
Клеточное строение листа — урок. Биология, Бактерии. Грибы. Растения (5–6 класс).
Главной функцией листа является осуществление фотосинтеза.
Фотосинтез — процесс, при котором зелёные растения, используя солнечный свет, из неорганических веществ производят органические вещества.
Фотосинтез происходит в хлоропластах.
Внутреннее строение листа
Изучение микроскопического строения листовой пластинки позволит понять механизм фотосинтеза и функции зелёных листьев.
Рис. \(1\). Лист под микроскопом
Если рассмотреть поперечный срез листовой пластинки под микроскопом, то можно заметить, что сверху и снизу он покрыт тонкой бесцветной кожицей, а внутри находятся зелёные клетки. Хорошо видна жилка, или проводящий пучок.
Рис. \(2\). Строение листовой пластинки
Строение кожицы
На верхней и нижней поверхностях листа находится кожица (эпидермис, эпидерма). Это разновидность покровной ткани, которая защищает клетки от механических повреждений и от высыхания, а также обеспечивает газообмен и испарение воды.
Это разновидность покровной ткани, которая защищает клетки от механических повреждений и от высыхания, а также обеспечивает газообмен и испарение воды.
Клетки эпидермиса бесцветные, чтобы солнечный свет свободно проникал внутрь листа.
На поверхности кожицы находится кутикула — восковой слой (восковой налёт), который предотвращает потерю воды. Растения с толстой кутикулой испаряют меньше воды, чем растения с тонкой кутикулой.
Связь тканей листа с внешней средой происходит через расположенные в кожице устьица.
Строение мякоти листа
Внутренняя часть листа (мякоть) образована фотосинтезирующей основной тканью.
В клетках мякоти имеются хлоропласты, в которых происходит фотосинтез. В мякоти присутствуют две разновидности основной ткани: столбчатая и губчатая. Столбчатая ткань находится под верхним эпидермисом.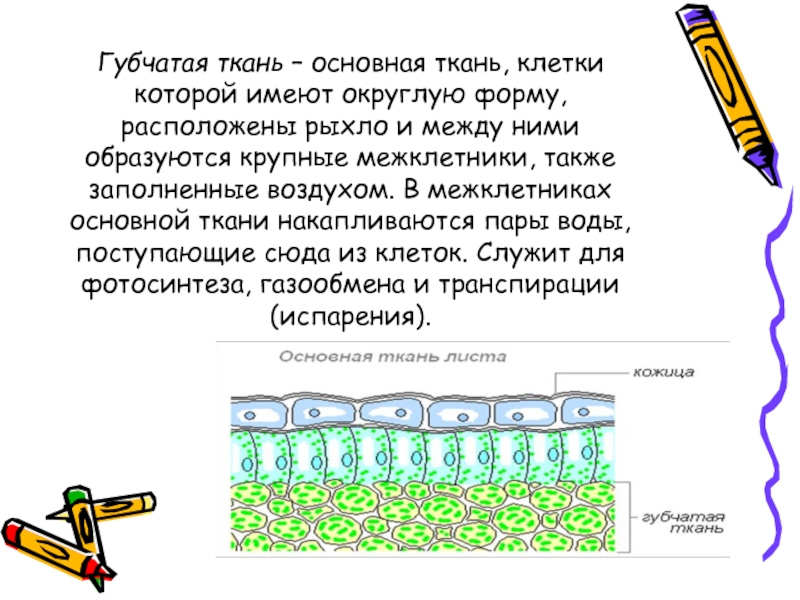 Она состоит из нескольких слоёв продолговатых клеток, содержащих большое количество хлоропластов.
Она состоит из нескольких слоёв продолговатых клеток, содержащих большое количество хлоропластов.
Под столбчатой тканью расположены клетки губчатой ткани. Эти клетки округлые и расположены рыхло. Между ними много межклетников, заполненных воздухом. В клетках губчатой ткани меньше хлоропластов по сравнению со столбчатой тканью.
Строение жилок листа
На поперечном срезе листовой пластинки под микроскопом хорошо видны жилки (проводящие пучки). Они образованы сосудами, ситовидными трубками и волокнами.
Рис. \(3\). Строение проводящего пучка
Волокна имеют толстые стенки. Они выполняют опорную функцию. Сосуды обеспечивают поступление к клеткам листа воды и минеральных солей, а ситовидные трубки — отток образовавшихся в ходе фотосинтеза органических веществ к другим органам растения.
Источники:
Рис. 1. Лист под микроскопом. https://image.shutterstock. com/image-photo/coffee-leaf-crosssection-under-microscope-600w-1672652050
com/image-photo/coffee-leaf-crosssection-under-microscope-600w-1672652050
Рис. 2. Строение листовой пластинки. https://img.freepik.com/free-vector/diagram-showing-anatomy-of-a-leaf_1308-38515.jpg?size=626&ext=jpg
Рис. 3. Строение проводящего пучка. © ЯКласс
ГДЗ биология 6 класс Лабораторные работы Задание: § Клеточное строение листа
Изображения обложек учебников приведены на страницах данного сайта исключительно в качестве иллюстративного материала (ст. 1274 п. 1 части четвертой Гражданского кодекса Российской Федерации)
Авторы: Лабораторные работы
Вид УМК: лабораторные работы
На данной странице представлено детальное решение § Клеточное строение листа по биологии для учеников 6 классa автор(ы) Лабораторные работы
§ Клеточное строение листа
Ход работы:
Изучаем готовые микропрепараты среза листа. Находим клетки верхней и нижней кожицы, устьица.
Рассматриваем клетки мякоти листа.
 Мякоть листа состоит из клеток основной ткани, которая лежит в 2-3 слоя. Слои плотно прилегают к верхней кожице и состоят из продолговатых клеток, которые похожи на столбики. Эта часть называется столбчатой. В цитоплазме клеток мы видим много хлоропластов. Чуть ниже столбчатой ткани можно увидеть уже округлые или неправильной формы клетки, прилегающие плотно друг у другу. В них меньше хлоропластов, но они образуют губчатую ткань.
Мякоть листа состоит из клеток основной ткани, которая лежит в 2-3 слоя. Слои плотно прилегают к верхней кожице и состоят из продолговатых клеток, которые похожи на столбики. Эта часть называется столбчатой. В цитоплазме клеток мы видим много хлоропластов. Чуть ниже столбчатой ткани можно увидеть уже округлые или неправильной формы клетки, прилегающие плотно друг у другу. В них меньше хлоропластов, но они образуют губчатую ткань.Находим на микропрепарате межклетники. Они расположены в губчатой ткани и заполнены воздухом. Их функция заключается в улучшении газообмена между окружающей средой и клетками. Также они могут накапливать продукты выделительных тканей, например, слизь или эфирные масла.
Находим проводящие пучки листа. Они образованы из сосудов, волокон и ситовидных трубок. Волокна – вытянутые клетки с толстыми клетками придают листу прочность, а сосуды выполняют транспортирующую функцию – переносят воду и растворенные в ней минеральные вещества. Ситовидные трубки состоят из живых длинных клеток с поперечными перегородками между ними, которые пронизаны узкими канальцами.
 По ним движутся растворы органических веществ.
По ним движутся растворы органических веществ.Зарисовываем поперечный срез листа и делаем все необходимые подписи.
Add
Новыe решебники
© 2021Copyright. Все права защищены. Правообладатель SIA Ksenokss.Адрес: 1069, Курземес проспект 106/45, Рига, Латвия.
Тел.: +371 29-851-888 E-mail: [email protected]
Губчатая паренхима листа — Справочник химика 21
Клетки губчатой паренхимы мезофилла листа (клетки имеют неправильную форму) [c.257] Мезофилл состоит обычно из двух главных типов паренхимы из палисадной паренхимы, клетки которой ориентированы перпендикулярно поверхности листовой пластинки, и губчатой паренхимы—клеток различного размера, образующих рыхлую трехмерную сетку под палисадной тканью или между ее слоями (фиг. 59). Палисадная ткань на поперечном срезе листа представляется значительно более плотной, чем губчатая однако на срезах, параллельных листовой пластинке, видно, что значительная часть поверхности каждой палисадной клетки также соприкасается с воздухом, наполняющим межклетники.
У яблони, пораженной ржавчиной, снижение транспирации связано с нарушениями в анатомическом строении листа. Рыхлая ткань губчатой паренхимы заменяется плотной, лишенной межклетников тканью, составленной из столбчатых клеток у больных листьев резко сокращается также число устьиц. [c.645]
Важнейшая ткань листа — мезофилл, где осуществляется фотосинтез (рис. 3,17). Покрывающий лист эпидермис, клетки которого, за исключением замыкающих клеток устьиц, не содержат хлоропластов, защищает ткани листа, регулирует газообмен и транспирацию. Система разветвленных проводящих пучков необходима для снабжения тканей листа водой, минеральными и некоторыми органическими веществами и для оттока ассимилятов в другие части растения. Мезофилл обычно дифференцирован на две ткани — палисадную (столбчатую), расположенную под верхним эпидермисом, и губчатую, находящуюся в нижней стороне листа.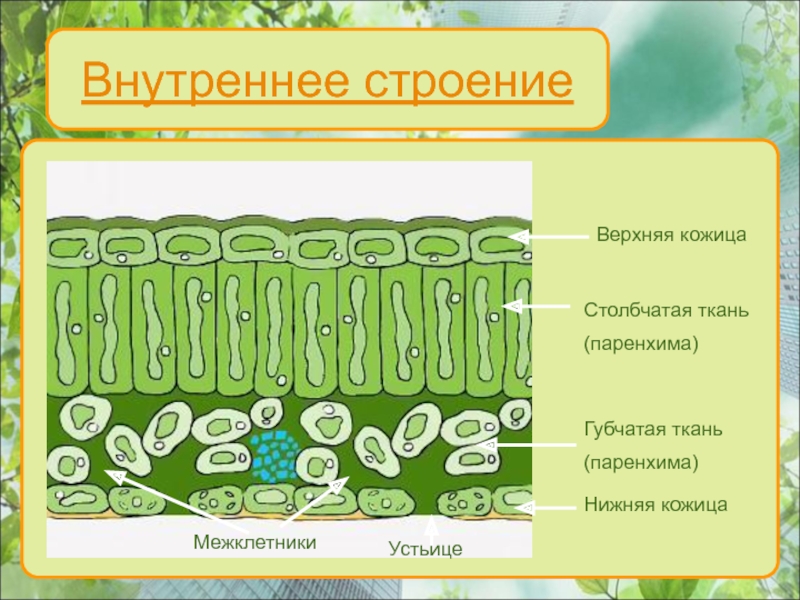
Большие различия в величине осмотического давления наблюдаются даже у клеток одной и той же ткани. Например, у палисадных клеток листа осмотическое давление в полтора-два раза выше, чем у губчатых. И в данном случае различия не случайны, так как установлено, что сильное развитие палисадной паренхимы наблюдается и в пределах одной и той же ткани (табл. 4).
Листья, выросшие на ярком свету, имеют, как правило, более развитый столбчатый мезофилл. У растений засушливых ареалов — ксерофитов — палисадная паренхима часто расположена по обеим сторонам листа, а губчатая сильно редуцирована или совсем отсутствует. [c.100]
[c.100]
Микроскопия. При рассмотрении листа с поверхности видны клетки эпидермиса с извилистыми стенками. Устьица только на нижней стороне листа с 4—7 околоустьичными клетками (аномоцитный тип). На нижней стороне листа по жилкам встречаются редкие, длинные простые волоски с тонкими стенками, часто оборванные, состоящие из 7—20 клеток, иногда перекрученные или с отдельными спавшимися члениками. На верхушках город-чатых зубцов при схождении жилок расположена гидатода с сосочковидным эпидермисом и 2—5 крупными водяными устьицами. Клетки губчатой паренхимы с крупными водяными устьицами. Клетки губчатой паренхимы с крупными межклетниками (аэренхима). Жилки сопровождаются млечными трубками с темно-бурым зернистым содержимым (после кипячения в щелочи). [c.310]
Таким образом, негомогенности нельзя избежать, даже применяя разбавленные клеточные суспензии, в которых освещенность одинакова для всех клеток, но не для всех молекул хлорофилла.
 В более плотных суспензиях можно достичь лишь средней по времени постоянной освещенности всех клеток, и то лишь при очень энергичном размешивании. В слоевищах многоклеточных водорослей или в листьях высших растений несоразмерность в скоростях поглощения света у различных клеток не может быть учтена совсем. Например, поглощение в клетках губчатой паренхимы при всех обстоятельствах бывает значительно слабее, чем в палисадных клетках (фиг. 136). Таким образом, на кривых, представляюнщх скорость фотосинтеза (Р) как функцию концентрации двуокиси углерода или интенсивности света, абсциссы являются средними значениями, усредненными для одной или
В более плотных суспензиях можно достичь лишь средней по времени постоянной освещенности всех клеток, и то лишь при очень энергичном размешивании. В слоевищах многоклеточных водорослей или в листьях высших растений несоразмерность в скоростях поглощения света у различных клеток не может быть учтена совсем. Например, поглощение в клетках губчатой паренхимы при всех обстоятельствах бывает значительно слабее, чем в палисадных клетках (фиг. 136). Таким образом, на кривых, представляюнщх скорость фотосинтеза (Р) как функцию концентрации двуокиси углерода или интенсивности света, абсциссы являются средними значениями, усредненными для одной или Выше было указано, что при опытах с листьями высших наземных растений возникает специальный вопрос о прохождении двуокиси углерода через устьица и воздушные ходы, по которым она должна течь, чтобы достигнуть фотосинтезирующих клеток полисадной ткани и губчатой паренхимы. [c.327]
Мозаика. При этом заболевании листья приобретают пеструю (мозаичную) расцветку, в них происходит уменьшение хлорофилла, уплотнение клеток губчатой паренхимы, исчезно вение межклеточных пространств, уменьшение клеток палисадной паренхимы. Все эти нарушения вызывают ослабление роста и развития растения. [c.39]
Все эти нарушения вызывают ослабление роста и развития растения. [c.39]
Последнее, очевидно, связано с нзлшнением анатомического строения лпстьев растении увеличивается губчатая паренхима межклеточного пространства н тем самым улучшается газообмен листьев. Кроме того, метка радпо-активным изотопом фосфора (Р ) суперфосфата, внесенного в почву, позволила установить более быстрое погло-щ,епие фосфора корнями растений, выросших из облученных семян. [c.289]
А И Ах—оболочка кожицы листа 77—палисадная паренхима Г — губчатая паренхима в клетках обоих видов паренхимы видны зернышки хлорофилла X—меж клетные ходы К — отложёну я кальциевой соли щавелевой кислоты (друзы) У—устьица В — дыхательная полость. [c.53]
Через систему заполненных воздухом межклетников идет газообмен между живыми клетками и внещней средой, с которой связывают эту систему устьица (особые поры листа) или чечевички (специализированные щели в стеблях древесных пород).
МЕЗОФИЛЛ (СМ. ТАКЖЕ РИС. 7.3. И 7.4). Эта выполняющая ткань располагается между двумя слоями эпидермиса листа (рис. 6.1) и состоит из модифицированных паренхимных клеток, осуществляющих фотосинтез. Фотосинтетическую паренхиму иногда называют хлоренхимой. Цитоплазма клеток хлоренхимы содержит большое число хлоропластов, в которых и протекают реакции фотосинтеза. У двудольных растений мезофилл состоит из двух четко различающихся слоев верхний слой составляет палисадная паренхима, клетки которой имеют столбчатую форму, а нижний — губчатая паренхима с клетками неправильной формы, содержащими меньше хлоропластов. Фотосинтез вдет главным образом в палисадной паренхиме, а воздухоносные межклетники губчатой паренхимы обеспечивают интенсивный газообмен. [c.224]
Паренхимные клетки мезофилла содержат специализированные органеллы — хлоропласты, — осуществляющие фотосинтез.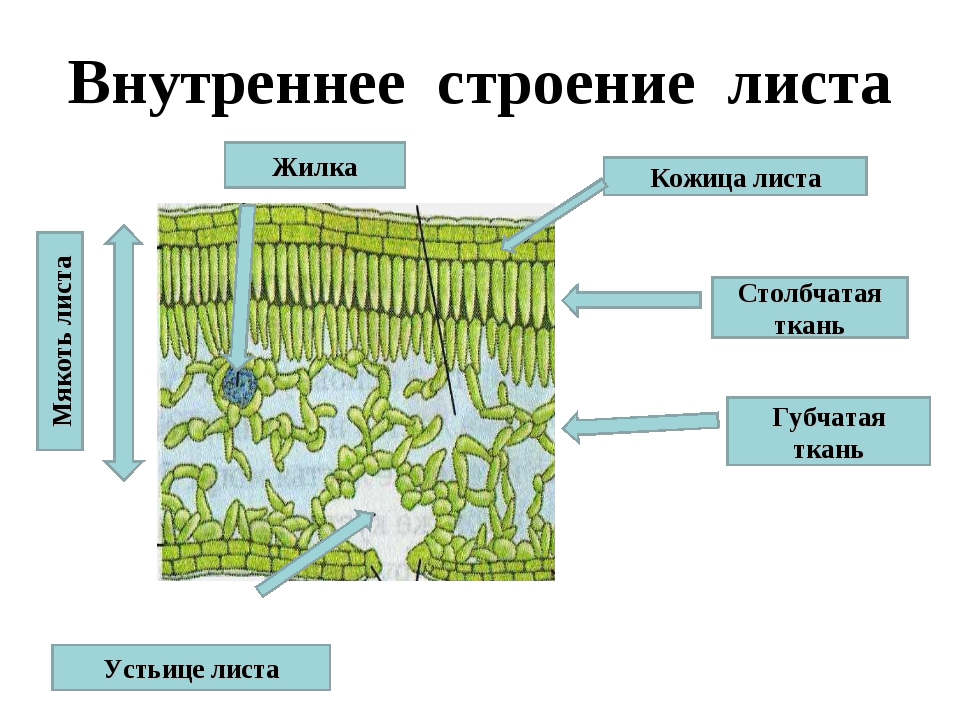 В хлоропластах находится хлорофилл. У двудольных клетки палисадной паренхимы мезофшша, содержащие больше хлоропластов, располагаются вблизи верхней поверхности листа, что обеспечивает максимальное улавливание света. Сравнительно большая длина этих клеток увеличивает возможности поглощения света. Хлоропласты находятся по периферии клеток палисадной паренхимы. Это позволяет им поглощать максимально возможное количество света и облегчает газообмен. Хлоропласты обладают фототаксисом, т. е. они перемещаются в клетке по направлению к свету. У двудольных губчатая паренхима мезофилла имеет обширные межклетники для эффективного газообмена. [c.341]
В хлоропластах находится хлорофилл. У двудольных клетки палисадной паренхимы мезофшша, содержащие больше хлоропластов, располагаются вблизи верхней поверхности листа, что обеспечивает максимальное улавливание света. Сравнительно большая длина этих клеток увеличивает возможности поглощения света. Хлоропласты находятся по периферии клеток палисадной паренхимы. Это позволяет им поглощать максимально возможное количество света и облегчает газообмен. Хлоропласты обладают фототаксисом, т. е. они перемещаются в клетке по направлению к свету. У двудольных губчатая паренхима мезофилла имеет обширные межклетники для эффективного газообмена. [c.341]
Микроскопия. При рассмотрении листа с поверхности видны клетки эпидермиса с извилистыми боковыми стенками, со складчатой кутикулой многочисленные устьица с 3—4 околоустьичными клетками различной величины преобладают на нижней стороне листа. Волоски редкие, головчатые и простые. Головчатые волоски двух типов одни с длинной многоклеточной ножкой и одноклеточной головкой, другие — с одноклеточной ножкой и головкой из 4—6 клеток.
Микроскопия. При рассмотрении листа с поверхности видны клетки эпидермиса с верхней стороны — с более прямыми стенками, с нижней — с более извилистыми устьица с обеих сторон с 3—4 околоустьичными клетками различной величины. Волоски двух типов простые — крупные из 2 (реже 5) клеток с тонкими стенками, с грубобородавчатой кутикулой, расположенные главным образом по жилкам и по краю листа и головчатые — более мелкие, с многоклеточной (реже одноклеточной) округлой притупленной головкой на короткой одноклеточной ножке. У молодых листьев головчатых волосков значительно больше, чем у старых. В клетках губчатой паренхимы имеются в большом количестве друзы оксалата кальция.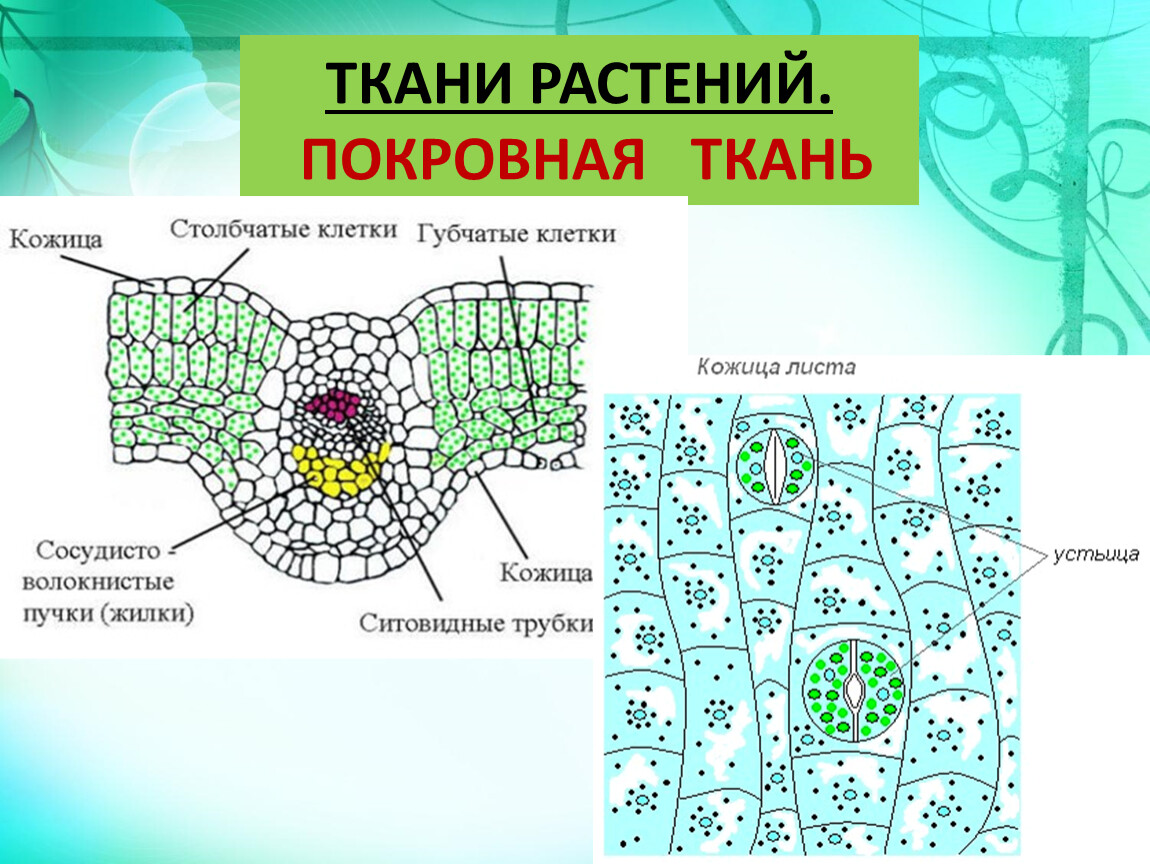 [c.307]
[c.307]
Необходимо иметь в виду, что этот артефакт (кажущаяся компактность расположения клеток мезофилла, наблюдаемая на типичных поперечных срезах листьев) привел к двум ошибочным представлениям относительно внутренней геометрии листа одно из них касается аэрации палисадной ткани, а другое — строения подустьичных полостей. Поэтому важно указать следующее. Во-первых, площадь внутренней поверхности листа обычно на порядок больше, чем площадь наружной его поверхности [774, 775], а палисадная ткань имеет обычно большую, а не меньшую открытую внутреннюю поверхность, чем губчатая паренхима [217]. Во-вторых, хотя непосредственно под каждым устьицем обычно находится более широкое свободное пространство, чем между большинством клеток мезофилла, представление о том, что подустьичные полости как бы выстланы клетка.ми (возникающее при взгляде на поперечный срез листа), совершенно неверно. Устьица — это только ворота, через которые осуществляется вентиляция всей внутренней поверхности мезофилла.
