
Биология для студентов — 11. Жгутики, фимбрии бактерий. Строение, функции, механизм движения
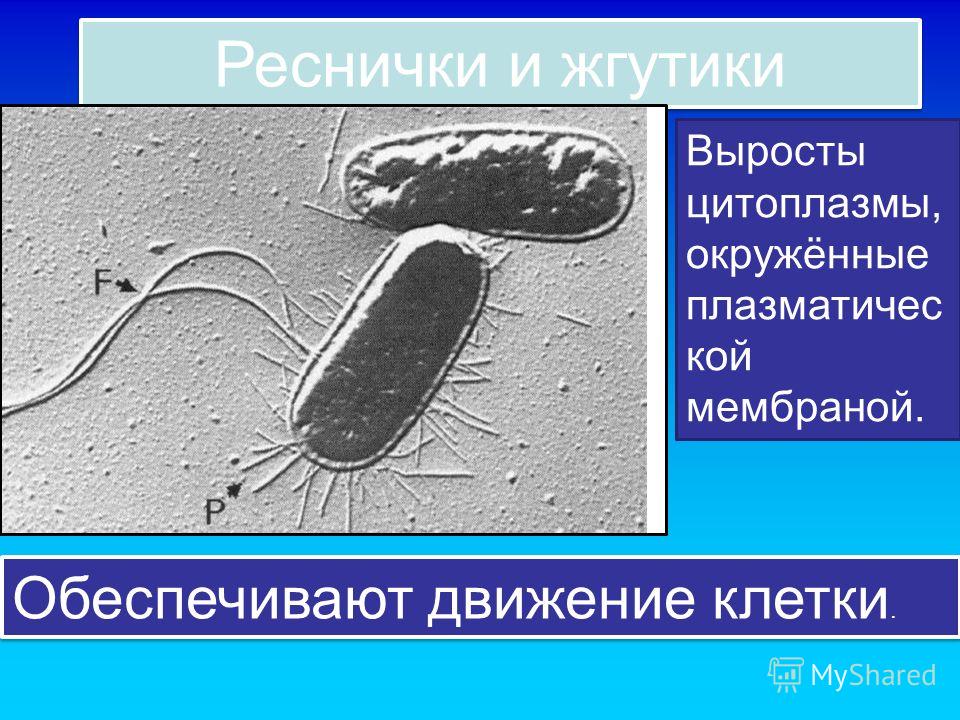
На клеточной поверхности многих прокариот имеются структуры, определяющие способность клетки к движению в жидкой среде. Это — жгутики. Их число, размеры, расположение, как правило, являются признаками, постоянными для определенного вида, и поэтому учитываются при систематике прокариот. Однако накапливаются данные о том, что количество и расположение жгутиков у одного и того же вида могут в значительной степени определяться условиями культивирования и стадией жизненного цикла, и, следовательно, не стоит переоценивать таксономическое значение этого признака.
Если жгутики находятся у полюсов или в полярной области клетки, говорят об их полярном или субполярном расположении, если вдоль боковой поверхности, говорят о латеральном расположении.
Жгутики представляют собой длинные отростки, которые отходят от одного (монотрихи, лофотрихи) или обоих (амфитрихи) полюсов бактериальной клетки либо распределены по всей ее поверхности (перитрихи).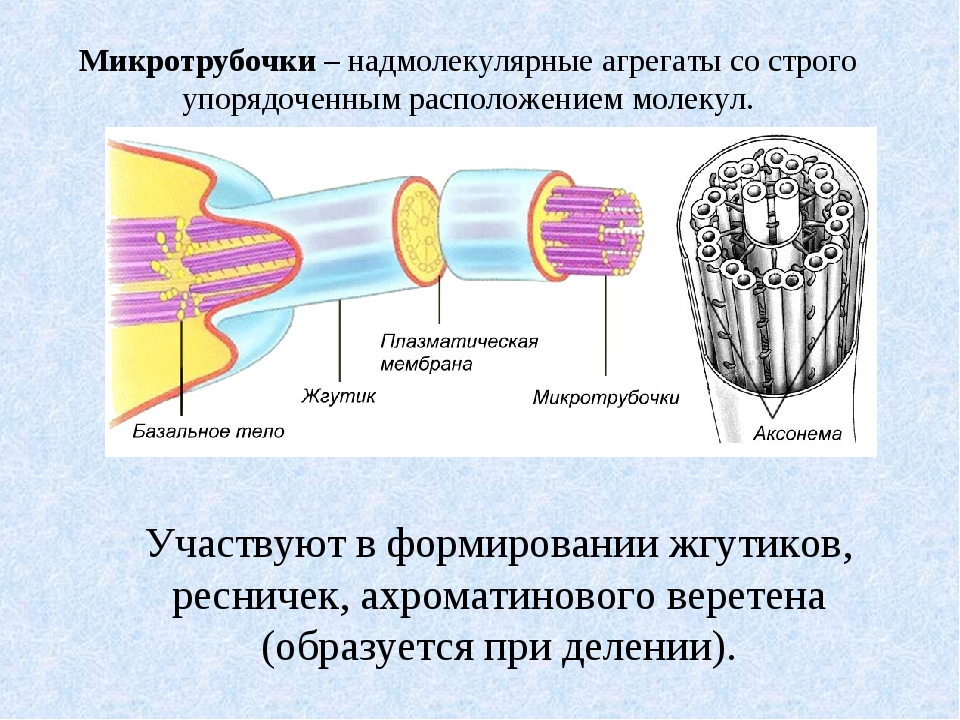
У некоторых спирохет, например, Treponema pallidum и Borrelia burgdorferi, продольно расположенные жгутики собраны в осевую нить. Благодаря этому образованию, спирально охватывающему клетку, спирохеты могут активно передвигаться при помощи вращательных движений. Некоторые бактерии могут перемещаться по субстрату без видимых двигательных структур.
В зависимости от числа жгутиков и их локализации на поверхности клетки различают:
- монополярные монотрихи(один жгутик прикреплен к одному полюсу клетки;
- монополярные политрихи(пучок жгутиков расположен на одном полюсе клетки), биполярные политрихи (на каждом полюсе — по пучку жгутиков;
- перитрихи(многочисленные жгутики расположены по всей поверхности клетки или вдоль ее боковой поверхности.
В последнем случае число жгутиков может достигать 1000 на клетку.
Обычная толщина жгутика — 10-20 нм, длина — от 3 до 15 мкм. У некоторых бактерий длина жгутика может на порядок превышать диаметр клетки. Как правило, полярные жгутики более толстые, чем перитрихиальные.
Жгутик представляет собой относительную жесткую спираль, обычно закрученную против часовой стрелки. Вращение жгутика также осуществляется против часовой стрелки с частотой от 40 до 60 об/с, что вызывает вращение клетки, но в противоположном направлении. Поскольку клетка намного массивнее жгутика, она вращается со значительно меньшей скоростью — порядка 12-14 об/мин. Вращательное движение жгутика преобразуется также в поступательное движение клетки, скорость которого в жидкой среде для разных видов бактерий составляет от 16 до 100 мкм/с.
Изучение строения жгутика под электронным микроскопом обнаружило, что он состоит из трех частей. Основную массу жгутика составляет длинная спиральная нить (фибрилла), у поверхности клеточной стенки переходящая в утолщенную изогнутую структуру — крюк. Нить с помощью крюка прикреплена к базальному телу, вмонтированному в ЦПМ и клеточную стенку. Белковые субъединицы уложены в виде спирали, внутри которой проходит полый канал. Наращивание жгутика происходит с дистального конца, куда субъединицы поступают по внутреннему каналу. У некоторых видов жгутик снаружи дополнительно покрыт чехлом особого химического строения или же являющимся продолжением клеточной стенки и, вероятно, построенным из того же материала.
Нить с помощью крюка прикреплена к базальному телу, вмонтированному в ЦПМ и клеточную стенку. Белковые субъединицы уложены в виде спирали, внутри которой проходит полый канал. Наращивание жгутика происходит с дистального конца, куда субъединицы поступают по внутреннему каналу. У некоторых видов жгутик снаружи дополнительно покрыт чехлом особого химического строения или же являющимся продолжением клеточной стенки и, вероятно, построенным из того же материала.
К поверхностным структурам бактериальной клетки относятся также фимбрии (пили, реснички, ворсинки) — жесткие прямые полые нити из белка пилина, локализованые на КС. Фимбрии короче и тоньше жгутиков: их диаметр 3–20 нм, длина 0,2–10,0 мкм.
Фимбрии — необязательная клеточная структура, так как и без них бактерии хорошо растут и размножаются. В отличие от жгутиков, фимбрии не выполняют двигательную функцию и обнаружены у подвижных и неподвижных форм. По своему функциональному назначению фимбрии подразделяются на 2 типа. Термин «фимбрии» чаще используется для обозначения общих пили, а термин «пили» — для обозначения секс-пили.
Термин «фимбрии» чаще используется для обозначения общих пили, а термин «пили» — для обозначения секс-пили.
Фимбрии 1 (общего) типа имеются у большинства бактерий. Они покрывают всю поверхность клетки, располагаются перитрихиально или полярно. Количество фимбрий велико — от нескольких сотен до нескольких тысяч на одну бактериальную клетку. Синтез фимбрий контролируется бактериальной хромосомой, утрата фимбрий приводит к их новому синтезу.
Покрывая всю клетку, фимбрии создают ворсистую поверхность. Иногда фимбрии сливаются в комки, придавая неопрятный вид клетке; в других случаях поверхность клеток покрыта войлокообразным чехлом, состоящим из сплетений тонких нитей.
Пили 2 типа (синонимы: конъюгативные, половые, секс-пили) образуются только мужскими клетками-донорами, содержащими трансмиссивные плазмиды (F, R, Col), в ограниченном количестве (1–4 на клетку), имеют терминальные вздутия.
Функции фимбрий.
Фимбрии обоих типов:
- Обладают антигенной активностью.

- На них адсорбируются бактериофаги (специфические вирусы бактерий).
- Адгезивная функция: обеспечивают прикрепление бактерий к клеткам слизистых оболочек организма хозяина и к другим субстратам (клеткам растений, грибов, неорганическим частицам и органическим остаткам).
- Механическая защита бактериальной клетки. Придают бактериям свойство гидрофобности и способствуют объединению клеток в группы.
- Увеличивают всасывательную поверхность клетки бактерий, участвуют в процессах питания, водно-солевого обмена и в транспорте метаболитов.
Половые пили: F–пили обеспечивают конъюгацию — передачу части генетического материала от донорской клетки к реципиентной.
:: Теория и практика :: Цитозоль и цитоскелет :: Функции цитоскелета :: функции микротрубочек
Клеточный центр, или центросома, представляет собой немембранный органоид, локализованный около ядра и состоящий из двух центриолей и центросферы. Постоянным и наиболее важным компонентом клеточного центра являются центриоли.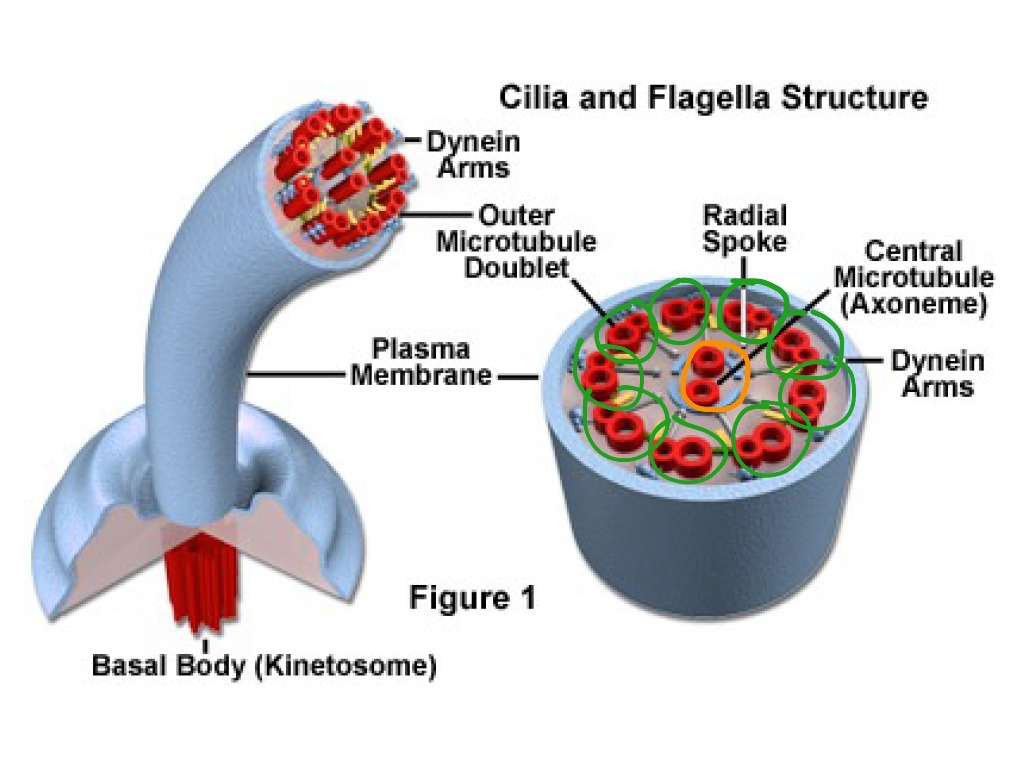 Этот органоид обнаружен в клетках животных, низших растений и грибов.
Этот органоид обнаружен в клетках животных, низших растений и грибов. Центриоли (от лат. centrum – срединная точка, центр) представляют собой два перпендикулярно расположенных друг к другу цилиндра, стенки которых образованы микротрубочками и соединены системой связок. Конец одного цилиндра (дочерняя центриоль) направлен к поверхности другого (материнская центриоль). Совокупность сближенных между собой материнской и дочерней центриолей называется диплосомой. Впервые центриоли были обнаружены и описаны в 1875 г. В. Флемингом. В интерфазных клетках центриоли часто располагаются возле комплекса Гольджи и ядра.
Стенка центриолей состоит из расположенных по окружности 9 триплетов микротрубочек, образующих полый цилиндр. Систему микротрубочек центриоли можно описать формулой (9X3) + 0, подчеркивая отсутствие микротрубочек в центральной части. Диаметр центриоли составляет около 0,2 мкм, длина — 0,3-0,5 мкм (однако, есть центриоли, достигающие в длину нескольких микрометров). Кроме микротрубочек в состав центриоли входят дополнительные структуры — «ручки», соединяющие триплеты.

Центросфера — плотный слой цитоплазмы вокруг центриолей, в котором часто содержатся микротрубочки, расположенные лучами.
Центриолярный цикл. Строение и активность центриолей меняются в зависимости от периода клеточного цикла. Это позволяет говорить о центриолярном цикле. В начале периода G1 от поверхности материнской центриоли начинается рост микротрубочек, которые растут и заполняют цитоплазму. По мере роста микротрубочки теряют связь с областью центриолей и могут находиться в цитоплазме длительное время.
В периоде S или G2 происходит удвоение числа центриолей. Этот процесс заключается в том, что центриоли в диплосоме расходятся и около каждой из них происходит закладка процентриолей. В начале вблизи и перпендикулярно исходной центриоли закладываются девять одиночных микротрубочек. Затем они преобразуются в девять дуплетов, а потом в девять триплетов микротрубочек новых центриолей. Этот способ увеличения числа центриолей был назван дупликацией. Следует отметить, что удвоение числа центриолей не связано с их делением, почкованием или фрагментацией, а происходит путем образования процентриолей.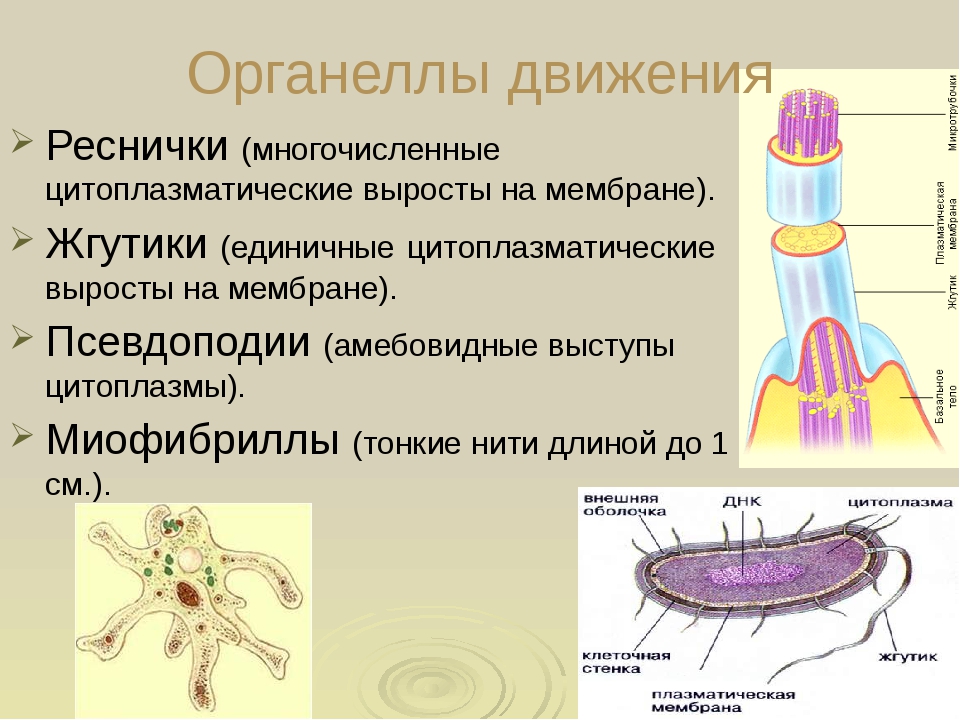 Таким образом, в результате дупликации в клетке содержатся четыре попарно связанные центриоли. В этом периоде материнская центриоль продолжает играть роль центра образования цитоплазматических микротрубочек.
