
2.6. Клеточный центр. Органоиды движения. Клеточные включения
Вопрос 1. Каковы функции клеточного центра?
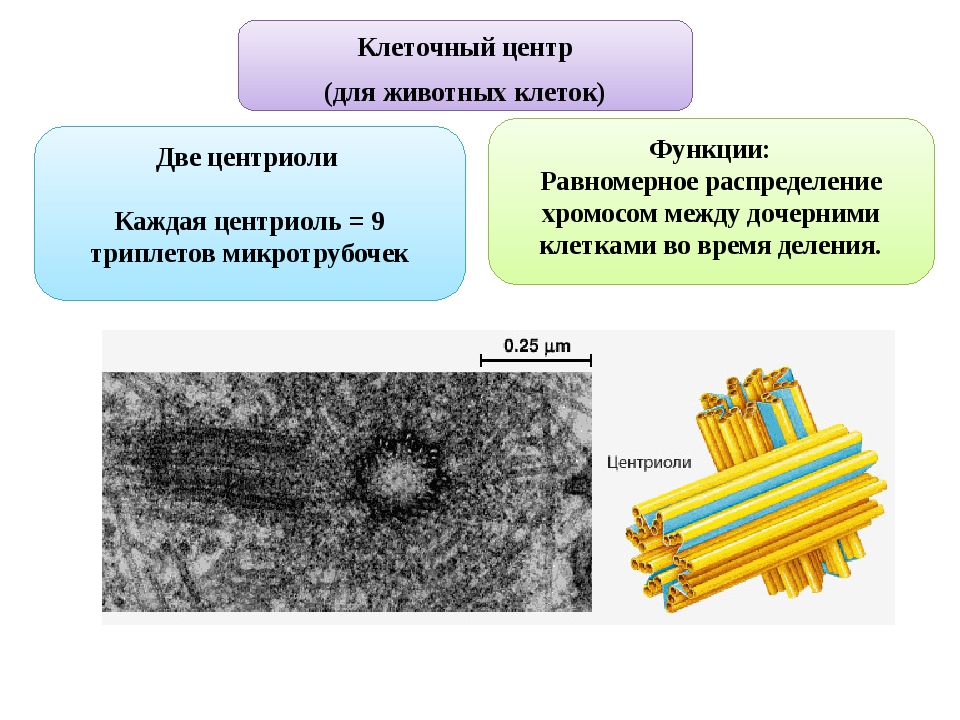
Клеточный центр выполняет функцию формирования внутреннего скелета клетки (цитоскелета). Цитоскелет представляет собой сеть микротрубочек, пронизывающих цитоплазму, поддерживающих форму клетки, обеспечивающих движение органоидов клетки, а также работу специализированных органоидов движения — ресничек и жгутиков.
Клеточный центр обеспечивает также и нормальное деление клетки. Центриоли клеточного центра расходятся к полюсам делящейся клетки и образуют веретено деления, благодаря которому из одной материнской впоследствии образуются две дочерние клетки.
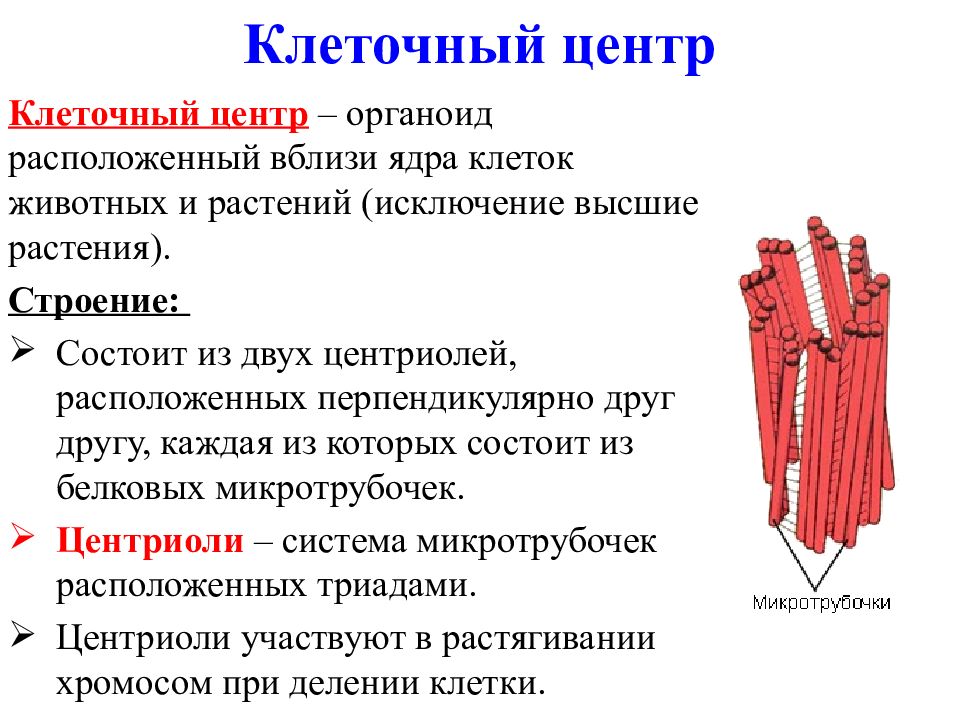
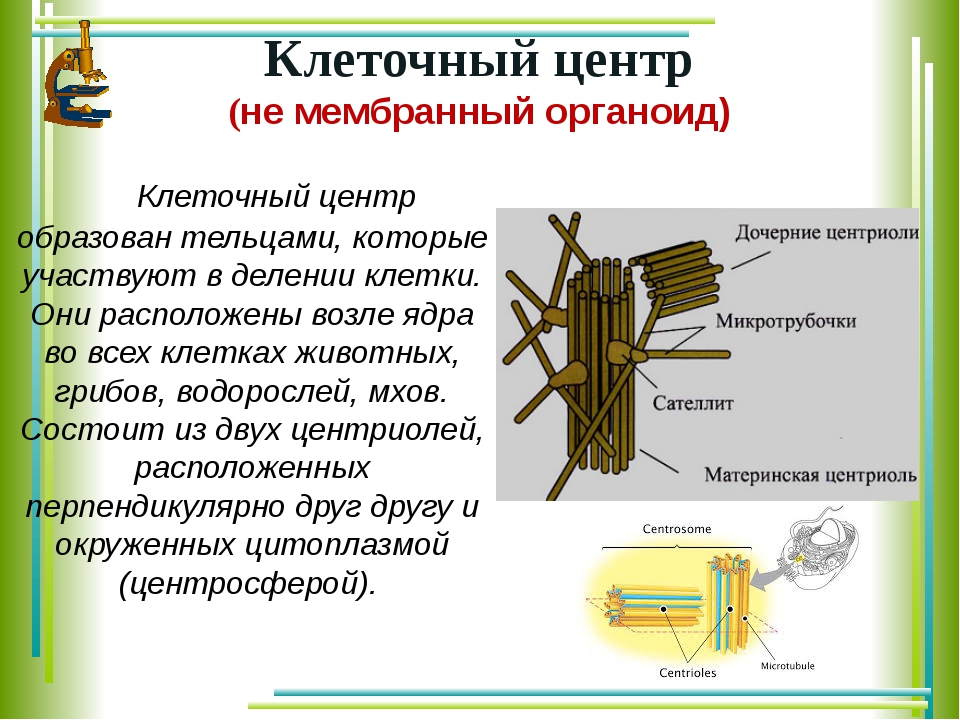
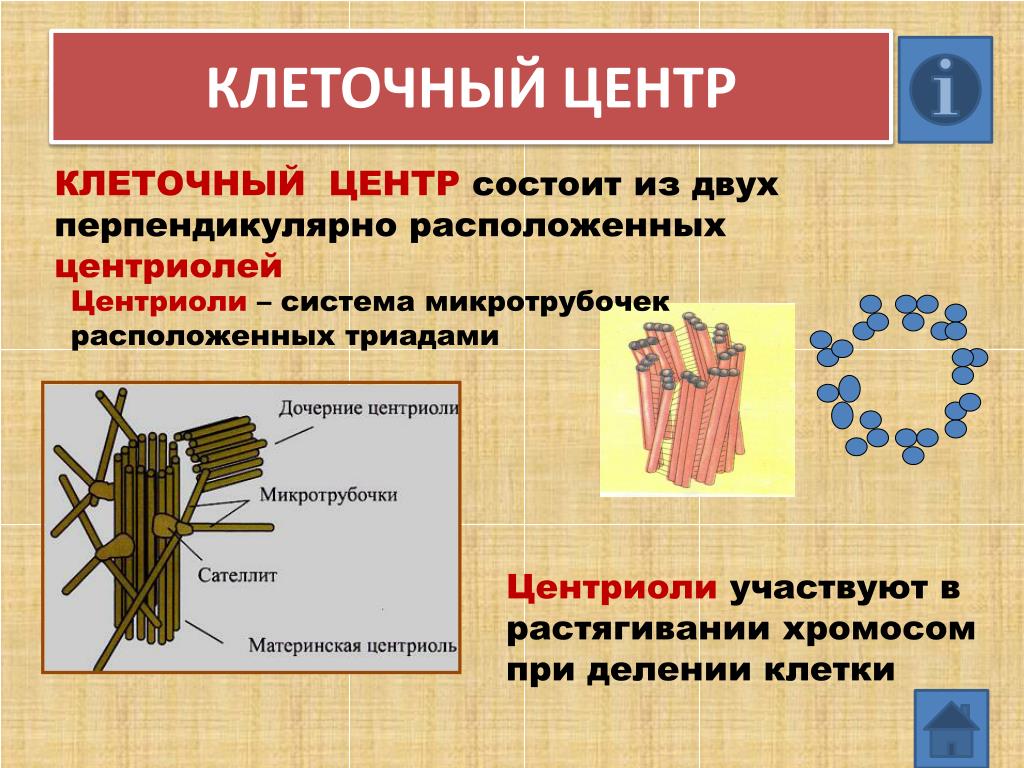
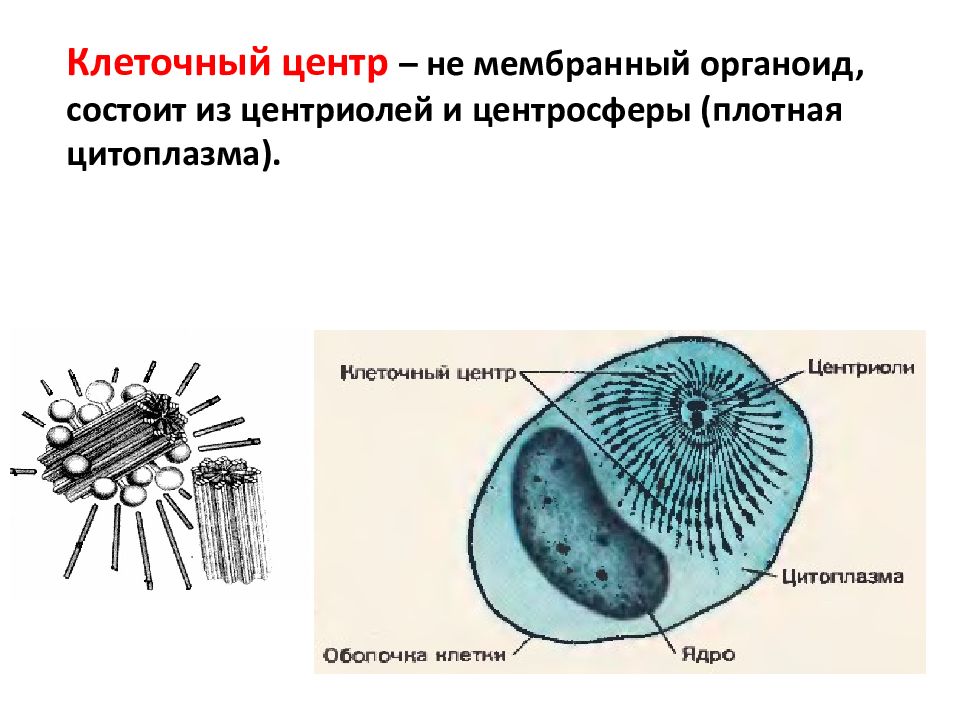
Центриоли представлены цилиндриками, образованными множеством микротрубочек. Центриоли, расположенные под прямым углом друг относительно друга, находятся вблизи от ядра и образуют клеточный центр.
Вопрос 2. Каковы функции центриолей в клетке?
Центриоли входят в состав клеточного центра и обеспечивают нормальное деление клетки.
Вопрос 3. В чем сходство и различие между ресничками и жгутиками?
У органоидов движения клетки много общего. Реснички и жгутики являются специализированными органоидами движения клетки, они образованы микротрубочками. В основании и жгутика, и реснички лежит базальное тельце, которое укрепляет их в цитоплазме клетки. Механизм движения ресничек и жгутиков одинаков, в его основе лежит скольжение микротрубочек друг относительно друга. Сходство этих органоидов движения заключается также и в том, что на их работу расходуется энергия АТФ.
Различаются реснички и жгутики размерами. Жгутики в несколько раз длиннее ресничек. Кроме того, реснички, изгибаясь волнообразно, обеспечивают клетке плавное, медленное передвижение. Жгутик же осуществляет вращательные движения, что позволяет клетке активно перемещаться.
Вопрос 4. Назовите примеры клеточных включений.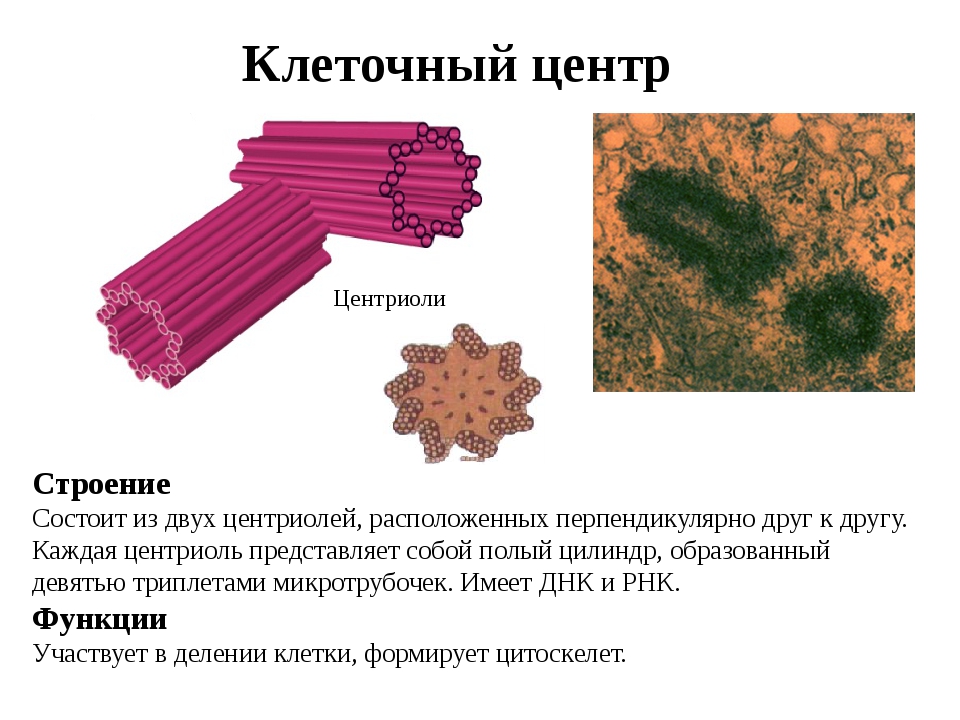
Временные образования в клетке называют клеточными включениями. К ним относятся гранулы крахмала, гликогена или белка, мелкие капли жира, кристаллы солей.
2.6. Клеточный центр. Органоиды движения. Клеточные включения
4.7 (94.84%) 31 votesНа этой странице искали :
- каковы функции клеточного центра
- функции клеточного центра
- клеточный центр функции
- каковы функции центриолей в клетке
- клеточный центр выполняет функции
Сохрани к себе на стену!
Open Library — открытая библиотека учебной информации
Биология Вакуоли
просмотров — 176
Клеточный центр
Органоиды, участвующие в делении и движении клеток
К ним относятся клеточный центр и его производные – реснички и жгутики.
Клеточный центр имеется в животных клетках и у некоторых низших растений.
Строение клеточного центра в интерфазе.
Клеточный центр образован двумя центриолями (диплосома). Центриоли окружены светлой зоной – центросферой, от которой отходит лучистость, образующая астросферу. Между центриолями находится удлиненное тельце – мостик (центродесмоза), который во время митотического деления принимает участие в построении ахроматинового веретена. Каждая центриоль представляет собой цилиндр, стенка которого состоит из девяти комплексов микротрубочек длиной около 0,5 и 0,25 мкм. Каждый комплекс состоит из трех микротрубочек и в связи с этим принято называть триплетом.
Центриоли расположены взаимно перпендикулярно: одна материнская, другая – дочерняя. Материнская центриоль окружена электронно-плотным ободком, образованным шаровидными сателлитами, соединенными плотным материалом с наружной стороной каждого триплета. Из них образуется новая центриоль.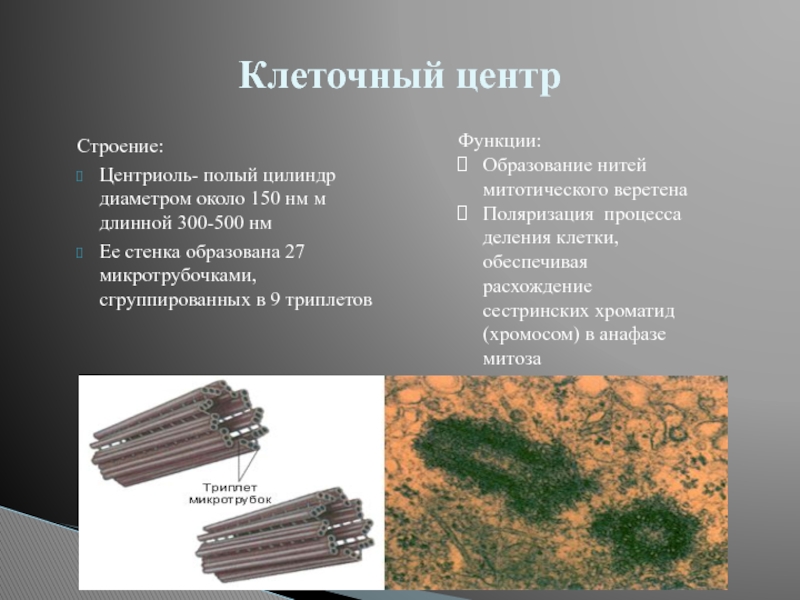
Средняя часть материнской центриоли может быть окружена комплексом фибриллярных структур, называемым гало.
К концу сателлитов и к области гало по цитоплазме транспортируются тубулины, и именно здесь происходит сборка микротрубочек.
Функции клеточного центра:
1) сборка микротрубочек;
2) клеточный центр играет важную роль при митотическом делении клеток; во время митоза центриоли удваиваются и вместе с окружающими их астросферами расходятся к полюсам клетки, участвуя в образовании митотического аппарата; с помощью этого аппарата осуществляется равномерное распределение хромосом между дочерними клетками;
3) центриоли принимают участие в образовании базальных телец, лежащих в основании мерцательных ресничек и жгутиков.
Вакуоли – одномембранные органоиды общего значения, имеются в растительных клетках. Молодые растительные клетки целиком заполнены цитоплазмой. Ядро в них довольно крупное и занимает центральное положение.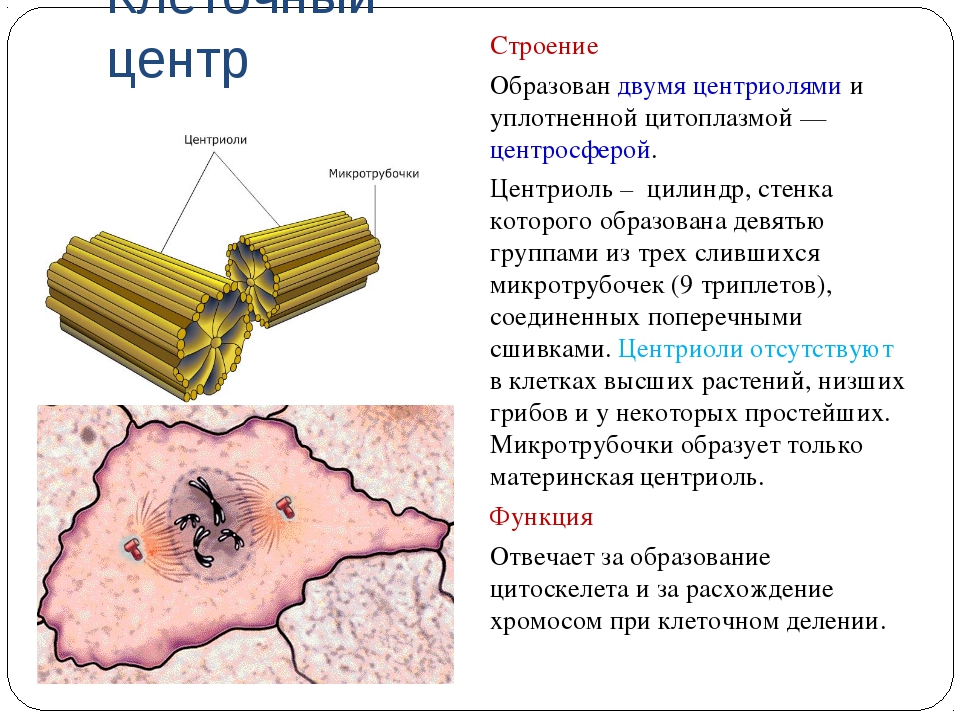
Клеточный сок образуется в результате обмена веществ в процессе жизнедеятельности всего растительного организма. Он является водным раствором различных органических и неорганических веществ. Основной частью клеточного сока является вода. Ее содержание доходит до 70 и даже 95%. Химический состав клеточного сока у растений различный, от него зависят их вкусовые качества. Клеточный сок обычно имеет кислую реакцию, реже нейтральную и еще реже – щелочную. В клеточном соке находятся в растворенном состоянии различные органические кислоты, сахара, соли, белки, дубильные вещества, гликозиды, алкалоиды, пигменты и другие вещества.
Функции вакуолей.
а) процесс вакуолизации – крайне важное условие роста клеток растяжением;
б) принимают участие в поддержании тургорного давления в клетках.
7. 3. Демонстрация преподавателем методики практических приемов по данной теме.
Преподаватель знакомит студентов с планом и методикой проведения практической работы.
7. 4. Самостоятельная работа студентов под контролем преподавателя.
Читайте также
В клетках растений, грибов и многих протистов содержатся вакуоли — крупные мембранные пузырьки или полости в цитоплазме, заполненные преимущественно водным содержимым. Вакуоли образуются из пузыревидных расширений эндоплазматического ретикулума или из пузырьков… [читать подробенее]
Вопрос 24. Вакуоли. Параплазматические (эргастические) включения
Функции гилоксисом
Гилоксисомы
Их характеристика
Микротельца — это гладкостенные пузырьки величиной 0,1-1,5 мкм с относительно проницаемой мембраной,. .. [читать подробенее]
.. [читать подробенее]
Пероксисомы Небольшие вакуоли, одетые одинарной мембраной, состоят из гранулярного матрикса, в центре которого располагается сердцевина. В зоне сердцевины видны кристаллоподобные структуры упакованных фибрилл. Пероксисомы обнаруживаются у человека главным… [читать подробенее]
Вакуоли содержатся почти во всех растительных клетках. Они представляют собой полости в клетке, заполненные водянистым содержимым – клеточным соком. От цитоплазмы клеточный сок изолирован избирательно проницаемой вакуолярной мембраной – тонопластом. Тонопласт… [читать подробенее]
Вакуоли содержатся практически во всех растительных клетках. Вакуоль – это полость в клетке, заполненная водянистым содержимым – клеточным соком и ограниченная от цитоплазмы избирательно проницаемой мембраной – тонопластом. Клеточный сок выделяется протопластом в.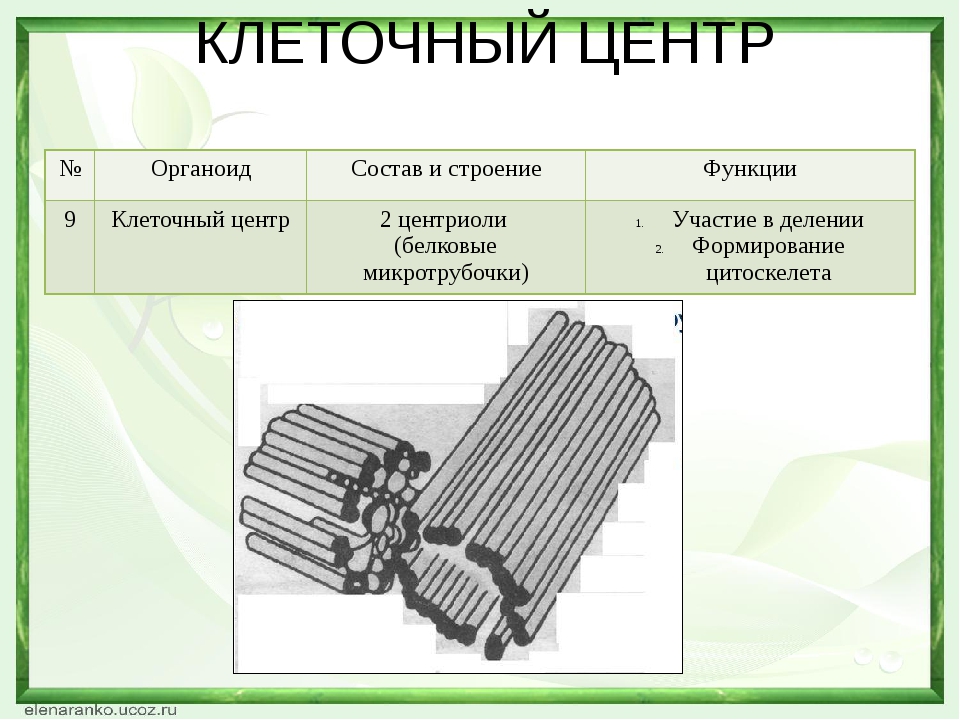 .. [читать подробенее]
.. [читать подробенее]
Лизосомы Лизосомы — одномембранные органоиды. Представляют собой мелкие пузырьки (диаметр от 0,2 до 0,8 мкм), содержащие набор гидролитических ферментов. Ферменты синтезируются на шероховатой ЭПС, перемещаются в аппарат Гольджи, где происходит их модификация и упаковка… [читать подробенее]