 Следовательно, хотя при диффузии газов от поверхности клетки до устьичного отверстия длина и извилистость пути в одной части листа могут быть значительно большими, чем в другой, нужно всегда помнить, что обмен как СО.,, так и водяного пара происходит через все открытые участки поверхности клеток, находящихся внутри листа. [c.233]
Следовательно, хотя при диффузии газов от поверхности клетки до устьичного отверстия длина и извилистость пути в одной части листа могут быть значительно большими, чем в другой, нужно всегда помнить, что обмен как СО.,, так и водяного пара происходит через все открытые участки поверхности клеток, находящихся внутри листа. [c.233] Рис, 3.9. Схематическое изображение листа в поперечном разрезе. Лист состоит нз двух защитных слоев — верхнего и нижнего эпидермиса, между которыми располагаются активно фотосинтезирующие клеткн. К верхнему эпидермису примыкает слой палисадной паренхимы, а к иижнему — губчатая паренхима. Эпидермис пронизан устьичными щелями, отверстость которых регулируется тургором замыкающих клеток. Межклеточные оздухоносиые пространства создают тот путь, по которому происходит газообмен, т. е. обмен СО, кисло- родом и водяными парами. По мелким сосудистым пучкам в лист поступают вода и минеральные вещества, а из листа оттекают синтезированные сахара. Как верхний, так и нижний эпидермис бывает обычно покрыт воскообразной, практически водонепроницаемой кутикулой. [c.96]
[c.96]
Мезофиты очень разнообразны по своим физиологическим и анатомо-морфологическим признакам. Такое разнообразие характерно не только для различных видов, но и для растений одного и того же вида, находящихся в неодинаковых экологических условиях. Так, известно явление усиления ксероформио-сти строения по мере перехода от нижних листьев к листьям расположенных выше ярусов (закон В. Р. Заленского). [234]. Объясняется это усложнением условий водоснабжения листьев верхних ярусов по сравнению с листьями нижних ярусов листья верхних ярусов подвергаются более интенсивным воздействиям солнечного света, температурных градиентов, режима влажности атмосферы и т. д. У световых листьев дифференцируются и палисадная паренхима, состоящая из плотно прилегающих друг к другу клеток, и губчатая паренхима, в то время как у теневых листьев имеется только рыхлая губчатая ткань. [c.214]
Для осуществления процесса фотосинтеза имеют значение особенности строения листа.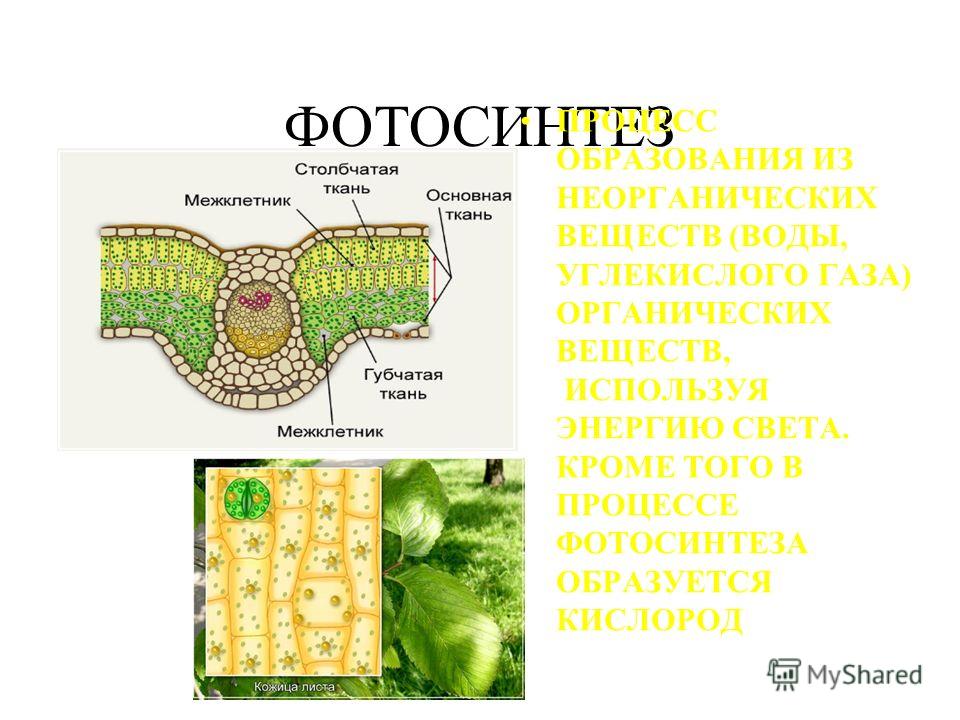 К верхней стороне листа прилегает палисадная ткань, клетки которой расположены перпендикулярно, плотно соприкасаются друг с другом и содержат много хлоропластов. Палисадная паренхима является преимущественно ассимиляционной тканью. К нижнему эпидермису прилегает губчатая паренхима с рыхлорасположенными клетками и межклетниками. Это приспособление у растений служит для лучшего проникновения газов в клетки (рис. 21). [c.151]
К верхней стороне листа прилегает палисадная ткань, клетки которой расположены перпендикулярно, плотно соприкасаются друг с другом и содержат много хлоропластов. Палисадная паренхима является преимущественно ассимиляционной тканью. К нижнему эпидермису прилегает губчатая паренхима с рыхлорасположенными клетками и межклетниками. Это приспособление у растений служит для лучшего проникновения газов в клетки (рис. 21). [c.151]
Каждый из перечисленных органов растительного организма построен из нескольких типов тканей, т. е. групп клеток, которые выполняют определенную физиологическую функцию и имеют сходное морфологическое строение, обеспечивающее реализацию этой функции. По функциональному значению в растениях различают следующие типы тканей образовательные (меристемы), ассимиляционные (хло-ренхима), запасающие, покровные, выделительные, механические (скелетные), проводящие и аэренхиму. Причем в каждом таком типе представлены ткани с более узкой специализацией. Например, к ассимиляционным тканям листа относятся столбчатая и губчатая паренхима, обкладка пучка. К покровным тканям — эпидермис, ризодерма, перидерма, эндодерма и др. [c.28]
К покровным тканям — эпидермис, ризодерма, перидерма, эндодерма и др. [c.28]
Микроскопия. Цельное и резаное сырье. Кусочки листоппй пластинки просветляют (метод приготовления препаратов, см. стр. 859) и рассматривают с поверхности. В зависимости от установки микрометрического винта виден или эпидермис (кожица) с выростами, или мезофилл (мякоть) с жилками, кристаллами и вместилишами. Палисадная ткань имеет вид равномерных, большей частью округлых клеток. Губчатая ткань состоит из разнообразных по форме клеток, часто с крупными межклетниками. В жилках видны тонкие спиральные (иногда пористые) сосуды в один или несколько рядов. Жилки и паренхима в таких препаратах обычно однообразны для различных листьев и поэтому диагностического значения почти не имеют. [c.296]
Порошок. При измельчении мякоть листа в значительной степени разрушается. Жилки обычно разрываются продольно и сопровождаются сохранившимися клетками паренхимы. Нередко можно встретить отдельные фрагменты листа в поперечном сечении с хорошо с0хранивн1имися тканями на таком фрагменте хорошо видны верхний эпидермис, палисадная ткань, губчатая ткань, нижний эпидермис и часто мелкие проводящие пучки. Основное диагностическое значение в порошках листьев обычно имеют те же элементы, что и в препарате листа с поверхности эпидермис, волоски, кристаллы, железки и т. д. Эпидермис в порошке лежит небольшими обрывками (по нескольку клеток), волоски и их обрывки чаще лежат отдельно и лишь изредка прикреплены к эпидермису кристаллы — отдельно и в клетках паренхимы, железки и вместилища — чаще вместе с обрывками окружающей их ткани листа. [c.296]
Основное диагностическое значение в порошках листьев обычно имеют те же элементы, что и в препарате листа с поверхности эпидермис, волоски, кристаллы, железки и т. д. Эпидермис в порошке лежит небольшими обрывками (по нескольку клеток), волоски и их обрывки чаще лежат отдельно и лишь изредка прикреплены к эпидермису кристаллы — отдельно и в клетках паренхимы, железки и вместилища — чаще вместе с обрывками окружающей их ткани листа. [c.296]
Согласно современным представлениям, токсический газ, попадая через устьица или эпидермис в лист, растворяется в воде клеточных оболочек и взаимодействует с щгтоплазмой. Первыми повреждаются клетки устьичных полостей, затем клетки губчатой палисадной паренхимы. Газ, растворяющийся в воде, образует кислоту илн щелочь, которые взаимодействуют с протопластом. Часть их нейтрализуется, а часть остается в свободном состоянии. Кислоты разрушают хлорофилл, изменяют pH тканей листа и устойчивость биоколлопдов цитоплазмы, повышают общую окисляемость, увеличивают дисперсность коллоидов и гигроскопичность ткаией, отрицательно влияют на энзиматический аппарат, нарушают обмен веществ в клетках листа [c. 523]
523]
Клеточное строение листа — презентация онлайн
2. Кожица листа
Кожица — один извидов
покровной
ткани растения.
Клетки
кожицы
бесцветны
и
прозрачны, но среди
бесцветных
клеток
встречаются
расположенные
парами
зеленые
замыкающие клетки.
Между
ними
находится щель.
Эти клетки и щель
между
ними
называют устьицем.
3. Устьице
Устьицемназываются
замыкающие клетки и
щель между ними.
Через устьичную щель в
лист проникает воздух и
происходит
испарение
воды.
У большинства растений
устьица находятся на
нижней стороне листа, на
листьях водных растений
на верхней стороне листа,
а у подводных их вообще
нет.
4. Строение мякоти листа
Состоит из клеток основнойткани.
Слои прилегающие к верхней
кожице
образованны
клетками удлиненной формы.

Это столбчатая основная
ткань.
В цитоплазме этих клеток
много хлоропластов.
Губчатая ткань состоит из
клеток
округлой
или
неправильной
формы
неплотно прилегающие друг
к другу.
Хлоропластов в этих клетках
меньше, чем в столбчатых.
5. Межклетники
Межклетники – это полости образованныеклетками губчатой ткани.
Межклетники заполнены воздухом.
Через межклетники осуществляется газообмен.
между
6. Строение жилок листа
7. Строение жилок листа
Волокна–
сильно
вытянутые
клетки
с
толстыми клетками. Они
придают
прочность
листу.
Сосуды – образованны
мертвыми вытянутыми
клетками.
По
ним
передвигаются вода и
растворенные
в
ней
минеральные вещества.
Ситовидные трубки — образованными живыми длинными
клетками, поперечные перегородки которых пронизаны узкими
каналами.
По С.
 Т. Из листьев передвигаются органические вещества.
Т. Из листьев передвигаются органические вещества.8. Домашнее задание
§ 24, 25. упр. 80, 82, 83Урок-исследование по теме «Клеточное строение листа»
Тип урока: урок изучения нового материала.
Форма проведения: урок-исследование.
Триединая цель урока:
1. Образовательная: познакомить учащихся с клеточным строением листа; установить связь особенностей строения клеток с выполняемыми им функциями; продолжить формирование умения приготовления микропрепарата и работа с ним.
2. Развивающая: развивать логическое мышление, память, внимание, познавательный интерес, исследовательские умения.
3. Воспитательная: воспитывать чувство сопереживания, самоконтроля, культуру труда.
Оборудование: презентация, готовые микропрепараты, микроскопы.
Планируемые результаты обучения:
Учащиеся должны знать:
— строение и функции кожицы листа;
— роль устьиц в жизни растений;
— строение устьиц;
— особенность строения столбчатой ткани;
— характерные признаки губчатой ткани;
— строение проводящих пучков жилок листа;
— значение слов кожица листа, устьица,
столбчатая и губчатая ткани, межклетники,
проводящие пучки, мякоть листа, осмос,
транспирация.
Учащиеся должны понимать:
— зависимость интенсивности транспирации от состояния замыкающих клеток устьиц;
— что осмотическое давление неодинаково у различных растений и в клетках разных органов одного и того же растения.
Структура урока
1. Оргмомент.
2. Введение в урок. Создание проблемной ситуации.
3. Изучение нового материала.
4. Закрепление.
5. Тестовый контроль.
6. Рефлексия.
7. Подведение итогов урока.
8. Домашнее задание.
Ход урока
1. Оргмомент (отметить отсутствующих учеников в журнале, визуально проверить готовность к уроку, словами “В добрый путь” психологически настроить на урок).
2. Введение в урок.
Вопросы учителя:
1. Какой орган растения Вы изучаете? (Лист)
2. Какую важную работу (функцию) выполняет лист? (Участвует в фотосинтезе, газообмене, испарении воды)
Создаю проблемную ситуацию. Вы хотите узнать,
какие особенности внутреннего строения листа
помогают выполнять ему эти функции? (“Да”)
Вы хотите узнать,
какие особенности внутреннего строения листа
помогают выполнять ему эти функции? (“Да”)
Для ответа на этот вопрос мы с Вами должны изучить клеточное строение листа, поэтому тема нашего урока “Клеточное строение листа”. (Слайд 2).
Презентация.
Прошу учащихся помочь мне сформировать цель урока.
Цель урока: выяснить, как клеточное строение листа соответствует его функции. (Слайд 2).
Для того, что бы изучить клеточное строение
листа, нам необходимо его исследовать. (Слайд 3).
Поэтому у нас не просто урок, а урок-исследование.
Представьте себе, что мы с Вами находимся в
научно-исследовательской лаборатории. Я главный
сотрудник лаборатории, а Вы мои помощники.
Действительно помощники, так как вместе с Вами в
нашей лаборатории мы заранее приготовили
микропрепарат кожицы листа для исследования на
уроке.
Вспомните, как Вы готовили микропрепарат (ответы учеников).
Напомните правила работы с микроскопом (ответы учеников).
Приступаем к исследованию.
Для того, что бы у Вас было представление, какие клетки входят в состав листа, откройте учебник страница 118 рис. 84. Ответьте на вопрос, какие клетки образуют внутреннее строение листа?
Исследование начнем с клеток кожицы листа.
1. Найдите бесцветные клетки кожицы. Рассмотрите их форму и строение.
- На какие уже известные вам клетки они похожи?
- Почему они бесцветные и прозрачные?
- Какую функцию они выполняют?
- Посмотрите, оболочки клетки плотно прилегают друг к другу?
- Какое это имеет значение для листка?
- Группы клеток имеют одинаковое строение и
выполняют одну и туже функцию – называют тканью.
Какой тканью образованы клетки кожицы?
(Покровная).

- Среди бесцветных и прозрачных клеток кожицы увидели другие клетки? (Да). Они получили название устьичные клетки. (Слайд 4).
2. Найдите устьичные клетки? Чем устьичные клетки отличаются от других клеток кожицы листа? (Слайд 5).
Я показываю слайд и рассказываю о строении устьиц, их количестве на разных листовых пластинках, тем самым подвожу к вопросу о значении устьиц для листка. (Слайд 6).
Я рассказываю о нахождении устьиц у сухопутных растений, водных растений, плавающих на поверхности воды, также на подводных листьях. (Слайд 7).
3. Физкультминутка для глаз.
4. Зарисуйте кожицу листа под микроскопом. Отдельно зарисуйте устьице. Сделайте надписи к рисункам. (Рабочая тетрадь 42 с. Задание 81).
5. Сделайте вывод о значении кожицы листа.
Продолжаем наше исследование.
- Какие клетки находятся под верхним слоем
кожицы? (Клетки мякоти).

- Верхние два слоя клеток получили название “столбчатая ткань”.
- Почему они получили такое название? (Клетки расположены вертикально в виде столбиков).
- Клетки, которые прилегают к нижней кожицы, образуют губчатую ткань. Как Вы думаете, почему ее называют губчатой тканью? (Клетки более круглые, неправильной формы, неплотно прилегают друг к другу).
- Как называется пространство между клетками? (Межклетники).
- Как Вы думаете, чем они заполнены? (Воздухом, парами воды).
- Какие клетки столбчатой или губчатой ткани имеют более зеленую окраску? (Клетки столбчатой ткани).
- Какое вещество придает им зеленую окраску? (Хлорофилл, который находится в хлоропластах).
Основная функция этих клеток – фотосинтез.
Столбчатая и губчатая ткани являются
разновидностью основной ткани. (Фотосинтезирующей).
(Фотосинтезирующей).
Клетки мякоти листа – основная ткань.
Вывод: мы выяснили, как строение клеток соответствует выполнению функции основной ткани?
Исследуем жилки листовой пластинки. (Слайд 8).
Вспоминаем особенности строения сосудов, ситовидных трубок, какую выполняют функцию и какой образованы тканью.
Жилки листка прочные, удерживают листковую пластинку, придают листку прочность. (Слайд 9). Как Вы думаете, с какими особенностями строения клеток это связано? (В каждой жилке проходит пучек волокон, клетке с крепкими оболочками, и толстыми стенками).
Исследование завершено, цель урока мы достигли? (Да).
Что мы выяснили? (Ответы учащихся).
Учитель подводит итоги исследования.
Закрепление пройденного материала.
1. Какие клетки образуют листовую пластинку?
2. Какое значение имеет кожица листа? Клетками
какой ткани она образована?
Какое значение имеет кожица листа? Клетками
какой ткани она образована?
3. Что такое устьица и где они расположены?
4. Какое строение имеют клетки мякоти листа? К какому типу тканей они относятся?
5. В каких клетках листа особенно много хлоропластов?
6. Какую функцию выполняют проводящие пучки листа? Клетками каких тканей они образованы?
Найди ошибки (Игра “капитан Врунгель”)
(Клеточное строение листа)
1. Клетки кожицы образованы механической тканью, они пропускают свет и выполняют защитную функцию.
2. Устьичные клетки выполняют защитную функцию.
3. Клетки мякоти листа состоят из сосудов и ситовидных трубок.
4. Клетки мякоти листа имеют зеленый цвет, так как в них содержится хлорофилл.
5. Межклетники заполнены воздухом.
6. Волокна, которые входят в состав жилок листа, выполняют проводящую функцию.
Тестовый контроль с последующей
взаимопроверкой.
Клеточное строение листа
Вариант 1
1. К какому типу тканей относится кожица листа?
А) покровная ткань;
Б) проводящая ткань;
В) механическая ткань.
2. Какую функцию выполняют устьица листа?
А) проведение жидкостей;
Б) газообмен;
В) опора.
3. Чем отличается губчатая ткань от столбчатой?
А) расположением клеток;
Б) цветом клеток.
4. Какие клетки листа пропускают свет?
А) сосуды;
Б) ситовидные клетки;
В) клетки кожицы.
5. В каких клетках находятся хлоропласты?
А) в клетках кожицы;
Б) в межклетниках;
В) в клетках мякоти листа.
Вариант 2
1. Где расположены клетки устьиц?
А) в межклетниках;
Б) в клетках мякоти;
В) в клетках кожицы.

2. Чем отличается губчатая ткань от столбчатой?
А) расположением клеток;
Б) цветом клеток.
3. В каких клетках находятся хлоропласты?
А) в клетках кожицы;
Б) в межклетниках;
В) в клетках мякоти листа.
4. К какому типу тканей относится кожица листа?
А) покровная ткань;
Б) проводящая ткань;
В) механическая ткань.
5. Какую функцию выполняют устьица листа?
А) проведение жидкостей;
Б) газообмен;
В) опора.
Ключ ответа:
Вариант 1
1А, 2Б, 3А, 4В,5В
Вариант 2
1В, 2А, 3В, 4А, 5Б
Рефлексия (что нового узнали на уроке).
Подведение итогов: Дать характеристику работы учащихся на занятии.
Домашнее задание: поставить опыт с последующим наблюдением с. 119. Выполнить творческое задание: текст из 4-5 предложений и в 2-х из них допущены ошибки.