Таким образом, в результате дупликации в клетке содержатся четыре попарно связанные центриоли. В этом периоде материнская центриоль продолжает играть роль центра образования цитоплазматических микротрубочек.
В периоде G2 обе материнские центриоли покрываются фибриллярным гало (зона тонких фибрилл), от которого в профазе начнут отрастать митотические микротрубочки. В этом периоде в цитоплазме происходит исчезновение микротрубочек и клетка стремится приобрести шаровидную форму. В профазе митоза диплосомы расходятся к противоположным полюсам клетки. От фибриллярного гало материнской центриоли отходят микротрубочки, из которых формируется веретено деления митотического аппарата. Таким образом, центриоли являются центрами организации роста микротрубочек. В телофазе происходит разрушение веретена деления.
Следует отметить, что в клетках высших растений, некоторых водорослей, грибов, ряда простейших центры организации роста микротрубочек центриолей не имеют. У некоторых простейших центрами индукции образования микротрубочек выступают плотные пластинки, связанные с мембраной.
сжатая характеристика, строение и роль в клетках. Из чего состоят жгутики
Как прокариотические, так и эукариотические могут содержат структуры, известные как реснички и жгутики. Эти выросты на поверхности клеток помогают в их .
Особенности и функции
Реснички и жгутики являются выростами из некоторых клеток необходимые для клеточной локомоции (передвижения). Они также помогают перемещать вещества вокруг клеток и направлять их к нужным участкам.
Реснички и жгутики образуются из специализированных групп микротрубочек, называемых базальными телами.
Если выросты короткие и многочисленные, их называют ресничками. Если они длиннее и менее многочисленны (обычно только один или два), они называются жгутиками.
Строение
Обычно реснички и жгутики имеют сердцевину, состоящую из микротрубочек, соединенных с , расположенных по схеме 9+2. Кольцо из девяти микротрубочек имеет в своем центре две особые микротрубочки, которые сгибают реснички или жгутики. Этот тип организации встречается в устройстве большинства ресничек и жгутиков .
Этот тип организации встречается в устройстве большинства ресничек и жгутиков .
Где встречаются?
Как реснички, так и жгутики встречаются во многих типах клеток. Например, у спермы многих животных, водорослей и даже папоротников есть жгутики. Реснички можно найти в клетках таких тканей, как дыхательные пути и женский репродуктивный тракт.
Для передвижения в водной среде некоторые микроорганизмы используют жгутиковидный орган – «жгутик». Этот орган, встроенный в мембрану клетки, позволяет микроорганизму по желанию передвигаться в выбранном им направлении с определённой скоростью.
Мужские половые клетки также используют жгутик для передвижения.
Определенной время учёные знали о жгутиках. Однако, знания об их структурных особенностях, которые появились лишь за последнее десятилетие или около того, явились для них огромным сюрпризом. Было установлено, что жгутик двигается посредством очень сложного «органического двигателя», а не простого вибрирующего механизма, как считалось раньше.
Этот двигатель сформирован по тем же механическим принципам, что и электрический двигатель. В нём есть две главные части: движущаяся часть («ротор») и станционарная часть («статор»).
Бактериальный жгутик отличается от всех органических систем, которые совершают механические движения. Клетка не использует имеющуюся в наличии энергию, сохраняемую в молекулах АТФ. Вместо этого, она имеет специальный энергетический ресурс: микроорганизм использует энергию потока ионов сквозь их внешние мембраны. Внутренняя структура двигателя чрезвычайно сложна. В создании жгутика принимают участие около 240 различных белков. Каждый из них занимает определённое место. Учёные установили, что эти белки проводят сигналы, включающие и выключающие двигатель, формируют соединения, облегчающие движения на атомном уровне, и активизируют другие белки, которые присоединяют жгутик к клеточной мембраны. Моделей, разработанных для резюмирования работы системы, вполне достаточно для описания сложной структуры системы.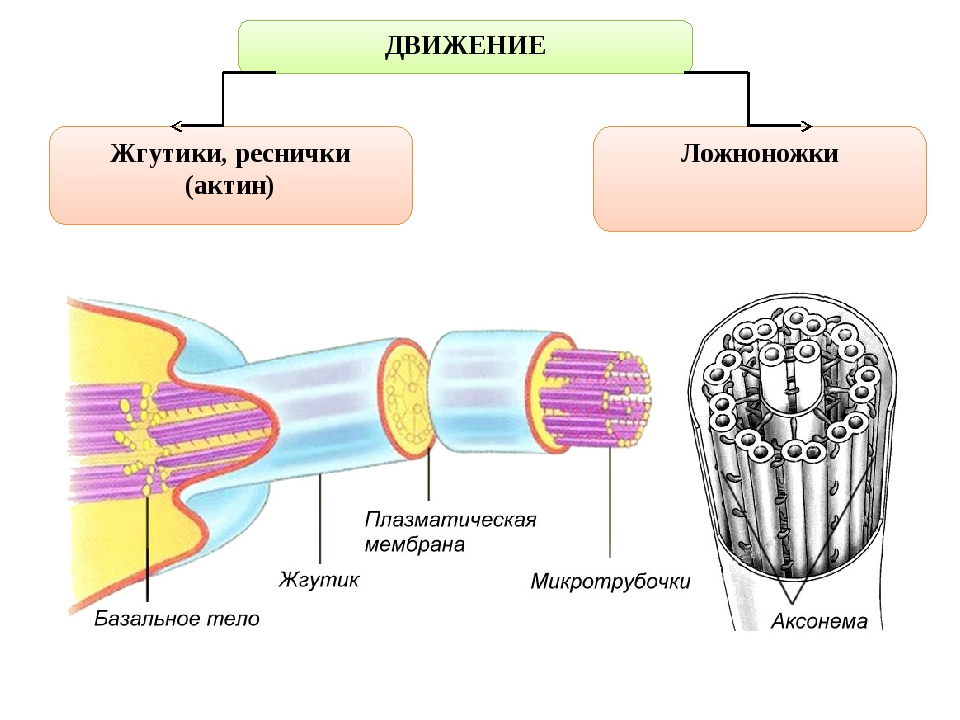
Сложной структуры бактериального жгутика самой по себе уже достаточно, чтобы опровергнуть теорию эволюции, поскольку жгутик имеет неснижаемо сложную структуру. Даже если одна единственная молекула этой невероятно сложной структуры исчезнет или повредится, жгутик не будет ни работать, ни представлять пользу для микроорганизма. Жгутик должен идеально работать с самого первого момента его существования. Этот факт ещё раз подчёркивает абсурдность утверждения теории эволюции о «ступенчатом развитии».
Даже у тех созданий, которых эволюционисты считают «простейшими», существует потрясающая структура. Бактериальный жгутик является одним из бесчисленных примеров. Этот микроорганизм передвигается в воде, двигая этим органом на своей оболочке. Когда были изучены внутренняя система этого хорошоизвестного органа, учёные всего мира были удивлены, обнаружив, что микроорганизм имеет чрезвычайно сложный по своему строению электрический двигатель. Этот электрический двигатель, который включает около пятидесяти разнообразных молекулярных субедениц, имеет достаточно замысловатую структуру, что изображено ниже.
Бактериальный жгутик является очевидным доказательством того, что даже предположительно «примитивные» создания имеют необычное строение. По мере того, как человечество всё более постигает детали, становится очевидным, что те организмы, которые учёные 19-го столетия, включая Дарвина, считали простейшими, на самом деле так же сложны, как и другие. Другими словами, с приходом понимания о совершенстве создания, очевидным становится бессмысленность попыток найти сотворению альтернативное объяснение.
Микороорганизм плавает в вязкой жидкой среде, вращая спиралевидными пропеллерами, называемыми жгутиками .
Бактериальный жгутик является наномашиной, состоящей из 25 разнообразных белков, в количестве от нескольких штук до десятков тысяч. Он состоит из собрания этого большого количества белков, каждый из которых в разных частях выполняет определённую функцию, такую как ротация двигателя, изоляция, приводной вал, очерёдность переключения регулятора, универсальная связка, спиралевидный пропеллер, и ротативный усилитель для самосборки.
Жгутиковые протеины синтезируются внутри тела клетки и транспортируются по длинному, узкому центральному каналу в жгутике к его периферическому (внешнему) окончанию, где они с помощью жгутикового наконечника в качестве установочного двигателя, эффективно и самостоятельно могут создавать сложные структуры наноразмером. Вращательный двигатель, диаметр которого всего от 30 до 40 нм, производит вращение жгутика с частотой около 300 Гц и мощностью в 10-16 Вт, с эффективностью преобразования энергии близкой к 100%.
Структурные дизайны и функциональные механизмы, обнаруженные в сложном механизме бактериального жгутика, могут обеспечить человечество множеством новаторских технологий, которые станут основой для будущей нанотехнологии, для которой мы сможем найти много полезных способов применения.(2)
Развитие микробиологии принесло в последние десятилетия множество открытий. И одно из них — это особенности движения жгутиковых бактерий. Устройство двигателей этих древнейших организмов оказалось очень сложным и по принципу своей работы очень отличающимся от жгутиков наших ближайших эукариотических родственников простейших.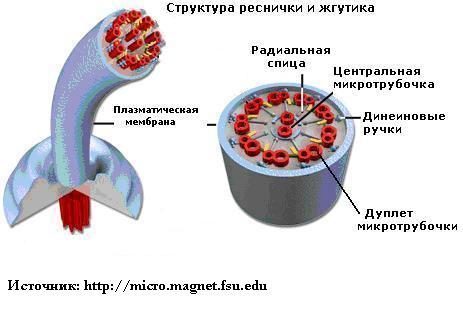 Вокруг двигателя жгутиковой бактерии разгорелись самые жаркие споры между креационистами и эволюционистами. О бактериях, их жгутиковых моторах и о многом другом — данная статья.
Вокруг двигателя жгутиковой бактерии разгорелись самые жаркие споры между креационистами и эволюционистами. О бактериях, их жгутиковых моторах и о многом другом — данная статья.
Общая биология
Для начала вспомним, что это за организмы и какое место они занимают в системе органического мира на нашей планете. Домен Bacteria объединяет огромное количество одноклеточных прокариотических (без оформленного ядра) организмов.
Эти живые клетки появились на арене жизни почти 4 миллиарда лет назад и были первыми поселенцами планеты. Они могут быть самой разной формы (кокки, палочки, вибрионы, спирохеты), но большинство из них — жгутиковые.
Где живут бактерии? Везде. На планете их обитает более 5×10 30 . В 1 грамме почвы их насчитывается порядка 40 миллионов, в нашем организме обитает до 39 триллионов. Их можно встретить на дне Марианской впадины, в горячих «черных курильщиках» на дне океанов, во льдах Антарктиды, а на ваших руках в данный момент находится до 10 миллионов бактерий.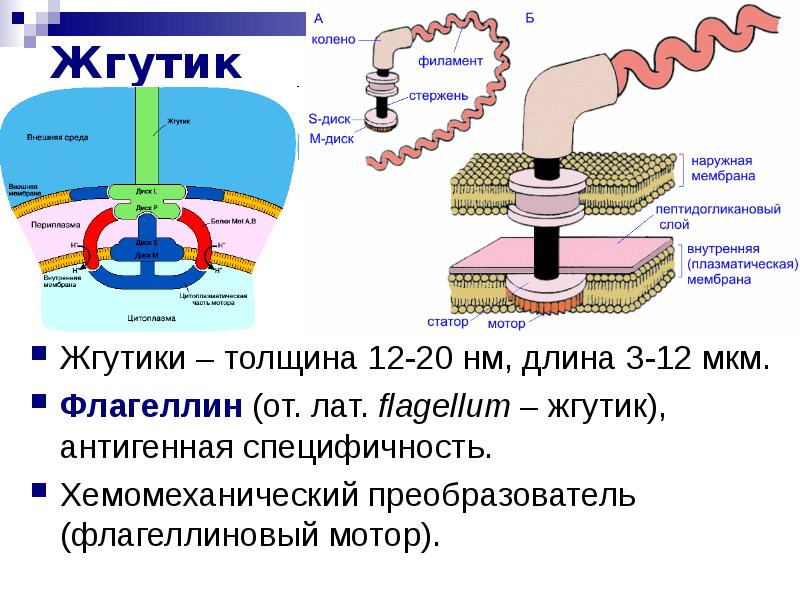
Значение неоспоримо
Несмотря на микроскопические размеры (0,5-5 мкм) их общая биомасса на Земле больше, чем биомасса животных и растений, вместе взятых. Их роль в круговороте веществ незаменима, а их свойства консументов (разрушителей органики) не позволяют планете покрыться горами трупов.
Ну и не стоит забывать о патогенах: возбудителями чумы, оспы, сифилиса, туберкулеза и множества других инфекционных заболеваний тоже являются бактерии.
Бактерии нашли применение и в хозяйственной деятельности человека. Начиная от пищевой промышленности (кисломолочные продукты, сыры, квашеные овощи, алкогольные напитки), «зеленой» экономике (биотопливо и биогаз) до методов клеточной инженерии и производства лекарственных препаратов (вакцины, сыворотки, гормоны, витамины).
Общая морфология
Как уже говорилось, у этих одноклеточных представителей жизни нет ядра, их наследственный материал (молекулы ДНК в виде кольца) расположены в определенном участке цитоплазмы (нуклеоид). Клетка их имеет плазматическую мембрану и плотную капсулу, образованную пептидогликаном муреином. Из клеточных органелл у бактерий есть митохондрии, могут быть хлоропласты и другие структуры с различными функциями.
Клетка их имеет плазматическую мембрану и плотную капсулу, образованную пептидогликаном муреином. Из клеточных органелл у бактерий есть митохондрии, могут быть хлоропласты и другие структуры с различными функциями.
Большинство бактерий — жгутиковые. Плотная капсула на поверхности клетки не позволяет им передвигаться при помощи изменения самой клетки, как это делают амебы. Их жгутики — это плотные белковые образования различной длины и диаметром около 20 нм. Одни бактерии имеют единственный жгутик (монотрихи), у других их два (амфитрихи). Иногда жгутики расположены пучками (лофотрихи) или же покрывают всю поверхность клетки (перитрихи).
Многие из них живут в виде одиночных клеток, но некоторые образуют скопления (пары, цепочки, филаменты, гифы).
Особенности движения
Жгутиковые бактерии могут двигаться по-разному. Одни движутся только вперед, а направление меняют путем кувыркания. Некоторые способны к подергиваниям, другие передвигаются путем скольжения.
Жгутики бактерий выполняют функции не только клеточного «весла», но могут быть и «абордажным» орудием.
Совсем до недавнего времени считали, что жгутик бактерии виляет как хвост у змеи. Последние исследования показали, что жгутик бактерий устроен куда сложнее. Он работает как турбина. Присоединенный к приводу, он вращается в одном направлении. Привод, или жгутиковый мотор бактерии, — это сложная молекулярная структура, которая работает как мышца. С тем отличием, что мышца после совершения сжатия должна расслабляться, а мотор бактерии работает постоянно.
Наномеханизм работы жгутика
Не углубляясь в биохимию движения, отметим, что в создании привода жгутика учувствует до 240 белков, которые делятся на 50 молекулярных составляющих с определенной функцией в системе.
В этой двигательной системе бактерии есть ротор, который движется, и статор, который обеспечивает это движение. Есть приводной вал, втулка, сцепление, тормоза и ускорители
Этот миниатюрный двигатель позволяет бактерии совершать перемещения на расстояния в 35 раз большие, чем ее собственный размер всего лишь за 1 секунду. При этом на работу самого жгутика, совершающего 60 тысяч оборотов в минуту, организм тратит всего 0,1 % всей энергии, которую расходует клетка.
При этом на работу самого жгутика, совершающего 60 тысяч оборотов в минуту, организм тратит всего 0,1 % всей энергии, которую расходует клетка.
Удивительно еще и то, что все запчасти своего двигательного механизма бактерия может заменять и ремонтировать «на ходу». Просто представьте, что вы летите в самолете. А техники меняют лопасти работающего мотора.
Жгутиковая бактерия против Дарвина
Двигатель, способный работать со скоростью до 60 000 оборотов в минуту, самочинящийся и использующий в виде топлива всего лишь углеводы (сахара), имеющий устройство сродни электрическому двигателю — могло ли такое устройство возникнуть в процессе эволюции?
Именно этот вопрос задал себе в 1988 году доктор биологических наук Майкл Бихи. Он ввел в биологию понятие несократимой системы — системы, в которой все ее части одновременно необходимы для обеспечения ее работы, а удаление хоть одной части приводит к полному нарушению ее функционирования.
С позиции эволюции Дарвина, все структурные изменения в организме происходят постепенно и отбираются естественным отбором только успешные.
Выводы М. Бихи, изложенные в книге «Черный ящик Дарвина» (1996): двигатель жгутиковой бактерии — неделимая система из более чем 40 частей, и отсутствие хоть одной приведет к полной нефункциональности системы, а значит, эта система не могла произойти путем естественного отбора.