Откладываются Откладываются Из обмена веществ обмена Временно выведенные Конечные продукты КЛЕТОЧНЫЕ ВКЛЮЧЕНИЯ Под воздействием света В темноте ВОЗМОЖНЫЕ ВЗАИМОПЕРЕХОДЫ ПЛАСТИД Строение хлоропластов Хлоропласты &… [читать подробенее]
Откладываются Откладываются
Из обмена веществ обмена
Временно выведенные Конечные продукты
КЛЕТОЧНЫЕ ВКЛЮЧЕНИЯ
Под воздействием света
В темноте
ВОЗМОЖНЫЕ ВЗАИМОПЕРЕХОДЫ ПЛАСТИД
Строение хлоропластов
Хлоропласты &.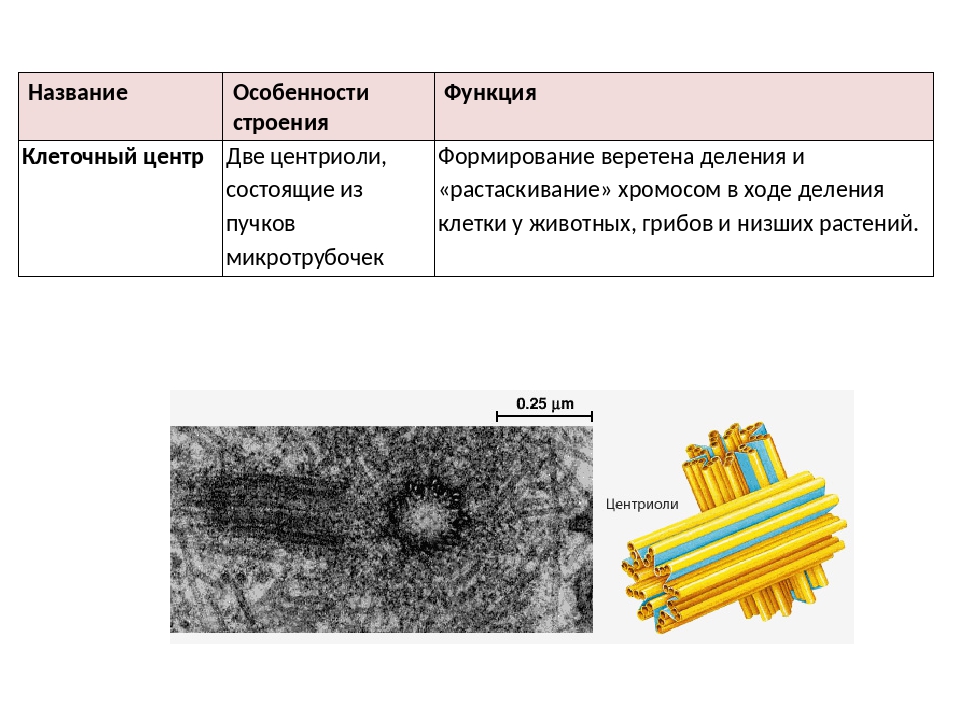
КОНТРОЛЬНІ ПИТАННЯ ТА ЗАВДАННЯ Перелік літератури з наведених питань Монтаж схем підключення ПЗВ Необхідною умовою нормального функціонування ПЗВ є відсутність у зоні дії ПЗВ любих з’єднань нульового робочого провідника N із заземленими елементами… [читать подробенее]
Жгутики и подвижность Хемотаксис – прямолинейное движение и кувыркание, реакция не зависит от способности микроорганизма утилизировать данный субстрат Наличие, число, расположение жгутиков является таксономически важным признаком. Монотрихи – один жгутик на… [читать подробенее]
Клеточный центр. Органоиды движения. Клеточные включения ❤️
Вопрос 1. Каковы функции клеточного центра?
Клеточный центр выполняет функцию формирования внутреннего скелета клетки. Цитоскелет представляет собой сеть микротрубочек, пронизывающих цитоплазму, поддерживающих форму клетки, обеспечивающих движение органоидов клетки, а также работу специализированных органоидов движения — ресничек и жгутиков.
Клеточный центр обеспечивает также и нормальное деление клетки. Центриоли клеточного центра расходятся к полюсам делящейся клетки и образуют веретено деления, благодаря которому из одной
материнской впоследствии образуются две дочерние клетки.Центриоли представлены цилиндриками, образованными множеством микротрубочек. Центриоли, расположенные под прямым углом друг относительно друга, находятся вблизи от ядра и образуют клеточный центр.
Вопрос 2. Каковы функции центриолей в клетке?
Центриоли входят в состав клеточного центра и обеспечивают нормальное деление клетки. Перед ее делением центриоли расходятся к полюсам, образуя веретено деления клетки.
Вопрос 3. В чем сходство и различие между ресничками и жгутиками?
У органоидов движения клетки много общего. Реснички
и жгутики являются специализированными органоидами движения клетки, они образованы микротрубочками. В основании и жгутика, и реснички лежит базальное тельце, которое укрепляет их в цитоплазме клетки. Механизм движения ресничек и жгутиков одинаков, в его основе лежит скольжение микротрубочек друг относительно друга. Сходство этих органоидов движения заключается также и в том, что на их работу расходуется энергия АТФ.
Механизм движения ресничек и жгутиков одинаков, в его основе лежит скольжение микротрубочек друг относительно друга. Сходство этих органоидов движения заключается также и в том, что на их работу расходуется энергия АТФ.Различаются реснички и жгутики размерами. Жгутики в несколько раз длиннее ресничек. Кроме того, реснички, изгибаясь волнообразно, обеспечивают клетке плавное, медленное передвижение. Жгутик же осуществляет вращательные движения, что позволяет клетке активно перемещаться.
Вопрос 4. Назовите примеры клеточных включений.
Временные образования в клетке называют клеточными включениями. К ним относятся гранулы крахмала, гликогена или белка, мелкие капли жира, кристаллы солей.
8. Строение и функции немембранных структур клетки . Карпатские орлы
В эту группу органоидов входят рибосомы, микротрубочки и микрофиламенты, клеточный центр. Рибосома
Это округлая рибонуклеопротеиновая частица. Диаметр ее составляет 20–30 нм. Состоит рибосома из большой и малой субъединиц, которые объединяются в присутствии нити м-РНК (матричной, или информационной, РНК). Комплекс из группы рибосом, объединенных одной молекулой м-РНК наподобие нитки бус, называется полисомой. Эти структуры либо свободно расположены в цитоплазме, либо прикреплены к мембранам гранулярной ЭПС (в обоих случаях на них активно протекает синтез белка).
Комплекс из группы рибосом, объединенных одной молекулой м-РНК наподобие нитки бус, называется полисомой. Эти структуры либо свободно расположены в цитоплазме, либо прикреплены к мембранам гранулярной ЭПС (в обоих случаях на них активно протекает синтез белка).
Полисомы гранулярной ЭПС образуют белки, выводимые из клетки и используемые для нужд всего организма (например, пищеварительные ферменты, белки женского грудного молока). Кроме этого, рибосомы присутствуют на внутренней поверхности мембран митохондрий, где также принимают активное участие в синтезе белковых молекул.
Микротрубочки
Это трубчатые полые образования, лишенные мембраны. Внешний диаметр составляет 24 нм, ширина просвета – 15 нм, толщина стенки – около 5 нм. В свободном состоянии представлены в цитоплазме, также являются структурными элементами жгутиков, цент-риолей, веретена деления, ресничек. Микротрубочки построены из стереотипных белковых субъединиц путем их полимеризации. В любой клетке процессы полимеризации идут параллельно процессам деполимеризации. Причем соотношение их определяется количеством микротрубочек. Микротрубочки имеют различную устойчивость к разрушающим их факторам, например, к колхицину (это химическое вещество, вызывающее деполимеризацию). Функции микротрубочек:
Причем соотношение их определяется количеством микротрубочек. Микротрубочки имеют различную устойчивость к разрушающим их факторам, например, к колхицину (это химическое вещество, вызывающее деполимеризацию). Функции микротрубочек:
1) являются опорным аппаратом клетки;
2) определяют формы и размеры клетки;
3) являются факторами направленного перемещения внутриклеточных структур.
Микрофиламенты
Это тонкие и длинные образования, которые обнаруживаются по всей цитоплазме. Иногда образуют пучки. Виды микро-филаментов:
1) актиновые. Содержат сократительные белки (актин), обеспечивают клеточные формы движения (например, амебоидные), играют роль клеточного каркаса, участвуют в организации перемещений органелл и участков цитоплазмы внутри клетки;
2) промежуточные (толщиной 10 нм). Их пучки обнаруживаются по периферии клетки под плазмалеммой и по окружности ядра. Выполняют опорную (каркасную) роль. В разных клетках (эпителиальных, мышечных, нервных, фибробластах) построены из разных белков.
Микрофиламенты, как и микротрубочки, построены из субъединиц, поэтому их количество определяется соотношением процессов полимеризации и деполимеризации.
Клетки всех животных, некоторых грибов, водорослей, высших растений характеризуются наличием клеточного центра. Клеточный центр обычно располагается рядом с ядром.
Он состоит из двух центриолей, каждая из которых представляет собой полый цилиндр диаметром около 150 нм, длиной 300–500 нм.
Центриоли расположены взаимоперпендикулярно. Стенка каждой центриоли образована 27 микротрубочками, состоящими из белка тубулина. Микротрубочки сгруппированы в 9 триплетов.
Из центриолей клеточного центра во время деления клетки образуются нити веретена деления.
Центриоли поляризуют процесс деления клетки, чем достигается равномерное расхождение сестринских хромосом (хроматид) в анафазе митоза.
90 000 охарактеризованных основных клеточных линий — начало работы Онкологический центрЗапросите или внесите клеточные линии через нашу страницу охарактеризованных клеточных линий Core iLab.

