерий, мы пока еще не знаем. С помощью электронных фотографий тонких срезов (слоистости) в стенках грамположительных бактерий не обнаружено. Вероятно, все эти вещества очень плотно связаны между собой.
Стенки грамотрицательных бактерий более сложные по химическому составу, в них содержится значительное количество липидов (жиров), связанных с белками и сахарами в сложные комплексы — липопротеиды и липополисахариды. Муреина в клеточных стенках грамотрицательных бактерий в целом меньше, чем у грамположительных бактерий. Структура стенки грамотрицательных бактерий также более сложная. С помощью электронного микроскопа было установлено, что стенки этих бактерий многослойные
Внутренний слой состоит из муреина. Над ним находится более широкий слой из неплотно упакованных молекул белка. Этот слой в свою очередь покрыт слоем липополисахарида. Самый верхний слой состоит из липопротеидов.
Клеточная стенка проницаема: через нее питательные вещества свободно проходят в клетку, а продукты обмена выходят в окружающую среду. Крупные молекулы с большим молекулярным весом не проходят через оболочку.
Крупные молекулы с большим молекулярным весом не проходят через оболочку.
Бактериальная клетка. Капсула.
Клеточная стенка многих бактерий сверху окружена слоем слизистого материала — капсулой. Толщина капсулы может во много раз превосходить диаметр самой клетки, а иногда она настолько тонкая, что ее можно увидеть лишь через электронный микроскоп, — микрокапсула.
Капсула не является обязательной частью клетки, она образуется в зависимости от условий, в которые попадают бактерии. Она служит защитным покровом клетки и участвует в водном обмене, предохраняя клетку от высыхания.
По химическому составу капсулы чаще всего представляют собой полисахариды. Иногда они состоят изгликопротеидов (сложные комплексы сахаров и белков) и полипептидов (род Bacillus), в редких случаях — из клетчатки (род Acetobacter).
Слизистые вещества, выделяемые в субстрат некоторыми бактериями, обусловливают, например, слизисто-тягучую консистенцию испорченного молока и пива.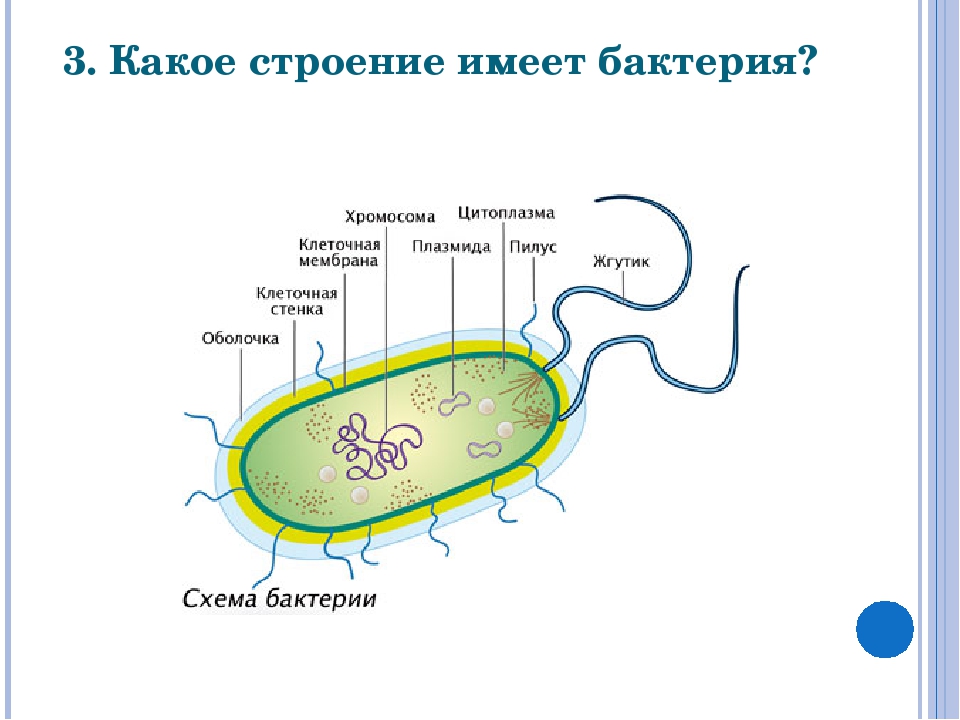
Бактериальная клетка. Цитоплазма.
Все содержимое клетки, за исключением ядра и клеточной стенки, называется цитоплазмой. В жидкой, бесструктурной фазе цитоплазмы (матриксе) находятся рибосомы, мембранные системы, митохондрии, пластиды и другие структуры, а также запасные питательные вещества. Цитоплазма обладает чрезвычайно сложной, тонкой структурой (слоистая, гранулярная). С помощью электронного микроскопа раскрыты многие интересные детали строения клетки.
Внешний липопротвидный слой протопласта бактерий, обладающий особыми физическими и химическими свойствами, называется цитоплазматической мембраной.
Внутри цитоплазмы находятся все жизненно важные структуры и органеллы.
Функции цитоплазматической мембраны:
- регулирует поступление веществ в клетку и выделение наружу продуктов обмена.
- питательные вещества могут поступать в клетку в результате активного биохимического процесса с участием ферментов.
- в мембране происходит синтез некоторых составных частей клетки, в основном компонентов клеточной стенки и капсулы.

- в цитоплазматической мембране находятся важнейшие ферменты (биологические катализаторы).
Упорядоченное расположение ферментов на мембранах позволяет регулировать их активность и предотвращать разрушение одних ферментов другими. С мембраной связаны рибосомы — структурные частицы, на которых синтезируется белок. Мембрана состоит из липопротеидов. Она достаточно прочна и может обеспечить временное существование клетки без оболочки. Цитоплазматическая мембрана составляет до 20% сухой массы клетки.
На электронных фотографиях тонких срезов бактерий цитоплазматическая мембрана представляется в виде непрерывного тяжа толщиной около 75A, состоящего из светлого слоя (липиды), заключенного между двумя более темными (белки). Каждый слой имеет ширину 20—30А. Такая мембрана называется элементарной.
Между плазматической мембраной и клеточной стенкой имеется связь в виде десмозов — мостиков. Цитоплазматическая мембрана часто дает инвагинации — впячивания внутрь клетки. Эти впячивания образуют в цитоплазме особые мембранные структуры, названные мезосомами. Некоторые виды мезосом представляют собой тельца, отделенные от цитоплазмы собственной мембраной.
Эти впячивания образуют в цитоплазме особые мембранные структуры, названные мезосомами. Некоторые виды мезосом представляют собой тельца, отделенные от цитоплазмы собственной мембраной.
Внутри таких мембранных мешочков упакованы многочисленные пузырьки и канальцы. Эти структуры выполняют у бактерий самые различные функции. Одни из этих структур — аналоги митохондрий. Другие выполняют функции зндоплазматической сети или аппарата Гольджи.
Путем инвагинации цитоплазматической мембраны образуется также фотосинтезирующий аппарат бактерий. После впячивания цитоплазмы мембрана продолжает расти и образует стопки, которые по аналогии с гранулами хлоропластов растений называют стопками тилакоидов. В этих мембранах, часто заполняющих собой большую часть цитоплазмы бактериальной клетки, локализуются пигменты (бактериохлорофилл, каротиноиды) и ферменты (цитохромы), осуществляющие процесс фотосинтеза.
В цитоплазме бактерий содержатся рибосомы— белок-синтезирующие частицы диаметром 200А.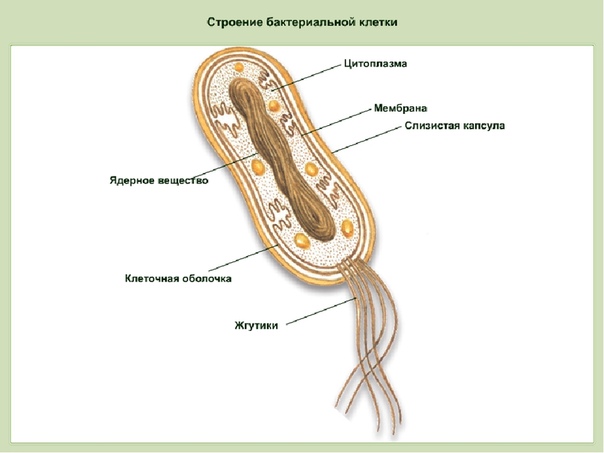 В клетке их насчитывается больше тысячи. Состоят рибосомы из РНК и белка. У бактерий многие рибосомы расположены в цитоплазме свободно, некоторые из них могут быть связаны с мембранами.
В клетке их насчитывается больше тысячи. Состоят рибосомы из РНК и белка. У бактерий многие рибосомы расположены в цитоплазме свободно, некоторые из них могут быть связаны с мембранами.
Рибосомы являются центрами синтеза белка в клетке. При этом они часто соединяются между собой, образуя агрегаты, называемые полирибосомами или полисомами.
В цитоплазме клеток бактерий часто содержатся гранулы различной формы и размеров. Однако их присутствие нельзя рассматривать как какой-то постоянный признак микроорганизма, обычно оно в значительной степени связано с физическими и химическими условиями среды. Многие цитоплазматические включения состоят из соединений, которые служат источником энергии и углерода. Эти запасные вещества образуются, когда организм снабжается достаточным количеством питательных веществ, и, наоборот, используются, когда организм попадает в условия, менее благоприятные в отношении питания.
У многих бактерий гранулы состоят из крахмала или других полисахаридов — гликогена и гранулезы.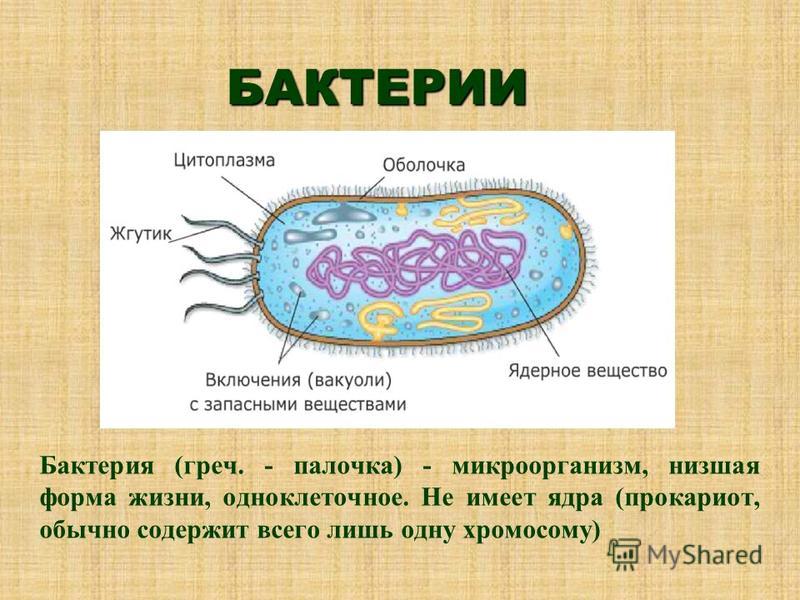 У некоторых бактерий при выращивании на богатой сахарами среде внутри клетки встречаются капельки жира. Другим широко распространенным типом гранулярных включений является волютин (метахроматиновые гранулы). Эти гранулы состоят из полиметафосфата (запасное вещество, включающее остатки фосфорной кислоты).
У некоторых бактерий при выращивании на богатой сахарами среде внутри клетки встречаются капельки жира. Другим широко распространенным типом гранулярных включений является волютин (метахроматиновые гранулы). Эти гранулы состоят из полиметафосфата (запасное вещество, включающее остатки фосфорной кислоты).
Полиметафосфат служит источником фосфатных групп и энергии для организма. Бактерии чаще накапливают волютин в необычных условиях питания, например на среде, не содержащей серы. В цитоплазме некоторых серных бактерий находятся капельки серы.
Помимо различных структурных компонентов, цитоплазма состоит из жидкой части — растворимой фракции. В ней содержатся белки, различные ферменты, т-РНК, некоторые пигменты и низкомолекулярные соединения — сахара, аминокислоты.
В результате наличия в цитоплазме низкомолекулярных соединений возникает разность в осмотическом давлении клеточного содержимого и наружной среды, причем у разных микроорганизмов это давление может быть различным.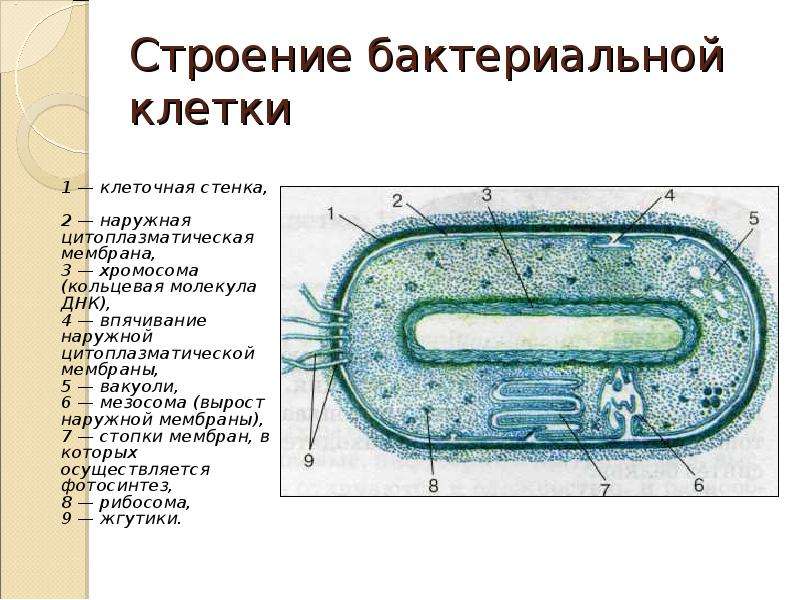 Наибольшее осмотическое давление отмечено у грамположительных бактерий — 30 атм, у грамотрицательных бактерий оно гораздо ниже — 4—8 атм.
Наибольшее осмотическое давление отмечено у грамположительных бактерий — 30 атм, у грамотрицательных бактерий оно гораздо ниже — 4—8 атм.
Бактериальная клетка. Ядерный аппарат.
В центральной части клетки локализовано ядерное вещество — дезоксирибонуклеиновая кислот а (ДНК).
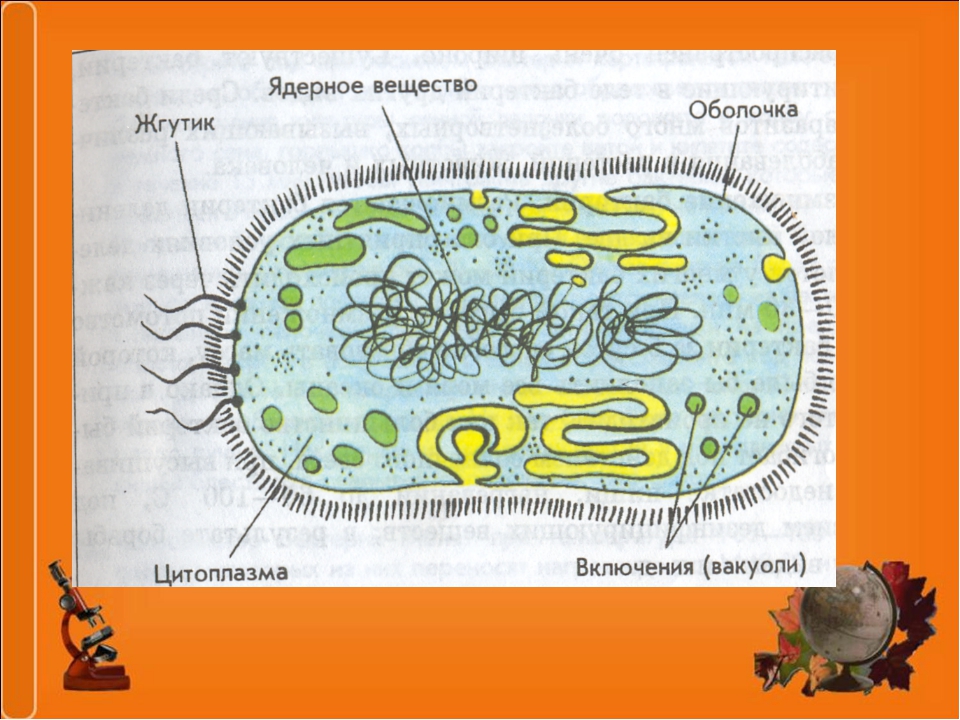
У бактерий нет такого ядра, как у высших организмов (эукариотов), а есть его аналог — «ядерный эквивалент» — нуклеоид (см. рис. 2, 8), который является эволюционно более примитивной формой организации ядерного вещества. Микроорганизмы, не имеющие настоящего ядра, а обладающие его аналогом, относятся к прокариотам.
Все бактерии — прокариоты. В клетках большинства бактерий основное количество ДНК сконцентрировано в одном или нескольких местах. В клетках эукариотов ДНК находится в определенной структуре — ядре. Ядро окружено оболочкой— мембраной.
У бактерий ДНК упакована менее плотно, в отличие от истинных ядер; нуклеоид не обладает мембраной, ядрышком и набором хромосом. Бактериальная ДНК не связана с основными белками — гистонами — ив нуклеоиде расположена в виде пучка фибрилл.
Бактериальная ДНК не связана с основными белками — гистонами — ив нуклеоиде расположена в виде пучка фибрилл.
Бактериальная клетка. Жгутики.
На поверхности некоторых бактерий имеются придаточные структуры; наиболее широко распространенными из них являются жгутики — органы движения бактерий.
Жгутик закрепляется под цитоплазматической мембраной с помощью двух пар дисков. У бактерий может быть один, два или много жгутиков. Расположение их различно: на одном конце клетки, на двух, по всей поверхности и т. д. Жгутики бактерий имеют диаметр 0,01—0,03 мкм, длина их может во много раз превосходить длину клетки. Бактериальные жгутики Состоят из белка — флагеллина — и представляют собой скрученные винтообразные нити.
На поверхности некоторых бактериальных клеток имеются тонкие ворсинки — фимбрии.
Бактериальная клетка. Структура бактериальной клетки
Бактериальная клетка – это целостный организм. Большинство бактерий одноклеточные организмы.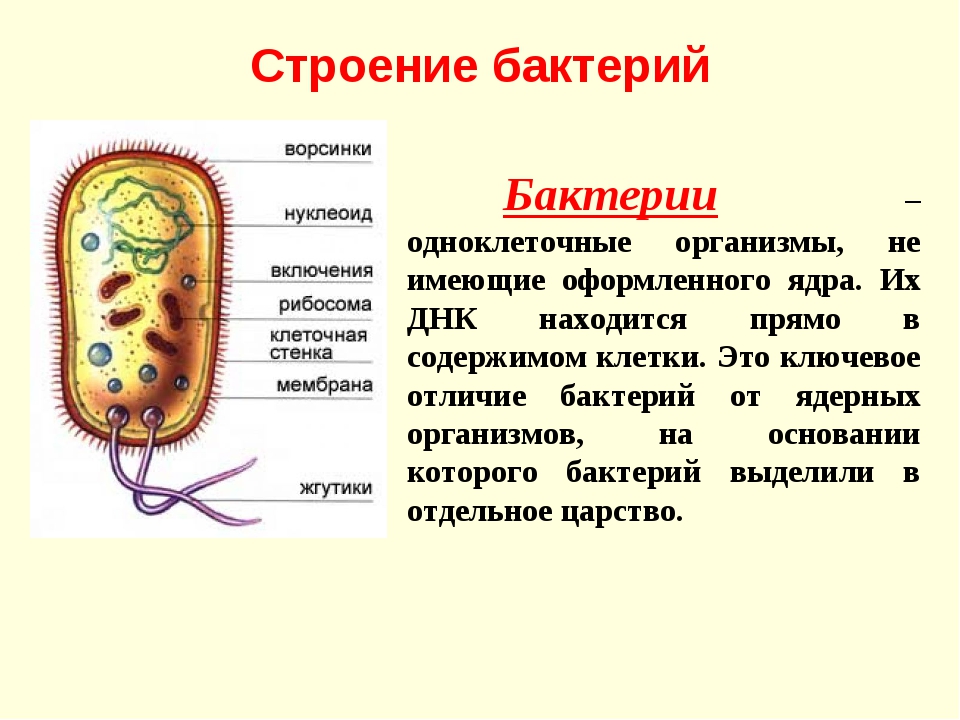 Их клетки относят к прокариотическим. Они не имеют оформленного ядра и мембранных органоидов. Большинство бактерий нуждаются в готовых органических веществах.
Их клетки относят к прокариотическим. Они не имеют оформленного ядра и мембранных органоидов. Большинство бактерий нуждаются в готовых органических веществах.
Бактериальная клетка. Структура.
Структура прокариотической клетки отличается от эукариотической отсутствием оформленного ядра (т.е. ядерной мембраны), отсутствием внутриклеточных мембран, ядрышек, комплекса Гольджи, лизосом, митохондрий. Основная структура бактериальной клетки: клеточная стенка, цитоплазматическая мембрана, цитоплазма с включениями и ядро (нуклеоид – наследственный материал, представлен 1 молекулой ДНК, замкнутой в кольцо и суперспирализованной, бактериальный нуклеоид не отделен от цитоплазмы мембраны). Отсутствуют: оформленное ядро, внутриклеточные мембраны, ядрышки, лизосомы, митохондрии, комплекс Гольджи.
Структура бактериальной клетки включает в себя:
- Нуклеоид – образование подобное ядру, расположен в центральной зоне бактерий, представляет собой наследственный (генетический) материал бактериальной клетки, представлен 1 молекулой ДНК, замкнутой в кольцо и суперспирализованной (скручена в рыхлый клубок).
 Длина ДНК около 1мм. Объем информации около 1000 генов (признаков). Нуклеоид не отделен от цитоплазмы мембраной. Нуклеоид не имеет ядерной оболочки, ядрышка, основных белков. Выявляется в световом микроскопе при окраске по Фельгену и Гимзе.
Длина ДНК около 1мм. Объем информации около 1000 генов (признаков). Нуклеоид не отделен от цитоплазмы мембраной. Нуклеоид не имеет ядерной оболочки, ядрышка, основных белков. Выявляется в световом микроскопе при окраске по Фельгену и Гимзе. - Цитоплазма – коллоид, т.е. водный раствор белков, липидов, минеральных веществ и углеводов. В цитоплазме находятся рибосомы, включения (полисахариды, поли-β-масляная кислота, полифосфаты-волютин), плазмиды. На рибосомах происходит синтез белка. Рибосомы прокариот отличаются от эукариотических более мелкими размерами (70 S).