Бальзам для креационистов
Теория сотворения в изложении ученого и профессора биологии, декана факультета биологических наук в Лихайском университете Бетлехема (США) М. Бихи сразу привлекла внимание служителей церкви и сторонников теории божественного происхождения жизни.
В 2005 году в США даже состоялся судебный процесс, где Бихи выступал свидетелем со стороны сторонников теории «разумного дизайна», на котором рассматривался вопрос введения в школах Дувра изучения креационизма на курсе «О пандах и людях». Процесс был проигран, преподавание такого предмета признали противоречащим действующей конституции.
Но споры креационистов и эволюционистов продолжаются и сегодня.
Оглавление темы «Анатомия бактериальной клетки. Физиология бактерий.»:
Физиология бактерий.»: 1. Анатомия бактериальной клетки. Поверхностные структуры бактерии. Капсула бактерий. Организация капсул. Окраска капсул бактерий. Состав капсул. Антигенные свойства капсул.
3. Микроворсинки бактерий. Фимбрии бактерий. F-пили (секс-пили) бактерии. Клеточная оболочка бактерий. Гликокаликс.
4. Клеточная стенка бактерий. Функции клеточной стенки. Строение клеточной стенки бактерии. Пептидогликан. Муреиновый мешок. Структура пептидогликана (муреина)
5. Грамотрицательные бактерии. Клеточная стенка грамотрицательных бактерий. Строение клеточной стенки грамотрицательных бактерий.
6. Грамположительные бактерии. Клеточная стенка грамположительных бактерий. Строение клеточной стенки грамположительных бактерий. Аутолизины бактерий. Сферопласты. Протопласты.
7. Цитоплазматическая мембрана (ЦПМ) бактерии. Состав цитоплазматической мембраны бактерий. Транспортные системы. Мезосомы. Периплазматическое пространство.
8. Цитоплазма бактерий. Бактериальный геном. Бактериальные рибосомы. Запасные гранулы бактерии.
Бактериальные рибосомы. Запасные гранулы бактерии.
9. Физиология бактерий. Питание бактерий. Тип питания бактерии. Голозои. Голофиты. Вода. Значимость воды для бактерий.
10. Усваиваемые бактериальной клеткой соединения. Пути поступления веществ в бактериальную клетку. Пассивный перенос. Диффузия.
По характеру движения подвижные бактерии разделяют на плавающие и скользящие (ползающие ). Орган движения плавающих бактерий — жгутики ; подвижность скользящих бактерий обеспечивают волнообразные сокращения тела.
Расположение жгутиков — характерный признак, имеющий таксономическое значение. Варианты расположения жгутиков приведены на рис. 4-1. У некоторых бактерий жгутики расположены по всей поверхности клеточной стенки (например, у бактерий рода Proteus), такие бактерии известны как перитрихи [от греч. peri-, вокруг, + trichos, волос]. Некоторые бактерии снабжены только одним толстым жгутиком (например, представители рода Vibrio), они известны как монотрихи .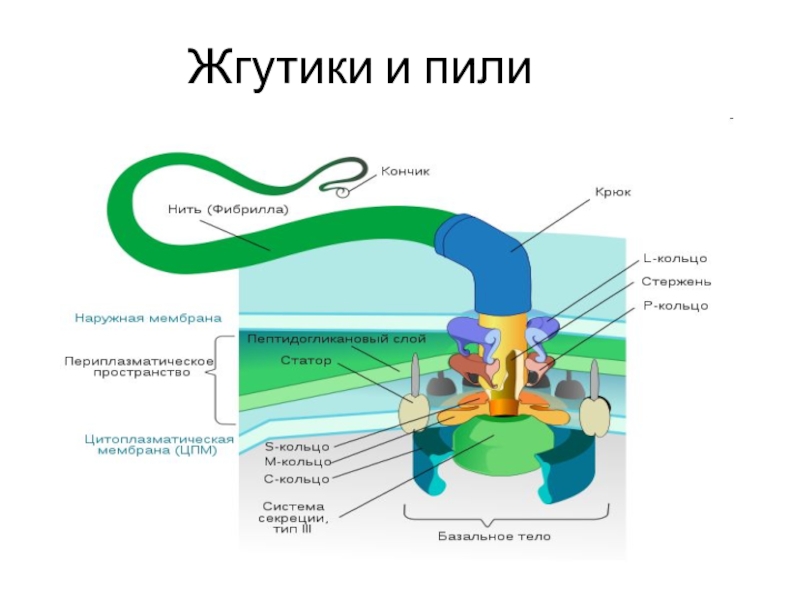 Политрихи — бактерии, имеющие одиночный по виду жгутик, образованный пучком из 2-50 жгутиков. Полярные жгутики прикреплены к одному или обоим концам бактерии. Монополярно-политрихиальное расположение жгутиков имеют лофотрихи [от греч. lophos, пучок, + trichos, волос], к ним, например, относят представителей рода Pseudomonas. Биполярно-политрихиальное жгутикование имеют амфитрихи [от греч. amphi-, двусторонний, + trichos, волос] (например, бактерии рода Spirillum).
Политрихи — бактерии, имеющие одиночный по виду жгутик, образованный пучком из 2-50 жгутиков. Полярные жгутики прикреплены к одному или обоим концам бактерии. Монополярно-политрихиальное расположение жгутиков имеют лофотрихи [от греч. lophos, пучок, + trichos, волос], к ним, например, относят представителей рода Pseudomonas. Биполярно-политрихиальное жгутикование имеют амфитрихи [от греч. amphi-, двусторонний, + trichos, волос] (например, бактерии рода Spirillum).
Жгутик — спирально изогнутая полая нить, образованная субъединицами флагеллина. У разных бактерий толщина жгутиков варьирует от 12 до 18 нм, что составляет не более 1/10 диаметра жгутиков водорослей и простейших. Жгутики также различают по длине и диаметру витка. Место прикрепления жгутика к бактериальной клетке имеет сложное строение и состоит из базальной структуры и так называемого «крюка» (рис. 4-2). У грамположительных бактерий в состав базальной структуры входит одна пара, а у грамотрицательных бактерий — две пары колец. Кольца играют роль «приводного диска» и «подшипника». Вся конструкция выполняет функцию хемомеханического преобразователя (флагеллиновый мотор). У спирохет за движение ответственна особая органел-ла — осевая нить, состоящая из двух рядов бактериальных жгутиков, расположенных продольно внутри клетки.
4-2). У грамположительных бактерий в состав базальной структуры входит одна пара, а у грамотрицательных бактерий — две пары колец. Кольца играют роль «приводного диска» и «подшипника». Вся конструкция выполняет функцию хемомеханического преобразователя (флагеллиновый мотор). У спирохет за движение ответственна особая органел-ла — осевая нить, состоящая из двух рядов бактериальных жгутиков, расположенных продольно внутри клетки.
Бактериальные жгутики совершают поступательные и вращательные движения, проталкивая бактерии через среду подобно корабельному винту. Они также могут изменять направление вращения и тянуть клетку подобно пропеллеру. Скорость обратного движения в четыре раза меньше скорости поступательного движения. Некоторые перитрихи могут перемещаться по поверхности агара, то есть плавающие бактерии способны к передвижению по поверхности твёрдых сред. В частности, Proteus vulgaris распространяется по поверхности агара, образуя тонкий налёт (напоминающий таковой при выдохе на холодное стекло), а неподвижные штаммы протея лишены такой способности.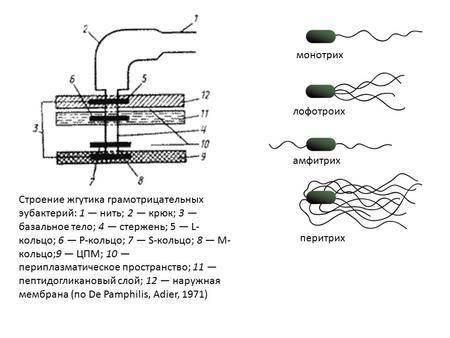 Это явление получило название «феномен роения », а наблюдение за ним легло в основу некоторых понятий бактериальной серодиагностики. Так, жгутиковые Аг называют Н-Аг [от нем. Hauch, выдох, налёт], а Аг клеточной поверхности — О-Аг [от нем. фпе Hauch, без налёта1.
Это явление получило название «феномен роения », а наблюдение за ним легло в основу некоторых понятий бактериальной серодиагностики. Так, жгутиковые Аг называют Н-Аг [от нем. Hauch, выдох, налёт], а Аг клеточной поверхности — О-Аг [от нем. фпе Hauch, без налёта1.
Рис. 4-2. Схема строения бактериального жгутика . БС — базальная структура, ВМ — внешняя мембрана, ЦПМ — цитоплазматическая мембрана, Р — ротор, О — ось, КО — кольцо жгутикового мотора, КР — крюк, С — цилиндрики-соединители, Н — нить жгутика, Ш — шапочка.
Способность бактерий к целенаправленному движению генетически обусловлена. Например, у Escherichia coli в регуляцию этого процесса вовлечено 3% генома (приблизительно 50 генов). Эти гены кодируют белки, образующие локомоторный аппарат, а также белки и ферменты, участвующие в преобразовании сигналов. Для жгутикового аппарата характерна периодическая изменчивость. Во многом этот процесс носит адаптивный характер и наиболее выражен у патогенных микроорганизмов. В частности, некоторые бактерии выработали систему вариабельности антигенных характеристик жгутиков, позволяющую им на какое-то время избегать направленных эффектов защитных иммунных механизмов.
В частности, некоторые бактерии выработали систему вариабельности антигенных характеристик жгутиков, позволяющую им на какое-то время избегать направленных эффектов защитных иммунных механизмов.
Лабораторная диагностика подвижности бактерий
Подвижность бактерий определяют микроскопией препаратов в «раздавленной » или «висячей » капле. Способность к движению можно определять также после внесения культуры бактерий уколом в столбик полужидкого агара (подвижные виды растут по всей толще среды, неподвижные — по уколу) или посевом бактерий в водный конденсат скошенного столбика агара (подвижные виды переплывают из конденсата на поверхность среды и колонизируют её), либо определяют способность бактерий давать «феномен роения ».
Клетки могут перемещаться при помощи специализированных органоидов, к которым относятся реснички и жгутики. Реснички клеток всегда многочисленны (у простейших их количество исчисляется сотнями и тысячами), а длина составляет 10-15мкм. Жгутиков же чаще всего 1-8, длина их — 20-50мкм.
Жгутиков же чаще всего 1-8, длина их — 20-50мкм.
Строение и функции органоидов движения
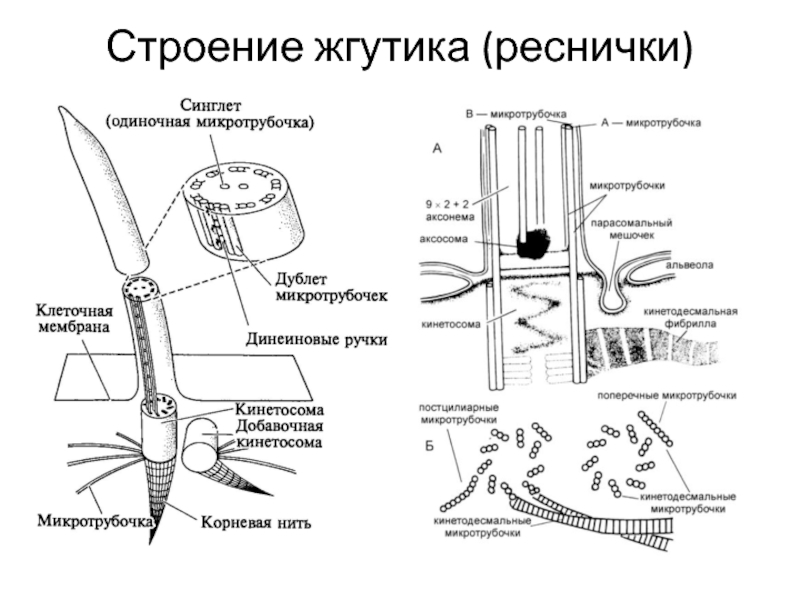
Строение ресничек и жгутиков, как у растительных, так и животных клеток сходно. Под электронным микроскопом обнаружено, что реснички и жгутики это немембранные органоиды, состоящие из микротрубочек. Две из них располагаются в центре, а вокруг них по периферии лежат еще 9 пар микротрубочек. Вся эта структура покрыта цитоплазматической мембраной, являющейся продолжением клеточной мембраны.
Жгутики и реснички обеспечивают не только передвижение клеток в пространстве, но и перемещение различных веществ на поверхности клеток, а также попадание пищевых частиц в клетку. У основания ресничек и жгутиков находятся базальные тельца, которые тоже состоят из микротрубочек.
Предполагают, что базальные тельца являются центром формирования микротрубочек жгутиков и ресничек. Базальные тельца, в свою очередь, нередко происходят из клеточного центра.
Большое количество одноклеточных организмов и некоторые клетки многоклеточных не имеют специальных органоидов движения и передвигаются при помощи псевдоподий (ложноножек), которое получило название амебоидного.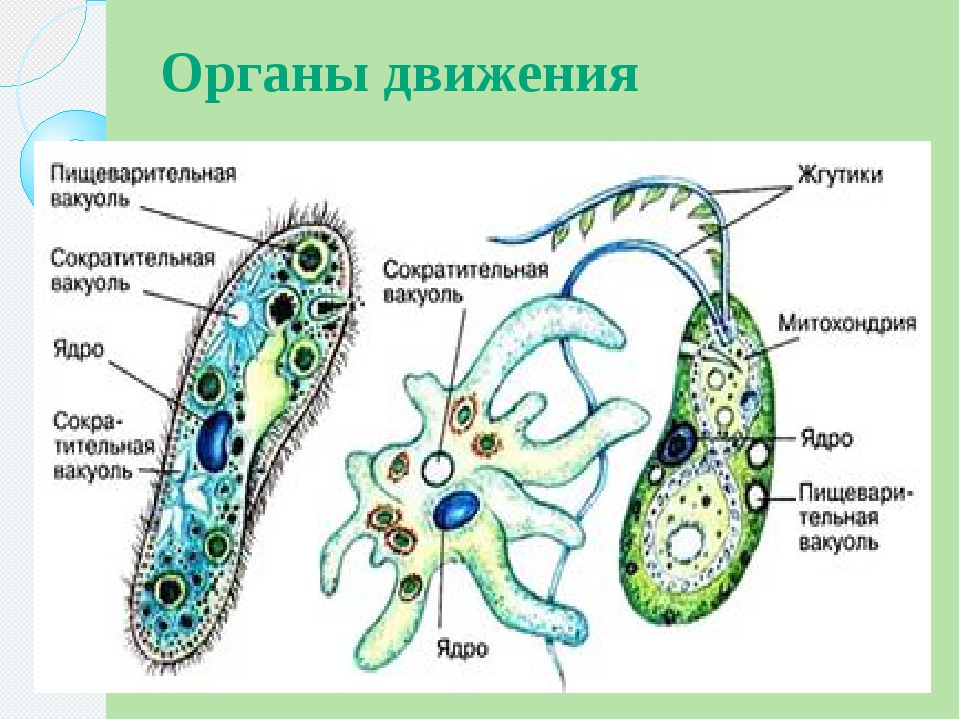 В основе его лежит движение молекул особых белков, называемых сократимыми.
В основе его лежит движение молекул особых белков, называемых сократимыми.
Особенности движения простейших
Одноклеточные организмы также способны передвигаться (инфузория туфелька, эвглена зеленая, амеба обыкновенная). Для перемещения в толще воды каждая особь наделена специфическими органоидами. У простейших такими органоидами являются реснички, жгутики, ложноножки.
Эвглена зелёная
Эвглена зелёная — представитель простейших из класса жгутиковых. Тело эвглены веретенообразной формы, удлиненное с заостренным концом. Органоиды движения эвглены зеленой представлены жгутиком, который находится на тупом конце. Жгутики — это тонкие выросты тела, число которых варьирует от одного до десятков.
Механизм движения при помощи жгутика отличается у разных видов. В основном это вращение в виде конуса, вершина которого обращена к телу. Перемещение наиболее эффективно при достижении углом вершины конуса 45°. Скорость колеблется в пределах от 10 до 40 оборотов за секунду. Часто наблюдается помимо вращательного движения жгутика, также его волнообразные покачивания.
Часто наблюдается помимо вращательного движения жгутика, также его волнообразные покачивания.
Такой характер движения свойствен для одножгутиковых видов. У многожгутиковых нередко жгутики располагаются в одной плоскости и не формируют конуса вращения.
Микроскопическое строение жгутиков довольно сложное. Они окружены тонкой оболочкой, которая является продолжением наружного слоя эктоплазмы — пелликулы. Внутреннее пространство жгутика заполнено цитоплазмой и продольно расположенными нитями — фибриллами.
Периферически расположенные фибриллы отвечают за осуществление движения, а центральные выполняют опорную функцию.
Инфузория туфелька
Передвигается инфузория туфелька за счет ресничек, осуществляя ими волнообразные движения. Направляется вперед тупым концом.
Реснички двигаются в одной плоскости и делают прямой удар после полного выпрямления, а возвратный — в выгнутом положении. Удары идут последовательно один за другим с небольшой задержкой. Во время плаванья, инфузория осуществляет вращательные движения вокруг продольной оси.
Перемещается туфелька со скоростью до 2,5мм/c. Направленность меняется за счёт перегибов тела. Если на пути будет преграда, то после столкновения инфузория начинает двигаться в противоположную сторону.