Запрос линий ячеек
Пожалуйста, ознакомьтесь с нашим Списком клеточных линий , доступных для покупки.
Для клиентов внутри MD Anderson: Когда линии сотовой связи будут готовы, вы получите уведомление по электронной почте, чтобы назначить встречу, чтобы забрать свои линии сотовой связи. В зависимости от клеточной линии время оборота составляет примерно одну неделю.
Для клиентов за пределами MD Anderson: После того как ваши формы iLab будут отправлены и полностью утверждены, MTA будет запущен Core. Процесс MTA может занять от двух до шести недель, в зависимости от загруженности юридического отдела MD Anderson и сотрудничества с внешним учреждением. После того, как полностью выполненный MTA будет возвращен, Core отправит клеточные линии.
**ВАЖНАЯ ИНФОРМАЦИЯ О КЛЕТОЧНЫХ ЛИНИЯХ, ПОЛУЧЕННЫХ ИЗ НАШЕГО ЯДРА** Все клеточные линии, распространяемые Core, проходят профилирование STR и тестируются на микоплазменное и бактериальное загрязнение непосредственно перед замораживанием. Мы также следим за тем, чтобы в каждом флаконе было достаточное количество клеток, чтобы гарантировать правильный рост клеток после получения и оттаивания. Если у вас возникли проблемы с выращиванием любой из клеточных линий, полученных от Core, свяжитесь с нами. Мы можем дать несколько советов по выращиванию ваших клеток, но мы не можем предоставить немедленную замену флакона. Существует несколько причин, по которым клеточные линии могут расти неправильно, например: перенос и хранение после получения, метод размораживания, соответствующая среда, концентрация FBS, уровень и температура инкубатора CO 2 , опыт выращивания клеток и другие.Таким образом, мы предоставим флакон на замену только после оценки всех вышеперечисленных параметров и заключения о том, что клиенты следовали передовым методам культивирования клеток.
Мы также следим за тем, чтобы в каждом флаконе было достаточное количество клеток, чтобы гарантировать правильный рост клеток после получения и оттаивания. Если у вас возникли проблемы с выращиванием любой из клеточных линий, полученных от Core, свяжитесь с нами. Мы можем дать несколько советов по выращиванию ваших клеток, но мы не можем предоставить немедленную замену флакона. Существует несколько причин, по которым клеточные линии могут расти неправильно, например: перенос и хранение после получения, метод размораживания, соответствующая среда, концентрация FBS, уровень и температура инкубатора CO 2 , опыт выращивания клеток и другие.Таким образом, мы предоставим флакон на замену только после оценки всех вышеперечисленных параметров и заключения о том, что клиенты следовали передовым методам культивирования клеток.
Депозитные клеточные линии
- Это для любого PI, который желает поделиться клеточными линиями с другими внутренними и/или внешними пользователями ядра охарактеризованных клеточных линий MD Anderson.

