
Строение клеточного центра — Науколандия
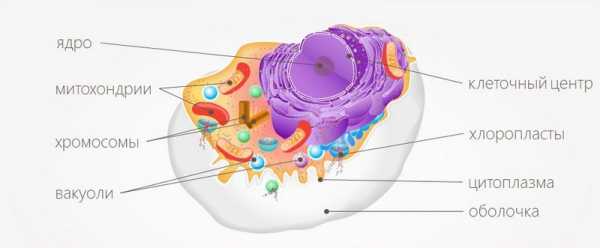
Клеточный центр — это органоид эукариотических клеток. По-другому клеточный центр называется центросомой. В большинстве клеток центросома включает две центриоли. Однако в клетках высших растений и некоторых других организмов клеточный центр есть, а центриолей (или центросомы) нет.
Обычно в неделящейся клетке бывает только одна центросома, и находится она в центральной ее области.
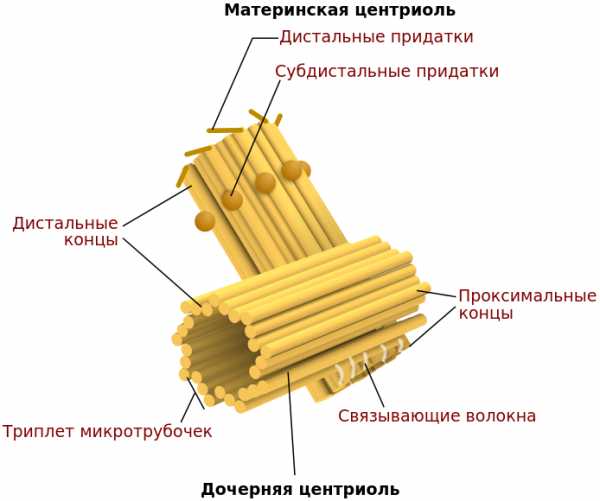
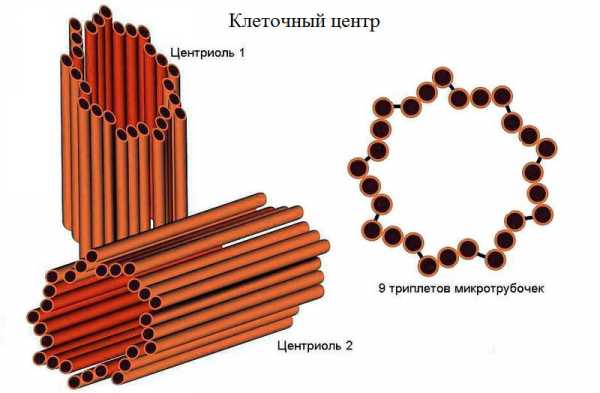
Центриоль — немембранный органоид. Каждая центриоль состоит из девяти триплетов микротрубочек, которые образует белок тубулин. Триплеты соединены между собой таким образом, что создается цилиндр. Высота цилиндра относится к его диаметру как 3 : 1. Средняя высота составляет около 0,3 мкм, а диаметр — около 0,1 мкм. Две центриоли располагаются под углом 90° друг к другу.
Однако строение клеточного центра несколько сложнее. Кроме пары центриолей в нем образуется сеть волокон и отходящих микротрубочек. Причем одна из центриолей является материнской и именно на ней формируются дополнительные образования.
Основная функция клеточного центра — это организация веретена деления. У животных и многих грибных клеток в процессе клеточного деления центриоли центросомы расходятся к различным полюсам клетки. Около каждой путем самосборки из тубулина образуется парная дочерняя центриоль (или она образуется позже, после деления).
Таким образом, в клетке оказывается два клеточных центра. От каждого в направлении к центру, к хромосомам, осуществляется сборка микротрубочек. Микротрубочки прикрепляются к центромерам хромосом и обеспечивают их равноценное расхождение к полюсам, или обеспечивают расхождение хроматид путем их отрыва друг от друга.
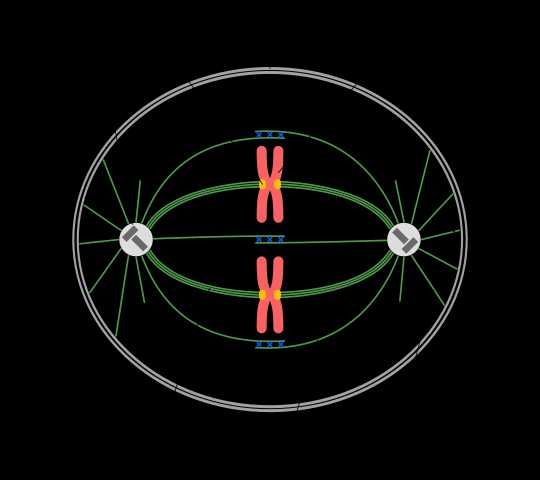
При расхождении происходит разборка микротрубочек с так называемого минус-конца, который находится в клеточном центре. Трубочка уменьшается и тем самым притягивает хромосому к своему полюсу клетки.
У растений веретено деления образуется без участия центриолей.
Кроме образования веретена деления клеточный центр выполняет и другие функции. В нем образуются микротрубочки для поддержания структуры клетки, базальные тельца ресничек и жгутиков.
scienceland.info
открытие в науке, значение, строение и функции
Клеточный центр (или центросома) — не мембранная органелла, которая находится в центре клетки, рядом с ядром. Отсюда и пошло название органоида. Присутствует только у низших растений и животных; высшие растения, грибы и некоторые простейшие лишены его.
Открытие в науке
Описание центросом на полюсах веретена деления, которые находятся в клетках во время митоза, сделали почти одновременно ученые-биологи Флеминг В. и Гертвиг О. Открытие сделано в 70-х годах XIX ст.
Ученые еще тогда установили, что после завершения митоза, центросомы не исчезают, а остаются в интерфазном периоде. Подробное строение удалось определить после появления электронной микроскопии в середине XX ст.
Функции и строение
Клеточный центр — органоид, видимый в оптический микроскоп в клетках животных и низших растений. Он находится обычно около ядра или в геометрическом центре клетки и состоит из двух палочковидных телец центриолей, размером около 0,3-1 мкм.
Под электронным микроскопом установлено, что центриоль представляет собой цилиндр, стенки которого построены девятью триплетами очень тонких трубочек. Каждый триплет включает 2 неполных набора — 11 протофибрил и 1 полный — 13 протофибрил.
Все центриоли имеют белковую ось, от которой к триплетам направляются тонкие нити из белка. Центриоли находятся в окружении бесструктурного вещества — центриолярного матрикса. Здесь происходит формирование микротрубочек, благодаря белку гамма-тубулину.
В клеточный центр входят две центриоли: дочерняя и материнская, которые взаимно перпендикулярны друг к другу и вместе формируют диплосому. Материнская центриоль в составе имеет дополнительные структурные элементы — сатиллиты, их количество постоянно меняется, и располагаются они на всем протяжении центриоли.
 Строение клеточного центра
Строение клеточного центраВ середине цилиндра находится полость, заполненная однородной массой. Пара центриолей, окружена более светлой зоной, называется центросферой.
Центросфера состоит из фибриллярных белков (основной — коллаген). Здесь располагаются микротрубочки, много микрофибрилл и скелетных фибрилл, которые обеспечивают фиксацию клеточного центра возле ядерной оболочки. Только в эукариотических клетках центриоли находятся под прямым углом относительно друг друга. Простейшим, нематодам не характерно такое строение.
| Цитологическая характеристика | ||
|---|---|---|
| Структурные элементы | Строение | Функции |
| Центриолярный матрикс | Немембранное образование, состоящее из белка гамма-тубулина | Принимает участие в создании микротрубочек |
| Центросома | Представлена парой сформированных центриолей, в составе которых имеется девять триплетов микротрубочек. Построены из белка коллагена и располагаются перпендикулярно относительно друг друга. | Отвечает за образование веретена деления, формирует цитоскелет |
Механизм распределения генетической информации
Перед митозом клеточный центр удваивается, при этом материнские центриоли рассоединяются и расходятся к противоположным полюсам.
Так в клетке появляется два клеточных центра. От них по направлению к центру, к хроматидам, идет сборка микротрубочек. Микротрубочки крепятся к центромерам пар хроматид и обеспечивают их равномерное распределение по дочерним клеткам.
Во время расхождения идет разборка микротрубочек с минус-конца, который расположен в центросоме. Микротрубочка укорачивается и, таким образом, тянет хромосому к определенному полюсу клетки. Каждая новообразованная клетка получает диплоидный набор хромосом и по одной центросоме.
Значение
Клеточный центр — главная структура, отвечающая за создание и управление микротрубочками клетки.
Выполняет такие функции:
- Формирование органоидов движения простейших организмов (жгутики), которые дают возможность перемещаться в водной среде.
- Образует реснички на поверхности эукариотических клеток, которые необходимы для восприятия внешних раздражителей (кожная рецепция).
- Формирует нити веретена деления во время непрямого, митотического деления клетки. Обеспечивает равное распределение генетической информации между дочерними клетками.
- Принимает участие в формировании микротрубочек, которые уходят или в цитоплазму, или становятся компонентом опорно-сократительного аппарата.
- Увеличение количества центросом характерно для опухолевых клеток.
Клеточный центр играет важную роль в процессе перемещения хромосом при митозе. С ним связана способность некоторых клеток к активному движению. Это доказывается тем, что в основании жгутиков или ресничек подвижных клеток (простейшие, сперматозооны) находятся образования такой же структуры, как и клеточный центр.
animals-world.ru
Пластиды. Клеточный центр. Органеллы движения
Пластиды
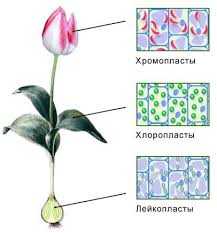
Пластиды свойственны лишь растительным клеткам. Они образуются у высших растений из пропластид – мелких телец меристематической зоны растений, круженных двойной мембраной, из которой в будущем сформируется оболочка пластиды.
Хлоропласты – это пластиды, содержащие хлорофилл и каротиноиды и способные к фотосинтезу. Расположены в основном в листьях растений. У высших растений на срезах хлоропласты обычно имеют вид двояковыпуклой линзіы а сверху — округлые.
Диаметр хлоропластов около 3-10 мкм (в среднем 5 мкм), потому их хорошо видно в световой микроскоп.
Хлоропласты всегда содержат хлорофилл и другие пигменты, участвующие в процессе фотосинтеза. Они локализированны в системе мембран, которые погружены в основное вещество хлоропласта – строму.