Клеточное строение листа. – Книга для чтения по ботанике – Kaz-Ekzams.ru
admin 24.08.2010
Книга для чтения по ботанике
Чтобы лучше понять значение зеленых листьев в жизни растений, познакомимся с внутренним строением листа.
Разрежем лист поперек. Срежем острой бритвой с поперечного разреза листа тончайшую пластиночку. Положим ее в каплю воды на предметном стекле, накроем покровным стеклом и рассмотрим под микроскопом.
Оказывается, лист — не сплошная зеленая пластинка. Он состоит из множества клеток различной величины и формы, то есть имеет клеточное строение.
Даже самая крупная клетка листа настолько мала, что простым глазом ее не различить. Если поставить точку на бумаге и разделить ее на 200— 300 частей, то каждая такая частичка по величине будет равна клетке. Приглядитесь: сверху и снизу листа вытянулись в ряд и плотно прилегают друг к другу клетки одинакового размера и формы. Это клетки кожицы. Она покрывает лист, предохраняет его от повреждений и высыхания. Клетки кожицы образуют покровную ткань.
Если поставить точку на бумаге и разделить ее на 200— 300 частей, то каждая такая частичка по величине будет равна клетке. Приглядитесь: сверху и снизу листа вытянулись в ряд и плотно прилегают друг к другу клетки одинакового размера и формы. Это клетки кожицы. Она покрывает лист, предохраняет его от повреждений и высыхания. Клетки кожицы образуют покровную ткань.
Клетки верхней кожицы прозрачны, как стекло, а в клетках нижней кое-где встречаются мелкие парные зеленые клетки. Эти парные клетки-называют устъичными. Устьичные клетки и щель между ними называют устьицем. Через устьица совершается проникновение углекислого газа воздуха внутрь листа и выход кислорода в атмосферу.
Через устьица из растения также испаряется избыток влаги.
У большинства растений устьица находятся только в нижней кожице листьев. Но у некоторых, например у капусты, они расположены и в верхней кожице. У растений, листья которых плавают на поверхности воды, например у белой лилии-кувшинки, устьица находятся только на верхней стороне листа.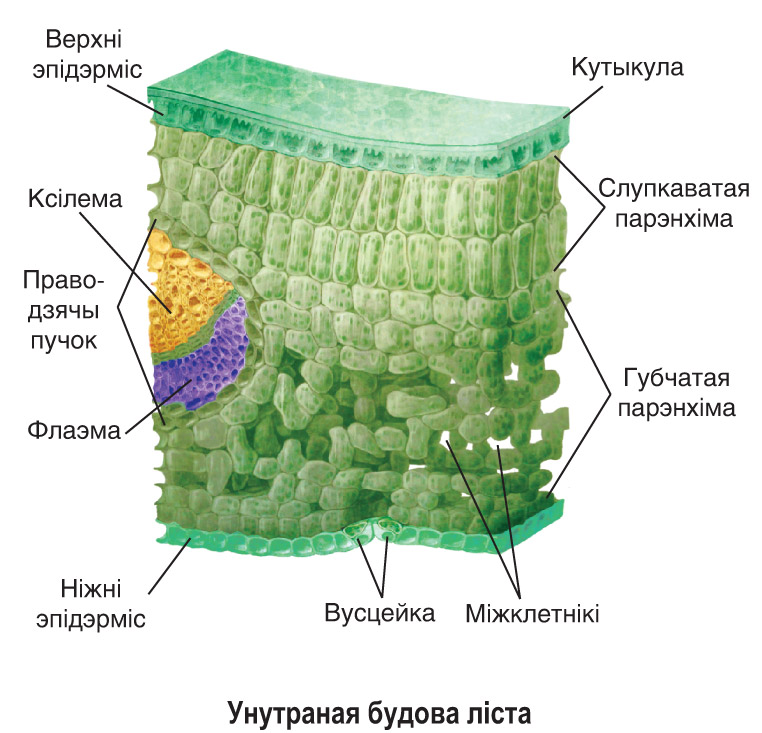 Число устьиц на листьях огромно. Ученые подсчитали, что на одном только листе подсолнечника на 1 мм2 220 устьиц, а на листе клена их на 1 мм2 — 550.
Число устьиц на листьях огромно. Ученые подсчитали, что на одном только листе подсолнечника на 1 мм2 220 устьиц, а на листе клена их на 1 мм2 — 550.
Под кожицей лежат клетки мякоти. Они зеленые, потому что в их цитоплазме содержатся очень мелкие зеленые пластиды — хлоропласты. Зеленый цвет хлоропластов объясняется присутствием в них зеленого красящего вещества — хлорофилла. Хлорофилл в хлоропластах образуется только на свету. Хлоропласты у цветковых растений по их форме часто называют хлорофилловыми зернами. О пластидах вы узнали еще при изучении строения клетки.
Мякоть листа состоит из нескольких рядов клеток. Клетки, примыкающие к верхней кожице, похожи на довольно ровные столбики. В них особенно много хлорофилловых зерен. Нижние клетки, более округлые или неправильной формы, неплотно прилегают друг к другу. Пространства между клетками называют межклетниками. Межклетники губчатой ткани заполнены воздухом.
Если рассматривать под микроскопом мякоть листа, в середине ее можно увидеть округлый разрез жилки. Присмотревшись, нетрудно заметить поперечные разрезы видоизмененных клеток — сосудов и волокон, они образуют сосудисто-волокнистые пучки жилки листа.
Присмотревшись, нетрудно заметить поперечные разрезы видоизмененных клеток — сосудов и волокон, они образуют сосудисто-волокнистые пучки жилки листа.
По жилкам передвигаются вода и растворенные в ней вещества, жилки придают прочность листу.
Сосудисто-волокнистые пучки жилки образованы, волокнами, сосудами и ситовидными трубками.
Волокна придают листу прочность, они состоят из отдельных сильно вытянутых клеток с утолщенными стенками.
Ситовидные трубки в отличие от сосудов образованы живыми клетками, поперечные перегородки между которыми продырявлены как сито. По ситовидным трубкам из листьев во все органы растения передвигаются органические вещества.
Клеточное строение листа водного растения элодеи, которую часто разводят в аквариумах, можно рассмотреть под микроскопом, даже не делая среза.
Рис. 53. Нижняя кожица листа с устьичными клетками: 1 — устьичные клетки; 2 — устьичная щель.
Рис. 54. Схема клеточного строения листа: 1 — клетки верхней кожицы; 2 — клетки мякоти листа; 3 — клетки жилки листа; 4 — межклетники; 5 — устьице; б — клетки нижней кожицы.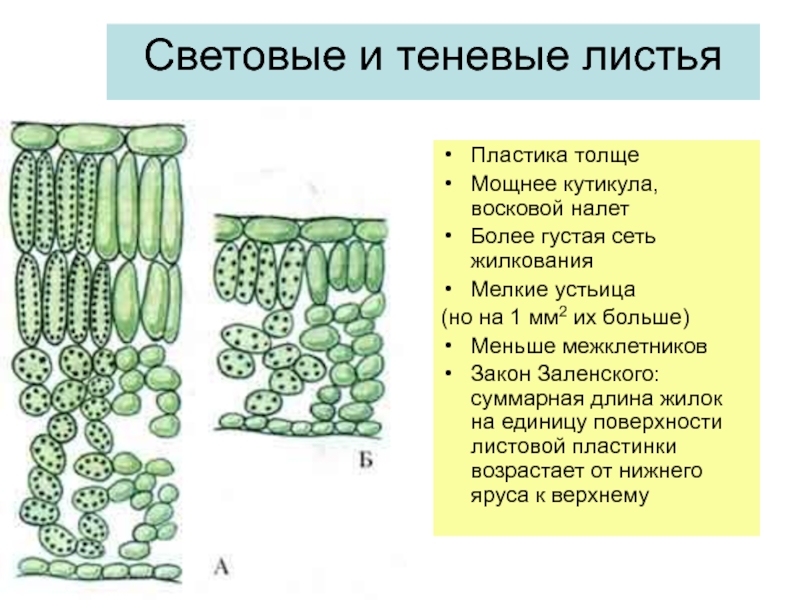
Хлорофилл легко выделить, положив лист в горячий спирт. Лист станет бесцветным, а спирт окрасится в ярко-зеленый цвет.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Просмотров: 4 015
Аэренхима
ОСНОВНЫЕ ТКАНИ
Основная ткань,
или паренхима, занимает большую часть пространства между другими постоянными
тканями стеблей, корней и других органов растения. Основные ткани состоят в
основном из живых клеток, разнообразных по форме. Клетки тонкостенные, но
иногда утолщенные и одревесневшие, с постенной цитоплазмой, простыми порами. Из
паренхимы состоят кора стеблей и корней, сердцевина стеблей, корневищ, мякоть
сочных плодов и листьев, она служит хранилищем питательных веществ в семенах. В
различных органах растения основная паренхима выполняет неодинаковые функции,
поэтому выделяют несколько подгрупп основных тканей: ассимиляционную,
запасающую, водоносную и воздухоносную.
В
различных органах растения основная паренхима выполняет неодинаковые функции,
поэтому выделяют несколько подгрупп основных тканей: ассимиляционную,
запасающую, водоносную и воздухоносную.
Ассимиляционная ткань, или хлорофиллоносная паренхима, или хлоренхима, — ткань, в которой осуществляется фотосинтез. Клетки тонкостенны, содержат хлоропласты, ядро. Хлоропласты, как и цитоплазма, расположены постенно. Хлоренхима находится непосредственно под кожицей. В основном хлоренхима сосредоточена в листьях и молодых зеленых побегах растений. В листьях различают палисадную, или столбчатую, и губчатую хлоренхиму (рис. 26). Клетки палисадной хлоренхимы удлиненные, цилиндрической формы, с очень узкими межклетниками. Губчатая хлоренхима имеет более
43
Рис. 26. Ассимиляционная ткань листа бука на поперечном срезе: А — световой лист; Б — теневой лист: 1 — эпидермис; 2 — устьице; 3 — палисадная ткань; 4 — губчатая паренхима; 5 — проводящий пучок
или менее
округлые рыхло расположенные клетки с большим количеством межклетников,
заполненных воздухом.
Аэренхима, или воздухоносная ткань, — паренхима со значительно развитыми межклетниками в разных органах характерна для водных, прибрежно-водных и болотных растений (камыши, ситники, кубышки, рдесты, водокрасы и др.), корни и корневища которых находятся в иле, бедном кислородом (см. рис. 89). Атмосферный воздух доходит до подводных органов через фотосинтетическую систему посредством передаточных клеток. Кроме того, воздухоносные межклетники сообщаются с атмосферой с помощью своеобразных пневматод — устьиц листьев и стеблей, пневматод воздушных корней некоторых растений (монстера, филодендрон, фикус баньян и др.), щелей, отверстий, каналов, окруженных клетками-регуляторами сообщений. Аэренхима уменьшает удельный вес растения, что, вероятно, способствует поддержанию вертикального положения водных растений, а водным растениям с плавающими на поверхности воды листьями — удержанию листьев на поверхности воды.
Водоносная ткань
запасает воду в листьях и стеблях суккулентных растений (кактусы, алоэ, агавы,
толстянки и др. ), а также растений засоленных местообитаний (солерос, биюргун,
сарсазан, солянки, гребенщик, черный саксаул и др.), как правило, в аридных
областях. Листья злаков также имеют крупные водоносные
), а также растений засоленных местообитаний (солерос, биюргун,
сарсазан, солянки, гребенщик, черный саксаул и др.), как правило, в аридных
областях. Листья злаков также имеют крупные водоносные
44
клетки со слизистыми веществами, удерживающими влагу. Хорошо развитые водоносные клетки имеет мох сфагнум (рис. 27).
Запасающие ткани — ткани, в которых в определенный период развития растения откладывают продукты обмена — белки, углеводы, жиры и др. Клетки запасающей ткани обычно тонкостенны, паренхима живая. Запасающие ткани широко представлены в клубнях, луковицах, утолщенных корнях, сердцевине стеблей, эндосперме и зародышах семян, паренхиме проводящих тканей (фасоль, ароидные), вместилищах смол и эфирных масел в листьях лавра, камфарного дерева и др. Запасающая ткань может превращаться в хлоренхиму, например, при прорастании клубней картофеля, луковиц луковичных растений.
Рис. 27. Водоносная ткань мха сфагнума: 1 — водоносные клетки; 2 — хлорофиллоносные клетки
30.
 4C: Структура листа, функция и адаптация
4C: Структура листа, функция и адаптацияЛистья имеют множество структур, которые предотвращают потерю воды, транспортируют соединения, способствуют газообмену и защищают растение в целом.
Цели обучения
- Описать внутреннюю структуру и функцию листа
Ключевые моменты
- Эпидермис состоит из верхнего и нижнего эпидермиса; он помогает регулировать газообмен через устьица.
- Эпидермис имеет толщину в один слой, но может иметь больше слоев для предотвращения транспирации.
- Кутикула расположена вне эпидермиса и защищает от потери воды; трихомы отпугивают хищников.
- Мезофилл находится между верхним и нижним эпидермисом; он помогает в газообмене и фотосинтезе через хлоропласты.
- Ксилема переносит воду и минеральные вещества к листьям; флоэма переносит продукты фотосинтеза в другие части растения.
- Растения в холодном климате имеют игольчатые листья, которые уменьшаются в размерах; растения в жарком климате имеют сочные листья, которые помогают экономить воду.

Основные термины
- трихома : волосовидный или чешуйчатый отросток эпидермиса растения
- кутикула : неклеточное защитное покрытие вне эпидермиса многих беспозвоночных и растений
- мезофилл : внутренняя ткань (паренхима) листа, содержащая много хлоропластов.
Структура листа и функция
Внешний слой листа — эпидермис. Он состоит из верхнего и нижнего эпидермиса, которые присутствуют с обеих сторон листа.Ботаники называют верхнюю сторону адаксиальной поверхностью (или адаксисом), а нижнюю — абаксиальной поверхностью (или абаксисом). Эпидермис участвует в регуляции газообмена. Он содержит устьица, которые представляют собой отверстия, через которые происходит газообмен. Каждую устьицу окружают две замыкающие клетки, регулирующие ее открытие и закрытие. Защитные клетки — единственные клетки эпидермиса, содержащие хлоропласты.
Эпидермис обычно имеет толщину в один слой клеток. Однако у растений, которые растут в очень жарких или очень холодных условиях, эпидермис может иметь толщину в несколько слоев для защиты от чрезмерной потери воды в результате транспирации.Восковидный слой, известный как кутикула, покрывает листья всех видов растений. Кутикула снижает скорость потери воды с поверхности листа. Другие листья могут иметь небольшие волоски (трихомы) на поверхности листа. Трихомы помогают предотвратить травоядность, ограничивая движения насекомых или накапливая токсичные или неприятные на вкус соединения. Они также могут снизить скорость транспирации, блокируя поток воздуха через поверхность листа.
Однако у растений, которые растут в очень жарких или очень холодных условиях, эпидермис может иметь толщину в несколько слоев для защиты от чрезмерной потери воды в результате транспирации.Восковидный слой, известный как кутикула, покрывает листья всех видов растений. Кутикула снижает скорость потери воды с поверхности листа. Другие листья могут иметь небольшие волоски (трихомы) на поверхности листа. Трихомы помогают предотвратить травоядность, ограничивая движения насекомых или накапливая токсичные или неприятные на вкус соединения. Они также могут снизить скорость транспирации, блокируя поток воздуха через поверхность листа.
Под эпидермой листьев двудольных находятся слои клеток, известные как мезофилл, или «средний лист». Мезофилл большинства листьев обычно содержит два типа паренхимных клеток: столбчатую паренхиму и губчатую паренхиму. Столбчатая паренхима (также называемая палисадным мезофиллом) способствует фотосинтезу и имеет плотно расположенные клетки в форме столбцов.Он может быть представлен в один, два или три слоя. Под палисадной паренхимой располагаются рыхло расположенные клетки неправильной формы. Это клетки губчатой паренхимы (или губчатого мезофилла). Воздушное пространство между клетками губчатой паренхимы обеспечивает газообмен между листом и внешней атмосферой через устьица. У водных растений межклеточные пространства губчатой паренхимы помогают листу плавать. Оба слоя мезофилла содержат много хлоропластов.
Мезофилл большинства листьев обычно содержит два типа паренхимных клеток: столбчатую паренхиму и губчатую паренхиму. Столбчатая паренхима (также называемая палисадным мезофиллом) способствует фотосинтезу и имеет плотно расположенные клетки в форме столбцов.Он может быть представлен в один, два или три слоя. Под палисадной паренхимой располагаются рыхло расположенные клетки неправильной формы. Это клетки губчатой паренхимы (или губчатого мезофилла). Воздушное пространство между клетками губчатой паренхимы обеспечивает газообмен между листом и внешней атмосферой через устьица. У водных растений межклеточные пространства губчатой паренхимы помогают листу плавать. Оба слоя мезофилла содержат много хлоропластов.
 (б) (внизу) Эти листовые слои хорошо видны на сканирующей электронной микрофотографии. Многочисленные мелкие бугорки в клетках палисадной паренхимы представляют собой хлоропласты. Бугорки, выступающие из нижней поверхности листа, представляют собой железистые трихомы.
(б) (внизу) Эти листовые слои хорошо видны на сканирующей электронной микрофотографии. Многочисленные мелкие бугорки в клетках палисадной паренхимы представляют собой хлоропласты. Бугорки, выступающие из нижней поверхности листа, представляют собой железистые трихомы.Подобно стеблю, лист содержит сосудистые пучки, состоящие из ксилемы и флоэмы.Ксилема состоит из трахеид и сосудов, которые транспортируют воду и минеральные вещества к листьям. Флоэма переносит продукты фотосинтеза от листа к другим частям растения. Один сосудистый пучок, независимо от того, большой он или маленький, всегда содержит ткани как ксилемы, так и флоэмы.
Рисунок \(\PageIndex{1}\): Ксилема и флоэма : На этой сканирующей электронной микрофотографии показаны ксилема и флоэма в сосудистом пучке листа.Адаптация листьев
Виды хвойных растений, которые хорошо себя чувствуют в холодных условиях, такие как ель, пихта и сосна, имеют листья уменьшенного размера и игольчатого вида. Эти игольчатые листья имеют затонувшие устьица и меньшую площадь поверхности — два свойства, помогающие уменьшить потерю воды. В жарком климате такие растения, как кактусы, имеют сочные листья, которые помогают экономить воду. Многие водные растения имеют листья с широкими пластинками, которые могут плавать на поверхности воды; толстая восковая кутикула на поверхности листа, отталкивающая воду.
Эти игольчатые листья имеют затонувшие устьица и меньшую площадь поверхности — два свойства, помогающие уменьшить потерю воды. В жарком климате такие растения, как кактусы, имеют сочные листья, которые помогают экономить воду. Многие водные растения имеют листья с широкими пластинками, которые могут плавать на поверхности воды; толстая восковая кутикула на поверхности листа, отталкивающая воду.
границ | Развитие схизогенных межклеточных пространств у растений
Введение
Газообмен жизненно важен для всех живых организмов.Одноклеточные или мелкие многоклеточные организмы осуществляют газообмен путем диффузии непосредственно между клетками и окружающей средой или через гастроваскулярную полость. Сложные многоклеточные организмы осуществляют газообмен через специализированные системы для поглощения и циркуляции газа. Например, у позвоночных замкнутая кровеносная система, в которой кровь перекачивается сердцем, а газообмен происходит через легкие или жабры. Хотя у растений нет системы кровообращения, у многих развиваются межклеточные пространства (МКП), которые связаны непосредственно с внешней средой и обеспечивают эффективный газообмен через устьица или воздушные поры (Raven, 1996; Jackson and Armstrong, 1999; Evans, 2004).Развитие ИКС для газообмена имеет решающее значение для фотосинтеза и транспирации растений, поскольку коэффициенты диффузии углекислого газа и кислорода в воде в 10 000 раз ниже, чем в воздухе (Voesenek et al., 2006). В листьях сосудистых растений губчатый мезофилл является типичным примером ткани, содержащей ИКС для эффективного газообмена. В результате образования ICS в мезофилле площадь внутренней поверхности листа может быть от 7 до 32 раз больше, чем площадь его внешней поверхности, в зависимости от вида и факторов окружающей среды, таких как интенсивность или качество света (Dale, 1988).
Хотя у растений нет системы кровообращения, у многих развиваются межклеточные пространства (МКП), которые связаны непосредственно с внешней средой и обеспечивают эффективный газообмен через устьица или воздушные поры (Raven, 1996; Jackson and Armstrong, 1999; Evans, 2004).Развитие ИКС для газообмена имеет решающее значение для фотосинтеза и транспирации растений, поскольку коэффициенты диффузии углекислого газа и кислорода в воде в 10 000 раз ниже, чем в воздухе (Voesenek et al., 2006). В листьях сосудистых растений губчатый мезофилл является типичным примером ткани, содержащей ИКС для эффективного газообмена. В результате образования ICS в мезофилле площадь внутренней поверхности листа может быть от 7 до 32 раз больше, чем площадь его внешней поверхности, в зависимости от вида и факторов окружающей среды, таких как интенсивность или качество света (Dale, 1988).