- Это приносит пользу PI, предоставляя им безопасный сайт для хранения своих клеточных линий, устраняя затраты на расширение и распространение клеточных линий среди запрашивающих лиц и увеличивая количество клеточных линий, доступных для PI.
- Ядро будет расширять, хранить и распространять депонированные клеточные линии. Ядро также бесплатно предоставит флакон с депонированными клеточными линиями по запросу депонирующего ИП.
- Хранение гарантируется на срок до 3 (трех) лет с даты сдачи на хранение.
- The Core не принимает клеточные линии, которые:
- получены из коммерческих источников,
- ограничено MTA и/или
- микоплазма положительная.(Все клеточные линии будут проверены сразу после внесения, а микоплазмоположительные клеточные линии будут уничтожены после уведомления ИП.)
Характеристики центра трансплантации и выживаемость после аллогенной трансплантации гемопоэтических клеток у взрослых

Pasquini MC, Xhu X. Текущее использование и результаты трансплантации гемопоэтических стволовых клеток: CIBMTR Summary Slides, 2015. http://www.cibmtr.org .
Lee SJ, Astigarraga CC, Eapen M, Artz AS, Davies SM, Champlin R, et al.Различия в методах поддерживающей терапии при трансплантации гемопоэтических клеток. Biol Blood Marrow Transpl. 2008; 14:1231–8.
Артикул Google Scholar
Lee SJ, Joffe S, Artz AS, Champlin RE, Davies SM, Jagasia M, et al. Вариации индивидуальной практики врачей при трансплантации гемопоэтических клеток. Дж. Клин Онкол. 2008;26:2162–70.
ПабМед Статья ПабМед Центральный Google Scholar
Lee SJ, Vogelsang G, Gilman A, Weisdorf DJ, Pavletic S, Antin JH, et al. Обзор диагностики, лечения и классификации хронической РТПХ. Biol Blood Marrow Transpl. 2002; 8: 32–9.
Артикул Google Scholar
Пидала Дж., Ли С.Дж., Куинн Г., Джим Х., Ким Дж., Анасетти С. Варианты лечения подавления иммунитета после аллогенной трансплантации гемопоэтических клеток. Biol Blood Marrow Transpl. 2011;17:1528–36.
Артикул Google Scholar
Трифилио С., Верма А., Мехта Дж. Антимикробная профилактика у реципиентов трансплантата гемопоэтических стволовых клеток: неоднородность современной клинической практики. Пересадка костного мозга. 2004; 33: 735–9.
КАС пабмед Статья ПабМед Центральный Google Scholar
Windrum P, Morris TC, Drake MB, Niederwieser D, Ruutu T. Изменение использования диметилсульфоксида при трансплантации стволовых клеток: обзор центров EBMT. Пересадка костного мозга.2005; 36: 601–3.
КАС пабмед Статья ПабМед Центральный Google Scholar
Majhail NS, Mau LW, Chitphakdithai P, Payton T, Eckrich M, Joffe S, et al. Национальный опрос персонала центров трансплантации гемопоэтических клеток, инфраструктуры и моделей оказания помощи. Biol Blood Marrow Transpl. 2015;21:1308–14.
Национальный опрос персонала центров трансплантации гемопоэтических клеток, инфраструктуры и моделей оказания помощи. Biol Blood Marrow Transpl. 2015;21:1308–14.
Артикул Google Scholar
Majhail NS, Murphy EA, Omondi NA, Robinett P, Gajewski JL, LeMaistre CF, et al. Аллогенный врач-трансплантолог и возможности центра в США. Biol Blood Marrow Transpl. 2011;17:956–61.
Артикул Google Scholar
Murphy EA, Ferguson SS, Omondi NA, Getzendaner LC, Gajewski JL, Goldstein GA, et al. Симпозиум Национальной программы доноров костного мозга по защите интересов пациентов в терапии клеточной трансплантацией: устранение препятствий для трансплантации гемопоэтических клеток.Biol Blood Marrow Transpl. 2010;16:147–56.
Артикул Google Scholar
Маджхейл Н.С. Оптимизация качества и эффективности оказания медицинской помощи при трансплантации гемопоэтических клеток.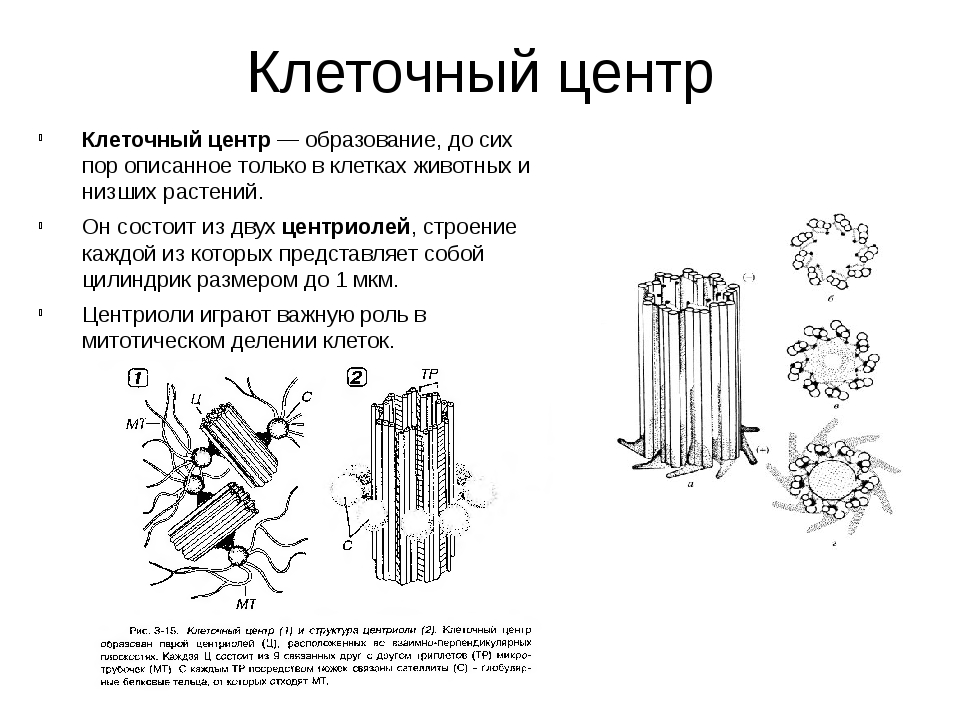 Curr Hematologic Malignancy Rep. 2015; 10:199–204.
Curr Hematologic Malignancy Rep. 2015; 10:199–204.
Артикул Google Scholar
Horowitz MM, Przepiorka D, Champlin RE, Gale RP, Gratwohl A, Herzig RH, et al. Должны ли HLA-идентичные трансплантаты костного мозга братьев и сестер при лейкемии быть ограничены крупными центрами? Кровь.1992; 79: 2771–4.
КАС пабмед Статья ПабМед Центральный Google Scholar
Loberiza FR Jr, Zhang MJ, Lee SJ, Klein JP, LeMaistre CF, Serna DS, et al. Ассоциация центров трансплантологии и врачей влияет на смертность после трансплантации гемопоэтических стволовых клеток в Соединенных Штатах. Кровь. 2005; 105: 2979–87.
КАС пабмед Статья ПабМед Центральный Google Scholar
Gratwohl A, Sureda A, Baldomero H, Gratwohl M, Dreger P, Kroger N, et al. Экономика и исход после трансплантации гемопоэтических стволовых клеток: ретроспективное когортное исследование. ЭБио Медицина. 2015;2:2101–9.
ЭБио Медицина. 2015;2:2101–9.
Google Scholar
Klingebiel T, Cornish J, Labopin M, Locatelli F, Darbyshire P, Handgretinger R, et al. Результаты и факторы, влияющие на результат после полностью гаплоидентичной трансплантации гемопоэтических стволовых клеток у детей с острым лимфобластным лейкозом очень высокого риска: влияние размера центра: анализ от имени рабочих групп по острым лейкозам и детским заболеваниям Европейской группы по пересадке крови и костного мозга.Кровь. 2010; 115:3437–46.
КАС пабмед Статья Google Scholar
Gratwohl A, Brand R, McGrath E, van Biezen A, Sureda A, Ljungman P, et al. Использование системы управления качеством «JACIE» и исход после трансплантации гемопоэтических стволовых клеток. Гематология. 2014; 99: 908–15.
ПабМед ПабМед Центральный Статья Google Scholar
Хан Т., Маккарти П.Л.-младший, Хассеброк А., Бредесон С., Гаевски Дж.Л., Хейл Г.А. и др. Значительное улучшение выживаемости после аллогенной трансплантации гемопоэтических клеток в период значительного увеличения использования, пожилого возраста реципиента и использования неродственных доноров. Дж. Клин Онкол. 2013;31:2437–49.
ПабМед ПабМед Центральный Статья Google Scholar
McCarthy PL Jr, Hahn T, Hassebroek A, Bredeson C, Gajewski J, Hale G, et al.Тенденции в использовании и выживаемости после трансплантации аутологичных гемопоэтических клеток в Северной Америке, 1995-2005 гг.: значительное улучшение выживаемости при лимфоме и миеломе в период увеличения возраста реципиента. Biol Blood Marrow Transpl. 2013;19:1116–23.
Артикул Google Scholar
Администрация ресурсов и услуг здравоохранения. Трансплантация клеток крови. http://bloodcell.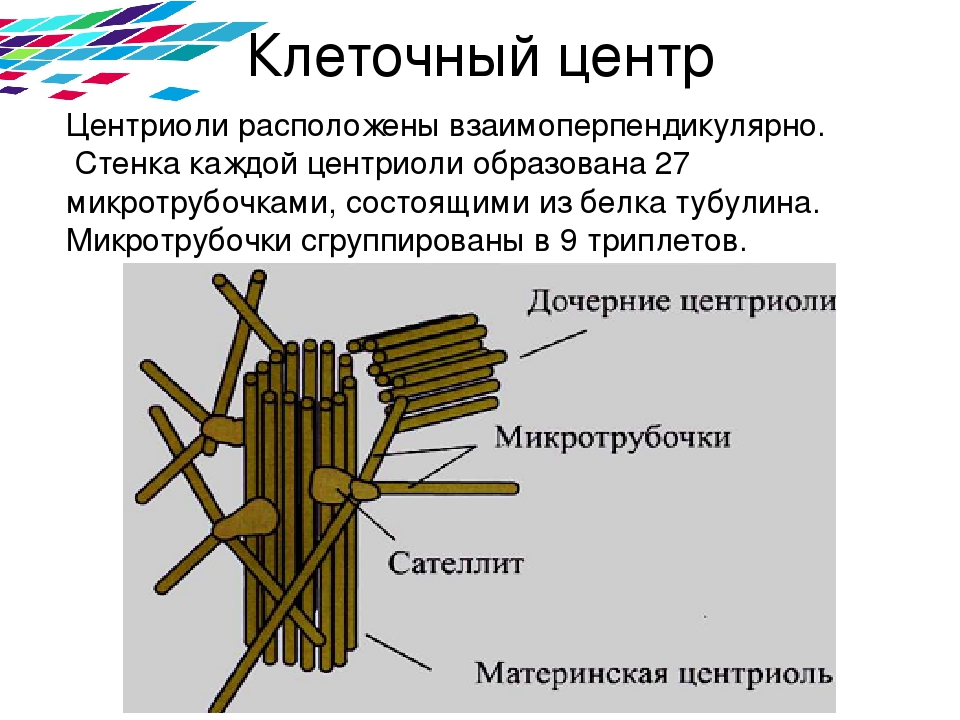 transplant.hrsa.gov/index.html. По состоянию на 27 января 2017 г.
transplant.hrsa.gov/index.html. По состоянию на 27 января 2017 г.
Логан Б.Р., Нельсон Г.О., Кляйн Дж.П. Центр анализа конкретных результатов трансплантации гемопоэтических клеток. Анализ данных за всю жизнь. 2008; 14: 389–404.
ПабМед ПабМед Центральный Статья Google Scholar
Центр международных исследований по пересадке крови и костного мозга: статистика пересадки и выживания в США на соответствующих сайтах. http://www.cibmtr.org/ReferenceCenter/SlidesReports/USStats. По состоянию на 27 января 2017 г.
Соррор М.Л., Марис М.Б., Сторб Р., Барон Ф., Сандмайер Б.М., Малони Д.Г. и др. Индекс коморбидности, специфичный для трансплантации гемопоэтических клеток (HCT): новый инструмент для оценки риска перед аллогенной HCT. Кровь. 2005; 106: 2912–9.
КАС пабмед ПабМед Центральный Статья Google Scholar

Контал С., О’Куигли Дж. Применение методов изменения точки при изучении влияния возраста на выживаемость при раке молочной железы.Анализ статистических данных. 1999; 30: 253–70.
Артикул Google Scholar
Majhail NS, Chitphakdithai P, Logan B, King R, Devine S, Rossmann SN, et al. Значительное улучшение выживаемости после трансплантации гемопоэтических клеток от неродственного донора в последнее время. Biol Blood Marrow Transpl. 2015;21:142–50.
Артикул Google Scholar
Horan JT, Logan BR, Agovi-Johnson MA, Lazarus HM, Bacigalupo AA, Ballen KK, et al.Снижение риска смертности, связанной с трансплантацией, после аллогенной трансплантации гемопоэтических клеток: насколько достигнут прогресс? Дж. Клин Онкол. 2011;29:805–13.
ПабМед ПабМед Центральный Статья Google Scholar

Чоудри Н.К., Флетчер Р.Х., Сумерай С.Б. Систематический обзор: взаимосвязь между клиническим опытом и качеством медицинской помощи. Энн Интерн Мед. 2005; 142: 260–73.
ПабМед Статья ПабМед Центральный Google Scholar
Рейд Р.О., Фридберг М.В., Адамс Дж.Л., МакГлинн Э.А., Мехротра А. Связь между характеристиками врача и качеством медицинской помощи. Arch Intern Med. 2010; 170:1442–9.
ПабМед Статья ПабМед Центральный Google Scholar
Груен Р.Л., Питт В., Грин С., Паркхилл А., Кэмпбелл Д., Джолли Д. Влияние объема медицинских услуг на смертность от рака: систематический обзор и метаанализ. CA: Рак J Clin. 2009; 59: 192–211.
Google Scholar
Halm EA, Lee C, Chassin MR. Связан ли объем с результатом в здравоохранении? Систематический обзор и методологическая критика литературы. Энн Интерн Мед. 2002; 137: 511–20.
Энн Интерн Мед. 2002; 137: 511–20.
ПабМед Статья ПабМед Центральный Google Scholar
Ауэрбах А.Д., Маселли Дж., Картер Дж., Пеков П.С., Линденауэр П.К. Взаимосвязь между объемом случаев, качеством лечения и результатами сложной онкологической хирургии. J Am Coll Surg. 2010; 211:601–8.
ПабМед ПабМед Центральный Статья Google Scholar
McAteer JP, LaRiviere CA, Drugas GT, Abdullah F, Oldham KT, Goldin AB. Влияние опыта хирурга, объема стационара и назначения специальности на исходы в детской хирургии: систематический обзор. JAMA Педиатр. 2013; 167: 468–75.
ПабМед Статья ПабМед Центральный Google Scholar
Чута Л.Н., Парк Х.С., Боффа Д.Д., Бласберг Д.Д., Деттербек Ф.К., Ким А.В. Госпитальный объем и результаты роботизированных лобэктомий. Грудь. 2017; 151:329–39.
ПабМед Статья ПабМед Центральный Google Scholar
Park HS, Detterbeck FC, Boffa DJ, Kim AW. Влияние госпитального объема торакоскопической лобэктомии на первичные исходы рака легкого. Энн Торак Серг. 2012;93:372–9.
ПабМед Статья ПабМед Центральный Google Scholar
Lin X, Tao H, Cai M, Liao A, Cheng Z, Lin H. Систематический обзор и метаанализ взаимосвязи между объемом больницы и результатами чрескожного коронарного вмешательства. Лекарство. 2016;95:e2687.
ПабМед ПабМед Центральный Статья Google Scholar
Кристиан К.К., Густафсон М.Л., Бетенский Р.А., Дейли Дж., Зиннер М.Дж. Отношение объем-результат: не верьте всему, что видите. Мир J Surg. 2005; 29:1241–4.
ПабМед Статья ПабМед Центральный Google Scholar

Месман Р., Вестерт Г.П., Берден Б.Дж., Фабер М.Дж. Почему больницы с большим объемом лечения достигают лучших результатов? Систематический обзор промежуточных факторов в отношениях объем-результат. Политика здравоохранения. 2015; 119:1055–67.
ПабМед Статья ПабМед Центральный Google Scholar
Poirel HA, Vanspauwen M, Macq G, De Geyndt A, Maertens J, Willems E, et al.Предоставление как аутологичных, так и аллогенных трансплантатов гемопоэтических стволовых клеток (ТГСК) может оказать более сильное влияние на исход аутологичных ТГСК у взрослых пациентов, чем уровень активности или внедрение JACIE в бельгийских центрах трансплантации. Пересадка костного мозга. 2019;54:1434–42.
ПабМед Статья ПабМед Центральный Google Scholar
Бейкер К.С., Дэвис С.М., Маджхайл Н.С., Хассебрук А., Кляйн Дж.П., Баллен К.К. и др. Раса и социально-экономический статус влияют на результаты трансплантации гемопоэтических клеток от неродственного донора.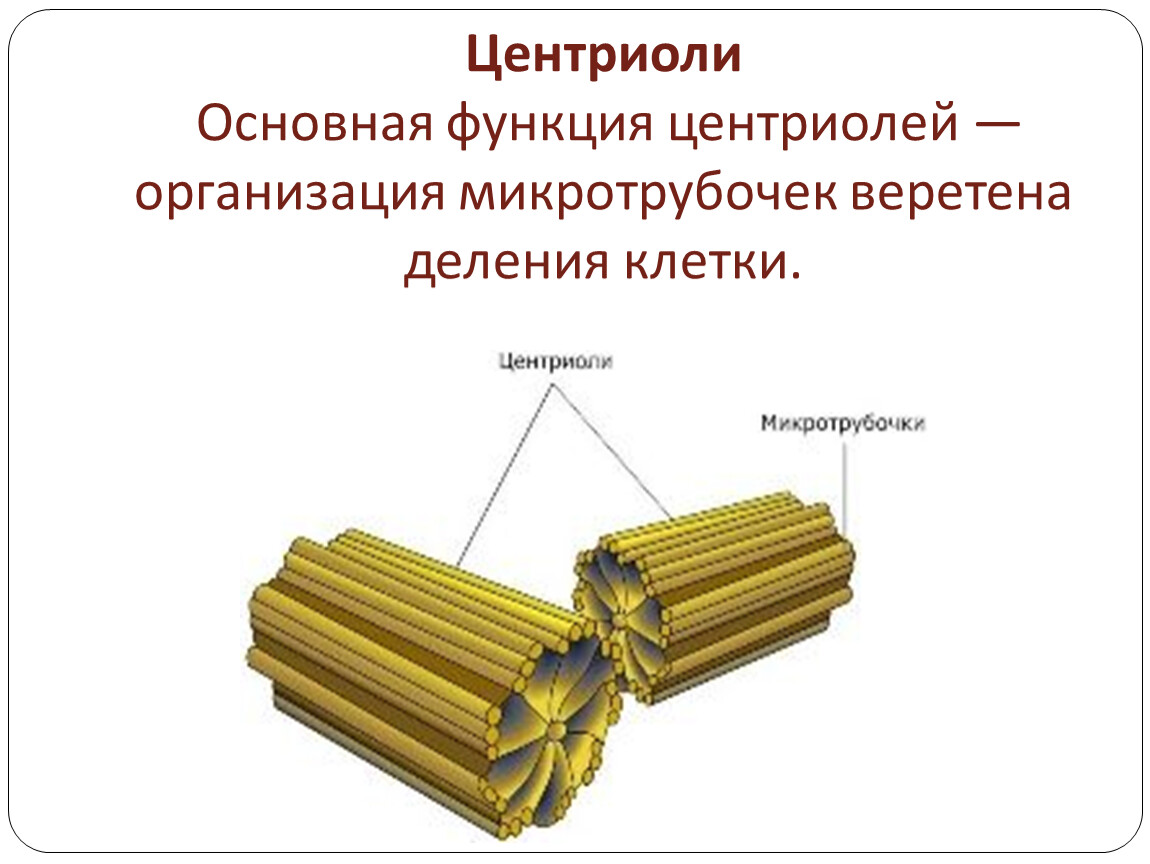 Biol Blood Marrow Transpl. 2009; 15:1543–54.
Biol Blood Marrow Transpl. 2009; 15:1543–54.
Артикул Google Scholar
Majhail NS, Omondi NA, Denzen E, Murphy EA, Rizzo JD. Доступ к трансплантации гемопоэтических клеток в США. Biol Blood Marrow Transpl. 2010;16:1070–5.
Артикул Google Scholar
Majhail NS, Nayyar S, Santibanez ME, Murphy EA, Denzen EM. Расовые различия в трансплантации гемопоэтических клеток в Соединенных Штатах.Пересадка костного мозга. 2012;47:1385–90.
КАС пабмед Статья ПабМед Центральный Google Scholar
Хашми С., Карпентер П., Кера Н., Тичелли А., Савани Б.Н. Затерянные в переходе: насущная потребность в клинике длительного наблюдения за выжившими после трансплантации крови и костного мозга. Biol Blood Marrow Transpl. 2015;21:225–32.
Артикул Google Scholar