Мембранная система – это то место, где осуществляется световая фаза фотосинтеза. Мембраны содержат хлорофилл и другие пигменты, ферменты.
Вся система образована большим количеством плоских мешочков, заполненных жидкостью –
Замечание 1
Одной из интересных особенностей у хлоропластов, кроме их способности к фотосинтезу, есть ещё одна интересная особенность — наличие системы, синтезирующей белки.
Хромопласты – это окрашенные пластиды, неспособные к фотосинтезу, которые содержат в основном красные, жёлтые и оранжевые пигменты (каротиноиды).Больше всего хромопластов в плодах и цветах.
Замечание 2
Оранжевый пигмент, от которого зависит окраска корня моркови, также находится в хромопластах.
Лейкопласты – это бесцветные пластиды, не содержащие пигментов. Они приспособлены для сохранения питательных веществ, потому их особенно много в запасающих органах растений – семенах, корнях, а также в молодых листьях. Амилопласты запасают крахмал, липидоплатиды – липиды в виде масел или жиров, протеинопласты – белки.
В 60-х годах $XX$ ст. было установлено, что в хлоропластах и митохондриях есть ДНК и рибосомы. На основании этого была выдвинута гипотеза о том, что в тех клетках, где есть эти органеллы, они могут быть полностью или частично независимыми от клеточного ядра. Было высказано предположение, что митохондрии и хлоропласты – это прокариотические организмы, которые вошли в клетки эукариот на ранних этапах эволюции жизни. Согласно с теорией эндосимбиоза, эти органеллы воплощают крайнюю форму симбиоза.
Клеточный центр
В клетках животных возле ядра расположена органелла, которую называют клеточным центром.
Основную часть клеточного центра составляют два маленьких тельца – центриоли, расположенные в небольшом участке уплотнённой цитоплазмы
Каждая центриоля имеет форму цилиндра длиной до 1 мкм.
Центриоли выполняют важную роль в делении клетки: они участвуют в образовании веретена деления.
Органеллы движения
Псевдоподии (ложноножки) – амёбообразные временные выпячивания цитоплазмы некоторых простейших (амёб, радиолярий) или клеток многоклеточных организмов (лейкоцитов).
Псевдоподии образуются благодаря движению цитоплазмы. В основе амебоидного движения – движение молекул сократительных белков. Кроме движения клеток, псевдоподии обеспечивают также процесс фагоцитоза — захват твёрдых питательных частиц.
Жгутики и реснички – нитевидные выросты клеточной поверхности.
Реснички обычно имеют в немного меньшую длину, чем жгутики, — она составляет до 15 мкм, тогда как у жгутиков – до 50-100 мкм.
С помощью электронного микроскопа установлено, что они имеют общую основную структуру:
- девять пар микротрубочек, расположенных кольцеобразно,
- две одиночные микротрубочки в центре.
В основании кольца расположено базальное тельце. У жгутиков этих телец два, а у ресничек – одно.
Жгутики двигаются волнообразно или винтообразно, осуществляется с помощью освобожденной энергии АТФ. Движения ресничек напоминают работу вёсел.
Замечание 3
Реснички помогают некоторым мелким организмам (инфузориям) двигаться в жидкой среде или образовывать у поверхности некоторых клеток поток жидкости, втягивающей за собой различные частицы.
Пример 1
Реснички клеток эпителия, выстилающих дыхательные пути, очищают воздух, проходящий по этих путях, от слизи и пыли.
Жгутики есть у многих одноклеточных организмов (хламидомонада, жгутиковые), также сперматозоидов.
spravochnick.ru
Глава 23. Клеточный центр
Итак, в клетках животных, растений и одноклеточных микротрубочки поляризованы, так что большей частью их растущие (+)-концы направлены к периферии клетки. Это связано с тем, что МТ начинают свой рост от специальных участков в клетке, от центров организации микротрубочек (ЦОМТ). Некоторые из ЦОМТ имеют сложную морфологическую организацию, другие устроены иначе. Различные ЦОМТ можно разделить на несколько групп: центросомные клеточные центры, и центры организации микротрубочек, не имеющие четкой локализации.
Так например, в клетках высших растений полимеризация МТ происходит по периферии клеточного ядра, от которого МТ расходятся радиально. Сходная картина наблюдается при регенерации МТ в гигантских клетках слюнных желез двукрылых. В ряде случаев новообразование МТ, их закладка, нуклеация, может происходить в цитоплазме вне связи со специальными зонами или структурами.
Но в большинстве случаев в интерфазных клетках животных организмов образование и рост МТ происходит от клеточного центра, содержащего специальные образования – центросомы, которые большей частью могут содержать сложно организованные центриоли, или же не иметь их.
Центросомы и центриоли
Центросомы были обнаружены и описаны сто лет назад (Флемминг, 1875; Бенеден, 1876) – это очень мелкие тельца, размер которых находится на границе разрешающей способности светового микроскопа, обычно располагающиеся в геометрическом центре клетки, откуда и их название. В некоторых объектах удавалось видеть, что мелкие плотные тельца (центриоли), обычно в паре (диплосома), окружены зоной более светлой цитоплазмы (собственно центросома), от которой отходят радиально тонкие фибриллы (центросфера) (рис. 277).
Центросомы характерны и обязательны для клеток животных, и нет у высших растений, у низших грибов и некоторых простейших. Было замечено, что центросомы в делящихся клетках принимают участие в формировании веретена деления и располагаются на его полюсах. В неделящихся клетках центросомы часть определяют полярность клеток эпителия и располагаются вблизи аппарата Гольджи. Такая связь центросом с аппаратом Гольджи характерна для многих клеток, в том числе для клеток крови и нервных клеток. Часто центросомы лежат рядом с ядром, располагаясь в зонах его впячивания. Например, в полиморфных лейкоцитах (нейрофилы) центросома лежит внутри подкововидного впячивания ядра (рис. 278).
Типичное строение клеточный центр имеет в клетках животных. Он представляет собой зону, состоящую из центриолей и окружающей их аморфной фибриллярной массы или матрикса. В ряде случаев в состав клеточного центра или центросомы входит только эта фибриллярная масса, от которой отходят микротрубочки (см. ниже).
Наиболее же часто кроме матрикса в состав клеточного центра входят центриоли, как мелкие тельца, с трудом наблюдаемые в световом микроскопе.
Тонкое строение центриолей удалось изучить только с помощью электронного микроскопа. Основу строения центриолей составляют расположенные по окружности девять триплетов микротрубочек, образующие таким образом полый цилиндр (рис. 279). Его ширина около 0, 15 мкм, а длина такого цилиндра 0,3-0,5 мкм (хотя встречаются центриоли, достигающие в длину несколько микрон) (рис. 280).
Первая микротрубочка триплета (А-микротрубочка) имеет диаметр около 25 нм и толщину стенки 5 нм, которая состоит из 13 глобулярных субъединиц. Длина каждого триплета равна длине центриоли. Вторая и третья (В и С) микротрубочки отличаются от А-микротрубочки тем, что они являются неполными, содержат 11 субъединиц и вплотную примыкают к своим соседям. Каждый триплет располагается к радиусу такого цилиндра под углом около 400. Кроме микротрубочек в состав центриоли входит ряд дополнительных структур. От А-микротрубочки отходят так называемые “ручки”, выросты, один из которых (внешний) направлен к С- микротрубочке соседнего триплета, а другой (внутренний) – к центру цилиндра.
Обычно в интерфазных клетках всегда присутствуют две центриоли, располагающиеся рядом друг с другом, образуя дуплет центриолей, или диплосому (рис. 281). В диплосоме центриоли располагаются под прямым углом по отношению друг к другу. Из двух центриолей различают “материнскую” и “дочернюю”, продольная ось последней перпендикулярна продольной оси материнской центриоли. Обе центриоли сближены своими концами так, что проксимальный конец дочерней центриоли как бы смотрит на поверхность материнской. В дистальном участке материнской центриоли располагается аморфный материал в виде выростов или шпор – это придатки. Их нет на дочерней центриоли (рис. ).
Дочерняя центриоль несколько отличается от материнской. Центральная часть цилиндра центриоли занята структурой, напоминающей тележное колесо; она имеет центральную “втулку” диаметром около 25 нм и 9 спиц, направленных по одной к А-микротрубочке каждого из триплетов. Такие структуры внутри центриоли расположены в одном из её концов, проксимальном, что делает строение цилиндра центриоли полярным. На дистальном конце центриоли внутри её нет таких структур. Объем, занимаемый внутри центриоли втулкой со спицами, может составлять у разных клеток от 3\4 до 1\5 длины центриоли. У некоторых видов втулка отсутствует или заменена скоплением аморфного материала. Торцы центриолярного цилиндра, кроме системы втулки и спиц на проксимальном конце, ничем не закрыты.
Систему микротрубочек центриоли обычно описывают формулой 9 + 0, или (9х3) + 0, подчеркивая отсутствие микротрубочек в её центральной части.
Вокруг каждой центриоли расположен бесструктурный, или тонковолокнистый матрикс. Сами микротрубочки триплетов погружены в аморфный материал т.н. муфты или оправы. Если выделенные центриоли обработать 0,6М раствором NaCl, то произойдет полная экстракция микротрубочек, но центриоль как таковая не растворится: вместо нее останется цилиндрическая структура, имеющая девять полых отверстий, некогда занимавшихся триплетами микротрубочек. Поэтому все схемы центриолей в этой книге, как и во многих других значительно упрощены и не включают материал муфты центриолярного цилиндра.
Часто около центриолей и в связи с ним можно обнаружить несколько дополнительных структур: сателлиты, фокусы схождения микротрубочек, исчерченные волокнистые корешки, дополнительные микротрубочки, образующие особую зону – центросферу вокруг центриоли (рис. 282).
При исследовании в электронном микроскопе интерфазных центриолей было найдено, что лучистое сияние центросферы, обнаруживаемое в световом микроскопе, представляет собой большое число микротрубочек, радиально расходящихся от зоны диплосомы. В диплосоме лишь одна из центриолей, материнская, содержит ряд дополнительных структур. Одни из них, перицентриолярные сателлиты, состоят из имеющей тонкое фибриллярное строение конусовидной ножки, расположенной на стенке центриоли, и головки, заканчивающейся на этой ножке. Ножки сателлитов часто имеют поперечную исчерченность (рис. 284). Количество таких перицентриолярных сателлитов непостоянно, они могут располагаться на разных уровнях по длине центриоли. Кроме этих структур рядом с диплосомой, но не связанные с ней структурно, могут располагаться плотные мелкие (20-40 нм) тельца, к которым подходят одна или несколько микротрубочек (фокусы схождения микротрубочек). Микротрубочки отходят и от головок сателлитов. Эти центросомные микротрубочки не отходят непосредственно от микротрубочек цилиндров центриолей, а связаны или с сателлитами, или с матриксом. Такие микротрубочки и образуют как бы лучистую сферу (центросферу) вокруг центриоли, где (-)-концы МТ связаны с ЦОМТ, а (+)-концы радиально расходятся на периферию клетки. При образовании центросферы в интерфазной клетке только специальные структуры центриоли, сателлиты и матрикс, каким-то образом связаны с образованием микротрубочек; микротрубочки самих центриолей в этом процессе не участвуют. Было найдено, что восстановление прицентриолярных микротрубочек после их деполимеризации на холоду происходит за счет появления новых микротрубочек, отходящих от головок сателлитов. Таким образом, можно считать, что эти дополнительные структуры являются центрами, на которых происходит сборка микротрубочек из тубулинов ( центры организации микротрубочек – ЦОМТ).