Межклеточные пространства обычны и хорошо развиты в корнях и стеблях водно-болотных растений, произрастающих в гипоксических почвах (Seago et al.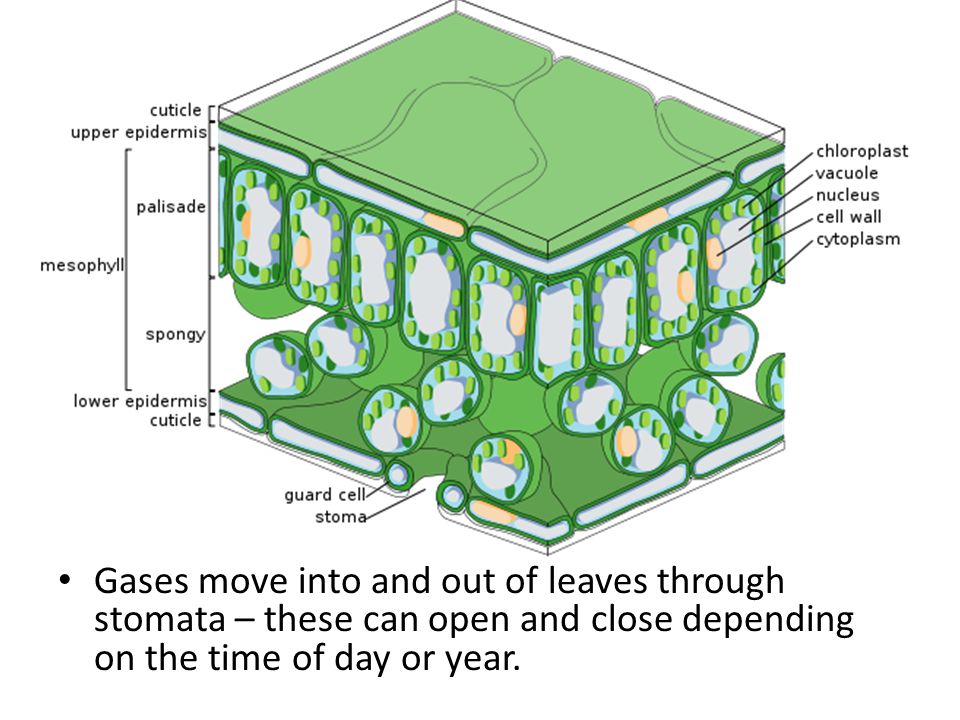 , 2005; Joshi, Kumar, 2012). ИКС также наблюдаются у мохообразных, например, в воздушных камерах гаметофита некоторых печеночников и в подустьичных полостях спорофита мхов и роголистников (Renzaglia et al., 2000). ICS и устьица зарегистрированы в окаменелостях ранних наземных растений, найденных в кремнях Райни, возраст которых составляет примерно 400–412 миллионов лет, что указывает на то, что ICS являются одними из наиболее консервативных признаков в эволюции наземных растений (Edwards et al., 1998). В дополнение к эволюции устьиц, кутикулы, лигнифицированных клеточных стенок и системы водной проводимости интернализация поверхности газообмена с помощью ICS, вероятно, является одной из наиболее важных адаптаций наземных растений к наземным условиям (Raven, 2002).
, 2005; Joshi, Kumar, 2012). ИКС также наблюдаются у мохообразных, например, в воздушных камерах гаметофита некоторых печеночников и в подустьичных полостях спорофита мхов и роголистников (Renzaglia et al., 2000). ICS и устьица зарегистрированы в окаменелостях ранних наземных растений, найденных в кремнях Райни, возраст которых составляет примерно 400–412 миллионов лет, что указывает на то, что ICS являются одними из наиболее консервативных признаков в эволюции наземных растений (Edwards et al., 1998). В дополнение к эволюции устьиц, кутикулы, лигнифицированных клеточных стенок и системы водной проводимости интернализация поверхности газообмена с помощью ICS, вероятно, является одной из наиболее важных адаптаций наземных растений к наземным условиям (Raven, 2002).
Межклеточные пространства у растений образуются путем лизогении или схизогении. Лизигенный ICS вызывается пространственно определенной гибелью клеток, чтобы покинуть пространство. Лизигенное образование ICS происходит в корнях многих видов сельскохозяйственных культур, включая рис (Justin and Armstrong, 1991), кукурузу (He et al. , 1996; Gunawardena et al., 2001) и пшеницы (Trought and Drew, 1980). Напротив, шизогенные ICS образуются путем разделения клеток, а не гибели клеток. Шизогенное образование ICS распространено в корнях и стеблях водно-болотных растений, таких как виды Rumex и Sagittaria . У большинства сосудистых растений ICS в мезофилле листа являются схизогенными, образующимися в результате частичного разделения клеток после разрушения компонентов клеточной стенки (Sachs et al., 1882; Dale, 1988). Шизогенные ICS могут образовываться путем дифференцированного роста, что приводит к отделению соседних клеток друг от друга в средней ламелле клеточных стенок.Этот процесс включает дифференцировку специализированных клеток, которые подвергаются разрушению клеточной стенки, а затем дифференцированно делятся и увеличиваются, чтобы генерировать ICS путем разделения клеток. Хотя процесс образования шизогенных ICS был описан на гистологическом уровне у некоторых видов, его регуляция развития и молекулярные механизмы остаются в значительной степени неизвестными (Jackson and Armstrong, 1999; Evans, 2004).
, 1996; Gunawardena et al., 2001) и пшеницы (Trought and Drew, 1980). Напротив, шизогенные ICS образуются путем разделения клеток, а не гибели клеток. Шизогенное образование ICS распространено в корнях и стеблях водно-болотных растений, таких как виды Rumex и Sagittaria . У большинства сосудистых растений ICS в мезофилле листа являются схизогенными, образующимися в результате частичного разделения клеток после разрушения компонентов клеточной стенки (Sachs et al., 1882; Dale, 1988). Шизогенные ICS могут образовываться путем дифференцированного роста, что приводит к отделению соседних клеток друг от друга в средней ламелле клеточных стенок.Этот процесс включает дифференцировку специализированных клеток, которые подвергаются разрушению клеточной стенки, а затем дифференцированно делятся и увеличиваются, чтобы генерировать ICS путем разделения клеток. Хотя процесс образования шизогенных ICS был описан на гистологическом уровне у некоторых видов, его регуляция развития и молекулярные механизмы остаются в значительной степени неизвестными (Jackson and Armstrong, 1999; Evans, 2004).
Ремоделирование клеточной стенки при разделении клеток
Разделение клеток является критическим аспектом различных процессов развития, включая появление боковых корней, осыпание поврежденных или стареющих органов, высвобождение пыльцы из пыльников и семян из стручков, размягчение плодов и образование ИКС в листьях. мезофилле или в аэренхиме переувлажненных корней.Процесс разделения клеток включает разрушение клеточных стенок в определенном участке ткани (Patterson, 2001). Образование ICS, по-видимому, требует специализированных модификаций клеточной стенки в большинстве зарегистрированных случаев (Knox, 1992). В тканях корня гороха начальной стадии образования ICS предшествует формирование определенного слоя, разделяющего клетки, чтобы открыть щель (Roland, 1978). В листьях фасоли образование ICS происходит схизогенно на вновь образованных клеточных соединениях и включает высоко локализованное разрушение клеточной стенки в родительских клетках, которое продолжается в области средней ламеллы родительских и дочерних клеток (Jeffree et al. , 1986). У семядолей гороха степень разделения в клеточных соединениях связана с локальными утолщениями стенок и электронно-плотными структурами (Kollöffel and Linssen, 1984). Локализованный лизис компонентов клеточной стенки инициирует развитие обширных ICSs в мезофилле листьев кукурузы. Впоследствии эти промежутки расширяются за счет механических напряжений, создаваемых характерными формами клеток. Этот процесс сопровождается локальным утолщением стенки (Apostolakos et al., 1991).
, 1986). У семядолей гороха степень разделения в клеточных соединениях связана с локальными утолщениями стенок и электронно-плотными структурами (Kollöffel and Linssen, 1984). Локализованный лизис компонентов клеточной стенки инициирует развитие обширных ICSs в мезофилле листьев кукурузы. Впоследствии эти промежутки расширяются за счет механических напряжений, создаваемых характерными формами клеток. Этот процесс сопровождается локальным утолщением стенки (Apostolakos et al., 1991).
Процесс абсциссии органов был изучен у различных видов покрытосеменных растений, включая рис, томат и Arabidopsis thaliana .Это обеспечивает модель хорошо организованного разделения клеток, которое включает разрушение материала клеточных стенок между соседними клетками (Patterson, 2001). При абсциссии органов разделение клеток ограничено узкой полосой клеток, составляющих слой абсциссии (Aalen et al., 2013). На первой стадии абсциссии органа клетки в зоне абсциссии (АЗ) дифференцируются с образованием мелких, изодиаметрических и цитоплазматически плотных клеток. Дифференцировка клеток АЗ происходит одновременно с развитием латеральных органов из апикальной меристемы (Sexton, Roberts, 1982; Roberts et al., 2002). Второй этап — активация абсциссии, при которой АЗ-клетки становятся способными реагировать на сигналы абсциссии, а органы готовятся к отделению от растения. Гормоны растений являются основными эндогенными регуляторами этого процесса. В целом этилен и жасмонат действуют как сигналы для ускорения абсциссии, тогда как ауксин, гиббереллины и брассиностероиды ингибируют этот процесс (Estornell et al., 2013). После активации абсциссии происходит расщепление пектиновых полисахаридов в клеточных стенках АЗ с последующим расширением клеток в слое АЗ.Различные ферменты действуют на структурные полисахариды, что приводит к гидролизу средней пластинки и клеточных стенок клеток АЗ. Эти ферменты включают экспансины, ксилоглюканэндотрансглюкозилазу/гидролазы, β -1,4-глюканазы (целлюлазы) и полигалактуроназы (ПГ; Estornell et al., 2013). Активация молекулярного механизма абсциссии включает регуляцию экспрессии многочисленных семейств генов, которые кодируют ферменты ремоделирования клеточной стенки (Cai and Lashbrook, 2008; Ogawa et al.
Дифференцировка клеток АЗ происходит одновременно с развитием латеральных органов из апикальной меристемы (Sexton, Roberts, 1982; Roberts et al., 2002). Второй этап — активация абсциссии, при которой АЗ-клетки становятся способными реагировать на сигналы абсциссии, а органы готовятся к отделению от растения. Гормоны растений являются основными эндогенными регуляторами этого процесса. В целом этилен и жасмонат действуют как сигналы для ускорения абсциссии, тогда как ауксин, гиббереллины и брассиностероиды ингибируют этот процесс (Estornell et al., 2013). После активации абсциссии происходит расщепление пектиновых полисахаридов в клеточных стенках АЗ с последующим расширением клеток в слое АЗ.Различные ферменты действуют на структурные полисахариды, что приводит к гидролизу средней пластинки и клеточных стенок клеток АЗ. Эти ферменты включают экспансины, ксилоглюканэндотрансглюкозилазу/гидролазы, β -1,4-глюканазы (целлюлазы) и полигалактуроназы (ПГ; Estornell et al., 2013). Активация молекулярного механизма абсциссии включает регуляцию экспрессии многочисленных семейств генов, которые кодируют ферменты ремоделирования клеточной стенки (Cai and Lashbrook, 2008; Ogawa et al. , 2009; Estornell et al., 2013). Мутационный анализ указывает на значимость PG в событиях разделения клеток, включая абсциссию (Ogawa et al., 2009). После фактического отделения происходит одревеснение раны от абсцисс, чтобы защитить растение от атаки патогенов.
, 2009; Estornell et al., 2013). Мутационный анализ указывает на значимость PG в событиях разделения клеток, включая абсциссию (Ogawa et al., 2009). После фактического отделения происходит одревеснение раны от абсцисс, чтобы защитить растение от атаки патогенов.
Пептид-лиганд и рецептор-подобная киназа в регуляции разделения клеток
Ключевые генетические компоненты, которые регулируют разделение клеток, были изучены с использованием мутантов с опадением органов цветка A. thaliana . У цветков дикого типа лепестки, чашелистики и нити опадают вскоре после опыления или цветения.Ген INFLORESCENCE DEFICIENT IN ABSCISSION ( IDA ), кодирующий короткий белок с N-концевым сигнальным пептидом, идентифицирован как причинный ген мутантного фенотипа, вызывающего сохранение цветков на растении на неопределенный срок (Бутенко). и др., 2003). Потеря функции генов, кодирующих богатую лейцином повторную рецептороподобную киназу (RLK) HAESA (HAE) и ее близкородственный дублирующий партнер HAESA-Like2 (HSL2), достаточна для блокирования абсциссии органа (Jinn et al.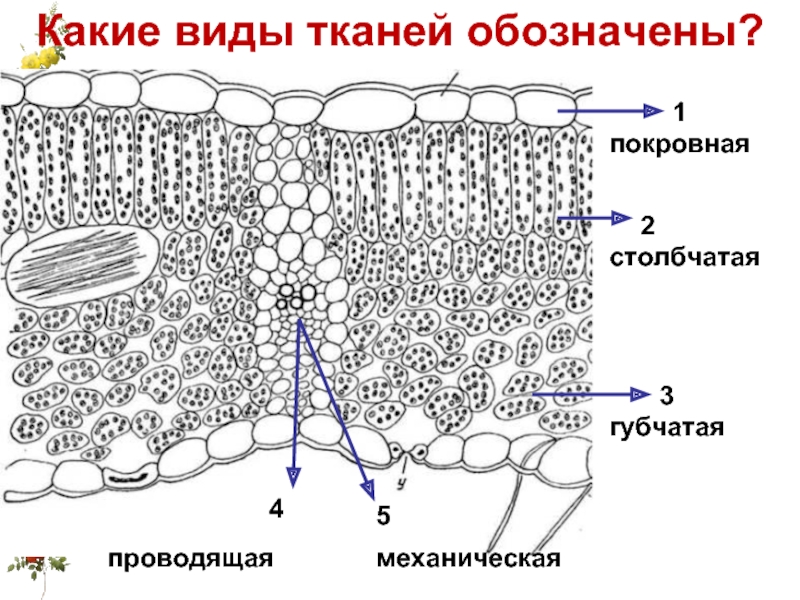 , 2000; Чо и др., 2008). Морфологические исследования органов цветка показывают, что спецификация AZ-клеток не нарушена ни у двойного мутанта ida , ни у hae hsl2 . Это открытие свидетельствует о том, что как IDA , так и HAE/HSL2 экспрессируются после спецификации клеток AZ и способствуют активации скоординированных ферментов ремоделирования клеточной стенки в клетках AZ, что приводит к разделению клеток. Значительные генетические и биохимические данные подтверждают связь лиганд-рецептор между IDA и HAE/HSL2 (Stenvik et al., 2006; Чо и др., 2008 г.; Бутенко и др., 2014). Несколько членов митоген-активируемого киназного каскада, а именно, МИТОГЕН-АКТИВИРОВАННАЯ ПРОТЕИНКИНАЗА-КИНАЗА4 (MKK4), MKK5, МИТОГЕН-АКТИВИРОВАННАЯ ПРОТЕИНКИНАЗА3 (MAPK3) и MAPK6, действуют после передачи сигналов IDA и HAE/HSL2 (Cho et al., 2008). Фактор транскрипции домена MADS AGAMOUS-подобный 15 (AGL15), который регулирует экспрессию HAE посредством прямого связывания с промотором HAE , является прямой мишенью фосфорилирования MAPK.
, 2000; Чо и др., 2008). Морфологические исследования органов цветка показывают, что спецификация AZ-клеток не нарушена ни у двойного мутанта ida , ни у hae hsl2 . Это открытие свидетельствует о том, что как IDA , так и HAE/HSL2 экспрессируются после спецификации клеток AZ и способствуют активации скоординированных ферментов ремоделирования клеточной стенки в клетках AZ, что приводит к разделению клеток. Значительные генетические и биохимические данные подтверждают связь лиганд-рецептор между IDA и HAE/HSL2 (Stenvik et al., 2006; Чо и др., 2008 г.; Бутенко и др., 2014). Несколько членов митоген-активируемого киназного каскада, а именно, МИТОГЕН-АКТИВИРОВАННАЯ ПРОТЕИНКИНАЗА-КИНАЗА4 (MKK4), MKK5, МИТОГЕН-АКТИВИРОВАННАЯ ПРОТЕИНКИНАЗА3 (MAPK3) и MAPK6, действуют после передачи сигналов IDA и HAE/HSL2 (Cho et al., 2008). Фактор транскрипции домена MADS AGAMOUS-подобный 15 (AGL15), который регулирует экспрессию HAE посредством прямого связывания с промотором HAE , является прямой мишенью фосфорилирования MAPK. Фосфорилирование с помощью MAPK снимает репрессию AGL15 экспрессии HAE , что приводит к увеличению передачи сигналов, опосредованной рецептором HAE, и, таким образом, завершает петлю положительной обратной связи, контролирующую опадение цветочных органов (Patharkar and Walker, 2015).Сверхэкспрессия IDA вызывает раннее опадение и образование белого вещества в цветочных AZ. Основными компонентами белого вещества являются моносахариды клеточной стенки арабиноза и галактоза, что позволяет предположить, что IDA действует как положительный регулятор разделения клеток, способствуя разрушению клеточной стенки в AZ (Stenvik et al., 2006; Cho et al., 2008). . Эти эффекты сверхэкспрессии IDA обращаются, когда активность HAE и HSL2 скомпрометирована (Stenvik et al., 2006; Cho et al., 2008). Данные микрочипа предполагают, что модуль передачи сигналов IDAHAE/HSL2 участвует в регуляции генов ремоделирования клеточной стенки (Cai and Lashbrook, 2008).Снижение транскрипции ферментов, модифицирующих клеточную стенку, наблюдается в AZs мутанта hae hsl2 (Cho et al.
Фосфорилирование с помощью MAPK снимает репрессию AGL15 экспрессии HAE , что приводит к увеличению передачи сигналов, опосредованной рецептором HAE, и, таким образом, завершает петлю положительной обратной связи, контролирующую опадение цветочных органов (Patharkar and Walker, 2015).Сверхэкспрессия IDA вызывает раннее опадение и образование белого вещества в цветочных AZ. Основными компонентами белого вещества являются моносахариды клеточной стенки арабиноза и галактоза, что позволяет предположить, что IDA действует как положительный регулятор разделения клеток, способствуя разрушению клеточной стенки в AZ (Stenvik et al., 2006; Cho et al., 2008). . Эти эффекты сверхэкспрессии IDA обращаются, когда активность HAE и HSL2 скомпрометирована (Stenvik et al., 2006; Cho et al., 2008). Данные микрочипа предполагают, что модуль передачи сигналов IDAHAE/HSL2 участвует в регуляции генов ремоделирования клеточной стенки (Cai and Lashbrook, 2008).Снижение транскрипции ферментов, модифицирующих клеточную стенку, наблюдается в AZs мутанта hae hsl2 (Cho et al. , 2008). Кроме того, сигнальный модуль IDA-HAE/HSL2 регулирует экспрессию генов ремоделирования клеточной стенки, что способствует деградации пектиновых полисахаридов во время процесса разделения клеток при появлении боковых корней (Kumpf et al., 2013). Эти данные указывают на то, что сигнальный модуль IDA-HAE/HSL2 был адаптирован для работы в различных процессах разделения клеток корней и побегов.
, 2008). Кроме того, сигнальный модуль IDA-HAE/HSL2 регулирует экспрессию генов ремоделирования клеточной стенки, что способствует деградации пектиновых полисахаридов во время процесса разделения клеток при появлении боковых корней (Kumpf et al., 2013). Эти данные указывают на то, что сигнальный модуль IDA-HAE/HSL2 был адаптирован для работы в различных процессах разделения клеток корней и побегов.
Убиквитинлигаза E3 регулирует разделение клеток при формировании ICS
Слоевище гаметофита печеночника Marchantia polymorpha содержит многослойную ткань с воздушными камерами на дорсальной поверхности (Smith, 1955). Воздушные камеры имеют ICS, который содержит филаменты, богатые хлоропластами, развившиеся из субэпидермиса, и ICS связан непосредственно с внешней атмосферой через воздушные поры в эпидермисе камеры (Barnes and Land, 1907; Apostolakos et al., 1982). Развитие воздушных пор и воздушных камер Marchantia начинается с образования схизогенных ИКС, называемых «инициальными апертурами», между антиклинальными стенками протодермальных клеток в апикальной области слоевища (Apostolakos et al. , 1982). Распространение ICS из начальных отверстий в результате деления клеток и роста протодермальных и субпротодермальных клеток вокруг начальных отверстий приводит к образованию воздушной камеры (рис. 1А; Apostolakos et al., 1982).
, 1982). Распространение ICS из начальных отверстий в результате деления клеток и роста протодермальных и субпротодермальных клеток вокруг начальных отверстий приводит к образованию воздушной камеры (рис. 1А; Apostolakos et al., 1982).
Рис. 1. Формирование схизогенного межклеточного пространства (МКП) у Marchantia polymorpha. (A) Схематическое изображение формирования ICS во время разработки воздушной камеры. После периклинального деления клеток с образованием слоев протодермальных и субпротодермальных клеток ICSs сначала появляются в виде начальной апертуры между антиклинальными стенками протодермальных клеток. Основание начальной апертуры расширяется. Первичная воздушная камера формируется в результате антиклинальных клеточных делений и роста протодермальных и субпротодермальных клеток, окружающих ICS. (B) Гипотетическая модель NOP1-опосредованной передачи сигналов, участвующих в формировании шизогенного ICS. Внеклеточные сигналы, такие как пептидные лиганды, которые воспринимаются рецептором Y, способствуют образованию ICS посредством модуляции регуляторного фактора X, который в противном случае подавляет активацию ферментов ремоделирования клеточной стенки. При модуляции регуляторного фактора X передача сигналов рецептора Y активирует лигазу E3 NOP1 посредством фосфорилирования домена ARM-repeat. NOP1 катализирует убиквитинирование и последующую деградацию регуляторного фактора X, что способствует отделению клеток посредством транскрипционной активации генов ремоделирования клеточной стенки.У nop1 разделение клеток конститутивно подавляется действием регуляторного фактора X.
При модуляции регуляторного фактора X передача сигналов рецептора Y активирует лигазу E3 NOP1 посредством фосфорилирования домена ARM-repeat. NOP1 катализирует убиквитинирование и последующую деградацию регуляторного фактора X, что способствует отделению клеток посредством транскрипционной активации генов ремоделирования клеточной стенки.У nop1 разделение клеток конститутивно подавляется действием регуляторного фактора X.
