
«Мель» фонетический разбор | Грамота
Для того чтобы осуществить фонетический анализ слова “мель”, нужно придерживаться правил русского языка по транскрипции. Без учёта этих правил звуко-буквенный разбор не осуществить.
Фонетический разбор
Сначала делим единицу языка “мель” на слоги.
- Количество слогов на прямую зависит от числа гласных букв.
- Анализируемое слово имеет 1 гласную, поэтому имеет 1 слог: мель. Буква “е” – ударная: мЕль.
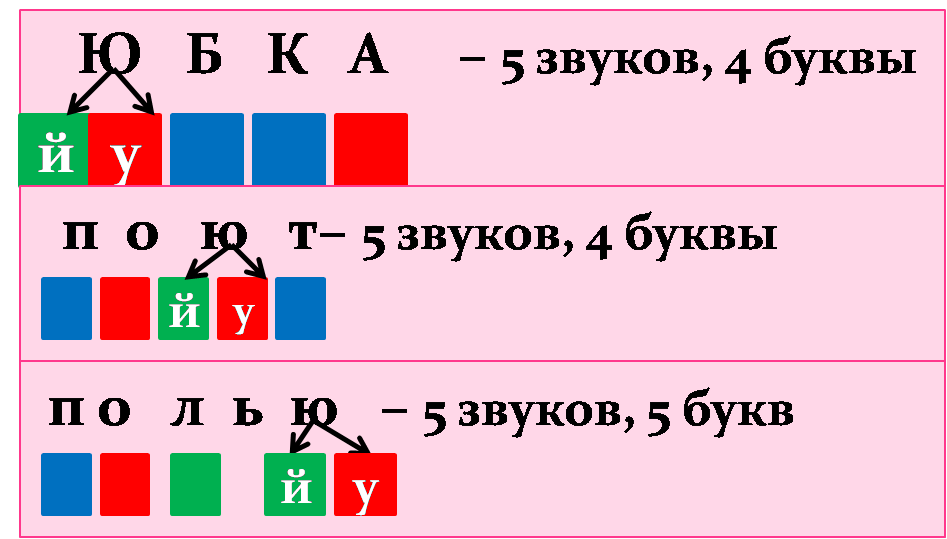
- Следующим шагом подсчитаем сколько единиц письма и речи в слове “мель”: 4 единиц письма (1 гласная, 2 согласных и мягкий знак), 3 звука.
- Данное слово нельзя перенести ни одним методом.
Транскрипция слова
Транскрипция имеет вид: [м’эл’].
Звуко-буквенный разбор
- м — [м’] — согласный, звонкий, мягкий, по второму признаку пара отсутствует, по третьему имеется
- е — [э] — гласный, ударность присутствует
- л — [л’] — согласный, звонкий, мягкий, по второму признаку пара отсутствует, по третьему имеется
- ь — не представляется как единица речи
Проверь себя: «Зимой» фонетический разбор
Гласные:
Единица письма “е” – ударная, поэтому можно представить её как звук [э].
Согласные:
Единица письма “е” - смягчающая, поэтому последующий звук можно представить как единицу речи [м’].
Раздел: ФонетикаПравильно Моллюск – единственно правильный вариант написания существительного, пишется с двумя буквами “л”. Слово является заимствованным (происходит от латинского molluscus – «мягковатый»), правильность написания следует запомнить. На дне моря я увидел моллюска в раковине. Моллюск с витой раковиной очень красивый. Хочу… Читать дальше »
| To reiterate the metaphor of steering, if 31 pilots argue over a ship’s steering wheel, the ship will only run aground. | Пользуясь образом из сферы мореплавания, можно сказать, что если место у штурвала корабля будет оспаривать 31 штурман, то такому кораблю суждено сесть на мель. |
| I hope they run aground and drown. | Надеюсь, что они сели на мель и утонули. |
| We don’t get too many visitors here. Except people whose ships have run aground. | Большинство людей здесь с кораблей, севших на мель в здешних водах. |
| His ship had run aground on an island way west of Sumatra. | Его судно потерпело крушение у острова на западе от Суматры. |
| May run aground because the helmsman’s thought Lacked force to balance opposites. | Бывает в бурю выкинут на риф Затем, что кормчий не сумел найти Для сил противных равновесья… |
| Do you want us to run aground, woman? | Хочешь, чтобы мы на мель сели, женщина? |
| ‘She’s run aground,’ I repeated. ‘I expect she’s got a hole in her bottom.’ | Наскочила на мель, — повторила я, — у нее, видно, дыра в днище. |
| Basically, we’ve run aground on a subspace sandbar. | В сущности, мы сели на подпространственную мель. |
| The San Esteban, Lope’s ship, will run aground in Ireland’s coast. | Сан-Эстебан, корабль Лопе, сядет на мель у берегов Ирландии. |
| To escape the pursuing hooligans, they take a boat out to sea, only to run aground and be left stranded on Grand Island. | Чтобы спастись от преследовавших их хулиганов, они берут лодку в море, но только для того, чтобы сесть на мель и остаться на Большом острове. |
| The following day, 26 April, he spotted two ships that had run aground. | На следующий день, 26 апреля, он заметил два корабля, которые сели на мель. |
| She reached Sydney at sunset on 17 April 2005, after having run aground a few hours earlier in Botany Bay. | Она достигла Сиднея на закате 17 апреля 2005 года, после того как несколькими часами ранее села на мель в Ботани-Бей. |
Portunato’s report on the hull concluded that the ship had not been involved in a collision or run aground. | В отчете портунато по корпусу корабля говорилось, что корабль не был вовлечен в столкновение или сел на мель. |
| The theory supposes that Briggs feared that his ship would run aground and launched the yawl in the hope of reaching land. | Теория предполагает, что Бриггс боялся, что его корабль сядет на мель, и спустил якорь в надежде достичь суши. |
| On February 21, 2007 Bourbon Dolphin succeeded in pulling the German mine hunter Grömitz to sea again after she had run aground near Florø. | 21 февраля 2007 года Bourbon Dolphin удалось снова вытащить в море немецкий минный охотник Grömitz после того, как он сел на мель около флоре. |
| Ferdinand is aboard the ship that is run aground due to the storm created by the sorcerer and old Duke, Prospero. | Фердинанд находится на борту корабля, который сел на мель из-за шторма, созданного колдуном и старым герцогом Просперо. |
A suitable spot being found, Cincinnati was run aground, a hawser tied to a tree, and gangplank laid out. | Когда было найдено подходящее место, Цинциннати сел на мель, привязал к дереву ястреба и спустил трап. |
| A suitable spot being found, the Cincinnati was run aground, a hawser tied to a tree, and gangplank laid out. | Найдя подходящее место, Цинциннати сел на мель, привязал к дереву ястреба и спустил трап. |
| The unusually low water-levels in the Gudenå, the longest Danish river, led to fears that the SS Hjejlen would run aground. | Необычно низкий уровень воды в Гудене, самой длинной датской реке, привел к опасениям, что судно SS Hjejlen сядет на мель. |
| Only for them to run aground on a beach. | Только для того, чтобы они сели на мель на пляже. |
| NOAA said the source of the sound was most likely a large iceberg that had run aground off Antarctica. | НОАА сказал, что источником звука, скорее всего, был большой айсберг, который сел на мель у берегов Антарктиды. |
Eighteen ships were sunk or run aground, including five battleships. | Восемнадцать кораблей были потоплены или сели на мель, в том числе пять линкоров. |
| Другие результаты | |
Краткий словарь финно-угорских топонимов
Краткий словарь финно-угорских топонимов
Автор: А. Тилипман
| ПРИРОДНЫЕ ОБЪЕКТЫ | ||||||
| карел. | фин. | вепс. | эст. | саам. | транскрипция | толкование |
| jogi | joki | jogi | jõgi | ёгк | йок, йоки, йоги, йыги, деги | река |
| oja | oja | oja | oja | вуэйй | ойя, оя, уя, уай, уй, ва | ручей |
| suo | suo | so | soo | шуэннь | суо, суу, соо, шуо | болото |
| neva | nova | нова, ново, нов, нева | трясина, топь | |||
| jarvi | järvi | järv | järv | яввьр | ярв, ярве, ярви, явр | озеро |
| lambi | lampi | ламба, ламбушка = озерцо | ||||
| vaara | vaara | ваара, вара, варака, ваари | гора, возвышенность (покрытая лесом) | |||
| selgy | selkä | sel’g | селльк | сельга, селькя | сельга = каменистая гряда, хребет (буквально: спина) | |
| mägi | mäki | mägi | mägi | мяги, мяки | холм, гора | |
| kallivo | kallio | kal’l’ | kalju | каллио, каливо, калливо, калья | скала, утёс | |
| kivi | kivi | kivi | kivi | киви | крупный камень, булыжник, валун | |
| čuuru | sora | чиввр | чур, чура, чивр, сор | мелкий камень, галька, гравий | ||
| koski | koski | kos’k | kosk | кушшк | коск, коски, кошки | речной порог, перекат, водопад |
| поадан | падун | падун = водопад | ||||
| karikko | куэррк | куэрк, корк, корга, корежка | корга = мель класса «подводная каменистая гряда» | |||
| luuda | luoto | lodo | луд, луда, луото | луда = каменистый островок, мель, отмель | ||
| suari | saari | sar’ | saar | саар, сарь, сари, саари, шуари | остров | |
| суэл | сол, суол | остров | ||||
| pudas | пудас, пудос, пудаш, пудож | протока, рукав реки | ||||
| salmi | salmi | sal’m | чуэлльм | салми, салма, солом, чёлм, чуолма | салма = пролив | |
| lahti | lahti | laht | луххт | лахт, лахта, лухт, лакша | лахта = залив, губа, бухта | |
| niemi | niemi | nem’ | neem | ниеми, немь, мень | мыс (наволок), полуостров | |
| нёаррк | нярк | мыс | ||||
| randu | ranta | rand | rand | рыннт | ранда, ранта, ранна, ранд | берег |
| mua | maa | ma | maa | ма, маа | земля, край, страна, местность | |
| nišku | niska | nišk | ниска | зашеек = исток реки из озера (буквально: затылок) | ||
| pohju | pohja | pohj | põhi | похья, пога | буквально: дно (вероятно, в значении «конец») | |
| muda | muta | muda | muda | моадть` | муд, муда | ил, иловая грязь, муть |
| savi | savi | savi | savi | чуййв | сави, савой | глина |
| hieta | хиета, хето, хетто, хед, хедо | мелкий песок | ||||
| АНТРОПОГЕННЫЕ ОБЪЕКТЫ | ||||||
карел. | фин. | вепс. | эст. | саам. | транскрипция | толкование |
| linnu | linna | lidn | linn | ланнь` | линна, линн | город (кроме фин.), крепость (карел., фин.) |
| kylä | kylä | külä | küla | кюля, кюла, кюлю, кула | деревня, село | |
| kodi | koti | kodi | коти, коди | дом, здание, хозяйство (кар. ) ) | ||
| taloi | talo | тало | дом, здание, хозяйство (кар.) | |||
| perti | pirtti | pert’ | пэррт | перти, пиртти, перт | изба, зимнее жилище (саам.) | |
| koda | kota | куэдть’ | кода, кодо | шалаш, летнее жилище (саам.) | ||
| matku | matka | матка, матк, мотк | путь, дорога (часто в значении «волок») | |||
| veski | вески | мельница | ||||
| sildu | silta | sild | sild | силта | мост | |
| ristu | risti | rist | рысст | ристи | крест | |
| kalmu | kalm | калльм | калма, калмо | могила | ||
| haudu | hauta | haud | хаута, хауда | могила | ||
| ФАУНА | ||||||
карел. | фин. | вепс. | эст. | саам. | транскрипция | толкование |
| kondii | kontio | kondi | контио, конди, конды | медведь | ||
| karhu | karu | карху, карг | медведь | |||
| naali | нёалл | ноль | песец | |||
| koira | koira | koir | koer | койра | собака | |
| sika | siga | siga | сика, сико | свинья | ||
| reboi | reboi | rebane | ребо, ребой, репо, рябо, ряпой | лиса, лисица | ||
| majai | majava | мая, май | бобр | |||
| mägry | mäyrä | mäger | мяг, мег, мягр, мегр, мюгр, маура | барсук | ||
| важь | важ, важе | важенка = самка оленя | ||||
| сарьвэсь | хирвас, хирваш, гирвас | гирвас = олень-производитель | ||||
| hirvi | hirvi | hirb’ | hirv | сэррв | хирви, кирбей | лось, олень (эст. ) ) |
| pedri | peura | pedr | педр, пеура, педра | олень | ||
| lammas | lammas | lambaz | lammas | ламьпесь | ламбас, ламмас | овца |
| orava | orava | orav | orava | вуэррев | орав, оров | белка |
| janoi | jänis | jänes | янис, яниш, янец, яни | заяц | ||
| lindu | lintu | lind | lind | линд, линда, линдо | птица | |
| haukku | haukka | хаукка, хаука | ястреб | |||
| kotku | kotka | kotkas | куэццькэмь | котк, кочк | орёл | |
| kaaren | карьнэсь | куарнис, карниз, корней, курней | ворон | |||
| korppi | корпи | ворон | ||||
| varoi | varis | variš | vares | варис, вариш, вареж, варь, вар | ворона | |
| joučen | joutsen | joucen | юцен, йоутсен, еутсо, евчен, девчен | лебедь | ||
| нюххч | нюхч | лебедь | ||||
| чуххч | чухч, чук | глухарь | ||||
| mečoi | metso | mecoi | metsis | мецо, метсо | глухарь | |
| tikku | tikka | tik | тик | дятел | ||
| чашэнь | чаш, часна | дятел | ||||
| hanhi | hanhi | hanh’ | hani | чуэнь | ханхи, ханга, ханка, чуна, чуно, чунь | гусь |
| kajoi | kajag | kajakas | кай | кай | озёрная чайка | |
| kana | kana | kana | kana | каннь` | кан, кана | курица |
| tadri | teeri | tedr | teder | тедри, тейри, тетро | тетерев | |
| kurgi | kurki | kurg | kurg | курки, курги, кург | журавль | |
| harakku | harakka | haraga | harakas | хараг | сорока | |
| muna | muna | muna | маннь` | муна, мун, манне | яйцо | |
| kala | kala | kala | kala | кулль | кала, кал, куэль | рыба |
| lohi | lohi | lõhi | лохи, логи, лого | лосось | ||
| ahven | ahven | ahven | ahven | ахвен, агвен | окунь | |
| haugi | hauki | haug’ | haug | хауги, хауки, кауге | щука | |
| särgi | särki | särg | сярги, сярки, сярг, сярк, серг, сарг | плотва | ||
| lahnu | lahna | лахн, лагн, лахна | лещ | |||
| uročču | uros | оаресь | урос, урас, орз | самец | ||
| kanzu | kansa | kanz | канза | люди, народ, семья (кар. , вепс.) , вепс.) | ||
| kalmu | kalma | калма, калмо | смерть, богиня смерти и разложения | |||
| hiisi | хиз, хис, хиж, хиден | злой (лесной) дух, демон; священная роща | ||||
Представленный словарь ни в коей мере не претендует на полноту и задуман как полезное приложение к топографическим картам некоторых российских регионов – преимущественно Карелии, Архангельской, Вологодской, Ленинградской и Мурманской областей. Во множестве случаев топонимы несут в себе дополнительную ценную информацию, далеко не всегда отображённую (и способную быть отображённой) на карте иными, классическими способами. С точки зрения составителя, владение этой информацией не может повредить никому.