Химия центриолей изучена слабо, потому что еще не разработаны методы получения этой структуры в виде чистой фракции. Трудности биохимического изучения центриолей связаны с тем, что это одиночная клеточная структура, имеющая объем всего 0, 03 мкм3. Для сравнения, вспомним, что в клетке имеется: около тысячи штук митохондрий, около миллиона рибосом, около сотни хромосом, около 1 мм2 мембран.
Есть все основания говорить о том, что в состав микротрубочек центриолей входят тубулины. Это доказывается тем, что колхицин прекращает рост микротрубочек в процентриолях, возникающих вблизи материнской центриоли. Предположения о возможной химической природе остальных элементов центриоли основаны главным образом на данных, полученных из химии ресничек и жгутиков, имеющих много сходных черт строения с центриолями.
Данные о химическом строении центриолей получены главным образом с помощью иммунохимических методов. В изолированных базальных тельцах простейшего хламидомонады обнаружено более 200 различных белков, среди которых выявлены четыре вида тубулинов, в том числе - тубулин, центрин, перицентрин, белок р210 и многие другие.
В интерфазных клетках центриоли оказываются связаны с ядром и с ядерной мембраной. При выделении ядер практически все центриоли клеток печени и селезенки крыс оказываются в этой фракции. Связь центриолей с ядром осуществляется главным образом промежуточными филаментами. Если живые клетки подвергнуть ультрацентрифугированию, то центриоли опускаются к центробежному полюсу вместе с ядрами.
Центросомный цикл
Было обнаружено, что строение и активность центросом меняются в зависимости от периода клеточного цикла, в течение которого клеточный центр претерпевает тоже циклические изменения (рис. 283).
Целесообразнее начать рассмотрение циклических изменений в структуре центросом с митоза. Начиная с профазы и кончая телофазой, центросомы имеют сходное строение, несмотря на то, что за время митоза происходит ряд существенных клеточных перестроек: конденсация хромосом, разрушение ядерной оболочки, образование веретена деления, расхождение хромосом. В митозе в клеточных центрах (их два, по одному на каждый полюс клетки) находится по диплосоме. Как полагается, дочерняя центриоль своим концом направлена на материнскую. Материнская центриоль на всех стадиях митоза окружена довольно широкой (до 0,3 мкм) зоной тонких фибрилл – центриолярное фибриллярное гало (рис. 279). От этого гало радиально отходят микротрубочки. Важно подчеркнуть, что у дочерних центриолей ни гало, ни отходящих от центриолей микротрубочек нет. В это время происходит формирование веретена митотического аппарата, состоящего из микротрубочек. Эта структура действительно имеет форму веретена, где на концах его на полюсах клетки располагаются диплосомы, окруженные радиальными микротрубочками (центросфера). В данном случае можно говорить о том, что зоны диплосом, клеточные центры, являются центрами организации (полимеризации) микротрубочек. В пользу этого говорят следующие факты: после исчезновения микротрубочек веретена и центросферы, которые происходят при действии холода или колхицина, новые микротрубочки возникают главным образом в районе материнских центриолей, диплосом, в каждом из полюсов клетки. Интересно, что рост новых микротрубочек не связан с микротрубочками триплетов центриолярного цилиндра, они начинают отрастать от зоны гало, расположенной на материнской центриоли. Важно отметить, что в это время на материнских центриолях (как и на дочерних) нет сателлитов, и в это же время цитоплазма теряет микротрубочки: микротрубочки цитоплазмы разбираются, а пул освободившихся тубулиновых мономеров идет на образование микротрубочек веретена и центросферы, которые образуются на фибриллярном гало, как на ЦОМТ. Этот процесс полимеризации митотических микротрубочек отражает первую форму активности центриолярного аппарата (рис. ). Если в профазе облучить центриоль лазерным микролучем, то образование веретена останавливается.
Примерно сходное строение имеют клеточные центры на всех стадиях митоза, но к телофазе толщина фибриллярного гало уменьшается.
К концу телофазы, когда произошло разделение клетки надвое, а хромосомы начали деконденсироваться и образовывать новые интерфазные ядра, происходит разрушение веретена деления, его микротрубочки деполимеризуются. Клеточные центры при этом меняют свою структуру. Материнская и дочерняя центриоли теряют взаимно перпендикулярное расположение и отходят друг от друга на небольшие (0,5-2мкм) расстояния, но все же держатся в одном месте. Вокруг материнской центриоли гало и микротрубочки не выявляются. В это время микротрубочек в цитоплазме также практически нет.
В начале G1-периода на поверхности материнской центриоли возникают сателлиты, имеющие ножку и головку, от которой радиально отходят микротрубочки, которые начинают расти в длину и заполнять собой цитоплазму (рис. 284а). Следовательно, вторая форма активности клеточного центра – образование цитоплазматических микротрубочек в интерфазных клетках. Надо подчеркнуть, что активной здесь является только материнская центриоль, которую легко узнать по придаткам в ее дистальной части.
Если считать клеточные центры основными (если не единственными) местами образования цитоплазматических микротрубочек, то общее количество последних должно быть равно числу микротрубочек, отходящих от центриолей. При исследовании в электронном микроскопе оказалось, что от клеточных центров в интерфазе отходит всего лишь несколько десятков микротрубочек, а в цитоплазме их так много, что с помощью иммунофлуоресцентного метода их трудно подсчитать. Это дает основание предполагать, что по мере роста микротрубочек часть из них теряет связь с областью центриолей и может находиться в цитоплазме долгое время. Центросомы же индуцируют полимеризацию новых микротрубочек, которые приходят на смену постепенно деполимеризующимся старым. Вероятно, в цитоплазме есть несколько генераций микротрубочек: “старые”, не связанные с клеточным центром, и новые, растущие от центросом. Таким образом, в клетке происходит как бы конвейерная смена и репродукция цитоплазматических микротрубочек.
Если клеткам запретить переходить в S-период, они могут существовать в фазе клеточного покоя (Go-период) (рис. 285). В это время материнская центриоль продолжает функционировать, как центр образования микротрубочек цитоскелета. Но одновременно она может проявить еще одну форму активности – образовать ресничку, вырост плазматической мембраны, заполненный аксонемой (осевой нитью), состоящей из девяти дублетов микротрубочек. Эти микротрубочки отрастают, как от затравок, от А- и В-микротрубочек триплетов материнской центриоли в дистальной ее части. Это – третья форма активности центриолей как центров организации микротрубочек (см. ниже).
При наступлении S-периода (или в середине его) клеточный центр приступает к четвертой форме своей активности: происходит удвоение числа центриолей. В это время около каждой из разошедшихся еще в конце телофазы центриолей, материнской и дочерней, происходит закладка новых центриолярных цилиндров – процентриолей (рис. 284б). В районе проксимальных концов каждой центриоли перпендикулярно длинной оси закладывается сначала девять синглетов (одиночных) микротрубочек, затем они преобразуются в девять дуплетов, а потом – в девять триплетов растущих микротрубочек новых центриолярных цилиндров.
Закладка процентриолей происходит на проксимальных концах центриолей; в этом месте растут новые поколения центриолей, тоже с проксимального конца. Во время роста процентриолей здесь можно видеть центральную “втулку” со спицами.
Благодаря такому росту структур образуется сначала короткая дочерняя центриоль – процентриоль — которая затем дорастает до размера материнской. Этот способ увеличения числа центриолей был назван дупликацией. Важно отметить, что размножение центриолей не связано с их делением, почкованием или фрагментацией, а происходит путем образования зачатка, процентриоли, вблизи и перпендикулярно к исходной центриоли. Правда, последнее условие соблюдается не во всех объектах, у некоторых оомицетов при дупликации центриоли происходит сначала расхождение центриолей, рост втулки, затем рост микротрубочек вдоль продолжения оси исходной центриоли, и центриоли располагаются конец в конец. Интересно, что триплеты в таких новых центриолях имеют угол наклона, противоположный таковому в материнской центриоли.
Факт удвоения центриолей привел некоторых исследователей к предположению, что центриоли, так же как митохондрии и пластиды, принадлежат к саморедуплицирующимся компонентам цитоплазмы, хотя прямых данных о наличии ДНК в составе центриолей нет.
В S-периоде во время удвоения (дупликации) центриолей материнская продолжает проявлять вторую форму активности: она продолжает быть центром образования цитоплазматических микротрубочек.
В результате процесса дупликации около каждой центриоли вырастает новая дочерняя центриоль (первая материнская центриоль и дочерняя на бывшей дочерней центриоли могут считаться как бы бабушкой и внучкой). Поэтому в клетке после завершения S-периода находятся уже две диплосомы (а всего четыре центриолярных цилиндра) (рис. 286).
После этого наступает следующий период клеточного цикла, постсинтетический (G2-период), когда в клетке начинается подготовка к очередному делению. В это время исчезают сателлиты на материнской диплосоме (так можно назвать старую материнскую центриоль с новой дочерней), а обе материнские центриоли в обеих диплосомах покрываются фибриллярным гало, от которого в профазе начинают отрастать митотические микротрубочки. Параллельно этому в цитоплазме происходит исчезновение микротрубочек, и клетка стремится приобрести шаровидную форму. Вся такая последовательность событий повторяется от цикла к циклу у клеток, способных к длительному размножению. В большинстве случаев клетки организма находятся в G0-периоде, поэтому у них центриоль участвует в полимеризации цитоплазматических микротрубочек и в образовании реснички (или множества ресничек). В последнем случае она входит в состав так называемого базального тельца.