Все реснички инфузорииимеют сходное строение с жгутиками эвглены зеленой. Ресничка у основания образует базальное зерно, которое играет важную роль в механизме движения организма.
У некоторых инфузорий реснички соединяются между собой и таким образом позволяют развить большую скорость.
Инфузории относятся к высокоорганизованным простейшим и свою двигательную активность они осуществляют с помощью сокращений. Форма тела простейшего может меняться, а после возвращаться в прежнее состояние. Быстрые сократительные движения возможны благодаря наличию особых волокон — мионем.
Амеба обыкновенная
Амеба — простейшее довольно крупных размеров (до 0,5мм). Форма тела полиподиальная, обусловлена наличием множественных псевдоподий — это выросты с внутренней циркуляцией цитоплазмы.
У амебы обыкновенной псевдоподии еще называют ложноножками. Направляя ложноножки в разные стороны, амёба развивает скорость в 0,2 мм/минуту.
К органоидам движения простейших не относятся цитоплазма, ядро, вакуоли, рибосомы, лизосомы, ЭПР, Аппарат Гольджи.
Специализированные органеллы и структуры клетки
Специализированные органеллы и структуры встречаются не во всех клетках.
Они характерны для зрелых клеток, являются признаками направления их дифференцировки и обеспечивают в них специфические функции. Примерами таких органелл являются микроворсинки, реснички, жгутики, миофибриллы, тонофибриллы, нейрофибриллы и некоторые другие органеллы.
Микроворсинки. Это структуры клетки, располагающиеся на ее внешней поверхности и выступающие во внеклеточное пространство. При световой микроскопии микроворсинки видны как тонкие выросты клетки. Если их много, то они формируют апикальную каемку на свободной поверхности. Эти выпячивания значительно расширяют площадь взаимодействия клетки с внешней средой.
Ферменты, прикрепленные к гликокаликсу и находящиеся в толще билипидного слоя мембраны микроворсинок, обеспечивают всасывание и/или переваривание веществ на поверхности клеток. В этом случае расширение контактной поверхности резко увеличивает эффективность подобных процессов, например комплекс микроворсинок в столбчатых эпителиоцитах тонкой кишки. В них микроворсинки тесно соприкасаются друг с другом, обильно покрывают внешнюю поверхность клетки. Толщина микроворсинок около 100 нм, а число и длина различны. Так, длина микроворсинок у столбчатых (призматических) клеток кишечника достигает 0,6…0,8 мкм.
Во многих клетках величина и размеры микроворсинок не постоянны. Так, в тироцигах щитовидной железы в период покоя они редкие и короткие, а при интенсивной нагрузке их высота и количество значительно увеличиваются.
Микроворсинки состоят из клеточной мембраны, гиалоплазмы и тонких микрофиламентов. Актиновые (тонкие) микрофиламенты располагаются параллельно поверхности мембраны в виде компактно упакованных, упорядоченных пучков. Внутри каждой микроворсинки располагается около 20…30 актиновых нитей. Положительный полюс микрофиламентов направлен к периферии и стыкуется с электронно-плотным аморфным веществом дистальной части (верхушкой), а в основании микроворсинки актиновые микрофиламенты вплетаются в сеть подобных им структур, формирующих кутикулу.
Внутри каждой микроворсинки располагается около 20…30 актиновых нитей. Положительный полюс микрофиламентов направлен к периферии и стыкуется с электронно-плотным аморфным веществом дистальной части (верхушкой), а в основании микроворсинки актиновые микрофиламенты вплетаются в сеть подобных им структур, формирующих кутикулу.
Тонкие микрофиламенты в микроворсинке лежат параллельно друг другу на расстоянии около 10 нм, регулярно соединяясь между собой с помощью белков — фимбрина и фасцина. Эти белковые комплексы образуют поперечные сцепления и объединяют тонкие микрофиламенты в компактные пучки. С мембранами тонкие микрофиламенты взаимодействуют с помощью минимиозина и виллина. Взаимодействие с минимиозинами позволяет микроворсинке сокращаться (уменьшать или увеличивать высоту).
Сходное строение с микроворсинками имеют стереоцилии. Они крупнее микроворсинок и не обладают всасывающей способностью.
Реснички и жгутики. Они представляют собой выпячивания цитоплазмы, окруженные клеточной мембраной, способные к активному движению. Органеллы хорошо заметны при большом увеличении микроскопа. Реснички и жгутики на ультраструктурном уровне имеют сходные принципы строения, но могут иметь разные функции. Реснички перемещают поверхностный субстрат полого органа, тогда как жгутик спермия позволяет передвигаться самой клетке.
Органеллы хорошо заметны при большом увеличении микроскопа. Реснички и жгутики на ультраструктурном уровне имеют сходные принципы строения, но могут иметь разные функции. Реснички перемещают поверхностный субстрат полого органа, тогда как жгутик спермия позволяет передвигаться самой клетке.
В многоклеточных организмах животных строение ресничек и жгутиков резко отличается от подобных органелл прокариот. Жгутики у бактерий образованы белком флагеллином, не имеющим отношения к комплексам микротрубочек у эукариот.
Реснички у эукариот — это специальные органеллы движения, встречающиеся лишь в некоторых клетках. Реснички находятся в однослойном эпителии органов дыхания и женских половых путей. В реснитчатом эпигелиоците дыхательных путей можно найти около 50…60 ресничек.
Ресничка представляет собой тонкий цилиндрический вырост цитоплазмы с постоянным диаметром 300 нм, покрытый плазматической мембраной.
В основании ресничек и жгутика в цитоплазме видны хорошо окрашивающиеся мелкие гранулы — базальные тельца.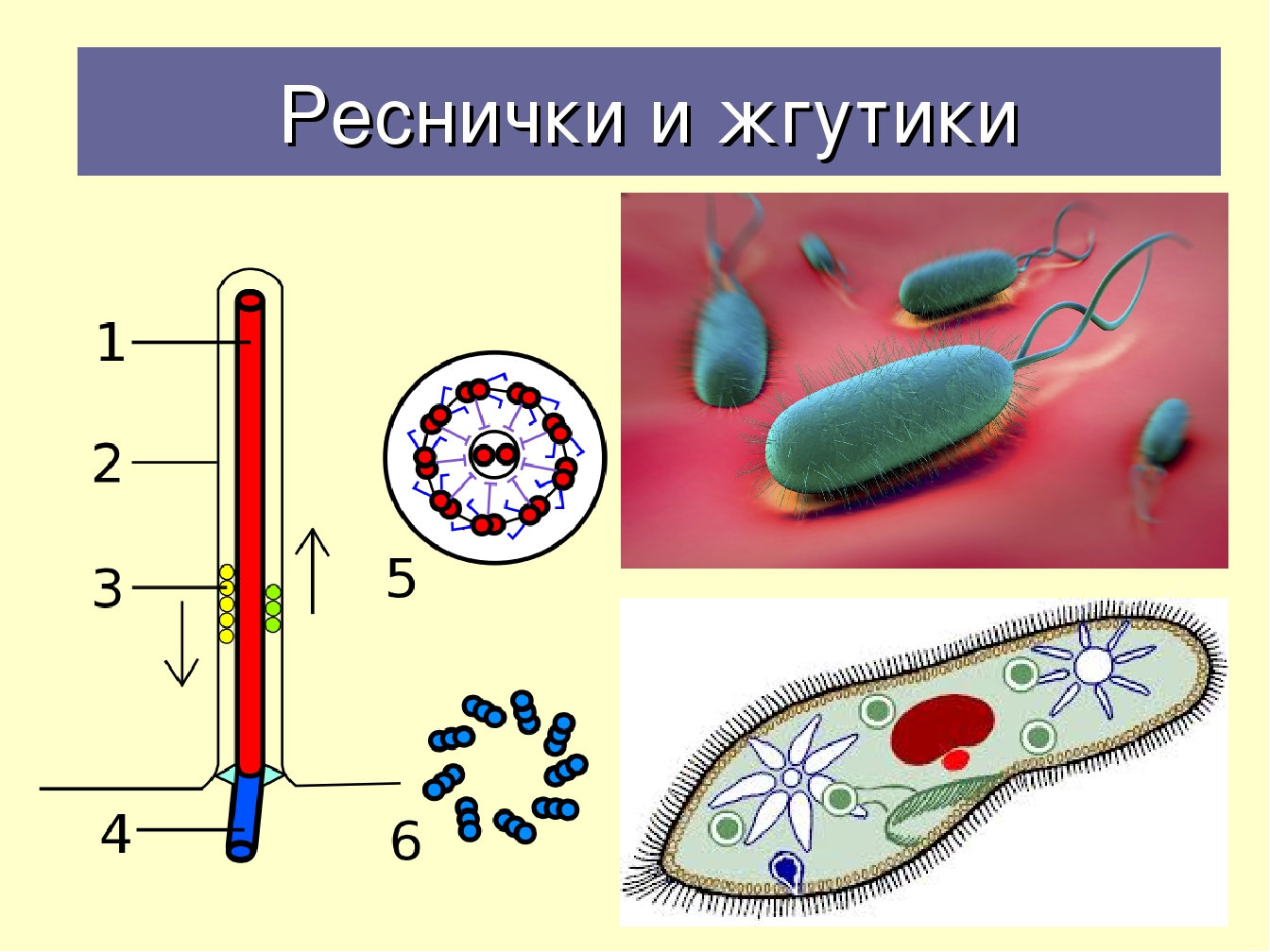 В этой области над клеткой выступает выпячивание — аксонема. Аксонема («осевая нить») — сложная структура, состоящая из микротрубочек и выступающая в просвет или полость органа, выстланного реснитчатым эпителием. Проксимальная часть реснички (базальное тело) погружена в цитоплазму. Диаметры аксонемы и базального тельца одинаковые.
В этой области над клеткой выступает выпячивание — аксонема. Аксонема («осевая нить») — сложная структура, состоящая из микротрубочек и выступающая в просвет или полость органа, выстланного реснитчатым эпителием. Проксимальная часть реснички (базальное тело) погружена в цитоплазму. Диаметры аксонемы и базального тельца одинаковые.
Базальное тельце по строению аналогично центриоли и состоит из девяти триплетов микротрубочек, между которыми расположены ручки, втулки и спицы. К базальному тельцу могут прикрепляться спутники, от которых отходят микротрубочки. Таким образом, базальное тельце, наряду с центросомой, является центром организации микротрубочек и может выполнять аналогичные функции.
Аксонема по составу опорного аппарата отличается от базального тельца. Стенку цилиндра аксонемы образуют 9 дуплетов микротрубочек. Кроме периферических дуплетов микротрубочек в центре аксонемы располагается пара центральных микротрубочек. Эти микротрубочки лежат отдельно на расстоянии примерно 25 нм.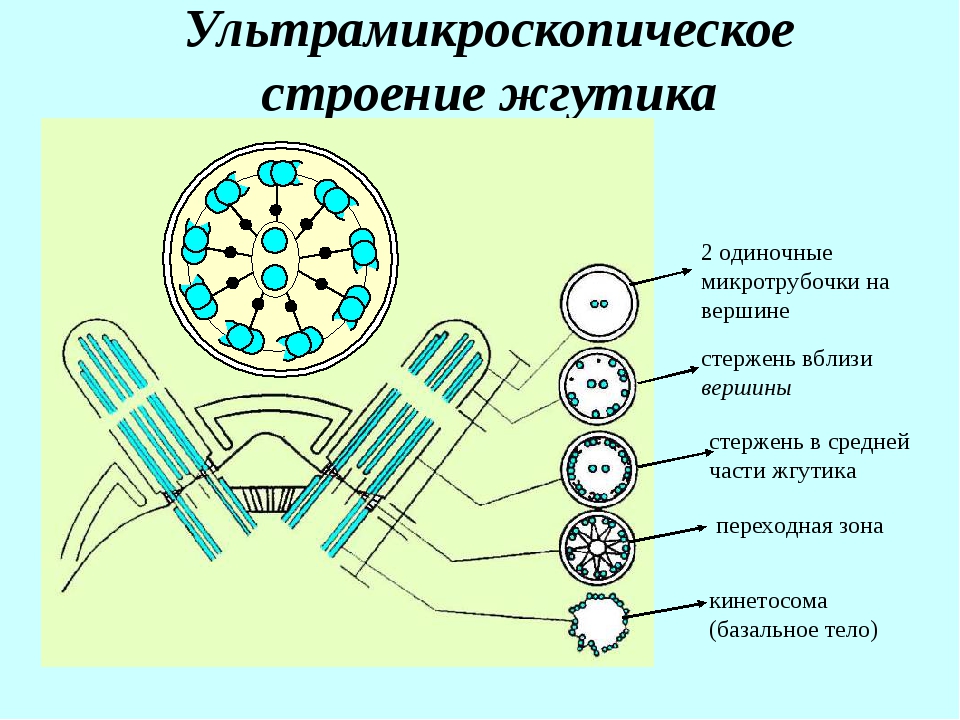 В целом систему микротрубочек реснички описывают как (9 х 2 + 2) в отличие от (9 х 3 + 0) системы центриолей и базальных телец.
В целом систему микротрубочек реснички описывают как (9 х 2 + 2) в отличие от (9 х 3 + 0) системы центриолей и базальных телец.
Базальное тельце и аксонема структурно связаны друг с другом и составляют единое целое. Каждая из двух первых микротрубочек девяти триплетов базального тельца является основой для дуплетов микротрубочек цитоплазматического выроста, таким образом, две микротрубочки триплетов базального тельца являются микротрубочками дуплетов аксонемы. Они полимеризуются от базального тельца на основании реснички.
Из дуплета а-микротрубочка полная и образована 13 тубулинами. Неполная b-микротрубочка составлена 11 тубулинами, так как две глобулы белка являются общими с а-микротрубочками. Центральные микротрубочки формируются от центральной втулки базального тельца. Все микротрубочки достигают своим дистальным отрицательным полюсом гомогенный матрикс аксонемы.
Соседние дуплеты соединены между собой «ручками», образованными белками динеинами. Динеины обладают АТФазной активностью, способны изменять свою стереологическую структуру.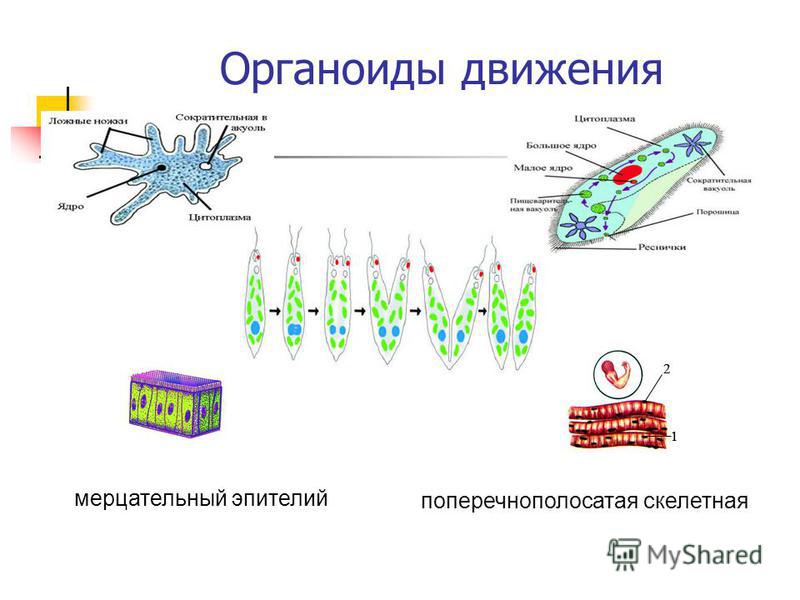 В присутствии ионов кальция динеины сокращаются с потреблением энергии. Это позволяет микротрубочкам скользить относительно друг друга. Кроме динеина в составе ресничек выделяют нидоген.
В присутствии ионов кальция динеины сокращаются с потреблением энергии. Это позволяет микротрубочкам скользить относительно друг друга. Кроме динеина в составе ресничек выделяют нидоген.
К центральным микротрубочкам от периферических дуплетов радиально направляются спицы. Центральные же микротрубочки объединены втулкой.
Свободные клетки, имеющие реснички и жгутики, способны передвигаться, а неподвижные клетки движением ресничек могут перемещать жидкость и различные частицы в полых органах. При движении ресничек и жгутиков длина их не уменьшается, поэтому неправильно называть это движение сокращением. Траектория движения ресничек очень разнообразна: маятникообразная, крючкообразная или волнообразная.
Основной белок ресничек — тубулин не способен к сокращению, укорочению, поэтому движение ресничек осуществляется за счет активности белка динеина. Незначительные смещения дуплетов микротрубочек относительно друг друга вызывают изгиб всей реснички, а если такое локальное смещение происходит вдоль жгутика, то возникает волнообразное движение.
Волнообразное движение жгутика спермия позволяет ему передвигаться с очень высокой скоростью — до 5 мм в минуту.
Волнообразное перемещение ресничек мерцательного эпителия происходит строго согласованно, но скорость движения в разных направлениях отличается. Обычно в какую-то одну сторону реснички сокращаются с большой скоростью, а в противоположном направлении их положение изменяется плавно. Это обеспечивает ток жидкости в сторону быстрого сокращения ресничек. Дефекты ресничек могут приводить к различным нарушениям, например к наследственному рецидивирующему бронхиту и хроническому синуситу, возникающим в результате нарушений функции ресничного эпителия.