На протяжении более века M. polymorpha был предметом исследований развития и физиологии. Он считается новым модельным организмом из-за легкости его роста, базового эволюционного положения среди наземных растений, простой генетической архитектуры и генетической трансформируемости (Ishizaki et al., 2008; Kubota et al., 2013), а также простоты генерирования. таргетинг (Ishizaki et al., 2013а; Сугано и др., 2014). Используя эффективный протокол трансформации, опосредованной Agrobacterium (Ishizaki et al., 2008), мутант nopperabo1 ( nop1 ), нарушенный в формировании воздушной камеры, был выделен из 10 000 линий, меченных Т-ДНК (Ishizaki et al. ., 2013б). У nop1 нарушено разделение клеточных стенок во время формирования начальной апертуры. Каузальный ген nop1 был идентифицирован как NOPPERABO1 ( NOP1 ), который кодирует растительный белок U-box (PUB), несущий тандемные повторы ARMADILLO (ARM) на С-конце и локализующийся на плазматической мембране ( Ишизаки и др., 2013б).
., 2013б). У nop1 нарушено разделение клеточных стенок во время формирования начальной апертуры. Каузальный ген nop1 был идентифицирован как NOPPERABO1 ( NOP1 ), который кодирует растительный белок U-box (PUB), несущий тандемные повторы ARMADILLO (ARM) на С-конце и локализующийся на плазматической мембране ( Ишизаки и др., 2013б).
Домен U-box похож по структуре на мотив пальца RING. Белки U-box функционируют как убиквитинлигазы E3 и катализируют перенос убиквитина от убиквитин-конъюгирующего фермента (E2) к мишени для убиквитинирования (Aravind and Koonin, 2000; Ohi et al., 2003). Белки PUB содержат различные организации доменов и во многих случаях несут дополнительные предсказанные домены, такие как домены серин/треонинкиназы, тетратрикопептидного повтора, повтора WD40 и повтора ARM. Повторный домен ARM представляет собой высококонсервативную правую суперспираль из α -спиралей, участвующих в межбелковых взаимодействиях (Samuel et al. , 2006; Тевари и др., 2010). Таким образом, расположение NOP1 в плазматической мембране может зависеть от его С-концевых повторов ARM посредством их взаимодействия с регуляторными белками, расположенными на плазматической мембране или рядом с ней.
, 2006; Тевари и др., 2010). Таким образом, расположение NOP1 в плазматической мембране может зависеть от его С-концевых повторов ARM посредством их взаимодействия с регуляторными белками, расположенными на плазматической мембране или рядом с ней.
Модель функции NOP1 в схизогенной формации ICS
Инициация ICS у Marchantia происходит в углах трех или более протодермальных клеток, расположенных только в области апикальной вырезки (Apostolakos et al., 1982). Чтобы определить положение ремоделирования клеточной стенки для образования ICS, межклеточная связь может участвовать в механизме определения относительного положения протодермальных клеток в области апикальной вырезки.
Недавние данные указывают на связь между белками PUB-ARM и RLK и предполагают роль белков PUB-ARM в качестве потенциальных сигнальных белков для RLK (Gu et al., 1998; Kim et al., 2003; Samuel et al., 2006). , 2008; Mbengue et al., 2010; Lu et al., 2011). Например, у A. thaliana белки PUB-ARM PUB12 и PUB13 снижают активность богатого лейцином повтора RLK FLAGELLIN-SENSING 2 путем убиквитинирования и последующей деградации (Lu et al. , 2011). У Brassica и Arabidopsis , S -рецепторная киназа фосфорилирует ARM-повторный домен белка PUB-ARM ARK1, который действует как позитивный регулятор реакции самонесовместимости (Gu et al., 1998; Самуэль и др., 2008).
, 2011). У Brassica и Arabidopsis , S -рецепторная киназа фосфорилирует ARM-повторный домен белка PUB-ARM ARK1, который действует как позитивный регулятор реакции самонесовместимости (Gu et al., 1998; Самуэль и др., 2008).
NOP1 может быть вовлечен в локализованный в плазматической мембране сигнальный путь RLK, который регулирует образование ICS в воздушных камерах (рис. 1В). Поскольку мутация с потерей функции NOP1 приводит к нарушению отделения клеточной стенки, NOP1 может катализировать убиквитинирование и последующую деградацию регуляторного фактора (факторов), которые подавляют отделение клеточной стенки (рис. 1В). Идентификация NOP1-ассоциированных белков будет иметь решающее значение для понимания молекулярных механизмов образования ICS.
Как обсуждалось выше, сигнальный модуль IDA-HAE/HSL2 регулирует разделение клеток посредством транскрипционной активации генов ремоделирования клеточной стенки, таких как экспансины, ксилоглюканэндотрансглюкозилаза/гидролазы, целлюлазы и PG у A. thaliana (Cai и Lashbrook, 2008; Cho et al., 2008; Kumpf et al., 2013). В начальном процессе формирования воздушной камеры M. polymorpha происходит локальное утолщение и последующая отслойка клеточной стенки с образованием начального отверстия на стыке трех-пяти протодермальных клеток (Apostolakos, Galatis, 1985).В этом процессе участвует E3-лигаза PUB-ARM NOP1 (Ishizaki et al., 2013b). Мохообразные демонстрируют некоторые отличительные черты с точки зрения состава клеточной стенки по сравнению с сосудистыми растениями, т. Е. Отсутствие лигнифицированной вторичной клеточной стенки и уменьшенное количество сшитого боратом рамногалактуронана II в первичной клеточной стенке. Однако первичная клеточная стенка мохообразных имеет некоторые общие компоненты с сосудистыми растениями, такие как микрофибриллы целлюлозы, маннозосодержащие гемицеллюлозы и ксилоглюканы (Matsunaga et al., 2004; Саркар и др., 2009). В геноме мха Physcomitrella patens были идентифицированы гомологи генов ремоделирования клеточной стенки для экспансинов, ксилоглюканэндотрансглюкозилазы/гидролазы, целлюлазы и PG (Davison and Blaxter, 2005; Carey and Cosgrove, 2007; Eklöf and Brumer, 2010; Маккарти и др.
thaliana (Cai и Lashbrook, 2008; Cho et al., 2008; Kumpf et al., 2013). В начальном процессе формирования воздушной камеры M. polymorpha происходит локальное утолщение и последующая отслойка клеточной стенки с образованием начального отверстия на стыке трех-пяти протодермальных клеток (Apostolakos, Galatis, 1985).В этом процессе участвует E3-лигаза PUB-ARM NOP1 (Ishizaki et al., 2013b). Мохообразные демонстрируют некоторые отличительные черты с точки зрения состава клеточной стенки по сравнению с сосудистыми растениями, т. Е. Отсутствие лигнифицированной вторичной клеточной стенки и уменьшенное количество сшитого боратом рамногалактуронана II в первичной клеточной стенке. Однако первичная клеточная стенка мохообразных имеет некоторые общие компоненты с сосудистыми растениями, такие как микрофибриллы целлюлозы, маннозосодержащие гемицеллюлозы и ксилоглюканы (Matsunaga et al., 2004; Саркар и др., 2009). В геноме мха Physcomitrella patens были идентифицированы гомологи генов ремоделирования клеточной стенки для экспансинов, ксилоглюканэндотрансглюкозилазы/гидролазы, целлюлазы и PG (Davison and Blaxter, 2005; Carey and Cosgrove, 2007; Eklöf and Brumer, 2010; Маккарти и др. , 2014). Чтобы достичь хорошо организованного образования ICS, NOP1 может быть вовлечен в передачу сигналов RLK сходным образом с IDA-HAE/HSL2, который регулирует транскрипционную активность генов ремоделирования клеточной стенки.Необходимы дальнейшие сравнительные геномные исследования, чтобы пролить свет на генетические и биохимические процессы ремоделирования клеточной стенки при образовании ICS M. polymorpha и способствовать нашему пониманию фундаментального механизма механизма ремоделирования клеточной стенки и его эволюции в наземные растения.
, 2014). Чтобы достичь хорошо организованного образования ICS, NOP1 может быть вовлечен в передачу сигналов RLK сходным образом с IDA-HAE/HSL2, который регулирует транскрипционную активность генов ремоделирования клеточной стенки.Необходимы дальнейшие сравнительные геномные исследования, чтобы пролить свет на генетические и биохимические процессы ремоделирования клеточной стенки при образовании ICS M. polymorpha и способствовать нашему пониманию фундаментального механизма механизма ремоделирования клеточной стенки и его эволюции в наземные растения.
Заключение
Развитие ICS считается важным событием в эволюции наземных растений, но мало что известно о задействованных молекулярных механизмах.Процесс образования шизогенных ICS включает хорошо организованное ремоделирование клеточной стенки и может иметь общий регуляторный механизм с процессом абсциссии органов. В последние годы был достигнут значительный прогресс в понимании многих регуляторных аспектов абсциссии органов с использованием модельного растения A. thaliana . Внеклеточный сигнальный модуль, опосредованный пептидными лигандами и молекулами RLK, активирует транскрипцию генов, которые кодируют ферменты, ремоделирующие клеточную стенку, тем самым способствуя отщеплению органов и появлению боковых корешков.Знание механизмов абсциссии органов может быть полезным для разработки генетических и молекулярных стратегий для понимания формирования шизогенного ICS. Поскольку M. polymorpha имеет простую архитектуру генома и поддается как трансформации (Ishizaki et al., 2008; Kubota et al., 2013), так и целенаправленной модификации генома (Ishizaki et al., 2013a; Sugano et al., 2014), он должен быть эффективным модельным организмом для изучения образования ИКС на молекулярном уровне путем изучения развития воздушной камеры.Дальнейшие исследования NOP1-опосредованного развития воздушных камер у M. polymorpha будут способствовать нашему пониманию фундаментального регуляторного механизма и эволюции разделения клеток у наземных растений.
thaliana . Внеклеточный сигнальный модуль, опосредованный пептидными лигандами и молекулами RLK, активирует транскрипцию генов, которые кодируют ферменты, ремоделирующие клеточную стенку, тем самым способствуя отщеплению органов и появлению боковых корешков.Знание механизмов абсциссии органов может быть полезным для разработки генетических и молекулярных стратегий для понимания формирования шизогенного ICS. Поскольку M. polymorpha имеет простую архитектуру генома и поддается как трансформации (Ishizaki et al., 2008; Kubota et al., 2013), так и целенаправленной модификации генома (Ishizaki et al., 2013a; Sugano et al., 2014), он должен быть эффективным модельным организмом для изучения образования ИКС на молекулярном уровне путем изучения развития воздушной камеры.Дальнейшие исследования NOP1-опосредованного развития воздушных камер у M. polymorpha будут способствовать нашему пониманию фундаментального регуляторного механизма и эволюции разделения клеток у наземных растений.
Заявление о конфликте интересов
Автор заявляет, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Я благодарю Мию Мизутани, Рюичи Нисихаму и Такаюки Кочи за полезные обсуждения.Эта работа была поддержана грантами MEXT KAKENHI для научных исследований в области инноваций (25114510 для KI).
Каталожные номера
Аален, Р. Б., Вильдхаген, М., Сто, И. М., и Бутенко, М. А. (2013). IDA: пептидный лиганд, регулирующий процессы разделения клеток у Arabidopsis . Дж. Экспл. Бот. 64, 5253–5261. doi: 10.1093/jxb/ert338
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Апостолакос П. и Галатис Б. (1985).Исследования развития воздушных пор и воздушных камер Marchantia Palaacea II. Ультраструктура образования начальной апертуры с особым упором на центры организации корковых микротрубочек. Кан. Дж. Бот. 63, 744–756.
Кан. Дж. Бот. 63, 744–756.
Академия Google
Апостолакос П., Галатис Б. и Митракос К. (1982). Исследования развития воздушных пор и воздушных камер Marchantiapalacea I. Световая микроскопия. Энн. Бот. 49, 377–396.дои: 10.1007/BF01276334
Полнотекстовая перекрестная ссылка | Академия Google
Апостолакос П., Галатис Б. и Пантерис Э. (1991). Микротрубочки в клеточном морфогенезе и формировании межклеточного пространства в мезофилле листа Zea mays и эпитеме Pilea cadierei . J. Физиол растений. 137, 591–601. doi: 10.1016/S0176-1617(11)80705-4
Полнотекстовая перекрестная ссылка | Академия Google
Барнс, Ч.Р., и Лэнд, В.Дж.Г. (1907). Бриологические документы.I. Происхождение воздушных камер. Бот. Газ. 44, 197–213. дои: 10.1086/329317
Полнотекстовая перекрестная ссылка | Академия Google
Бутенко М.А., Паттерсон С.Е., Грини П.Е., Стенвик Г.Е., Амундсен С.С. , Мандал А. и соавт. (2003). Соцветие с дефицитом абсциссии контролирует абсциссию органов цветка у Arabidopsis и идентифицирует новое семейство предполагаемых лигандов у растений. Растительная клетка 15, 2296–2307. doi: 10.1105/tpc.014365
, Мандал А. и соавт. (2003). Соцветие с дефицитом абсциссии контролирует абсциссию органов цветка у Arabidopsis и идентифицирует новое семейство предполагаемых лигандов у растений. Растительная клетка 15, 2296–2307. doi: 10.1105/tpc.014365
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бутенко М.A., Wildhagen, M., Albert, M., Jehle, A., Kalbacher, H., Aalen, R.B., et al. (2014). Инструменты и стратегии для сопоставления пар рецепторов пептид-лиганд. Растительная клетка 26, 1838–1847. doi: 10.1105/tpc.113.120071
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кай, С., и Лэшбрук, К.С. (2008). Профилирование транскриптома зоны абсциссии тычинок выявляет новых кандидатов для контроля абсциссии: усиленное сохранение цветочных органов в трансгенных растениях, сверхэкспрессирующих Arabidopsis ZINC FINGER PROTEIN2. Завод Физиол. 146, 1305–1321. doi: 10.1104/стр.107.110908
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Кэри, Р.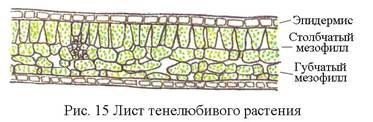 Э., и Косгроув, Д. Дж. (2007). Портрет надсемейства экспансинов у Physcomitrella patens : сравнение с экспансинами покрытосеменных. Энн. Бот. 99, 1131–1141. doi: 10.1093/aob/mcm044
Э., и Косгроув, Д. Дж. (2007). Портрет надсемейства экспансинов у Physcomitrella patens : сравнение с экспансинами покрытосеменных. Энн. Бот. 99, 1131–1141. doi: 10.1093/aob/mcm044
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чо, С. К., Ларю, К. Т., Шевалье, Д., Wang, H., Jinn, T.L., Zhang, S., et al. (2008). Регуляция опадения органов цветка у Arabidopsis thaliana . Проц. Натл. акад. науч. США 105, 15629–15634. doi: 10.1073/pnas.0805539105
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Дейл, Дж. Э. (1988). Контроль расширения листьев. год. Преподобный Завод Физиол. Завод Мол. биол. 39, 267–295. doi: 10.1146/annurev.pp.39.060188.001411
Полнотекстовая перекрестная ссылка | Академия Google
Эдвардс, Д., Керп, Х., и Хасс, Х. (1998). Устьица ранних наземных растений: анатомо-экофизиологический подход. Дж. Экспл. Бот. 49, 255–278. doi: 10. 1093/jxb/49.Special_Issue.255
1093/jxb/49.Special_Issue.255
Полнотекстовая перекрестная ссылка | Академия Google
Эклёф, Дж. М., и Брумер, Х. (2010). Семейство генов XTH: обновленная информация о структуре, функциях и филогении ферментов при ремоделировании ксилоглюкана. Завод Физиол. 153, 456–466. doi: 10.1104/стр.110.156844
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Эсторнелл, Л.Х., Агусти Дж., Мерело П., Талон М. и Тадео Ф. Р. (2013). Выяснение механизмов, лежащих в основе абсциссии органов. Растениевод. 199–200, 48–60. doi: 10.1016/j.plantsci.2012.10.008
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Gu, T., Mazzurco, M., Sulaman, W., Matias, D.D., and Goring, D.R. (1998). Связывание белка повтора плеча с киназным доменом киназы рецептора S-локуса. Проц. Натл. акад. науч. США 95, 382–387. дои: 10.1073/пнас.95.1.382
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гунавардена, А.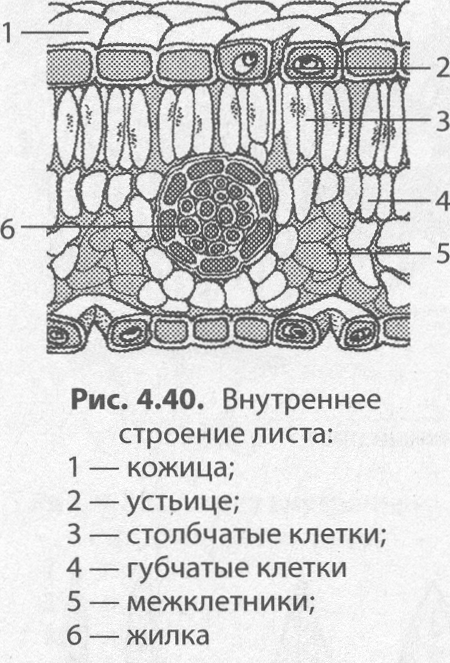 Х., Пирс, Д. М., Джексон, М. Б., Хоуз, Ч. Р., и Эванс, Д. Э. (2001). Характеристика запрограммированной гибели клеток при образовании аэренхимы, индуцированной этиленом или гипоксией, в корнях кукурузы ( Zea mays L.). Планта 212, 205–214. дои: 10.1007/s004250000381
Х., Пирс, Д. М., Джексон, М. Б., Хоуз, Ч. Р., и Эванс, Д. Э. (2001). Характеристика запрограммированной гибели клеток при образовании аэренхимы, индуцированной этиленом или гипоксией, в корнях кукурузы ( Zea mays L.). Планта 212, 205–214. дои: 10.1007/s004250000381
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Хе, К., Финлейсон, С.А., Дрю, М.С., Джордан, В.Р., и Морган, П.В. (1996). Биосинтез этилена при формировании аэренхимы в корнях кукурузы в условиях механического импеданса и гипоксии. Завод Физиол. 112, 1679–1685.
Реферат PubMed | Академия Google
Исидзаки, К., Тиёда, С., Ямато, К.Т., и Кочи, Т. (2008). Agrobacterium — опосредованная трансформация гаплоидного печеночника Marchantia polymorpha L., новая модель для биологии растений. Физиология клеток растений. 49, 1084–1091. doi: 10.1093/pcp/pcn085
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Исидзаки К. , Джозука-Хисатоми Ю., Исида С., Иида С. и Кочи Т. (2013a). Нацеливание генов, опосредованное гомологичной рекомбинацией, в печеночнике Marchantia polymorpha L. Sci. 3, 1532. doi: 10.1038/srep01532
, Джозука-Хисатоми Ю., Исида С., Иида С. и Кочи Т. (2013a). Нацеливание генов, опосредованное гомологичной рекомбинацией, в печеночнике Marchantia polymorpha L. Sci. 3, 1532. doi: 10.1038/srep01532
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Исидзаки К., Мизутани М., Шимамура М., Масуда А., Нишихама, Р., и Кохчи, Т. (2013b). Существенная роль убиквитинлигазы Е3 NOPPERABO1 в формировании схизогенного межклеточного пространства у печеночников Marchantia polymorpha . Растительная клетка 25, 4075–4084. doi: 10.1105/tpc.113.117051
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Джексон М.Б. и Армстронг В. (1999). Формирование аэренхимы и процессы вентиляции растений в связи с затоплением и затоплением почвы. Растение Биол. 1, 274–287. doi: 10.1111/j.1438-8677.1999.tb00253.x
Полнотекстовая перекрестная ссылка | Академия Google
Джеффри, К.Э., Дейл, Дж. Э., и Фрай, С.К. (1986). Генезис межклеточных пространств в развивающихся листьях Phaseolus vulgaris L. Protoplasma 132, 90–98. дои: 10.1007/BF01275795
Э., и Фрай, С.К. (1986). Генезис межклеточных пространств в развивающихся листьях Phaseolus vulgaris L. Protoplasma 132, 90–98. дои: 10.1007/BF01275795
Полнотекстовая перекрестная ссылка | Академия Google
Джинн, Т.Л., Стоун, Дж.М., и Уокер, Дж.К. (2000). HAESA, Arabidopsis , богатая лейцином киназа рецептора повторов, контролирует опадение цветочных органов. Гены Дев. 14, 108–117.