Khera N, Martin P, Edsall K, Bonagura A, Burns LJ, Juckett M, et al. Координация помощи пациенту при трансплантации гемопоэтических клеток. Кровь Adv. 2017;1:1617–27.
ПабМед ПабМед Центральный Статья Google Scholar
Battiwalla M, Hashmi S, Majhail N, Pavletic S, Savani BN, Shelburne N. Инициатива Национального института здравоохранения по трансплантации гемопоэтических клеток: разработка рекомендаций по улучшению выживаемости и долгосрочных результатов.Biol Blood Marrow Transpl. 2017;23:6–9.
Артикул Google Scholar
Hashmi SK, Bredeson C, Duarte RF, Farnia S, Ferrey S, Fitzhugh C, et al. Инициатива Национального института здравоохранения по отдаленным последствиям пересадки крови и костного мозга: отчет рабочей группы по оказанию медицинской помощи. Biol Blood Marrow Transpl. 2017;23:717–25.
Артикул Google Scholar

Majhail NS, Rizzo JD, Lee SJ, Aljuurf M, Atsuta Y, Bonfim C, et al.Рекомендуемые методы скрининга и профилактики для лиц с длительным выживанием после трансплантации гемопоэтических клеток. Biol Blood Marrow Transpl. 2012;18:348–71.
Артикул Google Scholar
Majhail NS, Murphy E, Laud P, Preussler JM, Denzen EM, Abetti B, et al. Рандомизированное контролируемое исследование индивидуального резюме лечения и планов ухода за выжившими после трансплантации гемопоэтических клеток. Гематология. 2019;104:1084–92.
ПабМед ПабМед Центральный Статья Google Scholar
Denzen EM, Preussler JM, Murphy EA, Baker KS, Burns LJ, Foster J, et al. Адаптация плана ухода за выжившими: предпочтения пациента и поставщика для реципиентов трансплантации гемопоэтических клеток. Biol Blood Marrow Transpl. 2019;25:562–9.
Артикул Google Scholar
Hashmi SK, Lee SJ, Savani BN, Burns L, Wingard JR, Perales MA, et al.Опрос комитета по практическим рекомендациям ASBMT по клиникам долгосрочного наблюдения за выжившими после трансплантации гемопоэтических клеток. Biol Blood Marrow Transpl. 2018;24:1119–24.
Артикул Google Scholar
Транскриптомы одиночных клеток выявляют характерные черты типов клеток островков поджелудочной железы человека
doi: 10.15252/embr.201540946. Epub 2015 21 декабря.Принадлежности Расширять
Принадлежности
- 1 Исследовательский центр молекулярной медицины CeMM Австрийской академии наук, Вена, Австрия.

- 2 Венский медицинский университет Анна Шпигель Forschungsgebäude, Вена, Австрия.
- 3 Отделение хирургии, Центр клеточной изоляции и трансплантации, Университетские больницы Женевы Женевский университет, Женева, Швейцария.
- 4 Исследовательский центр молекулярной медицины CeMM Австрийской академии наук, Вена, Австрия Факультет лабораторной медицины Венского медицинского университета, Вена, Австрия Институт информатики им. Макса Планка, Саарбрюккен, Германия [email protected] [email protected]. Тел. .ac.at [email protected].
Элемент в буфере обмена
Джин Ли и др. Представитель ЕМБО
2016 февраль.
Представитель ЕМБО
2016 февраль.
Показать варианты
Формат АннотацияPubMedPMID
дои: 10.15252/эмбр.201540946. Epub 2015 21 декабря.Принадлежности
- 1 Исследовательский центр молекулярной медицины CeMM Австрийской академии наук, Вена, Австрия.
- 2 Венский медицинский университет Анна Шпигель Forschungsgebäude, Вена, Австрия.

- 3 Отделение хирургии, Центр клеточной изоляции и трансплантации, Университетские больницы Женевы Женевский университет, Женева, Швейцария.
- 4 Исследовательский центр молекулярной медицины CeMM Австрийской академии наук, Вена, Австрия Факультет лабораторной медицины Венского медицинского университета, Вена, Австрия Институт информатики им. Макса Планка, Саарбрюккен, Германия [email protected] [email protected]. Тел. .ac.at [email protected].
Элемент в буфере обмена
Полнотекстовые ссылки Параметры отображения цитированияПоказать варианты
Формат АннотацияPubMedPMID
Абстрактный
Панкреатические островки Лангерганса содержат несколько специализированных типов эндокринных клеток, которые обычно идентифицируются по экспрессии отдельных маркерных генов. Однако установленные маркерные гены не могут охватить полный спектр клеточной гетерогенности в островках поджелудочной железы человека, а существующие объемные наборы данных транскриптома обеспечивают средние значения для нескольких клеточных популяций. Чтобы проанализировать клеточный состав островка поджелудочной железы человека и установить транскриптомы для всех основных типов клеток, мы провели секвенирование одноклеточной РНК на 70 клетках, отсортированных из первичной ткани человека. Мы использовали этот набор данных для проверки ранее описанных маркерных генов на уровне отдельных клеток и для определения специфически экспрессируемых факторов транскрипции для всех подтипов островковых клеток.Все данные доступны для просмотра и загрузки, что создает полезный ресурс профилей экспрессии одиночных клеток для эндокринных клеток в островках поджелудочной железы человека.
Однако установленные маркерные гены не могут охватить полный спектр клеточной гетерогенности в островках поджелудочной железы человека, а существующие объемные наборы данных транскриптома обеспечивают средние значения для нескольких клеточных популяций. Чтобы проанализировать клеточный состав островка поджелудочной железы человека и установить транскриптомы для всех основных типов клеток, мы провели секвенирование одноклеточной РНК на 70 клетках, отсортированных из первичной ткани человека. Мы использовали этот набор данных для проверки ранее описанных маркерных генов на уровне отдельных клеток и для определения специфически экспрессируемых факторов транскрипции для всех подтипов островковых клеток.Все данные доступны для просмотра и загрузки, что создает полезный ресурс профилей экспрессии одиночных клеток для эндокринных клеток в островках поджелудочной железы человека.
Ключевые слова: альфа-клетки; бета-клетки; сахарный диабет; маркерные гены; одноклеточная РНК-seq.
© 2015 Авторы.Публикуется на условиях лицензии CC BY NC ND 4.0.
Цифры
Рисунок 1. Транскриптомы одиночных клеток резюмируют основные…
Рис. 1.Транскриптомы одиночных клеток повторяют основные типы клеток поджелудочной железы
Рабочий процесс получения и анализа…
Рабочий процесс для получения и анализа данных секвенирования РНК отдельных клеток из островков поджелудочной железы человека.

Итеративный подход на основе PCA/biplot для идентификации типов клеток и транскриптов, определяющих тип клеток, по данным секвенирования РНК одиночных клеток.
Экспрессия (масштабированные значения RPKM) генов, определяющих тип клеток, как указано в (B), во всех отдельных клетках. Транскрипты и отдельные клетки сгруппированы по типу клеток, как указано в (B).
Отображение транскрипционного сходства между всеми отдельными клетками с помощью MDS. Схема окраски основана на типах клеток, указанных в (B).
Относительная экспрессия (масштабированное значение RPKM) канонических маркерных генов для 6 идентифицированных популяций клеток островков поджелудочной железы, представленная размером пузырьков и спроецированная на профиль MDS.
Рисунок EV1. Статистический анализ одноклеточной РНК…
Рисунок EV1. Статистический анализ данных секвенирования РНК одиночных клеток
Статистический анализ данных секвенирования РНК одиночных клеток
Количество обнаруженных транскриптов и общее…
Количество обнаруженных транскриптов и общее количество выровненных прочтений для каждой отдельной клетки. Красная линия обозначает 500 транскриптов, ниже которых образцы исключались из анализа.
Диаграммы рассеяния, отображающие корреляцию между количеством введенных молекул РНК ERCC и измеренными значениями RPKM в четырех репрезентативных одиночных клетках. Корреляция Пирсона ( r ) отмечена в верхнем левом углу.
Графики рассеяния, коррелирующие количество необработанных прочтений (слева) и нормализованные значения экспрессии RPKM (справа) с длиной транскрипта. В этот анализ были включены все адекватно покрытые транскрипты (> 25 прочтений) всех 64 образцов.
 Наблюдаемая отрицательная корреляция после нормализации RPKM лежит в диапазоне того, о чем сообщалось ранее 19.
Наблюдаемая отрицательная корреляция после нормализации RPKM лежит в диапазоне того, о чем сообщалось ранее 19.Необработанные значения подсчета и нормализованные значения экспрессии RPKM для 10 групп контрольной группы ERCC. Количество молекул, включенных в реакцию секвенирования, постоянно внутри одной группы, тогда как длина транскриптов значительно варьирует.Таким образом, в каждой группе одинаковые значения экспрессии для различных длин транскриптов подтверждают, что нормализация RPKM систематически не наказывает более длинные транскрипты в оцененном диапазоне длин транскриптов ERCC от ~200 до 2000 п.н. Контрольные всплески ERCC, охватывающие более 25 прочтений, обозначены синим цветом, а те, у кого ≤ 25 прочтений, — красным.
MDS, отображающий транскрипционное сходство между соответствующими типами клеток опубликованного набора данных 20 (префикс: ext) и текущего набора данных.
Рисунок EV2.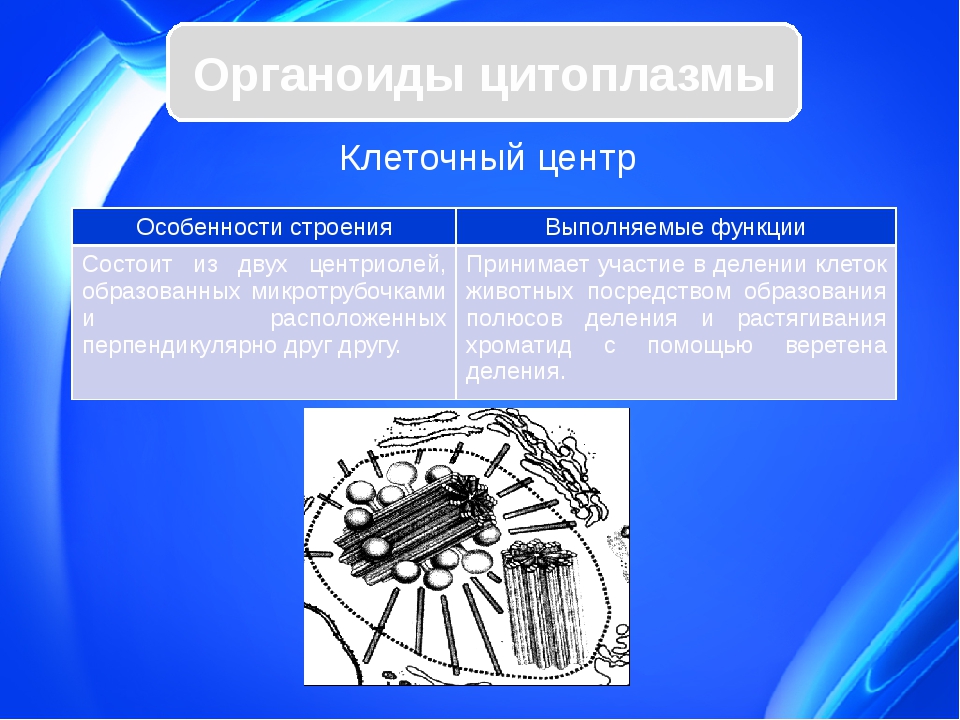 Оценка перекрестного загрязнения между альфа и…
Оценка перекрестного загрязнения между альфа и…
Рисунок EV2. Оценка перекрестного загрязнения между альфа- и бета-клетками
Точечная диаграмма, отображающая одну альфа-канал и…
Точечная диаграмма, показывающая отдельные альфа- и бета-клетки, образцы островков из 500 клеток, а также объемные образцы островков и бета-клеток из опубликованных наборов данных в соответствии с их средневзвешенным значением масштабированных значений экспрессии в специфических для альфа- и бета-клеток генах профиля. Три выбранные ячейки профиля для каждого типа ячеек обозначены их идентификатором образца.
Чистые и смешанные профили экспрессии, состоящие из 233 генов, специфичных для альфа-клеток, и 252 генов, специфичных для бета-клеток.
 Профили, специфичные для альфа- и бета-клеток, рассчитываются на основе значений экспрессии только трех выбранных клеток профиля, в то время как гены профиля были выбраны на основе всех отдельных клеток, классифицированных как альфа- или бета-клетки, поэтому градиенты экспрессии в смешанных профилях не совпадают. не всегда следовать одному и тому же направлению.
Профили, специфичные для альфа- и бета-клеток, рассчитываются на основе значений экспрессии только трех выбранных клеток профиля, в то время как гены профиля были выбраны на основе всех отдельных клеток, классифицированных как альфа- или бета-клетки, поэтому градиенты экспрессии в смешанных профилях не совпадают. не всегда следовать одному и тому же направлению.Кривые корреляции профиля для каждого отдельного образца. Максимум каждой кривой определяет максимальную дисперсию, которая может быть объяснена (ось y ) соответствующим профилем смеси (ось x ), обеспечивающим меру состава соответствующей выборки.
Диаграмма, поясняющая переход от кривых корреляции профилей к оценкам состава выборки. Состав профиля, который объясняет большую часть дисперсии, линейно масштабируется до максимальной объясненной дисперсии.
Оценки состава выборки для каждой оцениваемой выборки. Различия между образцами 500-клеточных островков и объемными образцами можно объяснить техническими эффектами, которые обогащают альфа-клетки во время культивирования островков, диссоциации и очистки FACS.

Рисунок 2. Экспрессия клеточной транскрипции…
Рисунок 2. Экспрессия факторов транскрипции, специфичных для типа клеток, при разрешении одной клетки
Фигура 2.Экспрессия факторов транскрипции, специфичных для типа клеток, при разрешении одной клетки- A
Объединенные треки UCSC Genome Browser для локусов PDX1 и ARX . Соответствующие дорожки для всех одиночных ячеек представлены на рис. S2 в Приложении.
- B
Относительная экспрессия (масштабированное значение RPKM) важных факторов транскрипции, представленная размером пузырьков и спроецированная на профиль MDS.
- C–F
Типоспецифичная экспрессия панэндокринных (C), альфа-клеточных (D), бета-клеточных (E) и РР-клеточных (F) факторов транскрипции (красная полоса: средняя экспрессия).Статистическая значимость экспрессии дифференциального гена представлена на рис. S6 в Приложении.
Рисунок EV3. Оценка парной корреляции эндокринной…
Рисунок EV3. Оценка парной корреляции эндокринных маркерных генов
Корреляционная матрица, отображающая все гены (…
Рисунок EV3.Оценка парной корреляции эндокринных маркерных генов Корреляционная матрица, отображающая все гены (ось y ), которые высоко коррелируют ( r > 0,9) по крайней мере с одним из эндокринных маркерных генов (ось x ). Различные транскрипты одного и того же гена обозначены «Tx».
Различные транскрипты одного и того же гена обозначены «Tx».
Рис. 3.Транскриптомы одиночных клеток обнаруживают уникальные особенности…
Рисунок 3. Транскриптомы одиночных клеток обнаруживают уникальные особенности островков человека
- A
Тепловая карта, отображающая P…
- A
Тепловая карта, отображающая P -значения, полученные с помощью парного анализа обогащения набора генов (GSEA) для REST-связывающего мотива.
- B
Относительная экспрессия (масштабированное значение RPKM) генов, содержащихся в наборе генов REST-связывающего мотива.