Обычно в клетку после деления попадают два центриолярных цилиндра в составе диплосомы. В различных экспериментальных условиях можно запретить разделение клетки надвое и получить клетки с удвоенным числом хромосом (полиплоидные клетки). Совершенно очевидно, что в таких клетках будет и удвоенное число центриолей. Клетки могут снова вступать в клеточный цикл, при этом будет удваиваться как количество ДНК, так и число центриолей. Было обнаружено, что у тетраплоидных (с четырехкратным набором хромосом) клеток печени в G0-периоде в цитоплазме видны не два, а четыре центриолярных цилиндра, а в полюсах при делении таких клеток было обнаружено по две диплосомы в каждом. Аналогичная ситуация замечена и у других полиплоидных клеток (мегакариоциты костного мозга, полиплоидные гибридные клетки и др.). В связи с этим предположили, что между числом плоидности клетки (числом хромосомных наборов) и числом центриолей существует прямая связь.
Нарушения центриолярного цикла могут вызвать ряд патологических изменений клеток, в первую очередь появление многополюсных митозов. Так, при действии β-меркаптоэтанола происходит блокада нормального митоза, при этом диплосомы расходятся на отдельные центриоли. При отмывании от этого вещества клетка снова приступает к делению, но в этом случае каждая центриоль активируется и образует полюс веретена. Таким образом, возникают трех- или четырехполюсные митозы, приводящие к неравномерному распределению хромосом между дочерними клетками. Это в свою очередь приводит к изменению числа хромосом (анэуплоидия), которое часто вызывает гибель клетки. Иногда при образовании многополюсных митозов в некоторых полюсах отсутствуют центриоли: в полюсе располагается только фибриллярный материал центросомы (бесцентриолярные полюса).
Итак, в подавляющем большинстве клеток млекопитающих центросомы участвуют в полимеризации тубулинов и являются структурами, играющими роль центров организации микротрубочек. Микротрубочки самих центриолей являются затравками для полимеризации тубулинов только в одном случае – при росте аксонемы реснички, когда центриоль становится базальным тельцем. Это временное состояние: при переходе клеток к делению реснички могут исчезать, а базальное тельце снова может выполнять роль центриоли, участвуя в организации цитоплазматических микротрубочек или микротрубочек митотического веретена. Только в этих случаях центрами организации микротрубочек являются не сами центриолярные цилиндры, а перицентриолярный материал (головка сателлитов, околоцентриолярный матрикс, гало и т.д.). Следовательно, центриоль как таковую нужно рассматривать как один из компонентов более сложной структуры – клеточного центра или центросомы. Эта оговорка связана с тем, что у всех высших растений ЦОМТ не содержит центриолей. Более того, в раннем эмбриогенезе позвоночных животных образуются веретена деления, не имеющие центриолей в полюсах. По всей вероятности в последних случаях центриоли возникают позже заново, а не образуются путем “репликации”. Вопрос о процессе образования центриолей далек от решения. Остается неясным процесс появления процентриолей. В процессе эмбриогенеза отмечены случаи возникновения центриолей de novo у морского ежа, у моллюсков, у мышей. Так, в эмбриогенезе мыши центриоли появляются только после 1-2 делений клеток бластулы, несмотря на то, что сами клеточные деления происходят нормально, за исключением того, что в полюсах деления в зоне бесструктурной центросомы центриоли отсутствуют. С другой стороны, если в соматических клетках культуры ткани уничтожить центросому с центриолью с помощью микрооблучения, то новые центриоли не возникают.
Базальные тельца. Строение и движение ресничек и жгутиков.
Как уже указывалось, у многих клеток животных, вышедших из клеточного цикла, в G0-стадии центриоли принимают участие в образовании аппарата движения – ресничек. Их две группы: кинетоцилии, характерные для специальных эпителиев (ресничные эпителии трахеи, яйцеводов) или свободно плавающих клеток (сперматозоиды, простейшие), и так называемые первичные реснички, встречающиеся во многих клетках, не обладающих способностью к движению.
Вначале рассмотрим строение кинетоцилей – подвижных ресничек и жгутиков. В световом микроскопе эти структуры видны как тонкие выросты клетки, в их основании в цитоплазме видны хорошо красящиеся мелкие гранулы – базальные тельца, аналоги центриолей (рис. 287). Клетки, имеющие реснички или жгутики, обладают способностью двигаться, будучи в свободном состоянии, или же перемещать жидкости в случае, если клетки неподвижны. Свободноживущие одноклеточные организмы, снабженные одним или несколькими жгутиками, обычно движутся тем концом вперед, который несет жгутики. Иной способ движения можно видеть у спермиев некоторых животных: жгутик, располагаясь сзади, толкает тело клетки вперед. Скорость движения клеток за счет работы жгутиков может достигать очень большой величины (до 5 мм / мин).
Множественные реснички также могут обеспечивать движение свободноживущих клеток, таких как инфузории или некоторые жгутиконосцы. Реснички эпителиальных клеток многих беспозвоночных и позвоночных животных обеспечивают поток жидкостей вдоль поверхности таких клеток. Число ресничек на клетку может достигать 300 в эпителии трахеи; у инфузории туфельки на клетку приходится 10-14 тыс. рядами расположенных ресничек.
При движении ресничек и жгутиков не происходит уменьшения их длины, поэтому неправильно называть это движение сокращением. Траектория движения ресничек очень разнообразна (рис. 288). В различных клетках это движение может быть маятникообразным, крючкообразным, воронкообразным или волнообразным.
У многоресничных клеток (инфузории, клетки ресничного эпителия) движение ресничек не хаотично, а строго упорядочено. В этом случае реснички расположены рядами. В продольном ряду отдельные реснички начинают движение и проходят отдельные его фазы по очереди, метахронно. В поперечном же ряду все реснички находятся в одной фазе движения (синхронны). Это создает движущую волну по поверхности клетки (рис. )289.
Общая архитектура реснички представлена на рис. 290, 291. Ресничка представляет собой тонкий цилиндрический вырост цитоплазмы с постоянным диаметром 300 нм. Этот вырост от основания до самой его верхушки покрыт плазматической мембраной. Внутри выроста расположена аксонема, сложная структура, состоящая в основном из микротрубочек. Нижняя, проксимальная часть реснички, базальное тельце, погружена в цитоплазму. Диаметры аксонемы и базального тельца одинаковы (около 200 нм).
На поперечном сечении реснички видна плазматическая мембрана, окружающая аксонему. Аксонема в своем составе имеет девять дублетов микротрубочек, образующих внешнюю стенку цилиндра аксонемы. Дублеты микротрубочек слегка повернуты (около 100) по отношению к радиусу аксонемы. Кроме периферических дублетов микротрубочек в центре аксонемы располагается пара центральных микротрубочек. В целом систему микротрубочек реснички описывают как (9х2)+2. В дублетах микротрубочек также различают А-микротрубочку, состоящую из 13 субъединиц, и В-микротрубочку, неполную, содержащую 11 субъединиц. А-микротрубочка несет на себе ручки, которые направлены к В-микротрубочке соседнего дуплета. От А-микротрубочки к центру аксонемы отходит радиальная связка, или спица, оканчивающаяся головкой, присоединяющейся к центральной муфте, имеющей диаметр около 70 нм, окружающей две центральные микротрубочки. Последние лежат отдельно друг от друга на расстоянии около 25 нм. Таким образом, в аксонеме располагается 20 продольных микротрубочек, в то время как в базальном тельце их 27 (рис. 291, 292).
Базальное тельце состоит из 9 триплетов микротрубочек (как и центриоль), имеет ручки, втулку и спицы, расположенные в проксимальной (нижней) ее части. На участке базального тельца, примыкающем к плазматической мембране, есть девять придатков, выступов, идущих от каждого триплета микротрубочек к плазматической мембране и связывающих его с клеточной поверхностью.
Базальное тельце и аксонема структурно связаны друг с другом и составляют единое целое: А- и В-микротрубочки триплетов базального тельца продолжаются в А- и В-микротрубочках дуплетов аксонемы. Однако внутренние части аксонемы и базального тельца значительно отличны друг от друга. Часто в зоне перехода базального тела в аксонему наблюдают аморфную поперечную пластинку, которая как бы отделяет эти две части. Центральные микротрубочки аксонемы начинаются от этой пластинки так же, как в этом месте начинается и центральная муфта (капсула) (рис. 290).
В основании ресничек и жгутиков часто встречаются исчерченные корешки, или кинетодесмы, представляющие собой пучки тонких (6 нм) фибрилл, обладающих поперечной исчерченностью (рис. 293). Часто такие исчерченные кинетодесмы простираются от базальных телец вглубь цитоплазмы по направлению к ядру. Роль этих структур не ясна. Они не изменяются при действии колхицина, могут встречаться и в составе центриолей интерфазных клеток, не принимающих участия в образовании ресничек.
При движении ресничек не происходит изменения их длины, они не “сокращаются”, а изгибаются, бьются. Оказалось, что механически отделенные реснички способны к биению в присутствии АТФ. При отделении ресничек базальные тельца остаются в теле клетки. Это означает, что для механической работы ресничек базальное тело не нужно, а только аксонема участвует в генерации движения. Удалось показать, что за движение ресничек отвечают “ручки”, сидящие на А-микротрубочках. При экстракции компонентов ручек реснички перестают биться в присутствии АТФ.
Было найдено, что в состав ручек входят белки динеины. Это большие белковые компоненты, состоящие из 9-12 полипептидных цепей, содержащие 2-3 глобулярные головки, связанные в общий корешок гибкими хвостами (рис. 294). Каждая головка динеина обладает АТФ-азной активностью, которая возрастает примерно в 6 раз при ассоциации с микротрубочками. В состав каждой ручки входит один белковый комплекс, одна молекула динеина. Так как экстракция ручек прекращает биение ресничек, то можно считать, что именно динеин ответственен за это движение, то есть динеин является мотором или двигателем при биении ресничек. Но каков механизм этого движения?
Этот вопрос был решен при использовании выделенных ресничек, лишенных плазматической мембраны, радиальных спиц и связок после частичной обработки аксонем протеазами. Оказалось, что такие аксонемы, содержащие динеиновые ручки, при добавлении к ним АТФ начинают увеличиваться в длину почти до девяти раз и одновременно утончаются. В электронном микроскопе видно, что такая аксонема увеличилась в длину за счет смещения пар микротрубочек одна относительно другой (рис. 295). Другими словами, произошло продольное скольжение дуплетов один относительно другого, аналогично тому, что происходит при сокращении саркомеров в мышце: скольжение миозиновых нитей относительно актиновых. В случае динеина повторные циклы ассоциации с субъединицами тубулина, изменения конформации при связывании АТФ и его гидролизе, вызывают перемещение головок вдоль микротрубочки от (+)-конца к (-)-концу. При этом соседний дуплет двигается к верхушке реснички. Когда ресничка содержит все компоненты, и дуплеты микротрубочек связаны друг с другом и с центральной парой микротрубочек, такие кооперативные смещения дуплетов микротрубочек приводят не к удлинению реснички, а к ее изгибу (рис. 296). Как регулируется последовательное перемещение дуплетов один относительно другого, еще не ясно.