Реферат PubMed | Академия Google
Джоши Р. и Кумар П. (2012). Лизигенное образование аэренхимы включает неапоптозную запрограммированную гибель клеток в корнях риса ( Oryza sativa L.). Физиол. Мол. биол. Завод 18, 1–9. doi: 10.1007/s12298-011-0093-3
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Джастин, С.Х.Ф.В., и Армстронг, В. (1991). Доказательства участия этена в формировании аэренхимы в придаточных корнях риса ( Oryza sativa L.). Новый фитол. 118, 49–62. doi: 10. 1111/j.1469-8137.1991.tb00564.x
1111/j.1469-8137.1991.tb00564.x
Полнотекстовая перекрестная ссылка | Академия Google
Ким, М., Чо, Х.С., Ким, Д.М., Ли, Дж.Х., и Пай, Х.С. (2003). CHRK1, родственная хитиназе рецептороподобная киназа, взаимодействует с NtPUB4, белком повторов броненосца, в табаке. Биохим. Биофиз. Acta 1651, 50–59. doi: 10.1016/S1570-9639(03)00234-6
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Нокс, Дж.П. (1992). Клеточная адгезия, разделение клеток и морфогенез растений. Завод J. 2, 137–141. doi: 10.1111/j.1365-313X.1992.00137.x
Полнотекстовая перекрестная ссылка | Академия Google
Коллёффель, К., и Линссен, П.В.Т. (1984). Формирование межклетников в семядолях развивающихся и прорастающих семян гороха. Протоплазма 120, 12–19. дои: 10.1007/BF01287613
Полнотекстовая перекрестная ссылка | Академия Google
Кубота А., Ишизаки К., Хосака, М., и Кохчи, Т. (2013). Эффективная Agrobacterium опосредованная трансформация печеночника Marchantia polymorpha с использованием регенерирующих талломов.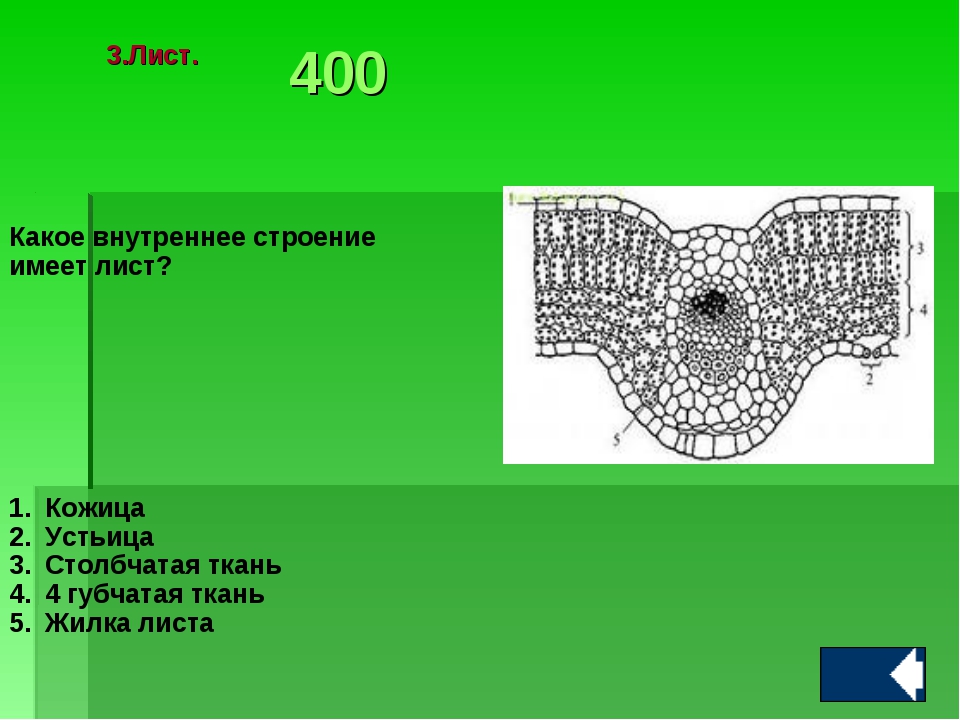 Биологи. Биотехнолог. Биохим. 77, 167–172. doi: 10.1271/bbb.120700
Биологи. Биотехнолог. Биохим. 77, 167–172. doi: 10.1271/bbb.120700
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Kumpf, R.P., Shi, C.L., Larrieu, A., Sto, I.M., Butenko, M.A., Peret, B., et al. (2013). Пептид абсциссии цветочных органов IDA и его рецепторы HAE/HSL2 контролируют разделение клеток во время появления боковых корней. Проц. Натл. акад. науч. США 110, 5235–5240. doi: 10.1073/pnas.1210835110
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лу, Д., Лин, В., Гао, X., Ву, С., Ченг, С., Авила, Дж., и др. (2011). Прямое убиквитинирование рецептора распознавания образов FLS2 ослабляет врожденный иммунитет растений. Наука 332, 1439–1442. doi: 10.1126/наука.1204903
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Мацунага, Т., Исии Т., Мацумото С., Хигучи М., Дарвилл А., Альбершейм П. и соавт. (2004). Наличие полисахарида первичной клеточной стенки рамногалактуронана II у папоротников и мохообразных. Значение для эволюции сосудистых растений. Завод Физиол. 134, 339–351. doi: 10.1104/стр.103.030072
Значение для эволюции сосудистых растений. Завод Физиол. 134, 339–351. doi: 10.1104/стр.103.030072
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Mbengue, M., Camut, S., De Carvalho-Niebel, F., Deslandes, L., Froidure, S., Klaus-Heisen, D., et al. (2010).Убиквитинлигаза E3 Medicago truncatula PUB1 взаимодействует с симбиотическим рецептором LYK3 и негативно регулирует инфекцию и образование клубочков. Растительная клетка 22, 3474–3488. doi: 10.1105/tpc.110.075861
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Маккарти, Т. В., Дер, Дж. П., Хонаас, Л. А., де Памфилс, К. В., и Андерсон, К. Т. (2014). Филогенетический анализ семейств генов, связанных с пектином, у Physcomitrella patens и девяти других видов растений дает представление об эволюции клеточных стенок. BMC Растение Биол. 14:79. дои: 10.1186/1471-2229-14-79
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Огава М. , Кей П., Уилсон С. и Суэйн С. М. (2009). ПОЛИГАЛАКТУРОНАЗЫ 1 (ADPG1), ADPG2 и QUARTET2 арабидопсиса DEHISCENCE ZONE являются полигалактуроназами, необходимыми для разделения клеток во время репродуктивного развития у Arabidopsis . Растительная клетка 21, 216–233. doi: 10.1105/tpc.108.063768
, Кей П., Уилсон С. и Суэйн С. М. (2009). ПОЛИГАЛАКТУРОНАЗЫ 1 (ADPG1), ADPG2 и QUARTET2 арабидопсиса DEHISCENCE ZONE являются полигалактуроназами, необходимыми для разделения клеток во время репродуктивного развития у Arabidopsis . Растительная клетка 21, 216–233. doi: 10.1105/tpc.108.063768
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Охи, М.Д., Вандер Коой, К.В., Розенберг, Дж.А., Чазин, В.Дж., и Гулд, К.Л. (2003). Структурное понимание U-box, домена, связанного с множественным убиквитинированием. Нац. Структура биол. 10, 250–255. doi: 10.1038/nsb906
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Патаркар, О. Р., и Уокер, Дж. К. (2015). Опадение цветочных органов регулируется петлей положительной обратной связи. Проц. Натл. акад. науч. США 112, 2906–2911. doi: 10.1073/pnas.1423595112
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Рэйвен, Дж.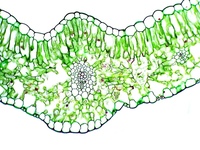 А. (1996). В пустоты: распределение, функции, развитие и поддержание газовых пространств в растениях. Энн. Бот. 78, 137–142. doi: 10.1006/anbo.1996.0105
А. (1996). В пустоты: распределение, функции, развитие и поддержание газовых пространств в растениях. Энн. Бот. 78, 137–142. doi: 10.1006/anbo.1996.0105
Полнотекстовая перекрестная ссылка | Академия Google
Рэйвен, Дж. А. (2002). Давление отбора на эволюцию устьиц. Новый фитол. 153, 371–386. doi: 10.1046/j.0028-646X.2001.00334.x
Полнотекстовая перекрестная ссылка | Академия Google
Ренцалья, К.С., Дафф, Р.Дж.Т., Никрент, Д.Л., и Гарбари, Д.Дж. (2000). Вегетативные и репродуктивные инновации ранних наземных растений: последствия для единой филогении. Филос. Транс. Р. Соц. Лонд. Б биол. науч. 355, 769–793. doi: 10.1098/rstb.2000.0615
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Робертс, Дж. А., Эллиотт, К. А., и Гонсалес-Карранса, З. Х. (2002). Опадение, расхождение и другие процессы разделения клеток. год. Преподобный завод биол. 53, 131–158. doi: 10.1146/annurev.arplant. 53.092701.180236
53.092701.180236
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Роланд, Дж. К. (1978). Дифференцировка клеточной стенки и этапы раскрытия межклеточного газового пространства. J. Cell Sci. 32, 325–336.
Реферат PubMed | Академия Google
Сакс, Дж., Беннетт, А.В., Вайнс, С.Х., и Тиселтон-Дайер, В.Т. (1882). Учебник ботаники: морфологический и физиологический .Оксфорд: Кларендон Пресс.
Академия Google
Самуэль, М.А., Мудгил, Ю., Солт, Дж.Н., Дельмас, Ф., Рамачандран, С., Чилелли, А., и др. (2008). Взаимодействия между рецепторными киназами S-домена и убиквитинлигазами AtPUB-ARM E3 указывают на консервативный сигнальный путь у Arabidopsis . Завод Физиол. 147, 2084–2095. doi: 10.1104/стр.108.123380
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Саркар, П., Босняга, Э.и Ауэр, М. (2009). Клеточные стенки растений на протяжении всей эволюции: к молекулярному пониманию принципов их строения. Дж. Экспл. Бот. 60, 3615–3635. doi: 10.1093/jxb/erp245
Дж. Экспл. Бот. 60, 3615–3635. doi: 10.1093/jxb/erp245
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Seago, J.L. Jr., Marsh, L.C., Stevens, K.J., Soukup, A., Votrubova, O., et al. (2005). Повторное исследование коры корня цветковых растений водно-болотных угодий на наличие аэренхимы. Энн. Бот. 96, 565–579. дои: 10.1093/аоб/mci211
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Секстон Р. и Робертс Дж. А. (1982). Клеточная биология абсциссии. год. Преподобный Завод Физиол. 33, 133–162. doi: 10.1146/annurev.pp.33.060182.001025
Полнотекстовая перекрестная ссылка | Академия Google
Смит, Г. М. (1955). Cryptogamic Botany , Vol. 2, Мохообразные и папоротникообразные . Лондон: McGraw-Hill Book Company, Inc.
.Академия Google
Стенвик Г.Э., Бутенко М.А., Урбанович Б.Р., Роуз Дж.К. и Аален Р.Б. (2006). Сверхэкспрессия INFLORESCENCE DEFICIENT IN ABSCISSION активирует разделение клеток в рудиментарных зонах абсциссии у Arabidopsis . Растительная клетка 18, 1467–1476. doi: 10.1105/tpc.106.042036
Растительная клетка 18, 1467–1476. doi: 10.1105/tpc.106.042036
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сугано С.С., Ширакава М., Такаги Дж., Мацуда Ю., Шимада Т., Хара-Нисимура И. и др. (2014). CRISPR/Cas9-опосредованный направленный мутагенез в печеночнике Marchantia polymorpha L. Физиология клеток растений. 55, 475–481. doi: 10.1093/pcp/pcu014
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тевари, Р., Бейлс, Э., Бантинг, К. А., и Коутс, Дж. К. (2010). Функции белка-повторителя броненосца: вопросы для маленьких существ. Тренд. Клеточная биол. 20, 470–481. doi: 10.1016/j.tcb.2010.05.003
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Trough, M.C.T., and Drew, M.C. (1980). Развитие повреждения от переувлажнения молодых растений пшеницы в анаэробных растворных культурах. Дж. Экспл. Бот. 31, 1573–1585. doi: 10.1093/jxb/31. 6.1573
6.1573
Полнотекстовая перекрестная ссылка | Академия Google
Voesenek, L.A., Colmer, T.D., Pierik, R., Millenaar, F.F., and Peeters, A.J. (2006). Как растения справляются с полным погружением в воду. Новый фитол. 170, 213–226. doi: 10.1111/j.1469-8137.2006.01692.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Функциональная анатомия листа | СпрингерЛинк
Действия
‘) var buybox = документ.querySelector(«[data-id=id_»+ метка времени +»]»).parentNode ;[].slice.call(buybox.querySelectorAll(«.вариант-покупки»)).forEach(initCollapsibles) функция initCollapsibles(подписка, индекс) { var toggle = подписка.querySelector(«.цена-варианта-покупки») подписка. classList.remove(«расширенный»)
var form = подписка.querySelector(«.форма-варианта-покупки») если (форма) {
var formAction = форма.получить атрибут («действие»)
form.setAttribute(«действие», formAction.replace(«/checkout», «/cart»))
document.querySelector(«#ecommerce-scripts»).addEventListener(«load», bindModal(form, formAction, timestamp, index), false)
} var priceInfo = подписка.querySelector(«.Информация о цене»)
var PurchaseOption = toggle.parentElement если (переключить && форма && priceInfo) {
переключать.setAttribute(«роль», «кнопка»)
toggle.setAttribute(«tabindex», «0») toggle.addEventListener («щелчок», функция (событие) {
var expand = toggle.
classList.remove(«расширенный»)
var form = подписка.querySelector(«.форма-варианта-покупки») если (форма) {
var formAction = форма.получить атрибут («действие»)
form.setAttribute(«действие», formAction.replace(«/checkout», «/cart»))
document.querySelector(«#ecommerce-scripts»).addEventListener(«load», bindModal(form, formAction, timestamp, index), false)
} var priceInfo = подписка.querySelector(«.Информация о цене»)
var PurchaseOption = toggle.parentElement если (переключить && форма && priceInfo) {
переключать.setAttribute(«роль», «кнопка»)
toggle.setAttribute(«tabindex», «0») toggle.addEventListener («щелчок», функция (событие) {
var expand = toggle. getAttribute(«aria-expanded») === «true» || ложный
toggle.setAttribute(«aria-expanded», !expanded)
form.hidden = расширенный
если (! расширено) {
покупкаВариант.classList.add («расширенный»)
} еще {
покупкаOption.classList.remove(«расширенный»)
}
priceInfo.hidden = расширенный
}, ложный)
}
} функция bindModal (форма, formAction, метка времени, индекс) {
var weHasBrowserSupport = window.fetch && Array.from функция возврата () {
var Buybox = EcommScripts ? EcommScripts.Ящик для покупок: ноль
var Modal = EcommScripts ? EcommScripts.Modal : ноль if (weHasBrowserSupport && Buybox && Modal) {
var modalID = «ecomm-modal_» + метка времени + «_» + индекс var modal = новый модальный (modalID)
modal.
getAttribute(«aria-expanded») === «true» || ложный
toggle.setAttribute(«aria-expanded», !expanded)
form.hidden = расширенный
если (! расширено) {
покупкаВариант.classList.add («расширенный»)
} еще {
покупкаOption.classList.remove(«расширенный»)
}
priceInfo.hidden = расширенный
}, ложный)
}
} функция bindModal (форма, formAction, метка времени, индекс) {
var weHasBrowserSupport = window.fetch && Array.from функция возврата () {
var Buybox = EcommScripts ? EcommScripts.Ящик для покупок: ноль
var Modal = EcommScripts ? EcommScripts.Modal : ноль if (weHasBrowserSupport && Buybox && Modal) {
var modalID = «ecomm-modal_» + метка времени + «_» + индекс var modal = новый модальный (modalID)
modal.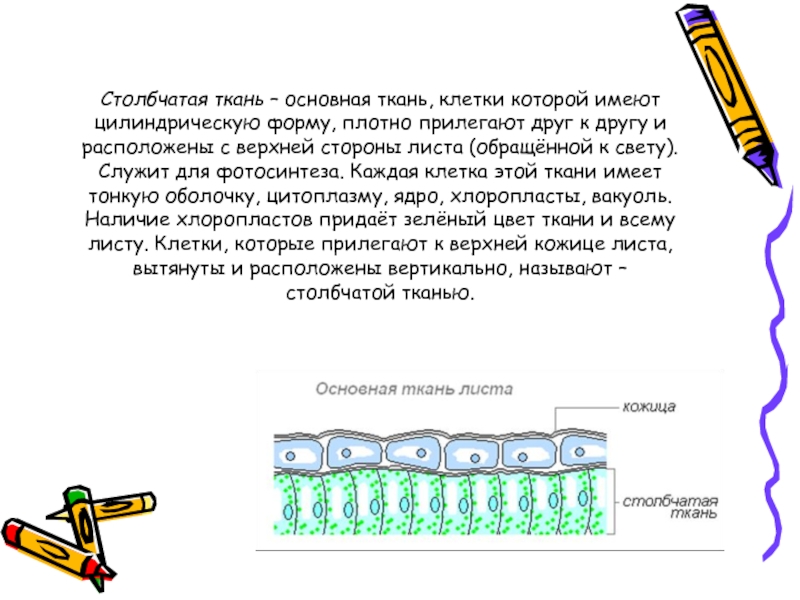 domEl.addEventListener («закрыть», закрыть)
функция закрыть () {
форма.querySelector(«кнопка[тип=отправить]»).фокус()
} форма.setAttribute(
«действие»,
formAction.replace(«/checkout», «/cart?messageOnly=1»)
) form.addEventListener(
«Отправить»,
Buybox.interceptFormSubmit(
Буйбокс.fetchFormAction(окно.fetch),
Buybox.triggerModalAfterAddToCartSuccess(модальный),
консоль.лог,
),
ложный
) document.
domEl.addEventListener («закрыть», закрыть)
функция закрыть () {
форма.querySelector(«кнопка[тип=отправить]»).фокус()
} форма.setAttribute(
«действие»,
formAction.replace(«/checkout», «/cart?messageOnly=1»)
) form.addEventListener(
«Отправить»,
Buybox.interceptFormSubmit(
Буйбокс.fetchFormAction(окно.fetch),
Buybox.triggerModalAfterAddToCartSuccess(модальный),
консоль.лог,
),
ложный
) document.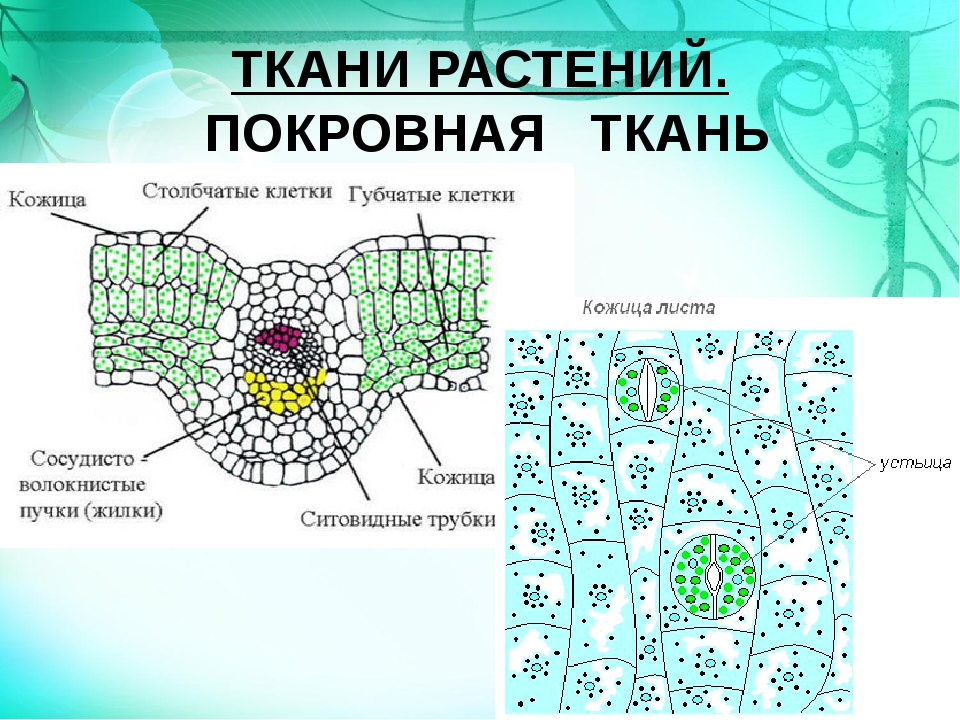 body.appendChild(modal.domEl)
}
}
} функция initKeyControls() {
документ.addEventListener(«keydown», функция (событие) {
if (document.activeElement.classList.contains(«цена-варианта-покупки») && (event.code === «Пробел» || event.code === «Enter»)) {
если (document.activeElement) {
событие.preventDefault()
документ.activeElement.click()
}
}
}, ложный)
} функция InitialStateOpen() {
var buyboxWidth = buybox.смещениеШирина
;[].slice.call(buybox.querySelectorAll(«.опция покупки»)).forEach(функция (опция, индекс) {
var toggle = option.querySelector(«.
body.appendChild(modal.domEl)
}
}
} функция initKeyControls() {
документ.addEventListener(«keydown», функция (событие) {
if (document.activeElement.classList.contains(«цена-варианта-покупки») && (event.code === «Пробел» || event.code === «Enter»)) {
если (document.activeElement) {
событие.preventDefault()
документ.activeElement.click()
}
}
}, ложный)
} функция InitialStateOpen() {
var buyboxWidth = buybox.смещениеШирина
;[].slice.call(buybox.querySelectorAll(«.опция покупки»)).forEach(функция (опция, индекс) {
var toggle = option.querySelector(«. цена-варианта-покупки»)
var form = option.querySelector(«.форма-варианта-покупки»)
var priceInfo = option.querySelector(«.Информация о цене»)
если (buyboxWidth > 480) {
переключить.щелчок()
} еще {
если (индекс === 0) {
переключать.щелчок()
} еще {
toggle.setAttribute («ария-расширенная», «ложь»)
form.hidden = «скрытый»
priceInfo.hidden = «скрытый»
}
}
})
} начальное состояниеОткрыть() если (window.buyboxInitialized) вернуть
window.buyboxInitialized = истина initKeyControls()
})()
цена-варианта-покупки»)
var form = option.querySelector(«.форма-варианта-покупки»)
var priceInfo = option.querySelector(«.Информация о цене»)
если (buyboxWidth > 480) {
переключить.щелчок()
} еще {
если (индекс === 0) {
переключать.щелчок()
} еще {
toggle.setAttribute («ария-расширенная», «ложь»)
form.hidden = «скрытый»
priceInfo.hidden = «скрытый»
}
}
})
} начальное состояниеОткрыть() если (window.buyboxInitialized) вернуть
window.buyboxInitialized = истина initKeyControls()
})()Листья однодольных и двудольных растений
1.
 Листья однодольных и двудольных содержат сходные типы кожных, наземных и сосудистых тканей, но они расположены по-разному в пределах каждого типа листа.
Листья однодольных и двудольных содержат сходные типы кожных, наземных и сосудистых тканей, но они расположены по-разному в пределах каждого типа листа.Листья цветковых растений имеют верхнюю и нижнюю поверхности, при этом верхняя поверхность обычно обращена от земли, а нижняя поверхность обращена к ней.
кожная ткань листа
Как однодольные, так и двудольные листья имеют внешний восковой слой, называемый кутикулой , который покрывает кожную ткань верхнего и нижнего эпидермиса .Кутикула защищает лист и помогает ему удерживать воду. Эпидермис, расположенный под кутикулой, также защищает лист. Он также играет ключевую роль в газообмене , поскольку содержит поры, называемые устьицами . Устьица также в некоторой степени присутствуют на стебле и цветках растения, но в первую очередь они характерны для листьев.