Движения ресничек и жгутиков зависят от содержания внутриклеточного кальция, АТФ, ионного состава межклеточного вещества, обеспечения кислородом, глюкозой и др.
Гормоны и биологически активные вещества регулируют движения ресничек и жгутиков. Влияние того или иного гормонального фактора зависит от специализации клетки и ее рецепторного аппарата.
Реснички образуются за счет центриолей. От материнской центриоли синтезируется дочерняя центриоль, но процесс этот не заканчивается на дуплете органелл, а вновь синтезированная органелла смещается на периферию (в субмембранное пространство). Процесс может повторяться многократно. В результате под мембраной образуется множество базальных телец, которые служат основой для образования ресничек.
Базальные инвагинации (впячивания). Это внедрения цитолеммы в цитоплазму, прилежащие к базальной мембране или иной плотной структуре. Чаще всего базальные впячивания встречаются в эпителии в базальной части клетки. В других тканях они встречаются гораздо реже, например впячивание цитолеммы на остеокласте со стороны разрушаемой (резорбцируемой) кости, так называемая гофрированная каемка. Базальные инвагинации существенно увеличивают внутреннюю (базальную) поверхность клетки.
Нередко в зонах инвагинации цитолеммы видны многочисленные митохондрии. Совокупность базальных инвагинаций и митохондрий формирует базальную исчерченность, которая хорошо заметна при большом увеличении микроскопа в дистальных и проксимальных канальцах нефронов почек. Базальная исчерченность в эпителиоцитах канальцев — это признак активных процессов трансмембранного переноса веществ; наряду с увеличением поверхности присутствует высокий уровень энергетического потребления, обеспечиваемый АТФ.
Базальная исчерченность в эпителиоцитах канальцев — это признак активных процессов трансмембранного переноса веществ; наряду с увеличением поверхности присутствует высокий уровень энергетического потребления, обеспечиваемый АТФ.
Базальные впячивания формируют сложный лабиринт каналов и ходов, взаимных переплетений. На поверхности клеточной мембраны обнаруживают много ионных каналов, рецепторов, значительную ферментативную активность.
Форму базальных впячиваний поддерживают структуры цитоскелета: промежуточные филаменты и тонкие микрофиламенты. Они соединяются с внутренней поверхностью мембраны с помощью интегральных мембранных белков, которые, в свою очередь, сцеплены друг с другом через гликокаликс. Состав промежуточных филаментов разнообразен и зависит от тканевой принадлежности клетки: в эпителии — это цитокератины, которые формируют микрофибриллы (гонофибриллы).
Миофибрилла. Это специализированная органелла мышечной ткани, основная функция которой — сокращение. При световой микроскопии миофибриллы заметны в скелетной и сердечной мышечной тканях, где они имеют вид мощных пучков волокон, расположенных строго упорядоченно и придающих симпластам или клеткам продольную и поперечную исчерченность.
При световой микроскопии миофибриллы заметны в скелетной и сердечной мышечной тканях, где они имеют вид мощных пучков волокон, расположенных строго упорядоченно и придающих симпластам или клеткам продольную и поперечную исчерченность.
Миофибрилла представляет собой систему взаимодействующих друг с другом тонких и толстых микрофиламентов (миофиламентов). Отдельные миофиламенты можно рассмотреть лишь при электронной микроскопии, но каждая миофибрилла (особенно в поперечнополосатых мышечных тканях) состоит из сотен таких нитей. Распределение миофиламентов может быть строго упорядочено в продольном направлении и придавать структуре поперечную и продольную исчерченность либо распределяться в виде сети (в гладких мышечных клетках). В последнем случае миофиламенты при световой микроскопии выявить не удается.
Сокращение в миофибрилле обеспечивается взаимодействием белков, образующих миофиламенты. Сокращение — это энергоемкий процесс, зависящий от внутриклеточного содержания ионов кальция. Сокращение может быть вызвано прямым нервным либо гуморальным влиянием. Любое из специфических возбуждений сопровождается деполяризацией мембраны с увеличением внутриклеточного содержания ионов кальция, что активизирует взаимодействие актина с миозином.
Сокращение может быть вызвано прямым нервным либо гуморальным влиянием. Любое из специфических возбуждений сопровождается деполяризацией мембраны с увеличением внутриклеточного содержания ионов кальция, что активизирует взаимодействие актина с миозином.
Рассмотрим строение миофибриллы на примере скелетного мышечного волокна (симпласта). В скелетном мышечном волокне миофибрилла продолжается на всю длину симпласта. Миофибриллы поперечнополосатых мышечных тканей имеют поперечную исчерченность, которая формируется при чередовании светлых (изотропных, I-дисков) и темных (анизотропных, A-дисков) дисков.
Анизотропные диски в поляризованном свете обеспечивают двойное лучепреломление, а изотропные такой способностью не обладают. Светлые диски при световой микроскопии имеют вид слабо окрашенных полос. На электронной микрофотографии видно, что светлые диски не содержат толстых нитей (миофиламентов). При очень большом увеличении светового микроскопа иногда в центре светлого I-диска заметна темная линия — телофрагма, или Z-линия (зона соединения между собой тонких нитей).
Темные диски при электронной микроскопии видны как зона параллельно лежащих толстых миофиламентов. Значительная часть темного диска содержит как толстые, так и тонкие миофиламенты. При световой и электронной микроскопиях эти участки просматриваются как наиболее темные. В центре темного А-диска под очень большим увеличением светового микроскопа иногда можно увидеть мезофрагму, или М-линию — область соединения толстых нитей. Она видна как тончайшая темная поперечная полоска. Участок A-диска, в котором отсутствуют тонкие нити, называется Н-зоной. В отличие от темного диска в целом Н-зона несколько светлее окрашена.
Соотношение длин анизотропного и изотропного дисков величина непостоянная и зависит от степени сокращения или расслабления. Так, в момент максимального сокращения длина изотропного диска минимальна, а Н-зона отсутствует. При расслаблении длина изотропного диска и Н-зоны максимальна. Размеры анизотропного диска при этом остаются относительно постоянными.
Толстые нити (миофиламенты) имеют поперечный диаметр около 10…12 нм, они образованы сложно устроенными белками — миозинами. Каждая молекула миозина содержит две тяжелые и две легкие цепи полипептидов меромиозинов, то есть миозин — это тетрамер. Он состоит из тела (хвоста), шейки и головки. Шейка и тело сформированы из двух взаимно переплетающихся полипептидных цепочек тяжелых цепей меромиозинов. Головка раздвоена и к ней присоединяются две легкие цепи меромиозина. Легкий меромиозин (миозин) способен разрушать АТФ, то есть обладает АТФазной активностью. Между шейкой и головкой миозина есть «шарнирное» соединение — место, легко изменяющее свою пространственную ориентацию или изгибающееся подобно суставу. Это происходит в момент взаимодействия головки с белками тонкого миофиламенга — актинами.
Каждая молекула миозина содержит две тяжелые и две легкие цепи полипептидов меромиозинов, то есть миозин — это тетрамер. Он состоит из тела (хвоста), шейки и головки. Шейка и тело сформированы из двух взаимно переплетающихся полипептидных цепочек тяжелых цепей меромиозинов. Головка раздвоена и к ней присоединяются две легкие цепи меромиозина. Легкий меромиозин (миозин) способен разрушать АТФ, то есть обладает АТФазной активностью. Между шейкой и головкой миозина есть «шарнирное» соединение — место, легко изменяющее свою пространственную ориентацию или изгибающееся подобно суставу. Это происходит в момент взаимодействия головки с белками тонкого миофиламенга — актинами.
Тонкий миофиламент по строению близок к тонким микрофиламентам и состоит из двойной цепочки актинов. Они спирально закручены. В отличие от обычных актиновых нитей тонкие миофиламенты достаточно стабильные структуры и не подвергаются постоянному распаду и полимеризации. Эта устойчивость объясняется присоединением к актиновым цепочкам их стабилизирующего белка — тропомиозина (фибриллярного белка).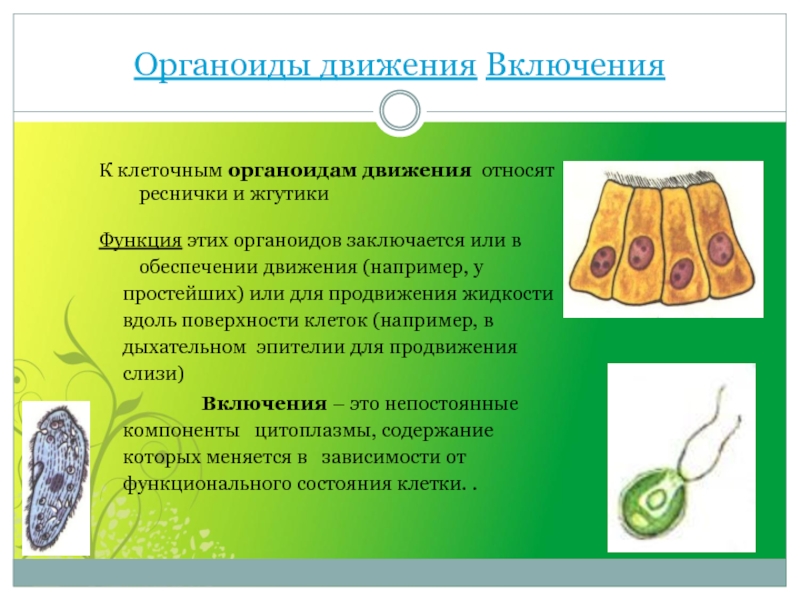
Кроме него в тонком миофиламенте имеются и другие белки — тропонины, составляющие комплекс из трех глобул. Эти глобулы представлены С-, I- и Т-тропонинами. С-тропонин связывается с ионами кальция, I-тропонин препятствует взаимодействию актина с головкой миозина, а Т-тропонин присоединяется к тропомиозину. В покое актины связаны с тропомиозином и тропонинами так, что актин блокирован и не может взаимодействовать с миозином.
При возбуждении мышечной клетки в матриксе цитоплазмы резко увеличивается содержание ионов кальция. Они соединяются с С-тропонином, к которому имеют высокую степень сродства. Это изменяет форму тропонинового комплекса, что сопровождается перестройкой пространственной конфигурации тропомиозина и изменением формы актиновой нити в целом. В результате молекулы актина могут взаимодействовать с головками миозина. Миозиновые головки соединяются с ближайшими актинами, но при этом происходит сокращение миозинов в зонах шарнирных соединений. В результате толстая нить слегка продвигается вперед в направлении центральной части изотропного диска.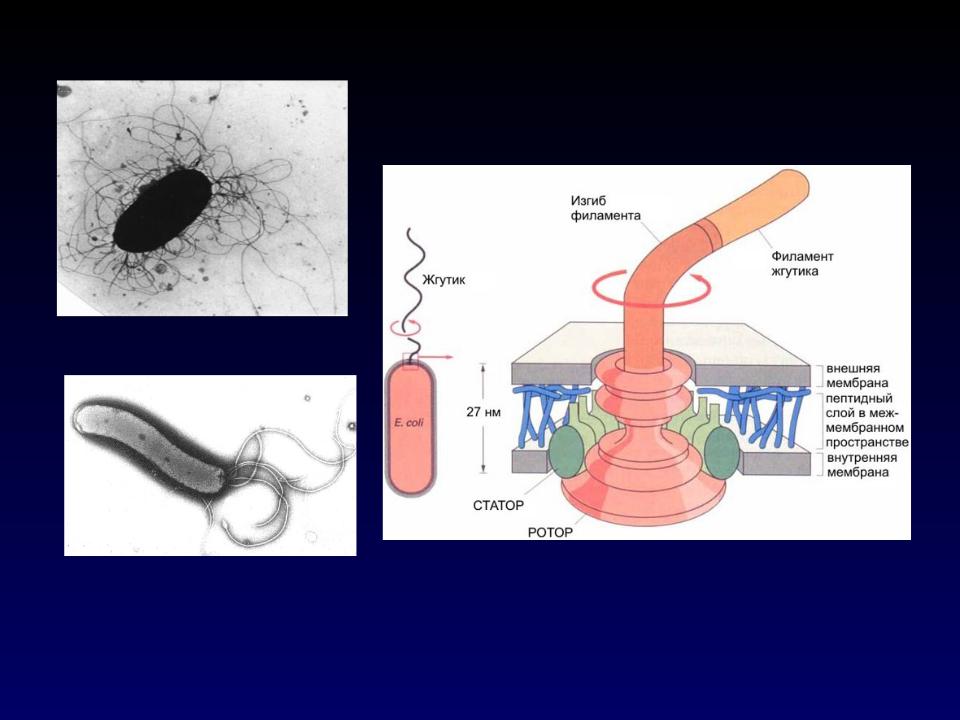
Следующим шагом является разрушение АТФ легкими цепями меромиозина. Этой энергии хватает, чтобы разорвать связь миозина с актином. Шарнирное соединение «выпрямляется», то есть занимает исходное положение, но оказавшаяся чуть впереди головка вновь связывается с последующими молекулами актина. Вновь происходит сокращение и движение вперед. Таким образом, миозиновые головки как бы «шагают» по актиновым нитям за счет шарнирного соединения и АТФазной активности миозина.
При прекращении возбуждения в мышечном волокне содержание ионов кальция вновь снижается, С-тропонин высвобождается, это приводит к тому, что актиновый (тонкий) миофиламент вновь восстанавливает свою прежнюю структуру, и актин «закрывается» тропомиозином. В этой ситуации взаимодействие миозина с актином становится вновь невозможным, и миофибрилла занимает исходное положение — происходит расслабление мышечного волокна.
Распределение миофибрилл в скелетной мышце отличается от сердечной. В кардиомиоцитах миофибриллы занимают в основном периферию клетки, тогда как в скелетном симпласте они располагаются центрально. В гладком миоците толстых миофиламентов фактически нет, и тонкие миофиламенты взаимодействуют с молекулами минимиозина.
В гладком миоците толстых миофиламентов фактически нет, и тонкие миофиламенты взаимодействуют с молекулами минимиозина.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Википедия — свободная энциклопедия
Избранная статья
Прохождение Венеры по диску Солнца — разновидность астрономического прохождения (транзита), — имеет место тогда, когда планета Венера находится точно между Солнцем и Землёй, закрывая собой крошечную часть солнечного диска. При этом планета выглядит с Земли как маленькое чёрное пятнышко, перемещающееся по Солнцу. Прохождения схожи с солнечными затмениями, когда наша звезда закрывается Луной, но хотя диаметр Венеры почти в 4 раза больше, чем у Луны, во время прохождения она выглядит примерно в 30 раз меньше Солнца, так как находится значительно дальше от Земли, чем Луна. Такой видимый размер Венеры делает её доступной для наблюдений даже невооружённым глазом (только с фильтрами от яркого солнечного света), в виде точки, на пределе разрешающей способности глаза. До наступления эпохи покорения космоса наблюдения этого явления позволили астрономам вычислить расстояние от Земли до Солнца методом параллакса, кроме того, при наблюдении прохождения 1761 года М. В. Ломоносов открыл атмосферу Венеры.
До наступления эпохи покорения космоса наблюдения этого явления позволили астрономам вычислить расстояние от Земли до Солнца методом параллакса, кроме того, при наблюдении прохождения 1761 года М. В. Ломоносов открыл атмосферу Венеры.
Продолжительность прохождения обычно составляет несколько часов (в 2004 году оно длилось 6 часов). В то же время, это одно из самых редких предсказуемых астрономических явлений. Каждые 243 года повторяются 4 прохождения: два в декабре (с разницей в 8 лет), затем промежуток в 121,5 года, ещё два в июне (опять с разницей 8 лет) и промежуток в 105,5 года. Последние декабрьские прохождения произошли 9 декабря 1874 года и 6 декабря 1882 года, а июньские — 8 июня 2004 года и 6 июня 2012 года. Последующие прохождения произойдут в 2117 и 2125 годах, опять в декабре. Во время прохождения наблюдается «явление Ломоносова», а также «эффект чёрной капли».
Хорошая статья
Резня в Благае (сербохорв. Масакр у Благају / Masakr u Blagaju) — массовое убийство от 400 до 530 сербов хорватскими усташами, произошедшее 9 мая 1941 года, во время Второй мировой войны. Эта резня стала вторым по счету массовым убийством после создания Независимого государства Хорватия и была частью геноцида сербов.
Эта резня стала вторым по счету массовым убийством после создания Независимого государства Хорватия и была частью геноцида сербов.
Жертвами были сербы из села Велюн и его окрестностей, обвинённые в причастности к убийству местного мельника-хорвата Йосо Мравунаца и его семьи. Усташи утверждали, что убийство было совершено на почве национальной ненависти и свидетельствовало о начале сербского восстания. Задержанных сербов (их число, по разным оценкам, составило от 400 до 530 человек) содержали в одной из школ Благая, где многие из них подверглись пыткам и избиениям. Усташи планировали провести «народный суд», но оставшаяся в живых дочь Мравунаца не смогла опознать убийц среди задержанных сербов, а прокуратура отказалась возбуждать дело против кого-либо без доказательства вины. Один из высокопоставленных усташей Векослав Лубурич, недовольный таким развитием событий, организовал новый «специальный суд». День спустя дочь Мравунаца указала на одного из задержанных сербов. После этого 36 человек были расстреляны. Затем усташи казнили остальных задержанных.
Затем усташи казнили остальных задержанных.