Рост ресничек, удлинение микротрубочек их аксонем происходит на вершине реснички. Следовательно, там локализованы (+)-концы микротрубочек.
Образование аксонемы ресничек происходит за счет роста А- и В-микротрубочек центриолей, которые в этом случае становятся базальным тельцем. В простейшем случае при образовании одиночных ресничек или так называемых первичных ресничек материнская центриоль подходит к плазматической мембране своим дистальным торцом, связывается с ней своими придатками. В это время начинается рост микротрубочек на (+)- концах А- и В-микротрубочек триплетов. Возникают девять дублетов микротрубочек аксонемы, которые, наращиваясь с (+)-концов на верхушке аксонемы как бы вытягивают плазматическую мембрану, образуя вырост – ресничку. Две центральные микротрубочки возникают в связи с плотным веществом, лежащим на границе бывшей центриоли и выроста плазматической мембраны (рис. 290а).
При образовании многоресничных клеток происходит многочисленная репликация центриолей и образование многочисленных ресничек.
В ресничном эпителии позвоночных множественные базальные тельца возникают вокруг так называемых дейтеросом – аморфных электронноплотных структур размером от 60 до 700 нм, по периферии которых происходит закладка множественных зачатков базальных телец. Вокруг одной дейтеросомы образуются до десятка новых базальных телец. Они затем мигрируют к плазматической мембране и принимают участие в образовании аксонем (рис. 298).
Необходимо отметить, что клетки с множеством ресничек теряют способность к делению и не могут выходить из G0-стадии клеточного цикла. На смену им из эпителиального пласта приходят стволовые недифференцированные клетки, которые могут делиться и давать новые поколения многоресничных клеток.
Микротрубочки аксонемы устойчивы к действию колхицина, но при росте реснички колхицин полностью прекращает включение новых молекул тубулина, что приводит к торможению роста ресничек.
Вторая категория ресничных клеток – клетки с так называемыми первичными ресничками, не обладающими способностью к движению. Практически все типы клеток, за исключением клеток крови, мышц и кишечного эпителия, в G0-периоде образуют первичные реснички, которые отличаются от настоящих ресничек, или киноцилий, тем, что они не имеют пары центральных микротрубочек и не способны к движению. Они образуются в результате того, что диплосома подходит к плазматической мембране и от материнской центриоли начинается рост аксонемы, но без двух центральных микротрубочек. Если клетки культуры фибробластов, обладающих в G0-периоде такими ресничками, стимулировать к делению, то эти реснички исчезают, а базальное тельце-центриоль начинает свой цикл как обычная центриоль в клетках, способных к делению.
Функциональное значение этих первичных ресничек не ясно. Но интересно отметить, что при развитии сенсорных клеток сетчатки их наружные сегменты палочек и колбочек возникают сначала за счет образования первичных ресничек. Возможно, что у нерецепторных клеток, имеющих такие первичные реснички, последние выполняют функции внешних анализаторов, являются как бы «антеннами», на поверхности которых рецепторные молекулы плазматической мембраны могут регистрировать механические и химические сигналы, поступающие из внешней межклеточной среды.
studfiles.net
Глава 23. Клеточный центр
Итак, в клетках животных, растений и одноклеточных микротрубочки поляризованы, так что большей частью их растущие (+)-концы направлены к периферии клетки. Это связано с тем, что МТ начинают свой рост от специальных участков в клетке, от центров организации микротрубочек (ЦОМТ). Некоторые из ЦОМТ имеют сложную морфологическую организацию, другие устроены иначе. Различные ЦОМТ можно разделить на несколько групп: центросомные клеточные центры, и центры организации микротрубочек, не имеющие четкой локализации.
Так например, в клетках высших растений полимеризация МТ происходит по периферии клеточного ядра, от которого МТ расходятся радиально. Сходная картина наблюдается при регенерации МТ в гигантских клетках слюнных желез двукрылых. В ряде случаев новообразование МТ, их закладка, нуклеация, может происходить в цитоплазме вне связи со специальными зонами или структурами.
Но в большинстве случаев в интерфазных клетках животных организмов образование и рост МТ происходит от клеточного центра, содержащего специальные образования – центросомы, которые большей частью могут содержать сложно организованные центриоли, или же не иметь их.
Центросомы и центриоли
Центросомы были обнаружены и описаны сто лет назад (Флемминг, 1875; Бенеден, 1876) – это очень мелкие тельца, размер которых находится на границе разрешающей способности светового микроскопа, обычно располагающиеся в геометрическом центре клетки, откуда и их название. В некоторых объектах удавалось видеть, что мелкие плотные тельца (центриоли), обычно в паре (диплосома), окружены зоной более светлой цитоплазмы (собственно центросома), от которой отходят радиально тонкие фибриллы (центросфера) (рис. 277).
Центросомы характерны и обязательны для клеток животных, и нет у высших растений, у низших грибов и некоторых простейших. Было замечено, что центросомы в делящихся клетках принимают участие в формировании веретена деления и располагаются на его полюсах. В неделящихся клетках центросомы часть определяют полярность клеток эпителия и располагаются вблизи аппарата Гольджи. Такая связь центросом с аппаратом Гольджи характерна для многих клеток, в том числе для клеток крови и нервных клеток. Часто центросомы лежат рядом с ядром, располагаясь в зонах его впячивания. Например, в полиморфных лейкоцитах (нейрофилы) центросома лежит внутри подкововидного впячивания ядра (рис. 278).
Типичное строение клеточный центр имеет в клетках животных. Он представляет собой зону, состоящую из центриолей и окружающей их аморфной фибриллярной массы или матрикса. В ряде случаев в состав клеточного центра или центросомы входит только эта фибриллярная масса, от которой отходят микротрубочки (см. ниже).
Наиболее же часто кроме матрикса в состав клеточного центра входят центриоли, как мелкие тельца, с трудом наблюдаемые в световом микроскопе.
Тонкое строение центриолей удалось изучить только с помощью электронного микроскопа. Основу строения центриолей составляют расположенные по окружности девять триплетов микротрубочек, образующие таким образом полый цилиндр (рис. 279). Его ширина около 0, 15 мкм, а длина такого цилиндра 0,3-0,5 мкм (хотя встречаются центриоли, достигающие в длину несколько микрон) (рис. 280).
Первая микротрубочка триплета (А-микротрубочка) имеет диаметр около 25 нм и толщину стенки 5 нм, которая состоит из 13 глобулярных субъединиц. Длина каждого триплета равна длине центриоли. Вторая и третья (В и С) микротрубочки отличаются от А-микротрубочки тем, что они являются неполными, содержат 11 субъединиц и вплотную примыкают к своим соседям. Каждый триплет располагается к радиусу такого цилиндра под углом около 400. Кроме микротрубочек в состав центриоли входит ряд дополнительных структур. От А-микротрубочки отходят так называемые “ручки”, выросты, один из которых (внешний) направлен к С- микротрубочке соседнего триплета, а другой (внутренний) – к центру цилиндра.
Обычно в интерфазных клетках всегда присутствуют две центриоли, располагающиеся рядом друг с другом, образуя дуплет центриолей, или диплосому (рис. 281). В диплосоме центриоли располагаются под прямым углом по отношению друг к другу. Из двух центриолей различают “материнскую” и “дочернюю”, продольная ось последней перпендикулярна продольной оси материнской центриоли. Обе центриоли сближены своими концами так, что проксимальный конец дочерней центриоли как бы смотрит на поверхность материнской. В дистальном участке материнской центриоли располагается аморфный материал в виде выростов или шпор – это придатки. Их нет на дочерней центриоли (рис. ).
Дочерняя центриоль несколько отличается от материнской. Центральная часть цилиндра центриоли занята структурой, напоминающей тележное колесо; она имеет центральную “втулку” диаметром около 25 нм и 9 спиц, направленных по одной к А-микротрубочке каждого из триплетов. Такие структуры внутри центриоли расположены в одном из её концов, проксимальном, что делает строение цилиндра центриоли полярным. На дистальном конце центриоли внутри её нет таких структур. Объем, занимаемый внутри центриоли втулкой со спицами, может составлять у разных клеток от 3\4 до 1\5 длины центриоли. У некоторых видов втулка отсутствует или заменена скоплением аморфного материала. Торцы центриолярного цилиндра, кроме системы втулки и спиц на проксимальном конце, ничем не закрыты.
Систему микротрубочек центриоли обычно описывают формулой 9 + 0, или (9х3) + 0, подчеркивая отсутствие микротрубочек в её центральной части.
Вокруг каждой центриоли расположен бесструктурный, или тонковолокнистый матрикс. Сами микротрубочки триплетов погружены в аморфный материал т.н. муфты или оправы. Если выделенные центриоли обработать 0,6М раствором NaCl, то произойдет полная экстракция микротрубочек, но центриоль как таковая не растворится: вместо нее останется цилиндрическая структура, имеющая девять полых отверстий, некогда занимавшихся триплетами микротрубочек. Поэтому все схемы центриолей в этой книге, как и во многих других значительно упрощены и не включают материал муфты центриолярного цилиндра.
Часто около центриолей и в связи с ним можно обнаружить несколько дополнительных структур: сателлиты, фокусы схождения микротрубочек, исчерченные волокнистые корешки, дополнительные микротрубочки, образующие особую зону – центросферу вокруг центриоли (рис. 282).
При исследовании в электронном микроскопе интерфазных центриолей было найдено, что лучистое сияние центросферы, обнаруживаемое в световом микроскопе, представляет собой большое число микротрубочек, радиально расходящихся от зоны диплосомы. В диплосоме лишь одна из центриолей, материнская, содержит ряд дополнительных структур. Одни из них, перицентриолярные сателлиты, состоят из имеющей тонкое фибриллярное строение конусовидной ножки, расположенной на стенке центриоли, и головки, заканчивающейся на этой ножке. Ножки сателлитов часто имеют поперечную исчерченность (рис. 284). Количество таких перицентриолярных сателлитов непостоянно, они могут располагаться на разных уровнях по длине центриоли. Кроме этих структур рядом с диплосомой, но не связанные с ней структурно, могут располагаться плотные мелкие (20-40 нм) тельца, к которым подходят одна или несколько микротрубочек (фокусы схождения микротрубочек). Микротрубочки отходят и от головок сателлитов. Эти центросомные микротрубочки не отходят непосредственно от микротрубочек цилиндров центриолей, а связаны или с сателлитами, или с матриксом. Такие микротрубочки и образуют как бы лучистую сферу (центросферу) вокруг центриоли, где (-)-концы МТ связаны с ЦОМТ, а (+)-концы радиально расходятся на периферию клетки. При образовании центросферы в интерфазной клетке только специальные структуры центриоли, сателлиты и матрикс, каким-то образом связаны с образованием микротрубочек; микротрубочки самих центриолей в этом процессе не участвуют. Было найдено, что восстановление прицентриолярных микротрубочек после их деполимеризации на холоду происходит за счет появления новых микротрубочек, отходящих от головок сателлитов. Таким образом, можно считать, что эти дополнительные структуры являются центрами, на которых происходит сборка микротрубочек из тубулинов ( центры организации микротрубочек – ЦОМТ).
