
Митохондрии особенности строения и функции таблица. Митохондрии
Митохондрии — это микроскопические мембранные органоиды, которые обеспечивают клетку энергией. Поэтому их называют энергетическими станциями (аккумулятором) клеток.
Митохондрии отсутствуют в клетках простейших организмов, бактерий, энтамеб, которые живут без использования кислорода. Некоторые зеленые водоросли, трипаносомы содержат одну большую митохондрию, а клетки сердечной мышцы, мозга имеют от 100 до 1000 данных органелл.
Особенности строения
Митохондрии относятся к двухмембранным органеллам, имеют внешнюю и внутреннюю оболочки, межмембранное пространство между ними и матрикс.
Внешняя мембрана . Она гладкая, не имеет складок, отграничивает внутреннее содержимое от цитоплазмы. Ширина ее равна 7нм, в составе находятся липиды и белки. Важную роль выполняет порин — белок, образующий каналы во внешней мембране. Они обеспечивают ионный и молекулярный обмен.
Межмембранное пространство .
Внутренняя мембрана . Построена в основном из белка, только треть отводится на липидные вещества. Большое количество белков являются транспортными, так как внутренняя мембрана лишена свободно проходимых пор. Она формирует много выростов – крист, которые выглядят, как приплюснутые гребни. Окисление органических соединений до CO 2 в митохондриях происходит на мембранах крист. Этот процесс кислородзависимый и осуществляется под действием АТФ-синтетазы. Высвобожденная энергия сохраняется в виде молекул АТФ и используется по мере необходимости.
Матрикс – внутренняя среда митохондрий, имеет зернистую однородную структуру. В электронном микроскопе можно увидеть гранулы и нити в клубках, которые свободно лежат между кристами. В матриксе находится полуавтономная система синтеза белка – здесь расположены ДНК, все виды РНК, рибосомы.
Расположение в клетке и деление
Хондриом – это группа митохондрий, которые сосредоточены в одной клетке. Они по-разному располагаются в цитоплазме, что зависит от специализации клеток. Размещение в цитоплазме также зависит от окружающих ее органелл и включений. В клетках растений они занимают периферию, так как к оболочке митохондрии отодвигаются центральной вакуолью. В клетках почечного эпителия мембрана образует выпячивания, между которыми находятся митохондрии.
В стволовых клетках, где энергия используется равномерно всеми органоидами, митохондрии размещены хаотично. В специализированных клетках они, в основном, сосредоточены в местах наибольшего потребления энергии. К примеру, в поперечно-полосатой мускулатуре они расположены возле миофибрилл. В сперматозоидах они спирально охватывают ось жгутика, так как для приведения его в движение и перемещения сперматозоида нужно много энергии.
Деление . Митохондрии способны к самостоятельному размножению, имея собственный геном. Органеллы делятся с помощью перетяжки или перегородок. Формирование новых митохондрий в разных клетках отличается периодичностью, например, в печеночной ткани они сменяются каждые 10 дней.
Функции в клетке
- Основная функция митохондрий – образование молекул АТФ.
- Депонирование ионов Кальция.
- Участие в обмене воды.
- Синтез предшественников стероидных гормонов.
Молекулярная биология – это наука, изучающая роль митохондрий в метаболизме. В них также идет превращение пирувата в ацетил-коэнзим А, бета-окисление жирных кислот.
| Таблица: строение и функции митохондрий (кратко) | ||
|---|---|---|
| Структурные элементы | Строение | Функции |
| Наружная мембрана | Гладкая оболочка, построена из липидов и белков | Отграничивает внутреннее содержимое от цитоплазмы |
| Межмембранное пространство | Находятся ионы водорода, белки, микромолекулы | Создает протонный градиент |
| Внутренняя мембрана | Образует выпячивания – кристы, содержит белковые транспортные системы | Перенос макромолекул, поддержание протонного градиента |
| Матрикс | Место расположения ферментов цикла Кребса, ДНК, РНК, рибосом | Аэробное окисление с высвобождением энергии, превращение пирувата в ацетил-коэнзим А. |
| Рибосомы | Объединённые две субъединицы | Синтез белка |
Сходство митохондрий и хлоропластов
Общие свойства для митохондрий и хлоропластов обусловлены, прежде всего, наличием двойной мембраны.
Признаки сходства также заключаются в способности самостоятельно синтезировать белок. Эти органеллы имеют свое ДНК, РНК, рибосомы.
И митохондрии и хлоропласты могут делиться с помощью перетяжки.
Объединяет их также возможность продуцировать энергию, митохондрии более специализированы в этой функции, но хлоропласты во время фотосинтезирующих процессов тоже образуют молекулы АТФ. Так, растительные клетки имеют меньше митохондрий, чем животные, потому что частично функции за них выполняют хлоропласты.
Опишем кратко сходства и различия:
- Являются двомембранными органеллами;
- внутренняя мембрана образует выпячивания: для митохондрий характерны кристы, для хлоропластов – тиллакоиды;
- обладают собственным геномом;
- способны синтезировать белки и энергию.
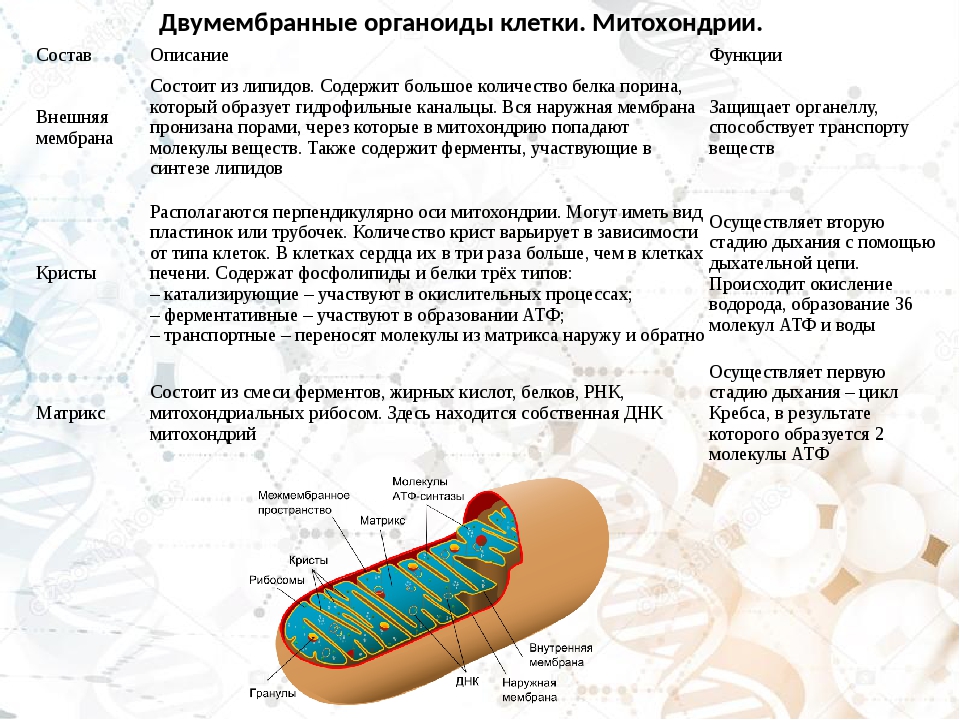
Различаются данные органоиды своими функциями: митохондрии предназначены для синтеза энергии, здесь осуществляется клеточное дыхание, хлоропласты нужны растительным клеткам для фотосинтеза.
Происхождение митохондрий
Межмембранное пространство
Межмембранное пространство представляет собой пространство между наружной и внутренней мембранами митохондрии. Его толщина — 10-20 нм. Так как наружная мембрана митохондрии проницаема для небольших молекул и ионов, их концентрация в периплазматическом пространстве мало отличается от таковой в цитоплазме. Напротив, крупным белкам для транспорта из цитоплазмы в периплазматическое пространство необходимо иметь специфические сигнальные пептиды; поэтому белковые компоненты периплазматического пространства и цитоплазмы различны. Одним из белков, содержащихся в периплазматическом пространстве, является цитохром c — один из компонентов дыхательной цепи митохондрий.
Внутренняя мембрана
Внутренняя мембрана образует многочисленные гребневидные складки — кристы, существенно увеличивающие площадь ее поверхности и, например, в клетках печени составляет около трети всех клеточных мембран. Характерной чертой состава внутренней мембраны митохондрий является присутствие в ней кардиолипина — особого фосфолипида , содержащего сразу четыре жирные кислоты и делающего мембрану абсолютно непроницаемой для протонов . Ещё одна особенность внутренней мембраны митохондрий — очень высокое содержание белков (до 70 % по весу), представленных транспортными белками , ферментами дыхательной цепи , а также крупными АТФ-синтетазными комплексами. Внутренняя мембрана митохондрии в отличие от внешней не имеет специальных отверстий для транспорта мелких молекул и ионов; на ней, на стороне, обращенной к матриксу, располагаются особые молекулы АТФ-синтазы , состоящие из головки, ножки и основания. При прохождении через них протонов происходит синтез АТФ . В основании частиц, заполняя собой всю толщу мембраны, располагаются компоненты дыхательной цепи . Наружная и внутренняя мембраны в некоторых местах соприкасаются, там находится специальный белок-рецептор, способствующий транспорту митохондриальных белков, закодированных в ядре, в матрикс митохондрии.
Характерной чертой состава внутренней мембраны митохондрий является присутствие в ней кардиолипина — особого фосфолипида , содержащего сразу четыре жирные кислоты и делающего мембрану абсолютно непроницаемой для протонов . Ещё одна особенность внутренней мембраны митохондрий — очень высокое содержание белков (до 70 % по весу), представленных транспортными белками , ферментами дыхательной цепи , а также крупными АТФ-синтетазными комплексами. Внутренняя мембрана митохондрии в отличие от внешней не имеет специальных отверстий для транспорта мелких молекул и ионов; на ней, на стороне, обращенной к матриксу, располагаются особые молекулы АТФ-синтазы , состоящие из головки, ножки и основания. При прохождении через них протонов происходит синтез АТФ . В основании частиц, заполняя собой всю толщу мембраны, располагаются компоненты дыхательной цепи . Наружная и внутренняя мембраны в некоторых местах соприкасаются, там находится специальный белок-рецептор, способствующий транспорту митохондриальных белков, закодированных в ядре, в матрикс митохондрии.
Матрикс
Матрикс — ограниченное внутренней мембраной пространство. В матриксе (розовом веществе) митохондрии находятся ферментные системы окисления пирувата , жирных кислот, а также ферменты цикла трикарбоновых кислот (цикла Кребса). Кроме того, здесь же находится митохондриальная ДНК , РНК и собственный белоксинтезирующий аппарат митохондрии.
Митохондриальная ДНК
Находящаяся в матриксе митохондриальная ДНК представляет собой замкнутую кольцевую двуспиральную молекулу , в клетках человека имеющую размер 16569 нуклеотидных пар, что приблизительно в 10 5 раз меньше ДНК, локализованной в ядре . В целом митохондриальная ДНК кодирует 2 рРНК , 22 тРНК и 13 субъединиц ферментов дыхательной цепи , что составляет не более половины обнаруживаемых в ней белков . В частности, под контролем митохондрального генома кодируются семь субъединиц АТФ-синтетазы, три субъединицы цитохромоксидазы и одна субъединица убихинол-цитохром-с -редуктазы. При этом все белки, кроме одного, две рибосомные и шесть тРНК транскрибируются с более тяжёлой (наружной) цепи ДНК, а 14 других тРНК и один белок транскрибируются с более лёгкой (внутренней) цепи.
На этом фоне геном митохондрий растений значительно больше и может достигать 370000 нуклеотидных пар, что примерно в 20 раз больше описанного выше генома митохондрий человека. Количество генов здесь также примерно в 7 раз больше, что сопровождается появлением в митохондриях растений дополнительных путей электронного транспорта, не сопряжённых с синтезом АТФ.
Таким образом, суммарная реакция, катализируемая ферментами дыхательной цепи, состоит в окислении НАДН кислородом с образованием воды. По сути этот процесс заключается в ступенчатом переносе электронов между атомами металлов , присутствующих в простетических группах белковых комплексов дыхательной цепи, где каждый последующий комплекс обладает более высоким сродством к электрону, чем предыдущий. При этом сами электроны передаются по цепи до тех пор, пока не соединятся с молекулярным кислородом, обладающим наибольшим сродством к электронам. Освобождаемая же при этом энергия запасается в виде электрохимического (протонного) градиента по обе стороны внутренней мембраны митохондрий.
Завершающим этапом функционирования митохондрии является генерация АТФ , осуществляемая встроенным во внутреннюю мембрану специальным макромолекулярным комплексом с молекулярной массой 500 кДа. Этот комплекс, называемый АТФ-синтетазой, как раз и катализирует синтез АТФ путём конверсии энергии трансмембранного электрохимического градиента протонов водорода в энергию макроэргической связи молекулы АТФ.
АТФ-синтеза
В структурно-функциональном плане АТФ-синтаза состоит из двух крупных фрагментов, обозначаемых символами F 1 и F 0 . Первый из них (фактор сопряжения F 1) обращён в сторону матрикса митохондрии и заметно выступает из мембраны в виде сферического образования высотой 8 нм и шириной 10 нм. Он состоит из девяти субъединиц, представленных пятью типами белков. Полипептидные цепи трёх субъединиц α и стольких же субъединиц β уложены в похожие по строению белковые глобулы , которые вместе образуют гексамер (αβ) 3 , имеющий вид слегка приплюснутого шара. Подобно плотно уложенным долькам апельсина, последовательно расположенные субъединицы α и β образуют структуру, характеризующуюся осью симметрии третьего порядка с углом поворота 120°. В центре этого гексамера находится субъединица γ, которая образована двумя протяжёнными полипептидными цепями и напоминает слегка деформированный изогнутый стержень длиной около 9 нм. При этом нижняя часть субъединицы γ выступает из шара на 3 нм в сторону мембранного комплекса F 0 . Также внутри гексамера находится минорная субъединица ε, связанная с γ. Последняя (девятая) субъединица обозначается символом δ и расположена на внешней стороне F 1 .
Подобно плотно уложенным долькам апельсина, последовательно расположенные субъединицы α и β образуют структуру, характеризующуюся осью симметрии третьего порядка с углом поворота 120°. В центре этого гексамера находится субъединица γ, которая образована двумя протяжёнными полипептидными цепями и напоминает слегка деформированный изогнутый стержень длиной около 9 нм. При этом нижняя часть субъединицы γ выступает из шара на 3 нм в сторону мембранного комплекса F 0 . Также внутри гексамера находится минорная субъединица ε, связанная с γ. Последняя (девятая) субъединица обозначается символом δ и расположена на внешней стороне F 1 .
Мембранная часть АТФ-синтазы, называемая фактором сопряжения F 0 , представляет собой гидрофобный белковый комплекс, пронизывающий мембрану насквозь и имеющий внутри себя два полуканала для прохождения протонов водорода. Всего в состав комплекса F 0 входит одна белковая субъединица типа а , две копии субъединицы b , а также от 9 до 12 копий мелкой субъединицы c .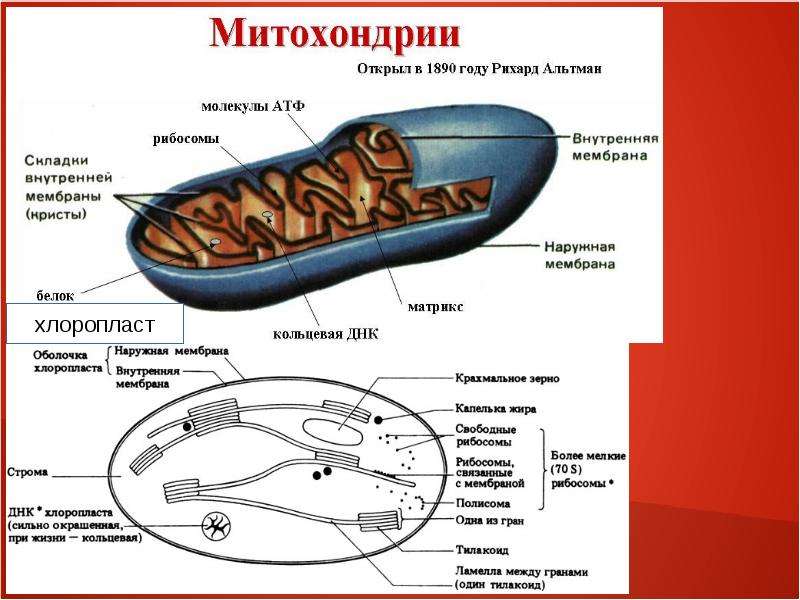 Субъединица а (молекулярная масса 20 кДа) полностью погружена в мембрану, где образует шесть пересекающих её α-спиральных участков. Субъединица b (молекулярная масса 30 кДа) содержит лишь один сравнительно короткий погружённый в мембрану α-спиральный участок, а остальная её часть заметно выступает из мембраны в сторону F 1 и закрепляется за расположенную на её поверхности субъединицу δ. Каждая из 9-12 копий субъединицы c (молекулярная масса 6-11 кДа) представляет собой сравнительно небольшой белок из двух гидрофобных α-спиралей, соединённых друг с другом короткой гидрофильной петлёй, ориентированной в сторону F 1 , а все вместе образуют единый ансамбль, имеющий форму погружённого в мембрану цилиндра. Выступающая из комплекса F 1 в сторону F 0 субъединица γ как раз и погружена внутрь этого цилиндра и достаточно прочно зацеплена за него.
Субъединица а (молекулярная масса 20 кДа) полностью погружена в мембрану, где образует шесть пересекающих её α-спиральных участков. Субъединица b (молекулярная масса 30 кДа) содержит лишь один сравнительно короткий погружённый в мембрану α-спиральный участок, а остальная её часть заметно выступает из мембраны в сторону F 1 и закрепляется за расположенную на её поверхности субъединицу δ. Каждая из 9-12 копий субъединицы c (молекулярная масса 6-11 кДа) представляет собой сравнительно небольшой белок из двух гидрофобных α-спиралей, соединённых друг с другом короткой гидрофильной петлёй, ориентированной в сторону F 1 , а все вместе образуют единый ансамбль, имеющий форму погружённого в мембрану цилиндра. Выступающая из комплекса F 1 в сторону F 0 субъединица γ как раз и погружена внутрь этого цилиндра и достаточно прочно зацеплена за него.
Таким образом, в молекуле АТФ-синтазы можно выделить две группы белковых субъединиц, которые могут быть уподоблены двум деталям мотора: ротору и статору . «Статор» неподвижен относительно мембраны и включает в себя шарообразный гексамер (αβ) 3 , находящуюся на его поверхности и субъединицу δ, а также субъединицы a и b мембранного комплекса F 0 . Подвижный относительно этой конструкции «ротор» состоит из субъединиц γ и ε, которые, заметно выступая из комплекса (αβ) 3 , соединяются с погружённым в мембрану кольцом из субъединиц c .
«Статор» неподвижен относительно мембраны и включает в себя шарообразный гексамер (αβ) 3 , находящуюся на его поверхности и субъединицу δ, а также субъединицы a и b мембранного комплекса F 0 . Подвижный относительно этой конструкции «ротор» состоит из субъединиц γ и ε, которые, заметно выступая из комплекса (αβ) 3 , соединяются с погружённым в мембрану кольцом из субъединиц c .