Изображение дня
Эхинопсисы, растущие на холме посреди солончака Уюни
Органоиды движения — строение, характеристики и функции
Органоиды движения — небольшие наросты на клеточной мембране, состоящие из системы микротрубочек. Они позволяют клеткам свободно перемещаться, что способствует росту живых организмов. Реснички, жгутики, псевдоподии и миофибриллы являются основными представителями органоидов движения. Строение и функции этих органелл изучаются на уроках биологии в 6 классе.
Краткая информация
Органоиды движения содержатся в растительных и животных клетках, входящих в состав многоклеточных организмов. Структура этих органелл формируется из молекул белков и фосфолипидов. Их средний размер составляет 0,25—100 мкм. В таблице перечислены основные особенности органоидов движения.
| Наименование органоида движения | Строение | Функции органоидов движения | Название одноклеточного организма |
| Жгутики | Цитоплазматические наросты, расположенные на поверхности мембраны | Передвижение клеток | Жгутиковые инфузории |
| Реснички | Тонкие выросты на эластичной структуре клетки | Очистка органов от пыли | Ресничные инфузории |
| Псевдоподии (ложноножки) | Выступы в цитоплазме клетки | Питание и передвижение организма | Саркодовые |
| Миофибриллы | Нити малой толщины | Сокращение мышц | Корненожки |
В человеческом организме присутствует большое количество ресничек и жгутиков.
Они предназначены для очищения легких, защиты эпителия и стабильного функционирования репродуктивной системы. Принцип работы этих органоидов движения заключается в установлении прочных связей с клеточной мембраной.
Жгутики бактерий и архей
Жгутик — органоид движения эукариотов, обеспечивающий передвижение клеточных организмов в жидкой среде. Они содержатся в протистах, зооспорах и половых клетках. Эти органеллы представляют собой небольшие наросты, окруженные эластичной пленкой. Жгутики имеют цитоскелет, где осуществляется процесс гидролиза АТФ. Второстепенные функции жгутиков:
- формирование биологических пленок;
- обеспечение контакта клеточных организмов с субстратами;
- облегчение проникания симбиотических бактерий в клетки;
- включение защитных механизмов иммунной системы;
- предотвращение заражения клетки инфекционными вирусами.

Жгутик эукариотических клеток представляет собой комплексный структурный элемент. Он включает в себя 9 пар микротрубочек, соединенных нексиновыми мостиками. Между ними присутствует переходная зона эксонемы. В центральной части жгутика располагается ось с центриолями. На следующем рисунке описано строение органоида в разрезе.
80% бактерий состоят из жгутиков. Они находятся на противоположных полюсах клеточного организма. Отличительной чертой жгутиков бактерий является их расположение в клетке. Они вмонтированы в оболочку клеточного организма. Вращение жгутика осуществляется при помощи энергии, получаемой при гидролизе АТФ. Органоид перемещается по часовой стрелке. Частотный диапазон вращения органеллы составляет от 200 до 1850 Гц. Бактериальный жгутик состоит из следующих компонентов:
 Представлено в виде муреинового чехла, окруженного мембранной оболочкой. Оно состоит из системы секреции и мотора.
Представлено в виде муреинового чехла, окруженного мембранной оболочкой. Оно состоит из системы секреции и мотора.Основным рабочим элементом жгутика бактерий является филамент. Этот компонент объединяет несколько тысяч субъединиц фосфолипидов и белка. При вращении органоида филамент приобретает форму спирали, закрученной в левую сторону.
Базальное тело бактериального жгутика состоит из следующих частей:
- оси клеточного центра;
- L-кольца;
- P-кольца;
- MS-кольца;
- ротора;
- C-кольца.
Для определения характера передвижения жгутика требуется знать количество ресурсов, поставляемых основными компонентами базального тела.
Жгутики архей состоят из археллума, включающего в себя 7—13 различных генов. Структура этого органоида формируется едиными оперонами. Функцией жгутиков архей является перемещение клеточных организмов во влажной среде, но они не принимают участия в формировании биологических пленок.
Механизм работы ресничек
Реснички представляют собой тонкие органеллы в форме волоса. Находясь в неподвижном состоянии, эти органоиды выполняют роль рецепторов. Поверхность ресничек покрыта плотной цитоплазматической мембраной. В состав органеллы входит большое количество микротрубочек. В центральной части ресничек расположено базальтовое тело с центриолями.
В микротрубочках присутствуют белковые структуры, обеспечивающие стабильное скольжение ресничек. Во время передвижения органоиды совершают удары. Они предназначены для деполяризации клеточной мембраны. Характер и направление ударов зависят от процентного содержания ионов кальция в структуре ресничек.
Псевдоподии и миофибриллы
Псевдоподии представляют собой цитоплазматические выросты без плотной клеточной оболочки. В школьных учебниках эти органеллы также могут называться ложноножками. Псевдоподии сдержат простейшие организмы:
- амебы;
- арцеллы;
- фораминиферы.
Ложноножки предназначены для всасывания питательных веществ. Также к их функциям относится перемещение клеточных организмов. Скорость движения клеток составляет не более 0,2 мм/мин. Во время перемещения псевдоподии закрепляются в субстрате и захватывают частицы пищи. В результате этого процесса формируется пищеварительная вакуоль.
Миофибриллы представляют собой органоиды, состоящие из тонких белковых филаментов. Они располагаются в мышечном волокне и не имеют клеточной оболочки. Миофибриллы окружены саркоплазматическим ретикулумом, состоящим из саркомеров. Связь этих органелл с мышцами обеспечивается при помощи белковых нитей.
Миофибриллы обеспечивают стабильное сокращение мышечных волокон. При воздействии нервных импульсов эти органоиды начинают уменьшаться. В результате сокращения числа органоидов образуется энергия, определяющая силу мышцы. Во время силовых тренировок количество миофибрилл увеличивается. Этот процесс называется гипертрофией. Большие органоиды начинают делиться на несколько маленьких, иначе мышцы не смогут получать достаточное количество энергии.
После травмирования мышечных волокон число миофибрилл уменьшается. Этот процесс ускоряется при наличии гипса, фиксирующего мышцы в определенном положении. Это обусловлено разрушением саркомеров. В результате сокращения количества миофибрилл мышцы получают меньше энергии.
ПредыдущаяБиологияГуморальный иммунитет — характеристика, механизм и факторы
СледующаяБиологияОрганы чувств у паукообразных — виды, характеристика и строение
Немембранные органоиды клетки. Строение рибосомы
Ст. 88
Рассмотрите рис. 60 — 62. В чем сходство и отличия строения немембранных органоидов клетки? Предположите, каковы их функции.
Рибосомы: Округлый органоид, состоящий из двух субъединиц
Функция: Синтез белка
Клеточный центр: Два цилиндрика – центриоли, образованные микротрубочками
Функция: Образуют цитоскелет клетки, нити веретена деления, жгутики и реснички
Органоиды движения – реснички, жгутики: Микротрубочки (белковые)
Функция: Передвижение клеток в пространстве; Перемещение вдоль клеток окружающей их жидкости и частиц
Микротрубочки: состоят из множества субъединиц сократительного белка тубулина.
Функция: формируют внутреннюю структуру – цитоскелет клетки, который выполняет опорную функцию. Они обеспечивают движение цитоплазмы, перемещение в клетке некоторых органоидов, например митохондрий, лизосом и пузырьков аппарата Гольджи.
Ст. 91
Вопросы и задания
1. Каково внешнее строение рибосомы? Из чего она состоит?
Это очень мелкие тельца грибовидной формы, на которых идёт синтез белка. Рибосома состоит из двух субъединиц: большой и малой. Каждая субъединица состоит из рибосомальной РНК (рРНК) и белка.
2. Где в клетке встречаются рибосомы, с какими клеточными структурами связана их деятельность? Какие функции выполняют рибосомы?
Рибосомы обычно находятся в диссоциированном состоянии, т. е. большая и малая субъединицы не связаны друг с другом. Во время синтеза белка две субъединицы рибосомы соединяются с иРНК и образуют единый комплекс, в котором происходит биосинтез белка. Одни рибосомы находятся на гранулярной ЭПС, и синтезируемый ими белок поступает внутрь ЭПС и транспортируется дальше по отсекам клетки. Другие, так называемые свободные, рибосомы находятся в цитоплазме и участвуют в синтезе белков цитоплазмы. Несколько рибосом способны во время синтеза белка соединяться с иРНК и образовывать длинные цепи – полисомы. На полисоме одновременно идёт синтез нескольких молекул одного и того же белка.
3. В каких клетках содержится больше рибосом: в клетках жировой ткани, волосяного фолликула или железистого эпителия? Ответ поясните.
Больше рибосом содержится в клетках волосяных луковиц, поскольку им необходимо синтезировать большое количество кератина и других белков, входящих в состав волос. Клетки жировой ткани являются своеобразными хранилищами липидов и не специализируются на синтезе белков.
4. Объясните строение микротрубочек. Какие функции они выполняют?
В морфологическом отношении микротрубочки представляют собой длинные полые цилиндры с внешним диаметром 25 нм. Стенка микротрубочек состоит из полимеризованных молекул белка тубулина. При полимеризации молекулы тубулина образуют 13 продольных протофиламентов, которые скручиваются в полую трубку. Размен мономера тубулина составляет около 5 нм, равного толщине стенки микротрубочки, в поперечном сечении которой видны 13 глобулярных молекул.
Функции:
1. Поддержание формы и полярности клетки, распределение ее компонентов.
2. Обеспечение внутриклеточного транспорта.
3. Обеспечение движений ресничек, хромосом в митозе.
4. Образование основы других органелл (центриолей, ресничек).
5. Какие клеточные структуры являются производными микротрубочек? В чём их сходства и отличия? Какие функции они выполняют?
Реснички и жгутики являются производными микротрубочек в клетках эпителия воздуховодных путей, женского полового тракта, семявыносяших путей, сперматозоидах. Являются органоидами движения в основном одноклеточных организмов
Центриоль – небольшой цилиндрический органоид, представляющий собой полый цилиндр, стенки которого образованы девятью триплетами микротрубочек. Участвует в процессе деления клетки.
Базальное тельце представлено одним цилиндром, состоящим из девяти триплетов микротрубочек. Способны восстанавливать жгутики и реснички.
Общее: микротрубочки и клеточные структуры имеют общее строение.
Различия: выполняют разные функции.
6. Опишите строение и принцип действия органоидов движения.
Жгутики и реснички. Это органоиды движения, характерные как для одноклеточных организмов (жгутиковые и инфузории), так и для некоторых клеток многоклеточных организмов (клетки некоторых эпителиев, сперматозоиды). Жгутики и реснички представляют собой цилиндр, стенку которого образуют 9 пар микротрубочек; в центре расположены две осевые микротрубочки. Эта часть полностью или на большем протяжении покрыта участком наружной цитоплазматической мембраны. В основании органоида, в наружном слое цитоплазмы, расположено базальное (основное) тельце, в котором к каждой паре микротрубочек, образующих наружную часть жгутики или реснички, прибавляется еще одна короткая микротрубочка. Таким образом, базальное тельце оказывается образованным из девяти триад трубочек и имеет сходство с компонентом клеточного центра – центриолью. Движение жгутиков и ресничек обусловлено скольжением микротрубочек каждой пары друг относительно друга, при котором затрачивается большое количество энергии в виде АТФ.
7. Какие органоиды из перечисленных ниже обязательно присутствуют в эукариотной клетке: микротрубочки, клеточный центр, центриоли, базальное тельце, цитоскелет? Объясните почему.
Клеточный центр необходим клетке, так как он принимает участие в её делении. В состав клеточного центра входит центриоль. Центриоль – парное образование. Она содержит 2 удлиненные гранулы, состоящие из микротрубочек и расположенные перпендикулярно друг другу. Цитоскелет в свою очередь придает клетке прочность, придает клетке форму. Цитоскелет включает в себя и базальные тельца. Каждый из этих органоидов необходим клетке в равной мере. Все эти органоиды тесно связано между собой.
Flagellum — Определение, функция, типы и тест
Flagellum Definition
Жгутик — это микроскопическая волосовидная органелла, используемая клетками и микроорганизмами для движения. Слово flagellum на латыни означает хлыст, точно так же, как хлестание жгутиков (множественное число) часто используется для передвижения. Специализированные жгутики у некоторых организмов также используются в качестве сенсорных органелл, которые могут обнаруживать изменения температуры и pH.
Функция жгутика
Жгутики — это нитчатые белковые структуры, обнаруженные у бактерий, архей и эукариот, хотя чаще всего они встречаются у бактерий.Обычно они используются для продвижения клетки через жидкость (например, через бактерии и сперму). Однако у жгутиков есть много других специализированных функций. Некоторые эукариотические клетки используют жгутик для увеличения скорости размножения. Другие эукариотические и бактериальные жгутики используются для определения изменений в окружающей среде, таких как изменение температуры или pH. Недавняя работа с зеленой водорослью Chlamydomonas reinhardtii показала, что жгутик также может использоваться в качестве секреторной органеллы, но для полного понимания этого открытия требуется больше времени.
Примеры жгутика
Жгутик может состоять из различных структур в зависимости от организма, особенно при сравнении жгутика эукариот и бактерий. Поскольку эукариоты обычно являются сложными организмами, прикрепленный жгутик также является более сложным. Жгутик состоит из микротрубочек, состоящих из белка тубулина. Девять пар микротрубочек окружают еще две пары микротрубочек в центре, образуя ядро жгутика; это известно как схема «девять плюс два».Вся структура девять плюс два закреплена в базальном теле внутри организма. Эти связанные микротрубочки используют АТФ, чтобы изгибаться вперед и назад вместе, подобно хлысту.
Хотя некоторые многоклеточные эукариоты имеют настоящий жгутик, почти половина человеческой популяции производит клетки с ними в форме сперматозоидов. Это единственная клетка в человеческом теле со жгутиком, и не зря. Чтобы двигаться по влагалищному тракту и встретиться с яйцеклеткой, сперматозоиды должны уметь плавать или перемещаться на очень большие расстояния (по сравнению с размером клетки и размером тела).Без жгутика было бы очень мало шансов на оплодотворение или стабильность популяции.
С другой стороны, бактериальные жгутики устроены и функционируют совершенно иначе, чем эукариотические аналоги. Эти жгутики состоят из белка, называемого флагеллином. АТФ не нужен, потому что бактериальный жгутик может использовать энергию протонодвижущей силы. Это означает, что энергия получена из ионных градиентов — обычно водорода или натрия — которые проходят через клеточные мембраны. Эти жгутики имеют форму спирали и быстро вращаются, как ветряная мельница, чтобы перемещать организм, а не хлестать вперед и назад.Бактерия Escherichia coli использует это движение, подобное ветряной мельнице, для продвижения вверх по уретре, вызывая инфекции мочевыводящих путей. Salmonella enterica , вредоносный патоген, использует несколько жгутиков, похожих на ветряные мельницы, для заражения человека-хозяина.
Сравнение движения жгутика у бактериальных (прокариотических) и эукариотических организмов:
Типы жгутика
Структура жгутика состоит из трех различных частей: колец, встроенных в базальное тело, крючка у поверхности организма, удерживающего его на месте, и белковых нитей жгутика.Эти три общие черты есть у каждого жгутика, независимо от организма. Тем не менее, существует четыре различных типа бактериального жгутика в зависимости от местоположения:
Жгутик
A. Однообразный: Одиночный жгутик на одном или другом конце организма.
B. Lophotrichous: Несколько жгутиков на одном или другом конце организма.
C. Амфитрихия: Один жгутик на обоих концах организма.
Д.Peritrichous: Несколько жгутиков прикреплены по всему организму.
Монотриховый, амфитриховый и лофотрихный жгутик считаются полярным жгутиком, потому что жгутик расположен строго на концах организма. Эти жгутики могут вращаться как по часовой стрелке, так и против часовой стрелки. Движение по часовой стрелке продвигает организм (или клетку) вперед, а движение против часовой стрелки — назад.
Перитрихозные жгутики не считаются полярными, так как они расположены по всему организму.Когда эти жгутики вращаются против часовой стрелки, они образуют пучок, который продвигает организм в одном направлении. Если несколько жгутиков оторвутся и начнут вращаться по часовой стрелке, организм начнет кувырок. В это время организм не может двигаться ни в каком реальном направлении.
Если какой-либо жгутик перестает вращаться, независимо от полярности, организм меняет направление. Это вызвано броуновским движением (постоянное движение жидких частиц) и потоками жидкости, которые догоняют организм и вращают его.Некоторые организмы, которые не могут изменить направление самостоятельно, полагаются на броуновское движение и жидкие потоки, чтобы сделать это за них.
- ATP — Аденозинтрифосфат, небольшая молекула, используемая в клетках как кофермент, передающий энергию.
- Микротрубочки — микроскопическая трубчатая структура, присутствующая в цитоплазме клеток, которая помогает формировать цитоскелет.
- Базальное тело — Органелла, образующая самое основание жгутика; По строению он похож на центриоль.
- Броуновское движение — Случайное движение частиц в жидкости (жидкости или газе), вызванное столкновением с другими молекулами внутри той же жидкости.
Викторина
1. Какова основная функция жгутика?
A. Производство энергии
B. Связь
C. Передвижение
D. Производство тепла
Ответ на вопрос № 1
C правильный.Хотя жгутик может многое делать, его основная функция — продвигать клетку или организм.
2. Что дает энергию жгутикам бактерий?
A. ATP
B. Градиенты ионов
C. Базальные тела
D. Flagellin
Ответ на вопрос № 2
B правильный. В отличие от жгутика эукариот, жгутик бактерий использует энергию ионных градиентов (обычно водород или натрий) для движения.