Химия центриолей изучена слабо, потому что еще не разработаны методы получения этой структуры в виде чистой фракции. Трудности биохимического изучения центриолей связаны с тем, что это одиночная клеточная структура, имеющая объем всего 0, 03 мкм3. Для сравнения, вспомним, что в клетке имеется: около тысячи штук митохондрий, около миллиона рибосом, около сотни хромосом, около 1 мм2 мембран.
Есть все основания говорить о том, что в состав микротрубочек центриолей входят тубулины. Это доказывается тем, что колхицин прекращает рост микротрубочек в процентриолях, возникающих вблизи материнской центриоли. Предположения о возможной химической природе остальных элементов центриоли основаны главным образом на данных, полученных из химии ресничек и жгутиков, имеющих много сходных черт строения с центриолями.
Данные о химическом строении центриолей получены главным образом с помощью иммунохимических методов. В изолированных базальных тельцах простейшего хламидомонады обнаружено более 200 различных белков, среди которых выявлены четыре вида тубулинов, в том числе - тубулин, центрин, перицентрин, белок р210 и многие другие.
В интерфазных клетках центриоли оказываются связаны с ядром и с ядерной мембраной. При выделении ядер практически все центриоли клеток печени и селезенки крыс оказываются в этой фракции. Связь центриолей с ядром осуществляется главным образом промежуточными филаментами. Если живые клетки подвергнуть ультрацентрифугированию, то центриоли опускаются к центробежному полюсу вместе с ядрами.
Центросомный цикл
Было обнаружено, что строение и активность центросом меняются в зависимости от периода клеточного цикла, в течение которого клеточный центр претерпевает тоже циклические изменения (рис. 283).
Целесообразнее начать рассмотрение циклических изменений в структуре центросом с митоза. Начиная с профазы и кончая телофазой, центросомы имеют сходное строение, несмотря на то, что за время митоза происходит ряд существенных клеточных перестроек: конденсация хромосом, разрушение ядерной оболочки, образование веретена деления, расхождение хромосом. В митозе в клеточных центрах (их два, по одному на каждый полюс клетки) находится по диплосоме. Как полагается, дочерняя центриоль своим концом направлена на материнскую. Материнская центриоль на всех стадиях митоза окружена довольно широкой (до 0,3 мкм) зоной тонких фибрилл – центриолярное фибриллярное гало (рис. 279). От этого гало радиально отходят микротрубочки. Важно подчеркнуть, что у дочерних центриолей ни гало, ни отходящих от центриолей микротрубочек нет. В это время происходит формирование веретена митотического аппарата, состоящего из микротрубочек. Эта структура действительно имеет форму веретена, где на концах его на полюсах клетки располагаются диплосомы, окруженные радиальными микротрубочками (центросфера). В данном случае можно говорить о том, что зоны диплосом, клеточные центры, являются центрами организации (полимеризации) микротрубочек. В пользу этого говорят следующие факты: после исчезновения микротрубочек веретена и центросферы, которые происходят при действии холода или колхицина, новые микротрубочки возникают главным образом в районе материнских центриолей, диплосом, в каждом из полюсов клетки. Интересно, что рост новых микротрубочек не связан с микротрубочками триплетов центриолярного цилиндра, они начинают отрастать от зоны гало, расположенной на материнской центриоли. Важно отметить, что в это время на материнских центриолях (как и на дочерних) нет сателлитов, и в это же время цитоплазма теряет микротрубочки: микротрубочки цитоплазмы разбираются, а пул освободившихся тубулиновых мономеров идет на образование микротрубочек веретена и центросферы, которые образуются на фибриллярном гало, как на ЦОМТ. Этот процесс полимеризации митотических микротрубочек отражает первую форму активности центриолярного аппарата (рис. ). Если в профазе облучить центриоль лазерным микролучем, то образование веретена останавливается.
Примерно сходное строение имеют клеточные центры на всех стадиях митоза, но к телофазе толщина фибриллярного гало уменьшается.
К концу телофазы, когда произошло разделение клетки надвое, а хромосомы начали деконденсироваться и образовывать новые интерфазные ядра, происходит разрушение веретена деления, его микротрубочки деполимеризуются. Клеточные центры при этом меняют свою структуру. Материнская и дочерняя центриоли теряют взаимно перпендикулярное расположение и отходят друг от друга на небольшие (0,5-2мкм) расстояния, но все же держатся в одном месте. Вокруг материнской центриоли гало и микротрубочки не выявляются. В это время микротрубочек в цитоплазме также практически нет.
В начале G1-периода на поверхности материнской центриоли возникают сателлиты, имеющие ножку и головку, от которой радиально отходят микротрубочки, которые начинают расти в длину и заполнять собой цитоплазму (рис. 284а). Следовательно, вторая форма активности клеточного центра – образование цитоплазматических микротрубочек в интерфазных клетках. Надо подчеркнуть, что активной здесь является только материнская центриоль, которую легко узнать по придаткам в ее дистальной части.
Если считать клеточные центры основными (если не единственными) местами образования цитоплазматических микротрубочек, то общее количество последних должно быть равно числу микротрубочек, отходящих от центриолей. При исследовании в электронном микроскопе оказалось, что от клеточных центров в интерфазе отходит всего лишь несколько десятков микротрубочек, а в цитоплазме их так много, что с помощью иммунофлуоресцентного метода их трудно подсчитать. Это дает основание предполагать, что по мере роста микротрубочек часть из них теряет связь с областью центриолей и может находиться в цитоплазме долгое время. Центросомы же индуцируют полимеризацию новых микротрубочек, которые приходят на смену постепенно деполимеризующимся старым. Вероятно, в цитоплазме есть несколько генераций микротрубочек: “старые”, не связанные с клеточным центром, и новые, растущие от центросом. Таким образом, в клетке происходит как бы конвейерная смена и репродукция цитоплазматических микротрубочек.
Если клеткам запретить переходить в S-период, они могут существовать в фазе клеточного покоя (Go-период) (рис. 285). В это время материнская центриоль продолжает функционировать, как центр образования микротрубочек цитоскелета. Но одновременно она может проявить еще одну форму активности – образовать ресничку, вырост плазматической мембраны, заполненный аксонемой (осевой нитью), состоящей из девяти дублетов микротрубочек. Эти микротрубочки отрастают, как от затравок, от А- и В-микротрубочек триплетов материнской центриоли в дистальной ее части. Это – третья форма активности центриолей как центров организации микротрубочек (см. ниже).
При наступлении S-периода (или в середине его) клеточный центр приступает к четвертой форме своей активности: происходит удвоение числа центриолей. В это время около каждой из разошедшихся еще в конце телофазы центриолей, материнской и дочерней, происходит закладка новых центриолярных цилиндров – процентриолей (рис. 284б). В районе проксимальных концов каждой центриоли перпендикулярно длинной оси закладывается сначала девять синглетов (одиночных) микротрубочек, затем они преобразуются в девять дуплетов, а потом – в девять триплетов растущих микротрубочек новых центриолярных цилиндров.
Закладка процентриолей происходит на проксимальных концах центриолей; в этом месте растут новые поколения центриолей, тоже с проксимального конца. Во время роста процентриолей здесь можно видеть центральную “втулку” со спицами.
Благодаря такому росту структур образуется сначала короткая дочерняя центриоль – процентриоль — которая затем дорастает до размера материнской. Этот способ увеличения числа центриолей был назван дупликацией. Важно отметить, что размножение центриолей не связано с их делением, почкованием или фрагментацией, а происходит путем образования зачатка, процентриоли, вблизи и перпендикулярно к исходной центриоли. Правда, последнее условие соблюдается не во всех объектах, у некоторых оомицетов при дупликации центриоли происходит сначала расхождение центриолей, рост втулки, затем рост микротрубочек вдоль продолжения оси исходной центриоли, и центриоли располагаются конец в конец. Интересно, что триплеты в таких новых центриолях имеют угол наклона, противоположный таковому в материнской центриоли.
Факт удвоения центриолей привел некоторых исследователей к предположению, что центриоли, так же как митохондрии и пластиды, принадлежат к саморедуплицирующимся компонентам цитоплазмы, хотя прямых данных о наличии ДНК в составе центриолей нет.
В S-периоде во время удвоения (дупликации) центриолей материнская продолжает проявлять вторую форму активности: она продолжает быть центром образования цитоплазматических микротрубочек.
В результате процесса дупликации около каждой центриоли вырастает новая дочерняя центриоль (первая материнская центриоль и дочерняя на бывшей дочерней центриоли могут считаться как бы бабушкой и внучкой). Поэтому в клетке после завершения S-периода находятся уже две диплосомы (а всего четыре центриолярных цилиндра) (рис. 286).
После этого наступает следующий период клеточного цикла, постсинтетический (G2-период), когда в клетке начинается подготовка к очередному делению. В это время исчезают сателлиты на материнской диплосоме (так можно назвать старую материнскую центриоль с новой дочерней), а обе материнские центриоли в обеих диплосомах покрываются фибриллярным гало, от которого в профазе начинают отрастать митотические микротрубочки. Параллельно этому в цитоплазме происходит исчезновение микротрубочек, и клетка стремится приобрести шаровидную форму. Вся такая последовательность событий повторяется от цикла к циклу у клеток, способных к длительному размножению. В большинстве случаев клетки организма находятся в G0-периоде, поэтому у них центриоль участвует в полимеризации цитоплазматических микротрубочек и в образовании реснички (или множества ресничек). В последнем случае она входит в состав так называемого базального тельца.