Устьица позволяют углекислому газу проникать в лист и обеспечивают выход водяного пара и кислорода из листа. Каждое устьице окаймлено двумя специализированными паренхиматозными клетками, называемыми замыкающими клетками . Эти клетки открывают и закрывают устьица. Когда тургорное давление в замыкающих клетках высокое, они изгибаются наружу, вызывая открытие устьичной поры. Когда тургорное давление в замыкающих клетках низкое из-за потери воды, устьичная пора закрывается.
Каждое устьице окаймлено двумя специализированными паренхиматозными клетками, называемыми замыкающими клетками . Эти клетки открывают и закрывают устьица. Когда тургорное давление в замыкающих клетках высокое, они изгибаются наружу, вызывая открытие устьичной поры. Когда тургорное давление в замыкающих клетках низкое из-за потери воды, устьичная пора закрывается.
Листовая ткань
Тип основной ткани, называемый мезофиллом , заполняет область между верхней и нижней эпидермисом листа.Клетки мезофилла содержат многочисленные хлоропласты , органеллы, которые осуществляют фотосинтез , превращая свет, воду и углекислый газ в сахар, который растение может расщепить для получения энергии. Кислород является основным побочным продуктом фотосинтеза, что отлично подходит для таких организмов, как люди, которым кислород нужен для дыхания!
Сосудистая ткань листа
В листьях однодольных и двудольных сосудистые пучки окружены одним или несколькими слоями паренхимных клеток, известными как оболочки пучков. Они защищают «жилки» листа. В листьях однодольных фотосинтез осуществляют клетки оболочки пучка, но не всегда в листьях двудольных.
Они защищают «жилки» листа. В листьях однодольных фотосинтез осуществляют клетки оболочки пучка, но не всегда в листьях двудольных.
Оба типа сосудистой ткани играют важную роль в листьях. Ксилема приносит воду и растворенные минералы от корней, а клетки мезофилла используют воду при фотосинтезе. Избыток воды выводится через транспирацию , выделение водяного пара через устьица.Флоэма переносит растворенные сахара, созданные в результате фотосинтеза, к стеблю и корням растения для использования или хранения.
2. Листья однодольных имеют устьица на верхней и нижней сторонах листа, а их жилки проходят параллельно друг другу.
растений | Вариации межклеточных воздушных пространств листа
В ходе эволюции вторжение растений на землю обнажило ткани растений на воздухе, что резко снизило сопротивление диффузии CO 2 в хлоропласты примерно в 10 000 раз.Эволюционное развитие межклеточного воздушного пространства листа было ключевой инновацией, которая позволила наземным растениям извлечь выгоду из пониженной устойчивости к диффузии CO 2 , обеспечиваемой земной средой. Однако наземное обитание также подвергало листья риску высыхания. Предположительно, растения нашли этот компромисс, разработав сложную пространственную клеточную структуру, чтобы создать более или менее извилистую трехмерную сеть межклеточного воздушного пространства, которая позволяла быстро доставлять CO 2 к хлоропластам, сохраняя при этом влажную среду внутри листа. для ограничения потерь воды при испарении.Листья, использующие метаболизм крассуловой кислоты (САМ), демонстрируют значительный контраст по своим свойствам межклеточного воздушного пространства по сравнению с листьями, использующими метаболический путь C 3 для фотосинтеза. САМ-растения часто занимают нижнюю часть спектра объема межклеточного воздушного пространства. По сравнению с листьями C 3 , листья CAM, как правило, имеют более крупные ячейки, которые более плотно упакованы, что обычно приводит к меньшей пористости. Тенденция САМ-растений к более высокой объемной доле клеток обеспечивает большее ночное хранение CO 2 в виде яблочной кислоты в вакуолях, которая затем декарбоксилируется и высвобождается в виде CO 2 , когда устьица закрыты в световой период.
Однако наземное обитание также подвергало листья риску высыхания. Предположительно, растения нашли этот компромисс, разработав сложную пространственную клеточную структуру, чтобы создать более или менее извилистую трехмерную сеть межклеточного воздушного пространства, которая позволяла быстро доставлять CO 2 к хлоропластам, сохраняя при этом влажную среду внутри листа. для ограничения потерь воды при испарении.Листья, использующие метаболизм крассуловой кислоты (САМ), демонстрируют значительный контраст по своим свойствам межклеточного воздушного пространства по сравнению с листьями, использующими метаболический путь C 3 для фотосинтеза. САМ-растения часто занимают нижнюю часть спектра объема межклеточного воздушного пространства. По сравнению с листьями C 3 , листья CAM, как правило, имеют более крупные ячейки, которые более плотно упакованы, что обычно приводит к меньшей пористости. Тенденция САМ-растений к более высокой объемной доле клеток обеспечивает большее ночное хранение CO 2 в виде яблочной кислоты в вакуолях, которая затем декарбоксилируется и высвобождается в виде CO 2 , когда устьица закрыты в световой период. С другой стороны, растения C 3 имеют тенденцию к уменьшению размера клеток, что, по-видимому, способствует большему воздействию поверхности мезофилла на межклеточное воздушное пространство, что позволяет Rubisco более эффективно фиксировать углерод. В этом выпуске Earles, Théroux-Rancourt et al. ( 10.1104/pp.18.00550 ) подробнее рассмотрим характеристики трехмерного (3D) межклеточного воздушного пространства, которые потенциально влияют на проводимость межклеточного воздушного пространства у 19 бромелиевых с CAM или C 3 путями фотосинтеза.Они пересмотрели стандартное уравнение для проводимости межклеточного воздушного пространства, включив в него извилистость, удлинение поперечного пути и связность межклеточного воздушного пространства. Они сообщают о значительных различиях в пористости, извилистости, удлинении бокового пути и связности межклеточного воздушного пространства во всех листьях бромелии.
С другой стороны, растения C 3 имеют тенденцию к уменьшению размера клеток, что, по-видимому, способствует большему воздействию поверхности мезофилла на межклеточное воздушное пространство, что позволяет Rubisco более эффективно фиксировать углерод. В этом выпуске Earles, Théroux-Rancourt et al. ( 10.1104/pp.18.00550 ) подробнее рассмотрим характеристики трехмерного (3D) межклеточного воздушного пространства, которые потенциально влияют на проводимость межклеточного воздушного пространства у 19 бромелиевых с CAM или C 3 путями фотосинтеза.Они пересмотрели стандартное уравнение для проводимости межклеточного воздушного пространства, включив в него извилистость, удлинение поперечного пути и связность межклеточного воздушного пространства. Они сообщают о значительных различиях в пористости, извилистости, удлинении бокового пути и связности межклеточного воздушного пространства во всех листьях бромелии. Их данные показывают, что признаки межклеточного воздушного пространства, которые лучше отражают трехмерную сложность листьев, сильно влияют на проводимость межклеточного воздушного пространства и что следует тщательно учитывать влияние межклеточного воздушного пространства на проводимость мезофилла с учетом анатомии листа.
Их данные показывают, что признаки межклеточного воздушного пространства, которые лучше отражают трехмерную сложность листьев, сильно влияют на проводимость межклеточного воздушного пространства и что следует тщательно учитывать влияние межклеточного воздушного пространства на проводимость мезофилла с учетом анатомии листа.
Внешние особенности, происхождение и внутренняя структура
Внешние особенности, происхождение и внутренняя структура
Таксономисты используют чрезмерное количество терминов в качестве средства для разделения и названия растений. Терминология, применяемая к способу прикрепления листьев к стеблю, например, включает чередующиеся — расположение, показанное на рисунке, — а также противоположные и мутовчатые и основанные на количестве листьев, прикрепленных к каждому узлу: один (чередующийся), два (противоположный) и три или более (мутовчатый).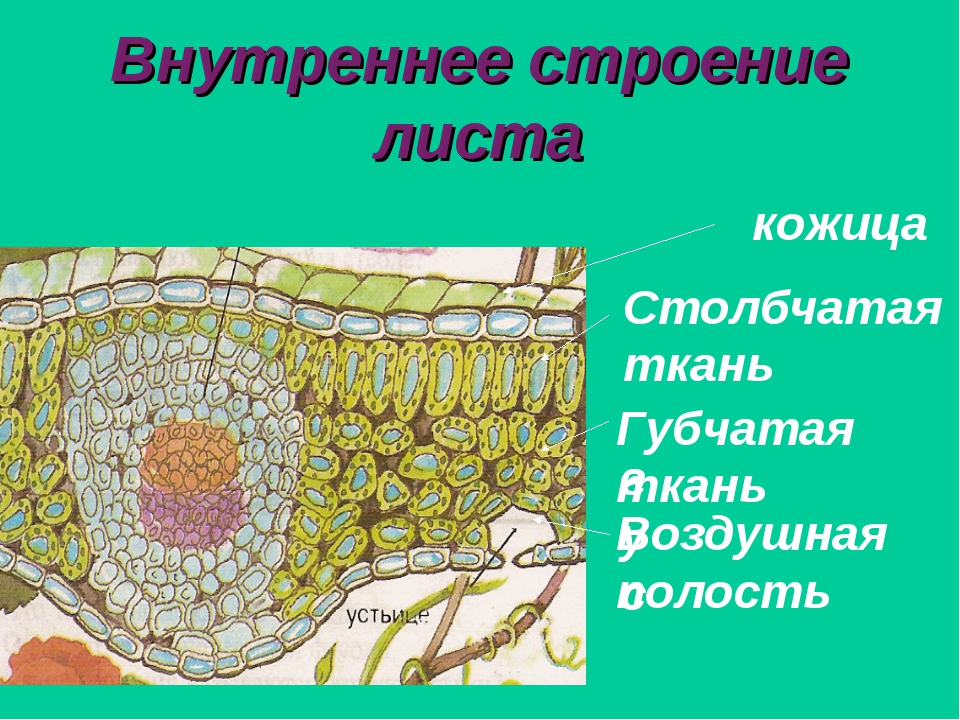 Если к черешку прикрепляется одиночная пластинка, как на рис. , лист простой; , если лист разделен на две или более отдельных частей, лист составной и может быть перистым или пальчатым , в зависимости от того, как листочки (отдельные отдельные части листа) прикреплены к листу. расширение черешка ( рахиса ). Другие стандартные термины используются для обозначения жилкования, общей формы, формы кончика, состояния края пластинки (зубчатый, гладкий, лопастный), волосистого (какие волоски) или гладкого (как на верхней, так и на нижней поверхностях или только на один) и многое другое.
Если к черешку прикрепляется одиночная пластинка, как на рис. , лист простой; , если лист разделен на две или более отдельных частей, лист составной и может быть перистым или пальчатым , в зависимости от того, как листочки (отдельные отдельные части листа) прикреплены к листу. расширение черешка ( рахиса ). Другие стандартные термины используются для обозначения жилкования, общей формы, формы кончика, состояния края пластинки (зубчатый, гладкий, лопастный), волосистого (какие волоски) или гладкого (как на верхней, так и на нижней поверхностях или только на один) и многое другое.
Листья возникают на верхушках побегов стеблей в клетках непосредственно под протодермой. Деление и расширение клеток в этой области приводит к образованию зачатка листа , в котором меристематические области вскоре становятся идентифицируемыми в верхней и нижней областях ткани, предназначенной для превращения в лезвие. Нить прокамбия из побега, след листа , соединяется с дифференцирующимися сосудистыми тканями зачатка, обеспечивая тем самым непрерывность проводящих тканей по всему растению. Область на сосудистом цилиндре стебля, где след листа расходится в зачаток листа, называется листовой щелью , название сбивает с толку; это не отверстие, а область, заполненная клетками паренхимы. «Пробел» относится к отсутствию клеток ксилемы и флоэмы в этой точке сосудистого цилиндра.
Область на сосудистом цилиндре стебля, где след листа расходится в зачаток листа, называется листовой щелью , название сбивает с толку; это не отверстие, а область, заполненная клетками паренхимы. «Пробел» относится к отсутствию клеток ксилемы и флоэмы в этой точке сосудистого цилиндра.
Ткани формирующейся пластинки развиваются быстрее на нижней ( абаксиальная поверхность ), чем на верхней ( адаксиальная поверхность ), в результате чего зачаток изгибается внутрь к верхушке побега.Удлиняющиеся зачатки изгибаются и защищают верхушечную меристему побега. В зачатке клетки делятся и удлиняются, дифференцируясь вниз от кончика, и среди тканей молодой пластинки вскоре появляются характерные для зрелого листа межклетники. Деление клеток прекращается, когда лист становится меньше полного размера, а последующее увеличение заключается в удлинении и расширении клеток и межклетников. Таким образом, листья имеют детерминантный рост, тогда как апикальная меристема с ее клетками, продолжающими бесконечно делиться, имеет неопределенный рост.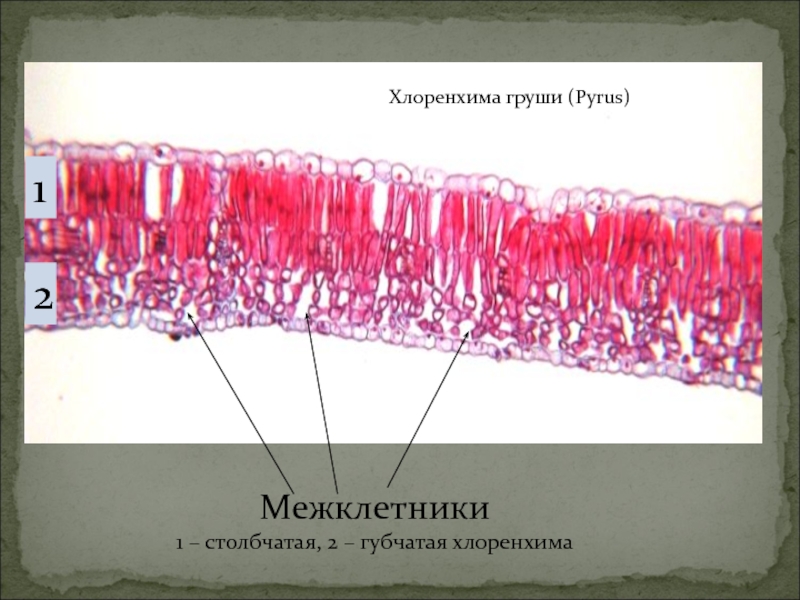
Стандартный лист имеет три участка ткани: эпидермис, мезофилл и сосудистые пучки или жилки (рис. ).
Рисунок 1Эпидермис
Эпидермис листьев представляет собой сплошной слой клеток на всех поверхностях листа, непрерывный, за исключением пор, устьиц ( устьиц , единственное число), которые облегчают газообмен между внутренней частью листа и атмосферой . Клетки паренхимы эпидермиса прилегают друг к другу, как брусчатка, и обычно не содержат хлоропластов, за исключением замыкающих клеток устьиц.Кутикула , состоящая из кутина и воска, откладывается на наружных первичных стенках эпидермальных клеток. У разных видов растений она разная по толщине. Волоски или чешуйки, называемые трихомами , являются отростками эпидермальных клеток и присутствуют на многих листьях. Железы , связанные с трихомами, часто производят вещества, вызывающие отвращение или токсичность для травоядных. Физическое присутствие клубка трихом на поверхности листа также удерживает многих животных от употребления в пищу или использования листа.
Физическое присутствие клубка трихом на поверхности листа также удерживает многих животных от употребления в пищу или использования листа.
Устьица состоят из двух почковидных защитных клеток , окружающих отверстие, устьица и обычно от двух до четырех вспомогательных клеток — обычных клеток паренхимы, форма которых соответствует замыкающим клеткам, поэтому в эпидермальном покрытии не остается отверстий. . (Обратите внимание, что «устьица» относится как к одной маленькой поре, так и ко всему аппарату замыкающих клеток плюс пора.) Стенки замыкающих клеток, обращенные к устьице, толще противоположных стенок и более эластичны.Когда замыкающие клетки наполняются водой (набухают), более тонкие стенки удлиняются быстрее, чем те, которые обращены к поре, тем самым оттягивая последние стенки друг от друга и открывая пору. И наоборот, когда клетки теряют воду и сокращаются (становятся дряблыми), стенки расслабляются и поры закрываются.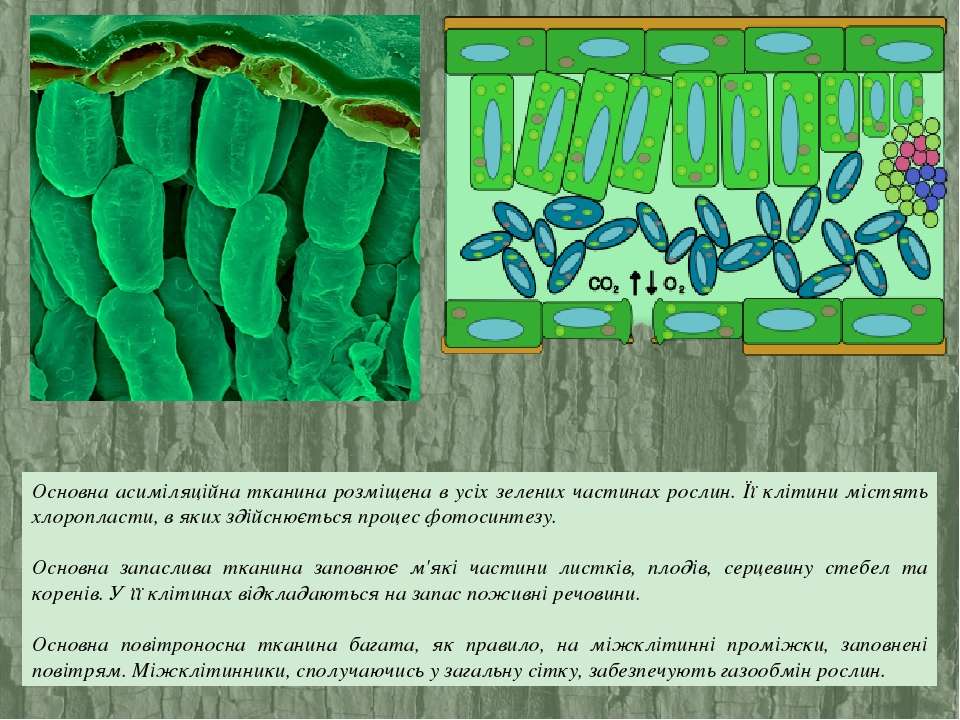 Устьица регулируют прохождение большей части воды из листьев и движение воздуха внутрь и наружу.
Устьица регулируют прохождение большей части воды из листьев и движение воздуха внутрь и наружу.
В зависимости от того, где обитает растение и как ориентированы его листья, устьица могут присутствовать как на верхней, так и на нижней поверхности листьев, исключительно на одной или другой стороне, или полностью отсутствовать на листьях, причем последний случай характерен для погруженных в воду растений. растения.
Мезофилл
Ткань мезофилла составляет основную часть большинства листьев, а хлоропласты в его клетках являются основными местами фотосинтеза. Мезофилл зажат между слоями эпидермиса. В листьях, расположенных горизонтально на стебле и в которых различимы верхняя и нижняя части, верхние и нижние клетки мезофилла имеют различную форму, тогда как в листьях, расположенных вертикально, мезофилл однороден на всем протяжении.
Если мезофилл дифференцирован, верхний слой называется палисадным мезофиллом и состоит из тесно расположенных столбчатых клеток, длинная ось которых проходит под прямым углом к поверхности листа.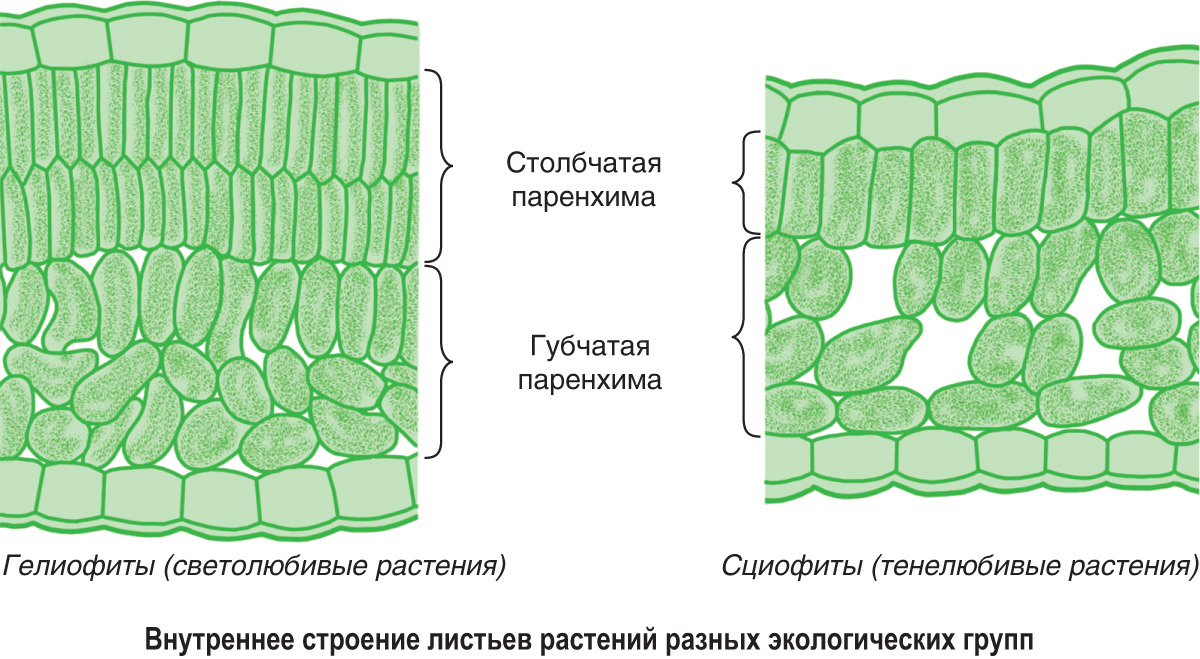 Нижняя ткань, называемая губчатым мезофиллом , состоит из клеток неправильной формы, свободно расположенных с большим межклеточным пространством. Хотя оба типа мезофилла содержат хлоропласты, в частоколе их больше, чем в губчатом мезофилле. Таким образом, мезофилл представляет собой разновидность хлоренхимы — паренхимы, содержащей хлоропласты. Губчатый мезофилл с его воздушными пространствами является, кроме того, аэренхимой.