- Цитоплазматическая мембрана – трехслойная структура, окружает наружную часть цитоплазмы бактерий. Состоит из двойного слоя липидов, больше фосфолипидов. ЦМ является динамической структурой, с подвижными компонентами, она участвует в регуляции осмотического давления, транспорте веществ, энергетическом метаболизме клетки.
- Включения – запасные питательные вещества бактериальной клетки, а также скопления пигментов.
 К запасным питательным веществам относятся: гранулы волютина (неорганического полифосфата), гликоген, гранулеза, крахмал, капли жира, скопления пигмента, серы, кальция. Включения, как правило, образуется при выращивании бактерий на богатых питательных средах и исчезает при голодании.
К запасным питательным веществам относятся: гранулы волютина (неорганического полифосфата), гликоген, гранулеза, крахмал, капли жира, скопления пигмента, серы, кальция. Включения, как правило, образуется при выращивании бактерий на богатых питательных средах и исчезает при голодании. - Плазмиды – небольшие кольцевые молекулы ДНК, паразитирующие внутри бактриальной клетки. Кроме собственной генетической информации (F-плазмиды) плазмиды могут нести дополнительную генетическую информацию, полезную для бактериальной клетки. Например, плазмиды могут кодировать фермент, разрушающий пенициллин (пенициллиназа). В этом случае бактерия, зараженная такой плазмидой будет устойчива к пенициллину. Плазмиды, кодирующие гены устойчивости к лекарственным препаратам, называются R-плазмидами (от resistance). Плазмиды, несущие только свою собственныю информацию, называют F- плазмидами.
- Клеточная мембрана – ограничивает цитоплазму, защищает от осмотического шока, выполняет функции защиты, транспорта, формообразования, деление клетки и транспорт метаболитов.
 Состоит из двойного слоя фосфолипидов и встроенных мембранных белков. КМ кроме барьерной и транспортной функций выполняют роль центра метаболической активности (в отличие от эукариотической клетки). Белки мембраны, ответственные за перенос необходимых веществ в клетку, называют пермеазами. На внутренней поверхности КМ находятся ферментные комплексы, т. е. упорядоченные скопления молекул ферментов, ответственных за синтез энергоносителей – молекул АТФ. КМ может образовывать выпячивания в цитоплазму, которые называют мезосомами, они усиливают клеточную мембрану.
Состоит из двойного слоя фосфолипидов и встроенных мембранных белков. КМ кроме барьерной и транспортной функций выполняют роль центра метаболической активности (в отличие от эукариотической клетки). Белки мембраны, ответственные за перенос необходимых веществ в клетку, называют пермеазами. На внутренней поверхности КМ находятся ферментные комплексы, т. е. упорядоченные скопления молекул ферментов, ответственных за синтез энергоносителей – молекул АТФ. КМ может образовывать выпячивания в цитоплазму, которые называют мезосомами, они усиливают клеточную мембрану.
Существует два вида мезосом:
- Септальные – образуют поперечные перегородки в процессе деления клетки.
- Латеральные – служат для увеличения поверхности КМ и повышения скорости обменных процессов.
Нуклеоид, ЦП и КМ образуют протопласт.
Одним из отличительных свойств бактерий является очень высокое внутриклеточное осмотическое давление (от 5 до 20 атм), что является результатом интенсивного обмена веществ. Поэтому для защиты от осмотического шока бактериальная клетка окружена прочной клеточной стенкой.
Поэтому для защиты от осмотического шока бактериальная клетка окружена прочной клеточной стенкой.
Бактериальная клетка. Классификации
По строению клеточной стенки все бактерии делятся на 2 группы:
- 1) Имеющие однослойную клеточную стенку – Грам-положительные.
- 2) Имеющие двухслойную клеточную стенку – Грам-отрицательные.
Названия Грам+ и Грам- имеют свою предисторию. В 1884 датский микробиолог Ганс Христиан Грам разработал оригинальный метод окраски микробов, в результате которого одни бактерии окрашивались в синий цвет (грам+), а другие в красный (грам-). Химическая основа различной окраски бактерий по методу Грама была выяснена сравнительно недавно – около 35 лет назад. Оказалось, что Г- и Г+ бактерии имеют разное строение клеточной стенки.
Структура бактериальной клетки включает клеточную стенку, они отличаются у граммположительных и граммотрицательных клеток.
Клеточная стенка Граммположительных (Г+) бактерий в основном состоит из 2 полимеров – пептидогликана и тейхоевых кислот.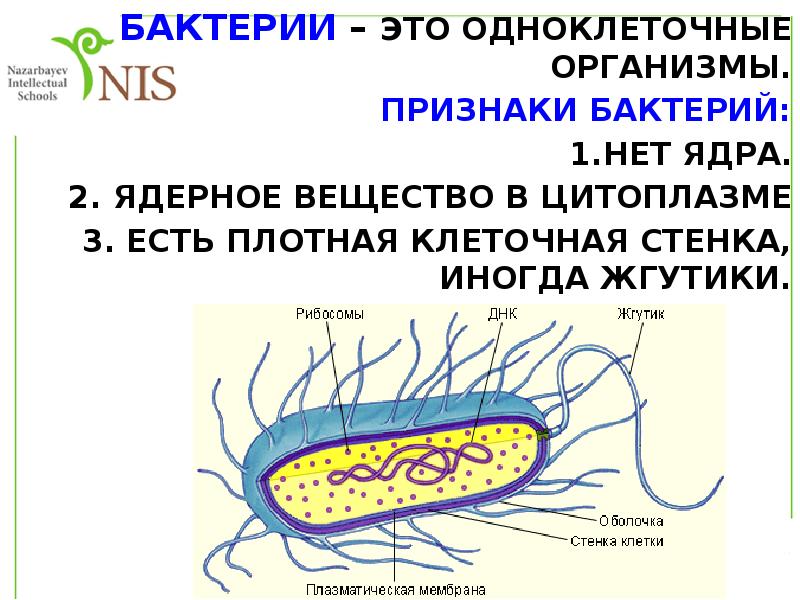 Пептидогликан представляет собой линейный полимер, в котором чередуются остатки мурамовой кислоты и ацетилглюкозамина. С мурамовой кислотой ковалентно связан тетрапептид (белок). Нити пептидогликана связаны между собой через пептиды и образуют прочный каркас – основу клеточной стенки. Между нитями пептидогликана находится другой полимер – тейхоевые кислоты (глицерол ТК и рибитол ТК) – полимер полифосфатов. Тейхоевые кислоты выступают на поверхности клеточной стенки и являются главными АГ Г+ бактерий. Кроме этого, в состав клеточной стенки Г+ бактерий входит рибонуклеат Mg.
Пептидогликан представляет собой линейный полимер, в котором чередуются остатки мурамовой кислоты и ацетилглюкозамина. С мурамовой кислотой ковалентно связан тетрапептид (белок). Нити пептидогликана связаны между собой через пептиды и образуют прочный каркас – основу клеточной стенки. Между нитями пептидогликана находится другой полимер – тейхоевые кислоты (глицерол ТК и рибитол ТК) – полимер полифосфатов. Тейхоевые кислоты выступают на поверхности клеточной стенки и являются главными АГ Г+ бактерий. Кроме этого, в состав клеточной стенки Г+ бактерий входит рибонуклеат Mg.
Клеточная стенка Граммотрицаительных (Гр-) бактерий состоит из 2-х слоев: внутренний слой представлен моно- или бислоем пептидогликана (тонкий слой) . Наружный слой состоит из липополисахаридов, липопротеина, белков, фосфолипидов. ЛПС всех Г- бактерий обладают токсическими и порогенными свойствами и называются эндотоксинами. При воздействии некоторых веществ, например пенициллина, нарушается синтез пептидогликанового слоя. При этом из Г+ бактерий образуется протопласт, а из Г- сферопласт ( т.к. сохраняется наружный слой клеточной стенки).
При этом из Г+ бактерий образуется протопласт, а из Г- сферопласт ( т.к. сохраняется наружный слой клеточной стенки).
При определенных условиях культивтрования клетки, лишенные клеточной стенки, сохраняют способность к росту и делению, и такие формы называют L- формами (по названию Института Листера, где было открыто это явление). В некоторых случаях после устранения фактора, тормозящего синтез клеточной стенки L-формы могут превратиться в исходные формы.
Многие бактерии синтезируют слизистое вещество, состоящее из мукополисахаридов, которое откладывается с наружной стороны клеточной стенки, окружая бактериальную клетку слизистым чехлом — это так называемая капсула. Функция капсулы – защита бактерий от фазоцитоза.
Жгутики – орган движения, спиральные нити, состоящие из белка флагеллина, основание связано с клеточной мембраной по средством базального тельца. Они выявляются с помощью электронной микроскопии препаратов, запыленных тяжелыми металлами или в световом после обработки спецметодами (серебрение).
По расположению жгутиков:
– монотрихи,
– лофотрихи,
– амфитрихи,
– перитрихи.
Бактериальная клетка. Поверхностные структуры.
1. Органы прикрепления к субстрату (адгезии) – пили (фимбрии) или реснички. Начинаются от мембраны клетки. Сосотоят из белка пилина. Число пилей может достигать 400 на 1 клетку.
2. Органы передачи наследственной информации – F-пили или sex-пили. F-пили образуются только в том случае, если клетка нечет плазмиду, т.к. белки F-пили кодирует ДНК плазмиды. Они представляют собой тонкую длинную трубочку, которая прикрепляется к другой бактериальной клетке. Через образовавшийся канал плазмида переходит в соседнюю бактериальную клетку.
3. Органы движения – жгутики – представляют собой спиральные нити. Их длина может превышать их диаметр в 10 и более раз. Жгутики состоят из белка флагеллина. Основание жгутика связано с клеточной мембраной посредством базального тельца. Базальное тельце состоит из системы колец, которые вращаясь передают вращательное движение жгутику.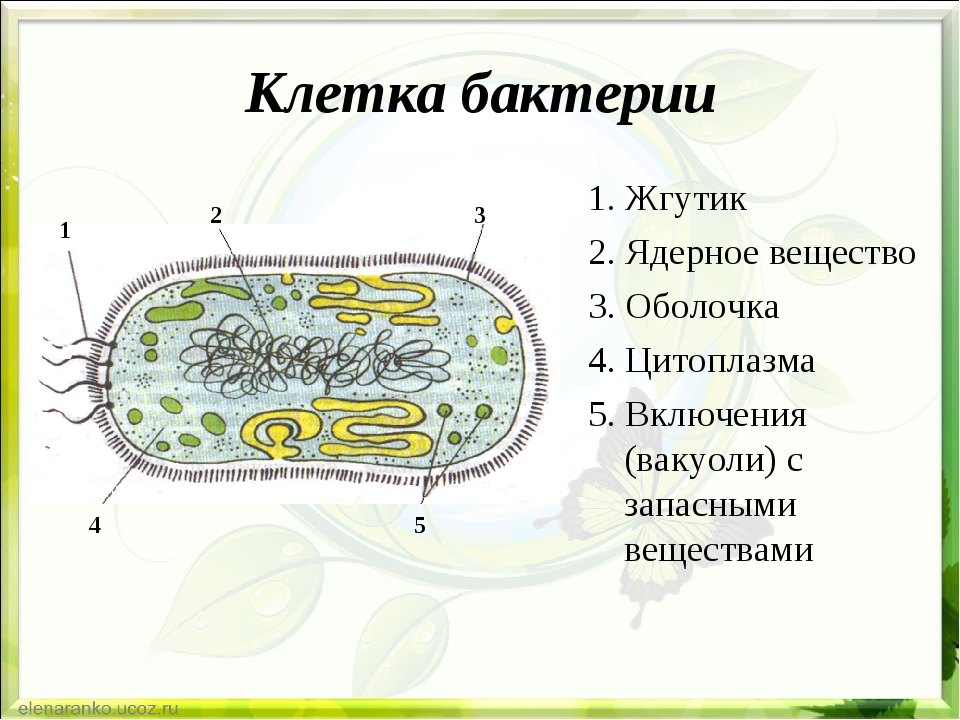 По расположению жгутиком бактерии делятся на моно-, лофо-, амфи-, перитрихи.
По расположению жгутиком бактерии делятся на моно-, лофо-, амфи-, перитрихи.
Бактериальная клетка. Спорообразование.
Споры – способ выживания в неблагоприятной ситуации, образуют только анаэробные бактерии, они выдерживают кипячение, холод и т.д. Внутри одной бактерии образуется одна спора, поэтому образование спор способствует сохранению вида и не является способом размножения как у грибов. При неблагоприятных условиях некоторые бактерии способны образовывать эндоспоры – покоящиеся клетки. Споры чрезвычайно устойчивы к высоким температурам, высушиванию, химическим веществам и сохраняют жизнеспособность в течение десятков лет. В отличие от вегетативных форм, в спорах почти полностью подавлен обмен, геном находится в репрессивном состоянии. Споры бактерий имеют многослойную оболочку, богатую кальцием, и очень небольшое количество свободной воды. При попадании в благоприятные условия спора прорастает, и из нее образуется вегетативная клетка.
Бациллы – спорообразующие анаэробные бактерии, у которых размер споры не превышает диаметр клетки. Клостридии – спорообразующие анаэробные бактерии, у которых размер споры превышает диаметр клетки, поэтому они принимают форму веретена. Процесс спорообразования происходит ряд стадий, в течение которых часть цитоплазмы и хромосома отделяются, окружаясь цитоплазматической мембраной, образуется проспора, затем образуется плохо проницаемая оболочка. Этот процесс сопровождается большим потреблением дипиколиновой кислоты и ионов кальция. После формования всех структур спора приобретает термоустойчивость, которую связывают с наличием дипиколината кальция.
Клостридии – спорообразующие анаэробные бактерии, у которых размер споры превышает диаметр клетки, поэтому они принимают форму веретена. Процесс спорообразования происходит ряд стадий, в течение которых часть цитоплазмы и хромосома отделяются, окружаясь цитоплазматической мембраной, образуется проспора, затем образуется плохо проницаемая оболочка. Этот процесс сопровождается большим потреблением дипиколиновой кислоты и ионов кальция. После формования всех структур спора приобретает термоустойчивость, которую связывают с наличием дипиколината кальция.
Морфологические особенности некоторых групп микроорганизмов.
Спирохеты.
Порядок Spirochartales
Cем. Spirichaetaceae и Leptospiraceae
Род Treponema, Leptospira и Borrelia
Описаны в 1868 Обермеййером. Г- тонкие подвижные извитые микроорганизмы.длина от 5 до 250мкм, спор и капсул не образуют. В неблагоприятных условиях некоторые спирохеты (T. pallidum) образуют покоящиеся формы – цисты. Содержат много липидов, поэтому плохо окрашиваются по Граму. Для окраски используется метод Романовского_Гимза. Подвижность обусловоена фибриллярным аппаратом, состоящим из сократительного белка.
pallidum) образуют покоящиеся формы – цисты. Содержат много липидов, поэтому плохо окрашиваются по Граму. Для окраски используется метод Романовского_Гимза. Подвижность обусловоена фибриллярным аппаратом, состоящим из сократительного белка.
Осевые фибриллы закреплены на противоположных концах клетки и обвивают цитоплазму. Количество фибрилл у трепонем 8-14, у боррелий 15-20, у лептоспир 2. Количество завитков: у трепонем 8-14 туго закрученных с одинаковой амплитудой, у боррелий до 20 неравномерных завитков, у лептоспир большое количество мелких витков, причем концы клетки образуют характерные крючки, отчего лептоспира непоминает по форме латинские буквы С или S.
Многие спирахеты не удается культивтровать на питательных средах.
Заболевания, вызываемые спирохетами:
Treponema pallidum – сифилис
Borrelia recurrentis – возвратный тиф
Borrelia burdorferi – болезнь Лайма (неспецифический полиартрит)
Leptospira interrogans – лептоспироз
Бактериальная клетка.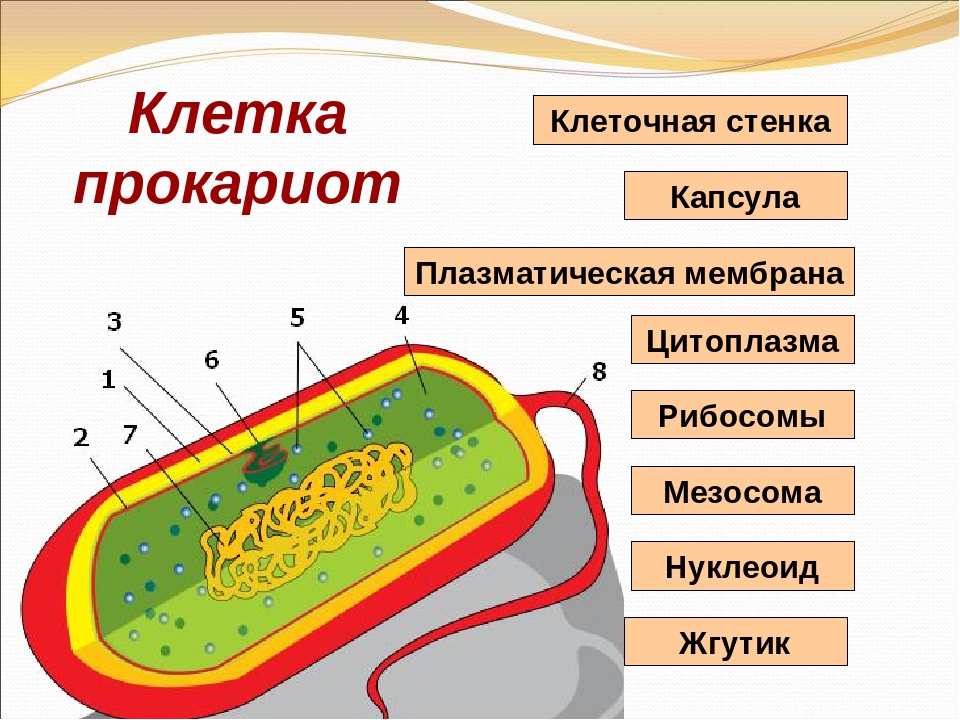 Микоплазмы.
Микоплазмы.
Относятся к классу Mollicates.
Виды, патогенные для человека: M.pneumoniae, M.hominis, M.urealyticum.
Широко распространенная в природе группа прокариот, не имеющая клеточной стенки. Не окрашивается по Граму. Клетка имеет неправильную, изменчивую форму и размеры. Могут приобретать шаровидную форму, нитевидную. Размножаются почкованием и фрагментацией. Имеют самый малый геном среди бактерий – кольцевидную молекулу ДНК. В каждой бактериальной клетке может присутствовать от нескольких до несколько десятков геномов. Микоплазмы размножаются почкованием. Почка микоплазмы называется «элементарное тельце», размеры которого сравнимы с размерами крупных вирусов.
Микоплазмы способны расти на плотных питательных средах сложного состава, требуют добавления факторов роста: глюкозы, аргинина, мочевины, холестерола. Образующиеся колонии имеют в центре возвышение (похожи на яичницу-глазунью).
Микоплазмы являются возбудителями артритов, пневмоний, заболеваний урогенитального тракта.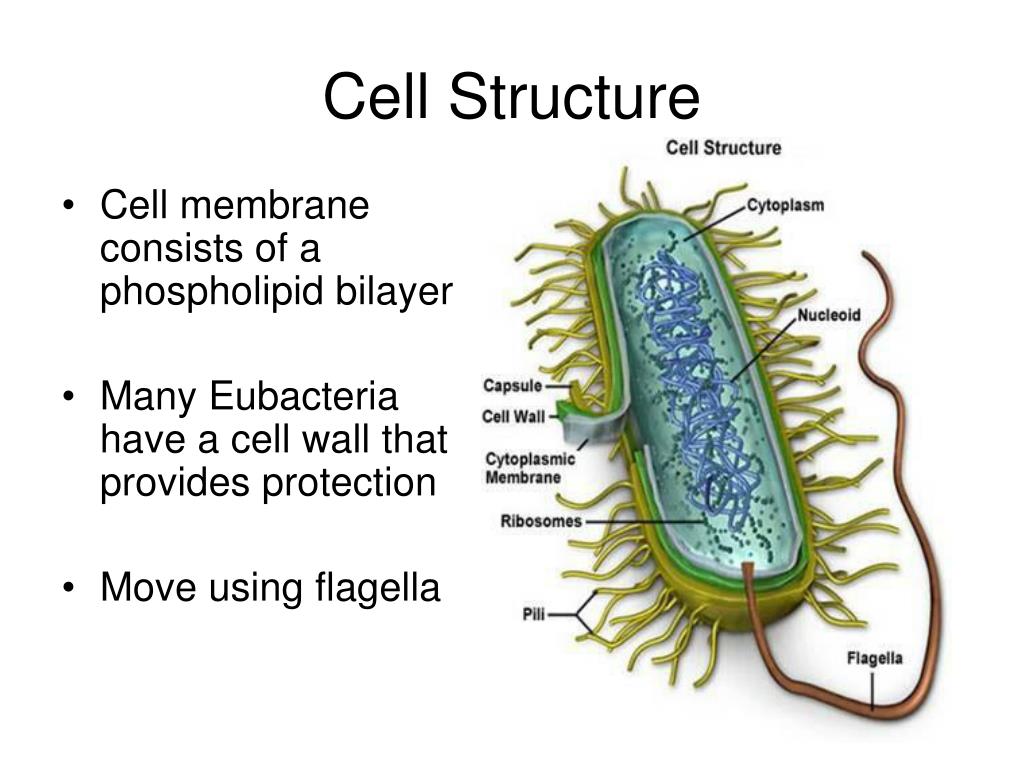
Бактериальная клетка. Рикетсии.
Рикетсии являются облигатными внутриклеточными паразитами. Культивируются на куриных эмбрионах, в культуре тканей. По Граму окрашиваются плохо. Метод окраски – по Романовскому-Гимза. По морфологии различают кокковидные, палочковидные, нитевидные формы. Естественным резервуаром в природе риккетсий являются членистоногие. Заболевания, вызываемые риккетсиями:
R.prowazekii – Сыпной тиф
C.burneti – Лихорадка Ку
Бактериальная клетка. Хламидии.
Описаны в 1907 Хальберштедтером.