Способность синтезировать АТФ — свойство единого комплекса F 0 F 1 , сопряжённого с переносом протонов водорода через F 0 к F 1 , в последнем из которых как раз и расположены каталитические центры, осуществляющие преобразование АДФ и фосфата в молекулу АТФ. Движущей же силой для работы АТФ-синтазы является протонный потенциал, создаваемый на внутренней мембране митохондрий в результате работы цепи электронного транспорта.
Сила, приводящая в движение «ротор» АТФ-синтазы, возникает при достижении разности потенциалов между наружной и внутренней сторонами мембраны > 220 мВ и обеспечивается потоком протонов, протекающих через специальный канал в F 0 , расположенный на границе между субъединицами a и c . При этом путь переноса протонов включает в себя следующие структурные элементы:
При этом путь переноса протонов включает в себя следующие структурные элементы:
- Два расположенных несоосно «полуканала», первый из которых обеспечивает поступление протонов из межмембранного пространства к существенно важным функциональным группам F 0 , а другой обеспечивает их выход в матрикс митохондрии;
- Кольцо из субъединиц c , каждая из которых в своей центральной части содержит протонируемую карбоксильную группу, способную присоединять H + из межмембранного пространства и отдавать их через соответствующие протонные каналы. В результате периодических смещений субъединиц с , обусловленных потоком протонов через протонный канал происходит поворот субъединицы γ, погружённой в кольцо из субъединиц с .
Таким образом, каталитическая активность АТФ-синтазы непосредственно связана с вращением её «ротора», при котором поворот субъединицы γ вызывает одновременное изменение конформации всех трёх каталитических субъединиц β, что в конечном счёте и обеспечивает работу фермента. При этом в случае образования АТФ «ротор» крутится по часовой стрелке со скоростью четыре оборота в секунду, а само подобное вращение происходит дискретными скачками по 120°, каждый из которых сопровождается образованием одной молекулы АТФ.
При этом в случае образования АТФ «ротор» крутится по часовой стрелке со скоростью четыре оборота в секунду, а само подобное вращение происходит дискретными скачками по 120°, каждый из которых сопровождается образованием одной молекулы АТФ.
Непосредственная функция синтеза АТФ локализована на β-субъединицах сопрягающего комплекса F 1 . При этом самым первым актом в цепи событий, приводящих к образованию АТФ, является связывание АДФ и фосфата с активным центром свободной β-субъединицы, находящейся в состоянии 1. За счёт энергии внешнего источника (тока протонов) в комплексе F 1 происходят конформационные изменения, в результате которых АДФ и фосфат становятся прочно связанными с каталитическим центром (состояние 2), где становится возможным образование ковалентной связи между ними, ведущей к образованию АТФ. На данной стадии АТФ-синтазы ферменту практически не требуется энергии, которая будет необходима на следующем этапе для освобождения прочно связанной молекулы АТФ из ферментативного центра.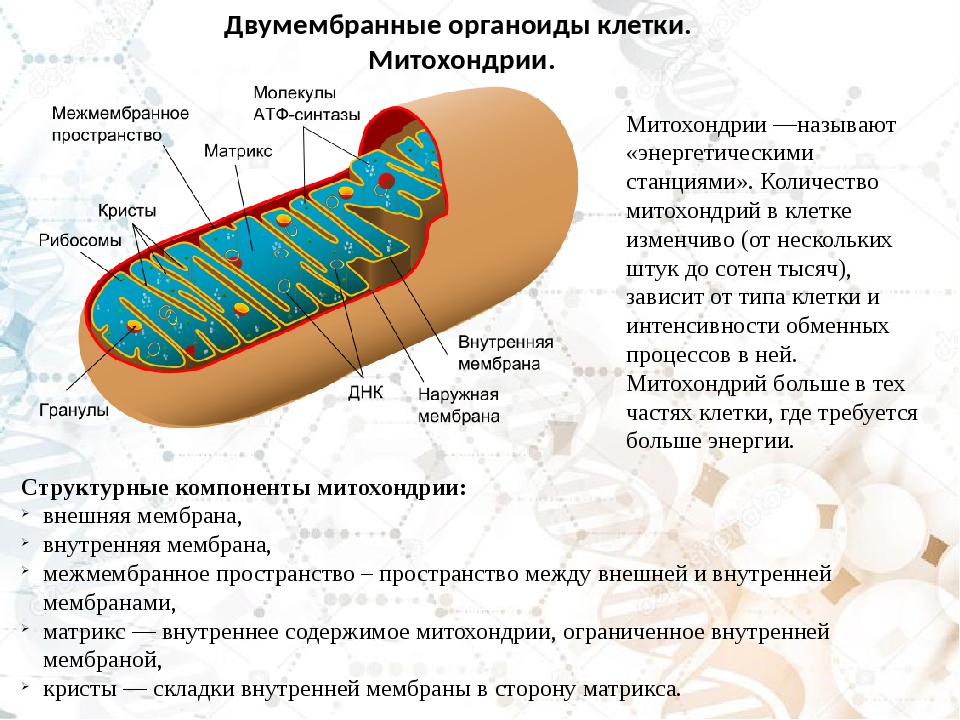 Поэтому следующий этап работы фермента заключается в том, чтобы в результате энергозависимого структурного изменения комплекса F 1 каталитическая β-субъединица, содержащая прочно связанную молекулу АТФ, перешла в состояние 3, в котором связь АТФ с каталитическим центром ослаблена. В результате этого молекула АТФ покидает фермент, а β-субъединица возвращается в исходное состояние 1, благодаря чему обеспечивается цикличность работы фермента.
Поэтому следующий этап работы фермента заключается в том, чтобы в результате энергозависимого структурного изменения комплекса F 1 каталитическая β-субъединица, содержащая прочно связанную молекулу АТФ, перешла в состояние 3, в котором связь АТФ с каталитическим центром ослаблена. В результате этого молекула АТФ покидает фермент, а β-субъединица возвращается в исходное состояние 1, благодаря чему обеспечивается цикличность работы фермента.
Работа АТФ-синтазы связана с механическими движениями её отдельных частей, что позволило отнести этот процесс к особому типу явлений, названных «вращательным катализом». Подобно тому, как электрический ток в обмотке электродвигателя приводит в движение ротор относительно статора, направленный перенос протонов через АТФ-синтетазу вызывает вращение отдельных субъединиц фактора сопряжения F 1 относительно других субъединиц ферментного комплекса, в результате чего это уникальное энергообразующее устройство совершает химическую работу — синтезирует молекулы АТФ.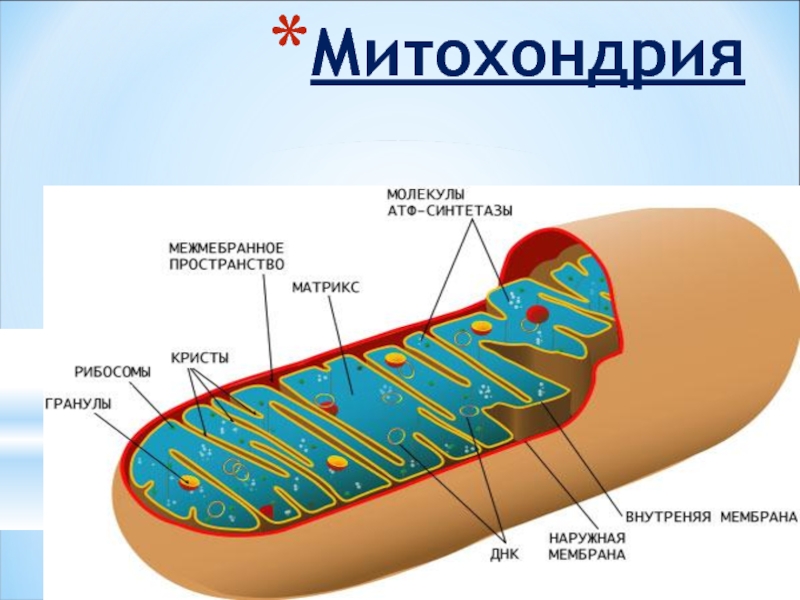 В дальнейшем АТФ поступает в цитоплазму клетки, где расходуется на самые разнообразные энергозависимые процессы. Подобный перенос осуществляется специальным встроенным в мембрану митохондрий ферментом АТФ/АДФ-транслоказой, который обменивает вновь синтезированную АТФ на цитоплазматическую АДФ, что гарантирует сохранность фонда адениловых нуклеотидов внутри митохондрий.
В дальнейшем АТФ поступает в цитоплазму клетки, где расходуется на самые разнообразные энергозависимые процессы. Подобный перенос осуществляется специальным встроенным в мембрану митохондрий ферментом АТФ/АДФ-транслоказой, который обменивает вновь синтезированную АТФ на цитоплазматическую АДФ, что гарантирует сохранность фонда адениловых нуклеотидов внутри митохондрий.
Словарь синонимов
Митохондрия. См. пластосома. (
Митохондрии (МТ) – одно из самых интересных мне направлений исследований. Объединение митохондрий с другой клеткой в ходе эндосимбиоза около 1,6 млрд лет назад стало основной всех многоклеточных эукариотов со сложной структурой. Предположительно митохондрии произошли от клеток, напоминающих α-протеобактерии.
Лучшее обзорное исследование последнего времени по митохондриях – работа Вернера Кулбрандта «Структура и функция митохондриальных белковых комплексов мембраны ». Если вы знаете английский язык и интересуетесь устройством этих органелл, то настоятельно рекомендую к прочтению.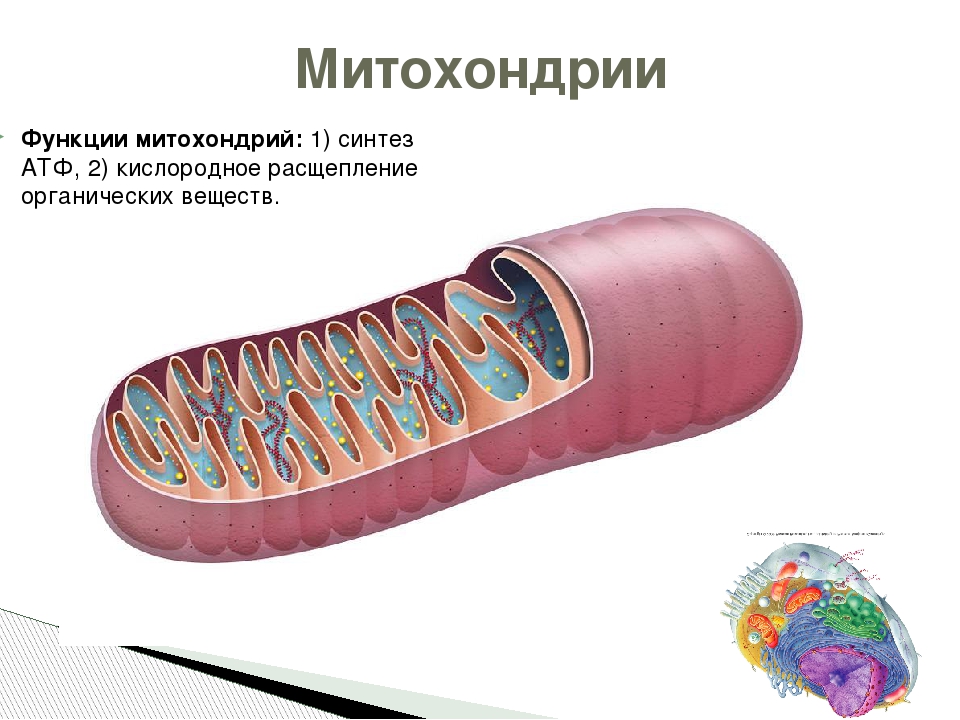 Эта статья так хороша, что может быть смело главой хорошего учебника по молекулярной биологии. Сначала я хотел перевести всю статью, но это бы заняло непростительно много времени и оторвало бы от других дел. Поэтому ограничусь тезисами и картинками. Периодически разбавляя все своими мыслями.
Эта статья так хороша, что может быть смело главой хорошего учебника по молекулярной биологии. Сначала я хотел перевести всю статью, но это бы заняло непростительно много времени и оторвало бы от других дел. Поэтому ограничусь тезисами и картинками. Периодически разбавляя все своими мыслями.
Митохондрия кодирует сама только 13 белков, не смотря на наличие отдельной от клетки ДНК (мтДНК) и всего «производственного» цикла по транскрипции белков. Изолированная митохондрия какое-то время может сохранять композицию и функционировать.
Рисунок 1. Компоненты мембраны митохондриона. Внешняя мембрана отделяет митохондрию от цитоплазмы. Она окружает внутреннюю мембрану, которая отделяет межмембранное пространство от богатого белками центрального матрикса. Внутреннюю мембрану разделяют на внутреннюю пограничную мембрану и кристы. Две эти части непрерывны в местах крепления крист (cristae junction ). Кристы простираются более или менее глубоко в матрикс и являются основным место митохондриального преобразования энергии.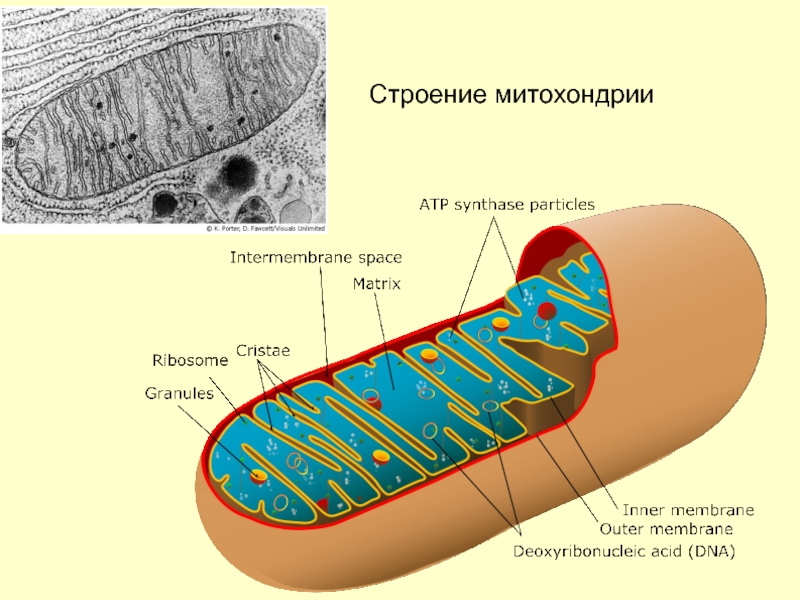 Небольшой протоновый градиент в межмембранном пространстве (pH 7,2-7,4) и матрикс (pH 7,9-8,0) приводят к образованию АТФ АТФ-синтазой в мембранах крист.
Небольшой протоновый градиент в межмембранном пространстве (pH 7,2-7,4) и матрикс (pH 7,9-8,0) приводят к образованию АТФ АТФ-синтазой в мембранах крист.
Внешняя мембрана пористая и позволяет веществам из цитоплазмы проходить через нее. Внутренняя мембрана плотная, для ее пересечения нужны транспортные белки [Гилберт Линг обоснованно не согласен], непрерывность барьера позволяет иметь внутренней мембране электрохимический потенциал в -180 mV. У матрикса довольно большой pH (7,9-8). Еще раз углублюсь в Линга. Щелочной (выше 7) pH способствует более развернутой конформации белков . Высокий pH нарушает водородные и солевые связи, делая поляризованные CO и NH доступными молекулам воды, там самым усиливая дипольный момент всей внутриклеточной воды и связывая ее. В этом ключе наличие мембраны нужно не для «удержания» протоплазмы внутри клетки (это делают сами белки при высоком pH), а для наличия потенциала.
мтДНК находится в нуклеотидах, которых примерно 1000 на клетку. Белковая плотность матрикса довольна высокая (до 500 мг/мл), что близко к кристаллизованным белкам.
Белковая плотность матрикса довольна высокая (до 500 мг/мл), что близко к кристаллизованным белкам.
Внутренняя мембрана образует инвагинации, называемые кристами, которые глубоко проникают в матрикс. Кристы определяют третий «отсек» митохондрий – просвет крист (cristae lumen). Мембраны кристы содержат большинство, если не все, полностью «собранные» комплексы цепи переноса электронов и АТФ-синтазы. Просвет кристы содержит большое количество маленького растворимого белкового переносчика электронов (цитохром с). Митохондриальные кристы, таким образом, основное место биологической конверсии энергии во всех не фотосинтетических эукариотах.
С кристами тоже много всего интересного. Оптические свойства кристы влияют на распространение и генерацию света в тканях. Я даже встречал идеи о том, что поверхность крист подобна (предположение) поверхностям топологических изоляторов (подразумевалась суперпроводимость без диссипации заряда).
Рисунок 2. Мембранные белковые комплексы дыхательной цепи. Комплекс I (NADH / убихинон оксидоредуктаза, синий), Комплекс II (сукцинат дегидрогеназа, розовы), Комплекс III (цитохром С редуктаза, оранжевый), Комплекс IV (цитохром С оксидаза, зеленый) и митохондриальная АТФ синтаза (известная как комплекс V , бежевая) работают вместе во время окислительного фосфорилирования, чтобы клетки могли использовать энергию. Комплексы I , III , IV выкачивают протоны вдоль мембраны кристы, создавая протоновый градиент, стимулирующий синтез АТФ.
Комплекс I (NADH / убихинон оксидоредуктаза, синий), Комплекс II (сукцинат дегидрогеназа, розовы), Комплекс III (цитохром С редуктаза, оранжевый), Комплекс IV (цитохром С оксидаза, зеленый) и митохондриальная АТФ синтаза (известная как комплекс V , бежевая) работают вместе во время окислительного фосфорилирования, чтобы клетки могли использовать энергию. Комплексы I , III , IV выкачивают протоны вдоль мембраны кристы, создавая протоновый градиент, стимулирующий синтез АТФ.
Теперь немного внимания на комплекс II. Вы помните, что жир (кето) делает упор в метаболизме на FADh3 и комплекс II. Они восстанавливают пару CoQ, в какой-то момент окисленного CoQ не хватает для транспортировки электронов на комплекс III и образует обратный поток электронов на комплекс I с образованием супероксида. При долгом HFLC-питании комплекс I будет обратимо разрушен, при этом это нормальная физиологическая оптимизация.
Еще прошу вас заметить, что комплекс II не выкачивает протоны. Что у нас рассеивает протоновый градиент, нарушает фосфорилирование и стимулирует сжигание жиров на тепло? Правильно, стресс холода. Термогенез связан с метаболизмом через комплекс, который не выкачивает протоны, тем самым не давая дополнительных протонов для АТФ-синтазы. Можно только удивляться как замечательно у нас продуман организм.
Крепления крист и MICOSМеста крепления крист (cristae junctions) – маленькие круглы отверстия примерно 25 нм диаметром. В митохондриях всех организмов есть система MICOS (mitochondria contact site and cristae to outer membrane), сборка из пяти мембранных и одного растворимого белков, прикрепляющих кристы к наружной мембране.
В клетках с повышенной потребностью в энергии, такие как скелетные и сердечные мышцы, кристы плотно заполняют большую часть объема митохондрии. В тканях с меньшими потребностями в энергии, таких как печень и почки, кристы находятся не так плотно по отношению друг к другу. Остается больше места в матрице для биосинтетических ферментов.
Остается больше места в матрице для биосинтетических ферментов.