- C
Объединенные треки браузера генома UCSC для REST . Соответствующие дорожки для всех одиночных ячеек представлены на рис. S3 в Приложении.
- D
Экспрессия DLK1 и GC в островковых клетках человека (красная полоса: средняя экспрессия). Статистическая значимость экспрессии дифференциального гена представлена на рис. S6 в Приложении.
- E–H
Совместное окрашивание DLK1 (E, F) или GC (G, H) инсулином и глюкагоном в репрезентативных островках человека (E, G) и мыши (F, H).
Рисунок EV4. Конкретное выражение выбранного маркера…
Рисунок EV4. Специфическая экспрессия выбранных маркерных генов
Относительное выражение (масштабированное значение RPKM) интересного…
Рисунок EV4. Специфическая экспрессия выбранных маркерных генов
Специфическая экспрессия выбранных маркерных генов Относительная экспрессия (масштабированное значение RPKM) интересующих генов во всех отдельных клетках, представленная размером пузырьков и спроецированная на профиль MDS, как показано на рис. 1D.
Рисунок EV5. Оценка специфичности типов клеток…
Рисунок EV5.Оценка специфичности типов клеток генов, идентифицированных в GWAS
, связанном с диабетом Рисунок EV5. Оценка специфичности клеточного типа генов, идентифицированных в GWAS , связанном с диабетомСпецифичность типа клеток для генов, зарегистрированных в GWAS, связанном с диабетом.
 Каждый ген, указанный в GWAS, связанном с диабетом (поиск «Диабет» в каталоге GWAS), был отнесен к типу клеток поджелудочной железы, в котором он экспрессировался наиболее специфически.Ранжирование проводилось, как описано в Приложении, рис. S7B и наборе данных EV6.
Каждый ген, указанный в GWAS, связанном с диабетом (поиск «Диабет» в каталоге GWAS), был отнесен к типу клеток поджелудочной железы, в котором он экспрессировался наиболее специфически.Ранжирование проводилось, как описано в Приложении, рис. S7B и наборе данных EV6.Тепловая карта, показывающая средние значения экспрессии для наиболее специфичных для типа клеток генов GWAS, связанных с диабетом, в различных идентифицированных здесь типах островковых клеток человека. В частности, перечислены только гены с рангом специфичности ниже 500 (пунктирная линия на панели А), а гены с одинаковой экспрессией во многих типах клеток не показаны. Цифры в цветных прямоугольниках обозначают количество исследований, в которых сообщалось о соответствующем гене.Тепловые карты окрашены средним значением ln(RPKM), средним значением натурального логарифма значений RPKM по всем ячейкам соответствующего типа ячеек.
Все фигурки (8)
Похожие статьи
- Восстановление жизнеспособных эндокринно-специфических клеток и транскриптомов у мышей с трансплантированными островками поджелудочной железы человека.
Редик С.Д., Лихи Л., Риттенхаус А.Р., Блоджетт Д.М., Дерр А.Г., Кучукурал А., Гарбер М.Г., Шульц Л.Д., Грейнер Д.Л., Ван Дж.П., Харлан Д.М., Бортелл Р., Юрчик А. Редик С.Д. и соавт. FASEB J. 2020 Jan; 34(1):1901-1911. doi: 10.1096/fj.201
2RR. Epub 2019 10 декабря. ФАСЭБ Дж. 2020. PMID: 31914605 Бесплатная статья ЧВК.
- Опрос островков в норме и болезни с помощью одноклеточных технологий.
Каррано А.С., Мулас Ф., Зенг С., Сандер М. Каррано А.С. и соавт. Мол метаб. 2017 5 мая; 6 (9): 991-1001. doi: 10.1016/j.molmet.2017.04.012. Электронная коллекция 2017 Сентябрь. Мол метаб. 2017. PMID: 28951823 Бесплатная статья ЧВК. Обзор.
- Анализ транскриптомов клеток поджелудочной железы у отдаленных видов выдвигает на первый план новые важные гены-регуляторы.

Тарифеньо-Сальдивия Э., Лавернь А., Бернар А., Падамата К., Бергеманн Д., Воз М.Л., Манфроид И., Пирс Б.Тарифеньо-Сальдивия Э. и др. БМС Биол. 2017 21 марта; 15 (1): 21. doi: 10.1186/s12915-017-0362-x. БМС Биол. 2017. PMID: 28327131 Бесплатная статья ЧВК.
- Одноклеточная РНК-Seq островков поджелудочной железы – еще не выполненное обещание?
Ван Ю.Дж., Кестнер К.Х. Ван Ю.Дж. и др. Клеточный метаб. 2019 5 марта; 29 (3): 539-544. doi: 10.1016/j.cmet.2018.11.016. Epub 2018 20 декабря.Клеточный метаб. 2019. PMID: 30581120 Бесплатная статья ЧВК. Обзор.
- Транскриптомы одиночных клеток идентифицируют сигнатуры островковых клеток человека и выявляют специфичные для типа клеток изменения экспрессии при диабете 2 типа.

Лоулор Н., Джордж Дж., Болисетти М., Курсаве Р., Сан Л., Сивакамасундари В., Кисия И., Робсон П., Ститцель М.Л. Лоулор Н. и др. Геном Res. 2017 фев; 27 (2): 208-222.doi: 10.1101/гр.212720.116. Epub 2016 18 ноября. Геном Res. 2017. PMID: 27864352 Бесплатная статья ЧВК.
Цитируется
100 статей- Аргиназа 2 и полиамины в бета-клетках поджелудочной железы человека: возможная роль в патогенезе диабета 2 типа.
Марселли Л., Боси Э., Де Лука С., Дель Герра С., Тези М., Сулейман М., Маркетти П.Марселли Л. и соавт. Int J Mol Sci. 2021 9 ноября; 22(22):12099. дои: 10.3390/ijms222212099. Int J Mol Sci. 2021. PMID: 34829980 Бесплатная статья ЧВК. Обзор.
- Партнеры в преступлении: бета-клетки и аутоиммунные реакции, сопричастные к патогенезу диабета 1 типа.