Порядок – Chlamydiales
Семейство – Chlamydiaceaea
Род Chlamydia и Сhlamydophila
Мелкие неподвижные Г- бактерии кокковидной формы. Облигатные внутриклеточные паразиты. Спор и капсул не образуют. В цитоплазме зараженных клеток хламидии существуют в виде ретикулярных телец – репродуктивных форм. Оазмеры РТ около 1 мкм. После нескольких циклов деления РТ конденсируются, уплотняясь и уменьшаясь в размерах до 0,3мкм, образуя т.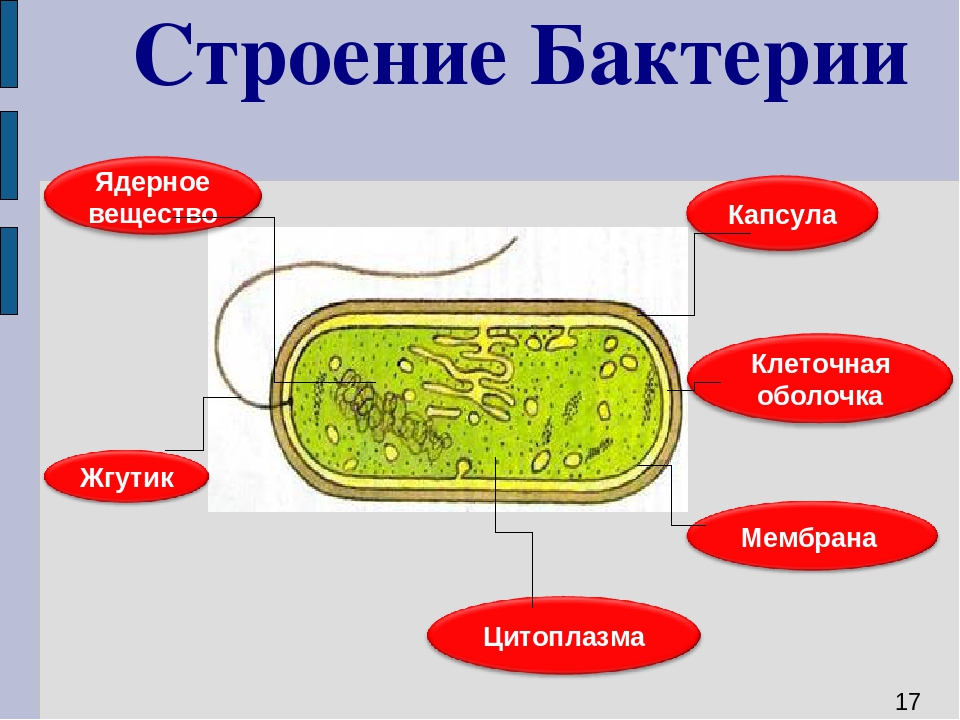 н. элементарные тельца – инфеционная форма хламидии. ЭТ после гибели клетки выходят в окружающую среду и способны заражать новые клетки.
н. элементарные тельца – инфеционная форма хламидии. ЭТ после гибели клетки выходят в окружающую среду и способны заражать новые клетки.
Бактериальная клетка. Патогенные хламидии.
Chlamydia trachomatis – трахома и урогенитальный хламидиоз.
Chlamydophila psittaci – пситтакоз
Chlamydophila pneumoniae – пневмония
Бактериальная клетка. Актиномицеты
Порядок Actinomycetales
Сем. Mycobacteriaceae, Nocardiaceae и Straptomycetaceae
Впервые описаны Харнем (?) в 1877. По внешним признакам сходны с мицелиальными грибами, однако относятся к бактериям, т.к. являются прокариотами. Актиномицеты имеют форму палочек или длинных ветвящихся нитей – гифов. Размножаются фрагментицией, некоторые (нокардии и стрептомицеты) – спорами.
Построению клеточной стенки Г+, капсул не образуют. Большинство актиномицетов обитаетв почве. Многие из них являются продуцентами антибиотиков.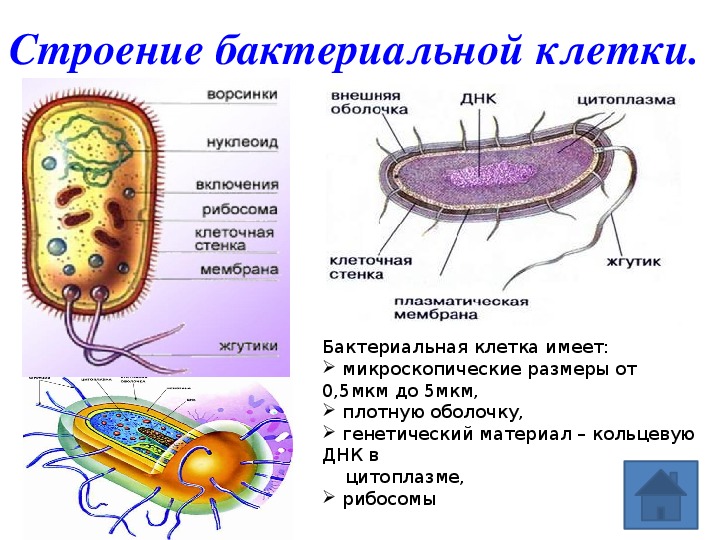
Бактериальная клетка. Патогенные виды.
Actinomyceti israelii – актиномикоз
Nocardia asteroides – нокардиоз
Механизм действия бактериофагов, как это работает. Фагочувствительность
открытие пенициллина – первого антибиотика природного происхождения, воспринималось поначалу как шанс раз и навсегда победить инфекционные заболевания, уносившие миллионы жизней. Однако во второй половине прошлого века первоначальная эйфория пошла на убыль: выяснилось, что бактерии благодаря мутациям могут приобретать устойчивость к любому антибиотику и даже обмениваться между собой этими генами. Так началась «гонка вооружений» человека и бактерий.
Между препаратами бактериофагов и химическими антибиотиками существует принципиальное различие: антибиотик, в отличие от фагового препарата, убивает всё на своем пути, включая содружество полезных микроскопических обитателей организма.
Cовместная эволюция бактерий и фагов за последние 3 миллиарда лет привела к тому, что практически для любой болезнетворной бактерии имеется соответствующий вирус-«убийца».
В отличие от обычных антибиотиков, бактериофаги модифицируют свои генетические программы сами, что снимает проблему устойчивости. Результат – высокая избирательность: конкретный бактериофаг обычно эффективен в отношении лишь одного определенного бактериального вида или даже штамма бактерий.
Процесс уничтожения бактериальной клетки бактериофагом включает в себя несколько строго запрограммированных шагов.
Действие бактериофагов
Очень важным является то, что бактериофаги не трогают не «свои» бактерии, поэтому не вызывают гибель «хорошей» микрофлоры и, конечно, абсолютно безопасны для клеток высших организмов, включая человека.
При фаготерапии отсутствуют побочные эффекты, такие как аллергия, дисбактериоз, вторичные инфекции (например, грибковые), что нередко наблюдается при приеме антибиотиков. Бактериофаги могут применяться в комбинированной терапии с любыми лекарственными препаратами, включая антибиотики.
Препараты бактериофагов представляют собой раствор, используют их либо местно (например, на кожу или слизистые), либо принимают внутрь. В организме бактериофаги концентрируются в местах наибольшего поражения и размножаются до тех пор, пока находят бактерии-«мишени». После того, как бактерии-«мишени» закончились, фаги выводятся из организма.
В организме бактериофаги концентрируются в местах наибольшего поражения и размножаются до тех пор, пока находят бактерии-«мишени». После того, как бактерии-«мишени» закончились, фаги выводятся из организма.
Главная цель фаготерапии – остановить и повернуть вспять инфекционный процесс, чтобы дать иммунной системе организма возможность справиться с болезнью.
С помощью бактериофагов лечат
В идеальном случае перед началом лечения у конкретного больного следует выделить болезнетворную бактерию в культуру и подобрать бактериофаг из «коллекции» уже известных бактериальных вирусов.
Препараты бактериофагов трудно поддаются стандартизации из-за специфики производства. Даже опытный специалист не всегда может заранее предсказать эффективность того или иного препарата у конкретного пациента, так как разные бактериальные штаммы бактерий могут вызывать схожие проявления болезни. Выход – производство «коктейлей» из фагов, способных убивать разные штаммы и даже виды возбудителей.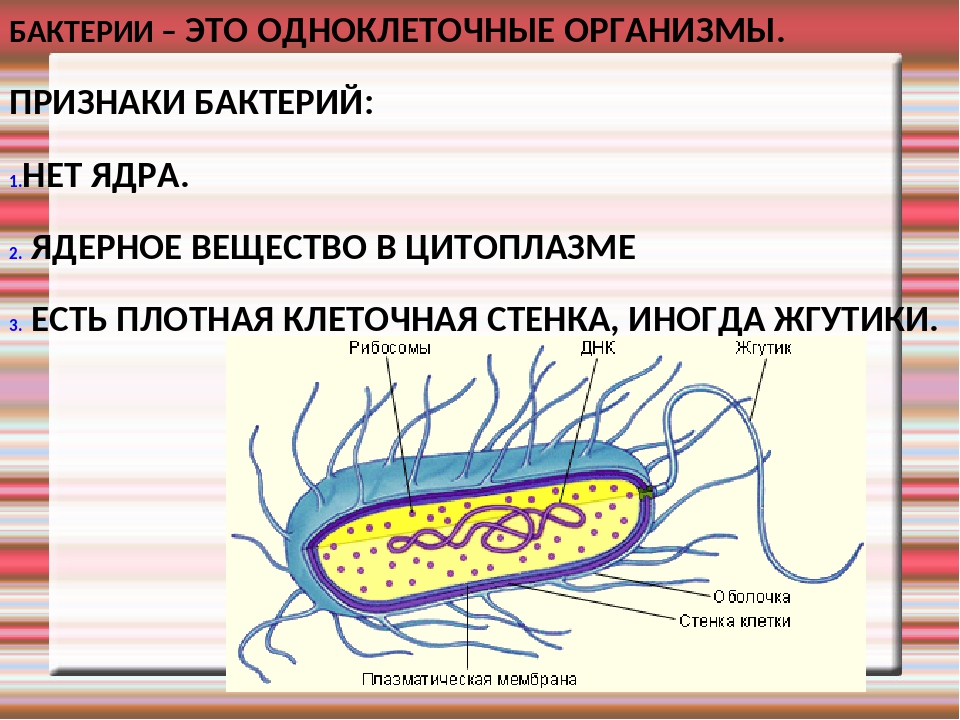 И такие препараты уже существуют — это комплексные, или поливалентные препараты бактериофагов
И такие препараты уже существуют — это комплексные, или поливалентные препараты бактериофагов
6 этапов работы бактериофагов
Условные обозначения
Адсорбция бактериофагов на бактериальных клетках
Специальные элементы фага, расположенные на поверхности в виде фибрилл или шипов, связываются со специфичными поверхностными молекулами — рецепторами на своей жертве — бактерии. Пока бактериофаг достаточно плотно не закрепится на поверхности бактерии, следующей стадии не происходит. Так как существует специальная система блокировки.
Инъекция нуклеиновой кислоты бактериофага внутрь клетки
После плотного прикрепления фага (адсорбции) происходит внедрение генетического материала бактериофага в тело бактерии. Для этого в структуре фага природа предусмотрела наличие специальных структур, которые действуют по типу шприца. Именно за счет этого фаг как бы делает инъекцию, растворяет оболочку и вводит свой генетический материал в бактерию.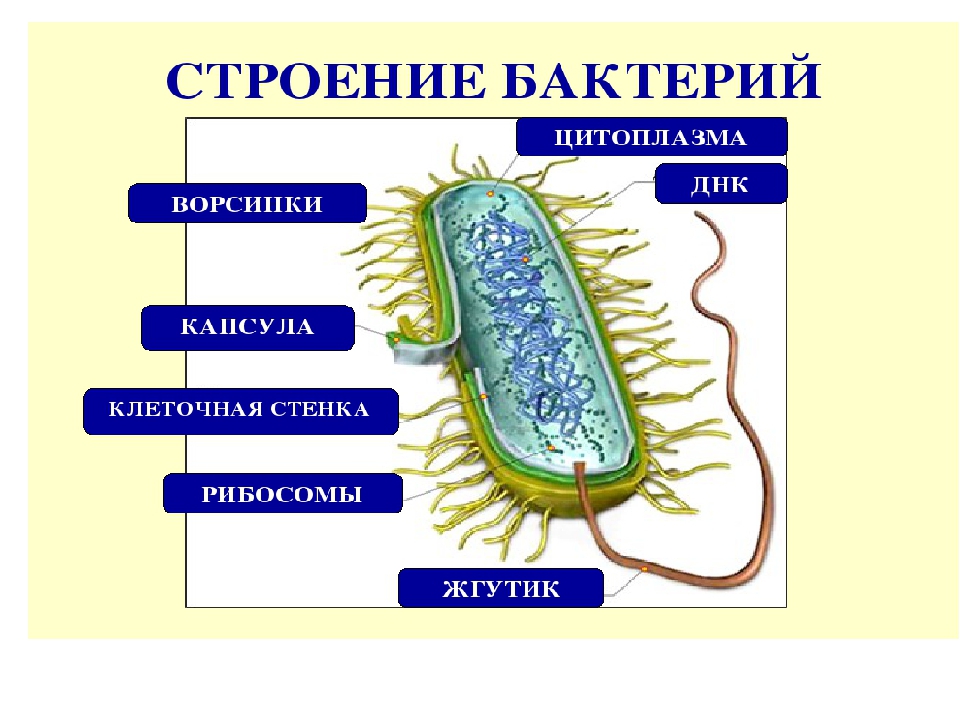 Для этого у фага есть агрессивный фермент для прокалывания бактерии.
Для этого у фага есть агрессивный фермент для прокалывания бактерии.
Сборка фаговых частиц
Сборка молодых фагов начинается с упаковывания генетического материала в икосаэдрические белковые оболочки. Далее к фагам присоединяется хвост, на головке фага появляются различные, необходимые для его жизнедеятельности белки. Количество молодых фагов внутри бактерии возрастает. Новое поколение готовится к выходу из бактериальной клетки.
Репликация копий нуклеиновой кислоты бактериофага
Когда ДНК фага попадает в бактерию она может раствориться ферментами бактерии. Однако фаг защищает свою ДНК специальными липкими белками, которые замыкают молекулу в кольцо, делая ее неуязвимой. Далее происходит тиражирование генетического материала (ДНК) фага прямо в клетке бактерии.
Синтез белковых и нуклеиновых частиц
После заражения бактерии начинается перестройка клеточного метаболизма под нужды фага: разрушаются некоторые клеточные белки.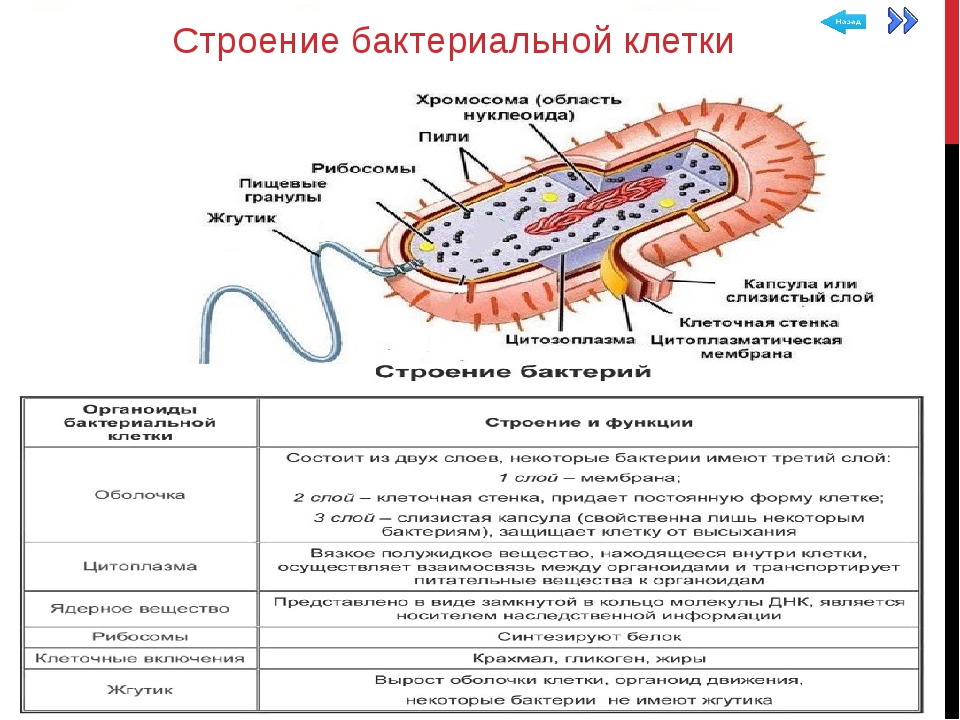 Далее происходит включение генетического материала фага в метаболизм бактерии и начинается сборка новых, молодых фагов.
Далее происходит включение генетического материала фага в метаболизм бактерии и начинается сборка новых, молодых фагов.
Выход зрелых фагов и смерть бактерии
Финал жизненного цикла фага — клеточный лизис. Молодые фаги используют для разрушения бактериальной клетки набор ферментов, которые расщепляют оболочки бактерий (лизины), и белков, создающих поры во внутренней мембране бактерии и обеспечивающих ускорение действия фермента лизина.
Противопоказания
Только индивидуальная непереносимость.
С клинической точки зрения фаги достаточно безопасны, поскольку люди с рождения встречаться с ними. Фаги – это постоянные спутники организма, они были обнаружены в желудочно кишечном тракте, коже, моче, во рту, где они содержаться в слюне. Бактериофаги являются безопасной альтернативой антибиотикам.
Сравнение профилактического и терапевтического использования фагов и антибиотиков
Воздействие на бактерии
Фаги эффективно убивают чувствительные клетки бактерий (их действие бактерицидное).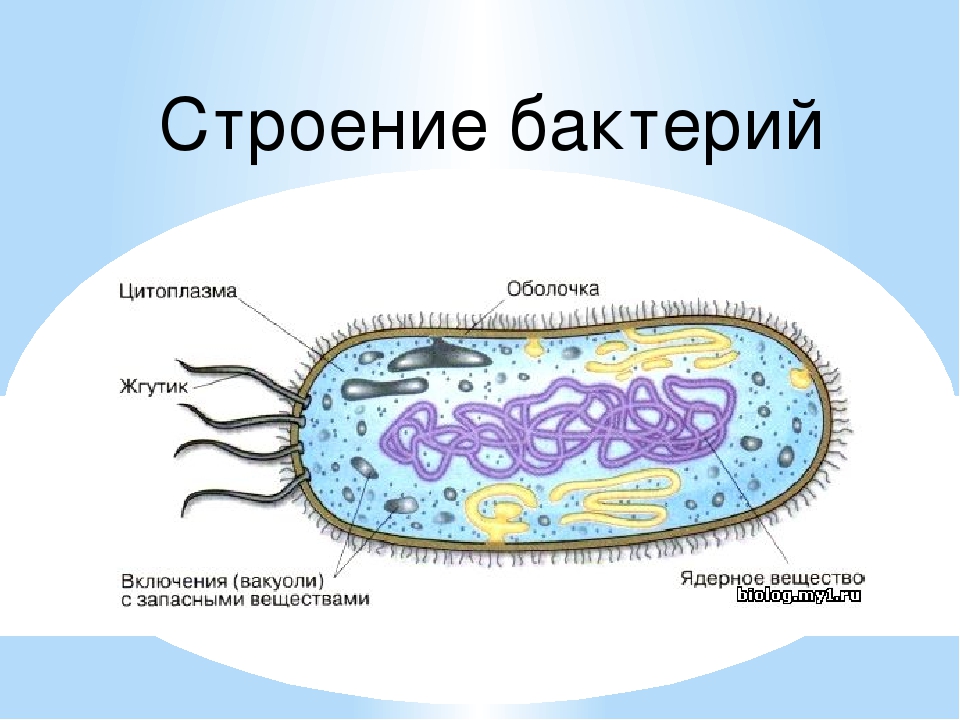
Некоторые антибиотики (например, хлорамфеникол) бактериостатические; они скорее ингибируют рост бактерий, чем убивают клетки.
Селективность действия
Высокая селективность бактериофагов позволяет нацеливаться на специфичных патогенов, не затрагивая нормальную бактериальную флору (например, маловероятно, что фаги будут враждебны по отношению к нормофлоре пациентов).
Антибиотики атакуют не только бактерий – возбудителей заболеваний, но также все чувствительные микроорганизмы, включая нормальную – и часто полезную – микрофлору хозяина. Поэтому их неселективное действие нарушает микробный баланс в организме пациента, что может привести к различным побочным эффектам.
Побочные эффекты
Незначительные побочные эффекты при применении лечебных фагов (могут быть вызваны высвобождением эндотоксинов из бактерий, лизированных in vivo фагами).
Для антибиотиков характерно множество побочных эффектов, включая кишечные расстройства, аллергии и вторичные инфекции (например, грибковые инфекции).
Разработка новых препаратов
Естественная коэволюция бактерий и фагов может облегчить получение новых литических фагов против фагоустойчивых бактерий, появляющихся в результате действия других фагов или естественных изменений в бактериальных популяциях.
Создание новых антибиотиков (например, против антибиотикоустойчивых бактерий) требует временных затрат и может занять много лет.
Устойчивость бактерий к действию препарата
Из-за специфичности фагов маловероятно появление устойчивости у других бактериальных видов, не являющихся мишенями терапии.
Из-за широкого спектра активности антибиотики могут приводить к отбору устойчивых мутантов многих видов бактерий, а не только у штаммов мишени.
Применение препарата
Из-за специфичности фагов их успешное использование для предупреждения и лечения бактериальных инфекций требует идентификации этиологического агента и определения его фагочувствительности к препарату in vivo перед началом фагового лечения.
Эмпирически назначенные до идентификации этиологического агента антибиотики с большей вероятностью будут эффективны нежели фаговые препараты.
Строение клеток бактерий | Биология
Первые бактерии появились, вероятно, более 3.5 млрд. лет назад и на протяжении почти миллиарда лет были единственными живыми существами на нашей планете. В настоящее время они распространены повсеместно и определяют различные процессы, происходящие в природе.