3. Что из перечисленного не считается жгутиком полярных бактерий?
A. Монотриховый
B. Амфитричный
C. Лофотриховый
D. Перитриховый
Ответ на вопрос № 3
D правильный. Перитрихозные жгутики не считаются полярными, потому что они расположены на всех частях организма, а не только на концах.
Строение и функции ресничек и жгутиков
Обзор
Реснички и жгутики представляют собой тонкие шиповидные / волосовидные структуры, которые отходят от тела множества клеток.Хотя они различаются по длине и количеству в разных типах клеток (а также по паттернам движения), реснички и жгутики в целом идентичны по структуре и составу.
В зависимости от типа клеток реснички и жгутики выполняют следующие функции:
· Двигательные клетки — Используя реснички или жгутики, клетки могут свободно перемещаться в окружающей среде, особенно в водной или влажной среде.
· Сенсорные функции — Некоторые реснички и жгутики позволяют клеткам ощущать изменения в своем окружении, что, в свою очередь, позволяет клеткам реагировать соответствующим образом.
· Транспортировка материала — Некоторые ячейки могут не только улавливать, но и направлять транспортировку данного материала. Это может служить для поглощения такого материала клеткой или предотвращения проникновения нежелательного материала / частиц / микроорганизмов в клетку или ткань.
* Жгутики прокариот имеют другую структуру по сравнению с жгутиками эукариотических клеток.
Реснички
За исключением большинства высших растений и грибов, реснички можно найти на поверхности многих эукариотических клеток.На этих клетках реснички отходят от базального тела.
В зависимости от типа клеток реснички выполняют несколько функций и поэтому делятся на две основные категории.
* Прокариоты (бактерии) не имеют ресничек.
Структура ресничек
Реснички — это микроскопические, похожие на волосы структуры, которые выступают с поверхности многих эукариотических клеток. Подобно другим органеллам эукариотических клеток, реснички представляют собой мембраносвязанные структуры, причем их мембрана непрерывна с плазматической мембраной.
Было показано, что в отличие от плазматической мембраны клеток, ресничная мембрана содержит отдельные липиды и белки.
Подвижные реснички
* Подвижные реснички были идентифицированы в 1640-х годах ван Левенгук, сделав их самыми ранними известными клеточными органеллами.
Подвижные реснички (9 + 2) можно найти как у высших животных, так и у одноклеточных эукариот. У микроскопических организмов (известных как инфузории) подвижные реснички используются для передвижения или перемещения жидкости по их поверхности, что способствует процессу питания.
У высших животных, таких как люди, подвижные реснички можно найти в ряде тканей (например, в респираторном эпителии и фаллопиевых трубах), где они либо участвуют в очистке, либо перемещении веществ.
В дыхательной системе реснички захватывают и удаляют грязь (а также слизь) из легких и других частей этой системы. С другой стороны, в маточной трубе реснички служат для перемещения яйцеклетки в матку.
На поверхности клетки подвижные реснички присутствуют в большом количестве, где они бьются скоординированным волнообразным образом, чтобы эффективно выполнять свои функции.
Некоторые примеры инфузорий включают:
Что касается структуры, подвижные реснички характеризуются радиальным паттерном, состоящим из девяти (9) дублетов внешних микротрубочек, которые окружают две синглетные микротрубочки.
Итак, здесь паттерн 9 + 2 относится к девяти дублетным микротрубочкам, окружающим две микротрубочки, которые расположены в центре. Кольцо каркаса микротрубочек в этом случае известно как аксонема.
В дополнение к микротрубочкам, которые являются основными компонентами структуры, подвижные реснички также состоят из динеиновых плеч и радиальных спиц, которые вносят вклад в общую подвижность структуры.
* Аксонема (пучок микротрубочек размером около 0,25 мкм в диаметре) окружена плазматической мембраной, и всю структуру (реснички) можно идентифицировать под микроскопом.
В своем основании (где она прикрепляется к клетке) аксонема прикреплена к цилиндрическим структурам, известным как базальные тела. Базальные тельца составляют около 0,4 мкм в длину и 0,2 мкм в ширину и состоят из A-канальца (девять (9) триплетных микротрубочек, состоящих из микротрубочек протофиламента), неполного B-канальца, а также неполного C-канальца.
Помимо закрепления ресничек в цитоплазме, базальные тельца также играют важную роль в сборке этих структур.
* Согласно исследованиям, базальные тельца либо являются продуктом центриолей, либо образуются в большом количестве до образования ресничек.
* Даже когда окружающая плазматическая мембрана была удалена, добавление АТФ позволяет аксонеме продолжать функционировать, что свидетельствует о том, что рабочий механизм структуры находится в аксонеме.
Механизм избиения ресничек
Как и в случае мышечного сокращения, биение / рабочий механизм ресничек (в частности, аксонемы), как было показано, является результатом скольжения белковых филаментов.
Хотя механизм в целом еще предстоит полностью понять, исследования показали, что динеины, которые действуют как молекулярные моторы, играют важную роль в обеспечении энергии биения ресничек.
Одной из моделей, которые использовались для описания изгиба и, следовательно, функционирования подвижных ресничек, является гипотеза модели переключателя.
Согласно модели переключения, каждая сторона изогнутых ресничек состоит из динеинов в заданном состоянии цикла генерации силы, который вносит вклад в асимметрию и изменяется с изменениями кривизны.
Здесь, согласно исследованиям, динеины на одной микротрубочке (в состоянии цикла генерации силы) скользят друг мимо друга, а на другой стороне — нет. Это приводит к изгибу аксонемы, в то время как переключение этой системы заставляет структуру изгибаться в другую сторону.
В конечном итоге повторение этого механизма заставляет подвижные реснички биться и, таким образом, выполнять свою функцию.
* Присоединение и высвобождение динеиновых плеч к соседнему дублету вызвано связыванием или гидролизом АТФ.
Первичные реснички (неподвижные реснички)
По сравнению с подвижными ресничками первичные реснички (9 + 0) выступают как одиночные структуры из тел клеток. Они обнаружены практически во всех клетках всех млекопитающих. Они в первую очередь участвуют в сенсорных функциях и, таким образом, позволяют тканям / органам организма реагировать должным образом.
Подобно подвижным ресничкам первичные реснички состоят из девяти дублетных микротрубочек, составляющих аксонему. Эти микротрубочки происходят из базального тела, что также обеспечивает стабильность.
В отличие от подвижных ресничек, однако, первичные реснички не обладают динеиновыми плечами и центральными синглетными микротрубочками (центральные парные микротрубочки). Это связано с тем, что первичные реснички неподвижны и, следовательно, не нуждаются в элементах, необходимых для подвижности.
* Первоначально считалось, что первичные реснички являются рудиментарными органеллами, которые не служат никакой цели.
Формирование первичных ресничек
Формирование первичных ресничек начинается, когда клетка входит в фазу G0 клеточного цикла. Здесь материнская центриоль центросомы сначала прикрепляется к пузырьку, после чего происходит рост аксонемы с поверхности центриоли.
Аксонемные микротрубочки также начинают полимеризоваться на кончике роста, куда груз доставляется посредством внутрижладжкового транспорта.
Эта двунаправленная транспортная система позволяет транспортировать белки к микротрубочкам во время их развития.В то время как везикула в конечном итоге экзоцитозируется, первичные реснички обнажаются на поверхности клетки и продолжают развиваться, пока не достигнут зрелости.
Внутрилагеллярный транспорт (IFT) все еще необходим для поддержания первичных ресничек.
* Было показано, что первичные реснички выровнены в одном направлении, что, в свою очередь, влияет на ориентацию клеток.
Функции первичных ресничек
Первичные реснички играют важную роль в передаче клеточных сигналов во время развития и гомеостаза.Учитывая, что первичные реснички (5-10 мкм в длину) подвергаются воздействию внеклеточной среды, они восприимчивы к различным стимулам, которые вносят вклад в их роль в передаче сигналов.
В дополнение к обнаружению различных химических факторов, морфогенов и факторов роста во внеклеточном матриксе, первичные реснички также обнаруживают изменения давления и движения жидкости по поверхности клетки.
Например, из-за потока мочи в почечных канальцах первичные реснички изгибаются, что, в свою очередь, приводит к притоку ионов кальция через соответствующие кальциевые каналы.
Помимо путей Hedgehog, пути передачи сигналов Wnt являются одними из наиболее документированных путей в отношении передачи сигналов от ресничек. По существу, путь передачи сигналов Wnt важен, потому что он участвует в ряде процессов, включая клеточную полярность, миграцию клеток, а также формирование нервного паттерна среди других.
Это происходит по двум основным путям, включая канонический путь Wnt и неканонический путь Wnt.
В то время как активация канонического пути Wnt способствует экспрессии гена, активация неканонического пути Wnt приводит к деградации b-катенина.Здесь связывание различных лигандов Wnt с рецепторами, расположенными на первичных ресничках, позволяет канонической передаче сигналов Wnt переключаться на неканоническую передачу сигналов и наоборот.
Роль первичных ресничек также очевидна в ряде др. Сигнальных путей, обеспечивающих соответствующие ответы. С другой стороны, дефекты функции ресничек связаны с различными заболеваниями, связанными с развитием, и дегенеративными заболеваниями.
Дефектные функции ресничек были связаны со следующими заболеваниями и синдромами заболеваний:
- Первичная дискинезия ресничек
- Синдром Альстрома
- Синдром Меккеля-Грубера
- Нефронофтизия
- Респираторная инфекция Мужское бесплодие
Жгутик
Жгутик (множественное число: Flagella) можно описать как нитевидную органеллу, которая в основном используется для передвижения.Как и реснички (обнаруженные в эукариотических клетках), жгутики также выступают из тела клетки, что позволяет им эффективно выполнять свои функции. Однако они длиннее, от 5 до 20 мкм.
Клетки, обладающие такой структурой, называются жгутиконосцами и включают как эукариотические, так и прокариотические клетки. Например, помимо большинства бактерий, которые используют жгутики для передвижения, структура также может быть найдена на таких одноклеточных организмах, как эвглена, и простейшие виды, такие как Trypanosoma evansi.Жгутики, а также лагеллы можно найти на гаметах различных организмов, включая слизевики, грибы и животных.
Структура жгутика
Хотя жгутики могут быть найдены как в эукариотических, так и в прокариотических клетках (и служат одной и той же цели), существуют различные различия в отношении их структур / состава, а также механизма, с помощью которого они функционируют между двумя типами клеток.
Жгутики прокариотических клеток состоят из глобулярного белка, известного как флагеллин.Здесь белок спирально оборачивается, образуя полый цилиндр по всей длине структуры. Этот белок отсутствует в жгутике эукариот, где он заменен белковыми филаментами, известными как микротрубочки.
Некоторые из других различий между ними включают:
- Жгутики прокариот, как правило, меньше и менее сложны по сравнению с жгутиками эукариот
- Жгутики эукариот питаются АТФ, а жгутики прокариот управляются протонами прокариотические жгутики характеризуются вращательным движением, в то время как у эукариотических клеток есть изгибы
- Прокариотические жгутики не имеют плазматической мембраны
Помимо длины, структура и состав жгутиков эукариот аналогичны ресничкам многих эукариот (описано выше).
Таким образом, в этом разделе основное внимание будет уделено структуре жгутиков прокариотических клеток.
Жгутик бактерий состоит из трех основных частей, которые включают:
- Базальная структура (роторный двигатель)
- Крюк (действует как универсальный шарнир)
- Нить (винтовой винт)
Базальное тело — У бактерий / прокариот базальное тело представляет собой стержень, состоящий из нескольких колец, расположенных внутри клеточной мембраны.У грамотрицательных бактерий кольца включают L-кольцо, которое расположено на внешней мембране липидного бислоя, и P-кольцо, которое расположено в слое пептидогликана.
Базальное тело обычно делится на несколько частей, которые включают:
- Белковые кольца (C-кольцо, MS-кольцо, P-кольцо и L-кольцо)
- Стержень
- Гильза
* Белок кольца служат протонными насосами, которые участвуют в движении ионов водорода через мембрану.Именно это движение ионов через мембрану в конечном итоге вращает кольца и, следовательно, жгутик.
* Базальное тело, как и крючок, также служит для закрепления нити конструкции на поверхности клетки.
Крючок
Состоящий из 120 субъединиц одного белка, крючок (короткий и изогнутый) действует как универсальный шарнир, соединяющий нить с базальным телом.
В отличие от базального тельца крючок не встроен в плазматическую мембрану.Однако он играет решающую роль в подвижности и такси бактерий, передавая крутящий момент двигателя на филаментную (пропеллерную) часть структуры.
Он состоит из 4 основных доменов, расположенных внутри и снаружи конструкции, природа которых допускает прямое соединение между крючком и стержнем.
* Соединение между крючком и филаментом состоит из двух белков (FlgK и FlgL), которые, как было показано, вносят вклад в формирование филаментной части структуры.
Нить
Нить — удлиненная часть жгутика. Он трубчатый и состоит из 11 протофиламентов, похожих на те, что находятся в стержневой и крюковой частях конструкции.
Флагеллин, который является основным компонентом филамента, также состоит из четырех доменов, которые образуют внутреннюю и внешнюю части структуры. Направление вращения нити зависит от вращения двигателя (по или против часовой стрелки).
В то время как жгутики, обнаруженные в таких организмах, как бактерии, археи и различные эукариотические клетки, используются для плавания в жидкости, а также для роения; Было также показано, что они выполняют ряд других важных функций.Например, в эукариотических клетках структура, как было показано, играет роль в увеличении продукции.
Было показано, что как в бактериях, так и в эукариотических клетках некоторые жгутики обладают сенсорными функциями, которые позволяют клеткам обнаруживать изменения в окружающей их среде и эффективно реагировать. Исследования показали, что у некоторых зеленых водорослей жгутики могут действовать как секреторные органеллы.
* Организмы можно классифицировать по количеству жгутиков на их поверхности.
К ним относятся:
- Монотрихий — один жгутик, исходящий с одного конца
- Лофотриховый — несколько жгутиков на одном полюсе
- Амфитрихий — один жгутик на обоих полюсах
- Перитрихий — несколько жгутиков на поверхности тела
Возвращение к инфузориям
Возвращение в прокариотические клетки
Возвращение в эукариотические клетки
Возвращение из ресничек и жгутика в MicroscopeMaster home
Список литературы
Brent W.Бисгроув и Х. Джозеф Йост. (2006). Роль ресничек в нарушениях развития и заболеваниях. Компания биологов.
Хироюки Терашима, Сэйдзи Кодзима и Мичио Хомма. (2008). Подвижность жгутиков у бактерий:
Структура и функция жгутикового мотора. Международный обзор клеточной и молекулярной биологии, том 270.
Такаши Исикава. (2017). Структура аксонемы из подвижных ресничек.
Стивен М. Кинг. (2018). Отключение динеинов приводит к изгибу ресничек.
Уитман Г.Б. (1990) Введение в реснички и жгутики. В: Бладгуд Р.А. (ред.) Ресничные и жгутиковые мембраны. Спрингер, Бостон, Массачусетс.
Юко Комия и Раймонд Хабас. (2008). Пути передачи сигнала Wnt. ncbi.
Ссылки
https://www.researchgate.net/publication/235374058_Cilia_Wnt_signaling_and_the_cytoskeleton
/10/article/academic.io/articles/articles/academic.info/ 250556
Жгутики: типы, функции и структура
Подвижность клеток — ключевой компонент выживания многих одноклеточных организмов, а также она может быть важна для более продвинутых животных.Клетки используют жгутики для передвижения , чтобы искать пищу и избегать опасности. Плетевидные жгутики могут вращаться, чтобы стимулировать движение посредством эффекта штопора, или они могут действовать как весла, перемещая клетки через жидкости.
Жгутики встречаются у бактерий и некоторых эукариот, но эти два типа жгутиков имеют разную структуру.
Жгутик бактерий помогает полезным бактериям перемещаться по организму и помогает болезнетворным бактериям распространяться во время инфекций.Они могут перемещаться туда, где они могут размножаться, и могут избежать некоторых атак со стороны иммунной системы организма. У продвинутых животных клетки, такие как сперматозоиды, перемещаются с помощью жгутика.
В каждом случае движение жгутиков позволяет клетке двигаться в общем направлении.
Структура жгутиков прокариотических клеток проста
Жгутики прокариот, например бактерий, состоят из трех частей:
- Нить жгутика представляет собой полую трубку, состоящую из жгутикового белка, называемого флагеллином .
- В основе нити накала находится гибкий крюк , который соединяет нить с основанием и действует как универсальный шарнир.
- Базальное тело состоит из стержня и ряда колец, которые прикрепляют жгутик к клеточной стенке и плазматической мембране.
Филамент жгутика создается путем транспортировки белка флагеллина от клеточных рибосом через полое ядро к кончику, где флагеллин прикрепляется и заставляет филамент расти.Базальное тело образует мотор жгутика, а крючок придает вращению эффект штопора.
Жгутики эукариот имеют сложную структуру
Движение жгутиков эукариот и прокариотических клеток схоже, но структура филамента и механизм вращения различны. Базальное тело эукариотических жгутиков прикреплено к телу клетки, но у жгутика нет стержня и дисков. Вместо этого филамент твердая и состоит из пар микротрубочек .
Канальцы расположены в виде девяти двойных трубок вокруг центральной пары трубок в форме 9 + 2. Канальцы состоят из линейных белковых цепочек вокруг полого центра. Двойные трубы имеют общую стенку, а центральные трубы независимы.