Записями в словаре, вопреки его названию, сложившемуся исторически, являются не сами топонимы, а кусочки, из которых они составляются, подобно детскому конструктору. К примеру, в Карелии есть остров, называющийся, «Кюляниемисуари». В дословном переводе это означает: «Остров с деревней на мысу». Топоним «Кюляниемисуари» не будет присутствовать в настоящем словаре, но составные его части – «остров», «деревня» и «мыс» – будут. Подобная описательная практика в изготовлении топонимов столь же характерна для рассматриваемых языков и местностей, сколь и не характерна для пары {русский язык, Московская область}, и именно эта особенность является причиной появления настоящего словаря на свет.
Далее, опять же, вопреки названию словаря, составитель рассматривает не всю обширную группу финно-угорских (угро-финских) языков, а лишь прибалтийско-финскую подгруппу финно-волжской группы финно-пермской подветви финно-угорских языков и некоторые языки подгруппы саамской (расположенной в классификации аналогичным образом). Нежелание давать словарю такое название, которое нельзя будет ни запомнить, ни выговорить, ни, тем более, уместить на одной строчке вашего экрана, привело составителя к выбору названия текущего – простого, но, строго говоря, некорректного. Составитель знает, что это название некорректно, ему можно об этом не напоминать. В отдаленной перспективе планируется охватить некоторые другие языки финно-волжской группы, в частности горно-марийский.
Нежелание давать словарю такое название, которое нельзя будет ни запомнить, ни выговорить, ни, тем более, уместить на одной строчке вашего экрана, привело составителя к выбору названия текущего – простого, но, строго говоря, некорректного. Составитель знает, что это название некорректно, ему можно об этом не напоминать. В отдаленной перспективе планируется охватить некоторые другие языки финно-волжской группы, в частности горно-марийский.
Словарь сведён в таблицу, состоящую из трёх логических частей. В левой части располагаются карельские, финские, вепсские, эстонские, а также саамские слова, участвующие в образовании известных составителю топонимов. Буквы саамского алфавита, не отображаемые в стандартных кодировках, заменены на более стандартные символы по принципу: «тот, кому очень надо, тот поймёт». В средней части сведены воедино все те буквосочетания, в которые могут превратиться исходные слова, в процессе переосмысления и перенаписания русскоязычным населением и русскоязычными картографами. Именно эти буквосочетания и следует искать в «теле» топонимов, записанных кириллицей. Наконец, правая часть объясняет смысл буквосочетаний из средней части.
Именно эти буквосочетания и следует искать в «теле» топонимов, записанных кириллицей. Наконец, правая часть объясняет смысл буквосочетаний из средней части.
Таким образом, просмотр таблицы рекомендуется начинать со средней части (второй широкий столбец справа), поглядывая, при необходимости, по сторонам. Если вы ищете конкретное буквосочетание, введите его в поисковую форму, возникающую после нажатия клавиш [Ctrl]+[F].
Ячейки в левой части таблицы могут быть пустыми по двум причинам. Главная из них – составителю неизвестно правильное написание того или иного слова. В первую очередь, это относится к словам саамского языка, письменность для которого удалось придумать всего 25 лет тому назад (в 1982 году). Вторая причина – по мнению составителя, пропущенное слово не участвует в образовании топонимов, а если и участвует, то составитель об этом, к сожалению, не знает.
В самом правом столбце таблицы периодически попадаются знаки равенства «=». Слева от них находятся поморские и иные диалектные слова, справа – их «перевод» на «современный русский язык».
Составитель предупреждает самыми серьёзными, из известных ему русских слов, – цель создания настоящего словаря не научная и даже не псевдонаучная, а сугубо утилитарная. Также оговорюсь, что мыслители, применяющие данный словарь для доказательства превосходства того или иного народа (языка) над другим, делают это в своей собственной, особой системе координат, не совпадающей с системой координат, в которой функционирует составитель.
www.ladoga-lake.ru (2003-2021)
Урок русского языка. Тема урока. Фонетика. Орфоэпия.
Тема урока. Фонетика. Орфоэпия.
Цель урока: повторить и обобщить сведения, изученные в разделе «Фонетика», «Орфоэпия».
Задачи: 1) показать значение фонетики для правописания и произношения;
2) показать роль правильного литературного произношения и ударения в устном общении;
3) освоить литературное произношение ряда слов, раскрыть смыслоразличительную роль ударения.
Формы работы: индивидуальная, фронтальная, групповая.
Оборудование: 1) карточки-информаторы;
2) карточки с заданиями
3)материалы презентации, выполненные в программе Power Point.
Ход урока
I. Сообщение темы, цели урока.
II. Актуализация знаний по теме «Фонетика».
1. Беседа с учащимися (учитель задает вопросы, учащиеся записывают ответы в тетрадь)
№
п/п Вопросы учителя Записи в тетради
1. В название какого раздела науки о языке входит часть слов телефон, магнитофон, микрофон? Фонетика.
Слайд 1
2. Что изучает фонетика Звуки речи.
Слайд 2
3. Звуки, при образовании которых воздушная струя проходит через рот без препятствий. Гласные звуки
Слайд 3
4. При образовании этих звуков воздушная струя проходит через рот, преодолевая различные препятствия. Согласные звуки
Слайд 4
Прочитайте, что у вас получилось?
Правильно. План урока (Слайд 5)
2. Беседа по плану:
— Что изучает фонетика?
— Чем отличаются звуки и буквы?
— Какими бывают гласные звуки? (слайд 6)
— Чем отличается произношение гласных звуков и согласных?
— Какие пары образуют согласные звуки? (слайд 7)
— Какие согласные звуки не имеют пары по глухости и звонкости?
— Какие согласные звуки не имеют пары по твердости и мягкости?
III.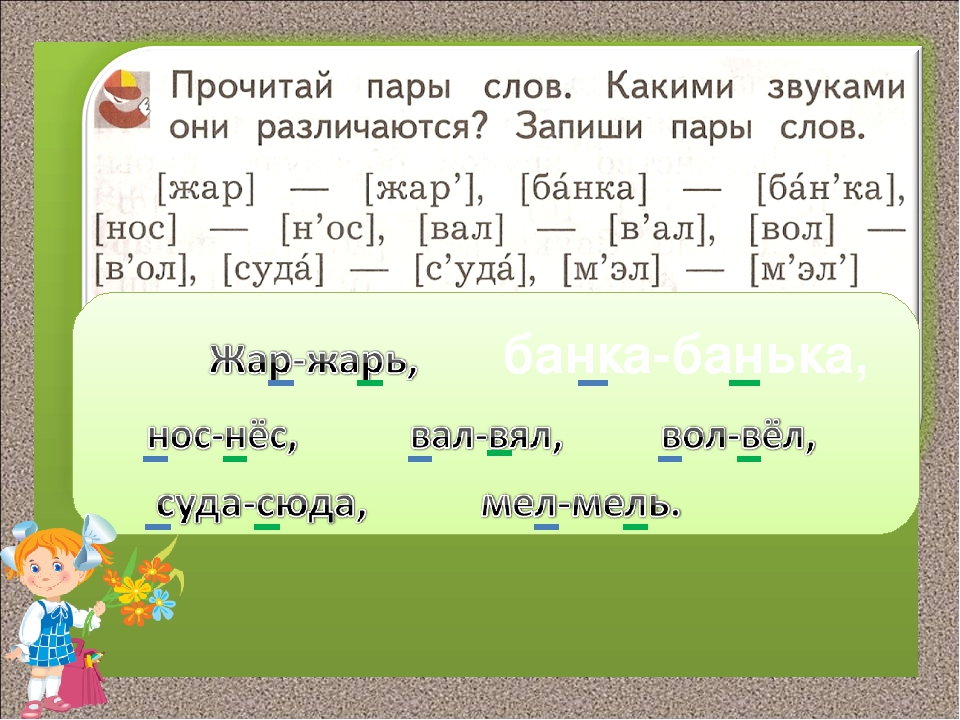 Тренировочные упражнения. (слайд 8)
Тренировочные упражнения. (слайд 8)
Задание 1. (слайд 9) Какими звуками и буквами различаются следующие пары слов?
Вол – вёл, плоты – плоды, супы – зубы, лук – люк, мол – мель.
Задание 2. (слайд 10) Запишите слова, которые образуются, если прочитать в обратном порядке транскрипцию следующих слов.
Лён, лей, люк, ток, шёл, шей, лёд, муж, араб, торг, суд.
Задание 3. (Ученикам раздаются карточки с заданиями) Какие слова затранскрибированы? Запишите их.
[ рыца ], [ в`ирнуца ], [ баица ], [ м`эснас`т ], [ б`эзна ],
[ рацтво ], [л`охкиj ], [ адгатка ], [ игзамин ],
[ дроп` ], [ пачиркн`и ], [ фтарова ], [ п`итно ], [ jэл`н`ик ].
Задание 4 (слайд 11) Запишите транскрипцию следующих слов, распределяя их по трем столбцам.
Количество букв и звуков в слове совпадает Звуков в слове больше, чем букв Звуков в слове меньше, чем букв
Кость, въезд, её, ёжик, женитьба, есть, короед, мозг, молоко, отбросил, подружиться, подсчет, приют, сбежал, ферзь, соевый, рушится, целиком, шведский, ясно, широкая.
Задание 5. (слайд 12) Определите, сколько раз встречается звук [ Ы ] в следующих строчках:
Аист жил у нас на крыше,
А в подвале жили мыши.
IV. Знакомство с основными фонетическими процессами. (Каждый ученик получает карточку-информатор).
Карточка-информатор
В потоке речи звуки влияют друг на друга, что приводит к их изменению. Эти изменения звуков называются фонетическими процессами.
Основные фонетические процессы, связанные с согласными звуками.
1. Оглушение звонких парных на конце слова:
Род [ рот ], флаг [ флак ].
2. Оглушение звонких парных перед глухими:
Селедка [ селетка ], в траву [ фтраву].
3. Озвончение глухих парных перед звонкими (кроме сонорных и [ в ], [ в` ]:
Косьба [каз`ба], к дереву [ гд`эр`иву ].
4. Расподобление звуков – усиление различий между звуками для облегчения произношения. НАПРИМЕР, в слове легкий вместо звуков [гк` ] произносим сочетание [ хк` ], что облегчает произношение слова.
5. Упрощение групп согласных происходит в словах, где есть непроизносимые согласные: в сочетании из трех согласных один не произносится:
снт — [ сн ]: устный — у[ сн ]ый;
здн — [ зн ]: поздний — по[ зн]ий;
лнц — [ нц ]: солнце — со[ нц ]е;
рдц — [ рц ]: сердце — се[ рц ]е.
V. Тренировочные упражнения (слайд 13).
Задание. Определите, какой согласный звук (глухой или звонкий) обозначен подчеркнутой буквой. Какие фонетические процессы наблюдаются в приведенных словах?
Забастовка, сдаться, от дома, любовь, что, указка, честный, счастливый, рябь, мороз, расчет, к берегу.
VI. ВЫВОД. Слово учителя.
Итак, первая тема, которую мы с вами повторили, называется фонетикой. Она изучает звуки нашей речи. Они шумят, поют, свистят, иногда даже спорят, но никогда не ссорятся, живут дружно. Рядом с фонетикой соседствует орфоэпия. Как вы думаете почему? Орфоэпия следит за тем, чтобы правильно произносили звуки в словах. Задача орфоэпии заключается в том, чтобы, минуя все индивидуальные особенности речи, а также особенности местных говоров, сделать язык наиболее совершенным средством широкого общения.
— Что изучает орфоэпия ? (Вопрос к классу)
VII. Произношение сочетаний ЧН, ЧТ. (слайд 14)
В 18 веке орфографическое сочетание ЧН произносилось как [ ШН]. Но со временем этот вариант начинает вытесняться произношением [Ч`Н] .
В современном языке слова с сочетанием ЧН можно разделить на три группы:
1. Упражнение. (слайд 15) Прочитайте данные слова обращая внимание на произношение ЧН.
Потому что, Кузьминична, табачный, молочный, нечто, булочная, девичник, очечник, нарочно, яичница, шуточный, троечник, Фоминична, подсолнечник, бесконечный, двоечник, скучно, конечный.
VIII. Произношение согласных перед Е в заимствованных словах.
В русском языке согласный перед Е звучит мягко (тень, дерево), но в словах, пришедших их других языков, эта закономерность может не соблюдаться. И в сочетаниях де, те, зе, се, не, ре, фе и др. произносится твердый согласный.
1. Упражнение. (слайд 16) Распределите данные слова в 2 столбца.
Согласный перед Е произносится твердо Согласный перед Е произносится мягко
Аттестат, бенефис, индекс, музей, антитеза, фонетика, темп, девиз, дебют, кафе, детектив, шинель, эпитет, синтез, эффект, музей, свитер, интервью, одеколон, пресса, компьютер, тире, термин, ремарка.