Форма и размеры бактерий
Бактерии — это одноклеточные микроскопические организмы. Они имеют форму палочек, шариков, спиралей. Некоторые виды образуют скопления но нескольку тысяч клеток. Длина палочковидных бактерий составляет 0,002—0,003 мм. Поэтому даже при помощи микроскопа отдельные бактерии увидеть очень трудно. Однако их легко заметить невооруженным глазом, когда они развиваются в большом количестве и образуют колонии. В лабораторных условиях колонии бактерий выращивают на специальных средах, содержащих необходимые питательные вещества.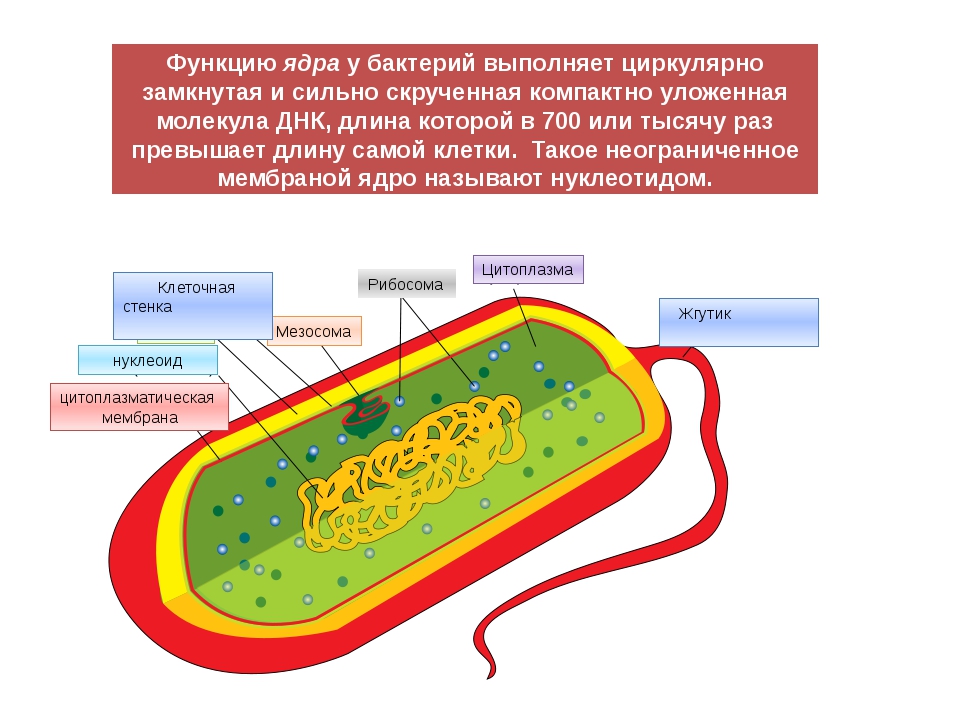
Строение бактериальной клетки
Бактериальная клетка, как и клетки растений, грибов и животных, покрыта плазматической мембраной. Но в отличие от них с внешней стороны мембраны расположена плотная клеточная оболочка. Она состоит из прочного вещества и выполняет одновременно защитную и опорную функции, придавая клетке постоянную форму. Через клеточную оболочку питательные вещества свободно проходят в клетку, а ненужные вещества выходят в окружающую среду. Часто поверх клеточной оболочки у бактерий вырабатывается дополнительный защитный слой слизи — капсула.
На поверхности клеточной оболочки некоторых бактерий имеются выросты — длинные жгутики (один, два и более) или короткие тонкие ворсинки. С их помощью бактерии передвигаются. В цитоплазме бактериальной клетки находится ядерное вещество — нуклеоид, которое несет наследственную информацию. Ядерное вещество в отличие от ядра не отделено от цитоплазмы. В связи с отсутствием оформленного ядра и другими особенностями строения клетки все бактерии объединяются в отдельное царство живой природы — царство Бактерий.
Распространение бактерий и их роль в природе
Бактерии — самые распространенные на Земле живые существа. Они обитают повсюду: в воде, воздухе, почве. Бактерии способны жить даже там, где не могут выжить другие организмы: в горячих источниках, во льдах Антарктиды, в подземных нефтяных месторождениях и даже внутри атомных реакторов. Каждая бактериальная клетка очень мала, но общее количество бактерий на Земле огромно. Это
связано с высокой скоростью размножения бактерий. Бактерии выполняют в природе самые разнообразные функции.
Велика роль бактерий в образовании топливных полезных ископаемых. Миллионы лет они разлагали останки морских организмов и наземных растений. В результате жизнедеятельности бактерий сформировались залежи нефти, природного газа, угля.
Цитоплазма — обзор | ScienceDirect Topics
4.1 Нацеливание на цитоплазму
Цитоплазма очень вязкая, что затрудняет перенос макромолекул внутри клетки. Некоторые исследования подтвердили участие актиновых филаментов в интернализации НЧ и определили роль микротрубочек в цитоплазматическом переносе полиплексов и липоплексов посредством везикулярного транспорта в перинуклеарную область (Sauer et al.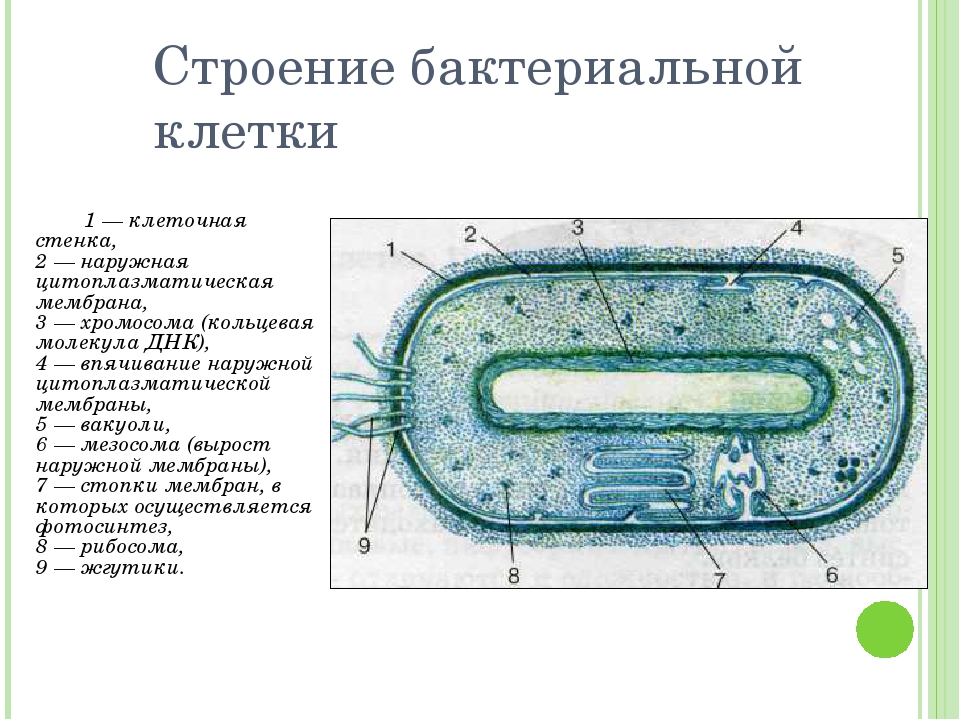 , 2009; Tammam et al., 2016). Несколько факторов могут влиять на способность НЧ диффундировать через цитоплазму.Например, эффективность диффузии НЧ зависит от их размера и формы. Меньшие сферические НЧ показали более высокую диффузию по сравнению с более крупными несферическими НЧ (Tammam et al., 2016). Более того, ПЭГилирование резко улучшило перемещение по цитоплазме из-за его слабого взаимодействия с цитоплазматическими органеллами. Однако для преодоления негативного эффекта ПЭГилирования на эндосомальное ускользание цепи ПЭГ конъюгируют с поверхностью НЧ с помощью расщепляемых линкеров (Jhaveri and Torchilin, 2016).
, 2009; Tammam et al., 2016). Несколько факторов могут влиять на способность НЧ диффундировать через цитоплазму.Например, эффективность диффузии НЧ зависит от их размера и формы. Меньшие сферические НЧ показали более высокую диффузию по сравнению с более крупными несферическими НЧ (Tammam et al., 2016). Более того, ПЭГилирование резко улучшило перемещение по цитоплазме из-за его слабого взаимодействия с цитоплазматическими органеллами. Однако для преодоления негативного эффекта ПЭГилирования на эндосомальное ускользание цепи ПЭГ конъюгируют с поверхностью НЧ с помощью расщепляемых линкеров (Jhaveri and Torchilin, 2016).
Различные типы наноносителей были изучены для доставки через цитоплазму (таблица 14.1). Однако липосомы являются наиболее широко изученным типом наносистем, и в литературе сообщалось о различных стратегиях, способствующих доставке активных молекул в цитоплазму клеток-мишеней. Использование фузогенных липидов в кислых условиях способствует взаимодействию и слиянию эндосомальных мембран, что приводит к доставке груза в цитоплазму (Jhaveri, Torchilin, 2016).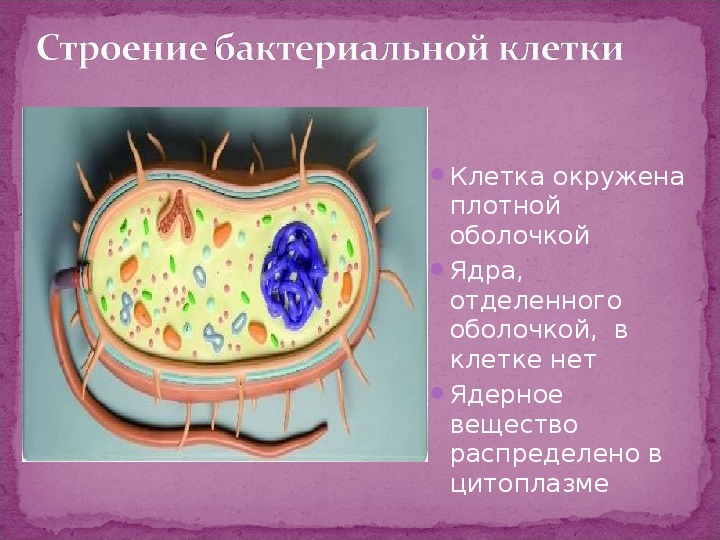 Было показано, что pH-чувствительные фузогенные липосомы являются эффективными системами доставки антигена для иммунотерапии рака.Мансурян и соавт. (2014) включили хелпер-фузогенный липид DOPE в катионные липосомы, состоящие из DOTAP-холестерина, для усиления доставки в цитозоль пептида p5 HER-2/neu (p5) и для стимуляции реакции цитотоксических Т-лимфоцитов (CTL). Эта сложная липосома усиливала ответ ЦТЛ и ингибировала прогрессирование опухоли.
Было показано, что pH-чувствительные фузогенные липосомы являются эффективными системами доставки антигена для иммунотерапии рака.Мансурян и соавт. (2014) включили хелпер-фузогенный липид DOPE в катионные липосомы, состоящие из DOTAP-холестерина, для усиления доставки в цитозоль пептида p5 HER-2/neu (p5) и для стимуляции реакции цитотоксических Т-лимфоцитов (CTL). Эта сложная липосома усиливала ответ ЦТЛ и ингибировала прогрессирование опухоли.
Другой подход к доставке НЧ по цитоплазме включает использование катионных наноносителей и СРР. Они также известны как домены белковой трансдукции и представляют собой короткие катионные пептиды с доменами из 20 аминокислот, с обильными остатками лизина и аргинина, которые опосредуют транслокацию через клеточные мембраны и усиливают доставку грузов, модифицированных CPP, внутрь клетки (Torchilin, 2008a). ).CPP может доставлять различные макромолекулы, включая наноносители, белки, нуклеиновые кислоты и лекарства.
Было предложено множество механизмов интернализации CPP; однако не было установлено единого пути в качестве механизма опосредованного CPP поглощения (Jhaveri and Torchilin, 2016).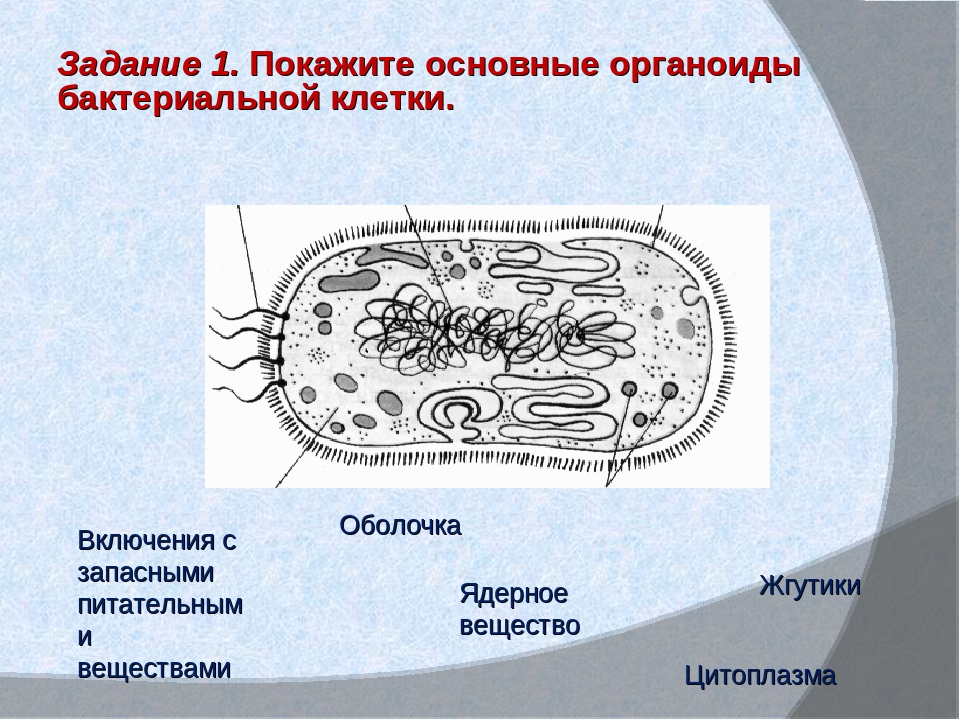 CPP или конъюгаты CPP-cargo могут использовать один или несколько, как энергозависимых, так и энергонезависимых, механизмов транслокации через клеточные мембраны (Jhaveri, Torchilin, 2016). CPP сами по себе или связанные с небольшими молекулами обычно взаимодействуют с анионной клеточной поверхностью посредством электростатического взаимодействия, быстро интернализируясь в клетки путем энергонезависимой трансдукции через липидный бислой (Herbig et al., 2005; Торчилин, 2008б). И наоборот, CPP, конъюгированные с большими грузами, в основном интернализуются посредством холестерин-зависимого макропиноцитоза (Al Soraj et al., 2012; Wadia et al., 2004). Также сообщалось о механизмах, опосредованных клатрином и кавеолином (Buhl et al., 2014; Mager et al., 2012). Было показано, что CPP, такие как ТАТ, полиаргинин, пенетратин, транспортан, антеннопедия и митоген-активируемый белок, доставляют широкий спектр терапевтических грузов к клеткам-мишеням, включая лекарства, белки, антитела, нуклеиновые кислоты, наноносители и контрастные вещества для МРТ.
CPP или конъюгаты CPP-cargo могут использовать один или несколько, как энергозависимых, так и энергонезависимых, механизмов транслокации через клеточные мембраны (Jhaveri, Torchilin, 2016). CPP сами по себе или связанные с небольшими молекулами обычно взаимодействуют с анионной клеточной поверхностью посредством электростатического взаимодействия, быстро интернализируясь в клетки путем энергонезависимой трансдукции через липидный бислой (Herbig et al., 2005; Торчилин, 2008б). И наоборот, CPP, конъюгированные с большими грузами, в основном интернализуются посредством холестерин-зависимого макропиноцитоза (Al Soraj et al., 2012; Wadia et al., 2004). Также сообщалось о механизмах, опосредованных клатрином и кавеолином (Buhl et al., 2014; Mager et al., 2012). Было показано, что CPP, такие как ТАТ, полиаргинин, пенетратин, транспортан, антеннопедия и митоген-активируемый белок, доставляют широкий спектр терапевтических грузов к клеткам-мишеням, включая лекарства, белки, антитела, нуклеиновые кислоты, наноносители и контрастные вещества для МРТ. , как in vitro, так и in vivo (Farkhani et al., 2014; Олсон и др., 2010 г.; Торчилин, 2008а). Наиболее часто используемым CPP является пептид ТАТ, полипептид из 86 аминокислот с богатой цистеином областью, полученный из белка ТАТ ВИЧ (Gump and Dowdy, 2007).
, как in vitro, так и in vivo (Farkhani et al., 2014; Олсон и др., 2010 г.; Торчилин, 2008а). Наиболее часто используемым CPP является пептид ТАТ, полипептид из 86 аминокислот с богатой цистеином областью, полученный из белка ТАТ ВИЧ (Gump and Dowdy, 2007).
Пептид ТАТ был конъюгирован с несколькими различными молекулами и наноносителями для адресной доставки в ткани и клетки. Несколько ТАТ-конъюгированных NP использовались для доставки siРНК, увеличивая поглощение и специфический нокдаун гена-мишени, опосредованный siRNA, например, NP золота (Huang et al., 2016b), липоплексы (Daniels et al., 2013), НЧ хитозана (Malhotra et al., 2013) и дендримеры. TATp-модифицированные липосомы (TATp-L) также использовались для доставки ДНК в культуры DC (Pappalardo et al., 2014). Включение последовательностей остатков гистидина и цистеина в структуру ТАТ для доставки ДНК увеличило эффективность генной трансфекции до 7000 раз, поскольку присутствие гистидина и цистеина, по-видимому, способствует эндосомальному ускользанию и стабильности комплексов ТАТ/ДНК (Lo and Wang). , 2008).Конъюгация CPPs с гистидинами способствует ускользанию из эндосом из-за эффекта «протонной губки» в кислых эндосомах, вызывая осмотическое набухание и лизис или слияние мембран, что приводит к образованию пор (Martin and Rice, 2007).
, 2008).Конъюгация CPPs с гистидинами способствует ускользанию из эндосом из-за эффекта «протонной губки» в кислых эндосомах, вызывая осмотическое набухание и лизис или слияние мембран, что приводит к образованию пор (Martin and Rice, 2007).
Другой CPP, октааргинин (R8), который имитирует ТАТ, использовался для модификации поверхности липосом, нагруженных пегилированным DOX, коммерчески доступных как DOXIL или Lipodox. Он усиливал противораковую активность и улучшал внутриклеточную доставку и выход DOX из эндосом по сравнению с немодифицированными липосомами, что приводило к более высокой доставке DOX в цитоплазму и ее конечному накоплению в ядре (Biswas et al., 2013а,б). В другом примере Steinbach et al. (2016) использовали CPP (MPG) для связывания клеток и интернализации PLGA NP. Все NP были интернализованы посредством клатрин-опосредованного процесса. Кроме того, две разные поверхностные группы НЧ (авидин и 1,2-дистеароил-sn-глицеро-3-фосфоэтаноламин [DSPE]) конъюгировали для оценки интернализации клеток.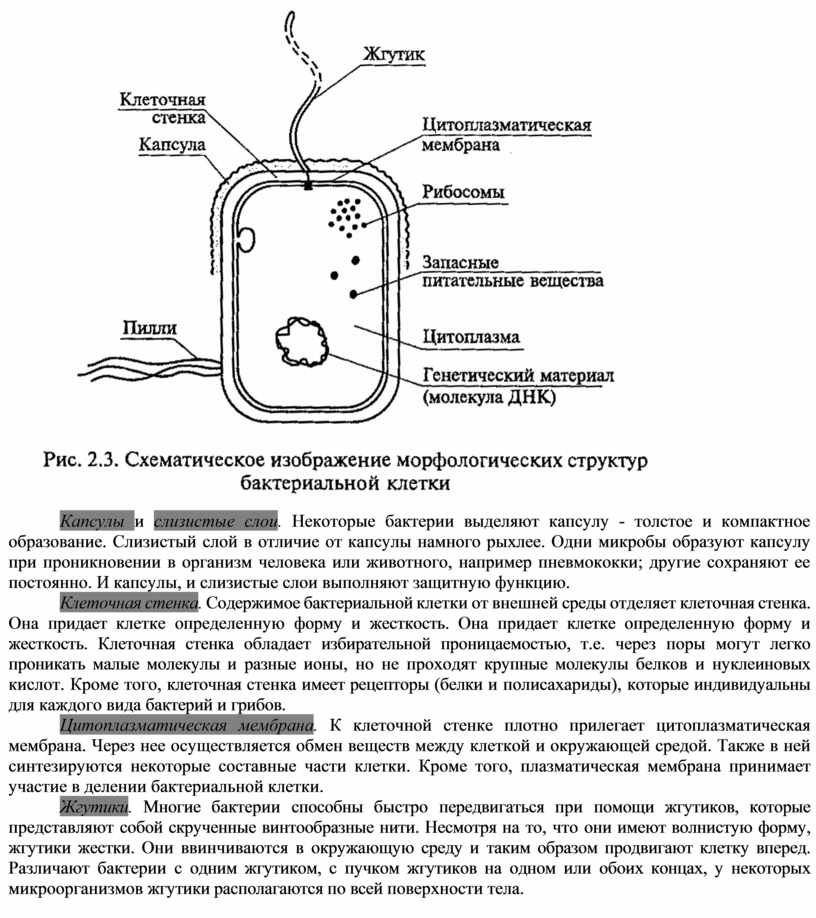 В целом, НЧ, модифицированные авидином и DSPE-CPP, значительно увеличивали интернализацию. Эти результаты демонстрируют многообещающие варианты реализации этой стратегии.
В целом, НЧ, модифицированные авидином и DSPE-CPP, значительно увеличивали интернализацию. Эти результаты демонстрируют многообещающие варианты реализации этой стратегии.
Другим механизмом эндосомального побега является дестабилизация эндосомальной мембраны фузогенными пептидами (Varkouhi et al., 2011). Они состоят из короткого пептидного домена из 20–30 аминокислот и обычно происходят из оболочечных вирусов. Они вызывают эндосомальное ускользание, претерпевая конформационные изменения за счет протонирования терминальных доменов при кислом pH и слияния с мембраной, вызывая дестабилизацию мембраны и транслокацию в цитоплазму (Shete et al., 2014). Аминоконцевая область субъединицы 2 гемагглютинина вируса гриппа содержит последовательность фузогенного пептида, которая использовалась в качестве модели для нескольких синтетических пептидов, включая пептиды, содержащие последовательности GALA или KALA (Rajendran et al., 2010). Эти пептиды используются для доставки в цитозоль различных составов лекарств и нуклеиновых кислот за счет конформационных изменений, обусловленных pH, что приводит к эффективному разрушению эндосомальных мембран (Lee et al. , 2001; Min et al., 2006; Parente et al., 1990; Сасаки и др., 2008). Другие фузогенные пептиды включают производный от гриппа diINF-7, который использовался для индуцирования эндосомального ускользания, конъюгированного с липидными носителями для доставки миРНК (Oliveira et al., 2007) и белка (Mastrobattista et al., 2002), а также для доставки генов. с полимерными векторами (Funhoff et al., 2004; Jiang et al., 2007). Т-домен дифтерийного токсина (Ariansen et al., 1993) использовался для повышения эффективности трансфекции полиплексов PEI (Kakimoto et al., 2009).
, 2001; Min et al., 2006; Parente et al., 1990; Сасаки и др., 2008). Другие фузогенные пептиды включают производный от гриппа diINF-7, который использовался для индуцирования эндосомального ускользания, конъюгированного с липидными носителями для доставки миРНК (Oliveira et al., 2007) и белка (Mastrobattista et al., 2002), а также для доставки генов. с полимерными векторами (Funhoff et al., 2004; Jiang et al., 2007). Т-домен дифтерийного токсина (Ariansen et al., 1993) использовался для повышения эффективности трансфекции полиплексов PEI (Kakimoto et al., 2009).