Нижняя ткань, называемая губчатым мезофиллом , состоит из клеток неправильной формы, свободно расположенных с большим межклеточным пространством. Хотя оба типа мезофилла содержат хлоропласты, в частоколе их больше, чем в губчатом мезофилле. Таким образом, мезофилл представляет собой разновидность хлоренхимы — паренхимы, содержащей хлоропласты. Губчатый мезофилл с его воздушными пространствами является, кроме того, аэренхимой.
Влажные поверхности клеток мезофилла являются местами потери воды и газообмена; устьица — это просто ворота, через которые вода и газы выходят наружу.
Мезофилл содержит укрепляющие ткани, в первую очередь вокруг жилок, но также и отдельными группами по всему мезофиллу. Особенно распространены склероиды и почти всегда для укрепления вен используются клетки колленхимы. Волокна распространены в листьях однодольных.
Вены (сосудистая ткань)
Жилки пронизывают все части листа, образуя сеть, соединяющую лист через черешок с сосудами стебля, а тем самым и с корнем.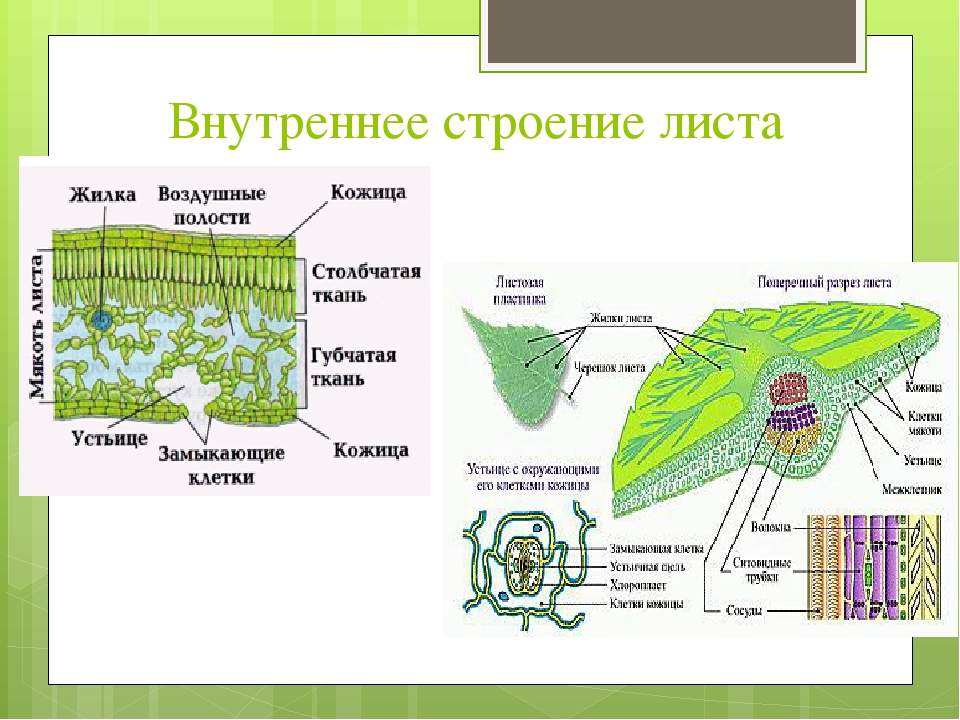 Первичная ксилема клеток занимают верхнюю часть жилки, а флоэма клеток нижнюю. Сосудистые ткани окружены пучковой оболочкой толщиной в один или два слоя, состоящей из волокон в меньших венах и паренхимы в больших.
Первичная ксилема клеток занимают верхнюю часть жилки, а флоэма клеток нижнюю. Сосудистые ткани окружены пучковой оболочкой толщиной в один или два слоя, состоящей из волокон в меньших венах и паренхимы в больших.
Волокна и колленхима присутствуют в жилках и вокруг них и придают прочность им и листу в целом. Расширения оболочки пучка соединяют оболочки пучка с одним или обоими эпидермисами, придавая дополнительную устойчивость лезвию.Крупные вены многократно разветвляются, становясь все меньше каждый раз, когда они делятся, пока в конечном итоге не заканчиваются только одной или двумя трахеидами на конце вены. Здесь клетки мезофилла находятся в непосредственном контакте или, самое большее, на расстоянии одной или двух клеток от сырья, переносимого в ксилему и используемого для фотосинтеза. Не менее удобна флоэма для вывоза фотосинтетов. Оболочки пучков изолируют проводящие ячейки и обеспечивают удержание материалов в трубопроводе.
Жилки тропических трав и других растений с фотосинтезом С4 окружены двумя цилиндрами, внутренний из толстостенных клеток обкладки пучка, внешний из тонкостенных клеток мезофилла.Говорят, что растения C4 имеют анатомию Кранца (от немецкого слова «венок») из-за этого. Кроме того, в листьях С4 отсутствуют отчетливые палисадные или губчатые зоны мезофилла.
%PDF-1.4 % 39 0 объект > эндообъект 38 0 объект >поток StampPDF Batch 2.7 для Solaris — SPDF 10452006-09-18T11:16:39Z2022-02-12T09:41:16-08:002022-02-12T09:41:16-08:00XPPapplication/pdf
uuid:71fa83fd-1dd2-11b2-0a00-ad08276d7200uuid:71fa8400-1dd2-11b2-0a00-b80000000000 0049 Тк 7 0 0 7 51,858 735,8835 Тм
[(17.)-423,6 (Jar)-59,6 (v)-39,8 (is)-337,6 (MC)-337,6 (\(1998\))]TJ
/T1_1 1 тс
9,714 0 тд
[(Завод)-337,6 (Ячейка)-337,6 (Окружающая среда)]TJ
/T1_0 1 тс
8.241 0 Тд
(21:1307\2261310.)Tj
-17,955 -1,2143 Тд
[(18.) -423,6 (Эванс) -337,6 (младший) -337,6 (фон) -337,6 (Кеммерер) -337,6 (S) -337,6 (\(1996\))]TJ
/T1_1 1 тс
17,827 0 тд
[(Растение)-337,6 (Физиол)]TJ
/T1_0 1 тс
5,8719 0 тд
[(110:339)-89,7 (\226346.)]ТД
-23,6989 -1,2143 Тд
[(19.)-423,6 (Гейслер)-337,6 (МДж,)-337,6 (Сак)-337,6 (ФД)-337.6 (\(2002\))]ТДЖ
/T1_1 1 тс
14.4209 0 Тд
[(Новый)-337,6 (Фитол)]TJ
/T1_0 1 тс
5.1522 0 Тд
[(153:469)-89,7 (\226)-89,7 (476.)]TJ
-19,5732 -1,2143 Тд
[(20.)-423,6 (Вебер)-337,6 (Х,)-337,6 (Борисюк)-337,6 (Л,)-337,6 (Вобус)-337,6 (У)-337,6 (\(2005\))]TJ
/T1_1 1 тс
19,2224 0 тд
[(Годовой)-337,6 (Обратный)-337,6 (Завод)-337,6 (Биол)]TJ
/T1_0 1 тс
9,1677 0 тд
(56:253\226279.)Tj
-28,3901 -1,2143 Тд
[(21.)-423,6 (Манц)-194,8 (Б,)-194,8 (Му)529,4 (\250)-24,3 (ллер)-194,8 (К,)-194,8 (Кучера)-194,8 (Б,)-194,8 (Волке)-194.
0049 Тк 7 0 0 7 51,858 735,8835 Тм
[(17.)-423,6 (Jar)-59,6 (v)-39,8 (is)-337,6 (MC)-337,6 (\(1998\))]TJ
/T1_1 1 тс
9,714 0 тд
[(Завод)-337,6 (Ячейка)-337,6 (Окружающая среда)]TJ
/T1_0 1 тс
8.241 0 Тд
(21:1307\2261310.)Tj
-17,955 -1,2143 Тд
[(18.) -423,6 (Эванс) -337,6 (младший) -337,6 (фон) -337,6 (Кеммерер) -337,6 (S) -337,6 (\(1996\))]TJ
/T1_1 1 тс
17,827 0 тд
[(Растение)-337,6 (Физиол)]TJ
/T1_0 1 тс
5,8719 0 тд
[(110:339)-89,7 (\226346.)]ТД
-23,6989 -1,2143 Тд
[(19.)-423,6 (Гейслер)-337,6 (МДж,)-337,6 (Сак)-337,6 (ФД)-337.6 (\(2002\))]ТДЖ
/T1_1 1 тс
14.4209 0 Тд
[(Новый)-337,6 (Фитол)]TJ
/T1_0 1 тс
5.1522 0 Тд
[(153:469)-89,7 (\226)-89,7 (476.)]TJ
-19,5732 -1,2143 Тд
[(20.)-423,6 (Вебер)-337,6 (Х,)-337,6 (Борисюк)-337,6 (Л,)-337,6 (Вобус)-337,6 (У)-337,6 (\(2005\))]TJ
/T1_1 1 тс
19,2224 0 тд
[(Годовой)-337,6 (Обратный)-337,6 (Завод)-337,6 (Биол)]TJ
/T1_0 1 тс
9,1677 0 тд
(56:253\226279.)Tj
-28,3901 -1,2143 Тд
[(21.)-423,6 (Манц)-194,8 (Б,)-194,8 (Му)529,4 (\250)-24,3 (ллер)-194,8 (К,)-194,8 (Кучера)-194,8 (Б,)-194,8 (Волке)-194. 8 (F,)-194,8 (Лейбнер-Мецгер)-194,8 (G)-194,8 (\(2005\))]TJ
/T1_1 1 тс
29,957 0 тд
[(Растение)-194,8 (Физиол)]TJ
/T1_0 1 тс
-28,2687 -1,2143 Тд
[(138:1538)-89,7 (\2261551.)]TJ
-1,6883 -1,2143 Тд
[(22.)-423,6 (Ходдад)-450,1 (WS,)-450,1 (Макналт)-29,6 (y)-450,1 (I,)-450,1 (Требе)19,9 (s)-450,1 (JE,)-450,1 ( А)-29,6 (ндерсон)-450,1 (ЭХ,)-450,1 (Лернк)-450,1 (П)-59,6 (А,)-450,1 (Ян)-450,1 (Л)]TJ
1,6883 -1,2143 Тд
(\(1994\))Tj
/T1_1 1 тс
3,1329 0 тд
(Наука) Tj
/T1_0 1 тс
3.4708 0 Тд
(2:1213\2261215.)Tj
-8,292 -1,2143 Тд
[(23.)-423,6 (Клотенс)-341,7 (П,)-341,7 (Баррет)-341,7 (Р,)-341,7 (Бар)-29,6 (учел,)-341,7 (Дж.)-341,7 (Гигай)- 341,7 (JP,)-341,7 (Schlenker)-341,7 (M)-341,7 (\(1996\))]TJ
/T1_1 1 тс
31,5862 0 тд
[(J)-341,7 (Phys)-341,7 (D)]TJ
-29,8978 -1,2143 Тд
[(Appl)-337,6 (Phys)]TJ
/T1_0 1 тс
4,5844 0 тд
(29:133\226146.)Tj
-6,2727 -1,2143 Тд
[(24.)-423,6(Сн)-29,6(игирёв)-436,3(А,)-436,3(Сн)-29,6(игирёв)-436,3(И,)-436,3(Кон)-436,3(В,)-436,3 (Кузнецов)-436,3 (С,)-436,3 (Щелок)-29.6(ов)-436,3(И)-436,3(\(1995\))]ТЖ
/T1_1 1 тс
32.
8 (F,)-194,8 (Лейбнер-Мецгер)-194,8 (G)-194,8 (\(2005\))]TJ
/T1_1 1 тс
29,957 0 тд
[(Растение)-194,8 (Физиол)]TJ
/T1_0 1 тс
-28,2687 -1,2143 Тд
[(138:1538)-89,7 (\2261551.)]TJ
-1,6883 -1,2143 Тд
[(22.)-423,6 (Ходдад)-450,1 (WS,)-450,1 (Макналт)-29,6 (y)-450,1 (I,)-450,1 (Требе)19,9 (s)-450,1 (JE,)-450,1 ( А)-29,6 (ндерсон)-450,1 (ЭХ,)-450,1 (Лернк)-450,1 (П)-59,6 (А,)-450,1 (Ян)-450,1 (Л)]TJ
1,6883 -1,2143 Тд
(\(1994\))Tj
/T1_1 1 тс
3,1329 0 тд
(Наука) Tj
/T1_0 1 тс
3.4708 0 Тд
(2:1213\2261215.)Tj
-8,292 -1,2143 Тд
[(23.)-423,6 (Клотенс)-341,7 (П,)-341,7 (Баррет)-341,7 (Р,)-341,7 (Бар)-29,6 (учел,)-341,7 (Дж.)-341,7 (Гигай)- 341,7 (JP,)-341,7 (Schlenker)-341,7 (M)-341,7 (\(1996\))]TJ
/T1_1 1 тс
31,5862 0 тд
[(J)-341,7 (Phys)-341,7 (D)]TJ
-29,8978 -1,2143 Тд
[(Appl)-337,6 (Phys)]TJ
/T1_0 1 тс
4,5844 0 тд
(29:133\226146.)Tj
-6,2727 -1,2143 Тд
[(24.)-423,6(Сн)-29,6(игирёв)-436,3(А,)-436,3(Сн)-29,6(игирёв)-436,3(И,)-436,3(Кон)-436,3(В,)-436,3 (Кузнецов)-436,3 (С,)-436,3 (Щелок)-29.6(ов)-436,3(И)-436,3(\(1995\))]ТЖ
/T1_1 1 тс
32.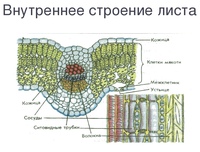 0817 0 тд
[(Rev)-436.3 (Sci)]TJ
-30,3934 -1,2143 Тд
[(Инстр)-29,6 (мкм)]TJ
/T1_0 1 тс
3,6504 0 тд
[(66:5486)-89,7 (\2265492.)]TJ
32,1613 13,3571 Тд
[(25.)-423,7 (Уилк)-29,6 (инс)-394,9 (ЮЗ,)-394,9 (Гуреев)-394,9 (ТЭ,)-394,9 (Гао)-394,9 (Д,)-394,9 (Погани)-394,9 (A,)-394,9 (Стивенсон)-394,9 (AW)-394,9 (\(1996\))]TJ
/T1_1 1 тс
32,4816 0 тд
(Природа)Tj
/T1_0 1 тс
-30,7933 -1,2143 Тд
(384:335\226338.)Tj
-1,6883 -1,2143 Тд
[(26.)-423,7 (Нюджент)-265 (КА,)-265 (Гуреев)-265 (ТЕ,)-265 (Куксон)-265 (ДФ,)-265 (Паган)-29.6(в)-265(Д,)-265(Барни)-265(З)-265(\(1996\))]TJ
/T1_1 1 тс
31,596 0 тд
[(Phys)-265 (Rev)]TJ
-29,9076 -1,2143 Тд
(Письмо) Tj
/T1_0 1 тс
2,0092 0 тд
(77:2961\2262964.)Tj
-3,6975 -1,2143 Тд
[(27.)-423,7 (Клотенс)-321,8 (П,)-321,8 (Людв)-39,8 (иг)-321,8 (Ж,)-321,8 (Бар)-29,6 (учель)-321,8 (Дж,)-321,8 (Ван)-321,8 (Дейк)-321,8 (Д,)-321,8 (Ван)-321,8 (Л)-29,6 (андуй)-29,6 (т)-321,8 (Ж,)-321,8 (Гигай)-321,8 (ЖП, )]ТДЖ
1,6883 -1,2143 Тд
[(Шленкер)-337,6 (М)-337,7 (\(1999\))]TJ
/T1_1 1 тс
9.
0817 0 тд
[(Rev)-436.3 (Sci)]TJ
-30,3934 -1,2143 Тд
[(Инстр)-29,6 (мкм)]TJ
/T1_0 1 тс
3,6504 0 тд
[(66:5486)-89,7 (\2265492.)]TJ
32,1613 13,3571 Тд
[(25.)-423,7 (Уилк)-29,6 (инс)-394,9 (ЮЗ,)-394,9 (Гуреев)-394,9 (ТЭ,)-394,9 (Гао)-394,9 (Д,)-394,9 (Погани)-394,9 (A,)-394,9 (Стивенсон)-394,9 (AW)-394,9 (\(1996\))]TJ
/T1_1 1 тс
32,4816 0 тд
(Природа)Tj
/T1_0 1 тс
-30,7933 -1,2143 Тд
(384:335\226338.)Tj
-1,6883 -1,2143 Тд
[(26.)-423,7 (Нюджент)-265 (КА,)-265 (Гуреев)-265 (ТЕ,)-265 (Куксон)-265 (ДФ,)-265 (Паган)-29.6(в)-265(Д,)-265(Барни)-265(З)-265(\(1996\))]TJ
/T1_1 1 тс
31,596 0 тд
[(Phys)-265 (Rev)]TJ
-29,9076 -1,2143 Тд
(Письмо) Tj
/T1_0 1 тс
2,0092 0 тд
(77:2961\2262964.)Tj
-3,6975 -1,2143 Тд
[(27.)-423,7 (Клотенс)-321,8 (П,)-321,8 (Людв)-39,8 (иг)-321,8 (Ж,)-321,8 (Бар)-29,6 (учель)-321,8 (Дж,)-321,8 (Ван)-321,8 (Дейк)-321,8 (Д,)-321,8 (Ван)-321,8 (Л)-29,6 (андуй)-29,6 (т)-321,8 (Ж,)-321,8 (Гигай)-321,8 (ЖП, )]ТДЖ
1,6883 -1,2143 Тд
[(Шленкер)-337,6 (М)-337,7 (\(1999\))]TJ
/T1_1 1 тс
9. 0491 0 Тд
[(Приложение)-337.6 (физ.)-337,6 (букв.)]TJ
/T1_0 1 тс
6,5935 0 тд
(75:2912\2262914.)Tj
-17,3309 -1,2143 Тд
[(28.)-423,7 (Заблер)-448 (С,)-448 (Клетенс)-448 (П,)-448 (Гигай)-448 (ДП,)-448 (Бар)-29,6 (учел)-448 (J,)-448 (Schlenker)-448 (M)-448 (\(2005\))]TJ
/T1_1 1 тс
32,071 0 тд
[(Rev)-448 (Sci)]TJ
-30,3827 -1,2143 Тд
[(Инстр)-29,6 (мкм)]TJ
/T1_0 1 тс
3,6504 0 тд
(76:073705.)Тдж
-5,3387 -1,2143 Тд
[(29.)-423,7 (Коен)-436,7 (Ж,)-436,7 (Янссен)-436,7 (Г,)-436,7 (Оп)-436,7 (де)-436,7 (Бек)-436,7 (М,)-436,7 (Фургон)-436.7 (Дейк)-436,7 (Д)-436,7 (\(1992\))]TJ
/T1_1 1 тс
29,3156 0 тд
[(Phys)-436.7 (Rev)-436.7 (Lett)]TJ
/T1_0 1 тс
-27,6273 -1,2143 Тд
(69:3743\2263746.)Tj
-1,6883 -1,2143 Тд
[(30.)-423,7(Л)-29,6(абиш)-251,2(ХК,)-251,2(Сегура-Пучаде)19,9(с)-251,2(Дж,)-251,2(Ван)-251,2(Бр)-29,6 (Ussel)-251,2 (D,)-251,2 (Moy)-251,2 (JP)-251,2 (\(1996\))]TJ
/T1_1 1 тс
29.0868 0 тд
[(ESRF)-251.2 (Информационный бюллетень)]TJ
/T1_0 1 тс
-27,3985 -1,2143 Тд
[(25:41\226)-89,7 (43.)]ТД
/T1_2 1 тс
0 Тк 6,5 0 0 6,5 51,6761 39,8835 Тм
(14630)Тж
/T1_3 1 тс
9.
0491 0 Тд
[(Приложение)-337.6 (физ.)-337,6 (букв.)]TJ
/T1_0 1 тс
6,5935 0 тд
(75:2912\2262914.)Tj
-17,3309 -1,2143 Тд
[(28.)-423,7 (Заблер)-448 (С,)-448 (Клетенс)-448 (П,)-448 (Гигай)-448 (ДП,)-448 (Бар)-29,6 (учел)-448 (J,)-448 (Schlenker)-448 (M)-448 (\(2005\))]TJ
/T1_1 1 тс
32,071 0 тд
[(Rev)-448 (Sci)]TJ
-30,3827 -1,2143 Тд
[(Инстр)-29,6 (мкм)]TJ
/T1_0 1 тс
3,6504 0 тд
(76:073705.)Тдж
-5,3387 -1,2143 Тд
[(29.)-423,7 (Коен)-436,7 (Ж,)-436,7 (Янссен)-436,7 (Г,)-436,7 (Оп)-436,7 (де)-436,7 (Бек)-436,7 (М,)-436,7 (Фургон)-436.7 (Дейк)-436,7 (Д)-436,7 (\(1992\))]TJ
/T1_1 1 тс
29,3156 0 тд
[(Phys)-436.7 (Rev)-436.7 (Lett)]TJ
/T1_0 1 тс
-27,6273 -1,2143 Тд
(69:3743\2263746.)Tj
-1,6883 -1,2143 Тд
[(30.)-423,7(Л)-29,6(абиш)-251,2(ХК,)-251,2(Сегура-Пучаде)19,9(с)-251,2(Дж,)-251,2(Ван)-251,2(Бр)-29,6 (Ussel)-251,2 (D,)-251,2 (Moy)-251,2 (JP)-251,2 (\(1996\))]TJ
/T1_1 1 тс
29.0868 0 тд
[(ESRF)-251.2 (Информационный бюллетень)]TJ
/T1_0 1 тс
-27,3985 -1,2143 Тд
[(25:41\226)-89,7 (43.)]ТД
/T1_2 1 тс
0 Тк 6,5 0 0 6,5 51,6761 39,8835 Тм
(14630)Тж
/T1_3 1 тс
9.