IX. Ударение.
Вопросы: 1) Что такое словесное ударение?
2) В чем заключаются особенности русского словесного ударения?
Упражнения: (слайд 17)
1. Составьте словосочетания с данными парами слов, обращая внимание на различное ударение:
Временный – временной, языковой – языковый, мелочный – мелочной, окружный – окружной, видение – видение, клубы – клубы, мокрота – мокрота.
2. Поставьте ударения в следующих словах. (слайд 18 )
Арахис, баловать, гербовый, донельзя, красивее, давнишний, квартал, иконопись, откупорить, еретик, диспансер, обеспечение, дремота, знамение, завидно, опека.
3. Прочитайте словосочетания, обращая внимание на правильную постановку ударения. (слайд 19 )
Подать ходатайство на гербовой бумаге, стала ещё красивее, огромные банты, сотрудники будут премированы, заключим договор, одолевала зевота, давнишний каталог, английская аристократия, средства воротятся, развитие логического мышления.
X. Подведение итогов урока.
1. Что нового вы узнали сегодня на уроке?
2. Какие задания показались наиболее сложными (вызвали затруднения)?
3. Какие умения и навыки совершенствовали?
XI. Домашнее задание.
Выполнить фонетический разбор следующих слов:
Счастливый, яркость, бьюсь, огонь, отбежать, вперед, невтерпеж.
На следующем уроке — контрольный тест.
Мягкие и твёрдые согласные / Согласные звуки и буквы, их обозначающие / Звуки и буквы / Справочник по русскому языку для начальной школы
- Главная
- Справочники
- Справочник по русскому языку для начальной школы
- Звуки и буквы
- Согласные звуки и буквы, их обозначающие
- Мягкие и твёрдые согласные
Согласные звуки могут быть твёрдыми и мягкими.
Многие согласные являются парными по мягкости-твёрдости, т.е. в разных словах могут смягчаться или становиться твёрдыми. В транскрипции мягкость согласного показывается значком [ ’ ].
В транскрипции мягкость согласного показывается значком [ ’ ].
Таблица согласных звуков, парных и непарных по твёрдости/мягкости:
| Группы согласных | Парные | Непарные |
| Твёрдые | [б], [в], [г], [д], [з], [к], [л], [м], [н], [п], [р], [с], [т], [ф], [х] | [ж], [ш], [ц] |
| Мягкие | [б’], [в’], [г’], [д’], [з’], [к’], [л’], [м’], [н’], [п’], [р’], [с’], [т’], [ф’], [х’] | [й’], [ч’], [щ’] |
На письме твёрдость согласных звуков обозначается гласными буквами а, о, у, ы, э (которые идут после согласного звука) — парк [парк], рота [рота], пума [пума], крынка [крынка], сэр [сэр].
Мягкость согласных звуков на письме обозначается:
- мягким знаком ь — мель [м’эл’], пальто [пал’то], письменный [п’ис’м’эный’]
- гласными буквами е, ё, и, ю, я — пень [п’эн’], свёкла [св’окла], пить [п’ит’], клюв [кл’ув], пляска [пл’аска].

Кроме этого парные по твёрдости/мягкости согласные звуки могут смягчаться и перед некоторыми мягкими согласными — з [з’], с [с’], н [н’], т [т’], л [л’].
Например: песня [п’эс’н’а], если [й’эс’л’и], хвостик [хвос’т’ик] и другие.
Также согласный [н’] будет мягким перед согласными буквами ч и щ, при этом мягкий знак между ними не пишется: пончик [пон’ч’ик], гонщик [гон’щ’ик].
Запомни, что согласные непарные по твёрдости-мягкости остаются только твёрдыми или только мягкими (вне зависимости от того, есть ли после них мягкий знак или какая за ними идёт гласная).
Твёрдые непарные согласные всегда остаются твёрдыми.
Например:
[ж] — жара [жара], жёлтый [жолтый’]
[ш] — шар [шар], шью [шй’у]
[ц] — цапля [цапл’а], цирк [цырк]
Мягкие непарные согласные всегда остаются мягкими.
Например:
[й’] — йогурт [й’огурт], мой [мой’]
[ч’] — час [ч’ас], чек [ч’эк]
[щ’] — щука [щ’ука], щель [щ’эл’]
Поделись с друзьями в социальных сетях:
Советуем посмотреть:
Глухие и звонкие согласные
Алфавит
Гласные звуки и буквы, их обозначающие
Согласные звуки и буквы, их обозначающие
Слог
Ударение
Фонетический разбор
Звуки и буквы
Правило встречается в следующих упражнениях:
1 класс
Страница 85, Канакина, Горецкий, Учебник
Страница 42, Канакина, Горецкий, Рабочая тетрадь
Упражнение 98, Климанова, Макеева, Учебник
Упражнение 160, Климанова, Макеева, Учебник
Упражнение 168, Климанова, Макеева, Учебник
Страница 26, Климанова, Рабочая тетрадь
Страница 27, Климанова, Рабочая тетрадь
Упражнение 5, Иванов, Евдокимова, Кузнецова, Учебник
Упражнение 5, Иванов, Евдокимова, Кузнецова, Учебник
Упражнение 2, Иванов, Евдокимова, Кузнецова, Учебник
2 класс
Упражнение 195, Канакина, Горецкий, Учебник, часть 1
Упражнение 205, Канакина, Горецкий, Учебник, часть 1
Упражнение 132, Канакина, Рабочая тетрадь, часть 1
Упражнение 136, Канакина, Рабочая тетрадь, часть 1
Упражнение 12, Канакина, Горецкий, Учебник, часть 2
Упражнение 76, Климанова, Бабушкина, Учебник, часть 1
Упражнение 88, Климанова, Бабушкина, Учебник, часть 1
Упражнение 110, Полякова, Учебник, часть 1
Упражнение 156, Полякова, Учебник, часть 1
Упражнение 185, Бунеев, Бунеева, Пронина, Учебник
3 класс
Упражнение 124, Канакина, Горецкий, Учебник, часть 1
Упражнение 52, Канакина, Рабочая тетрадь, часть 2
Упражнение 39, Климанова, Бабушкина, Учебник, часть 1
Упражнение 43, Климанова, Бабушкина, Учебник, часть 1
Упражнение 29, Климанова, Бабушкина, Рабочая тетрадь, часть 1
Упражнение 30, Климанова, Бабушкина, Рабочая тетрадь, часть 1
Упражнение 25, Климанова, Бабушкина, Учебник, часть 2
Упражнение 239, Климанова, Бабушкина, Учебник, часть 2
Упражнение 45, Климанова, Бабушкина, Рабочая тетрадь, часть 2
Упражнение 34, Песняева, Анащенкова, Рабочая тетрадь, часть 2
4 класс
Упражнение 57, Климанова, Бабушкина, Учебник, часть 1
Упражнение 54, Полякова, Учебник, часть 2
Упражнение 174, Бунеев, Бунеева, Пронина, Учебник, часть 2
5 класс
Упражнение 279, Ладыженская, Баранов, Тростенцова, Григорян, Кулибаба, Учебник, часть 1
Упражнение 280, Ладыженская, Баранов, Тростенцова, Григорян, Кулибаба, Учебник, часть 1
Упражнение 310, Ладыженская, Баранов, Тростенцова, Григорян, Кулибаба, Учебник, часть 1
Упражнение 187, Разумовская, Львова, Капинос, Учебник
Упражнение 193, Разумовская, Львова, Капинос, Учебник
Упражнение 33, Александрова, Рыбченкова, Глазков, Лисицин, Учебник, часть 1
Упражнение 49, Александрова, Рыбченкова, Глазков, Лисицин, Учебник, часть 1
Упражнение 50, Александрова, Рыбченкова, Глазков, Лисицин, Учебник, часть 1
Упражнение Задачка стр. 26,
Александрова, Рыбченкова, Глазков, Лисицин, Учебник, часть 1
26,
Александрова, Рыбченкова, Глазков, Лисицин, Учебник, часть 1
Упражнение 51, Александрова, Рыбченкова, Глазков, Лисицин, Учебник, часть 1
7 класс
Упражнение 481, Ладыженская, Баранов, Тростенцова, Григорян, Кулибаба, Александрова, Учебник
Упражнение 36, Разумовская, Львова, Капинос, Учебник
Упражнение 550, Разумовская, Львова, Капинос, Учебник
© budu5.com, 2021
Пользовательское соглашение
Copyright
Урок по русскому языку «Повторение изученного в начальной школе» | План-конспект урока по русскому языку (5 класс) по теме:
Тема: «Повторение изученного в начальной школе»
Цель: проверить умение владения навыком графического выделения орфограммы, определение морфологических признаков самостоятельных частей речи.
Ход урока:
1.Оргмомент.
2.Цель урока.
Изучение и сбережение русского языка является не праздным занятием от нечего делать, но насущной необходимостью.
А.И. Куприн
Обращение к эпиграфу:
- Убедили ли вас наши занятия в правомерности слов писателя Куприна?
- Попытайтесь обосновать свой ответ.
Сегодня мы будем повторять изученный материал, решая грамматические задачи.
Грамматические задачи даны в виде эпиграфов к каждому разделу повторения:
- УМЕЕТЕ ЛИ ВЫ РАЗЛИЧАТЬ ЧАСТИ РЕЧИ?
Для всего в русском языке есть великое множество слов.
К.Г. Паустовский
1)Словарь: искусство, искусный, багряный, адъютант, юннаты, юный, инженер, иллюстрация.
Слова обязательно являются какой-то частью речи. Расскажите о классификации частей речи. А какой части речи здесь нет? (глагол)
2)ИНЖЕНЕР (ТРАНСКРИПЦИЯ)
3)Записать предложение Опытный инженер чертил на бумаге. (синтаксический разбор, назвать орфограммы, подписать части речи, выписать словосочетания).
2.ПОВТОРИМ ОРФОГРАФИЮ.
Кто грамоте горазд, тому не пропасть.
Пословица
Что необходимо сделать в первую очередь при объяснении орфограмм? (Определить часть речи и часть слова.)
А. 1. В каком ряду в обоих словах пропущена безударная проверяемая гласная корня?
1) ключ..к, з..ма
2) зв..нок, кур..ца
3) в..сьмой, ч..рнеть
4) под..брать, к..рова
A2. В каком ряду в обоих словах пропущена одна и та же буква?
1) арбу.., сна..ть,
2) коро..ка, ша..ка
3) ко..точка, по..торить
4) наро.., скла..ка
А3. В каком слове нет непроизносимой согласной в корне?
1) вес..ник
2) чудес..ный
3) мес.ный
4) со..нце
ФИЗКУЛЬТМИНУТКА:
Если ВЫ согласны с утверждением, то стоите, если не согласны – садитесь.
- В русском языке 3 спряжения глагола.
- Существительные относятся к мужскому, женскому, среднему роду.
- Глаголы прошедшего времени изменяются по лицам и числам.
- К 3скл.
 относятся существительные жен. и муж. рода
относятся существительные жен. и муж. рода - Существительные склоняются.
- Прилагательные спрягаются.
- ВСПОМНИМ ПУНКТУАЦИЮ.
Знаки препинания – это как нотные знаки.
Они твердо держат текст и не дают ему рассыпаться.
К.Г. Паустовский
А1. В каком предложении знаки препинания расставлены неправильно? (слайд)
1) Приходит весна и птицы возвращаются в родные края.
2) Весною я пошел смотреть свои яблони и увидал, что мыши
объели их корни.
3) Трамвай показался из-за угла, медленно пересёк проспект
и остановился.
4) Хозяин заварил чай и позвал гостей к столу.
3.Работа с текстом (читает учитель- текст спроецирован на экран)
Дельфинья дружба.
По неизвес…ным причинам дельфины иногда сами выбрасываю(т,ть)ся на берег. Рыбаки говорят, что они увлекаются погоней (за)рыбой и садя(т,ть)ся на мель.Дельфины погибнут, если им не помогут люди.
Самое т.. желое препятствие для спасения дельфина – дельфин..я дружба. Если вы станете относить их в воду, то у вас ничего (не)получится. Дельфин вернется и снова выбр..сится на мель, если услышит крики своих товарищей.. Товарища в беде дельфин никогда (не)оставит.
желое препятствие для спасения дельфина – дельфин..я дружба. Если вы станете относить их в воду, то у вас ничего (не)получится. Дельфин вернется и снова выбр..сится на мель, если услышит крики своих товарищей.. Товарища в беде дельфин никогда (не)оставит.
— Что такое текст?
— Можно ли данную запись назвать текстом?
— Определите тему текста.
— Какова основная мысль? Что хотел автор подчеркнуть данным текстом?
— Выпишите слова с пропущенными буквами, обозначьте орфограмму .
Дифференцированная работа с текстом:
1 ряд – выписать 5 глаголов, выделить окончание. Определить спряжение
2 ряд – выписать 5 существительных, определить склонение.
3 ряд – выписать прилагательные, определить род, число, падеж.
Для всех морфемный разбор:
неизвестным, выбросится, рыбкой
Подведение итогов, возвращение к эпиграфу урока, записанному на доске.
.
шагов генетической транскрипции | Биология для майоров I
Результаты обучения
- Основные этапы транскрипции ДНК в РНК
Транскрипция происходит в ядре. Он использует ДНК в качестве матрицы для создания молекулы РНК (мРНК). Во время транскрипции создается цепь мРНК, которая комплементарна цепи ДНК. На рисунке 1 показано, как это происходит.
Он использует ДНК в качестве матрицы для создания молекулы РНК (мРНК). Во время транскрипции создается цепь мРНК, которая комплементарна цепи ДНК. На рисунке 1 показано, как это происходит.
Рисунок 1. Обзор транскрипции. Транскрипция использует последовательность оснований в цепи ДНК для создания комплементарной цепи мРНК.Триплеты — это группы из трех последовательных нуклеотидных оснований в ДНК. Кодоны — это комплементарные группы оснований в мРНК.
Вы также можете пройти этапы транскрипции по этой ссылке.Транскрипция происходит в три этапа: инициация, удлинение и завершение. Шаги показаны на рисунке 2.