Дополнительный многообещающий подход к цитозольной доставке основан на окислительно-восстановительных или ферментозависимых связях, которые разрушаются в цитоплазме из-за высокого окислительно-восстановительного потенциала или высоких внутриклеточных концентраций ферментов (Chang et al., 2016; Джавери и Торчилин, 2016). Zhao et al. (2014) разработали окислительно-восстановительную систему доставки на основе коллоидного мезопористого кремнезема (CMS), в которой 6-меркаптопурин был конъюгирован с CMS.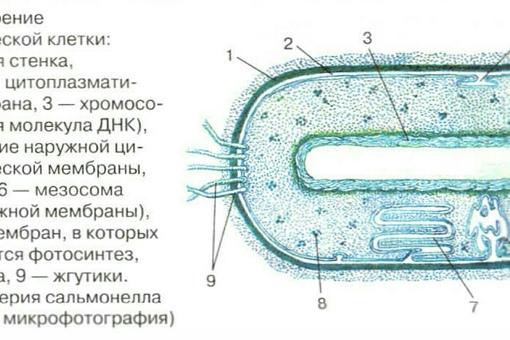 Комплекс показал более высокое клеточное поглощение посредством эндоцитоза, опосредованного рецептором CD44, в клетках колоректальной карциномы человека (HCT)-116. Это исследование предлагает новую стратегию для системы доставки лекарственных средств, реагирующей на стимул.
Комплекс показал более высокое клеточное поглощение посредством эндоцитоза, опосредованного рецептором CD44, в клетках колоректальной карциномы человека (HCT)-116. Это исследование предлагает новую стратегию для системы доставки лекарственных средств, реагирующей на стимул.
5: Бактерии — внутренние компоненты
Мы уже рассмотрели основные внутренние компоненты, присутствующие во всех бактериях, а именно цитоплазму, нуклеоид и рибосомы.Помните, что обычно считается, что у бактерий отсутствуют органеллы, связанные с билипидными мембранами компартменты, столь распространенные в эукариотических клетках (хотя некоторые ученые утверждают, что бактерии обладают структурами, которые можно рассматривать как простые органеллы). Но бактерии могут быть более сложными, с множеством дополнительных внутренних компонентов, которые могут способствовать их возможностям. Большинство этих компонентов являются цитоплазматическими, но некоторые из них являются периплазматическими, расположенными в пространстве между цитоплазматической и внешней мембраной у грамотрицательных бактерий.
Цитоскелет
Первоначально считалось, что у бактерий отсутствует цитоскелет , важный компонент эукариотических клеток. Однако за последние 20 лет ученые обнаружили бактериальные филаменты, состоящие из белков, которые являются аналогами белков цитоскелета, обнаруженных у эукариот. Также было установлено, что бактериальный цитоскелет играет важную роль в формировании формы клеток, делении клеток и целостности клеточной стенки.
ФЦЗ
FtsZ , гомологичный эукариотическому белку тубулину , образует кольцевую структуру в середине клетки во время клеточного деления, привлекая другие белки в эту область, чтобы построить перегородку, которая в конечном итоге разделит две образовавшиеся дочерние клетки.
МРЭБ
MreB , гомологичный эукариотическому белку актину , обнаружен в бациллах и спиралевидных бактериях и играет существенную роль в формировании формы клеток.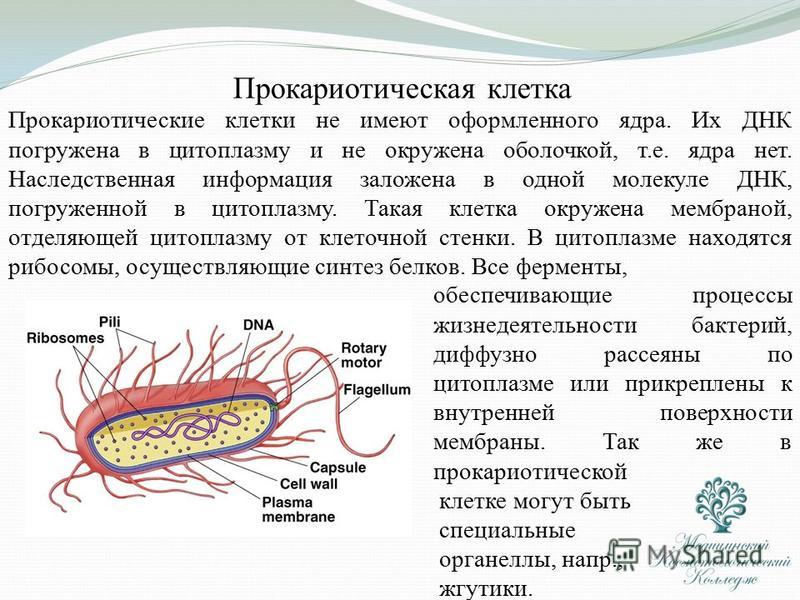 MreB принимает спиралевидную конфигурацию по всей длине клетки и диктует активность аппарата синтеза пептидогликана, обеспечивая несферическую форму.
MreB принимает спиралевидную конфигурацию по всей длине клетки и диктует активность аппарата синтеза пептидогликана, обеспечивая несферическую форму.
Крестинин
Кресцентин , гомологичный эукариотическим белкам ламин и кератин , обнаружен в спиралевидных бактериях с одиночной кривой.Белок собирается вдоль внутренней кривизны клетки, изгибая клетку и придавая ей окончательную форму.
Структуры цитоскелета.
Включения
Бактериальные включения обычно определяются как отдельная структура, расположенная либо в цитоплазме, либо в периплазме клетки. Они могут варьироваться по сложности, от простого набора химических веществ, таких как кристаллы, до довольно сложных структур, которые начинают конкурировать со структурами эукариотических органелл, дополненных мембранным внешним слоем.Их роль часто заключается в хранении компонентов в качестве метаболических резервов для клетки, когда вещество обнаруживается в избытке, но они также могут играть роль в подвижности и метаболических функциях.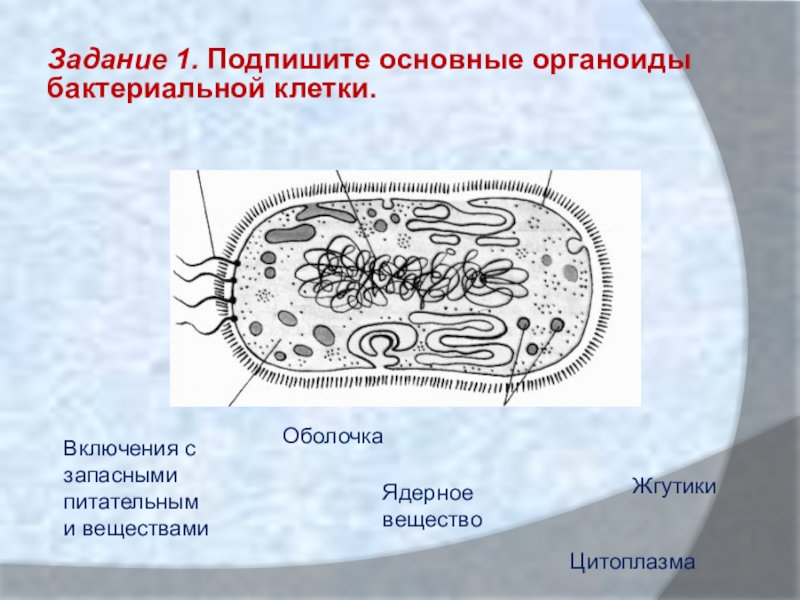
Хранение углерода
Углерод является наиболее распространенным веществом, запасаемым клеткой, поскольку все клетки основаны на углероде. Кроме того, соединения углерода часто могут быстро расщепляться клеткой, поэтому они также могут служить источниками энергии. Одним из самых простых и наиболее распространенных включений для хранения углерода является гликоген , в котором единицы глюкозы связаны друг с другом в многоразветвленной полисахаридной структуре.
Другим распространенным способом хранения углерода бактериями является форма поли-β-гидроксибутирата (PHB) , гранулы, которые образуются, когда звенья β-гидроксимасляной кислоты объединяются вместе. Этот липид по составу очень похож на пластик, что побудило некоторых ученых исследовать возможность его использования в качестве биоразлагаемого пластика. Гранулы ПОБ фактически имеют оболочку, состоящую как из белка, так и из небольшого количества фосфолипидов. И гликоген, и ПОБ образуются при избытке углерода, а затем расщепляются клеткой позже как на углерод, так и на энергию.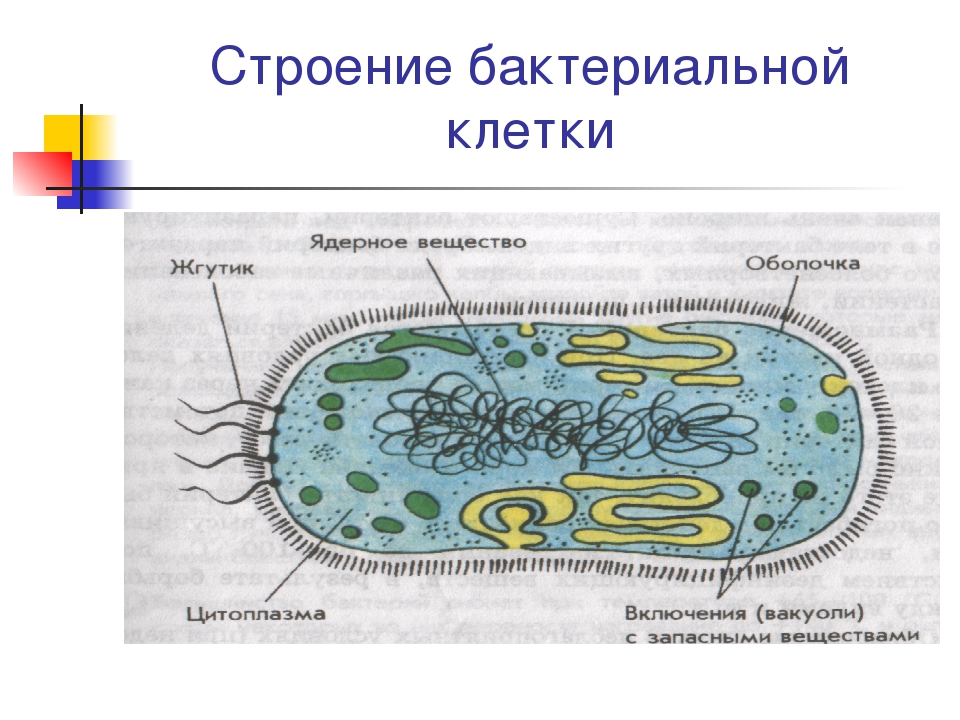
Неорганическое хранение
Часто бактериям требуется что-то другое, кроме углерода, либо для синтеза клеточных компонентов, либо в качестве альтернативного источника энергии. Гранулы полифосфата позволяют накапливать неорганический фосфат (PO43-), где фосфат может быть использован для производства нуклеиновой кислоты (помните сахар- фосфат основу?) или АТФ (конечно, аденозинтри фосфат ).
Другие клетки нуждаются в сере в качестве источника электронов для своего метаболизма и будут хранить избыток серы в виде глобул серы , которые образуются, когда клетка окисляет сероводород (h3S) до элементарной серы (S0), что приводит к образованию преломляющие включения.
Функции без сохранения
Бывают случаи, когда бактерия должна делать что-то помимо простого хранения органических или неорганических соединений для использования в метаболизме, и существуют включения, которые помогают с этими функциями, не связанными с хранением.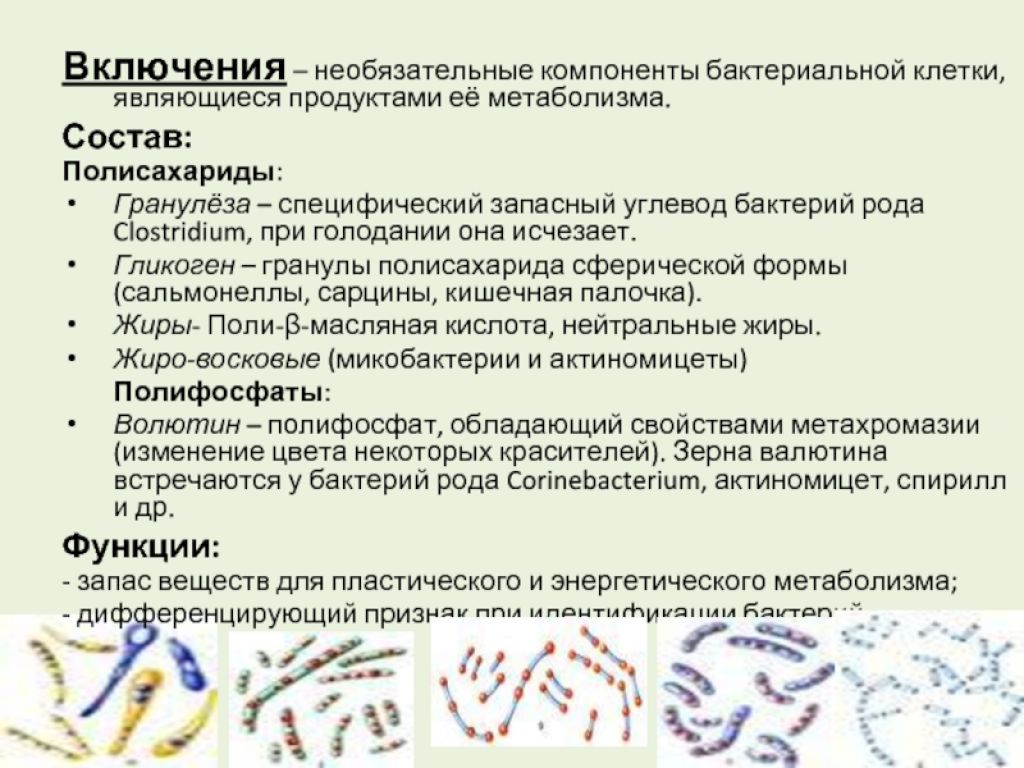 Одним из таких примеров являются газовые вакуоли , которые используются клеткой для управления плавучестью в толще воды, предоставляя клетке некоторый контроль над тем, где она находится в окружающей среде. Это ограниченная форма подвижности, только по вертикальной оси.Газовые вакуоли состоят из конгломератов газовых пузырьков , цилиндрических структур, полых и жестких. Газовые везикулы свободно проницаемы для всех типов газов за счет пассивной диффузии и могут быстро образовываться или схлопываться в зависимости от того, что необходимо клетке для подъема или опускания.
Одним из таких примеров являются газовые вакуоли , которые используются клеткой для управления плавучестью в толще воды, предоставляя клетке некоторый контроль над тем, где она находится в окружающей среде. Это ограниченная форма подвижности, только по вертикальной оси.Газовые вакуоли состоят из конгломератов газовых пузырьков , цилиндрических структур, полых и жестких. Газовые везикулы свободно проницаемы для всех типов газов за счет пассивной диффузии и могут быстро образовываться или схлопываться в зависимости от того, что необходимо клетке для подъема или опускания.
Магнитосомы представляют собой включения, содержащие длинные цепочки магнетита (Fe3O4), которые используются клеткой в качестве компаса в геомагнитных полях для ориентации в окружающей среде.Магнитотактические бактерии обычно микроаэрофильные , предпочитающие среду с более низким уровнем кислорода, чем атмосфера. Магенетосома позволяет клеткам найти оптимальную глубину для своего роста.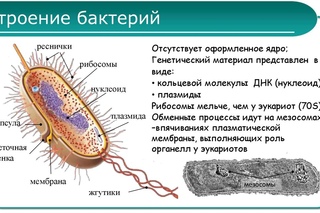 Магенетосомы имеют настоящий липидный бислой, напоминающий эукариотические органеллы, но на самом деле это инвагинация клеточной плазматической мембраны, модифицированная специфическими белками.
Магенетосомы имеют настоящий липидный бислой, напоминающий эукариотические органеллы, но на самом деле это инвагинация клеточной плазматической мембраны, модифицированная специфическими белками.
Микроотсеки
Бактериальные микрокомпартменты (BMC) отличаются от других включений своей структурой и функциональностью.Они имеют икосаэдрическую форму и состоят из белковой оболочки, состоящей из различных белков семейства BMC. Хотя их точная роль варьируется, все они участвуют в функциях, выходящих за рамки простого хранения веществ. Эти компартменты обеспечивают как местонахождение, так и вещества (обычно ферменты), необходимые для определенных метаболических процессов.
Наиболее изученным примером BMC является карбоксисома , которая обнаружена у многих бактерий, фиксирующих CO2. Карбоксисомы содержат фермент рибулозо-1,5-бисфосфаткарбоксилазу (к счастью, он также известен как RubisCO ), который играет решающую роль в превращении CO2 в сахар. Карбоксисома также играет роль в концентрации СО2, гарантируя тем самым, что все компоненты, необходимые для фиксации СО2, находятся в одном и том же месте в одно и то же время.
Карбоксисома также играет роль в концентрации СО2, гарантируя тем самым, что все компоненты, необходимые для фиксации СО2, находятся в одном и том же месте в одно и то же время.
Анаммоксосома
Анаммоксосома представляет собой большой мембранный компартмент, обнаруживаемый в бактериальных клетках, способных к проведению аннамокс-реакции ( аэробной ам мониевой окс идации), где аммоний (Nh5+) и нитрит (NO2- ) превращаются в газообразный азот (N2).Процесс осуществляется как способ получения клеткой энергии с использованием аммония в качестве донора электронов и нитрита в качестве акцептора электронов, в результате чего образуется газообразный азот. Это химическое превращение азота важно для круговорота азота.
Цикл азота. Шоу-Цин Ни и Цзянь Чжан [CC BY 3.0], Wikimedia Commons
Хлоросома
Обнаруженная у некоторых фототрофных бактерий хлоросома представляет собой высокоэффективную структуру для захвата низкой интенсивности света. Выстилая внутренний периметр клеточной мембраны, каждая хлоросома может содержать до 250 000 молекул бактериохлорофилла, расположенных плотными массивами. Собранный свет передается в реакционные центры в клеточной мембране, обеспечивая преобразование световой энергии в химическую энергию в форме АТФ. Хлоросома ограничена липидным монослоем.
Выстилая внутренний периметр клеточной мембраны, каждая хлоросома может содержать до 250 000 молекул бактериохлорофилла, расположенных плотными массивами. Собранный свет передается в реакционные центры в клеточной мембране, обеспечивая преобразование световой энергии в химическую энергию в форме АТФ. Хлоросома ограничена липидным монослоем.
Плазмида
Плазмида представляет собой внехромосомный фрагмент ДНК, который есть у некоторых бактерий в дополнение к генетическому материалу, обнаруженному в нуклеоиде.Он состоит из двухцепочечной ДНК и обычно имеет кольцевую форму, хотя были обнаружены линейные плазмиды. Плазмиды описываются как «несущественные» для клетки, где клетка может нормально функционировать в их отсутствие. Но хотя плазмиды имеют всего несколько генов, они могут придавать клетке важные свойства, например устойчивость к антибиотикам. Плазмиды реплицируются независимо от клетки и могут быть потеряны (известно как , излечивающий ) либо спонтанно, либо из-за воздействия неблагоприятных условий, таких как УФ-свет, тиминовое голодание или рост выше оптимальных условий.![]() Некоторые плазмиды, известные как эписомы , могут быть интегрированы в клеточную хромосому, где гены будут реплицироваться во время клеточного деления.
Некоторые плазмиды, известные как эписомы , могут быть интегрированы в клеточную хромосому, где гены будут реплицироваться во время клеточного деления.
Эндоспора
Еще есть эндоспора , чудо бактериальной инженерии. Это находится под заголовком «внутренние компоненты бактерий», но важно отметить, что эндоспора — это не внутренняя или внешняя структура, а превращение клетки в альтернативную форму. Клетки начинаются как вегетативные клетки, делая все, что должна делать клетка (метаболизм, размножение, стрижка газона…).Если они подвергаются воздействию враждебных условий (иссушение, высокая температура, сердитый сосед…) и , у них есть способность, они могут превратиться из вегетативной клетки в эндоспору. Эндоспора на самом деле образуется внутри вегетативной клетки (разве это не делает ее внутренней структурой?), а затем вегетативная клетка лизируется, высвобождая эндоспору (делает ли это внешнюю структуру?).
Слои эндоспор.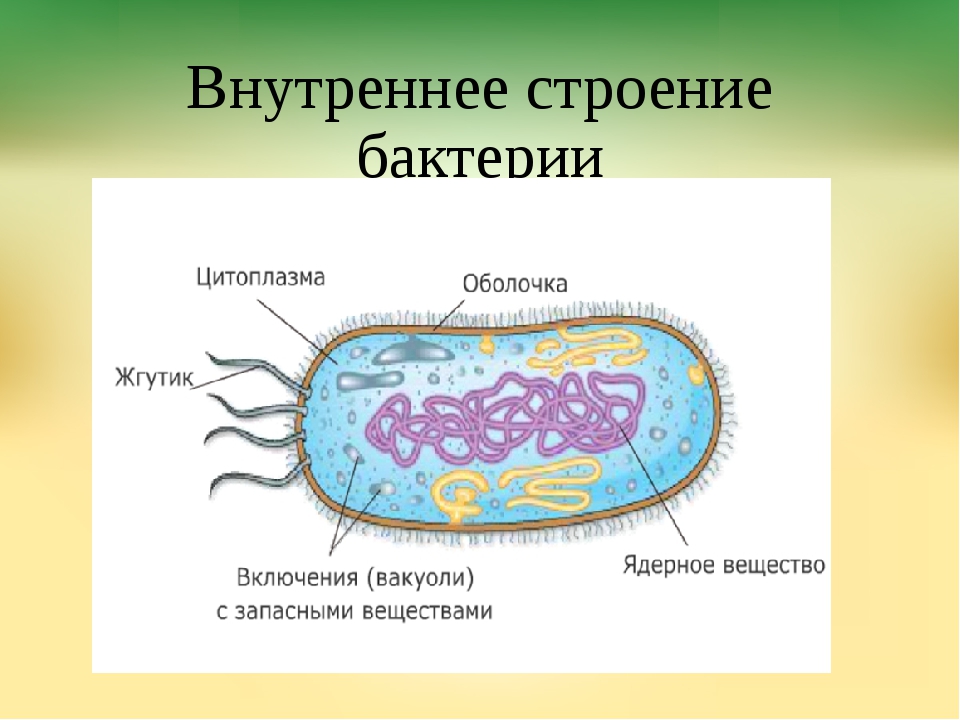
Эндоспоры образуются только у нескольких грамположительных родов и обеспечивают клетке устойчивость к широкому спектру суровых условий, таких как голодание, экстремальные температуры, воздействие высыхания, ультрафиолетового света, химических веществ, ферментов и радиации.В то время как вегетативная клетка является активной формой бактериальных клеток (растущая, метаболизирующая и т. д.), эндоспору можно рассматривать как спящую форму клетки. Он позволяет выжить в неблагоприятных условиях, но не позволяет клетке расти или воспроизводиться.