Торен Э., Бернетт К.С., Банерджи Р.Р., Хантер К.С., Це Х.М. Торен Э. и др. Фронт Иммунол. 2021 7 окт;12:756548.doi: 10.3389/fimmu.2021.756548. Электронная коллекция 2021. Фронт Иммунол. 2021. PMID: 34691077 Бесплатная статья ЧВК. Обзор.
- Применение одноклеточной геномики и вычислительных стратегий для изучения распространенных заболеваний и вариаций на уровне популяции.
Ауэрбах Б.Дж., Ху Дж., Рейли М.П., Ли М. Ауэрбах Б.Дж. и соавт. Геном Res. 2021 Окт;31(10):1728-1741. doi: 10.1101/гр.275430.121. Геном Res. 2021. PMID: 34599006 Бесплатная статья ЧВК.
- Генные сигнатуры эндокринных клеток-предшественников NEUROGENIN3+ в поджелудочной железе человека.
Yong HJ, Xie G, Liu C, Wang W, Naji A, Irianto J, Wang YJ.
 Йонг Х.Дж. и др.
Фронт Эндокринол (Лозанна). 2021 8 сент.; 12:736286. doi: 10.3389/fendo.2021.736286. Электронная коллекция 2021.
Фронт Эндокринол (Лозанна).2021.
PMID: 34566896
Бесплатная статья ЧВК.
Йонг Х.Дж. и др.
Фронт Эндокринол (Лозанна). 2021 8 сент.; 12:736286. doi: 10.3389/fendo.2021.736286. Электронная коллекция 2021.
Фронт Эндокринол (Лозанна).2021.
PMID: 34566896
Бесплатная статья ЧВК. - Анализ гетерогенности отдельных клеток и скрининг CRISPR в клеточной линии MIN6 выявили регуляторы транскрипции инсулина.
Чжао Р., Лу Дж., Ли К., Сюн Ф., Чжан Й., Чжу Дж., Пэн Г., Ян Дж. Чжао Р. и др. Клеточный цикл. 2021 Окт;20(19):2053-2065. дои: 10.1080/15384101.2021.1969204. Epub 2021 8 сентября. Клеточный цикл. 2021.PMID: 34494921
использованная литература
- Collombat P, Xu X, Ravassard P, Sosa-Pineda B, Dussaud S, Billestrup N, Madsen OD, Serup P, Heimberg H, Mansouri A (2009)Эктопическая экспрессия Pax4 в поджелудочной железе мыши превращает клетки-предшественники в альфа, а затем бета клетки.
 Ячейка 138: 449–462.
—
ЧВК
—
пабмед
Ячейка 138: 449–462.
—
ЧВК
—
пабмед
- Collombat P, Xu X, Ravassard P, Sosa-Pineda B, Dussaud S, Billestrup N, Madsen OD, Serup P, Heimberg H, Mansouri A (2009)Эктопическая экспрессия Pax4 в поджелудочной железе мыши превращает клетки-предшественники в альфа, а затем бета клетки.
- Chera S, Baronnier D, Ghila L, Cigliola V, Jensen JN, Gu G, Furuyama K, Thorel F, Gribble FM, Reimann F et al (2014)Выздоровление от диабета путем возрастного превращения дельта-клеток поджелудочной железы в продуценты инсулина.Природа 514: 503–507. — ЧВК — пабмед
- Fadista J, Vikman P, Laakso EO, Mollet IG, Esguerra JL, Taneera J, Storm P, Osmark P, Ladenvall C, Prasad RB et al (2014)Глобальный геномный и транскриптомный анализ островков поджелудочной железы человека выявляет новые гены, влияющие на метаболизм глюкозы.
 Proc Natl Acad Sci USA 111: 13924–13929
—
ЧВК
—
пабмед
Proc Natl Acad Sci USA 111: 13924–13929
—
ЧВК
—
пабмед
- Fadista J, Vikman P, Laakso EO, Mollet IG, Esguerra JL, Taneera J, Storm P, Osmark P, Ladenvall C, Prasad RB et al (2014)Глобальный геномный и транскриптомный анализ островков поджелудочной железы человека выявляет новые гены, влияющие на метаболизм глюкозы.
- Bramswig NC, Everett LJ, Schug J, Dorrell C, Liu C, Luo Y, Streeter PR, Naji A, Grompe M, Kaestner KH (2013)Эпигеномная пластичность позволяет человеку перепрограммировать альфа-клетки поджелудочной железы в бета-клетки.Дж. Клин Инвест 123: 1275–1284. — ЧВК — пабмед
- Kameswaran V, Bramswig NC, McKenna LB, Penn M, Schug J, Hand NJ, Chen Y, Choi I, Vourekas A, Won KJ et al (2014)Эпигенетическая регуляция кластера микроРНК DLK1-MEG3 в диабетических островках человека 2 типа.
 Cell Metab 19: 135–145
—
ЧВК
—
пабмед
Cell Metab 19: 135–145
—
ЧВК
—
пабмед
- Kameswaran V, Bramswig NC, McKenna LB, Penn M, Schug J, Hand NJ, Chen Y, Choi I, Vourekas A, Won KJ et al (2014)Эпигенетическая регуляция кластера микроРНК DLK1-MEG3 в диабетических островках человека 2 типа.
Показать все 34 ссылки
Типы публикаций
- Поддержка исследований, не-U.С. Правительство
термины MeSH
- Профилирование экспрессии генов / методы
- Профилирование экспрессии генов / стандарты
- Инсулинсекретирующие клетки / классификация
- Инсулинсекретирующие клетки/метаболизм*
- Одноклеточный анализ / методы
- Одноклеточный анализ / стандарты
LinkOut — больше ресурсов
Полнотекстовые источники
Прочие литературные источники
Укажите
КопироватьФормат: ААД АПА МДА НЛМ
Что такое серповидноклеточная анемия?
ВСС представляет собой группу наследственных заболеваний эритроцитов.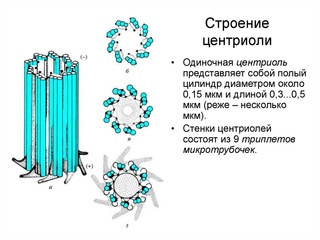 Здоровые эритроциты имеют круглую форму и перемещаются по мелким кровеносным сосудам, доставляя кислород ко всем частям тела. У человека с ВСС эритроциты становятся твердыми и липкими и напоминают С-образный сельскохозяйственный инструмент, называемый «серпом». Серповидные клетки рано погибают, что вызывает постоянную нехватку эритроцитов. Кроме того, когда они проходят через мелкие кровеносные сосуды, они застревают и блокируют кровоток. Это может вызвать боль и другие серьезные проблемы, такие как инфекция, острый грудной синдром и инсульт.
Здоровые эритроциты имеют круглую форму и перемещаются по мелким кровеносным сосудам, доставляя кислород ко всем частям тела. У человека с ВСС эритроциты становятся твердыми и липкими и напоминают С-образный сельскохозяйственный инструмент, называемый «серпом». Серповидные клетки рано погибают, что вызывает постоянную нехватку эритроцитов. Кроме того, когда они проходят через мелкие кровеносные сосуды, они застревают и блокируют кровоток. Это может вызвать боль и другие серьезные проблемы, такие как инфекция, острый грудной синдром и инсульт.
Типы SCD
Ниже приведены наиболее распространенные типы SCD:
ХбСС
Люди с этой формой ВСС наследуют два гена серповидно-клеточной анемии («S»), по одному от каждого родителя. Это обычно называется серповидно-клеточной анемией и обычно является наиболее тяжелой формой заболевания.
HbSC
Люди с этой формой ВСС наследуют ген серповидно-клеточной анемии («S») от одного родителя, а от другого родителя — ген аномального гемоглобина, называемый «С». Гемоглобин — это белок, который позволяет эритроцитам переносить кислород ко всем частям тела. Обычно это более легкая форма ВСС.
Гемоглобин — это белок, который позволяет эритроцитам переносить кислород ко всем частям тела. Обычно это более легкая форма ВСС.
HbS-бета-талассемия
Люди с этой формой ВСС наследуют один ген серповидно-клеточной анемии («S») от одного родителя и один ген бета-талассемии, другого типа анемии, от другого родителя. Различают два типа бета-талассемии: «0» и «+». Те, у кого HbS бета-талассемия, обычно имеют тяжелую форму ВСС. Люди с HbS бета +-талассемией, как правило, имеют более легкую форму ВСС.
Также есть несколько редких типов SCD:
HbSD, HbSE и HbSO
Люди с этими формами ВСС наследуют один ген серповидно-клеточной анемии («S») и один ген аномального типа гемоглобина («D», «E» или «O»). Гемоглобин — это белок, который позволяет эритроцитам переносить кислород ко всем частям тела. Тяжесть этих более редких типов ВСС варьируется.
Признак серповидноклеточной анемии (SCT)
ХБАС
Люди с SCT наследуют один ген серповидно-клеточной анемии («S») от одного родителя и один нормальный ген («A») от другого родителя. Это называется серповидноклеточным признаком (SCT). Люди с SCT обычно не имеют каких-либо признаков заболевания и живут нормальной жизнью, но они могут передать эту черту своим детям. Кроме того, есть несколько необычных проблем со здоровьем, которые потенциально могут быть связаны с чертой серповидно-клеточной анемии.
Это называется серповидноклеточным признаком (SCT). Люди с SCT обычно не имеют каких-либо признаков заболевания и живут нормальной жизнью, но они могут передать эту черту своим детям. Кроме того, есть несколько необычных проблем со здоровьем, которые потенциально могут быть связаны с чертой серповидно-клеточной анемии.
Узнайте больше о признаке серповидно-клеточной анемии »
Причина ВСС
SCD — это генетическое заболевание, присутствующее при рождении. Он передается по наследству, когда ребенок получает два гена серповидно-клеточной анемии — по одному от каждого родителя.
Диагностика
SCD диагностируется с помощью простого анализа крови. Чаще всего его обнаруживают при рождении во время рутинных скрининговых тестов новорожденных в больнице. Кроме того, ВСС можно диагностировать еще до рождения.
Поскольку дети с ВСС подвержены повышенному риску заражения и других проблем со здоровьем, важны ранняя диагностика и лечение.
Вы можете позвонить в местную организацию серповидно-клеточной анемии, чтобы узнать, как пройти тестирование.
Осложнения и лечение
У людей с ВСС первые признаки заболевания появляются в течение первого года жизни, обычно в возрасте около 5 месяцев.Симптомы и осложнения ВСС различны для каждого человека и могут варьироваться от легких до тяжелых.
Не существует единственного наилучшего лечения для всех людей с ВСС. Варианты лечения различны для каждого человека в зависимости от симптомов.
Узнайте об осложнениях и лечении »
Лечение
Единственным лекарством от ВСС является трансплантация костного мозга или стволовых клеток.
Костный мозг представляет собой мягкую жировую ткань в центре костей, где образуются клетки крови.Трансплантация костного мозга или стволовых клеток — это процедура, при которой здоровые клетки, образующие кровь, берутся у одного человека — донора — и пересаживаются человеку, чей костный мозг не работает должным образом.
Пересадка костного мозга или стволовых клеток очень опасна и может иметь серьезные побочные эффекты, включая смерть. Чтобы трансплантат сработал, костный мозг должен быть близким. Обычно лучшим донором является брат или сестра. Трансплантация костного мозга или стволовых клеток используется только в случаях тяжелой ВСС у детей с минимальным поражением органов от заболевания.
Клинические характеристики и исходы экстранодальной диффузной крупноклеточной В-клеточной лимфомы I стадии в эпоху ритуксимаба | Кровь
Добавил: С.Б. и А.Ю. задумал проект, проанализировал данные и написал рукопись; J.A.L. и В.Е.С. проанализировал данные, написал рукопись и предоставил цифры; и E.J., C.L.B., P.C.C., A.H., P.A.H., S.M.H., A.K., MJM, A.M., A.N., D.S., P.G., C.N.O., M.L.P., D.S., G.v.K., A.D.Z., J.Y. и A.Д. лечил пациентов и одобрил рукопись.
Раскрытие информации о конфликте интересов: J.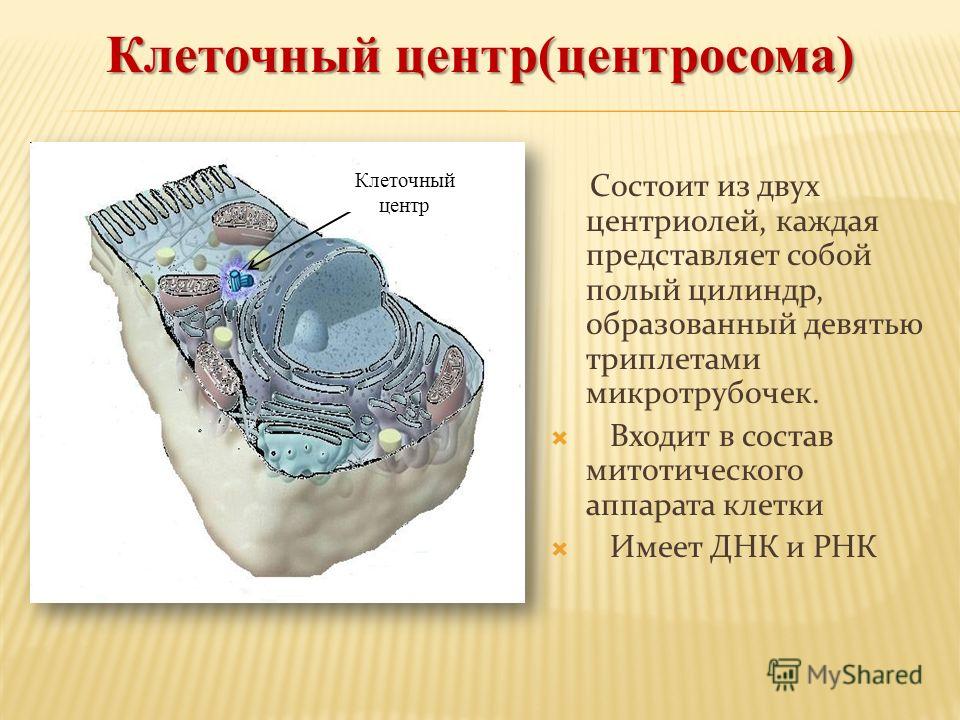 A.L. сообщает о поддержке заработной платы для проекта обмена информацией о неоплазии доказательств геномики (GENIE) через Американскую ассоциацию исследований рака. AD получила персональные гонорары от Roche, Corvus Pharmaceuticals, Physicians’ Education Resource, Seattle Genetics, Peerview Institute, Oncology Speciality Group, Pharmacyclos, Celgene, Novartis и Takeda, а также исследовательские гранты от Национального института рака и Roche.C.L.B. получил грантовое финансирование от Janssen, Novartis, Epizyme, Xynomics, Bayer и BMS; работал консультантом в Life Sci, GLG, Celgene, Seattle Genetics и Xynomics; и получает гонорар от Dava Oncology. П. Х. получает исследовательскую поддержку от Portola, Novartis/GSK, Molecular Templates и Janssen Pharmaceuticals и выступал в качестве консультанта для Karyopharm, Juno, Portola, Celgene и AstraZeneca. С.М.Х. получил финансирование исследований от ADCT Therapeutics, Aileron, Forty-Seven, Verastem, Kyowa Hakko Kirin, Millennium Pharmaceuticals Inc.
A.L. сообщает о поддержке заработной платы для проекта обмена информацией о неоплазии доказательств геномики (GENIE) через Американскую ассоциацию исследований рака. AD получила персональные гонорары от Roche, Corvus Pharmaceuticals, Physicians’ Education Resource, Seattle Genetics, Peerview Institute, Oncology Speciality Group, Pharmacyclos, Celgene, Novartis и Takeda, а также исследовательские гранты от Национального института рака и Roche.C.L.B. получил грантовое финансирование от Janssen, Novartis, Epizyme, Xynomics, Bayer и BMS; работал консультантом в Life Sci, GLG, Celgene, Seattle Genetics и Xynomics; и получает гонорар от Dava Oncology. П. Х. получает исследовательскую поддержку от Portola, Novartis/GSK, Molecular Templates и Janssen Pharmaceuticals и выступал в качестве консультанта для Karyopharm, Juno, Portola, Celgene и AstraZeneca. С.М.Х. получил финансирование исследований от ADCT Therapeutics, Aileron, Forty-Seven, Verastem, Kyowa Hakko Kirin, Millennium Pharmaceuticals Inc.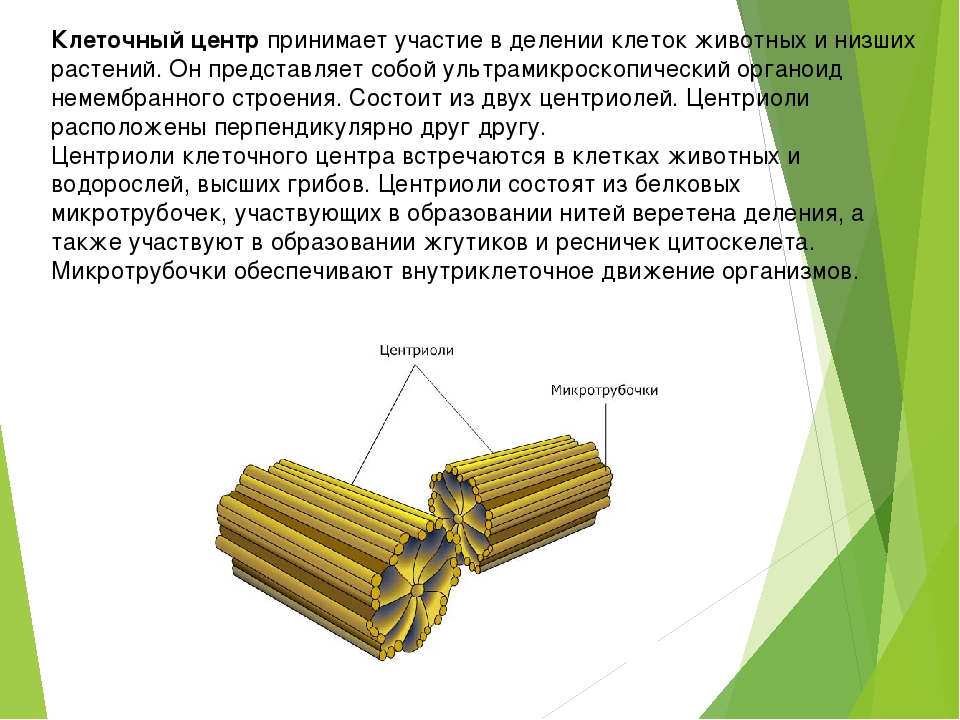 , Celgene, Trillium и Daiichii Sankyo, а также консультирует Astex, Affimed, Merck Sharp and Dome, Kyowa Hakko Kirin Pharma, Corvus Pharmaceuticals Inc., Celgene, Portola Pharmaceuticals, Takeda Millennium, Innate Pharma, Verastem, Miragen Therapeutics Inc., Seattle Genetics. и АДКТ. А.К. получает финансирование исследований от компаний AbbVie Pharmaceuticals, Adaptive Biotechnologies, Pharmacyclos и Seattle Genetics и входит в состав консультативного совета Celgene и Astra Zeneca. М.Л.П. получает гонорары от Flagship Ventures, Novartis, Evelo, Seres Therapeutics, Jazz Pharmaceuticals, Therakos, Amgen и Merck, а также консультирует Merck и Pharmacyclos.А.Н. получает гонорары от компаний «Янссен», «Фармацикликс» и «Прайм Онкология»; консультирует Medscape; входит в консультативный совет Janssen; работает в бюро докладчиков Prime Oncology; и получает финансирование исследований для Rafael Pharma и Pharmacyclos. ЯВЛЯЮСЬ. получает исследовательскую поддержку от Seattle Genetics, Merck, Bristol-Myers Squibb и Incyte и получает гонорар от Kyowa Hakko Kirin Pharma, Miragen Therapeutics, Takeda Pharmaceuticals, ADC Therapeutics, Seattle Genetics, Cell Medica, Bristol-Myers Squibb и Erytech Pharma.
, Celgene, Trillium и Daiichii Sankyo, а также консультирует Astex, Affimed, Merck Sharp and Dome, Kyowa Hakko Kirin Pharma, Corvus Pharmaceuticals Inc., Celgene, Portola Pharmaceuticals, Takeda Millennium, Innate Pharma, Verastem, Miragen Therapeutics Inc., Seattle Genetics. и АДКТ. А.К. получает финансирование исследований от компаний AbbVie Pharmaceuticals, Adaptive Biotechnologies, Pharmacyclos и Seattle Genetics и входит в состав консультативного совета Celgene и Astra Zeneca. М.Л.П. получает гонорары от Flagship Ventures, Novartis, Evelo, Seres Therapeutics, Jazz Pharmaceuticals, Therakos, Amgen и Merck, а также консультирует Merck и Pharmacyclos.А.Н. получает гонорары от компаний «Янссен», «Фармацикликс» и «Прайм Онкология»; консультирует Medscape; входит в консультативный совет Janssen; работает в бюро докладчиков Prime Oncology; и получает финансирование исследований для Rafael Pharma и Pharmacyclos. ЯВЛЯЮСЬ. получает исследовательскую поддержку от Seattle Genetics, Merck, Bristol-Myers Squibb и Incyte и получает гонорар от Kyowa Hakko Kirin Pharma, Miragen Therapeutics, Takeda Pharmaceuticals, ADC Therapeutics, Seattle Genetics, Cell Medica, Bristol-Myers Squibb и Erytech Pharma. Д.С. консультирует InPractice Elsevier и Seattle Genetics, а также входит в бюро докладчиков Medical Crossfire. А.Д.З. консультирует Genentech/Roche, Gilead, Celgene, Janssen, Amgen, Novartis, Adaptive Biotechnology и Verastem; входит в состав консультативного совета MorphoSys, Gilead, Genentech, AbbVie, AstraZeneca, Pharmacyclos; и получает финансирование исследований от MEI Pharmaceuticals, Roche, Gilead и Beigene. А.Ю. получает исследовательскую поддержку от Janssen, Curis, Merck, BMS, Syndax и Roche; получает гонорары от Janssen, AbbVie, Merck, Curis, Epizyme, Roche и Takeda; и консультирует Biopath, Xynomics, Epizyme, Roche, Celgene и HCM.Остальные авторы заявляют об отсутствии конкурирующих финансовых интересов.