Рис. 2. Транскрипция происходит в три этапа — инициацию, удлинение и завершение, — все они показаны здесь.
Шаг 1: Инициирование
Инициирование — начало транскрипции.Это происходит, когда фермент РНК-полимераза связывается с областью гена, называемой промотором. Это дает сигнал ДНК раскручиваться, чтобы фермент мог «прочитать» основания в одной из цепей ДНК. Теперь фермент готов к созданию цепи мРНК с комплементарной последовательностью оснований.
Теперь фермент готов к созданию цепи мРНК с комплементарной последовательностью оснований.
Шаг 2: удлинение
Удлинение представляет собой добавление нуклеотидов к цепи мРНК. РНК-полимераза считывает размотанную цепь ДНК и строит молекулу мРНК, используя комплементарные пары оснований.Во время этого процесса есть короткое время, когда вновь образованная РНК связывается с размотанной ДНК. Во время этого процесса аденин (A) в ДНК связывается с урацилом (U) в РНК.
Шаг 3: Прекращение действия
Терминация — это окончание транскрипции, которое происходит, когда РНК-полимераза пересекает стоп-последовательность (терминацию) в гене. Нить мРНК завершена и отделяется от ДНК.
В этом видео представлен обзор этих шагов. Вы можете прекратить просмотр видео в 5:35.(После этого обсуждается перевод, который мы обсудим в следующем результате.)
Внесите свой вклад!
У вас была идея улучшить этот контент? Нам очень понравится ваш вклад.
Улучшить эту страницуПодробнее
ТранскрипцияРНК | Микробиология
Цели обучения
- Объясните, как синтезируется РНК с использованием ДНК в качестве матрицы
- Различать транскрипцию у прокариот и эукариот
В процессе транскрипции информация, закодированная в последовательности ДНК одного или нескольких генов, транскрибируется в цепь РНК, также называемую транскриптом РНК .Полученная одноцепочечная молекула РНК, состоящая из рибонуклеотидов, содержащих основания аденин (A), цитозин (C), гуанин (G) и урацил (U), действует как мобильная молекулярная копия исходной последовательности ДНК. Транскрипция у прокариот и эукариот требует, чтобы двойная спираль ДНК частично раскручивалась в области синтеза РНК. Развернутая область называется пузырем транскрипции . Транскрипция определенного гена всегда происходит от одной из двух цепей ДНК, которая действует как матрица, так называемой антисмысловой цепи . Продукт РНК комплементарен матричной цепи ДНК и почти идентичен нетемплатной цепи ДНК или смысловой цепи . Единственное отличие состоит в том, что в РНК все нуклеотиды Т заменены на нуклеотиды U; во время синтеза РНК U включается, когда в комплементарной антисмысловой цепи присутствует A.
Продукт РНК комплементарен матричной цепи ДНК и почти идентичен нетемплатной цепи ДНК или смысловой цепи . Единственное отличие состоит в том, что в РНК все нуклеотиды Т заменены на нуклеотиды U; во время синтеза РНК U включается, когда в комплементарной антисмысловой цепи присутствует A.
Транскрипция бактерий
Бактерии используют одну и ту же РНК-полимеразу для транскрипции всех своих генов. Подобно ДНК-полимеразе, РНК-полимераза добавляет нуклеотиды один за другим к 3′-ОН группе растущей нуклеотидной цепи.Одним из критических различий в активности между ДНК-полимеразой и РНК-полимеразой является потребность в 3′-ОН, к которому можно добавить нуклеотиды: ДНК-полимеразе требуется такая 3′-ОН группа, что требует праймера, тогда как РНК-полимеразы нет. Во время транскрипции рибонуклеотид, комплементарный матричной цепи ДНК, добавляется к растущей цепи РНК, и ковалентная фосфодиэфирная связь образуется посредством синтеза дегидратации между новым нуклеотидом и последним добавленным нуклеотидом. В E. coli РНК-полимераза состоит из шести полипептидных субъединиц, пять из которых составляют основной фермент полимеразы, ответственный за добавление нуклеотидов РНК к растущей цепи.Шестая субъединица известна как сигма (σ). Фактор σ позволяет РНК-полимеразе связываться со специфическим промотором, тем самым обеспечивая транскрипцию различных генов. Существуют различные σ-факторы, которые позволяют транскрипцию различных генов.
В E. coli РНК-полимераза состоит из шести полипептидных субъединиц, пять из которых составляют основной фермент полимеразы, ответственный за добавление нуклеотидов РНК к растущей цепи.Шестая субъединица известна как сигма (σ). Фактор σ позволяет РНК-полимеразе связываться со специфическим промотором, тем самым обеспечивая транскрипцию различных генов. Существуют различные σ-факторы, которые позволяют транскрипцию различных генов.
Инициирование
Инициирование транскрипции начинается с промотора , последовательности ДНК, с которой аппарат транскрипции связывается и инициирует транскрипцию. Нуклеотидная пара в двойной спирали ДНК, которая соответствует сайту, из которого транскрибируется первый 5′-нуклеотид РНК, является сайтом инициации.Нуклеотиды, предшествующие сайту инициации, обозначены как «вышестоящие», тогда как нуклеотиды, следующие за сайтом инициации, называются «нижележащими» нуклеотидами. В большинстве случаев промоторы расположены непосредственно перед генами, которые они регулируют. Хотя промоторные последовательности различаются в зависимости от бактериального генома, некоторые элементы сохраняются. В положениях –10 и –35 внутри ДНК перед сайтом инициации (обозначенным +1) есть две консенсусные последовательности промоторов или области, которые схожи для всех промоторов и у разных видов бактерий.Консенсусная последовательность –10, называемая блоком TATA , является TATAAT. Последовательность –35 распознается и связывается с σ.
Хотя промоторные последовательности различаются в зависимости от бактериального генома, некоторые элементы сохраняются. В положениях –10 и –35 внутри ДНК перед сайтом инициации (обозначенным +1) есть две консенсусные последовательности промоторов или области, которые схожи для всех промоторов и у разных видов бактерий.Консенсусная последовательность –10, называемая блоком TATA , является TATAAT. Последовательность –35 распознается и связывается с σ.
Удлинение
Элонгация в фазе транскрипции начинается, когда σ-субъединица диссоциирует от полимеразы, позволяя коровому ферменту синтезировать РНК, комплементарную матрице ДНК, в направлении от 5 ‘к 3’ со скоростью примерно 40 нуклеотидов в секунду. По мере удлинения ДНК непрерывно разматывается перед основным ферментом и снова наматывается за ним (рис. 1).
Рис. 1. Во время элонгации бактериальная РНК-полимераза отслеживает ДНК-матрицу, синтезирует мРНК в направлении от 5 ‘к 3’ и раскручивает и перематывает ДНК по мере ее считывания.
Прекращение действия
После того, как ген транскрибируется, бактериальная полимераза должна отделиться от матрицы ДНК и высвободить вновь образованную РНК. Это называется прекращением транскрипции . Матрица ДНК включает повторяющиеся нуклеотидные последовательности, которые действуют как сигналы терминации, заставляя РНК-полимеразу останавливаться и высвобождаться из матрицы ДНК, высвобождая транскрипт РНК.
Подумай об этом
- Где σ-фактор РНК-полимеразы связывает ДНК, чтобы начать транскрипцию?
- Что происходит, чтобы инициировать полимеризационную активность РНК-полимеразы?
- Откуда исходит сигнал об окончании транскрипции?
Транскрипция в эукариотах
Прокариоты и эукариоты выполняют в основном одинаковый процесс транскрипции с некоторыми существенными различиями (см. Таблицу 1). Эукариоты используют три различных полимеразы, РНК-полимеразы I, II и III, все структурно отличные от бактериальной РНК-полимеразы . Каждый транскрибирует разные подмножества генов. Интересно, что архей содержат единственную РНК-полимеразу, которая более близка к эукариотической РНК-полимеразе II, чем к ее бактериальному аналогу. Эукариотические мРНК также обычно являются моноцистронными, что означает, что каждая из них кодирует только один полипептид, тогда как прокариотические мРНК бактерий и архей обычно являются полицистронными , что означает, что они кодируют несколько полипептидов.
Каждый транскрибирует разные подмножества генов. Интересно, что архей содержат единственную РНК-полимеразу, которая более близка к эукариотической РНК-полимеразе II, чем к ее бактериальному аналогу. Эукариотические мРНК также обычно являются моноцистронными, что означает, что каждая из них кодирует только один полипептид, тогда как прокариотические мРНК бактерий и архей обычно являются полицистронными , что означает, что они кодируют несколько полипептидов.
Наиболее важным различием между прокариотами и эукариотами является мембраносвязанное ядро последних, которое влияет на простоту использования молекул РНК для синтеза белка.Когда гены связаны в ядре, эукариотическая клетка должна транспортировать кодирующие белок молекулы РНК в цитоплазму для трансляции. Кодирующие белки первичные транскрипты , молекулы РНК, непосредственно синтезируемые РНК-полимеразой, должны пройти несколько этапов процессинга, чтобы защитить эти молекулы РНК от деградации во время их переноса из ядра в цитоплазму и транслирования в белок. Например, мРНК эукариот может длиться несколько часов, тогда как типичная мРНК прокариот длится не более 5 секунд.
Например, мРНК эукариот может длиться несколько часов, тогда как типичная мРНК прокариот длится не более 5 секунд.
Первичный транскрипт (также называемый пре-мРНК) сначала покрывается РНК-стабилизирующими белками, чтобы защитить его от деградации, пока он обрабатывается и экспортируется из ядра. Обработка первого типа начинается, пока синтезируется первичная расшифровка; специальный 7-метилгуанозиновый нуклеотид, называемый 5′-кэп , добавляется к 5′-концу растущего транскрипта. Помимо предотвращения деградации, факторы, участвующие в последующем синтезе белка, распознают кэп, что помогает инициировать трансляцию рибосомами.После завершения удлинения другой процессирующий фермент затем добавляет цепочку из примерно 200 адениновых нуклеотидов к 3′-концу, называемую хвостом поли-А . Эта модификация дополнительно защищает пре-мРНК от деградации и сигналов клеточным факторам о том, что транскрипт необходимо экспортировать в цитоплазму.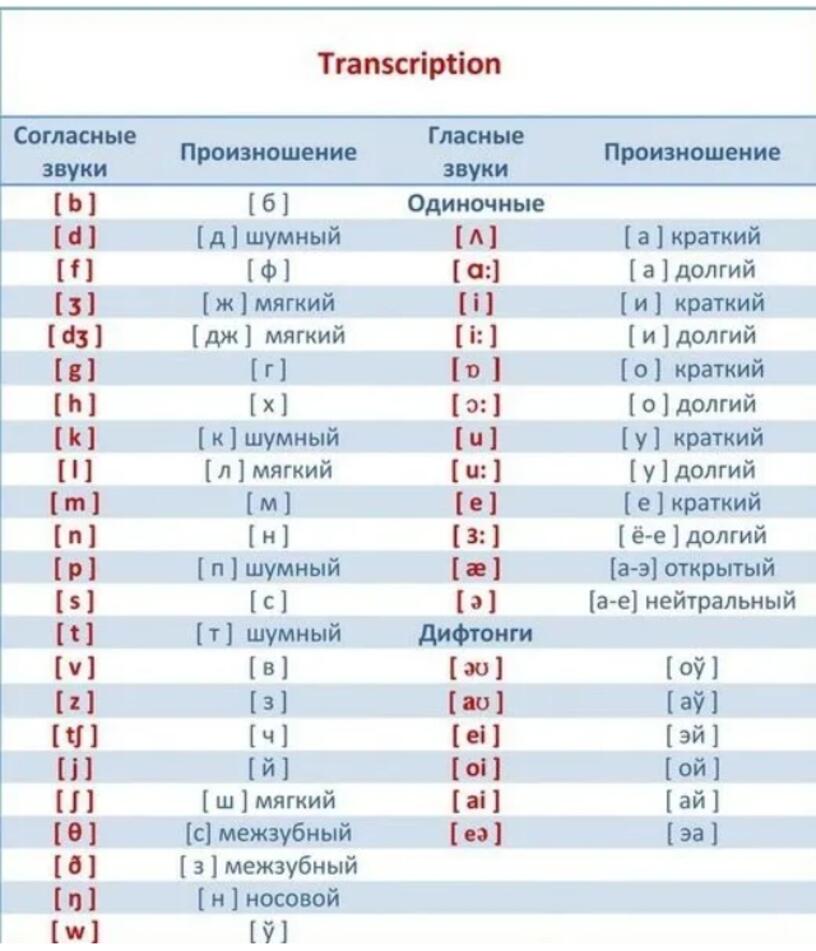
Эукариотические гены, кодирующие полипептиды, состоят из кодирующих последовательностей, называемых экзонами ( ex -on означает, что они сжаты ex ) и промежуточных последовательностей, называемых интронами ( int -ron обозначает их исходную роль int . ).Транскрибируемые последовательности РНК, соответствующие интронам, не кодируют участки функционального полипептида и удаляются из пре-мРНК во время процессинга. Важно, чтобы все последовательности РНК, кодируемые интроном, были полностью и точно удалены из пре-мРНК перед синтезом белка, чтобы последовательности, кодируемые экзонами, были правильно соединены вместе для кодирования функционального полипептида. Если процесс ошибается даже на один нуклеотид, последовательности соединенных экзонов будут сдвинуты, и полученный полипептид окажется нефункциональным.Процесс удаления последовательностей РНК, кодируемых интроном, и повторного соединения последовательностей, кодируемых экзонами, называется сплайсингом РНК и облегчается действием сплайсосомы , содержащей малые ядерные рибонуклеопротеины (мяРНП). Последовательности РНК, кодируемые интроном, удаляются из пре-мРНК, пока она еще находится в ядре. Хотя они не транслируются, интроны, по-видимому, выполняют различные функции, включая регуляцию генов и транспорт мРНК. По завершении этих модификаций зрелый транскрипт , мРНК, кодирующая полипептид, транспортируется из ядра и направляется в цитоплазму для трансляции.Интроны можно сплайсировать по-разному, в результате чего различные экзоны включаются или исключаются из конечного продукта мРНК. Этот процесс известен как альтернативное сращивание . Преимущество альтернативного сплайсинга заключается в том, что могут быть созданы различные типы транскриптов мРНК, все из которых происходят из одной и той же последовательности ДНК. В последние годы было показано, что некоторые археи также обладают способностью сплайсировать свою пре-мРНК.