Структура
Чтобы быть невероятно устойчивым к стольким различным веществам и условиям окружающей среды, необходимо много различных слоев. Бактериальная эндоспора имеет множество различных слоев, начиная с ядра в центре.Ядро — это местонахождение нуклеоида, рибосом и цитоплазмы клетки в крайне обезвоженном виде. Обычно он содержит только 25% воды, содержащейся в вегетативной клетке, что повышает термостойкость. ДНК дополнительно защищена присутствием малых кислоторастворимых белков (SASPs) , которые стабилизируют ДНК и защищают ее от деградации. Стабилизация ДНК усиливается за счет присутствия дипиколиновой кислоты в комплексе с кальцием ( Ca-DPA ), который встраивается между основаниями ДНК.Ядро обернуто внутренней мембраной , которая обеспечивает барьер проницаемости для химических веществ, который затем окружен корой , толстым слоем, состоящим из пептидогликана с меньшим количеством поперечных связей, чем в вегетативной клетке. Кора покрыта внешней мембраной . И, наконец, несколько споровых оболочек , сделанных из белка, которые обеспечивают защиту от воздействия окружающей среды, такого как химические вещества и ферменты.
ДНК дополнительно защищена присутствием малых кислоторастворимых белков (SASPs) , которые стабилизируют ДНК и защищают ее от деградации. Стабилизация ДНК усиливается за счет присутствия дипиколиновой кислоты в комплексе с кальцием ( Ca-DPA ), который встраивается между основаниями ДНК.Ядро обернуто внутренней мембраной , которая обеспечивает барьер проницаемости для химических веществ, который затем окружен корой , толстым слоем, состоящим из пептидогликана с меньшим количеством поперечных связей, чем в вегетативной клетке. Кора покрыта внешней мембраной . И, наконец, несколько споровых оболочек , сделанных из белка, которые обеспечивают защиту от воздействия окружающей среды, такого как химические вещества и ферменты.
Спороношение: превращение вегетативной клетки в эндоспору
Споруляция , превращение вегетативной клетки в эндоспору с высокой степенью защиты, обычно происходит, когда выживанию клетки каким-либо образом угрожает опасность. Фактический процесс очень сложен и обычно занимает несколько часов до завершения. Первоначально спорулирующие клетки реплицируют свою ДНК, как если бы они собирались подвергнуться клеточному делению. Перегородка формируется асимметрично, изолируя одну копию хромосомы на одном конце клетки (называемую предспорой ). Происходит синтез специфичных для эндоспор веществ, видоизменяющий предспору и приводящий к развитию слоев, специфичных для эндоспоры, а также к обезвоживанию. В конце концов «материнская клетка» лизируется, что позволяет высвобождать зрелую эндоспору в окружающую среду.
Фактический процесс очень сложен и обычно занимает несколько часов до завершения. Первоначально спорулирующие клетки реплицируют свою ДНК, как если бы они собирались подвергнуться клеточному делению. Перегородка формируется асимметрично, изолируя одну копию хромосомы на одном конце клетки (называемую предспорой ). Происходит синтез специфичных для эндоспор веществ, видоизменяющий предспору и приводящий к развитию слоев, специфичных для эндоспоры, а также к обезвоживанию. В конце концов «материнская клетка» лизируется, что позволяет высвобождать зрелую эндоспору в окружающую среду.
Спороношение.
Превращение эндоспоры в вегетативную клетку
Эндоспора остается бездействующей до тех пор, пока условия окружающей среды не улучшатся, вызывая химическое изменение, которое инициирует экспрессию генов. В превращении эндоспоры в метаболически активные вегетативные клетки есть три различных этапа: 1) активация , этап подготовки, который может быть инициирован применением тепла; 2) прорастание , когда эндоспора становится метаболически активной и начинает поглощать воду; 3) вырост , когда вегетативная клетка полностью выходит из оболочки эндоспоры.
Ключевые слова
цитоскелет, FtsZ, тубулин, MreB, актин, кресцентин, ламин, кератин, включение, гликоген, поли-β-гидроксибутират (ПГБ), гранулы полифосфата, глобулы серы, газовые вакуоли, газовые везикулы, магнитосомы, микроаэрофильные, микрокомпартменты, бактериальные микрокомпартменты (BMCs), карбоксисома, рибулозо-1,5-бисфосфаткарбоксилаза, RubisCO, анаммоксосома, аннамокс-реакция, хлоросома, плазмида, отверждение, эписома, эндоспора, вегетативная клетка, ядро, малые кислоторастворимые белки (SASP), дипиколиновая кислота, Ca -DPA, внутренняя мембрана, кора, наружная оболочка, споровая оболочка, спороношение, предспора, активация, прорастание, отросток.
Основные вопросы/цели
- Каковы функции и состав бактериального цитоскелета? Чем он отличается от цитоскелета эукариот? Каковы конкретные белки бактериального цитоскелета и какие подробности известны о каждом из них?
- Каково назначение включений, обнаруживаемых в бактериях? Каковы их характеристики?
- Каковы конкретные примеры запасающих включений, обнаруженных у бактерий? Уметь описать каждый тип с точки зрения структуры и назначения.

- Какие еще включения встречаются у бактерий? Уметь описать каждый тип с точки зрения структуры и назначения.
- Чем микрокомпартменты отличаются от включений? Какие конкретные примеры? Каков состав и назначение?
- Что такое анаммоксосомы? Каков их состав и назначение?
- Что такое плазмиды и какими характеристиками они обладают? Что такое эписомы? Что лечит и что его вызывает?
- Что такое бактериальные эндоспоры? Какова их цель? Какие характеристики у них есть? Каковы различные слои эндоспоры и какую роль играет каждый слой?
Исследовательские вопросы (НЕОБЯЗАТЕЛЬНО)
- Какие бактериальные структуры могут быть полезны ученым при решении социальных проблем?
Бактериальная цитоплазма как эффективный клеточный компартмент для получения функциональных аффинных реагентов на основе VHH и рекомбинантных антител, подобных IgG Camelidae | Фабрики микробных клеток
Bradbury ARM, Sidhu S, Dübel S, McCafferty J: Помимо естественных антител: мощь технологий отображения in vitro . Nat Biotechnol 2011, 29:245-254.
Артикул Google Scholar
de Marco A: Методики выделения альтернативных связующих веществ с улучшенными клиническими возможностями по сравнению с обычными антителами. Критик Rev Biotech. 2013, 33: 40-48. 10.3109/07388551.2012.665353.
Артикул Google Scholar
Abulrob A, Sprong H, Van Bergen en Henegouwen P, Stanimirovic D: Трансмигрирующее однодоменное антитело через гематоэнцефалический барьер: механизмы транспорта и антигенные эпитопы в эндотелиальных клетках головного мозга человека. Дж. Нейрохим. 2005, 95: 1201-1214. 10.1111/j.1471-4159.2005.03463.х.
Артикул Google Scholar
Vaneycken I, D’Huyvetter M, Hernot S, De Vos J, Xavier C, Devoogdt N, Caveliers V, Lahoutte T: Immuno-imaging с использованием нанотел. Curr Op Biotechnol. 2011, 22: 1-5. 10.1016/j.copbio.2011.06.009.
Curr Op Biotechnol. 2011, 22: 1-5. 10.1016/j.copbio.2011.06.009.
Артикул Google Scholar
Oliveira S, van Dongen GA, Stigter-van Walsum M, Roovers RC, Stam JC, Mali W, van Diest PJ, van Bergen en Henegouwen PM: Быстрая визуализация ксенотрансплантатов опухоли человека с помощью оптической визуализации с ближним — инфракрасное флуоресцентное нанотело рецептора антиэпидермального фактора роста. Мол изображения. 2012, 11: 33-46.
Google Scholar
Палмер Д.Б., Джордж А.Дж., Риттер М.А.: Селекция антител к детерминантам клеточной поверхности на эпителиальных клетках тимуса мыши с использованием библиотеки фагового дисплея. Иммунология. 1997, 91: 473-478. 10.1046/j.1365-2567.1997.00262.х.
Артикул Google Scholar
de Marco A: Выбор подходящих меток повышает эффективность применения выбранных связующих веществ: создание удобных для пользователя экспрессионных плазмид. Мет Мол Биол.2012, 911: 507-522. 10.1007/978-1-61779-968-6_32.
Мет Мол Биол.2012, 911: 507-522. 10.1007/978-1-61779-968-6_32.
Артикул Google Scholar
Mazor Y, Van Blarcom T, Iverson BL, Georgiou G: E-клональные антитела: отбор полноразмерных IgG-антител с использованием бактериального периплазматического дисплея. Нат Проток. 2008, 3: 1766-1777. 10.1038/нпрот.2008.176.
Артикул Google Scholar
Де Марни М., Монегал А., Вентурини С., Винати С., Карбоне Р., де Марко А. Микроматрицы, независимые от очистки антител (PIM), путем прямого нанесения бактерий на предметные стекла, обработанные TiO 2 . Методы 2012, 56:317-325.
Артикул Google Scholar
Arbabi-Ghahroudi M, Tanha J, MacKenzie R: Прокариотическая экспрессия антител. Метастаз рака Rev. 2005, 24: 501-519. 10.1007/s10555-005-6193-1.
Артикул Google Scholar
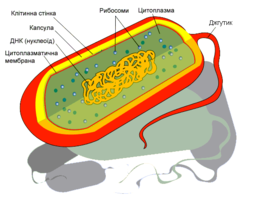
de Marco A: Стратегии успешной рекомбинантной экспрессии белков, зависимых от дисульфидных связей, в Escherichia coli . Факты о микробных клетках 2009, 8:26.
Артикул Google Scholar
Katsuda T, Sonoda H, Kumada Y, Yamaji H: Производство фрагментов антител в Escherichia coli . Methods Mol Biol 2012, 907:305-324.
Артикул Google Scholar
Заршлер К., Витеси С., Капплюш Ф., Ферстер С., Стефан Х.: Высокопродуктивная продукция функциональных растворимых однодоменных антител в цитоплазме Escherichia coli . Факты о микробных клетках 2013, 12:97.
Артикул Google Scholar
Simmons LC, Reilly D, Klimowski L, Raju S, Meng G, Sims P, Hong S, Shields RL, Damico LA, Rancatore P, Yansura DG: Экспрессия полноразмерных иммуноглобулинов в Escherichia coli : быстрое и эффективное производство агликозилированных антител.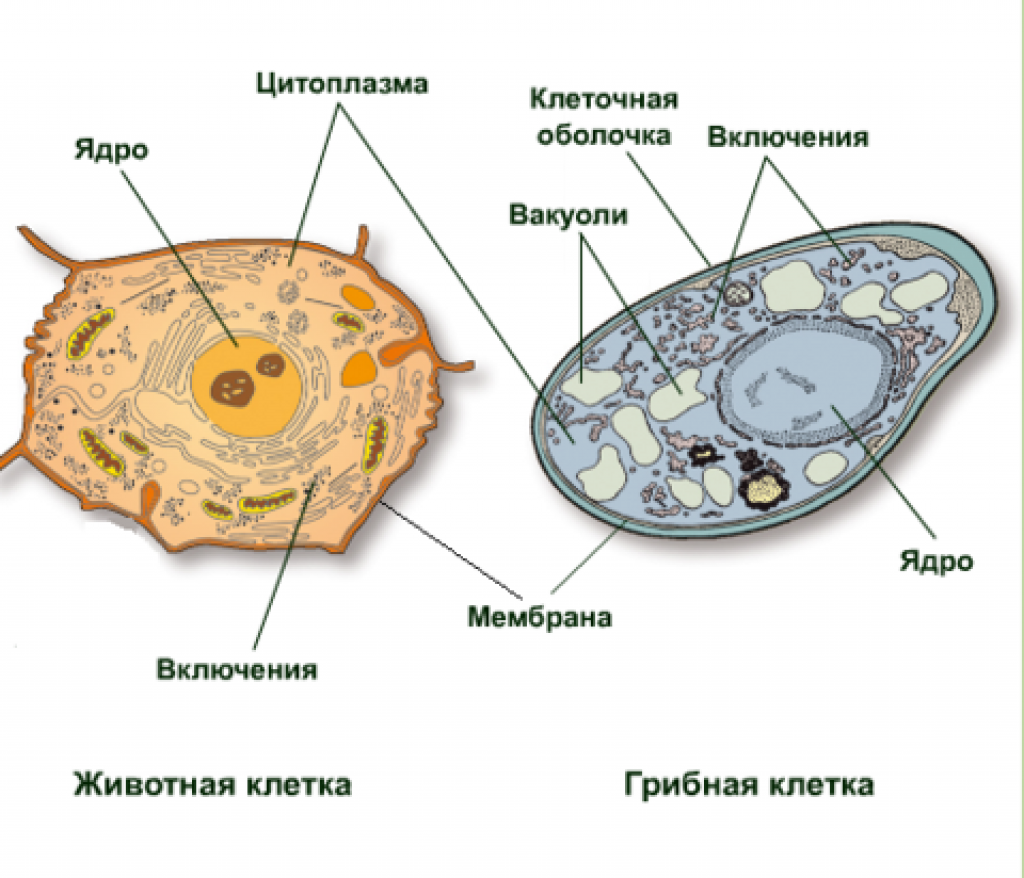 J Immunol Meth 2002, 263:133-147.
J Immunol Meth 2002, 263:133-147.
Артикул Google Scholar
Makino T, Skretas G, Kang T-H, Georgiou G: Комплексная разработка Escherichia coli для усиления экспрессии антител IgG. Metab Eng 2011, 13:241-251.
Артикул Google Scholar
Nguyen VD, Hatahet F, Salo KE, Enlund E, Zhang C, Ruddock LW: предварительная экспрессия сульфгидрилоксидазы значительно увеличивает выход эукариотических белков, содержащих дисульфидные связи, экспрессируемых в цитоплазме E.палочка . Microb Cell Fact 2011, 10:1-13.
Артикул Google Scholar
Veggiani G, de Marco A: Улучшенное количественное и качественное производство однодоменных интрател, опосредованное совместной экспрессией сульфгидрилоксидазы Erv1p. Prot Expr Purif. 2011, 79: 111-114. 10.1016/j.pep. 2011.03.005.
2011.03.005.
Артикул Google Scholar
de Marco A: Недавний вклад в области рекомбинантной экспрессии дисульфидно-связанного белка в бактериях.Факт микробной клетки. 2012, 11: 129-10.1186/1475-2859-11-129.
Артикул Google Scholar
Matos CF, Branston SD, Albiniak A, Dhanoya A, Freedman RB, Keshavarz-Moore E, Robinson C: Высокопродуктивный экспорт нативного гетерологичного белка в периплазму путем транслокации tat у Escherichia coli . Biotechnol Bioeng 2012, 109:2533-2542.
Артикул Google Scholar
Аронсон Д.Е., Костантини Л.М., Снапп Э.Л.: Superfolder GFP флуоресцирует в окисляющих средах, если на него нацелить через транслокон Sec. Движение. 2011, 12: 543-548. 10.1111/j.1600-0854.2011.01168.х.
Артикул Google Scholar
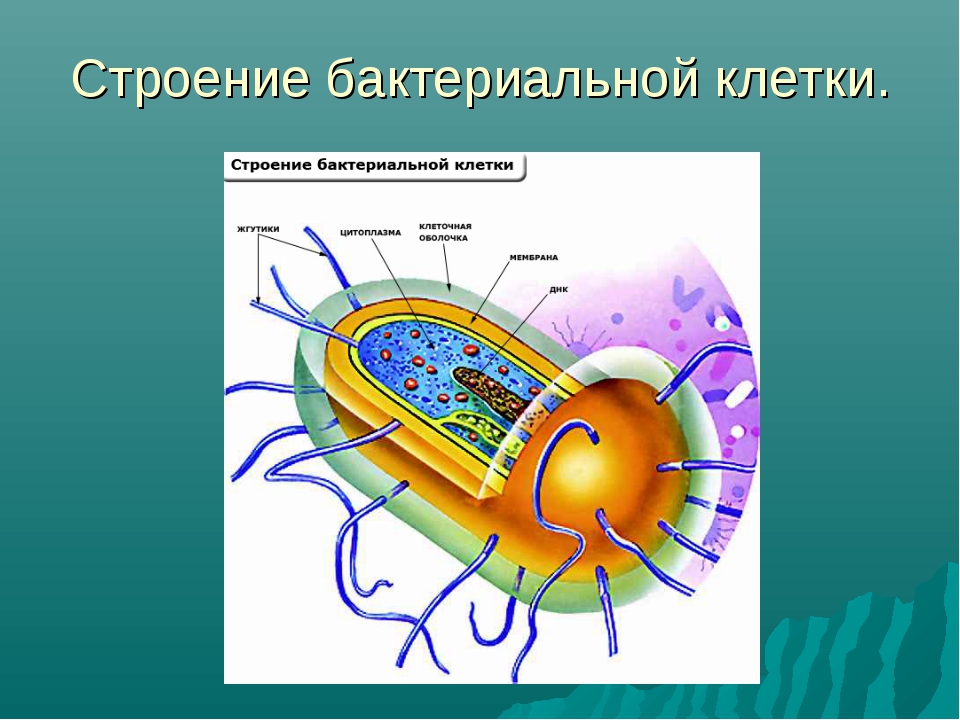
Dammeyer T, Timmis KN, Tinnefeld P: Векторы широкого круга хозяев для экспрессии белков с (Twin-)Strep-меткой, His-меткой и сконструированным, оптимизированным для экспорта желтым флуоресцентным белком. Факт микробной клетки.2013, 12: 49-10.1186/1475-2859-12-49.
Артикул Google Scholar
Mazmanian SK, Liu G, Ton-That H, Schneewind O: Staphylococcus aureus сортаза: фермент, который прикрепляет поверхностные белки к клеточной стенке. Наука 1999, 285:760-763.
Артикул Google Scholar
Мао Х., Харт С.А., Шинк А., Поллок Б.А. Лигирование белков, опосредованное сортазой: новый метод инженерии белков.J Am Chem Soc. 2004, 126: 2670-2671. 10.1021/ja039915e.
Артикул Google Scholar
Ta HT, Prabhu S, Leitner E, Jia F, von Elverfeldt D, Jackson KE, Heidt T, Nair AK, Pearce H, von Zur Muhlen C, Wang X, Peter K, Hagemeyer CE: Enzymatic single Мечение цепями антител: универсальный подход к целенаправленной молекулярной визуализации и поиску клеток при сердечно-сосудистых заболеваниях. Цирк рез. 2011, 109: 365-373. 10.1161/ЦИРКРЕСАХА.111.249375.
Цирк рез. 2011, 109: 365-373. 10.1161/ЦИРКРЕСАХА.111.249375.
Артикул Google Scholar
Botosoa EP, Maillasson M, Mougin-Degraef M, Remaud-Le Saëc P, Gestin JF, Jacques Y, Barbet J, Faivre-Chauvet A: Распознавание антитело-гаптена на поверхности функционализированных липосом, изученное с помощью SPR : Стерические затруднения пегилированных фосфолипидов в липосомах-невидимках, приготовленных для адресной доставки радионуклидов. Джей Друг Делив. 2011, 2011: 368535-10.1155/2011/368535.
Артикул Google Scholar
Trilling AK, Harmsen MM, Ruigrok VJ, Zuilhof H, Beekwilder J: Влияние однородной ориентации молекул захвата на чувствительность биосенсора: зависимость от свойств аналита. Биосенс Биоэлектрон. 2013, 40: 219-226. 10.1016/j.bios.2012.07.027.
Артикул Google Scholar
Gurszka A, Martinelli C, Sparacio E, Pelicci PG, de Marco A: одновременное использование N- и C-концевых антител против нуклеофосмина 1 в иммунофлуоресцентных экспериментах позволяет точно оценить его субклеточную локализацию при остром больных миелоидным лейкозом. Лейкемия. 2012, 26: 159-163. 10.1038/лей.2011.177.
Лейкемия. 2012, 26: 159-163. 10.1038/лей.2011.177.
Артикул Google Scholar
Парра-Палау Дж.Л., Педерсен К., Пег В., Скалтрити М., Ангелини П.Д., Эскорихуэла М., Мансилла С., Санчес Пла А., Рамони Чьял С., Басельга Дж., Аррибас Дж.: главная роль p95/611 -CTF, карбоксиконцевой фрагмент HER2, в понижающей модуляции рецептора эстрогена при HER2-положительном раке молочной железы. Рак рез. 2010, 70: 8537-8546. 10.1158/0008-5472.CAN-10-1701.
Артикул Google Scholar
M#x00FC;ller KM, Arndt KM, Pl#x00FC;ckthun A: Модель и имитация мультивалентного связывания с фиксированными лигандами. Анальная биохимия. 1998, 261: 149-158. 10.1006/abio.1998.2725.
Артикул Google Scholar
Kijanka M, Warnders FJ, El Khattabi M, Lub de Hooge M, van Dam GM, Ntziachristos V, de Vries L, Oliveira S, van Bergen en Henegouwen PMP: быстрая оптическая визуализация ксенотрансплантатов опухоли молочной железы человека с использованием анти-HER2 VHH, сайт-прямая конъюгация с IRDye800CW для хирургии под визуальным контролем. Eur J Nucl Med Mol Imaging. 2013, 40: 1718-1729. 10.1007/s00259-013-2471-2.
Eur J Nucl Med Mol Imaging. 2013, 40: 1718-1729. 10.1007/s00259-013-2471-2.
Артикул Google Scholar
Jörgensen ML, Friis NA, Just J, Madsen P, Petersen SV, Kristensen P: Экспрессия одноцепочечных вариабельных фрагментов, слитых с Fc-областью кроличьего IgG в Leishmania tarantolae . Microb Cell Fact 2014, 13:9.
Артикул Google Scholar
Лебендикер М., Даниэли Т., де Марко А. Путеводитель Trip Adviser по миру белковой науки; предложение повысить осведомленность о качестве рекомбинантных белков. Примечания BMC Res. 2014, 7: 585-10.1186/1756-0500-7-585.
Артикул Google Scholar
Hatalet F, Nguyen VD, Salo KE, Ruddock LW: Нарушение восстановительных путей не является существенным для эффективного образования дисульфидных связей в цитоплазме E. coli.Факт микробной клетки. 2010, 9: 67-
2010, 9: 67-
Google Scholar
Djender S, Beugnet A, Schneider A, de Marco A: Биотехнологические применения рекомбинантных однодоменных антител оптимизируются путем слияния C-конца с последовательностью EPEA (C-метка). Антитела. 2014, 3: 182-191. 10.3390/антиб3020182.