Обычно в клетку после деления попадают два центриолярных цилиндра в составе диплосомы. В различных экспериментальных условиях можно запретить разделение клетки надвое и получить клетки с удвоенным числом хромосом (полиплоидные клетки). Совершенно очевидно, что в таких клетках будет и удвоенное число центриолей. Клетки могут снова вступать в клеточный цикл, при этом будет удваиваться как количество ДНК, так и число центриолей. Было обнаружено, что у тетраплоидных (с четырехкратным набором хромосом) клеток печени в G0-периоде в цитоплазме видны не два, а четыре центриолярных цилиндра, а в полюсах при делении таких клеток было обнаружено по две диплосомы в каждом. Аналогичная ситуация замечена и у других полиплоидных клеток (мегакариоциты костного мозга, полиплоидные гибридные клетки и др.). В связи с этим предположили, что между числом плоидности клетки (числом хромосомных наборов) и числом центриолей существует прямая связь.
Нарушения центриолярного цикла могут вызвать ряд патологических изменений клеток, в первую очередь появление многополюсных митозов. Так, при действии β-меркаптоэтанола происходит блокада нормального митоза, при этом диплосомы расходятся на отдельные центриоли. При отмывании от этого вещества клетка снова приступает к делению, но в этом случае каждая центриоль активируется и образует полюс веретена. Таким образом, возникают трех- или четырехполюсные митозы, приводящие к неравномерному распределению хромосом между дочерними клетками. Это в свою очередь приводит к изменению числа хромосом (анэуплоидия), которое часто вызывает гибель клетки. Иногда при образовании многополюсных митозов в некоторых полюсах отсутствуют центриоли: в полюсе располагается только фибриллярный материал центросомы (бесцентриолярные полюса).
Итак, в подавляющем большинстве клеток млекопитающих центросомы участвуют в полимеризации тубулинов и являются структурами, играющими роль центров организации микротрубочек. Микротрубочки самих центриолей являются затравками для полимеризации тубулинов только в одном случае – при росте аксонемы реснички, когда центриоль становится базальным тельцем. Это временное состояние: при переходе клеток к делению реснички могут исчезать, а базальное тельце снова может выполнять роль центриоли, участвуя в организации цитоплазматических микротрубочек или микротрубочек митотического веретена. Только в этих случаях центрами организации микротрубочек являются не сами центриолярные цилиндры, а перицентриолярный материал (головка сателлитов, околоцентриолярный матрикс, гало и т.д.). Следовательно, центриоль как таковую нужно рассматривать как один из компонентов более сложной структуры – клеточного центра или центросомы. Эта оговорка связана с тем, что у всех высших растений ЦОМТ не содержит центриолей. Более того, в раннем эмбриогенезе позвоночных животных образуются веретена деления, не имеющие центриолей в полюсах. По всей вероятности в последних случаях центриоли возникают позже заново, а не образуются путем “репликации”. Вопрос о процессе образования центриолей далек от решения. Остается неясным процесс появления процентриолей. В процессе эмбриогенеза отмечены случаи возникновения центриолей de novo у морского ежа, у моллюсков, у мышей. Так, в эмбриогенезе мыши центриоли появляются только после 1-2 делений клеток бластулы, несмотря на то, что сами клеточные деления происходят нормально, за исключением того, что в полюсах деления в зоне бесструктурной центросомы центриоли отсутствуют. С другой стороны, если в соматических клетках культуры ткани уничтожить центросому с центриолью с помощью микрооблучения, то новые центриоли не возникают.
Базальные тельца. Строение и движение ресничек и жгутиков.
Как уже указывалось, у многих клеток животных, вышедших из клеточного цикла, в G0-стадии центриоли принимают участие в образовании аппарата движения – ресничек. Их две группы: кинетоцилии, характерные для специальных эпителиев (ресничные эпителии трахеи, яйцеводов) или свободно плавающих клеток (сперматозоиды, простейшие), и так называемые первичные реснички, встречающиеся во многих клетках, не обладающих способностью к движению.
Вначале рассмотрим строение кинетоцилей – подвижных ресничек и жгутиков. В световом микроскопе эти структуры видны как тонкие выросты клетки, в их основании в цитоплазме видны хорошо красящиеся мелкие гранулы – базальные тельца, аналоги центриолей (рис. 287). Клетки, имеющие реснички или жгутики, обладают способностью двигаться, будучи в свободном состоянии, или же перемещать жидкости в случае, если клетки неподвижны. Свободноживущие одноклеточные организмы, снабженные одним или несколькими жгутиками, обычно движутся тем концом вперед, который несет жгутики. Иной способ движения можно видеть у спермиев некоторых животных: жгутик, располагаясь сзади, толкает тело клетки вперед. Скорость движения клеток за счет работы жгутиков может достигать очень большой величины (до 5 мм / мин).
Множественные реснички также могут обеспечивать движение свободноживущих клеток, таких как инфузории или некоторые жгутиконосцы. Реснички эпителиальных клеток многих беспозвоночных и позвоночных животных обеспечивают поток жидкостей вдоль поверхности таких клеток. Число ресничек на клетку может достигать 300 в эпителии трахеи; у инфузории туфельки на клетку приходится 10-14 тыс. рядами расположенных ресничек.
При движении ресничек и жгутиков не происходит уменьшения их длины, поэтому неправильно называть это движение сокращением. Траектория движения ресничек очень разнообразна (рис. 288). В различных клетках это движение может быть маятникообразным, крючкообразным, воронкообразным или волнообразным.
У многоресничных клеток (инфузории, клетки ресничного эпителия) движение ресничек не хаотично, а строго упорядочено. В этом случае реснички расположены рядами. В продольном ряду отдельные реснички начинают движение и проходят отдельные его фазы по очереди, метахронно. В поперечном же ряду все реснички находятся в одной фазе движения (синхронны). Это создает движущую волну по поверхности клетки (рис. )289.
Общая архитектура реснички представлена на рис. 290, 291. Ресничка представляет собой тонкий цилиндрический вырост цитоплазмы с постоянным диаметром 300 нм. Этот вырост от основания до самой его верхушки покрыт плазматической мембраной. Внутри выроста расположена аксонема, сложная структура, состоящая в основном из микротрубочек. Нижняя, проксимальная часть реснички, базальное тельце, погружена в цитоплазму. Диаметры аксонемы и базального тельца одинаковы (около 200 нм).
На поперечном сечении реснички видна плазматическая мембрана, окружающая аксонему. Аксонема в своем составе имеет девять дублетов микротрубочек, образующих внешнюю стенку цилиндра аксонемы. Дублеты микротрубочек слегка повернуты (около 100) по отношению к радиусу аксонемы. Кроме периферических дублетов микротрубочек в центре аксонемы располагается пара центральных микротрубочек. В целом систему микротрубочек реснички описывают как (9х2)+2. В дублетах микротрубочек также различают А-микротрубочку, состоящую из 13 субъединиц, и В-микротрубочку, неполную, содержащую 11 субъединиц. А-микротрубочка несет на себе ручки, которые направлены к В-микротрубочке соседнего дуплета. От А-микротрубочки к центру аксонемы отходит радиальная связка, или спица, оканчивающаяся головкой, присоединяющейся к центральной муфте, имеющей диаметр около 70 нм, окружающей две центральные микротрубочки. Последние лежат отдельно друг от друга на расстоянии около 25 нм. Таким образом, в аксонеме располагается 20 продольных микротрубочек, в то время как в базальном тельце их 27 (рис. 291, 292).
Базальное тельце состоит из 9 триплетов микротрубочек (как и центриоль), имеет ручки, втулку и спицы, расположенные в проксимальной (нижней) ее части. На участке базального тельца, примыкающем к плазматической мембране, есть девять придатков, выступов, идущих от каждого триплета микротрубочек к плазматической мембране и связывающих его с клеточной поверхностью.
Базальное тельце и аксонема структурно связаны друг с другом и составляют единое целое: А- и В-микротрубочки триплетов базального тельца продолжаются в А- и В-микротрубочках дуплетов аксонемы. Однако внутренние части аксонемы и базального тельца значительно отличны друг от друга. Часто в зоне перехода базального тела в аксонему наблюдают аморфную поперечную пластинку, которая как бы отделяет эти две части. Центральные микротрубочки аксонемы начинаются от этой пластинки так же, как в этом месте начинается и центральная муфта (капсула) (рис. 290).
В основании ресничек и жгутиков часто встречаются исчерченные корешки, или кинетодесмы, представляющие собой пучки тонких (6 нм) фибрилл, обладающих поперечной исчерченностью (рис. 293). Часто такие исчерченные кинетодесмы простираются от базальных телец вглубь цитоплазмы по направлению к ядру. Роль этих структур не ясна. Они не изменяются при действии колхицина, могут встречаться и в составе центриолей интерфазных клеток, не принимающих участия в образовании ресничек.
При движении ресничек не происходит изменения их длины, они не “сокращаются”, а изгибаются, бьются. Оказалось, что механически отделенные реснички способны к биению в присутствии АТФ. При отделении ресничек базальные тельца остаются в теле клетки. Это означает, что для механической работы ресничек базальное тело не нужно, а только аксонема участвует в генерации движения. Удалось показать, что за движение ресничек отвечают “ручки”, сидящие на А-микротрубочках. При экстракции компонентов ручек реснички перестают биться в присутствии АТФ.
Было найдено, что в состав ручек входят белки динеины. Это большие белковые компоненты, состоящие из 9-12 полипептидных цепей, содержащие 2-3 глобулярные головки, связанные в общий корешок гибкими хвостами (рис. 294). Каждая головка динеина обладает АТФ-азной активностью, которая возрастает примерно в 6 раз при ассоциации с микротрубочками. В состав каждой ручки входит один белковый комплекс, одна молекула динеина. Так как экстракция ручек прекращает биение ресничек, то можно считать, что именно динеин ответственен за это движение, то есть динеин является мотором или двигателем при биении ресничек. Но каков механизм этого движения?
Этот вопрос был решен при использовании выделенных ресничек, лишенных плазматической мембраны, радиальных спиц и связок после частичной обработки аксонем протеазами. Оказалось, что такие аксонемы, содержащие динеиновые ручки, при добавлении к ним АТФ начинают увеличиваться в длину почти до девяти раз и одновременно утончаются. В электронном микроскопе видно, что такая аксонема увеличилась в длину за счет смещения пар микротрубочек одна относительно другой (рис. 295). Другими словами, произошло продольное скольжение дуплетов один относительно другого, аналогично тому, что происходит при сокращении саркомеров в мышце: скольжение миозиновых нитей относительно актиновых. В случае динеина повторные циклы ассоциации с субъединицами тубулина, изменения конформации при связывании АТФ и его гидролизе, вызывают перемещение головок вдоль микротрубочки от (+)-конца к (-)-концу. При этом соседний дуплет двигается к верхушке реснички. Когда ресничка содержит все компоненты, и дуплеты микротрубочек связаны друг с другом и с центральной парой микротрубочек, такие кооперативные смещения дуплетов микротрубочек приводят не к удлинению реснички, а к ее изгибу (рис. 296). Как регулируется последовательное перемещение дуплетов один относительно другого, еще не ясно.