Последовательности РНК, кодируемые интроном, удаляются из пре-мРНК, пока она еще находится в ядре. Хотя они не транслируются, интроны, по-видимому, выполняют различные функции, включая регуляцию генов и транспорт мРНК. По завершении этих модификаций зрелый транскрипт , мРНК, кодирующая полипептид, транспортируется из ядра и направляется в цитоплазму для трансляции.Интроны можно сплайсировать по-разному, в результате чего различные экзоны включаются или исключаются из конечного продукта мРНК. Этот процесс известен как альтернативное сращивание . Преимущество альтернативного сплайсинга заключается в том, что могут быть созданы различные типы транскриптов мРНК, все из которых происходят из одной и той же последовательности ДНК. В последние годы было показано, что некоторые археи также обладают способностью сплайсировать свою пре-мРНК.
| Таблица 1. Сравнение транскрипции у бактерий и эукариот | ||
|---|---|---|
| Недвижимость | Бактерии | Эукариоты |
| Количество полипептидов, кодируемых на мРНК | Моноцистронный или полицистронный | Исключительно моноцистронные |
| Удлинение прядей | ядро + σ = холоэнзим | РНК-полимеразы I, II или III |
| Добавление крышки 5 ‘ | № | Есть |
| Добавление 3 ‘хвоста поли-А | № | Есть |
| Сплайсинг пре-мРНК | № | Есть |
 In bacteria the holoenzyme (core + sigma) is responsible for strand elongation. In eukaryotes RNA polymerase I, II, or III are responsible for strand elongation. Eukaryotes have the addition of a 5′ cap, bacteria do not. Eukaryotes have the addition of a 3′ poly-A tail, bacteria do not. Eukaryotes have splicing of pre-mRNA, bacteria do not.»>
In bacteria the holoenzyme (core + sigma) is responsible for strand elongation. In eukaryotes RNA polymerase I, II, or III are responsible for strand elongation. Eukaryotes have the addition of a 5′ cap, bacteria do not. Eukaryotes have the addition of a 3′ poly-A tail, bacteria do not. Eukaryotes have splicing of pre-mRNA, bacteria do not.»> Визуализируйте, как происходит сплайсинг мРНК, наблюдая за процессом в действии в этом видео.
Посмотрите, как удаляются интроны во время сплайсинга РНК.
Подумай об этом
- В эукариотических клетках, как транскрипт РНК из гена белка изменяется после его транскрипции?
- Содержат ли экзоны или интроны информацию о белковых последовательностях?
Клиническая направленность: Трэвис, часть 2
Этот пример продолжает историю Трэвиса, начатую в книге «Функции генетического материала».
В отделении неотложной помощи медсестра сказала Трэвису, что он принял правильное решение — поехать в больницу, потому что его симптомы указывают на инфекцию, которая вышла из-под контроля.Симптомы Трэвиса прогрессировали, пораженный участок кожи и опухоль увеличивались. Внутри пораженного участка появилась сыпь, образовались пузыри и небольшие газовые карманы под самым внешним слоем кожи, и некоторые участки кожи стали серыми.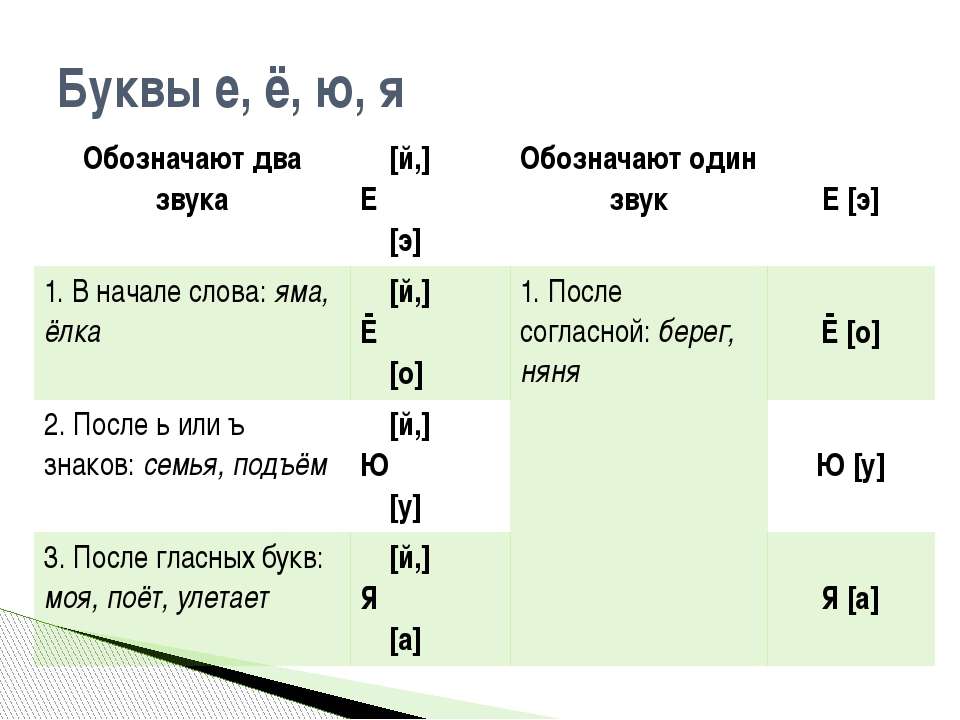 Учитывая гнилостный запах гноя, вытекающего из одного из пузырей, быстрое прогрессирование инфекции и внешний вид пораженной кожи, врач немедленно начал лечение некротического фасциита.Врач Трэвиса назначил посев жидкости, вытекающей из волдыря, а также назначил анализ крови, в том числе количество лейкоцитов.
Учитывая гнилостный запах гноя, вытекающего из одного из пузырей, быстрое прогрессирование инфекции и внешний вид пораженной кожи, врач немедленно начал лечение некротического фасциита.Врач Трэвиса назначил посев жидкости, вытекающей из волдыря, а также назначил анализ крови, в том числе количество лейкоцитов.
Трэвис был помещен в отделение интенсивной терапии и начал внутривенное введение антибиотика широкого спектра действия, чтобы попытаться минимизировать дальнейшее распространение инфекции. Несмотря на лечение антибиотиками, состояние Трэвиса быстро ухудшилось. Трэвис смутился, у него закружилась голова. В течение нескольких часов после поступления в больницу его кровяное давление значительно упало, а дыхание стало более поверхностным и учащенным.Вдобавок волдыри увеличивались, волдыри становились пурпурно-черными, а сама рана, казалось, быстро прогрессировала вверх по ноге Трэвиса.
- Какие возможные возбудители некротического фасциита Трэвиса?
- Каковы возможные объяснения того, почему лечение антибиотиками не работает?
Мы вернемся к примеру Трэвиса на следующих страницах.
Ключевые концепции и резюме
- Во время транскрипции информация, закодированная в ДНК, используется для создания РНК.
- РНК-полимераза синтезирует РНК, используя антисмысловую цепь ДНК в качестве матрицы путем добавления нуклеотидов комплементарной РНК к 3′-концу растущей цепи. РНК-полимераза
- связывается с ДНК в последовательности, называемой промотором , во время инициации транскрипции .
- Гены, кодирующие белки родственных функций, часто транскрибируются под контролем одного промотора в прокариотах, что приводит к образованию полицистронной молекулы мРНК , которая кодирует несколько полипептидов.
- В отличие от ДНК-полимеразы, РНК-полимеразе не требуется 3′-ОН группа для добавления нуклеотидов, поэтому праймер не требуется во время инициации.
- Прекращение транскрипции у бактерий происходит, когда РНК-полимераза сталкивается со специфическими последовательностями ДНК, которые приводят к остановке полимеразы.
 Это приводит к высвобождению РНК-полимеразы из цепи ДНК-матрицы, высвобождая транскрипт РНК .
Это приводит к высвобождению РНК-полимеразы из цепи ДНК-матрицы, высвобождая транскрипт РНК . - Эукариоты имеют три различных РНК-полимеразы.У эукариот также есть моноцистронная мРНК, каждая из которых кодирует только один полипептид.
- Первичные транскрипты эукариот обрабатываются несколькими способами, включая добавление 5′-кэпа и 3′- поли-A-хвоста , а также сплайсинг с образованием зрелой молекулы мРНК, которая может быть транспортирована. ядра, и это защищено от деградации.
Множественный выбор
На какой стадии бактериальной транскрипции участвует σ-субъединица РНК-полимеразы?
- инициирование
- удлинение
- прекращение
- сварка
Ответ а.Субъединица σ РНК-полимеразы участвует в инициации.
Какой из следующих компонентов участвует в инициации транскрипции?
- грунтовка
- происхождение
- промоутер
- стартовый кодон
Ответ c. Промотор участвует в инициации транскрипции.
Промотор участвует в инициации транскрипции.
Что из следующего не является функцией 5′-кэпа и 3′-поли-A-хвоста молекулы зрелой эукариотической мРНК?
- для облегчения сварки
- для предотвращения деградации мРНК
- для экспорта зрелого транскрипта в цитоплазму
- для облегчения связывания рибосомы с транскриптом
Ответ а.Облегчение сращивания не зависит от 5′-колпачка и 3′-хвоста поли-А.
Зрелая мРНК эукариота будет содержать каждую из этих функций, кроме какой из следующих?
- РНК, кодируемая экзоном
- РНК, кодируемая интроном
- 5 ‘крышка
- 3 ′ хвостовик поли-А
Ответ б. Зрелая мРНК эукариота будет , а не , содержать РНК, кодируемую интроном.
Заполните бланк
A ________ мРНК — это мРНК, которая кодирует несколько полипептидов.
Покажи ответ Полицистронная мРНК — это мРНК, кодирующая несколько полипептидов.
Белковый комплекс, отвечающий за удаление кодируемых интроном последовательностей РНК из первичных транскриптов у эукариот, называется ________.
Покажи ответБелковый комплекс, ответственный за удаление кодируемых интроном последовательностей РНК из первичных транскриптов у эукариот, называется сплайсосомой .
Подумай об этом
- Какова цель процессинга РНК у эукариот? Почему прокариотам не нужна подобная обработка?
- Ниже представлена последовательность ДНК.Представьте, что это часть молекулы ДНК, которая отделилась при подготовке к транскрипции, поэтому вы видите только антисмысловую цепь. Постройте последовательность мРНК, транскрибируемую с этой матрицы. Антисмысловая цепь ДНК: 3′-T A C T G A C T G A C G A T C-5 ‘
- Предсказать эффект изменения последовательности нуклеотидов в области –35 бактериального промотора.
Активация транскрипции с одноцепочечной ДНК.
Proc Natl Acad Sci U S A. 1996 11 июня; 93 (12): 5830–5835.
1996 11 июня; 93 (12): 5830–5835.
Лаборатория патологии, Национальный институт рака, Национальные институты здравоохранения, Бетесда, Мэриленд 20892, США.
Эта статья цитируется в других статьях в PMC.Abstract
Последовательно-специфические регуляторы экспрессии эукариотических генов аксиоматически действуют через мишени двухцепочечной ДНК. Белки, которые распознают цис-элементы ДНК как одиночные цепи, но для которых не хватало убедительных доказательств, указывающих на участие in vivo в транскрипции, остаются в этой схеме «сиротами».Мы стремились определить, могут ли специфичные для последовательности одноцепочечные связывающие белки находить свои родственные элементы и изменять транскрипцию in vivo, изучая гетерогенный ядерный рибонуклеопротеин K (hnRNP K), который связывает одноцепочечную последовательность (CCCTCCCCA; CT-element) человеческого c- ген myc in vitro. Чтобы контролировать его связывание с ДНК in vivo, способность hnRNP K активировать репортерный ген была усилена путем слияния с доменом трансактивации VP16. Было обнаружено, что этот химерный белок трансактивирует репортеры, управляемые кольцевыми, но не линейными CT-элементами, что позволяет предположить, что hnRNP K распознает одноцепочечный участок, образованный в результате отрицательной суперспирализации в кольцевой плазмиде.Когда CT-элементы были сконструированы так, чтобы перекрываться с операторами lexA, добавление белка lexA, либо in vivo, либо in vitro, аннулировало связывание hnRNP K, скорее всего, путем предотвращения образования одиночной цепи. Эти результаты не только показывают, что hnRNP K является однонитевым ДНК-связывающим белком in vivo, но и демонстрируют, как сегмент ДНК может изменять транскрипционную активность соседнего гена посредством взаимного преобразования дуплекса и одиночных цепей.
Было обнаружено, что этот химерный белок трансактивирует репортеры, управляемые кольцевыми, но не линейными CT-элементами, что позволяет предположить, что hnRNP K распознает одноцепочечный участок, образованный в результате отрицательной суперспирализации в кольцевой плазмиде.Когда CT-элементы были сконструированы так, чтобы перекрываться с операторами lexA, добавление белка lexA, либо in vivo, либо in vitro, аннулировало связывание hnRNP K, скорее всего, путем предотвращения образования одиночной цепи. Эти результаты не только показывают, что hnRNP K является однонитевым ДНК-связывающим белком in vivo, но и демонстрируют, как сегмент ДНК может изменять транскрипционную активность соседнего гена посредством взаимного преобразования дуплекса и одиночных цепей.
Полный текст
Полный текст доступен в виде отсканированной копии оригинальной печатной версии.Получите копию для печати (файл PDF) полной статьи (2,1 Мбайт) или щелкните изображение страницы ниже, чтобы просмотреть страницу за страницей. Ссылки на PubMed также доступны для Избранные ссылки .
Ссылки на PubMed также доступны для Избранные ссылки .
Изображения в этой статье
Щелкните изображение, чтобы увидеть его в увеличенном виде.
Избранные ссылки
Эти ссылки находятся в PubMed. Это может быть не полный список ссылок из этой статьи.