Рисунок 3. Томографический объем митохондрии сердца мыши. А) Трехмерный объем митохондрии сердца мыши, снятый cryo-ET. Наружная мембрана (серая) окутывает внутреннюю мембрану (светло-синяя). Внутренняя мембрана плотно наполнена кристами б) Томографический срез объема. Плотно заполненный матрикс, содержащий большую часть митохондриальных белков, выглядит темным на электронном микроскопе. В то время как межмембранное пространство и просветы крист выглядят светлыми из-за низкой концентрации белков.
Димеры АТФ синтазыМитохондриальная F1-F0 АТФ синтаза является самым заметным белковым комплексом кристы. АТФ синтаза – это древняя наномашина, которая использует электрохимический протновых градиент вокруг внутренней мембраны для создания АФТ посредством вращательного катализа
. Протоны, двигающиеся через F0 комплекс мембраны, вращают ротор из 8 (у млекопитающих) или 10 (у дрожжей) с-узлов. Центральный стебель передает крутящий момент c-ротора каталитической головке F1, где АТФ образуется из АДФ и фосфата через последовательность конформационных изменения. Периферийный стебель предотвращает непродуктивное вращение Головы F1 против комплекса F0.
Центральный стебель передает крутящий момент c-ротора каталитической головке F1, где АТФ образуется из АДФ и фосфата через последовательность конформационных изменения. Периферийный стебель предотвращает непродуктивное вращение Головы F1 против комплекса F0.
Многие годы считалось, что АТФ синтаза случайным образом располагается на внутренней мембране. Но оказалось, что АТФ синтаза располагается двойными рядами . Причем линейные ряды АТФ синтазы – фундаментальный атрибут всех живых митохондрий.
Рисунок 4. Двойные ряды АТФ синтазы у семи разных видов.
Ряды АТФ синтазы располагаются в основном вдоль хребтов крист. Димеры изгибают липидный бислой и как следствие само-организуются в ряды. Когда у митохондрий дрожжей выбивали узлы e и g АФТ синтазы, то штамм рос на 60% медленней диких собратьев, и потенциал мембран их митохондрий был снижен вдвое. У АФТ синтазы прокариотов недостает нескольких узлов, связанных с димерами, ряды димеров не были найдены у бактерий и архей. Кристы и ряды димеров АФТ синтазы, таким образом, являются адаптацией к большим энергетическим потребностям организма.
Кристы и ряды димеров АФТ синтазы, таким образом, являются адаптацией к большим энергетическим потребностям организма.
Рисунок 5. Структура димера АТФ синтазы митохондрии polymella sp. Вид сбоку на V-образный димер АТФ синтазы.
Комплексы и суперкомплексы дыхательной цепиПротоновый градиент вокруг внутренней мембраны создается тремя крупными мембранными комплексами, известными как комплекс I, комплекс III и комплекс IV (см. рисунок 2). Комплекс I кормится электронами из NADH, высвобождаемая при передаче электрона энергия выкачивает четыре протона. Комплекс III получает электрон от восстановленного хинола и передает его носителю электронов (цитохрому с), выкачивая в процессе один протон. Комплекс IV получает электрон из цитохрома с и передает его молекулярному кислороду, выкачивая 4 протона за каждую молекулу кислорода, превращенную в воду. Комплекс II не выкачивает протоны, напрямую передавая электроны хинолу. Как перенос электронов из NADH в хинол связан с транслокацией протонов пока не ясно. Комплекс I – крупнее III и IV вместе взятых.
Комплекс I – крупнее III и IV вместе взятых.
Рисунок 6. Комплекс I митохондрии коровьего сердца. Матриксная часть содержит ряд из восьми железно-серных (Fe-S) кластеров, которые направляют электроны из NADH в хинол на пересечении матрикса и мембраны. Мембранная часть состоит из 78 лопастей, включая выкачивающие протоны молекулы.
Комплексы I, III и IV соединяются в суперкомплексы или респирасомы. У пекарских дрожжей (saccharomyces cerevisiae ) нет комплекса I, их суперкомплексы состоят из III и IV. Роль суперкомплексов пока еще не ясна. Предполагают, что это делает транспорт электронов более эффективным, но прямых доказательств этому пока нет.
Рисунок 7. Суперкомплекс митохондрии коровьего сердца. Обратите внимание на дистанцию между комплексами I и III, который надо проделать хинолу. Стрелки – движения электрона в суперкомплексе.
Основным белком просвета кристы является цитохром с, который переносит электрон из комплекса III в комплекс IV. Если цитохром с высвобождается в цитоплазму клетки, то вызывает апоптоз .
Если цитохром с высвобождается в цитоплазму клетки, то вызывает апоптоз .
Рисунок 8. Ряды димеров АТФ синтазы задают форму кристам. У хребта кристы АФТ синтаза (желтый) образует слив для протонов (красный), протоновые насосы электронной цепи (зеленый) находятся по обоим сторона рядов димеров. Направляя протоны от источника к АТФ синтазе, кристы работают как протоновые направляющие, позволяющие эффективное производство АТФ. Красные стрелки показывают направление потока протонов.
Реорганизация мембраны во время старенияСтарение – фундаментальный и плохо понимаемый процесс всех эукариотов. Исследовали старение митохондрий на грибах Podospora anserina , которые живут всего 18 дней. В нормальной митохондрии кристы проникают глубоко в матрикс. Для этого нужны ряды димеров АТФ синтазы и MICOS комплекс у мест крепления крист. С возрастом кристы начинают все ближе подходить к поверхности мембраны, димеры АФТ синтазы превщаются в мономеры, и все заканчивается высвобождением цитохрома с и клеточной смертью.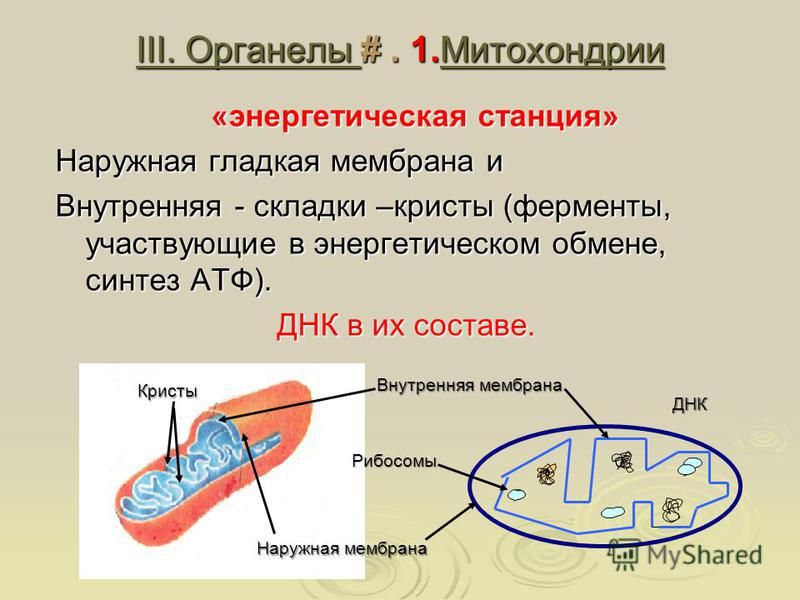
Транспорт электронов создает супероксид в комплексах I и III. Это побочный продукт метаболизма. Одновременно необходимый и смертельно опасный. Во время старения деление (fission) начинает превалировать над сращением (fussion). Это не дает поврежденным митохондриям «спастись» путем сращения и ускоряет неизбежное.
Рисунок 9. Изменения морфологии внутренней мембраны и димеров АТФ синтазы во время старения митохондрии.
Как видите, полей для будущих исследований очень много. Я предполагаю стык физики и биологии, где физики будут пытаются объяснить почему такая структура более энергетически эффективна. Тем более публикации по квантовой биологии
Митохондрии –
преобразователи энергии и её поставщики
для обеспечения клеточных функций –
занимают значительную часть цитоплазмы
клеток и сосредоточены в местах высокого
потребления АТФ (например, в эпителии
канальцев почки они располагаются
вблизи плазматической мембраны
(обеспечение реабсорбции), а в нейронах
– в синапсах (обеспечение электрогенеза
и секреции).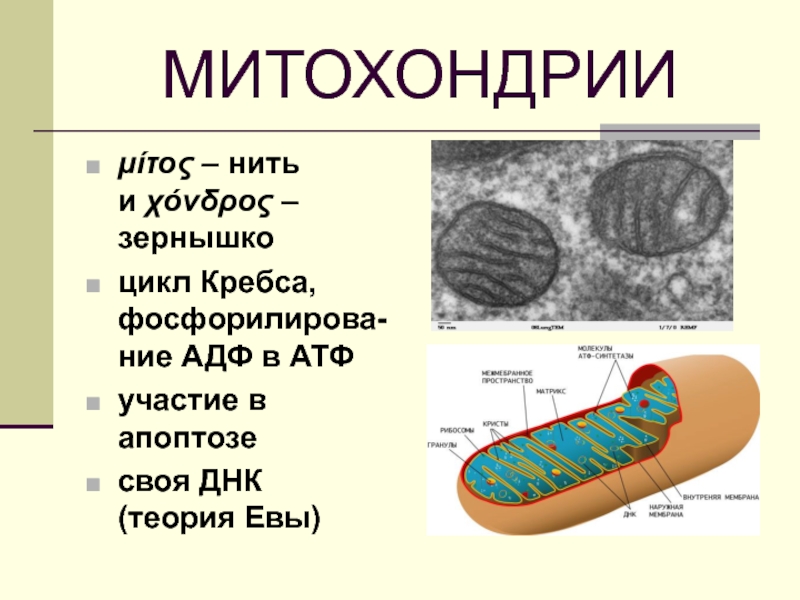 Количество митохондрий в
клетке измеряется сотнями. Митохондрии
имеют собственный геном. Органелла
функционирует в среднем 10 суток,
обновление митохондрий происходит
путем их деления.
Количество митохондрий в
клетке измеряется сотнями. Митохондрии
имеют собственный геном. Органелла
функционирует в среднем 10 суток,
обновление митохондрий происходит
путем их деления.
Морфология митохондрии
Митохондрии чаще имеют форму цилиндра диаметром 0,2-1 мкм и длиной до 7 мкм (в среднем около 2 мкм). У митохондрий две мембраны – наружная и внутренняя; последняя образует кристы. Между наружной и внутренней мембранами находится межмембранное пространство. Внемембранный объем митохондрии – матрикс.
∙ Наружная мембрана проницаема для многих мелких молекул.
∙ Межмембранное пространство. Здесь накапливаются ионы Н + , выкачиваемые из матрикса, что создает протонный градиент концентрации по обе стороны внутренней мембраны.
∙ Внутренняя
мембрана избирательно
проницаема; содержит транспортные
системы для переноса веществ (АТФ, АДФ,
Р 1 ,
пирувата, сукцината, α-кетоглурата,
малата, цитрата, цитидинтрифосфата,
ГТФ, дифосфатов) в обоих направлениях
и комплексы цепи переноса электронов,
связанные с ферментами окислительного
фосфорилирования, а также с
сукцинатдегидрогеназой (СДГ).
∙ Матрикс. В матриксе присутствуют все ферменты цикла Кребса (кроме СДГ), ферменты β-окисления жирных кислот и некоторые ферменты других систем. В матриксе находятся гранулы с Mg 2+ и Ca 2+ .
∙ Цитохимические маркёры митохондрий – цитохромоксидаза и СДГ.
Функции митохондрий
Митохондрии выполняют в клетке множество функций: окисление в цикле Кребса, транспорт электронов, хемиосмотическое сопряжение, фосфорилирование АДФ, сопряжение окисления и фосфорилирования, функцию контроля внутриклеточной концентрации кальция, синтез белков, образование тепла. Велика роль митохондрий в программированной (регулируемой) гибели клеток.
∙ Теплорепродукция. Естественный
механизм разобщения окислительного
фосфорилирования функционирует в
клетках бурого жира. В этих клетках
митохондрии имеют атипичную структуру
(уменьшен их объем, увеличена плотность
матрикса, расширены межмембранные
пространства) – конденсированные
митохондрии. Такие митохондрии могут
усиленно захватывать воду и набухать
в ответ на тироксин, увеличение
концентрации Ca 2+
в цитозоле, при этом усиливается
разобщение окислительного фосфорилирования,
и происходит выделение тепла.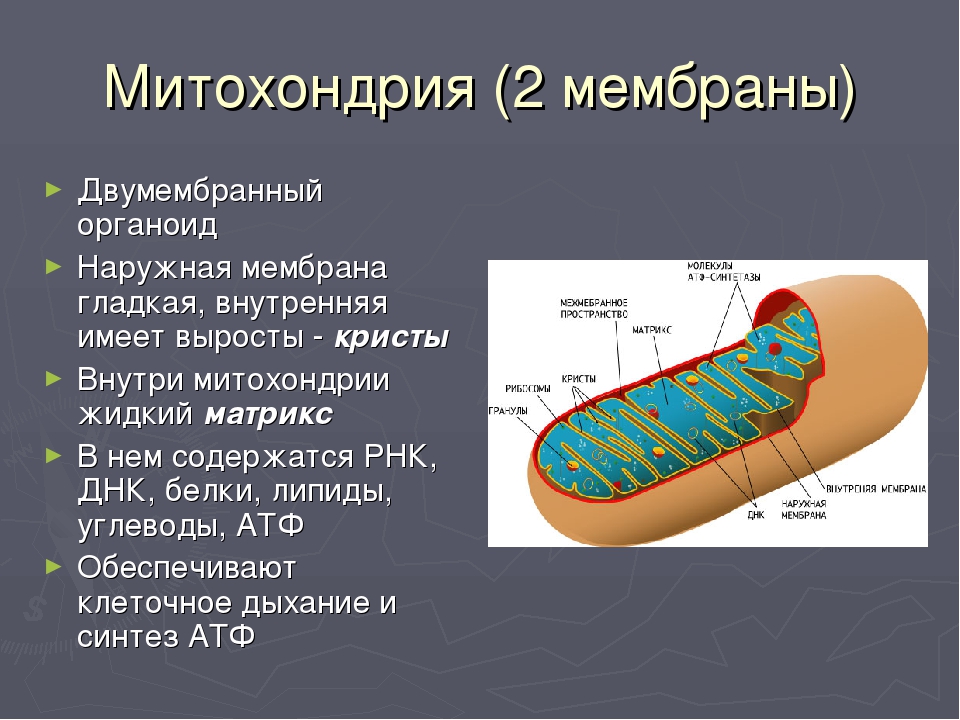 Эти
процессы обеспечивает специальный
разобщающий белок термогенин. Норадреналин
из симпатического отдела вегетативной
нервной системы усиливает экспрессию
разобщающего белка и стимулирует
теплопродукцию.
Эти
процессы обеспечивает специальный
разобщающий белок термогенин. Норадреналин
из симпатического отдела вегетативной
нервной системы усиливает экспрессию
разобщающего белка и стимулирует
теплопродукцию.
∙ Апоптоз. Митохондрии играют важную роль в регулируемой (программированной) гибели клеток – апоптозе, выделяя их в цитозоль факторы, повышающие вероятность гибели клетки. Одним из них является цитохром С – белок, переносящий электроны между белковыми комплексами во внутренней мембране митохондрий. Выделяясь из митохондрий, цитохром С включается в состав апоптосомы, активирующей каспазы (представители семейства киллерных протеаз).
Характерные для подавляющего большинства клеток. Основная функция – это окисление органических соединений и производство АТФ-молекул из освободившейся энергии. Маленькая митохондрия является главной энергетической станцией всего организма.
Происхождение митохондрий
Среди ученых на сегодняшний день весьма популярно мнение, что митохондрия появилась в клетке не самостоятельно в ходе эволюции. Скорее всего, это произошло из-за захвата примитивной клеткой, не способной на тот момент самостоятельно использовать кислород, бактерии, которая умела это и, соответственно, была превосходным источником энергии. Подобный симбиоз оказался успешным и закрепился в последующих поколениях. В пользу этой теории указывает наличие в митохондриях собственной ДНК.
Скорее всего, это произошло из-за захвата примитивной клеткой, не способной на тот момент самостоятельно использовать кислород, бактерии, которая умела это и, соответственно, была превосходным источником энергии. Подобный симбиоз оказался успешным и закрепился в последующих поколениях. В пользу этой теории указывает наличие в митохондриях собственной ДНК.
Как устроены митохондрии
Митохондрия обладает двумя мембранами: наружной и внутренней. Главная функция наружной мембраны – это отделение органоида от цитоплазмы клетки. Она состоит из билипидного слоя и белков, пронизывающих его, через которые и осуществляется транспорт молекул и ионов, необходимых для работы. В то время как гладкая, внутренняя образует многочисленные складки – кристы, которые существенно увеличивают ее площадь. Внутренняя мембрана по большей части состоит из белков, среди которых присутствуют ферменты дыхательной цепи, транспортные белки и крупные АТФ-синтетазные комплексы. Именно в этом месте происходит синтез АТФ.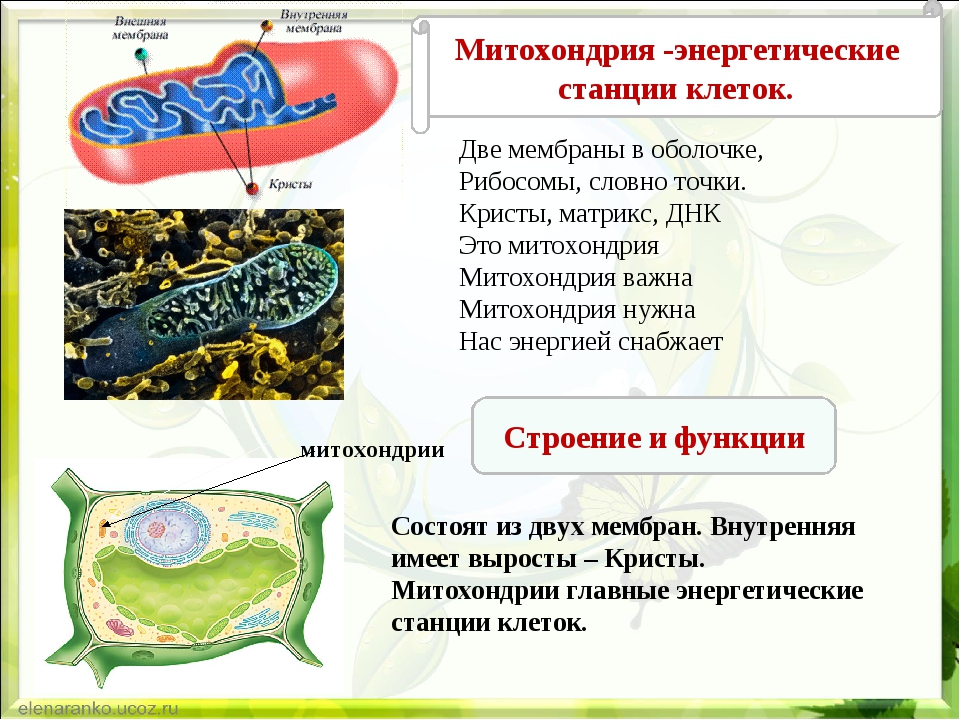 Между наружной и внутренней мембраной находится межмембранное пространство с присущими ему ферментами.
Между наружной и внутренней мембраной находится межмембранное пространство с присущими ему ферментами.
Внутреннее пространство митохондрий называется матрикс. Здесь расположены ферментные системы окисления жирных кислот и пирувата, ферменты цикла Кребса, а также наследственный материал митохондрий – ДНК, РНК и белоксинтезирующий аппарат.