Рост ресничек, удлинение микротрубочек их аксонем происходит на вершине реснички. Следовательно, там локализованы (+)-концы микротрубочек.
Образование аксонемы ресничек происходит за счет роста А- и В-микротрубочек центриолей, которые в этом случае становятся базальным тельцем. В простейшем случае при образовании одиночных ресничек или так называемых первичных ресничек материнская центриоль подходит к плазматической мембране своим дистальным торцом, связывается с ней своими придатками. В это время начинается рост микротрубочек на (+)- концах А- и В-микротрубочек триплетов. Возникают девять дублетов микротрубочек аксонемы, которые, наращиваясь с (+)-концов на верхушке аксонемы как бы вытягивают плазматическую мембрану, образуя вырост – ресничку. Две центральные микротрубочки возникают в связи с плотным веществом, лежащим на границе бывшей центриоли и выроста плазматической мембраны (рис. 290а).
При образовании многоресничных клеток происходит многочисленная репликация центриолей и образование многочисленных ресничек.
В ресничном эпителии позвоночных множественные базальные тельца возникают вокруг так называемых дейтеросом – аморфных электронноплотных структур размером от 60 до 700 нм, по периферии которых происходит закладка множественных зачатков базальных телец. Вокруг одной дейтеросомы образуются до десятка новых базальных телец. Они затем мигрируют к плазматической мембране и принимают участие в образовании аксонем (рис. 298).
Необходимо отметить, что клетки с множеством ресничек теряют способность к делению и не могут выходить из G0-стадии клеточного цикла. На смену им из эпителиального пласта приходят стволовые недифференцированные клетки, которые могут делиться и давать новые поколения многоресничных клеток.
Микротрубочки аксонемы устойчивы к действию колхицина, но при росте реснички колхицин полностью прекращает включение новых молекул тубулина, что приводит к торможению роста ресничек.
Вторая категория ресничных клеток – клетки с так называемыми первичными ресничками, не обладающими способностью к движению. Практически все типы клеток, за исключением клеток крови, мышц и кишечного эпителия, в G0-периоде образуют первичные реснички, которые отличаются от настоящих ресничек, или киноцилий, тем, что они не имеют пары центральных микротрубочек и не способны к движению. Они образуются в результате того, что диплосома подходит к плазматической мембране и от материнской центриоли начинается рост аксонемы, но без двух центральных микротрубочек. Если клетки культуры фибробластов, обладающих в G0-периоде такими ресничками, стимулировать к делению, то эти реснички исчезают, а базальное тельце-центриоль начинает свой цикл как обычная центриоль в клетках, способных к делению.
Функциональное значение этих первичных ресничек не ясно. Но интересно отметить, что при развитии сенсорных клеток сетчатки их наружные сегменты палочек и колбочек возникают сначала за счет образования первичных ресничек. Возможно, что у нерецепторных клеток, имеющих такие первичные реснички, последние выполняют функции внешних анализаторов, являются как бы «антеннами», на поверхности которых рецепторные молекулы плазматической мембраны могут регистрировать механические и химические сигналы, поступающие из внешней межклеточной среды.
studfiles.net
Клеточный центр Википедия
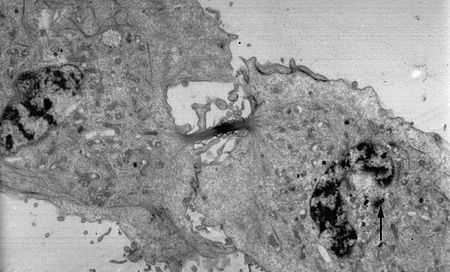 Телофаза митоза (электронная микрофотография). Стрелка указывает на центросому. Четко видны две центриоли, расположенные под прямым углом друг к другу: одна перерезана поперек, другая вдоль.
Телофаза митоза (электронная микрофотография). Стрелка указывает на центросому. Четко видны две центриоли, расположенные под прямым углом друг к другу: одна перерезана поперек, другая вдоль.Клеточный центр, или центросома (от др.-греч. σῶμα — тело) — немембранный органоид в клетках эукариот, состоит из двух центриолей и перицентриолярного материала. Является главным центром организации микротрубочек (ЦОМТ) эукариотической клетки, играет важнейшую роль в клеточном делении, участвуя в формировании веретена деления. Из центросомы образуются реснички и жгутики. Центросомы характерны для клеток животных, их нет у высших растений, у высших грибов, у некоторых простейших[1][2]. Ряд наследственных заболеваний человека вызван мутациями в генах, ирующих центросомные белки[3].
История открытия и изменений наименований[ | ]
Центросомы на полюсах веретена деления в делящихся клетках практически одновременно описали B. Флемминг, O. Гертвиг и Э. ван Бенеден в середине 70-х годов XIX в. Этим структурам было дано название «центросфера», а гранулы, которые удавалось разглядеть в фокусе центросферы, получили название «полярные корпускулы». Эдвард ван Бенеден и Теодор Бовери независимо друг от друга смогли показать, что центросферы не исчезают по завершению митоза, а сохраняются в интерфазной клетке, часто располагаясь в районе геометрического центра клетки. В 1887 году Эдвард ван Бенеден предложил переименовать центросферы в «центральные корпускулы» или «центральные тельца». В 1888 году Теодор Бовери предложил для этой структуры название «центросома», а позднее в 1895 году — «центриоль». Следует отметить, что многочисленные названия одной структуры породили терминологическую путаницу, а термины «центросома» и «центриоль» до изобретения электронной микроскопии использовали как синонимы[4]. С середины 50-х годов XX века, когда благодаря электронной микроскопии была изучена тонкая структура этой органеллы, название «центриоль» стали ассоциировать с центриолярными цилиндрами. В 1984 году американcкий клеточный биолог Даниэль Мезиа предложил использовать термин «центросома» для обозначения совокупности центриолей и окружающих их структур[5][6].
Строение центросомы[ | ]
У многих живых организмов (животных и ряда простейших) центросома содержит пару центриолей, цилиндрических структур, расположенных под прямым углом друг к другу. Каждая центриоль образована девятью триплетами микротрубочек, расположенными по кругу, а т
ru-wiki.ru
Клеточный центр — это… Что такое Клеточный центр?
Телофаза митоза (электронная микрофотография). Стрелка указывает на центросому. Четко видны две центриоли, расположенные под прямым углом друг к другу: одна перерезана поперек, другая вдоль.Центросома (от центр и греч. soma — тело), центросфера, центроплазма, участок цитоплазмы клетки, окружающий центриоли. В Центросоме отсутствуют клеточные органоиды. Она плотнее остальной части цитоплазмы, её можно растягивать и передвигать при помощи микрургических операций. В старой литературе термин «Центросома» часто употреблялся как синоним центриолей.
Центросома или клеточный центр — главный центр организации микротрубочек (ЦОМТ) и регулятор хода клеточного цикла в клетках эукариот. Впервые обнаружена в 1888 г. Теодором Бовери, который назвал её «особым органом клеточного деления». Хотя центросома играет важнейшую роль в клеточном делении, недавно было показано, что она не является необходимой. В подавляющем большинстве случаев в клетке в норме присутствует только одна центросома. Аномальное увеличение числа центросом характерно для раковых клеток. Более одной центросомы в норме характерно для некоторых полиэнергидных простейших и для синцитиальных структур.
У многих живых организмов (животных и ряда простейших) центросома содержит пару центриолей, цилиндрических структур, расположенных под прямым углом друг к другу. Каждая центриоль образована девятью триплетами микротрубочек, расположенными по кругу, а также ряда структур, образованных центрином, ценексином и тектином.
В интерфазе клеточного цикла центросомы ассоциированы с ядерной мембраной. В профазе митоза ядерная мембрана разрушается, центросома делится, и продукты ее деления (дочерние центросомы) мигрируют к полюсам делящегося ядра. Микротрубочки, растущие из дочерних центросом, крепятся другим концом к так называемым кинетохорам на центромерах хромосом, формируя веретено деления. По завершении деления в каждой из дочерних клеток оказывается только по одной центросоме.
Помимо участия в делении ядра, центросома играет важную роль в формировании жгутиков и ресничек. Центриоли, расположенные в ней, выполняют функцию центров организации для микротрубочек аксонем жгутиков. У организмов, лишенных центриолей (например, у сумчатых и базидиевых грибов, покрытосеменных растений), жгутики не развиваются.
Шесть стадий клеточного деления
На всех стадиях кариокинеза важнейшую роль играют Микротрубочки (далее МТ) – их образование и пространственная ориентация, взаимодействие с кинетохорами хромосом, структурные изменения, создающие силы, необходимые для разделения хромосом, и, наконец, их разрушение. МТ входят в состав цитоскелета и играют важнейшую роль в поддержании и изменении формы клетки и направленном переносе внутриклеточных компонентов (везикул, органелл, белков и т.п.) в цитоплазме. В клетках животных несколько тысяч МТ. Все они растут из специальных образований, называемых центрами организации МТ (ЦОМТ). В клетке может быть 1–2 ЦОМТ. Исследования показали, что от центросомы отходят всего несколько десятков МТ, следовательно, МТ не обязательно связаны с центросомой. Центриоли же дают начало новым МТ, которые приходят на смену постепенно деполимеризующимся старым. МТ представляет собой очень маленькую трубочку длиной несколько микрометров при наружном диаметре 25 нм. Она построена из 13 длинных «палочек» – протофиламентов, параллельных оси трубочки и расположенных по кругу. Протофиламент составлен из чередующихся глобул альфа- и бета-тубулина, причем в каждой паре таких глобул (димере тубулина) альфа-тубулин взаимодействует с бета-тубулином, а бета-тубулин – с альфа-тубулином ближайших соседних димеров, что и позволяет образоваться очень прочной цилиндрической конструкции. Как же такая конструкция может обеспечивать перемещение чего-либо внутри клетки? Что касается органелл, белков и других компонентов клетки, то они перемещаются по МТ, прикрепляясь к белкам-моторам: динеинам и кинезинам, которые способны буквально «шагать» по МТ в определенном направлении, потребляя в качестве топлива АТФ. Хромосомы же прикрепляются к концам МТ, которые затем каким-то образом быстро растаскивают их к полюсам веретена деления.
Ссылки
- Reider, CL, S Faruki and A Khodjakov (2001) TRENDS in Cell Biology. 11. 10: 413-418.
Wikimedia Foundation. 2010.
dic.academic.ru