- Крик Ф. Общая модель хромосом высших организмов.Природа. 1971, 5 ноября, 234 (5323): 25–27. [PubMed] [Google Scholar]
- Ларсен А., Вайнтрауб Х. Измененная конформация ДНК, обнаруженная нуклеазой S1, встречается в определенных областях в активном хроматине куриного глобина. Клетка. Июнь 1982 г., 29 (2): 609–622. [PubMed] [Google Scholar]
- Шон Э., Эванс Т., Уэлш Дж., Эфстратиадис А. Конформация промоторной ДНК: точное картирование S1-гиперчувствительных сайтов. Клетка. 1983 декабрь; 35 (3, часть 2): 837–848. [PubMed] [Google Scholar]
- Grosso LE, Pitot HC. Регуляция транскрипции c-myc во время химически индуцированной дифференцировки культур HL-60.
 Cancer Res. 1985 Февраль; 45 (2): 847–850. [PubMed] [Google Scholar]
Cancer Res. 1985 Февраль; 45 (2): 847–850. [PubMed] [Google Scholar] - Раджавашист Т. Б., Тейлор А. К., Андалиби А., Свенсон К. Л., Лусис А. Дж.. Идентификация белка цинкового пальца, который связывается с регуляторным элементом стерола. Наука. 1989, 11 августа; 245 (4918): 640–643. [PubMed] [Google Scholar]
- Бергеманн А.Д., Джонсон Э.М. Фактор HeLa Pur связывает одноцепочечную ДНК в конкретном элементе, консервативном во фланкирующих областях генов и источниках репликации ДНК. Mol Cell Biol. Март 1992 г., 12 (3): 1257–1265.[Бесплатная статья PMC] [PubMed] [Google Scholar]
- Tada H, Khalili K. Новый ДНК-связывающий белок, специфичный к последовательности, LCP-1, взаимодействует с одноцепочечной ДНК и дифференциально регулирует экспрессию ранних генов нейротропного JC вирус. J Virol. 1992 декабрь; 66 (12): 6885–6892. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Алтиок С., Гронер Б. Взаимодействие двух специфичных для последовательности одноцепочечных ДНК-связывающих белков с важной областью промотора гена бета-казеина регулируется лактогенными гормонами.
 Mol Cell Biol. 1993 декабрь; 13 (12): 7303–7310. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Mol Cell Biol. 1993 декабрь; 13 (12): 7303–7310. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Gaillard C, Cabannes E, Strauss F. Идентичность РНК-связывающего белка K частиц hnRNP с белком h26, специфичным для последовательности одноцепочечным ДНК-связывающим белком. Nucleic Acids Res. 1994, 11 октября; 22 (20): 4183–4186. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Takai T, Nishita Y, Iguchi-Ariga SM, Ariga H. Молекулярное клонирование MSSP-2, однонитевого связывающего белка гена c-myc: характеристика специфичности связывания и активность репликации ДНК.Nucleic Acids Res. 1994 25 декабря; 22 (25): 5576–5581. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Negishi Y, Nishita Y, Saëgusa Y, Kakizaki I, Galli I, Kihara F, Tamai K, Miyajima N, Iguchi-Ariga SM, Ariga H. Идентификация и клонирование кДНК одноцепочечных ДНК-связывающих белков, которые взаимодействуют с областью выше человеческого гена c-myc. Онкоген. 1994 Апрель; 9 (4): 1133–1143. [PubMed] [Google Scholar]
- Michelotti EF, Tomonaga T, Krutzsch H, Levens D.
 Белок, связывающий клеточные нуклеиновые кислоты, регулирует CT-элемент протоонкогена c-myc человека.J Biol Chem. 1995 21 апреля; 270 (16): 9494–9499. [PubMed] [Google Scholar]
Белок, связывающий клеточные нуклеиновые кислоты, регулирует CT-элемент протоонкогена c-myc человека.J Biol Chem. 1995 21 апреля; 270 (16): 9494–9499. [PubMed] [Google Scholar] - Гроссманн М.Э., Тиндалл ди-джей. Рецептор андрогена транскрипционно подавляется белками, связывающими одноцепочечную ДНК. J Biol Chem. 1995 5 мая; 270 (18): 10968–10975. [PubMed] [Google Scholar]
- Такимото М., Томонага Т., Матунис М., Авиган М., Крутч Х., Дрейфус Г., Левенс Д. Специфическое связывание гетерогенного белка К частиц рибонуклеопротеина с промотором c-myc человека in vitro. J Biol Chem. 1993 25 августа; 268 (24): 18249–18258.[PubMed] [Google Scholar]
- Томонага Т., Левенс Д. Гетерогенный ядерный рибонуклеопротеин К представляет собой ДНК-связывающий трансактиватор. J Biol Chem. 1995 г., 3 марта; 270 (9): 4875–4881. [PubMed] [Google Scholar]
- Swanson MS, Dreyfuss G. Классификация и очистка белков гетерогенных ядерных рибонуклеопротеиновых частиц по специфичности связывания РНК.
 Mol Cell Biol. 1988 Май; 8 (5): 2237–2241. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Mol Cell Biol. 1988 Май; 8 (5): 2237–2241. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Matunis MJ, Michael WM, Dreyfuss G.Характеристика и первичная структура поли (C) -связывающего гетерогенного ядерного рибонуклеопротеинового комплекса K-белка. Mol Cell Biol. 1992 Янв; 12 (1): 164–171. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Микелотти Э. Ф., Микелотти Г. А., Аронсон А. И., Левенс Д. Гетерогенный ядерный рибонуклеопротеин К является фактором транскрипции. Mol Cell Biol. 1996 Май; 16 (5): 2350–2360. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Taylor SJ, Shalloway D. РНК-связывающий белок, связанный с Src через его домены Sh3 и Sh4 в митозе.Природа. 1994 28 апреля; 368 (6474): 867–871. [PubMed] [Google Scholar]
- Садовски И., Пташне М. Вектор для экспрессии слитых белков GAL4 (1-147) в клетках млекопитающих. Nucleic Acids Res. 1989 25 сентября; 17 (18): 7539–7539. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Lillie JW, Green MR.
 Активация транскрипции белком E1a аденовируса. Природа. 2 марта 1989 г., 338 (6210): 39–44. [PubMed] [Google Scholar]
Активация транскрипции белком E1a аденовируса. Природа. 2 марта 1989 г., 338 (6210): 39–44. [PubMed] [Google Scholar] - Дункан Р., Базар Л., Микелотти Г., Томонага Т., Крутч Х., Авиган М., Левенс Д.Последовательно-специфический одноцепочечный связывающий белок активирует расположенный далеко вверх по течению элемент c-myc и определяет новый ДНК-связывающий мотив. Genes Dev. 1994 15 февраля; 8 (4): 465–480. [PubMed] [Google Scholar]
- Michelotti GA, Michelotti EF, Pullner A, Duncan RC, Eick D, Levens D. Множественные одноцепочечные цис-элементы связаны с активированным хроматином гена c-myc человека in vivo. Mol Cell Biol. 1996 июн; 16 (6): 2656–2669. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Garrity PA, Wold BJ.Эффекты различных ДНК-полимераз в ПЦР, опосредованной лигированием: усиленное геномное секвенирование и футпринтинг in vivo. Proc Natl Acad Sci U S. A. 1992, 1 февраля; 89 (3): 1021–1025. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Лю Л.
 Ф., Ван Дж.С. Суперспирализация матрицы ДНК во время транскрипции. Proc Natl Acad Sci U S. A. 1987, октябрь; 84 (20): 7024–7027. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Ф., Ван Дж.С. Суперспирализация матрицы ДНК во время транскрипции. Proc Natl Acad Sci U S. A. 1987, октябрь; 84 (20): 7024–7027. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Hirt B. Селективная экстракция ДНК полиомы из инфицированных культур клеток мыши. J Mol Biol.14 июня 1967 г .; 26 (2): 365–369. [PubMed] [Google Scholar]
- Вайнтрауб Х., Ченг П.Ф., Конрад К. Экспрессия трансфицированной ДНК зависит от топологии ДНК. Клетка. 4 июля 1986 г.; 46 (1): 115–122. [PubMed] [Google Scholar]
- DesJardins E, Hay N. Повторяющиеся CT-элементы, связанные белками цинковых пальцев, контролируют абсолютную и относительную активность двух основных промоторов c-myc человека. Mol Cell Biol. 1993 сентябрь; 13 (9): 5710–5724. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Saffer JD, Jackson SP, Thurston SJ.SV40 стимулирует экспрессию трансактирующего фактора Sp1 на уровне мРНК. Genes Dev. 1990 апр; 4 (4): 659–666. [PubMed] [Google Scholar]
- Gilinger G, Alwine JC.
 Активация транскрипции большим Т-антигеном вируса обезьяны 40: требования для простых промоторных структур, содержащих либо ТАТА, либо инициаторные элементы с вариабельными сайтами связывания вышестоящих факторов. J Virol. 1993 ноябрь; 67 (11): 6682–6688. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Активация транскрипции большим Т-антигеном вируса обезьяны 40: требования для простых промоторных структур, содержащих либо ТАТА, либо инициаторные элементы с вариабельными сайтами связывания вышестоящих факторов. J Virol. 1993 ноябрь; 67 (11): 6682–6688. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Rice PW, Cole CN. Эффективная активация транскрипции многих простых модульных промоторов большим Т-антигеном вируса обезьяны 40.J Virol. 1993 ноябрь; 67 (11): 6689–6697. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Duncan R, Collins I, Tomonaga T, Zhang T, Levens D. Уникальный мотив последовательности трансактивации обнаружен в карбоксиконцевом домене одноцепочечного связывающего белка FBP. Mol Cell Biol. 1996 Май; 16 (5): 2274–2282. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Kahn JD, Yun E, Crothers DM. Обнаружение локальной гибкости ДНК. Природа. 1994 10 марта; 368 (6467): 163–166. [PubMed] [Google Scholar]
- Парвин Дж. Д., Шарп, Пенсильвания.
 Топология ДНК и минимальный набор базальных факторов для транскрипции РНК-полимеразой II. Клетка. 1993 7 мая; 73 (3): 533–540. [PubMed] [Google Scholar]
Топология ДНК и минимальный набор базальных факторов для транскрипции РНК-полимеразой II. Клетка. 1993 7 мая; 73 (3): 533–540. [PubMed] [Google Scholar] - Парвин Дж. Д., Шикинд Б. М., Мейерс Р. Э., Ким Дж., Шарп П. А.. Множественные наборы базальных факторов инициируют транскрипцию РНК-полимеразой II. J Biol Chem. 15 июля 1994 г .; 269 (28): 18414–18421. [PubMed] [Google Scholar]
Статьи из материалов Национальной академии наук Соединенных Штатов Америки любезно предоставлены Национальной академией наук
расшифровка | Определение, шаги и биология
Транскрипция , синтез РНК из ДНК.Генетическая информация перетекает из ДНК в белок, вещество, придающее организму его форму. Этот поток информации происходит через последовательные процессы транскрипции (ДНК в РНК) и трансляции (РНК в белок). Транскрипция происходит, когда есть потребность в конкретном генном продукте в определенное время или в определенной ткани.
Гены состоят из промоторных областей и чередующихся областей интронов (некодирующие последовательности) и экзонов (кодирующие последовательности).Производство функционального белка включает транскрипцию гена из ДНК в РНК, удаление интронов и сплайсинг экзонов, трансляцию сплайсированных последовательностей РНК в цепочку аминокислот и посттрансляционную модификацию белковой молекулы.
Британская энциклопедия, Inc.Подробнее по этой теме
Нуклеиновая кислота: Транскрипция
Небольшие сегменты ДНК транскрибируются в РНК ферментом РНК-полимеразой, которая осуществляет это копирование в строго контролируемом процессе….
Во время транскрипции обычно копируется только одна цепь ДНК. Это называется цепочкой-матрицей, а полученные молекулы РНК представляют собой одноцепочечные информационные РНК (мРНК). Нить ДНК, которая соответствовала бы мРНК, называется кодирующей или смысловой цепью.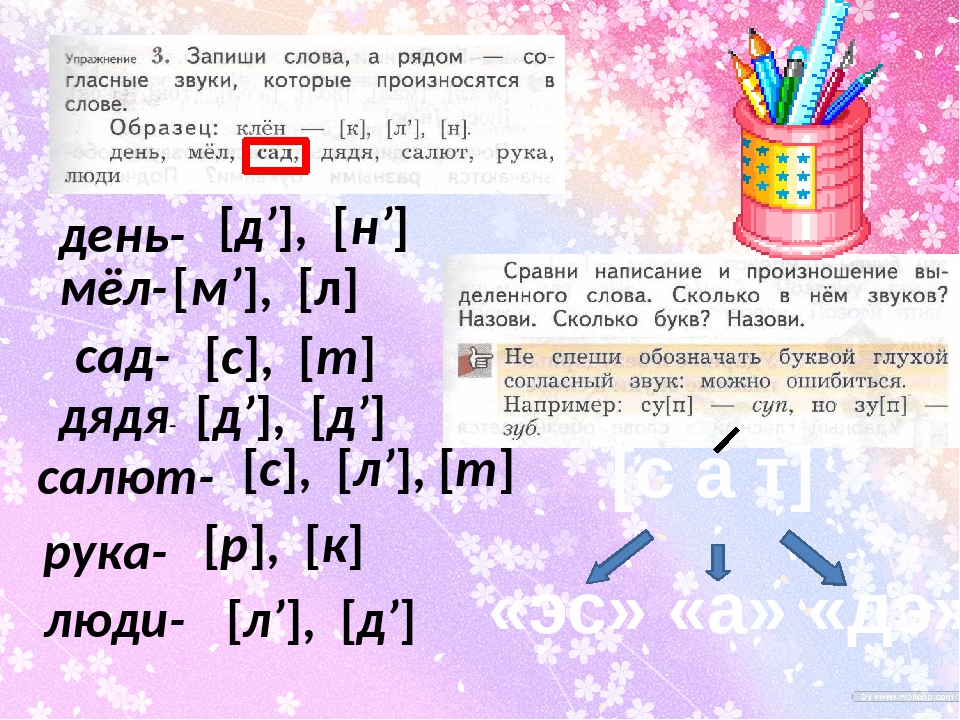 У эукариот (организмов, обладающих ядром) исходный продукт транскрипции называется пре-мРНК. Пре-мРНК широко редактируется посредством сплайсинга до того, как зрелая мРНК будет продуцирована и готова к трансляции рибосомой, клеточной органеллой, которая служит местом синтеза белка.Транскрипция любого гена происходит в хромосомном месте этого гена, который является относительно коротким сегментом хромосомы. Активная транскрипция гена зависит от необходимости активности этого конкретного гена в конкретной клетке или ткани или в данное время.