Для чего нужны митохондрии
Главной функцией митохондрий является синтез универсальной формы химической энергии – АТФ. Также они принимают участие в цикле трикарбоновых кислот, превращая пируват и жирные кислоты в ацетил-CoA, а затем окисляя его. В этом органоиде хранится и передается по наследству митохондриальная ДНК, кодирующая воспроизводство тРНК, рРНК и некоторых белков, необходимых для нормального функционирования митохондрий.
Ядро, митохондрии, хлоропласты, ЭПС, аппарат Гольджи. Строение и функции | Биология. Реферат, доклад, сообщение, краткое содержание, конспект, сочинение, ГДЗ, тест, книга
1. Каковы строение и функции ядра?
Каковы строение и функции ядра?
Ядро — наиболее важный компонент эукариотических клеток. Ядро состоит из ядерной оболочки, ядерного сока (кариоплазмы), содержащего хроматин и ядрышки.
Ядерная оболочка образована двумя мембранами (наружной и внутренней) и содержит многочисленные поры, через которые между ядром и цитоплазмой происходит обмен различными веществами.
Ядерный сок — полужидкое вещество, которое находится под ядерной оболочкой. В ядерном соке находятся ядрышки (1-2) и хромосомы. Хромосомы видны только в делящихся клетках. В неделящихся клетках — это нити, каждая из которых состоит из ДНК и белка. Число хромосом постоянно для каждого вида.
Ядрышко — небольшое округлое тельце. Его функция — синтез молекул рРНК и соединение с их с белками, т.е. формирование больших и малых частиц — рибосом. Ядрышки видны только в неделящихся клетках, во время деления они разрушаются.
Хромосомы — (от греч. «хрома» — краска, «сома» — тело) — важнейшая составная часть ядра, образованы ДНК и белками.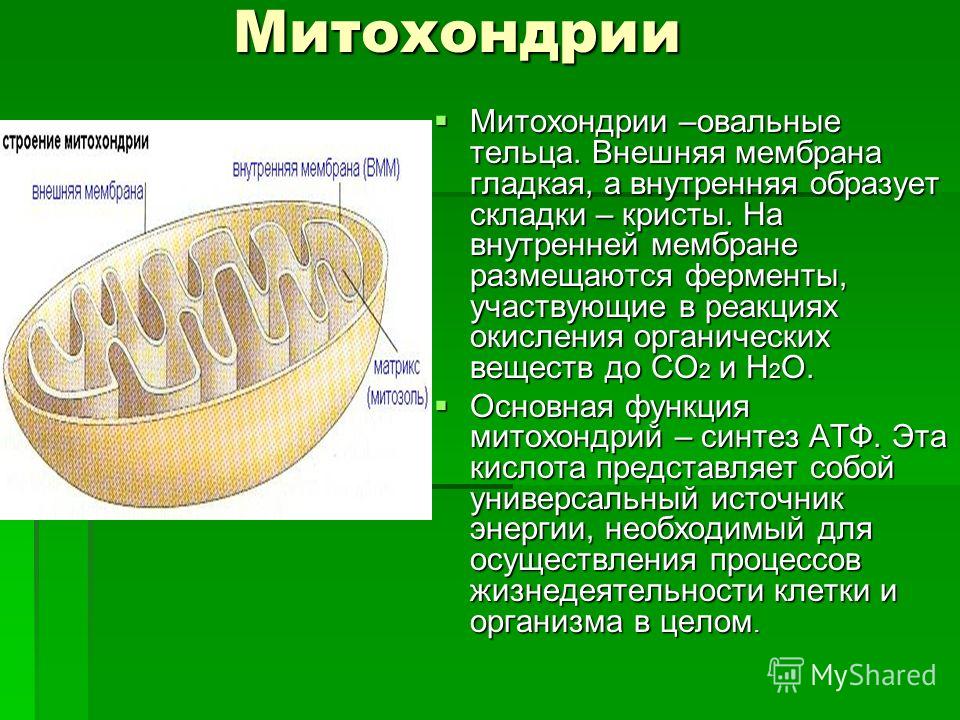 В неделящейся клетке хромосомы в световой микроскоп не видны, они имеют форму длинных, очень тонких нитей. Число, размер и форма хромосом в клетках каждого биологического вида постоянно.
В неделящейся клетке хромосомы в световой микроскоп не видны, они имеют форму длинных, очень тонких нитей. Число, размер и форма хромосом в клетках каждого биологического вида постоянно.
Функции ядра:
а) Осуществляет хранение и реализацию генетической информации, управление процессом биосинтеза белка.
б) Участвует в репликации и распределении наследственной информации между дочерними клетками, а следовательно, в регуляции клеточного деления и процессов развития организма.
в) Образование рибосом из рРНК и белков.
2. Каково строение митохондрий, какие функции они выполняют?
Митохондрии — внутриклеточные органоиды, оболочка которых состоит из двух мембран. Наружная мембрана гладкая, внутренняя образует выросты, называемые кристами. Внутри митохондрий находится полужидкий матрикс, который содержит РНК, ДНК, белки, липиды, углеводы, ферменты, АТФ и др. вещества. В матриксе имеются рибосомы.
Функции митохондрий связаны с тем, что на внутренней мембране имеются дыхательные ферменты и ферменты синтеза АТФ.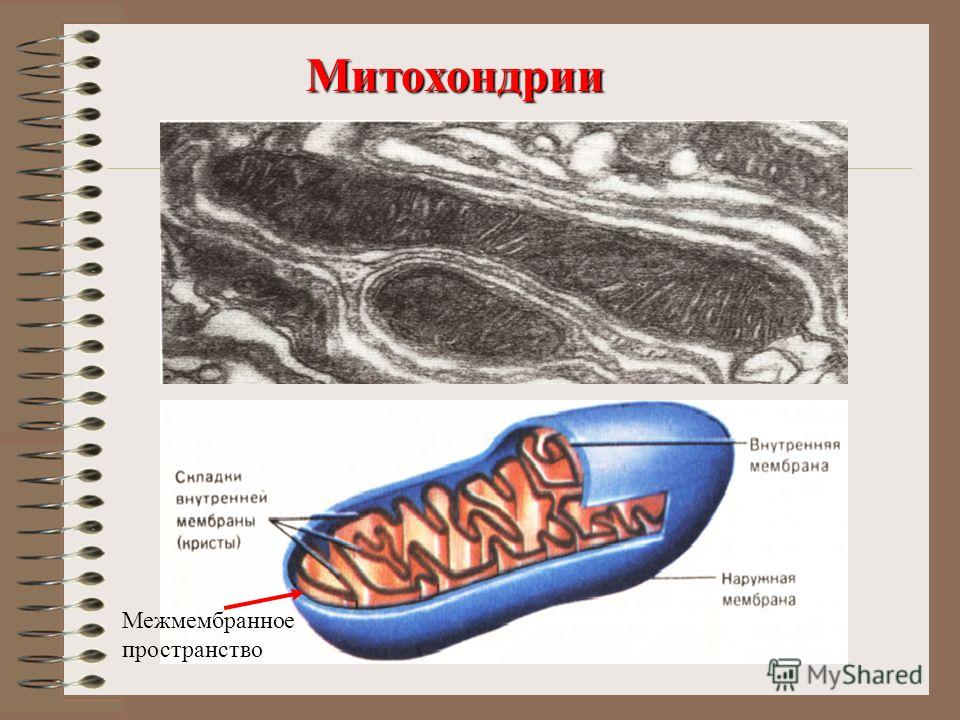 Благодаря этому митохондрии обеспечивают клеточное дыхание и синтез АТФ.
Благодаря этому митохондрии обеспечивают клеточное дыхание и синтез АТФ.
Митохондрии могут сами синтезировать белки, т.к. в них есть собственные ДНК, РНК и рибосомы. Размножаются митохондрии делением надвое.
3. Какое строение имеют хлоропласты, в клетках каких организмов они находятся, какие функции выполняют?
Хлоропласты — органоиды, присущие только растительным клеткам. Имеют две мембраны. Внутренняя мембрана образует выросты — тилакоиды. Тилакоиды формируют стопки — граны, которые объединяются друг с другом внутренней мембраной. В мембранах тилакоидов находится хлорофилл, а в промежутках между гранами в матриксе (строме) хлоропласта находятся рибосомы, РНК, ДНК. Рибосомы хлоропластов синтезируют белки. Основная функция хлоропластов — обеспечение процесса фотосинтеза, в мембранах тилакоидов идет световая фаза, а в строме хлоропластов — темновая фаза фотосинтеза.
4. Каково строение и функции ЭПС?
ЭПС (эндоплазматическая сеть) — это сеть каналов, пронизывающая всю цитоплазму. Она есть во всех клетках кроме клеток бактерий и эритроцитов. Стенки этих каналов представляют собой мембраны, контактирующие со всеми органоидами клетки. ЭПС и органоиды вместе составляют единую внутриклеточную систему, которая осуществляет обмен веществ и энергии в клетке и обеспечивает внутриклеточный транспорт веществ. Различают гладкую и гранулярную (шероховатую) ЭПС. Гранулярная ЭПС состоит из мембранных мешочков (цистерн), покрытых рибосомами, благодаря чему она кажется шероховатой. ЭПС без рибосом — гладкая. На рибосомах синтезируются белки. На мембранах гладкой ЭПС синтезируются липиды и углеводы, которые также поступают внутрь каналов ЭПС.
Она есть во всех клетках кроме клеток бактерий и эритроцитов. Стенки этих каналов представляют собой мембраны, контактирующие со всеми органоидами клетки. ЭПС и органоиды вместе составляют единую внутриклеточную систему, которая осуществляет обмен веществ и энергии в клетке и обеспечивает внутриклеточный транспорт веществ. Различают гладкую и гранулярную (шероховатую) ЭПС. Гранулярная ЭПС состоит из мембранных мешочков (цистерн), покрытых рибосомами, благодаря чему она кажется шероховатой. ЭПС без рибосом — гладкая. На рибосомах синтезируются белки. На мембранах гладкой ЭПС синтезируются липиды и углеводы, которые также поступают внутрь каналов ЭПС.
ЭПС выполняет Функции:
а) Участвует в синтезе органических веществ. Материал с сайта //iEssay.ru
б) Транспортирует синтезированные вещества в аппарат Гольджи.
в) Разделяет клетку на отсеки.
5. Каковы строение и функции аппарата Гольджи?
Комплекс Гольджи — это сложная сеть полостей, трубочек и пузырьков вокруг ядра. Он состоит из трех основных компонентов: группы мембранных полостей, системы трубочек, отходящих от полостей, и пузырьков на концах трубочек.
Он состоит из трех основных компонентов: группы мембранных полостей, системы трубочек, отходящих от полостей, и пузырьков на концах трубочек.
Выполняет следующие функции:
а) В пузырьках накапливаются вещества, которые синтезируются и транспортируются по ЭПС, здесь они подвергаются химическим изменениям.
б) Измененные вещества упаковываются в мембранные пузырьки, которые выделяются клеткой в виде секретов.
в) Часть пузырьков выполняет функцию лизосом, которые участвуют в переваривании частиц, попавших в клетку в результате фаго- и пиноцитоза.
На этой странице материал по темам:- эпс пластиды функцыи
- функция лизосом
- эпс ядро митохондрии
- каково строение и функции эндоплазматической сети
- функции ядрышка кратко
структура, функции, особенности строения в связи с биологическими функциями
Митохондрии — органеллы энергообеспечения
метаболических процесов в клетке.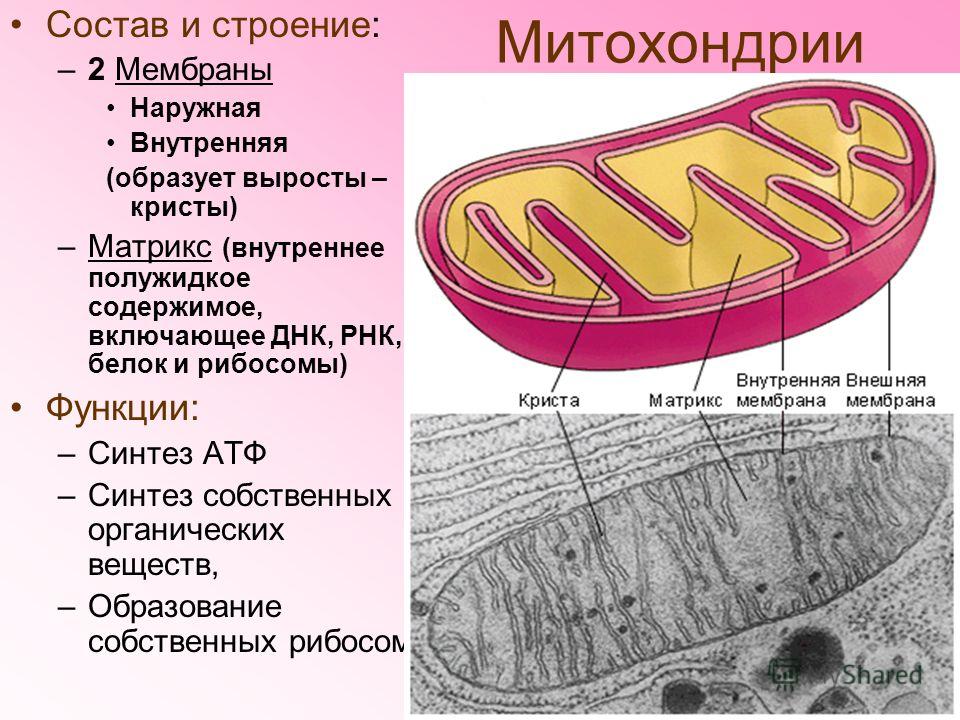 Размеры их варьируют от 0,5 до 5-7 мкм,
количество в клетке составляет от 50 до
1000 и более. В гиалоплазме митохондрии
распределены обычно диффузно, однако
в специализированных клетках сосредоточены
в тех участках, где имеется наибольшая
потребность в энергии. Например, в
мышечных клетках и симпластах большие
количества митохондрий сосредоточены
вдоль рабочих элементов — сократительных
фибрилл. В клетках, функции которых
сопряжены с особо высокими энергозатратами,
митохондрии образуют множественные
контакты, объединяясь в сеть, или
кластеры (кардиомиоциты и симпласты
скелетной мышечной ткани). В клетке
митохондрии выполняют функцию дыхания.
Клеточное дыхание — это последовательность
реакций, с помощью которых клетка
использует энергию связей органических
молекул для синтеза макроэргических
соединений типа АТФ. Образующиеся
внутри митохондрии молекулы АТФ
переносятся наружу, обмениваясь на
молекулы АДФ, находящиеся вне митохондрии.
В живой клетке митохондрии могут
передвигаться с помощью элементов
цитоскелета.
Размеры их варьируют от 0,5 до 5-7 мкм,
количество в клетке составляет от 50 до
1000 и более. В гиалоплазме митохондрии
распределены обычно диффузно, однако
в специализированных клетках сосредоточены
в тех участках, где имеется наибольшая
потребность в энергии. Например, в
мышечных клетках и симпластах большие
количества митохондрий сосредоточены
вдоль рабочих элементов — сократительных
фибрилл. В клетках, функции которых
сопряжены с особо высокими энергозатратами,
митохондрии образуют множественные
контакты, объединяясь в сеть, или
кластеры (кардиомиоциты и симпласты
скелетной мышечной ткани). В клетке
митохондрии выполняют функцию дыхания.
Клеточное дыхание — это последовательность
реакций, с помощью которых клетка
использует энергию связей органических
молекул для синтеза макроэргических
соединений типа АТФ. Образующиеся
внутри митохондрии молекулы АТФ
переносятся наружу, обмениваясь на
молекулы АДФ, находящиеся вне митохондрии.
В живой клетке митохондрии могут
передвигаться с помощью элементов
цитоскелета.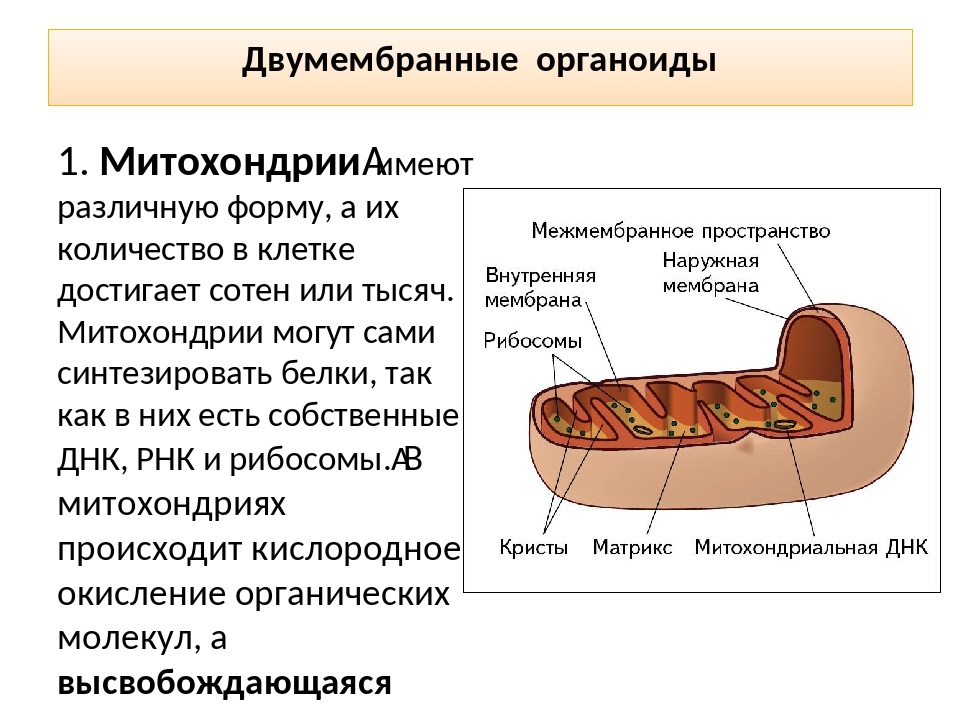 На ультрамикроскопическом
уровне стенка митохондрии состоит из
двух мембран — наружной и внутренней.
Наружная мембрана имеет относительно
ровную поверхность, внутренняя —
образует направленные в центр складки,
или кристы. Между наружной и внутренней
мембранами возникает неширокое (около
15 нм) пространство, которое называется
наружной камерой митохондрии; внутренняя
мембрана ограничивает внутреннюю
камеру. Содержимое наружной и внутренней
камер митохондрии различно, и так же,
как и сами мембраны, существенно
отличается не только по рельефу
поверхности, но и по ряду биохимических
и функциональных признаков. Наружная
мембрана по химическому составу и
свойствам близка к другим внутриклеточным
мембранам и плазмолемме.
На ультрамикроскопическом
уровне стенка митохондрии состоит из
двух мембран — наружной и внутренней.
Наружная мембрана имеет относительно
ровную поверхность, внутренняя —
образует направленные в центр складки,
или кристы. Между наружной и внутренней
мембранами возникает неширокое (около
15 нм) пространство, которое называется
наружной камерой митохондрии; внутренняя
мембрана ограничивает внутреннюю
камеру. Содержимое наружной и внутренней
камер митохондрии различно, и так же,
как и сами мембраны, существенно
отличается не только по рельефу
поверхности, но и по ряду биохимических
и функциональных признаков. Наружная
мембрана по химическому составу и
свойствам близка к другим внутриклеточным
мембранам и плазмолемме.
Ее характеризует
высокая проницаемость, благодаря
наличию гидрофильных белковых каналов.