Артикул Google Scholar
Schlegel S, Rujas E, Ytteberg AJ, Zubarev RA, Luirink J, de Gier J-W: Оптимизация продукции гетерологичных белков в периплазме E.coli, регулируя уровни экспрессии генов. Факт микробной клетки. 2013, 12: 24-10.1186/1475-2859-12-24.
Артикул Google Scholar
Де Мейер Т., Мюйлдерманс С., Депикер А.: Продукты на основе нанотел как средства исследования и диагностики. Тенденции биотехнологии. 2014, 32: 263-270. 10.1016/j.tibtech.2014.03.001.
Артикул Google Scholar
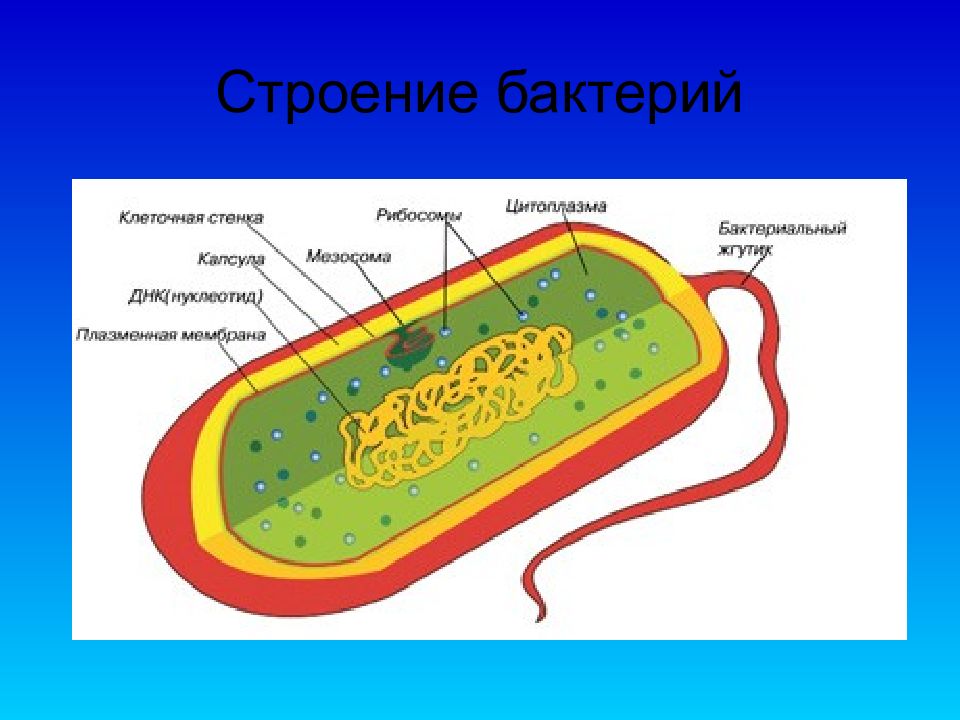
Ponsel D, Neugebauer J, Ladetzki-Baehs K, Tissot K: Высокая аффинность, возможность разработки и функциональный размер: Святой Грааль создания комбинаторной библиотеки антител.Молекулы. 2011, 16: 3675-3700. 10,3390/молекул 16053675.
Артикул Google Scholar
Ollis AA, Zhang S, Fisher AC, DeLisa MP: Сконструированные олигосахарилтрансферазы со значительно ослабленной специфичностью акцепторного сайта. Nat Chem Biol 2014, doi: 10.1038/nchembio.1609.
Google Scholar
Moutel S, El Marjou A, Vielemeyer O, Nizak C, Benaroch P, Dübel , Perez F: Многовидовая система Fc для производства рекомбинантных антител.БМС Биотехнология. 2009, 9: 14-10.1186/1472-6750-9-14.
Артикул Google Scholar
Монегал А., Ами Д., Мартинелли С., Хуанг Х., Алипранди М., Капассо П., Франкавилла С., Оссоленго Г. , де Марко А.: Иммунологические применения однодоменных рекомбинантных антител ламы, выделенных из наивной библиотеки. Заместитель инженера Дес Сел. 2009, 22: 273-280. 10.1093/белок/gzp002.
, де Марко А.: Иммунологические применения однодоменных рекомбинантных антител ламы, выделенных из наивной библиотеки. Заместитель инженера Дес Сел. 2009, 22: 273-280. 10.1093/белок/gzp002.
Артикул Google Scholar
Sala E, de Marco A: Скрининг оптимизированных протоколов очистки белков путем сочетания маломасштабной экспрессии и эксклюзионной хроматографии малого размера. Prot Expr Purif. 2010, 74: 231-235. 10.1016/j.pep.2010.05.014.
Артикул Google Scholar
Физика бактериальной цитоплазмы и ее влияние на клеточную физиологию
Область проекта, которую мы описываем ниже, находится на стыке экспериментальной работы в лабораторных условиях и математического моделирования.
Физические свойства бактерий до сих пор плохо изучены, несмотря на их важность для клеточных процессов. Обычно считается, что цитоплазма, в которой происходит большинство клеточных процессов, ведет себя как простая (вязкая) жидкость. Совместная работа лабораторий Джейкобса-Вагнера, Дюфресна и О’Херна свидетельствует об обратном.
Совместная работа лабораторий Джейкобса-Вагнера, Дюфресна и О’Херна свидетельствует об обратном.
Мы обнаружили, что бактериальная цитоплазма ведет себя как коллоидная взвесь на пороге стеклования. Стекловидные материалы характеризуются динамической остановкой и могут быть псевдоожижены при относительно незначительных возмущениях, роль, которую, по-видимому, выполняет клеточный метаболизм в бактериальной цитоплазме.
Эти открытия открывают новые области исследований, поскольку они затрагивают множество интересных биологических и физических вопросов. С физической точки зрения коллоидные стекла представляют собой интересные материалы, но обычно их изучают с использованием монодисперсных частиц. Напротив, бактериальная цитоплазма представляет собой высоко полидисперсный материал (размеры ее компонентов варьируются более чем на 3 порядка), и мы ожидаем, что дальнейшая работа над бактериальной цитоплазмой проинформирует нас о стеклообразной динамике в сложных средах.С биологической точки зрения мы ожидаем, что стеклообразные свойства будут иметь широкое значение для физиологии бактерий.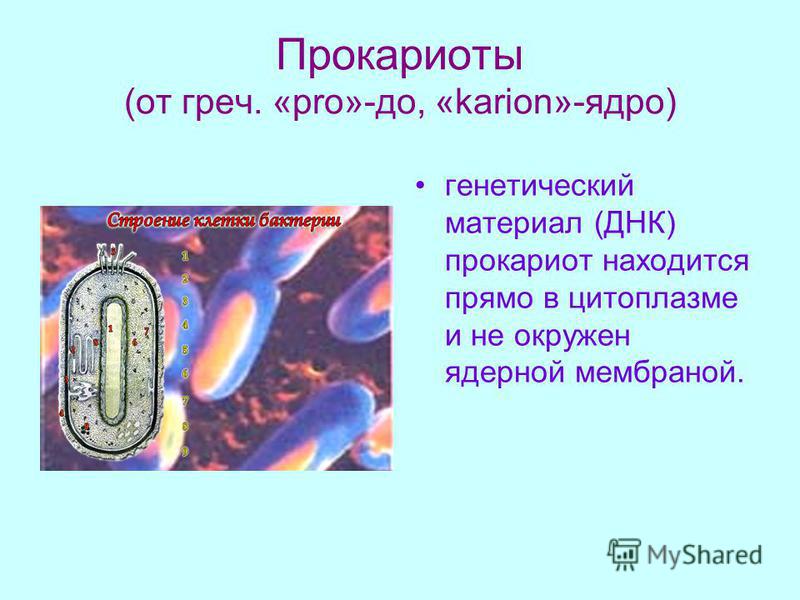
Например, метаболический покой является распространенным состоянием бактерий в природе (включая клинические условия), и мы исследуем, как бактерии могли использовать стекловидные свойства в ходе эволюции, чтобы противостоять стрессовым условиям, таким как голодание и лечение антибиотиками. Конкретные проекты будут включать эксперименты, моделирование или и то, и другое в зависимости от интересов учащегося.
Изображение выше: Бактериальная цитоплазма демонстрирует резко различающиеся физические свойства в зависимости от метаболического состояния клетки, о чем свидетельствует удержание зонда в неактивных клетках (красная траектория на изображении выше) и исследование зондом цитоплазматического пространства (зеленая траектория). ). В этом эксперименте зонды (частицы GFP-µNS) отслеживались с частотой кадров 1,3 с в течение 650 с.
Изображение выше: Отслеживание отдельных частиц в генетически идентичном E.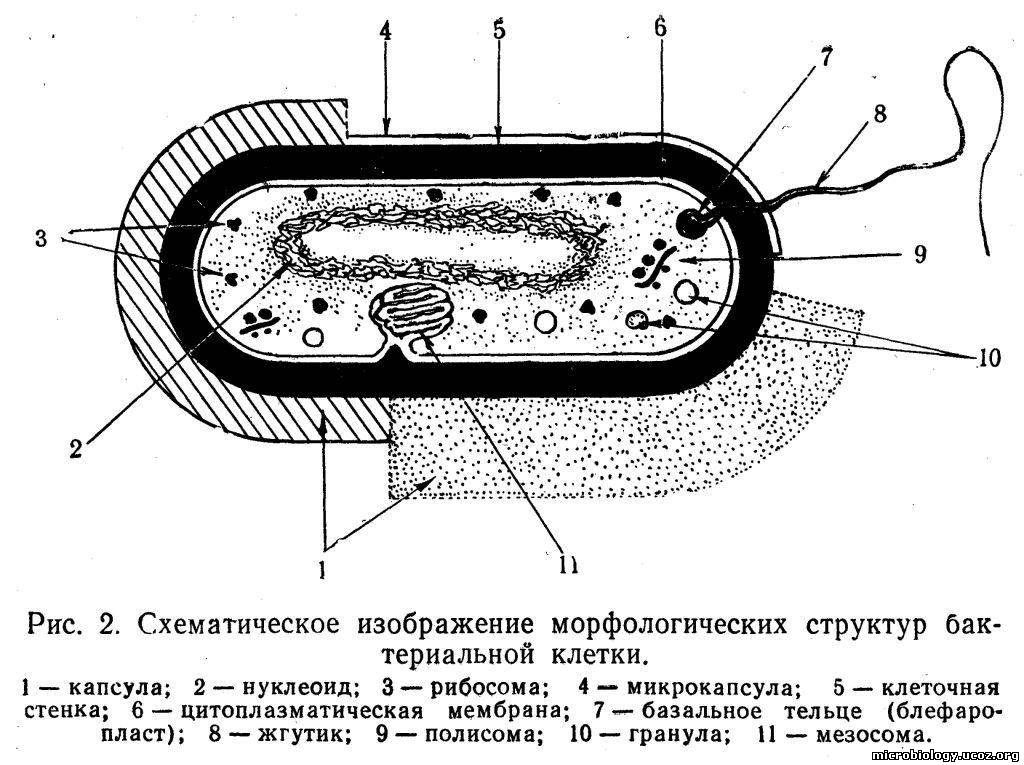 coli , растущие в тех же условиях, обнаруживают удивительно большую неоднородность в движении частиц, как показано на приведенных выше среднеквадратических смещениях (СКО) одной траектории. Мы заинтересованы в понимании происхождения этой гетерогенности и ее роли в биологии (например, фенотипическая межклеточная изменчивость).
coli , растущие в тех же условиях, обнаруживают удивительно большую неоднородность в движении частиц, как показано на приведенных выше среднеквадратических смещениях (СКО) одной траектории. Мы заинтересованы в понимании происхождения этой гетерогенности и ее роли в биологии (например, фенотипическая межклеточная изменчивость).
Изображение выше: Мы разрабатываем модели бактериальной цитоплазмы. На рисунке выше показаны три смоделированные траектории частицы, диффундирующей через цитоплазму E.палочка . Клетка была смоделирована в трех измерениях с твердой мембраной и непроницаемой внутренней сфероцилиндрической областью, которая представляет собой область ДНК (нуклеоид) клетки.
Изображение выше: Мы также используем моделирование, чтобы понять влияние скученности и полидисперсности в цитоплазме. На изображении выше показан пример упаковки 1000 сферических частиц двух разных размеров.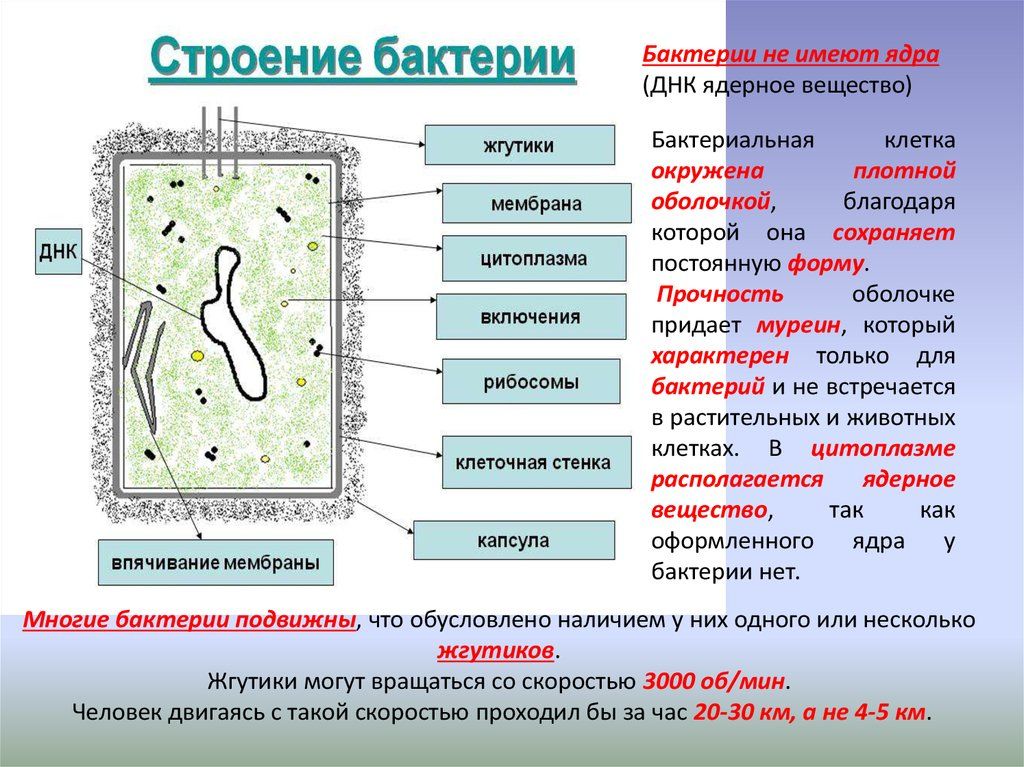
Сотрудничество в области исследований раскрывает структуру и функцию PspA и обнаруживает эволюционную связь бактериального белка с белками ESCRT-III человека — ScienceDaily
Клетки простых организмов, таких как бактерии, а также клетки человека окружены мембраной, которая выполняет различные задачи, в том числе защищает клетку от стресса.В рамках совместного проекта группы из Университета им. Иоганна Гутенберга в Майнце (JGU) и Исследовательского центра в Юлихе при участии Дюссельдорфского университета имени Генриха Гейне (HHU) обнаружили, что мембранный белок, обнаруженный в бактериях, имеет сходную структуру и функцию с группой белков. которые отвечают за ремоделирование и восстановление клеточной мембраны у человека. Ранее о связи между двумя группами белков не было известно. Исследовательская работа команды была недавно опубликована в журнале Cell .
PspA играет ключевую роль в реакции бактериального стресса
Система белков фагового шока (Psp) была обнаружена у бактерий примерно 30 лет назад.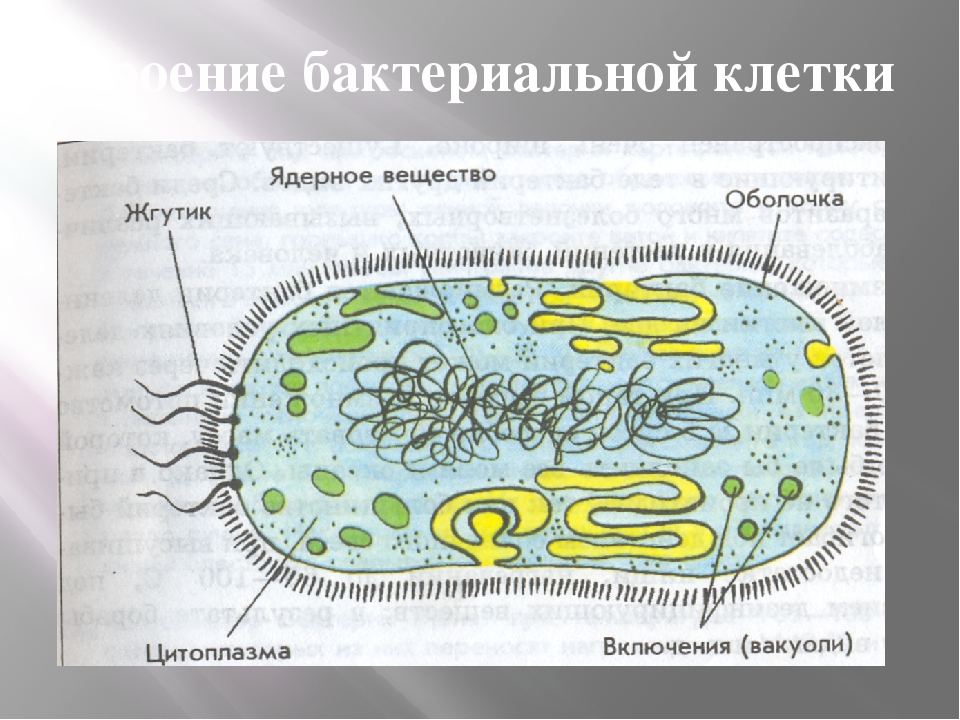 В то время было установлено, что это реакция бактерий Escherichia coli на заражение особыми вирусами, называемыми бактериофагами. Позже стало ясно, что его функция по защите клеточной мембраны превосходит специфический ответ на бактериофаговую инфекцию. Осмотический стресс, тепло, клеточные токсины или дефекты мембранной оболочки также могут вызвать стрессовую реакцию.
В то время было установлено, что это реакция бактерий Escherichia coli на заражение особыми вирусами, называемыми бактериофагами. Позже стало ясно, что его функция по защите клеточной мембраны превосходит специфический ответ на бактериофаговую инфекцию. Осмотический стресс, тепло, клеточные токсины или дефекты мембранной оболочки также могут вызвать стрессовую реакцию.
«Сегодня мы знаем, что система Psp активируется в ответ на многочисленные типы мембранного стресса. Однако некоторые молекулярные детали все еще остаются загадочными», — объяснил профессор Дирк Шнайдер, глава группы мембранных белков в JGU. «Вот почему мы решили поближе изучить основные белки системы Psp». Вместе со своей командой он недавно обнаружил, как представитель Psp IM30 образует защитную коврообразную структуру на клеточной мембране, чтобы справиться с мембранным стрессом.
В своей новой работе ученые тщательно изучили фаговый шоковый белок А (PspA), играющий ключевую роль в системе Psp. В частности, с помощью криоэлектронной микроскопии стало видно, как PspA образует длинные спиралевидные трубки, которые могут окружать биомембрану во внутренней полости.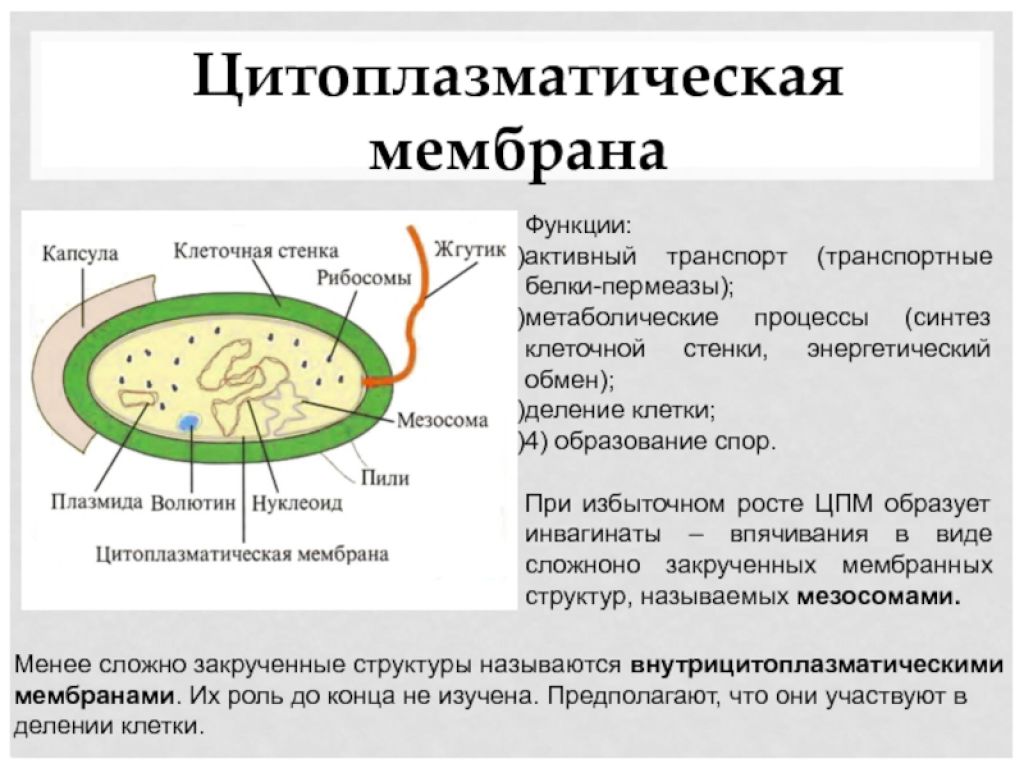 Изображения с высоким разрешением теперь впервые показывают, как PspA локально растворяет отдельные мембраны, а затем преобразует их в более крупные единицы или даже способствует формированию новых мембранных структур.
Изображения с высоким разрешением теперь впервые показывают, как PspA локально растворяет отдельные мембраны, а затем преобразует их в более крупные единицы или даже способствует формированию новых мембранных структур.