У эукариот (организмов, обладающих ядром) исходный продукт транскрипции называется пре-мРНК. Пре-мРНК широко редактируется посредством сплайсинга до того, как зрелая мРНК будет продуцирована и готова к трансляции рибосомой, клеточной органеллой, которая служит местом синтеза белка.Транскрипция любого гена происходит в хромосомном месте этого гена, который является относительно коротким сегментом хромосомы. Активная транскрипция гена зависит от необходимости активности этого конкретного гена в конкретной клетке или ткани или в данное время.
Небольшие сегменты ДНК транскрибируются в РНК ферментом РНК-полимеразой, которая осуществляет это копирование в строго контролируемом процессе. Первым шагом является распознавание определенной последовательности ДНК, называемой промотором, которая означает начало гена.В этот момент две нити ДНК разделяются, и РНК-полимераза начинает копирование с определенной точки на одной нити ДНК, используя особый тип сахаросодержащего нуклеозида, называемого рибонуклеозид-5’-трифосфатом, для начала растущей цепи. Дополнительные рибонуклеозидтрифосфаты используются в качестве субстрата, и за счет расщепления их высокоэнергетической фосфатной связи монофосфаты рибонуклеозидов включаются в растущую цепь РНК. Каждый последующий рибонуклеотид управляется правилами комплементарного спаривания оснований ДНК.Например, C (цитозин) в ДНК направляет включение G (гуанина) в РНК. Точно так же G в ДНК копируется в C в РНК, T (тимин) в A (аденин) и A в U (урацил; РНК содержит U вместо T ДНК). Синтез продолжается до тех пор, пока не будет достигнут сигнал обрыва, после чего РНК-полимераза отпадет от ДНК, и молекула РНК высвободится.
Дополнительные рибонуклеозидтрифосфаты используются в качестве субстрата, и за счет расщепления их высокоэнергетической фосфатной связи монофосфаты рибонуклеозидов включаются в растущую цепь РНК. Каждый последующий рибонуклеотид управляется правилами комплементарного спаривания оснований ДНК.Например, C (цитозин) в ДНК направляет включение G (гуанина) в РНК. Точно так же G в ДНК копируется в C в РНК, T (тимин) в A (аденин) и A в U (урацил; РНК содержит U вместо T ДНК). Синтез продолжается до тех пор, пока не будет достигнут сигнал обрыва, после чего РНК-полимераза отпадет от ДНК, и молекула РНК высвободится.
Перед многими генами прокариот (организмов, лишенных ядра) существуют сигналы, называемые «операторами» ( см. Опероны ), где специализированные белки, называемые репрессорами, связываются с ДНК непосредственно перед начальной точкой транскрипции и препятствуют доступу к ней. ДНК с помощью РНК-полимеразы.Таким образом, эти репрессорные белки предотвращают транскрипцию гена, физически блокируя действие РНК-полимеразы. Обычно репрессоры освобождаются от своего блокирующего действия, когда они получают сигналы от других молекул в клетке, указывающие на необходимость экспрессии гена. Перед некоторыми прокариотическими генами идут сигналы, с которыми связываются белки-активаторы, чтобы стимулировать транскрипцию.
Обычно репрессоры освобождаются от своего блокирующего действия, когда они получают сигналы от других молекул в клетке, указывающие на необходимость экспрессии гена. Перед некоторыми прокариотическими генами идут сигналы, с которыми связываются белки-активаторы, чтобы стимулировать транскрипцию.
Модель оперона и ее связь с регулятором гена.
Encyclopædia Britannica, Inc. Получите подписку Britannica Premium и получите доступ к эксклюзивному контенту. Подпишитесь сейчас Транскрипция у эукариот сложнее, чем у прокариот. Во-первых, РНК-полимераза высших организмов является более сложным ферментом, чем относительно простой фермент из пяти субъединиц прокариот. Кроме того, существует множество дополнительных факторов, которые помогают контролировать эффективность отдельных промоторов. Эти дополнительные белки называются факторами транскрипции и обычно отвечают на сигналы внутри клетки, которые указывают, требуется ли транскрипция.Во многих генах человека могут потребоваться несколько факторов транскрипции, прежде чем транскрипция сможет протекать эффективно. Фактор транскрипции может вызывать репрессию или активацию экспрессии генов у эукариот.
Фактор транскрипции может вызывать репрессию или активацию экспрессии генов у эукариот.
Транскрипция
Создано Джорджем Райсом, Государственный университет Монтаны
В общем, ДНК реплицируется путем раскручивания спирали, разделения цепей путем разрыва водородных связей между комплементарными цепями и синтеза двух новых цепей путем спаривания комплементарных оснований.Репликация начинается в определенном участке ДНК, который называется точкой начала репликации. Репликация ДНК является двунаправленной от точки начала репликации. Чтобы начать репликацию ДНК, раскручивающие ферменты, называемые ДНК-геликазами, заставляют две родительские цепи ДНК раскручиваться и отделяться друг от друга в точке начала репликации с образованием двух Y-образных вилок репликации. Эти репликационные вилки и являются фактическим местом копирования ДНК. Белки, дестабилизирующие спираль, связываются с одноцепочечными областями, поэтому две цепи не соединяются повторно. Ферменты, называемые топоизимеразами, производят разрывы в ДНК, а затем воссоединяются с ними, чтобы снять напряжение в спиральной молекуле во время репликации. Поскольку цепи продолжают раскручиваться и разделяться в обоих направлениях вокруг всей молекулы ДНК, водородная связь свободных нуклеотидов ДНК с нуклеотидами на каждой родительской цепи дает новые комплементарные цепи. Поскольку новые нуклеотиды выстраиваются в линию напротив каждой родительской цепи за счет водородных связей, ферменты, называемые ДНК-полимеразами, присоединяются к нуклеотидам посредством фосфодиэфирных связей.Фактически, нуклеотиды, выстраивающиеся путем комплементарного спаривания оснований, представляют собой дезоксинуклеозидтрифосфаты, состоящие из азотистого основания, дезоксирибозы и трех фосфатов. Когда фосфодиэфирная связь образуется между 5′-фосфатной группой нового нуклеотида и 3′-ОН последнего нуклеотида в цепи ДНК, два фосфата удаляются, обеспечивая энергию для связывания. В конце концов, каждая родительская цепь служит шаблоном для синтеза комплементарной копии самой себя, в результате чего образуются две идентичные молекулы ДНК.
Ферменты, называемые топоизимеразами, производят разрывы в ДНК, а затем воссоединяются с ними, чтобы снять напряжение в спиральной молекуле во время репликации. Поскольку цепи продолжают раскручиваться и разделяться в обоих направлениях вокруг всей молекулы ДНК, водородная связь свободных нуклеотидов ДНК с нуклеотидами на каждой родительской цепи дает новые комплементарные цепи. Поскольку новые нуклеотиды выстраиваются в линию напротив каждой родительской цепи за счет водородных связей, ферменты, называемые ДНК-полимеразами, присоединяются к нуклеотидам посредством фосфодиэфирных связей.Фактически, нуклеотиды, выстраивающиеся путем комплементарного спаривания оснований, представляют собой дезоксинуклеозидтрифосфаты, состоящие из азотистого основания, дезоксирибозы и трех фосфатов. Когда фосфодиэфирная связь образуется между 5′-фосфатной группой нового нуклеотида и 3′-ОН последнего нуклеотида в цепи ДНК, два фосфата удаляются, обеспечивая энергию для связывания. В конце концов, каждая родительская цепь служит шаблоном для синтеза комплементарной копии самой себя, в результате чего образуются две идентичные молекулы ДНК.
Транскрипция — это процесс, посредством которого последовательность ДНК ферментативно копируется РНК-полимеразой с образованием комплементарной РНК. Или, другими словами, перенос генетической информации из ДНК в РНК. В случае ДНК, кодирующей белок, транскрипция является началом процесса, который в конечном итоге приводит к трансляции генетического кода (через промежуточное звено мРНК ) в функциональный пептид или белок. Транскрипция имеет некоторые механизмы проверки, но они менее эффективны и менее эффективны, чем контроль ДНК; следовательно, транскрипция имеет более низкую точность копирования, чем репликация ДНК.Подобно репликации ДНК, транскрипция происходит в направлении от 5 ‘до 3’ (т. Е. Старый полимер читается в направлении от 3 ‘до 5’, а новые комплементарные фрагменты генерируются в направлении от 5 ‘до 3’). Транскрипция делится на 3 стадии: инициация, элонгация и завершение.
Типы РНК:
- мРНК — информационная РНК является копией гена.
 Он действует как фотокопия гена, имея последовательность, комплементарную одной цепи ДНК и идентичную другой цепи.МРНК действует как помощник по телефону, чтобы нести информацию, хранящуюся в ДНК в ядре, в цитоплазму, где рибосомы могут превратить ее в белок.
Он действует как фотокопия гена, имея последовательность, комплементарную одной цепи ДНК и идентичную другой цепи.МРНК действует как помощник по телефону, чтобы нести информацию, хранящуюся в ДНК в ядре, в цитоплазму, где рибосомы могут превратить ее в белок. - тРНК — транспортная РНК — это небольшая РНК, которая имеет очень специфическую вторичную и третичную структуру, так что она может связывать аминокислоту на одном конце и мРНК на другом конце. Он действует как адаптер для переноса аминокислотных элементов белка в соответствующее место, как это закодировано мРНК.
- рРНК — рибосомная РНК является одним из структурных компонентов рибосомы.Его последовательность представляет собой набор участков мРНК, так что рибосома может соответствовать и связываться с мРНК, из которой она будет производить белок.
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно.Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом.Если часы вашего компьютера показывают дату до 1 января 1970 г.,
браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.

- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу.Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт
не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к
остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Например, сайт
не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к
остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
| Thermo Fisher Scientific
Для экспериментов в области молекулярной биологии обратная транскрипция в первую очередь выполняется для создания комплементарных ДНК (кДНК), представляющих популяции тканевых или клеточно-специфичных РНК.Чтобы эксперименты были успешными, необходимо учитывать критические факторы, связанные с шаблоном, реагентами и условиями реакции.
Видео: Упрощенная обратная транскрипция
Узнайте, как работает обратная транскрипция и как выбрать правильные обратные транскриптазы и праймеры для вашего эксперимента.
Подготовка матрицы РНК
РНК служит матрицей при обратной транскрипции. Общая РНК обычно используется в синтезе кДНК для последующих приложений, таких как ОТ- (q) ПЦР, тогда как определенные типы РНК (например, РНК). например, информационная РНК (мРНК) и малые РНК, такие как миРНК) могут быть обогащены для определенных приложений, таких как создание библиотеки кДНК и профилирование миРНК.
например, информационная РНК (мРНК) и малые РНК, такие как миРНК) могут быть обогащены для определенных приложений, таких как создание библиотеки кДНК и профилирование миРНК.
Поддержание целостности РНК имеет решающее значение и требует особых мер предосторожности при извлечении, обработке, хранении и экспериментальном использовании. Лучшие методы предотвращения деградации РНК включают ношение перчаток, пипетирование с наконечниками, защищающими от аэрозоля, использование лабораторного оборудования и реагентов, не содержащих нуклеазы, а также дезинфекцию рабочих зон.
Для выделения и очистки РНК доступны различные стратегии в зависимости от типа исходных материалов (например,ж., кровь, ткани, клетки, растения) и цели экспериментов. Основными целями рабочих процессов выделения являются стабилизация молекул РНК, ингибирование РНКаз и максимальное увеличение урожая с помощью надлежащих методов хранения и экстракции. Оптимальные методы очистки удаляют из тканей растений эндогенные соединения, такие как сложные полисахариды и гуминовую кислоту, которые мешают активности ферментов, а также обычные ингибиторы обратных транскриптаз, такие как соли, ионы металлов, этанол и фенол. После очистки РНК следует хранить при –80 ° C с минимальными циклами замораживания-оттаивания.
После очистки РНК следует хранить при –80 ° C с минимальными циклами замораживания-оттаивания.
Видео: Ингибиторы в реакции обратной транскрипции
Узнайте об ингибиторах обратной транскриптазы, их механизмах ингибирования и советах по преодолению ингибиторов обратной транскрипции.
Существует несколько методов оценки качества и количества РНК после очистки. Распространенным подходом является УФ-спектроскопия для измерения оптической плотности на определенных длинах волн. Количество РНК можно определить по оптической плотности при 260 нм с использованием закона Бера-Ламберта, а наличие или отсутствие конкретных примесей можно сделать вывод из соотношений оптической плотности на разных длинах волн (, таблица 1, ).Обратите внимание, что УФ-поглощение не является специфическим только для РНК, поскольку все нуклеиновые кислоты поглощают УФ-излучение на одинаковых длинах волн. Для более специфичного и чувствительного анализа РНК могут быть рассмотрены флуоресцентные анализы с красителями, которые излучают сигнал только при специфическом связывании с молекулами-мишенями.
Видео: Что такое коэффициент чистоты?
Узнайте об определении чистоты образца по оптической плотности и вычислении соотношений A260 / A280.