Эта мембрана имеет в своем составе
рецепторные комплексы, распознающие
и связывающие вещества, поступающие в
митохондрию. Ферментный спектр наружной
мембраны небогат: это ферменты метаболизма
жирных кислот, фосфолипидов, липидов
и др.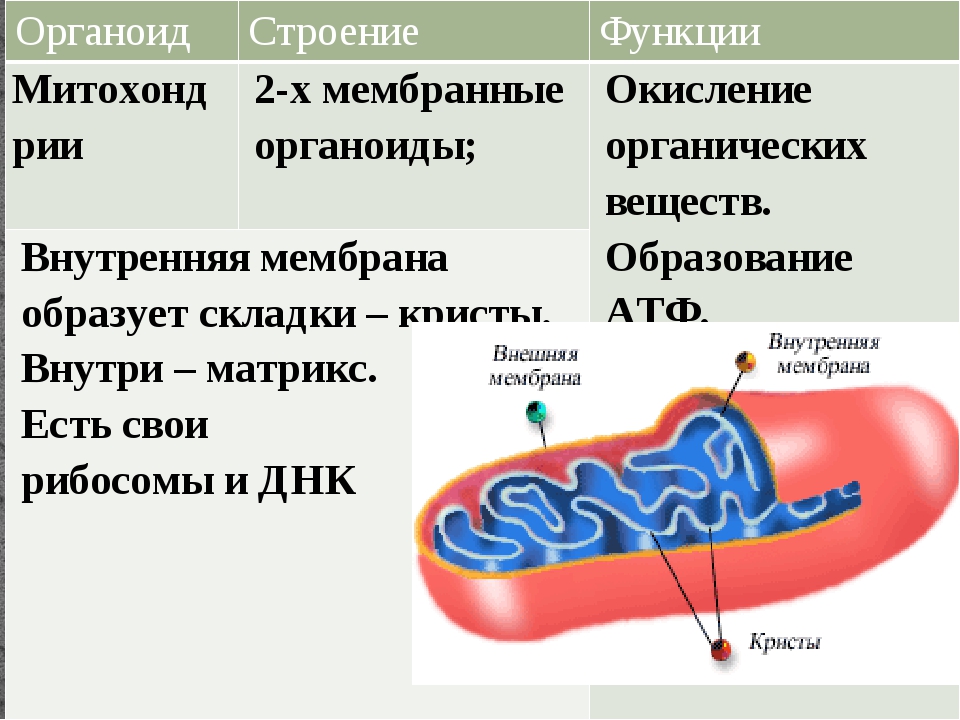 Главной функцией наружной
мембраны митохондрии является
отграничение органеллы от гиалоплазмы
и транспорт необходимых для осуществления
клеточного дыхания субстратов. Внутренняя
мембрана митохондрий в большинстве
клеток тканей различных органов
формирует кристы в виде пластин
(ламеллярные кристы), что значительно
увеличивает площадь поверхности
внутренней мембраны. В последней 20-25 %
всех белковых молекул составляют
ферменты дыхательной цепи и окислительного
фосфорилирования. В эндокринных клетках
надпочечников и половых желез митохондрии
участвуют в синтезе стероидных гормонов.
В этих клетках митохондрии имеют кристы
в виде трубочек (тубул), упорядоченно
расположенных в определенном направлении.
Поэтому кристы митохондрий в
стероидпродуцирующих клетках названных
органов именуются тубулярными. Матрикс
митохондрии, или содержимое внутренней
камеры, представляет собой гелеобразную
структуру, содержащую около 50 % белков.
Осмиофильные тельца, описанные при
электронной микроскопии, — это резервы
кальция.
Главной функцией наружной
мембраны митохондрии является
отграничение органеллы от гиалоплазмы
и транспорт необходимых для осуществления
клеточного дыхания субстратов. Внутренняя
мембрана митохондрий в большинстве
клеток тканей различных органов
формирует кристы в виде пластин
(ламеллярные кристы), что значительно
увеличивает площадь поверхности
внутренней мембраны. В последней 20-25 %
всех белковых молекул составляют
ферменты дыхательной цепи и окислительного
фосфорилирования. В эндокринных клетках
надпочечников и половых желез митохондрии
участвуют в синтезе стероидных гормонов.
В этих клетках митохондрии имеют кристы
в виде трубочек (тубул), упорядоченно
расположенных в определенном направлении.
Поэтому кристы митохондрий в
стероидпродуцирующих клетках названных
органов именуются тубулярными. Матрикс
митохондрии, или содержимое внутренней
камеры, представляет собой гелеобразную
структуру, содержащую около 50 % белков.
Осмиофильные тельца, описанные при
электронной микроскопии, — это резервы
кальция. Матрикс содержит ферменты
цикла лимонной кислоты, катализирующие
окисление жирных кислот, синтез рибосом,
ферменты, участвующие в синтезе РНК и
ДНК. Общее число ферментов превышает
40. Помимо ферментов, матрикс митохондрии
содержит митохондриальную ДНК (митДНК)
и митохондриальные рибосомы. Молекула
митДНК имеет кольцевидную форму.
Возможности внутримитохондриального
белкового синтеза ограничены — здесь
синтезируются транспортные белки
митохондриальных мембран и некоторые
ферментные белки, участвующие в
фосфорилировании АДФ. Все остальные
белки митохондрии кодируются ядерной
ДНК, и их синтез осуществляется в
гиалоплазме, и в дальнейшем они
транспортируются в митохондрию.
Жизненный цикл митохондрий в клетке
короткий, поэтому природа наделила их
двойственной системой воспроизводства
— помимо деления материнской митохондрии,
возможно образование нескольких
дочерних органелл путем почкования.
Матрикс содержит ферменты
цикла лимонной кислоты, катализирующие
окисление жирных кислот, синтез рибосом,
ферменты, участвующие в синтезе РНК и
ДНК. Общее число ферментов превышает
40. Помимо ферментов, матрикс митохондрии
содержит митохондриальную ДНК (митДНК)
и митохондриальные рибосомы. Молекула
митДНК имеет кольцевидную форму.
Возможности внутримитохондриального
белкового синтеза ограничены — здесь
синтезируются транспортные белки
митохондриальных мембран и некоторые
ферментные белки, участвующие в
фосфорилировании АДФ. Все остальные
белки митохондрии кодируются ядерной
ДНК, и их синтез осуществляется в
гиалоплазме, и в дальнейшем они
транспортируются в митохондрию.
Жизненный цикл митохондрий в клетке
короткий, поэтому природа наделила их
двойственной системой воспроизводства
— помимо деления материнской митохондрии,
возможно образование нескольких
дочерних органелл путем почкования.
Строение и функции митохондрий представляют собой довольно сложный вопрос. Наличие органеллы характерно почти для всех ядерных организмов – как для автотрофов (растений, способных к фотосинтезу), так и для гетеротрофов, которыми являются почти все животные, некоторые растения и грибы.
Наличие органеллы характерно почти для всех ядерных организмов – как для автотрофов (растений, способных к фотосинтезу), так и для гетеротрофов, которыми являются почти все животные, некоторые растения и грибы.
Главное предназначение митохондрий – окисление органических веществ и последующее использование освободившейся в результате этого процесса энергии. По этой причине органеллы имеют также и второе (неофициальное) название – энергетические станции клетки. Иногда их называют «пластидами катаболизма».
Что такое митохондрии
Термин имеет греческое происхождение. В переводе это слово означает «нить» (mitos), «зернышко» (chondrion). Митохондрии являются постоянными органоидами, которые имеют огромное значение для нормального функционирования клеток и делают возможным существование всего организма в целом.
«Станции» имеют специфическую внутреннюю структуру, которая изменяется в зависимости от функционального состояния митохондрии. Их форма может быть двух видов – овальная или продолговатая. Последняя нередко имеет ветвящийся вид. Число органоидов в одной клетке колеблется от 150 до 1500.
Последняя нередко имеет ветвящийся вид. Число органоидов в одной клетке колеблется от 150 до 1500.
Особый случай – половые клетки. В сперматозоидах присутствует всего лишь одна спиральная органелла, в то время как женских гаметах содержится в сотни тысяч больше митохондрий. В клетке органоиды не зафиксированы в одном месте, а могут передвигаться по цитоплазме, совмещаться друг с другом. Их размер составляет 0,5 мкм, длина может достигать 60 мкм, в то время как минимальный показатель – 7 мкм.
Определить размер одной «энергетической станции» – непростая задача. Дело в том, что при рассмотрении в электронный микроскоп на срез попадает только часть органеллы. Случается так, что спиральная митохондрия имеет несколько сечений, которые можно принять за отдельные, самостоятельные структуры.
Только объемное изображение позволит выяснить точное клеточное строение и понять, идет речь о 2-5 отдельных органоидах или же об одной, имеющей сложную форму митохондрии.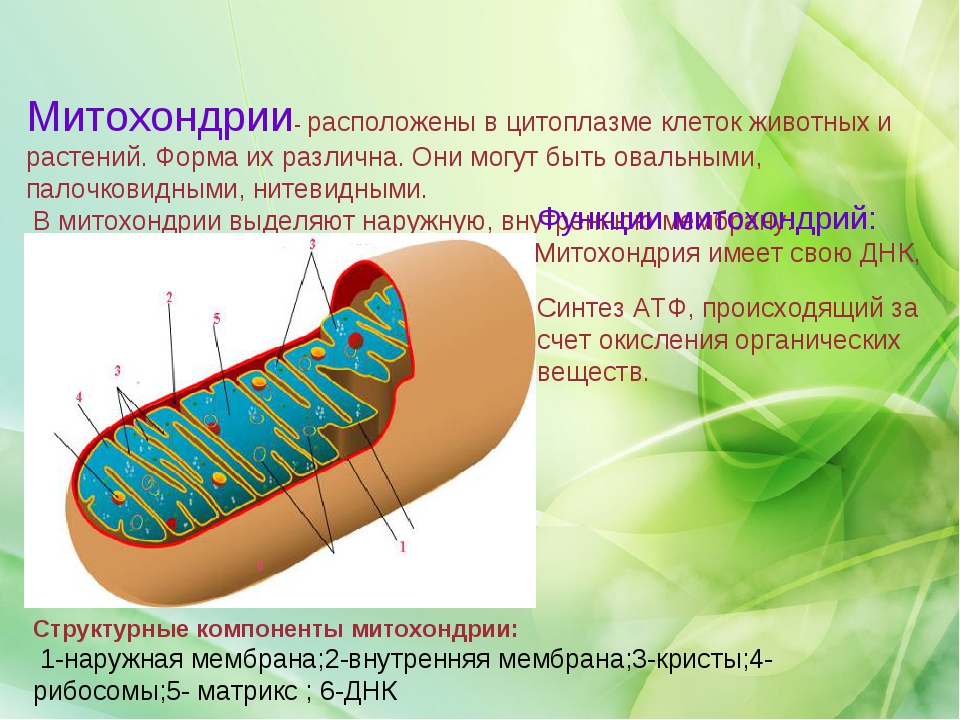
Особенности строения
Оболочка митохондрии состоит из двух слоев: наружного и внутреннего. Последний включает в себя различные выросты и складки, которые имеют листовидную и трубчатую форму.
Каждая мембрана имеет особенный химический состав, определенное количество тех или иных ферментов и конкретное предназначение. Наружную оболочку от внутренней отделяет межмембранное пространство толщиной 10-20 нм.
Весьма наглядно выглядит строение органеллы на рисунке с подписями.
Схема строения митохондрии
Посмотрев на схему строения, можно сделать следующее описание. Вязкое пространство внутри митохондрии называется матриксом. Его состав создает благоприятную среду для протекания в ней необходимых химических процессов. В его составе присутствуют микроскопические гранулы, которые содействуют реакциям и биохимическим процессам (например, накапливают ионы гликогена и других веществ).
В матриксе находятся ДНК, коферменты, рибосомы , т-РНК, неорганические ионы. На поверхности внутреннего слоя оболочки располагаются АТФ-синтаза и цитохромы. Ферменты способствуют таким процессам, как цикл Кребса (ЦТК), окислительное фосфорилирование и т. д.
На поверхности внутреннего слоя оболочки располагаются АТФ-синтаза и цитохромы. Ферменты способствуют таким процессам, как цикл Кребса (ЦТК), окислительное фосфорилирование и т. д.
Таким образом, главная задача органоида выполняется как матриксом, так и внутренней стороной оболочки.
Функции митохондрий
Предназначение «энергетических станций» можно охарактеризовать двумя основными задачами:
- выработка энергии: в них осуществляются окислительные процессы с последующим выделением молекул АТФ;
- хранение генетической информации;
- участие в синтезе гормонов, аминокислот и других структур.
Процесс окисления и выработки энергии проходят в несколько стадий:
Схематичный рисунок синтеза АТФ
Стоит отметить: в результате цикла Кребса (цикл лимонной кислоты) не образуются молекулы АТФ, происходит окисление молекул и выделение углекислого газа. Это промежуточный этап между гликолизом и электронтранспортной цепью.
Таблица «Функции и строение митохондрий»
От чего зависит число митохондрий в клетке
Превалирующее число органоидов скапливается рядом с теми участками клетки, где возникает необходимость в энергетических ресурсах. В частности, большое количество органелл собирается в зоне нахождения миофибрилл, которые являются частью мышечных клеток, обеспечивающих их сокращение.
В мужских половых клетках структуры локализуются вокруг оси жгутика – предполагается, что потребность в АТФ обусловлена постоянным движением хвоста гаметы. Точно так же выглядит расположение митохондрий у простейших, которые для передвижения используют специальные реснички – органеллы скапливаются под мембраной у их основания.
Что касается нервных клеток, то локализация митохондрий наблюдается вблизи синапсов, через которые передаются сигналы нервной системы. В клетках, синтезирующих белки, органеллы скапливаются в зонах эргастоплазмы – они поставляют энергию, которая обеспечивает данный процесс.
Кто открыл митохондрии
Свое название клеточная структура обрела в 1897-1898 годах благодаря К. Бренду. Связь процессов клеточного дыхания с митохондриями сумел доказать Отто Вагбург в 1920 году.
Заключение
Митохондрии являются важнейшей составляющей живой клетки, выступая в роли энергетической станции, которая производит молекулы АТФ, обеспечивая тем самым процессы клеточной жизнедеятельности.
Работа митохондрий основана на окислении органических соединений, в результате чего происходит генерация энергетического потенциала.
Митохондрии имеются во всех эукариотических клетках. Эти органеллы — главное место аэробной дыхательной активности клетки. Впервые митохондрии были обнаружены в виде гранул в мышечных клетках в 1850 г.
Число митохондрий в клетке очень непостоянно; оно зависит от вида организма и от природы клетки. В клетках, в которых потребность в энергии велика, содержится много митохондрий (водной печеночной клетке, например, их может быть около 1000).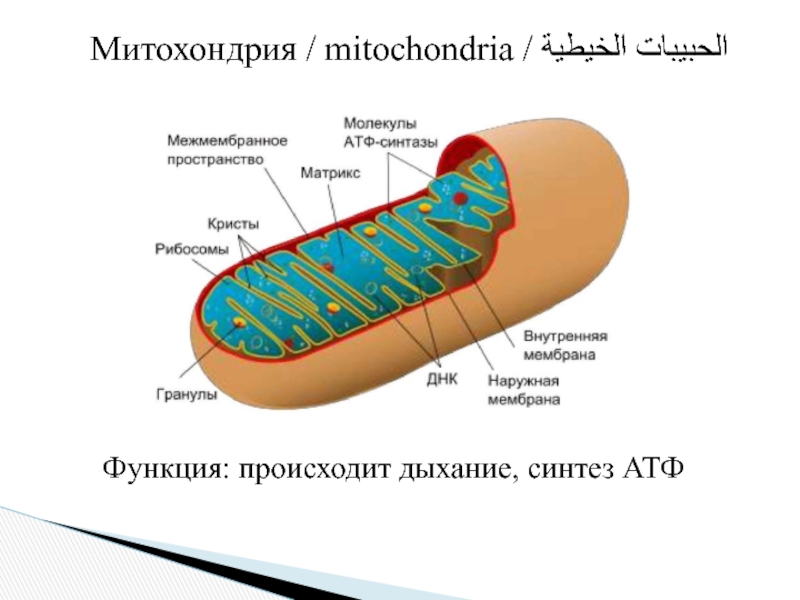 В менее активных клетках митохондрий гораздо меньше. Чрезвычайно сильно варьируют также размеры и форма митохондрий. Митохондрии могут быть спиральными, округлыми, вытянутыми, чашевидными и даже разветвленными: в более активных клетках они обычно крупнее. Длина митохондрий колеблется в пределах 1,5-10 мкм, а ширина — в пределах 0,25-1,00 мкм, но их диаметр не превышает 1 мкм.
В менее активных клетках митохондрий гораздо меньше. Чрезвычайно сильно варьируют также размеры и форма митохондрий. Митохондрии могут быть спиральными, округлыми, вытянутыми, чашевидными и даже разветвленными: в более активных клетках они обычно крупнее. Длина митохондрий колеблется в пределах 1,5-10 мкм, а ширина — в пределах 0,25-1,00 мкм, но их диаметр не превышает 1 мкм.
Митохондрии способны изменять свою форму, а некоторые могут также перемещаться в особо активные участки клетки. Такое перемещение позволяет клетке сосредоточить большое число митохондрий в тех местах, где выше потребность в АТФ. В других случаях положение митохондрий более постоянно (как, например, в летательных мышцах насекомых).
Строение митохондрий
Митохондрии выделяют из клеток в виде чистой фракции с помощью гомогенизатора и ультрацентрифуги, как описано в статье. После этого их можно исследовать в электронном микроскопе, используя для этого различные методики, например изготовление срезов или негативный контраст,. ..
..
Каждая митохондрия окружена оболочкой, состоящей из двух мембран. Наружную мембрану отделяет от внутренней небольшое расстояние — внутримембранное пространство. Внутренняя мембрана образует многочисленные гребневидные складки, так называемые кристы. Кристы существенно увеличивают поверхность внутренней мембраны, обеспечивая место для размещения компонентов дыхательной цепи. Через внутреннюю митохондри-альную мембрану осуществляется активный транспорт АДФ и АТФ. Метод негативного контрастирования, при котором окрашенными оказываются не сами структуры, а пространство вокруг них, позволил выяоить присутствие особых «элементарных частиц» на той стороне внутренней митохондриальной мембраны, которая обращена к матриксу. Каждая такая частица состоит из головки, ножки и основания.
Хотя микрофотографии свидетельствуют, казалось бы, о том, что элементарные частицы выступают из мембраны в матрикс, считается, что это артефакт, обусловленный самой процедурой приготовления препарата, и что в действительности они полностью погружены в мембрану. Головки частиц ответственны за синтез АТФ; в них находится фермент АТФаза, обеспечивающий сопряжение фосфорилирования АДФ с реакциями в дыхательной цепи. В основании частиц, заполняя собой всю толщу мембраны, располагаются компоненты самой дыхательной цепи. В митохондриальном матриксе содержится большая часть ферментов, участвующих в цикле Кребса, и протекает окисление жирных кислот. Здесь же находятся митохондриальные ДНК, РНК и 70S-рибосомы.
Головки частиц ответственны за синтез АТФ; в них находится фермент АТФаза, обеспечивающий сопряжение фосфорилирования АДФ с реакциями в дыхательной цепи. В основании частиц, заполняя собой всю толщу мембраны, располагаются компоненты самой дыхательной цепи. В митохондриальном матриксе содержится большая часть ферментов, участвующих в цикле Кребса, и протекает окисление жирных кислот. Здесь же находятся митохондриальные ДНК, РНК и 70S-рибосомы.