«Тысячи строительных блоков PspA могут быть собраны в большие спиральные структуры. Поэтому они являются идеальным объектом исследования для нашего криоэлектронно-микроскопического структурного анализа», — сказал профессор Карстен Саксе из Forschungszentrum Jülich и HHU Düsseldorf. Исследования проводились в Юлихе вместе с доктором Бенедиктом Юнгласом, бывшим докторантом профессора Дирка Шнайдера в JGU. В Центре микроскопии и спектроскопии с использованием электронов имени Эрнста Руска (ER-C) Forschungszentrum Jülich используются одни из самых мощных электронных микроскопов в Европе, а в последнее время — криомикроскопы для изучения быстрозамороженных биологических образцов.
PspA восстанавливает мембраны
Под микроскопом исследователи смогли распознать или, выражаясь жаргоном, «разрешить» структуру PspA. Структура белка необходима для его функции, и дефект в структуре может нарушить функцию белка. «Под микроскопом мы поняли, что PspA имеет структуру, аналогичную белкам ESCRT-III, над которыми уже работала наша лаборатория. Это стало полной неожиданностью и показало, насколько важно детально выяснить структуру белков», — сказал Сакс. .«Спустя миллиарды лет две группы белков генетически разошлись настолько, что их сходство можно было обнаружить только на основе их структуры».
Структура белка необходима для его функции, и дефект в структуре может нарушить функцию белка. «Под микроскопом мы поняли, что PspA имеет структуру, аналогичную белкам ESCRT-III, над которыми уже работала наша лаборатория. Это стало полной неожиданностью и показало, насколько важно детально выяснить структуру белков», — сказал Сакс. .«Спустя миллиарды лет две группы белков генетически разошлись настолько, что их сходство можно было обнаружить только на основе их структуры».
белка ESCRT-III обнаружены во всех живых организмах с истинным клеточным ядром, включая клетки человека. Здесь белковые комплексы ESCRT-III участвуют в ремоделировании и восстановлении клеточной мембраны, но также играют ключевую роль в ряде других клеточных процессов. У бактерий ранее не были известны белки семейства ESCRT-III. «Следовательно, PspA и ESCRT-III принадлежат к одной группе белков.Эти два вещества выполняют схожие задачи на мембранах внутри клеток», — сказал Шнайдер.
«Основываясь на схожих структурных и функциональных свойствах PspA и эукариотических белков ESCRT-III, мы идентифицировали PspA как бактериальный член эволюционно консервативного суперсемейства белков ремоделирования мембран ESCRT-III», — написали авторы в своей статье для .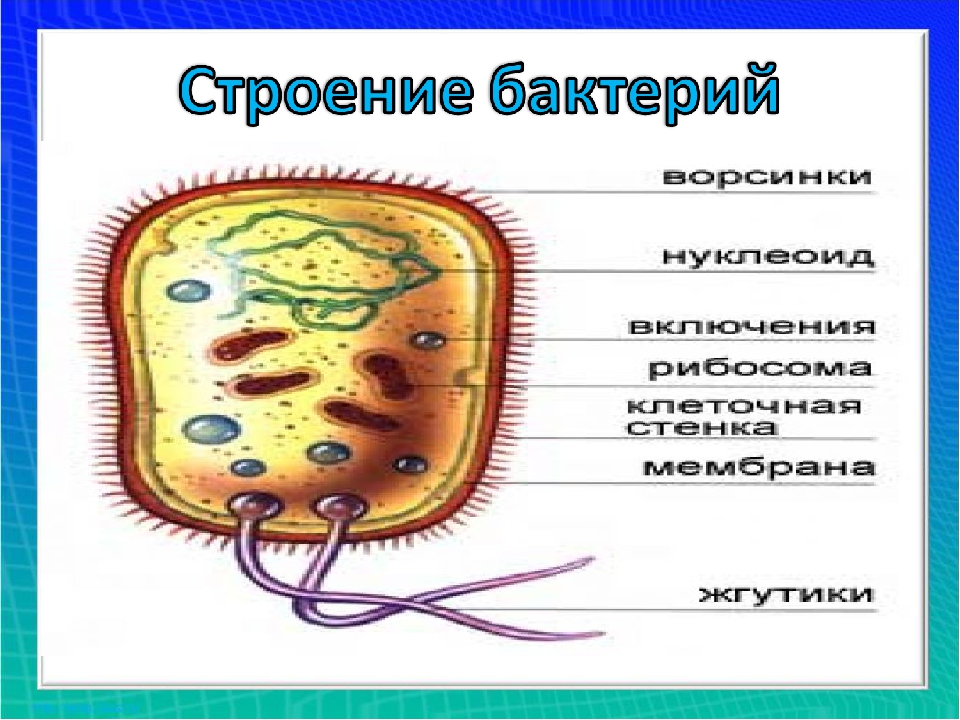 Сотовый .
Сотовый .
Источник истории:
Материалы предоставлены Johannes Gutenberg Universitaet Mainz . Примечание. Содержимое можно редактировать по стилю и длине.
Цитоплазма прокариотическая | Encyclopedia.com
Цитоплазма прокариотической клетки — это все, что находится внутри бактерии. В отличие от эукариотической клетки внутри бактерий нет функциональной сегрегации . Цитоплазма содержит все химические вещества и компоненты, которые используются для поддержания жизни бактерии, за исключением тех компонентов, которые находятся в мембране(ах) и периплазме грамотрицательных бактерий.
Цитоплазма ограничена цитоплазматической мембраной. Грамотрицательные бактерии содержат еще одну наружную мембрану. Между двумя мембранами находится периплазма.
При осмотре в световом микроскопе цитоплазма бактерий прозрачна. Только при большем увеличении, доступном с помощью просвечивающего электронного микроскопа , становится очевидным зернистый характер цитоплазмы. Точная структура цитоплазмы вполне может отличаться от этой точки зрения, поскольку цитоплазма состоит в основном из воды.Дегидратация, необходимая для обычной электронной микроскопии, вероятно, влияет на структуру цитоплазмы.
Точная структура цитоплазмы вполне может отличаться от этой точки зрения, поскольку цитоплазма состоит в основном из воды.Дегидратация, необходимая для обычной электронной микроскопии, вероятно, влияет на структуру цитоплазмы.
Цитоплазма прокариот и эукариот сходна по текстуре. Вместо того, чтобы быть свободно текущей жидкостью, цитоплазма больше похожа на гель. Консистенция сравнима с консистенцией десертного геля, за исключением того, что бактериальный гель способен течь. Способность к течению жизненно важна, поскольку молекулы, находящиеся в цитоплазме, должны быть способны перемещаться внутри бактерии, а также в цитоплазму и из нее.
Генетический материал бактерий рассеян по всей цитоплазме. Иногда геном дезоксирибонуклеиновой кислоты может агрегировать во время подготовки к микроскопии. Затем геном проявляется как более размытая область в зернистой цитоплазме. Эта искусственная структура была названа нуклеоидом. Также могут присутствовать меньшие кольцевые расположения генетического материала, называемые плазмидами . Рассредоточение бактериального генома по всей цитоплазме — одна из основных отличительных черт прокариотических и эукариотических клеток.
Рассредоточение бактериального генома по всей цитоплазме — одна из основных отличительных черт прокариотических и эукариотических клеток.
В цитоплазме также присутствует рибонуклеиновая кислота , различные ферменты , аминокислоты, углеводы, липиды, ионы и другие соединения, функционирующие в бактериях. Составляющие мембраны производятся в цитоплазме, а затем транспортируются к месту назначения.
Некоторые бактерии содержат специализированные области, известные как цитоплазматические включения, которые выполняют специализированные функции. В этих включениях могут храниться продукты, которые используются для питания бактерий.Примерами таких включений являются гликоген, поли-В-гидроксибутират и гранулы серы. Кроме того, некоторые бактерии содержат заполненные газом везикулы, которые удерживают бактерию на определенной глубине в воде, или мембранные структуры, содержащие хлорофилла . Последние служат для сбора света для получения энергии фотосинтезирующими бактериями.
Цитоплазма прокариотических клеток также содержит рибосом , необходимых для производства белка. В цитоплазме может быть много рибосом.Например, быстрорастущая бактерия может содержать более 15 000 рибосом.
В цитоплазме происходят процессы транскрипции , трансляции , импорта и экспорта белков, а также хоть какая-то деградация соединений. У грамотрицательных бактерий некоторые из этих функций также осуществляются в периплазматической жидкости. Механизмы, лежащие в основе правильной последовательной организации этих функций, еще предстоит полностью определить.
См. также Ультраструктура бактерий
лекция09
лекция0931 января 2005
Лекция 9
Чтение, Глава 4 и стр. 318-319
IV.Ячейки
B. Прокариоты и эукариоты
1. Прокариоты
Общая ячейка, которую мы
обсуждалось ранее, была эукариотическая клетка, как и ваши собственные клетки. Прокариотические клетки отличаются от эукариотических
следующими способами:
Прокариотические клетки отличаются от эукариотических
следующими способами:
а. Они очень маленькие, около 1 мкм. 1 мкм равен 1/1000 миллиметра. Эукариотические клетки, такие как ваша, имеют размер от 30 до 100 мкм.
б. У них есть нет ядра. Это означает, что их хромосома, где находятся гены, не заключен в мембрану.
г. У них есть одиночная кольцевая хромосома (у вас 46 линейных хромосом в ядра ваших клеток). Эта единственная хромосома имеет относительно мало гены и немного ДНК, которая не кодирует гены. Напротив, ваш хромосомы имеют много генов, и большая часть ДНК не кодирует гены.На самом деле большая часть ДНК в ваших хромосомах не похожа на другую. Делать что-нибудь.
д. Прокариотический
клетки не имеют хлоропластов и митохондрий. Несмотря на это, многие из
они могут осуществлять аэробное дыхание того же типа, что и митохондрии.
делать. Некоторые из них могут осуществлять фотосинтез так же, как это делают хлоропласты.
Обратите внимание, что вместо означает «до», а karyon означает «ядро». Прокариот означает «перед ядрами». Eu означает «истинный», поэтому эукариотический означает «истинное ядро».
2. Гипотеза эндосимбионта
Первые ячейки считались прокариотами, судя по микрофоссилиям и тот факт, что прокариоты проще, чем эукариоты. Эукариоты в настоящее время предположительно возникли впоследствии из комбинаций прокариотических клетки, концепция, известная как гипотеза эндосимбионтов.
Эта гипотеза была
впервые предложено более ста лет назад, но не было воспринято всерьез
до 1970-х годов, когда его популяризировала Линн Маргулис. Изначально,
Доктор Маргулис и гипотеза эндосимбионта подверглись резкой критике.
Сейчас в биологии общепринята гипотеза эндосимбионтов.
сообщество. Со временем накопление доказательств преодолело раннее
скептицизм. В частности, гипотеза эндосимбионта
ответы на многие вопросы, которые возникали о клетках, например
Почему у митохондрий есть собственная ДНК.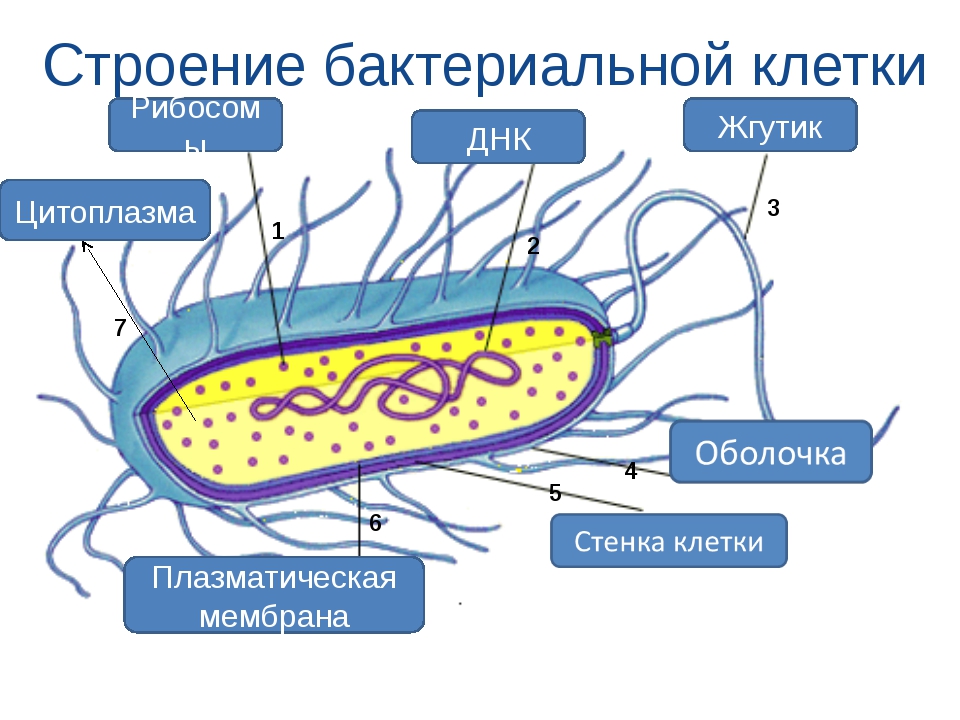
Чтобы лучше понять ячеек, мы кратко рассмотрим наименее спорную часть эндосимбионтная гипотеза, заключающаяся в том, что митохондрии и хлоропласты когда-то были свободноживущими бактериями, которые были поглощены более крупными клетками и остаются симбиотическими партнерами.
Доказательства эндосимбионтное происхождение митохондрий и хоропластов включает далее:
— Хлоропласты и митохондрии являются прокариотами.У них есть свои гены на маленькой кольцевой хромосоме, но без ядра. Этот Хромосома имеет небольшое количество некодирующей ДНК, как у бактерии. Хлоропласты и митохондрии также производят некоторые свои собственные белки из своих гены. Частью этого процесса является чтение РНК-копий генов. и правильная сборка аминокислот в белки. Это чтение сборка осуществляется очень маленькими органеллами, называемыми рибосомы. Хлоропласты и митохондрии имеют собственные рибосомы. похожие на бактерии и отличные от остальных клетки.По этой причине они чувствительны к антибиотикам. которые убивают бактерии путем связывания и инактивации бактериальных рибосомы.
— ДНК последовательности некоторых генов в хлоропластах и митохондриях больше похоже на последовательности ДНК для тех же самых генов у бактерий чем к тем же генам в ядре клетки. Этот указывает на то, что хлоропласты и митохондрии более тесно относятся к бактериям, чем к ядрам клеток в которым они живут.
— Эндосимбиоз происходящие сегодня. Коралловые животные, несколько моллюсков, морские слизни и растения все корни образуют симбиозы с водорослями или бактериями, в которых симбиотический партнер живет внутри клеток хозяина. Разные существуют виды динофлагеллят, которые находятся на разных стадиях приобретения хлоропласта путем поглощения и сохранения одноклеточные водоросли.
Изучение генов
присутствующие в хлоропластах и митохондриях, предполагают, что хлоропласт
предком была цианобактерия, а митохондриальным предком была
бактерия, способная к аэробному дыханию.
C. Мембраны и органеллы
Есть много видов клеток. Мы очень кратко обсудили клетки животных, клетки растений, и бактериальные (прокариотические) клетки. Сейчас мы сосредоточимся на общем особенности типичной животной клетки. (растительные и бактериальные клетки будут более подробно рассматривается позже в этом семестре, когда мы проводим обзор многообразие живых существ).
В многоклеточном организма, такого как человек, клетки специализированы для многих различных функции и, следовательно, имеют различные формы.Почти все эукариотические клетки обладают всеми следующими структурными особенностями, однако:
1. Клетка Мембрана
Это граница
клетки. Клеточная мембрана также известна как плазматическая мембрана
или иногда «плазмалемма». Он создает внутреннее пространство, которое может
быть организованным иначе, чем внешняя среда. Космос
внутри клеточной мембраны называется «цитоплазмой».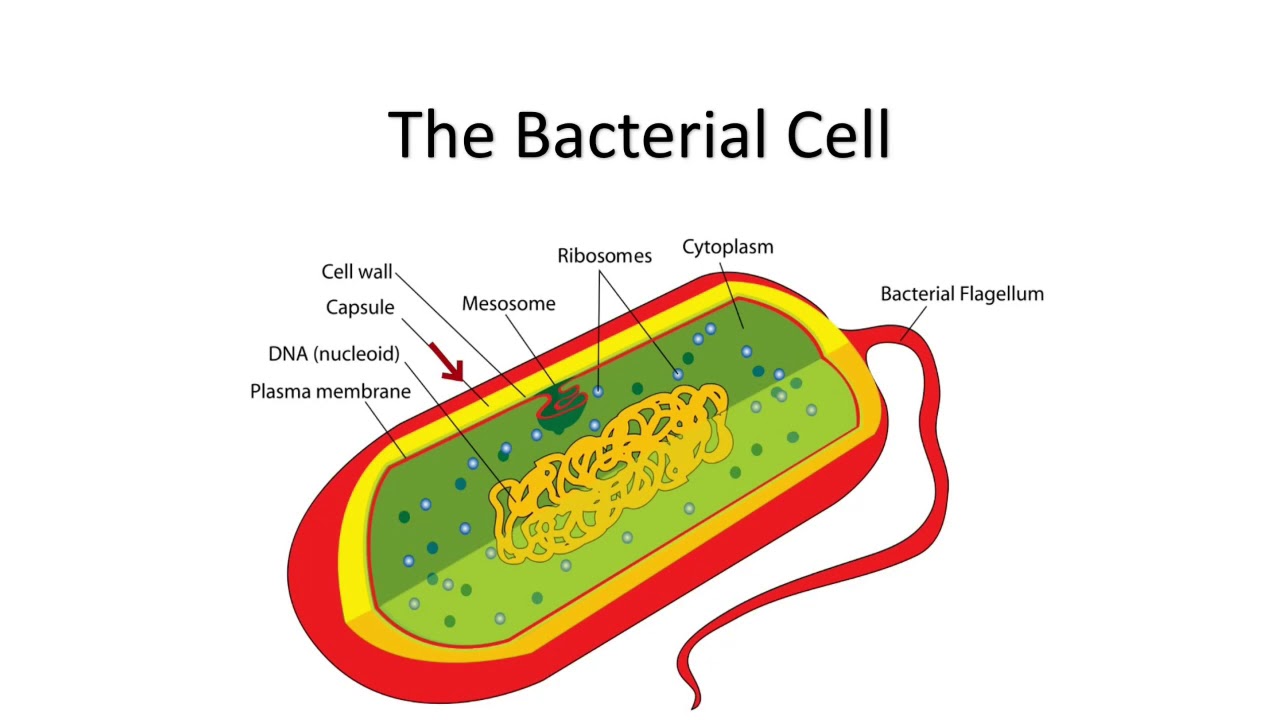 Это ставит под угрозу
густой раствор воды с большим количеством растворенных молекул и органелл
подвешен в нем.
Это ставит под угрозу
густой раствор воды с большим количеством растворенных молекул и органелл
подвешен в нем.
Клеточная мембрана состоит из липидного двойного слоя мембраны. Мембранные липиды имеют полярную, гидрофильная головная группа, которая часто включает фосфат (ПО 4 ). К головной группе присоединены две жирные кислоты «хвосты», которые неполярны и гидрофобны. В водных растворах, такие, которые встречаются внутри и снаружи клеток, эти липиды образуют двойную слой, в котором полярные гидрофобные головные группы обращены к внутренние и внешние водные растворы, в то время как неполярные, гидрофобные хвост обращен внутрь, от воды.
Липидный бислой
клеточная мембрана является барьером для многих видов молекул и ионов
растворяются в водном растворе внутри и снаружи клеток.
Крупные незаряженные молекулы (например, сахара) не проходят легко
через бислой. Небольшие заряженные растворенные вещества (ионы) также
не проходят легко (например, H + , Na + , K + ,
Кл —).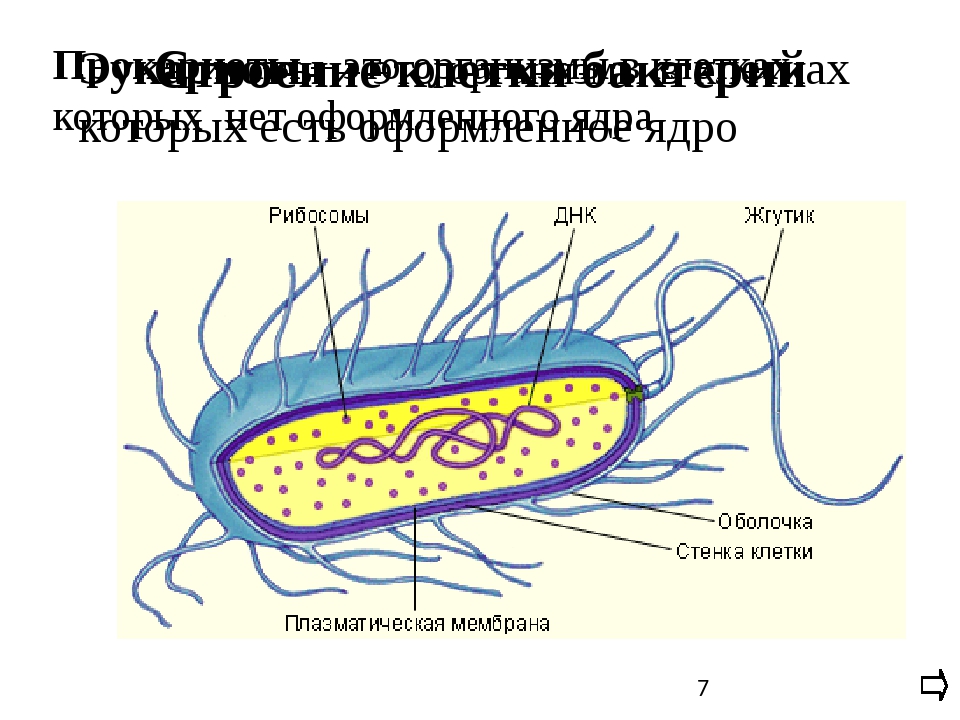 Небольшие незаряженные молекулы (например, O 2 ,
CO 2 , H 2 O) быстро проходят через бислой,
тем не мение.
Небольшие незаряженные молекулы (например, O 2 ,
CO 2 , H 2 O) быстро проходят через бислой,
тем не мение.
Встраивается в липид бислой клеточной мембраны (и других мембран в клетке) многие мембранные белки. К ним относятся следующие:
Ферменты — Белки, ускоряющие определенные химические реакции.Транспорт белки — Белки, которые перемещают ионы или большие молекулы через мембрана.
Рецептор белки — Белки, обнаруживающие гормоны или другие внешние сигналы, посылаемые в клетку другими клетками.
Признание белки — это белки, которые идентифицируют клетки в организме к другим клеткам. Инородные клетки, у которых отсутствует эта идентифицирующая мембрана белки, напр. патогены, уничтожаются вашим иммунным система.
Адгезия белки — это белки, которые соединяют клетки в ткани.