Таблица 1. Рекомендации по измерению УФ-излучения для анализа РНК
| Поглощение | Указывает на присутствие: | Целевые отношения | ||
|---|---|---|---|---|
| 230 нм | Органические соединения, сахара, мочевина, соли | 260617 A 260617 A 90 А 230 > 1.8|||
| 260 нм | Все нуклеиновые кислоты | A 260 ≈ 0,1–1,0 | ||
| 270 нм | Фенол | A 260 / A 270 > 1,0 | Белок | РНК: A 260 / A 280 ≈ 2,0 ДНК: A 260 / A 280 ≈ 1,8 |
| 330 нм | Рассеяние света | 23 A 330 330 |
Целостность РНК может быть оценена путем сравнения 28S и 18S рибосомных РНК (рРНК) как представления общей РНК.Общая РНК денатурируется, а затем разделяется по размеру с помощью гель-электрофореза, который носит качественный характер. Затем оценивается отношение интенсивностей 28S рРНК к 18S рРНК с соотношением 2: 1, указывающим на интактную РНК (, фиг. 1A, ). Более количественный метод, разработанный Agilent Technologies, объединяет микрофлюидику и собственный алгоритм для оценки целостности РНК. Этот метод производит цифровое считывание, называемое числом целостности РНК, или RIN, где значения в диапазоне от 8 до 10 указывают на высокое качество РНК [1,2] (, рис. 1B, ).
Рисунок 1. Анализ целостности РНК с помощью (A) гель-электрофореза и (B) микрофлюидики.
Удаление геномной ДНК
Иногда следовые количества геномной ДНК (гДНК) могут быть очищены совместно с РНК. Загрязняющая гДНК может мешать обратной транскрипции и может привести к ложноположительным результатам, более высокому фону или более низкому обнаружению в чувствительных приложениях, таких как RT-qPCR.
ДНКаза I обычно добавляется к выделенной РНК для удаления гДНК.Перед ОТ-ПЦР необходимо полностью удалить ДНКазу I, поскольку любой остаточный фермент может разрушить одноцепочечную ДНК, такую как праймеры и синтезированную кДНК. Часто инактивация ДНКазы I (например, обработка ЭДТА и нагреванием) или процедуры удаления ферментов приводят к деградации РНК или потере образца.
В качестве альтернативы ДНКазе I доступны двухцепочечные ДНКазы, которые устраняют загрязняющую гДНК без воздействия на РНК или одноцепочечные ДНК. Их термолабильные свойства позволяют просто инактивировать их при относительно умеренной температуре (например,г., 55 ° C) без негативных воздействий. Такие двухцепочечные термолабильные ДНКазы можно инкубировать с РНК в течение 2 минут при 37 ° C перед реакциями обратной транскрипции, чтобы упростить рабочий процесс (, рис. 2, ).
Рис. 2. Процедура удаления гДНК: ДНКаза I против фермента Invitrogen ezDNase. По сравнению с ДНКазой I фермент ezDNase предлагает более короткий рабочий процесс, более простую процедуру и меньшее повреждение РНК. Инактивация фермента ezDNase перед обратной транскрипцией необязательна, поскольку фермент не расщепляет праймеры, ssRNA или комплексы кДНК: РНК.Соображения по поводу обратной транскриптазы
Класс ферментов, известных как обратные транскриптазы, синтезирует комплементарную ДНК с использованием РНК в качестве матрицы, но они могут различаться по функциональной активности и свойствам. Их свойства влияют на их способность к обратной транскрипции длинных транскриптов РНК, GC-богатой РНК, РНК со значительными вторичными структурами и РНК субоптимального качества. ( White paper: Сравнение производительности обратных транскриптаз)
Большинство обратных транскриптаз, используемых в молекулярной биологии, происходят от гена pol вируса миелобластоза птиц (AMV) или вируса лейкемии мышей Молони (MMLV).Обратная транскриптаза AMV была одним из первых ферментов, выделенных для синтеза кДНК в лаборатории. Фермент представляет собой гетеродимер массой 170 кДа с оптимальным диапазоном температур реакции 42–48 ° C. Обратная транскриптаза AMV обладает сильной активностью РНКазы H, которая расщепляет РНК в гибридах РНК: кДНК, что приводит к более коротким фрагментам кДНК (<5 т.п.н.).
Обратная транскриптаза MMLV стала популярной альтернативой благодаря своей мономерной структуре, которая позволила упростить клонирование и модификации рекомбинантного фермента.Обратная транскриптаза MMLV представляет собой фермент массой 75 кДа с оптимальной температурой реакции около 37 ° C. Хотя она менее термостабильна, чем обратная транскриптаза AMV, обратная транскриптаза MMLV способна синтезировать более длинную кДНК (<7 т.п.н.) с более высокой эффективностью из-за более низкой активности РНКазы H [3].
Для дальнейшего улучшения синтеза кДНК была разработана обратная транскриптаза MMLV, обеспечивающая еще более низкую активность РНКазы H (т. Е. Мутированный домен РНКазы H или RNaseH —), более высокую термостабильность (до 55 ° C) и повышенную процессивность (65 раз выше).Эти свойства приводят к увеличению длины и выхода кДНК, более высокой чувствительности, повышенной устойчивости к ингибиторам и более быстрому времени реакции (, таблица 2, ) [4]. ( Подробнее: атрибутов обратной транскриптазы)
Таблица 2. Распространенные обратные транскриптазы и их атрибуты.| Обратная транскриптаза AMV | Обратная транскриптаза MMLV | Инженерная обратная транскриптаза MMLV (например, обратная транскриптаза Invitrogen SuperScript IV) | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 6 Высокая активность RNase H4 | ||||||||||||||||||||||||||
| Температура реакции (максимальная рекомендуемая) | 42 ° C | 37 ° C | 55 ° C | |||||||||||||||||||||||
| Время реакции | 60 мин | 60 мин | ||||||||||||||||||||||||
| Целевая длина | ≤5 kb | ≤7 kb | ≤12 kb | |||||||||||||||||||||||
| Относительный выход (со сложной или субоптимальной РНК) | Средний | Низкий | Высокий
| Oligo (dT) | Случайные гексамеры | Oligo (dT) + случайные гексамеры | Праймер для генов | |
|---|---|---|---|---|
| Типичная конечная концентрация98 2 –5 мкМ | 1–2 мкМ каждый | 0.5–1 мкМ | ||
| Удлинение препраймера при 25 ° C | — | — | ||
| Ключевые преимущества | Полноразмерная обратная транскрипция RNA ) tail | Обратная транскрипция большинства видов РНК, включая деградированную РНК | Комбинированные преимущества олиго (dT) и случайных праймеров | Обратная транскрипция, специфичная для интересующего гена |
Основные компоненты реакции
Помимо фермента и праймеров, основные компоненты реакции для обратной транскрипции включают матрицу РНК (предварительно обработанную для удаления геномной ДНК), буфер, dNTP, DTT, ингибитор РНКазы и воду, не содержащую РНКаз ( Рисунок 5 ).
Рис. 5. Реакция обратной транскрипции и ее основные компоненты.
- Шаблон РНК получают, как описано в предыдущих разделах. Таблица 4 предоставляет рекомендуемый диапазон входной РНК для реакций обратной транскрипции, а оптимальное количество зависит от распространенности целевой последовательности и чувствительности обратной транскриптазы.
| Шаблон РНК | Рекомендуемый диапазон |
|---|---|
| Общая РНК | 10 мкг – 5 мкг |
| Поли (A) РНК | 10 мкг – 0,5 мкг |
- Реакционный буфер поддерживает подходящие для реакции pH и ионную силу. Поставляемый буфер может также содержать добавки для повышения эффективности обратной транскрипции.
- dNTP обычно должен быть в концентрации 0,5–1 мМ каждый, предпочтительно в эквимолярных концентрациях. Для обеспечения эффективной обратной транскрипции рекомендуются свежеразведенные высококачественные дНТФ.
- DTT , восстанавливающий реагент, часто включается для оптимальной активности ферментов. Эффективность реакции может быть снижена, если DTT или другие добавки выпадают в осадок; следовательно, компоненты реакции должны быть растворены и хорошо перемешаны.
- Ингибитор РНКазы обычно включается в реакционный буфер или добавляется в реакцию обратной транскрипции для предотвращения деградации РНК.РНКазы могли быть совместно очищены во время выделения или введены во время настройки реакции. Существует ряд известных РНКаз, и соответствующие ингибиторы РНКаз следует выбирать в зависимости от способа их действия и требований к реакции.
- Вода, используемая в реакциях обратной транскрипции, не должна содержать нуклеаз. Рекомендуется вода, не содержащая нуклеаз, из коммерческого источника или вода, обработанная DEPC (диэтилпирокарбонатом) для удаления любых РНКаз. Загрязняющие РНКазы не могут быть удалены простой фильтрацией, а автоклавированной воды недостаточно, поскольку РНКазы термостабильны.
Соображения температуры и времени реакции
Реакции обратной транскрипции включают три основных этапа: отжиг праймера, полимеризацию ДНК и дезактивацию фермента ( Рисунок 6 ). Температура и продолжительность этих этапов зависят от выбора праймера, целевой РНК и обратной транскриптазы.
Рисунок 6. Три основных этапа синтеза кДНК.
- Отжиг праймеров: Праймер смешивают с РНК-матрицей, нагревают до 65 ° C в течение 5 минут, затем инкубируют на льду не менее 1 минуты.Это помогает гарантировать, что РНК является одноцепочечной и что праймер эффективно отжигается с мишенью. После отжига добавляют обратную транскриптазу и необходимые компоненты (например, буфер, dNTP, ингибитор РНКазы).
- Полимеризация ДНК: На этой стадии температура и продолжительность реакции могут варьироваться в зависимости от выбора праймера и используемой обратной транскриптазы. С олиго (dT) праймером (Tm ~ 35-50 ° C) реакцию можно инкубировать непосредственно при оптимальной температуре обратной транскриптазы (37-50 ° C).Случайные гексамеры обычно имеют более низкую Tm (~ 10–15 ° C) из-за их более короткой длины. Поэтому, когда используются случайные гексамеры (отдельно или в комбинации с олиго (dT)), мы рекомендуем инкубировать реакцию обратной транскрипции при комнатной температуре (~ 25 ° C) в течение 10 минут после добавления фермента для удлинения праймеров.
Среди обратных транскриптаз есть различия в термостабильности, которая, в свою очередь, определяет наивысшую оптимальную температуру полимеризации для каждого. Использование термостабильной обратной транскриптазы позволяет повысить температуру реакции (например,g., 50 ° C), чтобы помочь денатурировать РНК с высоким содержанием GC или вторичных структур, не влияя на активность фермента ( Рисунок 7 ). С такими ферментами высокотемпературная инкубация может привести к увеличению выхода, длины и представительства кДНК. ( Белая книга: Исследования производительности обратной транскриптазы SuperScript IV)
Время полимеризации зависит от процессивности обратной транскриптазы, которая относится к числу нуклеотидов, включенных в одно событие связывания.Например, обратной транскриптазе MMLV дикого типа с низкой процессивностью часто требуется> 60 мин для синтеза кДНК. Напротив, сконструированная обратная транскриптаза с высокой процессивностью может занять всего 10 минут для синтеза кДНК размером 9 т.п.н. ( Узнайте больше о процессивности обратной транскриптазы ).
Рисунок 7. Влияние термостабильности и ее влияние на активность обратной транскриптазы. Образцы, содержащие РНК различной длины, подвергали обратной транскрипции с использованием олиго (dT) праймеров и радиоактивно меченных dNTP.Продукты реакции разделяли гель-электрофорезом и визуализировали с помощью авторадиографии. Термостабильная обратная транскриптаза дает высокие выходы кДНК даже при температуре выше 50 ° C.
Обратная транскрипция за 10 минут
Узнайте, как можно достичь обратной транскрипции за 10 минут с помощью высокопроизводительной обратной транскриптазы.
- Деактивация фермента: Последний шаг в реакциях обратной транскрипции — это деактивировать обратную транскриптазу.Температура дезактивации может находиться в диапазоне 70–85 ° C, в зависимости от термостабильности фермента. Деактивация обычно проводится на 5–15 минут, при более высокой температуре требуется более короткое время.
Синтез первой и второй цепей кДНК
Синтез кДНК из матрицы РНК, как описано в предыдущем разделе, дает гибрид кДНК: РНК. Этот процесс упоминается как синтез первой цепи кДНК . Если присутствует активность РНКазы H (как в обратных транскриптазах AMV и MMLV дикого типа), РНК гибрида кДНК: РНК расщепляется во время синтеза первой цепи.КДНК первой цепи (с отожженной к ней РНК или без нее) может использоваться непосредственно в некоторых приложениях, таких как ОТ-ПЦР, где термостабильная ДНК-полимераза (например, ДНК-полимераза Taq ) реплицирует комплементарную цепь кДНК.
При конструировании библиотеки кДНК и секвенировании кДНК первой цепи используется в качестве матрицы для создания двухцепочечной кДНК, представляющей мишени РНК. Этот процесс известен как синтез второй цепи кДНК . При синтезе второй цепи кДНК рекомендуются обратные транскриптазы с минимальной активностью РНКазы Н, чтобы максимизировать длину и выход кДНК.
Синтез двухцепочечной кДНК часто использует другую ДНК-полимеразу (например, ДНК-полимеразу Т7, ДНК-полимеразу I, ДНК-полимеразу Taq ) для получения комплементарной цепи первой цепи кДНК. Дополнительные ферменты могут быть включены в синтез двухцепочечной кДНК, например, перечисленные ниже для модифицированного метода Габлера и Хоффмана [8] (, фиг. 8, ).
Рис. 8. Синтез двухцепочечной кДНК с помощью процедуры Габлера-Хоффмана.
- E. coli РНКаза H разрывает цепи РНК комплексов кДНК: РНК, обеспечивая 3′-ОН прайминговые сайты для синтеза ДНК.
- E. coli ДНК-полимераза I удлиняет разорванные цепи РНК за счет 5’3′-полимеразной активности и заменяет цепь РНК в направлении синтеза на 5’3′-экзонуклеазную активность в процессе, известном как перевод ник .
- E. coli ДНК-лигаза закрывает зазоры между вновь синтезированными сегментами кДНК.(ДНК-лигаза Т4 не должна использоваться в качестве замены, поскольку она может лигировать фрагменты двухцепочечной кДНК с тупым концом и образовывать химерные структуры.)
- ДНК-полимераза Т4 притупляет концы двухцепочечной кДНК (необязательно на последнем этапе).
В заключение, успех обратной транскрипции сильно зависит от компонентов реакции и условий.