От д-ра Меркола
Митохондрии: вы можете не знать, что это такое, но они жизненно важны для вашего здоровья. Доктор наук Ронда Патрик – биомедик, которая изучила взаимодействие митохондриального метаболизма, аномального метаболизма и рака.
Часть ее работы предполагает выявление ранних биомаркеров заболевания. Например, повреждение ДНК – это ранний биомаркер рака. Затем она пытается определить, какие питательные микроэлементы помогают восстановить это повреждение ДНК.
Она также исследовала митохондриальную функцию и метаболизм, которыми и я увлекаюсь с недавних пор.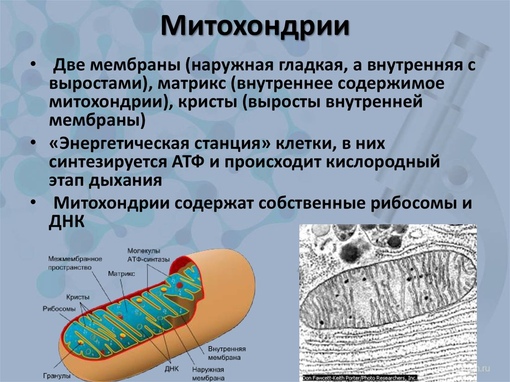 Если, прослушав это интервью, вы захотите узнать об этом побольше, рекомендую начать с книги д-ра Ли Ноу «Жизнь — эпическая история наших митохондрий».
Если, прослушав это интервью, вы захотите узнать об этом побольше, рекомендую начать с книги д-ра Ли Ноу «Жизнь — эпическая история наших митохондрий».
Митохондрии обладают огромным влиянием на здоровье, особенно на рак, и я начинаю верить, что оптимизация митохондриального метаболизма может лежать в основе эффективного лечения рака.
Важность оптимизации митохондриального метаболизма
Митохондрии представляют собой крошечные органеллы, которые, как первоначально считалось, мы унаследовали от бактерий. В красных кровяных тельцах и клетках кожи их почти нет, зато в зародышевых клетках их по 100 000, но в большинстве клеток их от одной до 2 000. Они – главный источник энергии для вашего организма.
Чтобы органы могли функционировать должным образом, им нужна энергия, и эта энергия вырабатывается митохондриями.
Поскольку митохондриальная функция лежит в основе всего, что происходит в организме, то оптимизация митохондриальной функции, и предотвращение нарушения функции митохондрий путем получения всех необходимых питательных веществ и прекурсоров, необходимых митохондриям, чрезвычайно важна для здоровья и профилактики заболеваний.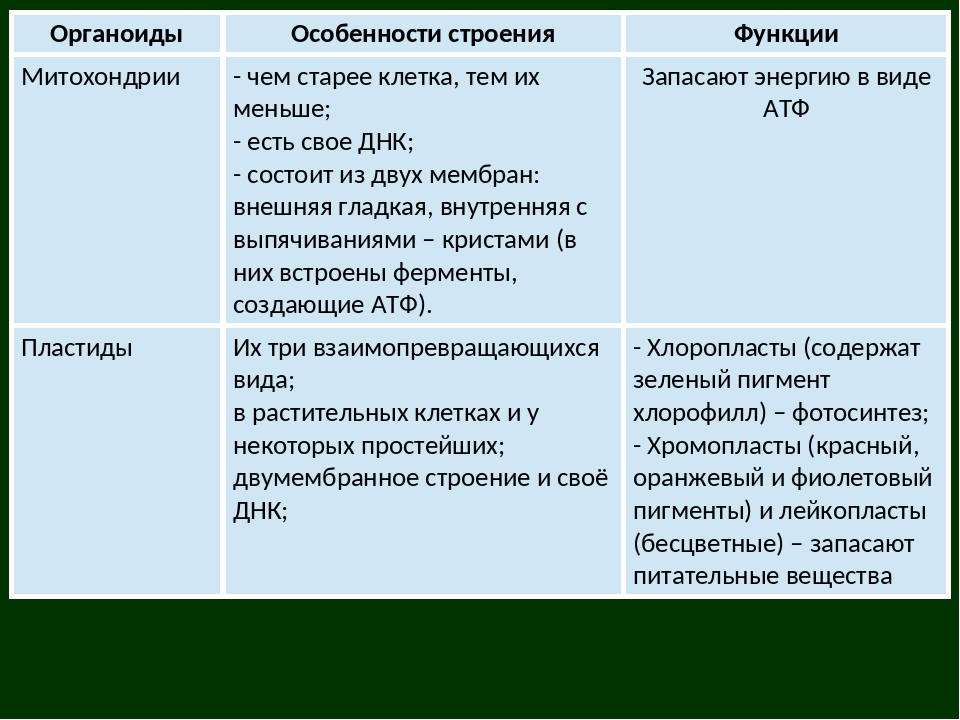
Так, одной из универсальных характеристик раковых клеток является серьезное нарушение функции митохондрий, при котором радикально снижено количество функциональных митохондрий.
Д-р Отто Варбург был врачом с научной степенью по химии и тесно дружил с Альбертом Эйнштейном. Большинство экспертов признают Варбурга величайшим биохимиком 20-го века.
В 1931 году он получил Нобелевскую премию – он открыл, что в качестве источника производства энергии раковые клетки используют глюкозу. Это назвали «эффектом Варбурга» но, к сожалению, это явление и по сей день игнорируется почти всеми.
Я убежден, что кетогенная диета, которая радикально улучшает здоровье митохондрий, может помочь при большинстве видов рака, особенно в сочетании с поглотителем продуктов брожения глюкозы, таким как 3-бромопируват.
Как митохондрии вырабатывают энергию
Чтобы производить энергию, митохондриям нужен кислород из воздуха, которым вы дышите, и жира и глюкоза из пищи, которую вы едите.
Эти два процесса — дыхания и приема пищи – соединяются друг с другом в процессе, который называется окислительное фосфорилирование. Именно он используется митохондриями для производства энергии в виде АТФ.
Митохондрии обладают рядом электронных транспортных цепочек, по которым они передают электроны из восстановленной формы съедаемой вами пищи, чтобы объединить их с кислородом из воздуха, которым вы дышите, и в конечном счете, образовать воду.
Этот процесс приводит протоны через митохондриальную мембрану, подзаряжая АТФ (аденозинтрифосфат) из АДФ (аденозина дифосфат). АТФ переносит энергию по всему организму
Но в ходе этого процесса образуются побочные продукты, такие как активные формы кислорода (АФК), которые повреждают клетки и митохондриальную ДНК, перенося их затем в ДНК ядра.
Таким образом, происходит компромисс. Вырабатывая энергию, организм стареет из-за возникающих в процессе разрушительных аспектов АФК. Скорость старения организма в значительной степени зависит от того, насколько хорошо функционируют митохондрии, и объема повреждений, который можно компенсировать с помощью оптимизации диеты.
Роль митохондрий при раковых заболеваниях
Когда появляются раковые клетки, активные формы кислорода, полученные в качестве побочного продукта производства АТФ, посылают сигнал, запускающий процесс клеточного самоубийства, также известный как апоптоз.
Поскольку клетки рака образуются каждый день, это хорошо. Убивая поврежденные клетки, организм избавляется от них и заменяет их здоровыми.
Раковые клетки, однако, устойчивы к этому протоколу самоубийства – у них против него встроена защита, как объяснил д-р Варбург и, впоследствии, Томас Сейфрид, который глубоко исследовал рак как заболевание обмена веществ.
Как поясняет Патрик:
«Одним из механизмов действия химиотерапевтических препаратов является образование активных форм кислорода. Они создают повреждения, и этого достаточно, чтобы подтолкнуть раковую клетку к смерти.
Думаю, причина этого в том, что раковая клетка, которая не использует свои митохондрии, то есть, больше не производит активные формы кислорода, и вдруг вы ее заставляете пользоваться митохондриями, и получается всплеск активных форм кислорода (ведь именно это делают митохондрии), и — бум, смерть, потому что раковая клетка уже готова к этой смерти.
Она готова умереть».
Почему полезно не есть по вечерам
Уже довольно долгое время я – поклонник чередующегося голодания по целому ряду причин, разумеется, из соображений долголетия и здоровья, а также потому, что оно, как представляется, обеспечивает мощную профилактику рака и благотворное влияние, как от лечения. А механизм этого связан с эффектом, который голодание оказывает на митохондрии.
Как уже упоминалось, основной побочный эффект переноса электронов, в котором участвуют митохондрии, состоит в том, что некоторые утекают из цепи переноса электронов и вступают в реакцию с кислородом, образуя свободные радикалы супероксида.
Анион супероксида (результат уменьшения кислорода на один электрон), является предшественником большинства активных форм кислорода и медиатором окислительных цепных реакций. Свободные радикалы кислорода атакуют липиды клеточных мембран, белковых рецепторов, ферментов и ДНК, что может преждевременно убивать митохондрии.
Некоторые свободные радикалы, вообще-то, даже полезные, необходимые организму для регулирования клеточных функций, но при избыточном образовании свободных радикалов возникают проблемы. К сожалению, именно поэтому у большинства населения развивается большинство заболеваний, особенно рак. Решить эту проблему можно двумя способами:
- Увеличить антиоксиданты
- Уменьшить выработку митохондриальных свободных радикалов
По моему мнению, одной из наиболее эффективных стратегий снижения митохондриальных свободных радикалов является ограничение количества топлива, которым вы заправляете организм. Это совсем непротиворечащее положение, ведь ограничение калорий последовательно демонстрирует много терапевтических преимуществ. Это одна из причин эффективности чередующегося голодания, поскольку оно ограничивает период времени, в который принимается пища, что автоматически уменьшает количество калорий.
Это особенно эффективно, если не есть за несколько часов до сна, потому что это – самое метаболически низкое состояние.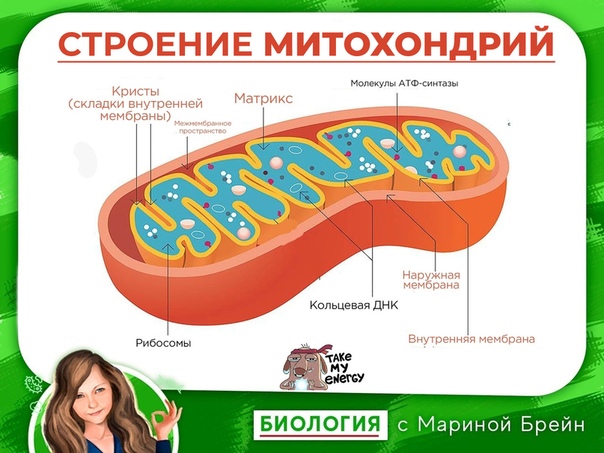
Возможно, неспециалистам все это покажется слишком сложным, но следует понять одно: поскольку во время сна организм использует наименьшее количество калорий, то следует избегать еды перед сном, ведь избыточное количество топлива в это время приведет к образованию избыточного количества свободных радикалов, которые разрушают ткани, ускоряют старение и способствуют возникновению хронических заболеваний.
Как еще голодание помогает здоровью функции митохондрий
Патрик также отмечает, что частично механизм эффективности голодания объясняется тем, что энергию организм вынужден получать из липидов и запасов жира, а это означает, что клетки вынуждены использовать свои митохондрии.
Митохондрии – это единственный механизм, с помощью которых организм может создавать энергию из жира. Таким образом, голодание помогает активировать митохондрии.
Она также считает, что это играет огромную роль в механизме, с помощью которого чередующееся голодание и кетогенная диета убивают раковые клетки, и объясняет, почему некоторые препараты, активирующие митохондрии, способны убивать раковые клетки.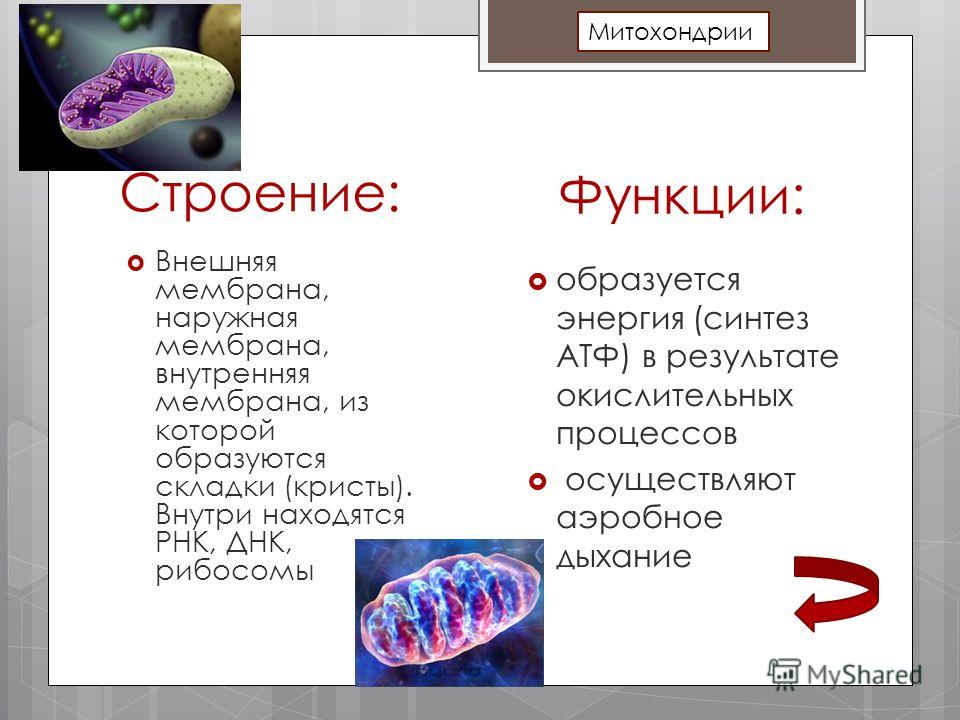 Опять же, это потому, что образуется всплеск активных форм кислорода, ущерб от которых и решает исход дела, вызывая гибель раковых клеток.
Опять же, это потому, что образуется всплеск активных форм кислорода, ущерб от которых и решает исход дела, вызывая гибель раковых клеток.
Питание митохондрий
С точки зрения питания, Патрик подчеркивает значение следующих питательных веществ и важных сопутствующих факторов, необходимых для правильного функционирования митохондриальных ферментов:
- Коэнзим Q10 или убихинол (восстановленная форма)
- L-карнитин, который переносит жирные кислоты в митохондрии
- D-рибоза, которая является сырьем для молекул АТФ
- Магний
- Все витамины группы В, в том числе рибофлавин, тиамин и B6
- Альфа-липоевая кислота (АЛК)
Как замечает Патрик:
«Я предпочитаю получать как можно больше питательных микроэлементов из цельных продуктов по целому ряду причин. Во-первых, они образуют между собой комплекс с волокнами, благодаря которому облегчается их всасывание.
Кроме того, в этом случае обеспечивается их правильное соотношение.
Получить их с избытком не удастся. Соотношение именно такое, как нужно. Есть и другие компоненты, которые, вероятно, еще предстоит определить.
Нужно быть очень бдительными, следя за тем, чтобы есть широкий спектр [продуктов] и получать правильные питательные микроэлементы. Я думаю, по этой причине полезно принимать добавки с комплексом витаминов В.
По этой причине их принимаю я. Другая причина заключается в том, что с возрастом мы перестаем так же легко усваивать витамины группы В, в основном, из-за увеличивающейся жесткости клеточных мембран. Это изменяет способ, которым витамины группы В транспортируются в клетку. Они водорастворимые, поэтому не хранятся в жире. Ими невозможно отравиться. В крайнем случае, будете мочиться чуть больше. Но я уверена в том, что они очень полезны».
Сохранить молодость митохондрий помогут физические упражнения
Физические упражнения тоже способствуют митохондриальному здоровью, поскольку они заставляют митохондрии трудиться. Как упоминалось ранее, одним из побочных эффектов усиленной работы митохондрий является создание активных форм кислорода, выступающих в качестве сигнальных молекул.
Как упоминалось ранее, одним из побочных эффектов усиленной работы митохондрий является создание активных форм кислорода, выступающих в качестве сигнальных молекул.
Одна из функций, сигнализируемых ими, является образование большего количества митохондрий. Поэтому, когда вы тренируетесь, организм реагирует, создавая больше митохондрий, чтобы удовлетворять повышенные запросы в энергии.
Старение неизбежно. Но ваш биологический возраст может сильно отличаться от хронологического, причем митохондрии имеют много общего с биологическим старением. Патрик цитирует недавнее исследование, которое показывает, как люди могут биологически стареть очень разными темпами.
Исследователи измерили более десятка различных биомаркеров, таких как длина теломера, повреждение ДНК, холестерин ЛПНП, метаболизм глюкозы и чувствительность к инсулину, в трех точках жизни людей: в возрасте 22, 32 и 38 лет.
«Мы обнаружили, что кто-то в возрасте 38 лет биологически мог выглядеть на 10 лет моложе или старше, судя по биологическим маркерам.
Несмотря на одинаковый возраст, биологическое старение происходит совершенно разными темпами.
Интересно, что когда этих людей сфотографировали и показали их фотографии прохожим с просьбой угадать хронологический возраст изображенных людей, то люди угадывали биологический, а не хронологической возраст».
Таким образом, независимо от фактического возраста, на сколько лет вы выглядите, соответствует вашим биологическим биомаркерам, которые в значительной степени обусловлены здоровьем митохондрий. Поэтому, хотя старения и не избежать, вы в значительной степени можете управлять тем, как вы стареете, а это, согласитесь, дает очень много возможностей. И одним из ключевых факторов является поддержание митохондрий в хорошем рабочем состоянии.
Как считает Патрик, «молодость» — это не столько хронологический возраст, сколько то, на какой возраст вы себя чувствуете, и насколько хорошо работает ваш организм:
«Я хочу знать, как оптимизировать свою мыслительную деятельность и свои спортивные результаты.
Я хочу продлить молодость. Я хочу дожить до 90. И когда я доживу, хочу заниматься серфингом в Сан-Диего точно так же, как и в свои 20. Я хотела бы угасать не так быстро, как некоторые люди. Мне нравится оттягивать это угасание и продлевать молодость столько, сколько получится, чтобы я как можно дальше радовалась жизни».
Что такое митохондрии? Если ответ на этот вопрос вызывает у вас затруднения, то наша статья как раз для вас. Мы рассмотрим особенности строения этих органелл во взаимосвязи с выполняемыми функциями.