Белковые спицы, оси и звенья соединяются с микротрубочками по длине филамента. Вместо движения, создаваемого в основании вращающимися кольцами, движение жгутика происходит за счет взаимодействия микротрубочек.
Жгутики работают за счет вращательного движения нити
Хотя бактериальные жгутики и жгутики эукариотических клеток имеют разную структуру, они оба работают за счет вращательного движения нити, чтобы продвигать клетку или перемещать жидкости мимо клетки. Более короткие нити будут двигаться вперед и назад, в то время как более длинные нити будут двигаться по круговой спирали.
У бактериальных жгутиков крючок в нижней части филамента вращается там, где он прикреплен к клеточной стенке и плазматической мембране.Вращение крючка приводит к движению жгутиков, как у пропеллера. У эукариотических жгутиков вращательное движение происходит из-за последовательного изгиба нити.
Результирующее движение может быть не только вращательным, но и хлыстовым.
Прокариотические жгутики бактерий приводятся в действие с помощью жгутикового двигателя
Под крючком бактериальных жгутиков основание жгутика прикреплено к клеточной стенке и плазматической мембране клетки серией колец, окруженных белковыми цепями.Протонный насос создает протонный градиент в самом нижнем из колец, а электрохимический градиент обеспечивает вращение за счет протонной движущей силы .
Когда протоны диффундируют через нижнюю границу кольца из-за движущей силы протонов, кольцо вращается, и прикрепленный крючок нити вращается. Вращение в одном направлении приводит к контролируемому поступательному движению бактерии. Вращение в обратном направлении заставляет бактерии двигаться беспорядочно кувыркающимся образом.
Результирующая подвижность бактерий в сочетании с изменением направления вращения создает своего рода случайное блуждание, которое позволяет клетке покрывать большую территорию в общем направлении.
Эукариотические жгутики используют АТФ для изгиба
Основание жгутика эукариотических клеток прочно прикреплено к клеточной мембране, и жгутики скорее изгибаются, чем вращаются. Белковые цепи, называемые динеином , прикреплены к некоторым двойным микротрубочкам, расположенным вокруг нитей жгутиков радиальными спицами.
Молекулы динеина заставляют жгутики изгибаться, перемещая микротрубочки вверх и вниз друг относительно друга. Они отделяют одну из фосфатных групп от молекул АТФ и используют высвободившуюся химическую энергию, чтобы захватить одну из микротрубочек и переместить ее к канальцу, к которому они прикреплены.
Путем координации такого действия изгиба результирующее движение нити может быть вращательным или возвратно-поступательным.
Прокариотические жгутики важны для размножения бактерий
Хотя бактерии могут выжить в течение длительного времени на открытом воздухе и на твердых поверхностях, они растут и размножаются в жидкостях. Типичная жидкая среда — это богатые питательными веществами растворы и внутренности развитых организмов.
Многие из этих бактерий, например, бактерии в кишечнике животных , полезны, но они должны уметь находить необходимые им питательные вещества и избегать опасных ситуаций.
Жгутики позволяют им двигаться к еде, прочь от опасных химикатов и распространяться при размножении.
Не все кишечные бактерии полезны. H. pylori , например, жгутиковые бактерии, вызывающие язву желудка. Он полагается на жгутики, чтобы перемещаться через слизь пищеварительной системы и избегать участков с повышенной кислотностью. Когда он находит подходящее место, он размножается и использует жгутики для распространения.
Исследования показали, что модель H.pylori жгутики являются ключевым фактором инфекционности бактерий.
Бактерии можно классифицировать по номеру и местоположению их жгутиков. Монотрихия бактерий имеют единственный жгутик на одном конце клетки. Lophotrichous бактерий имеют на одном конце пучок из нескольких жгутиков.
Перитрихийные бактерии имеют как боковые жгутики, так и жгутики на концах клетки, тогда как амфитрихов бактерий могут иметь один или несколько жгутиков на обоих концах.
Расположение жгутиков влияет на скорость и направление движения бактерии.
Эукариотические клетки используют жгутики для перемещения внутри и снаружи организмов
Эукариотические клетки с ядром и органеллами встречаются у высших растений и животных, но также как одноклеточные организмы. Эукариотические жгутики используются примитивными клетками для передвижения, но их можно найти и у продвинутых животных.
В случае одноклеточных организмов жгутики используются для поиска пищи, распространения и спасения от хищников или неблагоприятных условий.У продвинутых животных специфические клетки используют жгутик эукариот для особых целей.
Например, зеленые водоросли Chlamydomonas reinhardtii используют два жгутика водорослей для передвижения по воде озер и рек или по почве. Он полагается на это движение для распространения после размножения и широко распространен по всему миру.
У высших животных сперматозоид является примером мобильной клетки, использующей для движения жгутик эукариот. Вот как сперма движется по женскому репродуктивному тракту, чтобы оплодотворить яйцеклетку и начать половое размножение.
Жгутики: определение, структура и функции — видео и стенограмма урока
Структуры и типы жгутиков
Жгутик эукариот не является простой волосковой структурой, а имеет сложное поперечное сечение. По структуре он похож на реснички, хотя обычно реснички движутся вперед и назад, в отличие от движения штопора жгутика.
Жгутик эукариот представляет собой длинную палочковидную структуру, которая окружена расширением клеточной мембраны в виде оболочки.Основная часть структуры представляет собой филамент, называемый аксонемой . Необходимые материалы транспортируются по жгутику. Вся конструкция закреплена в базальном теле , которое по структуре похоже на центриоль.
Аксонема имеет девять пар микротрубочек, поддерживающих ее изнутри. Эти дублеты микротрубочек окружают две одиночные микротрубочки. Такое расположение называется структурой 9 + 2. Эукариотические реснички также имеют такую структуру; реснички просто короче.
Дублеты из девяти микротрубочек имеют динеиновых плеч , которые питаются от АТФ. Плечи заставляют микротрубочки в каждой паре скользить друг относительно друга. Это заставляет жгутик изгибаться, позволяя клетке двигаться. Радиальные спицы обращены к центральным микротрубочкам. Их роль неизвестна, но они могут играть роль в стабилизации жгутика.
Жгутики доменов бактерий и архей различны. Они по-прежнему перемещают клетку, но делают это путем вращения, а не изгиба изнутри, как жгутики эукариот.
Жгутики архей внешне похожи на жгутики бактерий. Однако есть ключевые отличия. Начнем с того, что два типа жгутиков передвигаются по-разному. Жгутики бактерий питаются потоком ионов, обычно водорода, но иногда натрия. Они состоят из отдельных волокон, которые вращаются индивидуально. Напротив, жгутиков achaeal , по-видимому, питаются АТФ, тем же химическим веществом, которое питает жгутики эукариот. Волокна архей также связаны и вращаются как одно целое.
Их рост и внутреннее строение также различаются. Жгутик бактерий имеет относительно полую внутреннюю часть, которая способствует росту на кончике жгутика. Новые подузлы продвигаются вверх через внутреннюю полость и прикрепляются к концу. Жгутики архей не имеют такой большой полой внутренней трубки и растут за счет добавления субъединиц в основании. Существуют также различные другие химические различия.
Функции, отличные от передвижения
Помимо помощи клетке в движении, жгутики могут служить сенсорными органеллами.Поскольку они окружены расширением клеточной мембраны, они могут обнаруживать разницу температур, а также химические вещества вне клетки. Эта сенсорная роль продолжается в euglena , где глазное пятно, светочувствительная органелла, располагается близко к жгутику, помогая определить, в каком направлении должна двигаться клетка.
Итоги урока
Давайте рассмотрим.
Жгутик представляет собой плетевидную структуру, которая позволяет клетке двигаться. Они встречаются во всех трех областях живого мира: бактерии, археи и эукариоты.Хотя все три типа жгутиков используются для передвижения, они очень различаются по своей структуре. Жгутик эукариот представляет собой длинную палочковидную структуру, которая окружена расширением клеточной мембраны, наподобие оболочки. Плечи динеина, которые сидят на девяти дублетах микротрубочек аксонемы, заставляют жгутик изгибаться, позволяя клетке двигаться. Жгутики доменов бактерий и архей по-прежнему перемещают клетку, но делают это за счет вращения, а не изгиба изнутри, как жгутики эукариот.
Жгутики не только помогают клетке двигаться, но и служат сенсорными органеллами. Поскольку они окружены расширением клеточной мембраны, они могут обнаруживать разницу температур, а также химические вещества вне клетки.
Результаты обучения
Изучив этот видеоурок по жгутикам, вы можете получить следующие результаты:
- Охарактеризуйте жгутик и сравните его с ресничками
- Обсудите три различных типа жгутиков
- Назовите функции жгутиков
Молекулярные выражения Биология клетки: Структура клеток животных
Реснички и жгутики
Реснички и жгутики — подвижные клеточные придатки, обнаруженные у большинства микроорганизмов и животных, но не у высших растений.У многоклеточных организмов реснички функционируют, чтобы перемещать клетку или группу клеток или помогать транспортировать жидкость или материалы мимо них. Дыхательные пути человека выстланы ресничками, которые предотвращают попадание вдыхаемой пыли, смога и потенциально вредных микроорганизмов в легкие. Помимо прочего, реснички также генерируют потоки воды, чтобы переносить пищу и кислород через жабры моллюсков и транспортировать пищу через пищеварительную систему улиток. Жгутики находятся в основном на гаметах, но также создают потоки воды, необходимые для дыхания и циркуляции губок и кишечнополостных.Для одноклеточных эукариот реснички и жгутики необходимы для передвижения отдельных организмов. Простейшие, принадлежащие к типу Ciliophora , покрыты ресничками, тогда как жгутики являются характеристикой группы простейших Mastigophora .
В эукариотических клетках реснички и жгутики содержат моторный белок динеин и микротрубочки, которые состоят из линейных полимеров глобулярных белков, называемых тубулином .Ядро каждой из структур называется аксонемой и содержит две центральные микротрубочки, которые окружены внешним кольцом из девяти микротрубочек дублета . Одна полная микротрубочка и одна частичная микротрубочка, последняя из которых разделяет стенку канальца с другой микротрубочкой, составляют каждую дублетную микротрубочку (см. Рисунок 1). Молекулы динеина расположены по окружности аксонемы через равные промежутки по ее длине, перекрывая промежутки между соседними дублетами микротрубочек.
Плазматическая мембрана окружает весь комплекс аксонемы, который прикреплен к клетке в структуре, называемой базальным тельцем (также известной как кинетосома ). Базальные тельца поддерживают основную структуру внешнего кольца аксонемы, но каждый из девяти наборов кольцевых филаментов состоит из трех микротрубочек, а не из дублета микротрубочек. Таким образом, базальное тело структурно идентично центриолям, которые находятся в центросоме, расположенной рядом с ядром клетки.У некоторых организмов, таких как одноклеточный Chlamydomonas , базальные тельца локально и функционально изменяются в центриоли, а их жгутики резорбируются перед делением клеток.
Реснички и жгутики эукариот обычно различаются по размеру и количеству: реснички обычно короче и встречаются вместе в гораздо большем количестве, чем жгутики, которые часто бывают одиночными. Структуры также демонстрируют несколько разные типы движения, хотя в обоих случаях движение генерируется активацией динеина и, как следствие, изгибом аксонемы.Движение ресничек часто описывают как движение плети или сравнивают с взмахом груди при плавании. Соседние реснички движутся почти одновременно (но не совсем), так что в группах ресничек возникают волнообразные паттерны движения. Однако у эукариот жгутики демонстрируют плавные, независимые волнообразные движения. Прокариотические жгутики, которые имеют совершенно иную структуру, построенную на основе белка , флагеллина , движутся во вращательном режиме с помощью базального двигателя.
Дефекты ресничек и жгутиков клеток человека связаны с некоторыми заметными проблемами со здоровьем.Например, наследственное состояние, известное как синдром Картагенера, вызвано проблемами с плечами динеина, которые проходят между микротрубочками, присутствующими в аксонеме, и характеризуется повторяющимися респираторными инфекциями, связанными с неспособностью ресничек дыхательных путей избавляться от бактерий или бактерий. другие материалы. Заболевание также приводит к мужскому бесплодию из-за неспособности сперматозоидов продвигаться через жгутики. Повреждение респираторных ресничек также может быть приобретенным, а не унаследованным, и чаще всего связано с курением сигарет.Например, бронхит часто вызывается скоплением слизи и дегтя в легких, которые невозможно удалить должным образом из-за повреждения ресничек, вызванного курением.
НАЗАД К СТРУКТУРЕ КЛЕТКИ ЖИВОТНЫХ
Вопросы или комментарии? Отправить нам письмо.
© 1995-2021, автор — Майкл В. Дэвидсон и Государственный университет Флориды. Все права защищены. Никакие изображения, графика, программное обеспечение, сценарии или апплеты не могут быть воспроизведены или использованы каким-либо образом без разрешения правообладателей.Использование этого веб-сайта означает, что вы соглашаетесь со всеми юридическими положениями и условиями, изложенными владельцами.
Этот веб-сайт обслуживается нашим
Команда разработчиков графики и веб-программирования
в сотрудничестве с оптической микроскопией в Национальной лаборатории сильного магнитного поля
.
Последнее изменение: пятница, 13 ноября 2015 г., 14:18
Счетчик доступа с 1 октября 2000 г .: 463134
Микроскопы предоставлены:
| Распечатать эту страницу
|
Строение и роль жгутиков у прокариот
Жгутики в основном используются для движения клеток и обнаруживаются у прокариот, а также у некоторых эукариот.Прокариотический жгутик вращается, создавая движение вперед нитью в форме штопора. Прокариот может иметь один или несколько жгутиков, расположенных на одном полюсе или разбросанных по клетке.
Структура и происхождение
Жгутик состоит из тела у основания, которое встроено в клеточную мембрану, нити или стержня, который является главным штопором вне клетки, и крючка для соединения тела и нити. Жгутик также имеет свой собственный экспортный аппарат, так как он может собираться самостоятельно.Крючок имеет заданную длину, до которой он растет, тогда как нить более изменчива. Считается, что установленная длина определяется кольцом C, которое измеряет крючок перед его экспортом. Для облегчения самосборки центр жгутика полый, от основания до кончика нити.
Иллюстрация клеточной структуры бактерий. Кредит изображения: BlueRingMedia / Shutterstock
Строение жгутика различается в зависимости от того, у прокариот он или у эукариот.У эукариот жгутик бьется подобно хлысту, тогда как у прокариот жгутик представляет собой неподвижную пробковидную сущность, крутящий момент которой зависит от двигателя в его основании.
Строение жгутика сложное. В основании ротор находится внутри клеточной мембраны. Белки MotA и MotB образуют кольца, расположенные внутри и над внутренней мембраной, чтобы объединить энергию движения протонов через мембрану с генерацией крутящего момента. MotA и MotB — это статическая невращающаяся часть конструкции.
MS, P и L кольца представляют собой белковые кольца, которые обеспечивают вращение главного внеклеточного стержня штопора через мембраны. Кольцо MS прикрепляет жгутики к цитоплазматической мембране, и именно здесь, вероятно, расположена экспортная система. Кольцо P облегчает вращение внутри пептидогликана, а кольцо L внутри внешних мембран. У грамположительных бактерий кольца P и L. Кольцо С расположено внутриклеточно и регулирует направление вращения. Направление вращения контролируется хемотаксической системой, на которую реагирует С-кольцо.
Роль
Жгутик — это в основном органелла для движения. Однако он также может участвовать в образовании биопленок, экспорте белков и адгезии. Адгезия важна для многих жизненных циклов бактерий, и у них есть несколько механизмов, таких как фимбрии, пили и другие белки, которые помогают в этом. Жгутики и адгезивные структуры обычно не экспрессируются одновременно, а скорее бактерии переключаются из подвижной формы в стационарную.
Бактериальные жгутики могут играть важную роль и с точки зрения человека.Жгутики обычно необходимы для инфекции, и из-за этого они имеют молекулярные структуры, ассоциированные с патогенами (PAMP). PAMPs могут распознаваться иммунной системой человека через toll-подобный рецептор 5 (TLR5). N- и C-концы флагеллина, составляющего основной стержень, являются высококонсервативными и являются тем, с чем связывается TLR5. Существуют также полимерные формы флагеллина, которые не связывают TLR5.
В отличие от концов, центральная область флагеллина имеет более высокую вариацию последовательности. Эта открытая область определяет различия в адгезивной функции флагеллинов разных штаммов или видов бактерий. Pseudomonas aeruginosa может вызывать инфекции мочевыводящих путей (ИМП) и другие состояния, например пневмонию. Его связыванию, по-видимому, способствуют флагеллин (FliC) и / или FliD, белки, которые покрывают концы стержня.
Бактерия Pseudomonas aeruginosa — нозокомиальная бактерия, устойчивая к антибиотикам. На рисунке показано полярное расположение жгутиков и наличие пилей на поверхности бактерий. Кредит изображения: Катерина Кон / Shutterstock
Роль жгутиков в образовании биопленок также важна с точки зрения человека из-за опасного образования биопленок на катетерах и оборудовании.Исследования Escherichia coli показали, что не флагеллированные бактерии (то есть без основного стержня, без флагеллина и FliD) демонстрируют пониженную адгезию и, следовательно, способность к образованию биопленок. Напротив, жгутиковые бактерии, которые не могли двигаться (то есть с отсутствием мотора, без MotB), были способны колонизировать так же, как и штаммы дикого типа.