Д.С. консультирует InPractice Elsevier и Seattle Genetics, а также входит в бюро докладчиков Medical Crossfire. А.Д.З. консультирует Genentech/Roche, Gilead, Celgene, Janssen, Amgen, Novartis, Adaptive Biotechnology и Verastem; входит в состав консультативного совета MorphoSys, Gilead, Genentech, AbbVie, AstraZeneca, Pharmacyclos; и получает финансирование исследований от MEI Pharmaceuticals, Roche, Gilead и Beigene. А.Ю. получает исследовательскую поддержку от Janssen, Curis, Merck, BMS, Syndax и Roche; получает гонорары от Janssen, AbbVie, Merck, Curis, Epizyme, Roche и Takeda; и консультирует Biopath, Xynomics, Epizyme, Roche, Celgene и HCM.Остальные авторы заявляют об отсутствии конкурирующих финансовых интересов.
Корреспонденция: Анас Юнес, Мемориальный онкологический центр имени Слоана Кеттеринга, 1275 York Ave, Box 330, New York, NY 10065; электронная почта: [email protected].
Синдромы ортостатической непереносимости после трансплантации гемопоэтических клеток: клиническая характеристика и терапевтические вмешательства в условиях моноцентра | Кардиоонкология
Синдромы ортостатической непереносимости, такие как ОГ и СПОТ, встречаются как у населения в целом, так и у пациентов с конкретными заболеваниями, в том числе у перенесших трансплантацию органов [7, 8, 12]. Синдромы ортостатической непереносимости связаны с повышенным риском сердечно-сосудистых заболеваний и более высокой смертностью [6, 12] и снижением качества жизни, что подчеркивает необходимость лучшего распознавания. В последние годы вегетативная дисфункция стала признанным сердечно-сосудистым расстройством, поражающим пациентов с раком и выживших после рака в большей степени, чем здоровые контроли [13].
Синдромы ортостатической непереносимости связаны с повышенным риском сердечно-сосудистых заболеваний и более высокой смертностью [6, 12] и снижением качества жизни, что подчеркивает необходимость лучшего распознавания. В последние годы вегетативная дисфункция стала признанным сердечно-сосудистым расстройством, поражающим пациентов с раком и выживших после рака в большей степени, чем здоровые контроли [13].
В текущем исследовании мы впервые описали распространенность ОГ и СПОТ у пациентов, направленных в кардиоонкологическую клинику после ТГСК по поводу нескольких гематологических заболеваний.ОГ возникала у 23% пациентов, СПОТ — у 9% после ТГСК. В этой когорте пациентов развитие ОГ или СПОТ не было связано с факторами риска, ранее описанными в других популяциях, такими как возраст, пол и прием антигипертензивных препаратов. В то время как лучевая терапия и использование специфических противоопухолевых схем могут быть факторами риска развития ОГ и СПОТ, возникновение этих синдромов, скорее всего, связано с процедурой трансплантации самой по себе, учитывая, что у пациентов не было диагностировано это до ТГСК и ни один из этих факторов риска не был значимым в многомерном анализе.
Распространенность ОГ, сходная с таковой у нашей популяции, ранее была описана Schuurmans et al. в небольшой группе реципиентов трансплантата легкого [8]. В другом недавнем исследовании ОГ возникала у 28% пациентов с трансплантацией почки и поджелудочной железы [7]. История периферической невропатии, связанной с длительно текущим сахарным диабетом, не коррелировала с развитием АГ в этой группе больных [7]. Гиперинсулинемия после трансплантации и сосудорасширяющие нейропептиды, высвобождаемые из поджелудочной железы, были описаны как возможные факторы, ответственные за развитие ОГ, но биологическая причина остается неясной.
Бергер и др. сообщили о более высоком присутствии дизавтономии в группе детей, перенесших рак, по сравнению со здоровыми контрольными группами [14]. Между двумя группами не было обнаружено различий в факторах, которые могут вызывать дизавтономию, но авторы не смогли изучить возможную связь с лечением рака из-за небольшого размера выборки [14]. Недавно в исследовании Deuring et al. вегетативная функция сердца была измерена в группе пациентов, перенесших HCT, и в группе здоровых людей, соответствующих контрольной группе, путем оценки ЧСС и респираторной синусовой аритмии (RSA) [15].Реципиенты трансплантата показали более высокую ЧСС и более низкую ОСА по сравнению с контрольной группой, что указывает на наличие сердечной вегетативной дисфункции [15].
вегетативная функция сердца была измерена в группе пациентов, перенесших HCT, и в группе здоровых людей, соответствующих контрольной группе, путем оценки ЧСС и респираторной синусовой аритмии (RSA) [15].Реципиенты трансплантата показали более высокую ЧСС и более низкую ОСА по сравнению с контрольной группой, что указывает на наличие сердечной вегетативной дисфункции [15].
В общей популяции заболеваемость ОГ экспоненциально увеличивается с возрастом, что не является неожиданным, поскольку некоторые факторы риска, связанные с развитием ОГ, такие как невропатия, сахарный диабет, болезни сердца, болезнь Паркинсона и потребность в политерапии, чаще встречаются у пожилые [16, 17]. POTS также часто диагностируется у подростков, особенно в течение двух лет с начала полового созревания, но часто имеет более доброкачественное течение и часто самокупируется [18].POTS в два-три раза чаще встречается у девочек-подростков по сравнению с мужчинами, и почти 90% взрослых пациентов с POTS — женщины [18], при этом не наблюдается явного предпочтения женского пола у пожилых людей. В нашей когорте мы наблюдали более высокую частоту POTS и OH у мужчин по сравнению с женщинами. Это было неожиданно и нелегко объяснимо. Необходимы дополнительные исследования, чтобы изучить эту очевидную разницу в зависимости от пола.
В нашей когорте мы наблюдали более высокую частоту POTS и OH у мужчин по сравнению с женщинами. Это было неожиданно и нелегко объяснимо. Необходимы дополнительные исследования, чтобы изучить эту очевидную разницу в зависимости от пола.
Лечение антигипертензивными препаратами является общепризнанным фактором риска ОГ и СПОТ [19].В этом ретроспективном анализе мы не обнаружили статистически значимых различий по возрасту, полу, применению антигипертензивных препаратов, между пациентами с ОГ или СПОТ и без них. Мы наблюдали связь между лучевой терапией и некоторыми классами химиотерапевтических агентов и развитием дизавтономии. Однако ни один из этих методов лечения не сохранил статистическую значимость в многомерной модели, предполагая, что другие, еще не идентифицированные факторы могут способствовать заболеваемости ОГ и СПОТ после трансплантации.Более того, пациентов с гематологическими злокачественными новообразованиями часто лечат несколькими химиотерапевтическими препаратами перед ПХТ, что усложняет оценку эффекта одного препарата. Предыдущее исследование пациентов с раком молочной железы выявило связь между вегетативной дисфункцией сердца и антрациклинами [20], подтверждая потенциальную роль этого класса препаратов в развитии ОГ и СПОТ. Лучевая терапия головы и шеи была связана со снижением значений АД после лечения, но ОГ не было зарегистрировано [21].В другой когорте из 282 выживших после рака, направленных на кардиоонкологическую программу, у 22 человек была диагностирована вегетативная дисфункция [22]. Из них у большинства были гематологические нарушения, а одной трети была проведена ТГСК [22]. У испытуемых было выявлено нарушение многих компонентов вегетативной функции (судомоторной, адренергической и сердечно-сосудистой) [22] [22]..
Предыдущее исследование пациентов с раком молочной железы выявило связь между вегетативной дисфункцией сердца и антрациклинами [20], подтверждая потенциальную роль этого класса препаратов в развитии ОГ и СПОТ. Лучевая терапия головы и шеи была связана со снижением значений АД после лечения, но ОГ не было зарегистрировано [21].В другой когорте из 282 выживших после рака, направленных на кардиоонкологическую программу, у 22 человек была диагностирована вегетативная дисфункция [22]. Из них у большинства были гематологические нарушения, а одной трети была проведена ТГСК [22]. У испытуемых было выявлено нарушение многих компонентов вегетативной функции (судомоторной, адренергической и сердечно-сосудистой) [22] [22]..
Учитывая своевременную связь с HCT, возможно, что сама HCT является ведущей причиной OH и POTS в этой группе больных.Действительно, трансплантация приводит к значительному бремени воспаления, которое может быть триггером развития ОГ и СПОТ [23]. Среднее время от HCT до постановки диагноза OH или POTS составило 82 (24–248) дней. Необходимы дальнейшие исследования для лучшего изучения причин ОГ и СПОТ после ТГСК.
Необходимы дальнейшие исследования для лучшего изучения причин ОГ и СПОТ после ТГСК.
ОГ связана с суточной вариабельностью АД и, часто, с гипертензией в положении лежа, что способствует интермиттирующему увеличению постнагрузки, способствуя развитию поражения органов-мишеней [5]. Это наблюдение было подтверждено и в нашей когорте, в которой у 1/3 пациентов, у которых развилась ОГ после ТГСК, также наблюдалась артериальная гипертензия в положении лежа.
Лечение ОГ и СПОТ в основном определяется симптомами, а немедикаментозное лечение обычно считается терапией первой линии [5, 24]. Однако одних немедикаментозных методов лечения в тяжелых случаях может быть недостаточно. Одним из ограничений этого анализа является невозможность зафиксировать использование немедикаментозных методов лечения с точки зрения назначений и соблюдения. Большинству (91%) пациентов в нашей когорте требовалось назначение лекарств для лечения ОГ и СПОТ. Мидодрин, селективный и прямой агонист а1-адренорецепторов, показал свою эффективность в уменьшении симптомов и улучшении качества жизни [25]. Флудрокортизон является эффективным средством для увеличения объема, но его применение противопоказано пациентам с сердечной недостаточностью, почечной недостаточностью или артериальной гипертензией в положении лежа [26]. Сообщалось, что при СПОТ β-адреноблокаторы улучшают симптомы [27, 28]. Однако не было доказано, что ни один из этих методов лечения не превосходит другие по снижению исходов, и ни один из этих методов лечения специально не изучался у пациентов после ТГСК. По нашему опыту, мидодрин был наиболее часто используемым фармакологическим средством при ОГ, с добавлением флудрокортизона у пациентов, не ответивших на лечение, а β-адреноблокаторы использовались как у пациентов с ОГ, так и у пациентов с СПОТ.Средняя продолжительность лечения составила 120, хотя треть пациентов продолжала лечение в течение длительного времени.
Флудрокортизон является эффективным средством для увеличения объема, но его применение противопоказано пациентам с сердечной недостаточностью, почечной недостаточностью или артериальной гипертензией в положении лежа [26]. Сообщалось, что при СПОТ β-адреноблокаторы улучшают симптомы [27, 28]. Однако не было доказано, что ни один из этих методов лечения не превосходит другие по снижению исходов, и ни один из этих методов лечения специально не изучался у пациентов после ТГСК. По нашему опыту, мидодрин был наиболее часто используемым фармакологическим средством при ОГ, с добавлением флудрокортизона у пациентов, не ответивших на лечение, а β-адреноблокаторы использовались как у пациентов с ОГ, так и у пациентов с СПОТ.Средняя продолжительность лечения составила 120, хотя треть пациентов продолжала лечение в течение длительного времени.
Выжившие после HCT подвержены риску нескольких острых и хронических сердечно-сосудистых осложнений [29, 30]. OH и POTS, однако, редко считаются частью этих возможных неблагоприятных последствий, и пациенты могут не подвергаться активному скринингу на это осложнение после трансплантации, поэтому точная частота этих осложнений у невыбранных реципиентов HCT неизвестна. OH и POTS могут быть изнурительными, ухудшать качество жизни пациентов и могут задерживать выздоровление после трансплантации [31].Несколько наблюдений связывают ОГ с отрицательными сердечно-сосудистыми исходами [32], а недавний метаанализ подтвердил связь между ОГ и риском всех причин — смерти, ишемической болезни сердца, сердечной недостаточности и инсульта [33].
OH и POTS могут быть изнурительными, ухудшать качество жизни пациентов и могут задерживать выздоровление после трансплантации [31].Несколько наблюдений связывают ОГ с отрицательными сердечно-сосудистыми исходами [32], а недавний метаанализ подтвердил связь между ОГ и риском всех причин — смерти, ишемической болезни сердца, сердечной недостаточности и инсульта [33].
Наши результаты особенно интересны тем, что мы впервые описываем распространенность ОГ и СПОТ у пациентов, направленных в кардиоонкологию после ТГСК. Диагноз OH и POTS был поставлен кардиологом, экспертом в области кардиоонкологии, и подтвержден тщательным просмотром медицинской карты.Следует признать некоторые ограничения. Во-первых, это ретроспективное исследование с очевидным потенциальным смещением отбора в сторону включения пациентов с симптомами и/или предполагаемым сердечно-сосудистым риском, что ограничивает нашу возможность применить эти результаты к невыбранным реципиентам HCT. Во-вторых, отсутствие систематической оценки показателей жизнедеятельности могло привести к неточностям в выявлении OH/POTS и требует проведения будущих проспективных исследований.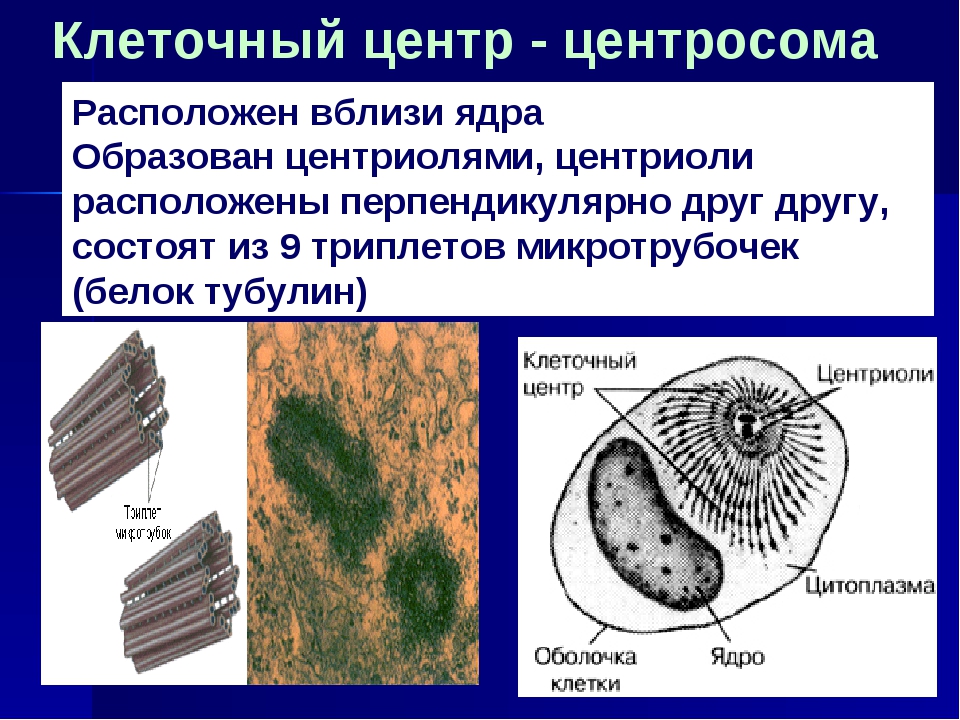 В-третьих, это одноцентровое исследование, хотя оно и проводится в крупной больнице, обслуживающей различные группы пациентов.Наконец, ограниченное количество случаев не позволило нам оценить возможную большую пользу лечения по сравнению с другими.
В-третьих, это одноцентровое исследование, хотя оно и проводится в крупной больнице, обслуживающей различные группы пациентов.Наконец, ограниченное количество случаев не позволило нам оценить возможную большую пользу лечения по сравнению с другими.
какие части клетки поддерживают какие характеристики жизни?
Это всего лишь несколько интересных способов запомнить важные клеточные органеллы и их функции в клетке.
клеточная мембрана — окружает межклеточное содержимое и цитоплазму вместе с органеллами, обычно полупроницаемыми липидными бислоями, особенно в клетках животных
гладкий эндоплазматический ретикулум — здесь производятся и упаковываются многие стероиды и жиры и готовятся к отправке из клетки
* в сравните и сопоставьте гладкость и шероховатость: подумайте о том, что гладкая ткань аналогична жиру, а шероховатая – прочной, структурной…белок строит мышцы, а мышцы делают вас сильным
грубый эндоплазматический ретикулум — здесь упаковываются и подготавливаются многие белки, а также подготавливаются к выходу из клетки готовится к дублированию с помощью первого шага, называемого транскрипцией
митрохондии — похожие на бобы структуры, которые отвечают за производство АТФ и электростанцию, энергию клетки
микротрубочки — считайте их инструментами, проходит через разделение и нуждается в помощи с выравниванием. ..в любом случае они обычно появляются только в это время
..в любом случае они обычно появляются только в это время
лизосомы — подумайте о таких процессах, как очищение клетки от мусора
рибосомы — подумайте о синтезе белка
клеточная стенка — обычно встречается только у растений клетки, твердая оболочка, окружающая растительную клетку
хлорофилл — важная пигментная структура зеленого цвета, которая помогает растениям получать энергию через солнечный свет от солнца
Я думаю, что это основные.Если есть какие-либо другие, которые вы хотите обсудить, дайте мне знать.
.