Что такое органеллы
Но для начала давайте вспомним, что такое органеллы. Так называют постоянные клеточные структуры. Митохондрии, рибосомы, пластиды, лизосомы… Все это органеллы. Подобно самой клетке, каждая подобная структура имеет общий план строения. Органеллы состоят из поверхностного аппарата и внутреннего содержимого — матрикса. Каждую из них можно сравнить с органами живых существ. Органеллы также имеют свои характерные черты, обусловливающие их биологическую роль.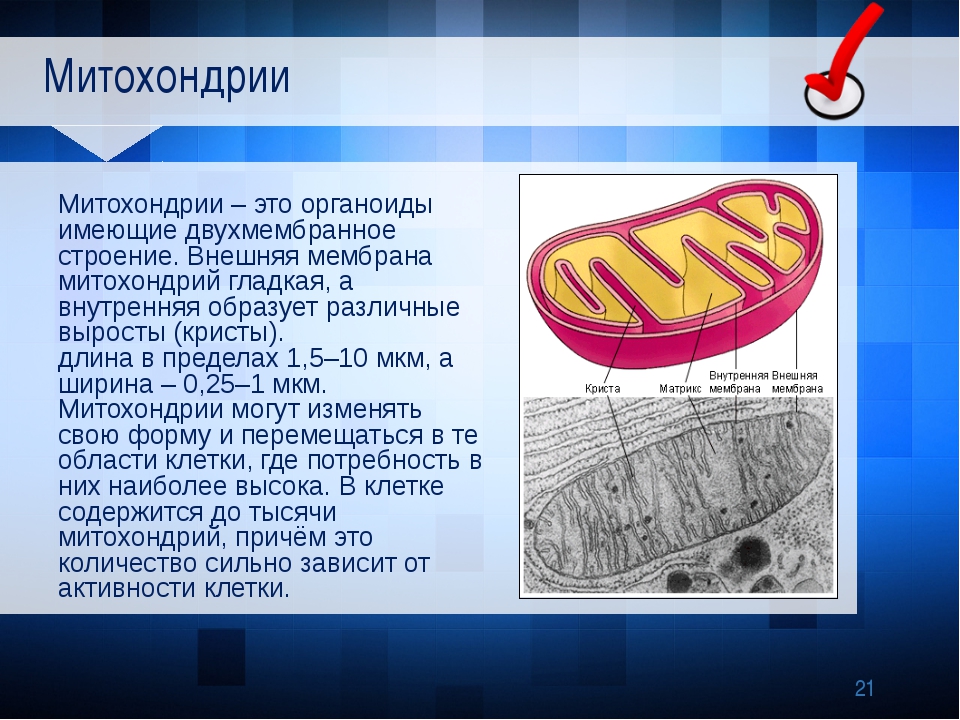
Классификация клеточных структур
Органеллы объединяют в группы по признаку строения их поверхностного аппарата. Различают одно-, дву- и немембранные постоянные клеточные структуры. К первой группе относятся лизосомы, комплекс Гольджи, эндоплазматический ретикулум, пероксисомы и различные виды вакуолей. Ядро, митохондрия и пластиды — двумембранные. А рибосомы, клеточный центр и органеллы движения полностью лишены поверхностного аппарата.
Теория симбиогенеза
Что такое митохондрии? Для эволюционного учения это не просто структуры клетки. Согласно симбиотической теории, митохондрии и хлоропласты являются результатом метаморфоз прокариот. Вполне возможно, что митохондрии произошли от аэробных бактерий, а пластиды — от фотосинтезирующих. Доказательством этой теории является тот факт, что данные структуры имеют собственный генетический аппарат, представленный кольцевой молекулой ДНК, двойную мембрану и рибосомы. Существует также предположение, что в дальнейшем от митохондрий произошли животные эукариотические клетки, а от хлоропластов — растительные.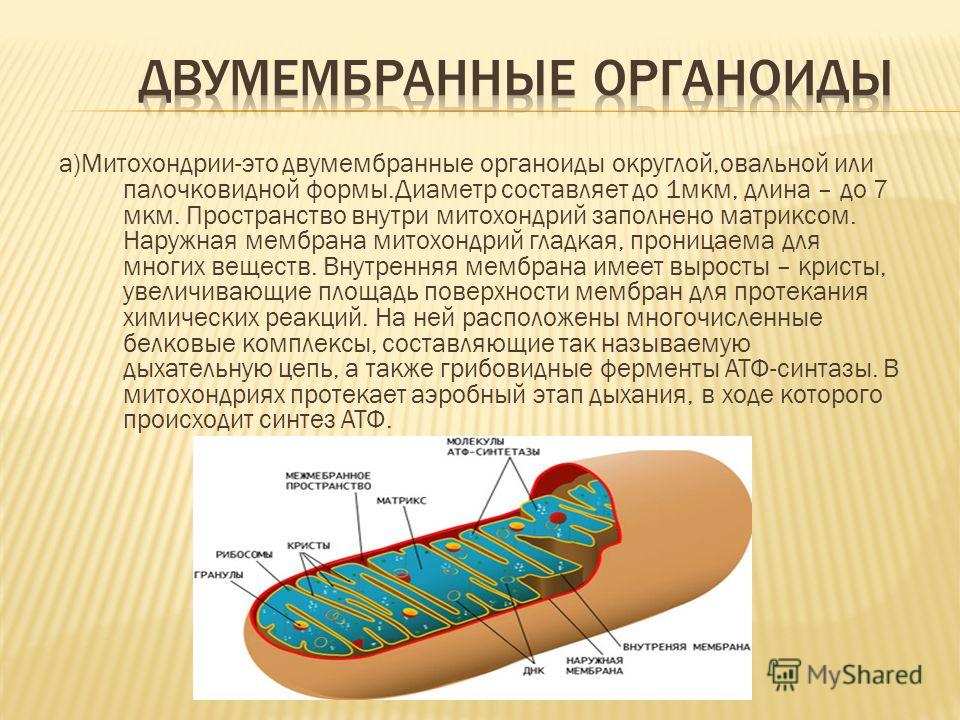
Расположение в клетках
Митохондрии являются составляющей частью клеток преобладающей части растений, животных и грибов. Отсутствуют они только у анаэробных одноклеточных эукариот, обитающих в бескислородной среде.
Строение и биологическая роль митохондрий долгое время оставались загадкой. Впервые при помощи микроскопа их удалось увидеть Рудольфу Келликеру в 1850 году. В мышечных клетках ученый обнаружил многочисленные гранулы, которые на свету были похожи на пух. Понять, какова роль этих удивительных структур, стало возможно благодаря изобретению профессора Пенсильванского университета Бриттона Ченса. Он сконструировал прибор, который позволял видеть сквозь органеллы. Так была определена структура и доказана роль митохондрий в обеспечении энергией клеток и организма в целом.
Форма и размер митохондрий
Общий план строения
Рассмотрим, что такое митохондрии с точки зрения особенностей их строения. Это двумембранные органеллы. Причем наружная — гладкая, а внутренняя имеет выросты.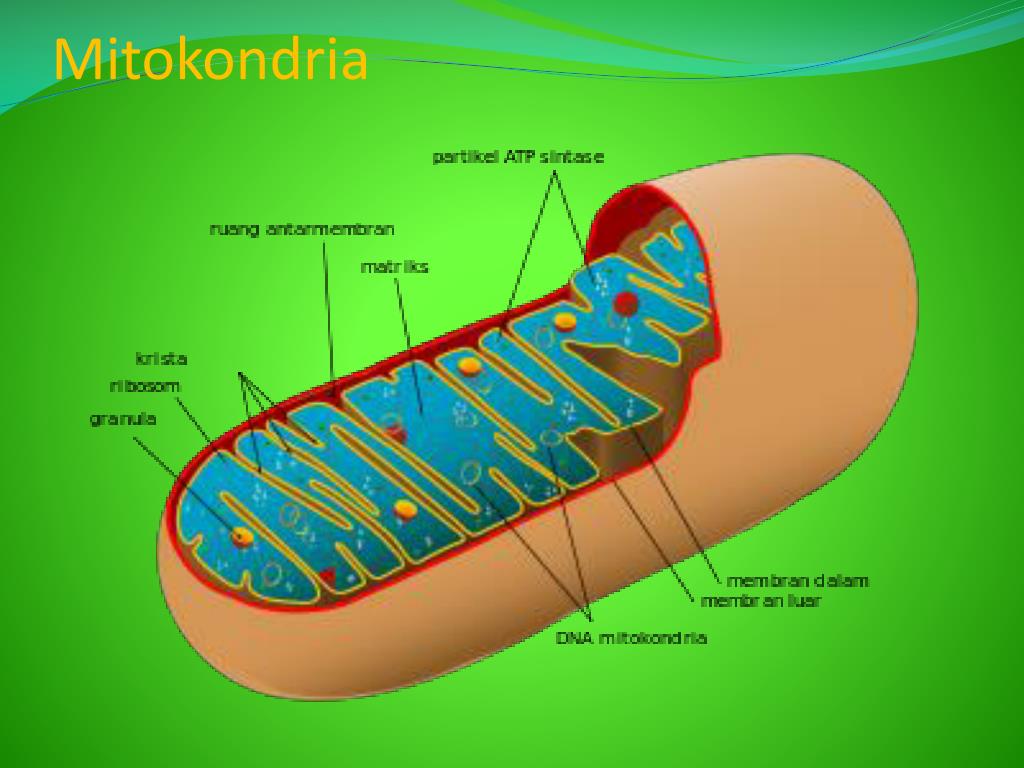 Матрикс митохондрий представлен различными ферментами, рибосомами, мономерами органических веществ, ионами и скоплениями кольцевых молекул ДНК. Такой состав делает возможным протекание важнейших химических реакций: цикла трикарбоновых кислот, мочевины, окислительного фосфорилирования.
Матрикс митохондрий представлен различными ферментами, рибосомами, мономерами органических веществ, ионами и скоплениями кольцевых молекул ДНК. Такой состав делает возможным протекание важнейших химических реакций: цикла трикарбоновых кислот, мочевины, окислительного фосфорилирования.
Значение кинетопласта
Мембрана митохондрии
Мембраны митохондрий не одинаковы по своему строению. Замкнутая наружная является гладкой. Она образована бислоем липидов с фрагментами белковых молекул. Его общая толщина составляет 7 нм. Данная структура выполняет функции отграничения от цитоплазмы, а также взаимосвязи органеллы с окружающей средой. Последняя возможна благодаря наличию белка порина, который формирует каналы. По ним посредством активного и пассивного транспорта передвигаются молекулы.
Химическую основу внутренней мембраны составляют белки. Она образует внутри органоида многочисленные складки — кристы. Эти структуры в значительной степени увеличивают активную поверхность органеллы.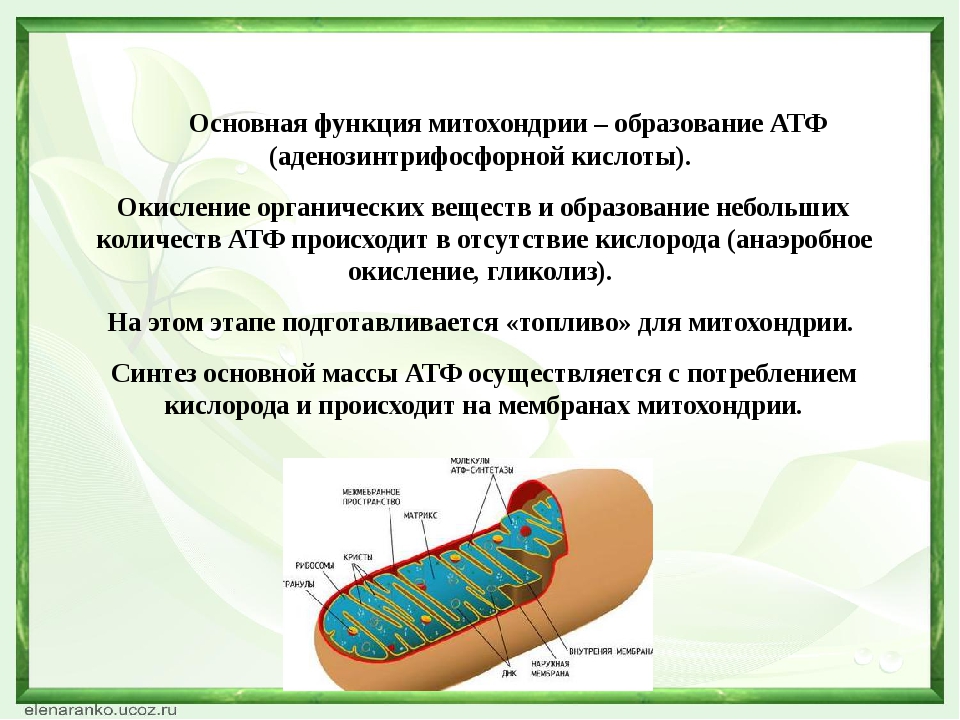 Главной особенностью строения внутренней мембраны является полная непроницаемость для протонов. В ней не образуются каналы для проникновения ионов извне. В отдельных местах наружная и внутренняя соприкасаются. Здесь расположен особый рецепторный белок. Это своеобразный проводник. С его помощью митохондриальные белки, которые закодированы в ядре, проникают внутрь органеллы. Между мембранами находится пространство, толщиной до 20 нм. В нем расположены различные виды белков, которые являются обязательными компонентами дыхательной цепи.
Главной особенностью строения внутренней мембраны является полная непроницаемость для протонов. В ней не образуются каналы для проникновения ионов извне. В отдельных местах наружная и внутренняя соприкасаются. Здесь расположен особый рецепторный белок. Это своеобразный проводник. С его помощью митохондриальные белки, которые закодированы в ядре, проникают внутрь органеллы. Между мембранами находится пространство, толщиной до 20 нм. В нем расположены различные виды белков, которые являются обязательными компонентами дыхательной цепи.
Функции митохондрий
Строение митохондрии напрямую взаимосвязано с выполняемыми функциями. Основная из них заключается в осуществлении синтеза аденозинтрифосфата (АТФ). Это макромолекула, которая случит основным переносчиком энергии в клетке. В ее состав входит азотистое основание аденин, моносахарид рибоза и три остатка фосфорной кислоты. Именно между последними элементами заключено основное количество энергии. При разрыве одной из них максимально ее может выделиться до 60 кДж. В целом прокариотическая клетка содержит 1 млрд молекул АТФ. Эти структуры постоянно находятся в работе: существование каждой из них в неизменном виде не продолжается больше одной минуты. Молекулы АТФ постоянно синтезируются и расщепляются, обеспечивая организм энергией в тот момент, когда это необходимо.
В целом прокариотическая клетка содержит 1 млрд молекул АТФ. Эти структуры постоянно находятся в работе: существование каждой из них в неизменном виде не продолжается больше одной минуты. Молекулы АТФ постоянно синтезируются и расщепляются, обеспечивая организм энергией в тот момент, когда это необходимо.
По этой причине митохондрии называют «энергетическими станциями». Именно в них происходит окисление органических веществ под действием ферментов. Энергия, которая при этом образуется, запасается и хранится в виде АТФ. К примеру, при окислении 1 г углеводов образуется 36 макромолекул этого вещества.
Строение митохондрии позволяет им выполнять еще одну функцию. Благодаря своей полуавтономности они являются дополнительным носителем наследственной информации. Ученые установили, что ДНК самих органелл не могут функционировать самостоятельно. Дело в том, что они не содержат всех необходимых для своей работы белков, поэтому заимствуют их в наследственном материале ядерного аппарата.
Итак, в нашей статье мы рассмотрели, что такое митохондрии. Это двумембранные клеточные структуры, в матриксе которых осуществляется ряд сложных химических процессов. Результатом работы митохондрий является синтез АТФ — соединение, которое обеспечивает организм необходимым количеством энергии.
Митохондрии строение — Справочник химика 21
Форма и строение митохондрий у различных микроорганизмов неодинаковы. Даже у одной и той же культуры при различных условиях и фазах роста форма и величина митохондрий меняется. В клетках дрожжей, перенесенных из аэробных условий в анаэробные, митохондрии теряют выраженную форму и образуются мембраны неопределенной формы. В бактериях функцию митохондрий выполняют особые образования цитоплазматической мембраны — мезосомы. Следовательно, в клетках бактерий аналогами митохондрий являются мезосомы. Как число митохондрий, так и число мезосом меняется, оно резко возрастает перед процессом деления клетки.
 Мезосомы бактерий специализируются в выполнении различных функций. Некоторые из них [c.19]
Перенос электронов от субстратов цикла трикарбоновых кислот к кислороду, сопровождающийся образованием воды, осуществляется сложной полиферментной системой, локализованной во внутренней мембране митохондрий. Последовательность функционирования отдельных дыхательных переносчиков в значительной мере была выяснена благодаря применению ингибиторного анализа, а также спектрофотометрических исследований. В настоящее время строение дыхательной цепи может быть представлено схемой на рис. 51. [c.435]
Мезосомы бактерий специализируются в выполнении различных функций. Некоторые из них [c.19]
Перенос электронов от субстратов цикла трикарбоновых кислот к кислороду, сопровождающийся образованием воды, осуществляется сложной полиферментной системой, локализованной во внутренней мембране митохондрий. Последовательность функционирования отдельных дыхательных переносчиков в значительной мере была выяснена благодаря применению ингибиторного анализа, а также спектрофотометрических исследований. В настоящее время строение дыхательной цепи может быть представлено схемой на рис. 51. [c.435] Вторая основная категория живых существ — это эукариоты, т. е. организмы, клетки которых содержат истинное ядро. Клетки эукариот крупнее и сложнее по строению, чем клетки прокариот. В ядре, окруженном мембраной, заключена большая часть ДНК, которая таким образом отделена от цитоплазмы. В цитоплазме содержатся различные органеллы, каждая из которых обладает характерной структурой, — митохондрии, лизосомы, центриоли. Клетки эукариот так разнообразны ло размерам и форме и настолько специализированы, что описать типичную клетку практически невозможно. Все же на рис. 1-3 мы попытались изобразить некую усредненную клетку, отчасти животную, отчасти растительную. [c.26]
Клетки эукариот так разнообразны ло размерам и форме и настолько специализированы, что описать типичную клетку практически невозможно. Все же на рис. 1-3 мы попытались изобразить некую усредненную клетку, отчасти животную, отчасти растительную. [c.26]
РИС. 10-9. А. Схема строения митохондрии. . Модель организации частиц в митохондриальных мембранах, основанная на электронно-микроскопических снимках сколов [c.395]
Другая цепь, называемая отстающей, имеет противоположное направление и обозначается 3 5. Двойная спираль характерна для большинства молекул ДНК. Тем не менее молекула ДНК может иметь не только двухспиральное строение, но и односпиральное, кольцевое, например, в вирусах, митохондриях. [c.46]
Размер митохондрий чаще равен 2—3 мкм в длину и около 1 мкм в диаметре, однако овальную форму митохондрий не следует Рис. 15.3. Схема строения [c.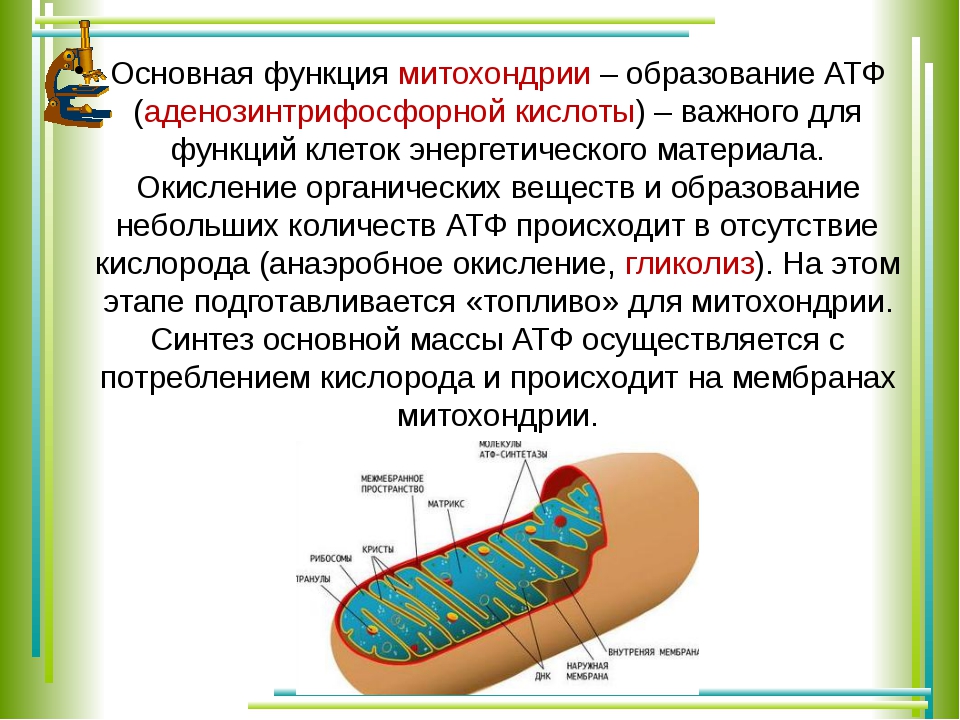 197]
197]
Содержимое всех живых клеток отделено от окружающей среды специальными структурами — биомембранами, которые обычно называют прото-плазматическими мембранами. У растений и бактерий наряду с такими мембранами снаружи клетки еще имеется клеточная стенка. Для эукариотических клеток характерно деление внутреннего содержимого клетки на отдельные отсеки, или компартменты. Они представляют собой субклеточные органеллы, ограниченные мембранами, например, ядро митохондрии, аппарат Гольджи. Однако мембраны служат не только поверхностями раздела. По существу, мембраны представляют собой сложные по строению и разнообразные по функциям биохимические системы. [c.106]
Строение и свойства митохондрий [c.429]
Изучение структуры мембран митохондрий методами рант-гено-структурного анализа и электронной микроскопии позволило сделать заключение, что темные слои стенок мембраны (рис. 3,1/) соответствуют слоям белка, а более светлые — бимолекулярным слоям липоидов. Общая картина строения мембранных стенок митохондрий в настоящее время представляется такой, какой она показана а рисунке 2>,VI. Каждая мембрана состоит из двух слоев белковых молекул и заключенных между ними двух слоев липидов. На этих белковых и липидных слоях адсорбированы ферменты, которые катализируют биохимические реакции в митохондриях. [c.30]
Общая картина строения мембранных стенок митохондрий в настоящее время представляется такой, какой она показана а рисунке 2>,VI. Каждая мембрана состоит из двух слоев белковых молекул и заключенных между ними двух слоев липидов. На этих белковых и липидных слоях адсорбированы ферменты, которые катализируют биохимические реакции в митохондриях. [c.30]
При рассмотрении строения клеточных структур (глава И) мы видели, что митохондрии представляют трехслойные мембраны с внутренними перегородками (рис. 2 и 3). На этих мембранах адсорбированы ферменты, катализирующие различные процессы обмена веществ. В настоящее время считается, что перенос электронов и водорода от НАД Нг на кислород может происходить в митохондриях двумя путями, из которых только один сопряжен с фосфорилированием, а другой не сопровождается синтезом АТФ. Эти два пути окисления НАД-Нг в митохондриях пространственно разграничены процессы окислительного фосфорилирования локализованы внутри митохондрий, а свободное окисление без фосфорилирования — на их поверхности. В соответствующих участках митохондрий имеются и соответствующие ферментные системы, катализирующие тот или иной процесс. Эти два пути окисления веществ отличаются не только местоположением, но и различной чувствительностью к специфическим ингибиторам и активаторам. [c.174]
В соответствующих участках митохондрий имеются и соответствующие ферментные системы, катализирующие тот или иной процесс. Эти два пути окисления веществ отличаются не только местоположением, но и различной чувствительностью к специфическим ингибиторам и активаторам. [c.174]
Регенерация АТР из ADP и Pj. Синтез АТР из ADP и неорганического фосфата (Pi) катализируется АТР-синтазой. Этот фермент преобразует доставляемую потоком электронов энергию в энергию фосфо-эфирных связей АТР. Фермент найден во всех мембранах, участвующих в преобразовании энергии, а именно в мембранах митохондрий, хлоропластов и бактерий. Он достаточно велик (мол. масса 350-10 ) и имеет сложное строение (рис. 7.12, Г)-состоит из головки, построенной из нескольких субъединиц, ножки и основания последнее погружено в липидный слой плазматической мембраны. АТР-синтаза катализирует присоединение фосфата к ADP с отщеплением молекулы воды, в результате чего образуется АТР. Каким образом поток протонов или протонный градиент осуществляет эту реакцию фосфорилирования, пока еще неизвестно возможно, что протоны по какому-то каналу или поре в молекуле фермента оттекают обратно внутрь митохондрии или бактерии, а освобождающаяся при этом энергия используется для фосфорилирования.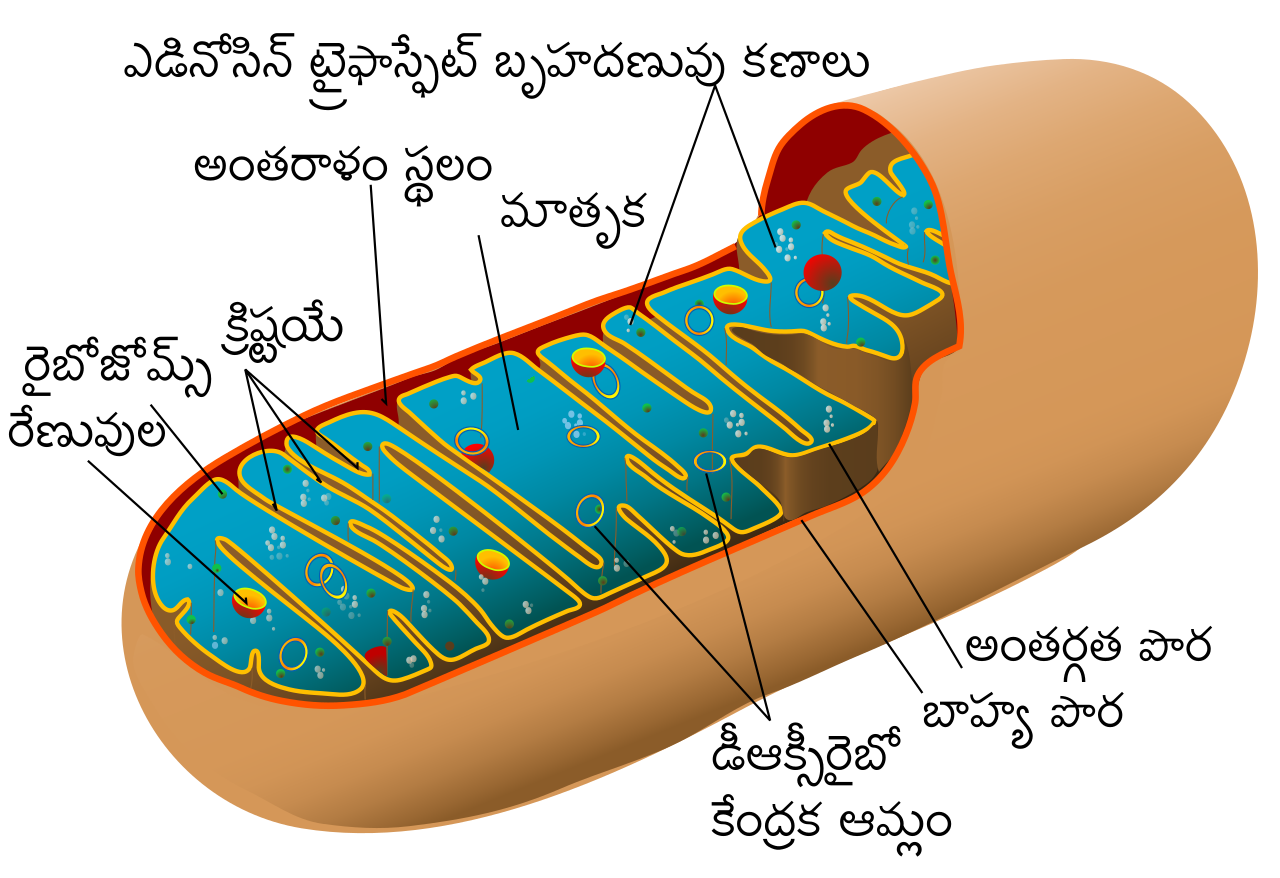 [c.245]
[c.245]
Химическое строение активной формы уксусной кислоты долгое время оставалось неясным только в последние годы удалось расшифровать структуру этого соединения. Вместе с тем был выяснен и механизм окислительного декарбоксилирования пировиноградной кислоты у некоторых микроорганизмов. [Установлено, что декарбоксилирование пировиноградной кислоты, сопровождающееся поглощением кислорода, катализируется сложной системой, в состав которой входит особая дегидрогеназа, коферменты (тиаминпирофосфат, липоевая кислота, коэнзим А, НАД) и система ферментов — катализаторов тканевого дыхания. Вся эта система локализована в митохондриях. [c.274]
РИС. 13-28. А. Схема строения палочки сетчатки позвоночных [135]. НЧ — наружный членнк СР — соединительная ресничка М — платно упакованные митохондрии Я — ядро СО — синаптическое окончание. Б. Электронная микрофотография продольного среза наружного членика палочки в сетчатке крысы (с любезного разрешения [c.
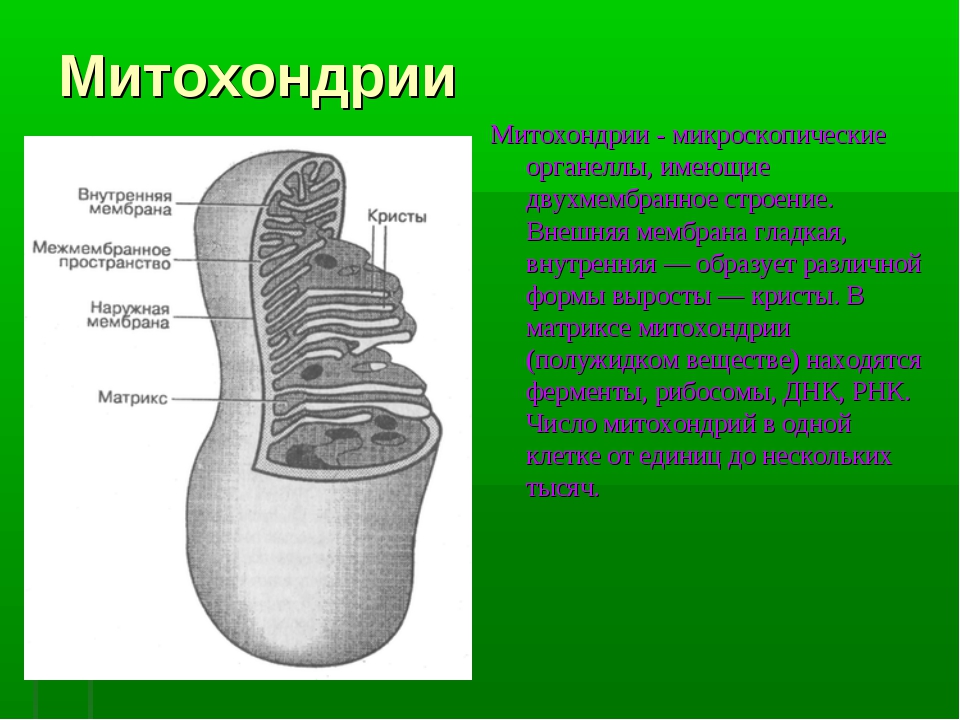 62]
62]Хлоропласты и митохондрии эукариотич. клеток содержат Р., отличные от типа 80S. Р. хлоропластов высших растений принадлежат к истинному 70S типу. Митохондриальные Р. более разнообразны их строение находится в зависимости от таксономич. принадлежности организма (т.е. от принадлежности к определенному виду, роду шш семейству). Напр., митохондриальные Р. млекопитающих существенно мельче типичных 70S Р. коэф. седиментации этих Р. составляет ок. 55S (т. наз. минирибосомы). [c.264]
В препаративной энзимологии чаще пользуются методом дифференциального центрифугирования гомогенатов тканей (рис. 4.26). Для этого сначала разрушают клеточную структуру с помощью подходящего дезинтегратора и полученную квазиоднородную (гомогенизированную) массу подвергают дифференциальному центрифугированию при температуре О—4°С. Обычно распределение ферментов изучают в последовательных индивидуальных фракциях, изолированных при дробном центрифугировании гомогенатов, в частности во фракции ядер, которую получают при низкой скорости центрифугирования, во фракции митохондрий, которая осаждается при средней скорости центрифугирования, во фракции микросом (или рибосом), для изолирования которой требуется высокая скорость центрифугирования, и, наконец, в оставшейся прозрачной надосадочной жидкости (супернатант), представляющей собой растворимую фракцию цитоплазмы. Следует отметить, что фракция митохондрий не является гомогенной, поскольку из нее удается изолировать частицы, известные как лизосомы, размер которьгх занимает промежуточное место между размерами митохондрий и микросом. В свою очередь микросомальная фракция также является гетерогенной, поскольку состоит в основном из элементов эндоплазматической сети неоднородного строения. [c.158]
Следует отметить, что фракция митохондрий не является гомогенной, поскольку из нее удается изолировать частицы, известные как лизосомы, размер которьгх занимает промежуточное место между размерами митохондрий и микросом. В свою очередь микросомальная фракция также является гетерогенной, поскольку состоит в основном из элементов эндоплазматической сети неоднородного строения. [c.158]
Число остатков изопрена в боковой цепи убихинона из разных источников варьирует от 6 до 10, что обозначают как KoQ , KoQ, и т.д. В митохондриях клеток человека и животных встречается убихинон только с 10 изопреновыми звеньями. Как и близкие к нему по структуре витамины К и Е, убихинон нерастворим в воде. В хлоропластах растений открыто близкое к убихинону соединение пластохинон, который отличается строением бензольного кольца вместо двух метоксильных остатков содержатся две метальные группы и отсутствует Hj-rpynna у 5-го углеродного атома. [c.243]
Строение и функциональная организация митохондрий явились предметом интенсивного изучения.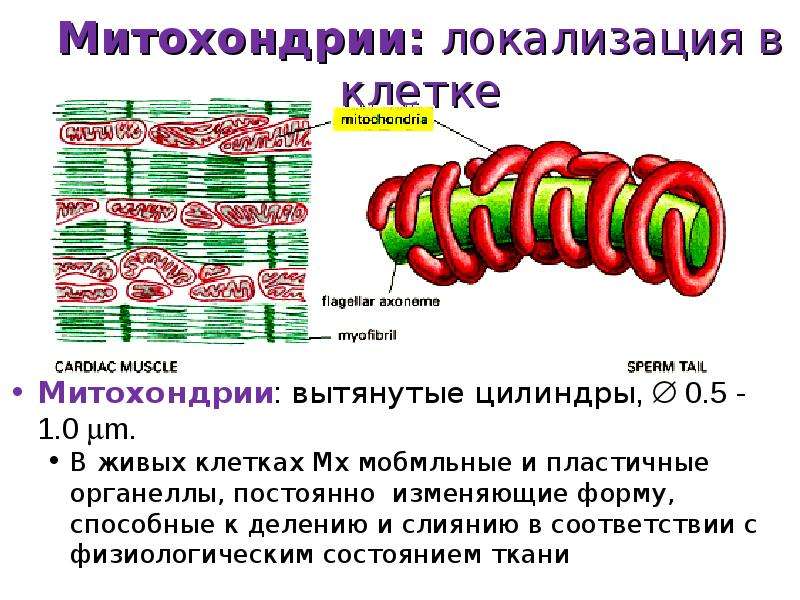 Тем не менее, многие важнейшие вопросы, сюда относящиеся, пока не имеют ответа. Специфическая мембранная структура митохондрий, присутствие в них автономной программы синтеза белков (ДНК), механохи-мйческая активность митохондрий непосредственно связаны с их ролью силовых станций клетки. [c.429]
Тем не менее, многие важнейшие вопросы, сюда относящиеся, пока не имеют ответа. Специфическая мембранная структура митохондрий, присутствие в них автономной программы синтеза белков (ДНК), механохи-мйческая активность митохондрий непосредственно связаны с их ролью силовых станций клетки. [c.429]
Электроны с восстановленных переносчиков (НАД Нз, НАДФ Нз, ФАД Нз), образующихся при функционировании ЦТК или окислительного пентозофосфатного цикла, поступают в дыхательную цепь, где проходят через ряд этапов, опускаясь постепенно на все более низкие энергетические уровни, и акцептируются соединением, служащим конечным акцептором электронов. Перенос электронов приводит к значительному изменению свободной энергии в системе. В наиболее соверщенном виде и единообразии дыхательная цепь предстает у эукариот, где она локализована во внутренней мембране митохондрий. У эубактерий дыхательные цепи поражают разнообразием своей конкретной организации при сохранении принципиального сходства в строении и функционировании. [c.360]
[c.360]
Помимо ядра и митохондрий в клетках эукариот существует и функционирует значительное число других органелл. Среди них следует в первую очередь упомянуть систему эндоплаэматическою ретикулума, которая представляет собой большое число ограниченных мембранами слоев, пузырьков, трубочек. Мембраны андоплазматического ретикулума oбpaзyютJ по-видимому, единую систему, связанную также с ядерной мембраной. Различают гладкий эндоплазматический ретикулум, имеющий трубчатое строение, в котором происходит в основном метаболизм липиДов. Кроме того, широко представлен шероховатый эндоплазматический ретикулум, на внешних поверхностях которого закреплены рибосомы. На этих рибосомах происходит синтез многочисленных мембранных белков. [c.434]
Липопротеины составляют большую группу сложных белков. Эти макромолекулы в значительных количествах находятся в митохондриях, из них в основном состоит эндоплазматический ретикулум, их обнаруживают и в плазме крови, и в молоке. Как правило, липопротеины — это большие молекулы. Их молекулярная масса достигает миллиона дальтон. Гидрофильность белковой и гидрофобность простетической группы липопротеинов определяют ту роль, которую они играют в процессах избирательной проницаемости. Липиды, входящие в состав липопротеинов, отличаются по строению и биологическим свойствам. В частности, в составе липопротеинов открыты нейтральные липиды, фосфолипиды, холестерин и др. Липидный компонент соединяется с белком при помощи нековалентных связей различной природы. Так, нейтральные липиды соединяются с белком посредством гидрофобных связей. Если же в образовании липопротеина участвует фосфолипид, то он взаимодействует с белком при помощи ионных связей. [c.48]
Как правило, липопротеины — это большие молекулы. Их молекулярная масса достигает миллиона дальтон. Гидрофильность белковой и гидрофобность простетической группы липопротеинов определяют ту роль, которую они играют в процессах избирательной проницаемости. Липиды, входящие в состав липопротеинов, отличаются по строению и биологическим свойствам. В частности, в составе липопротеинов открыты нейтральные липиды, фосфолипиды, холестерин и др. Липидный компонент соединяется с белком при помощи нековалентных связей различной природы. Так, нейтральные липиды соединяются с белком посредством гидрофобных связей. Если же в образовании липопротеина участвует фосфолипид, то он взаимодействует с белком при помощи ионных связей. [c.48]
У эукариот вьщеляют следующие основные группы мембран плазматическую, ядерную, эндоплазматического ретикулума, митохондрий, хлоропластов, возбудимые мембраны, миелиновые оболочки аксонов, нейронов и др. Несмотря на то что каждый тип мембран отличается по химическому составу и строению, выполняет специфичекие функции, мембраны имеют общие структурные особенности и построены по единому типу [c.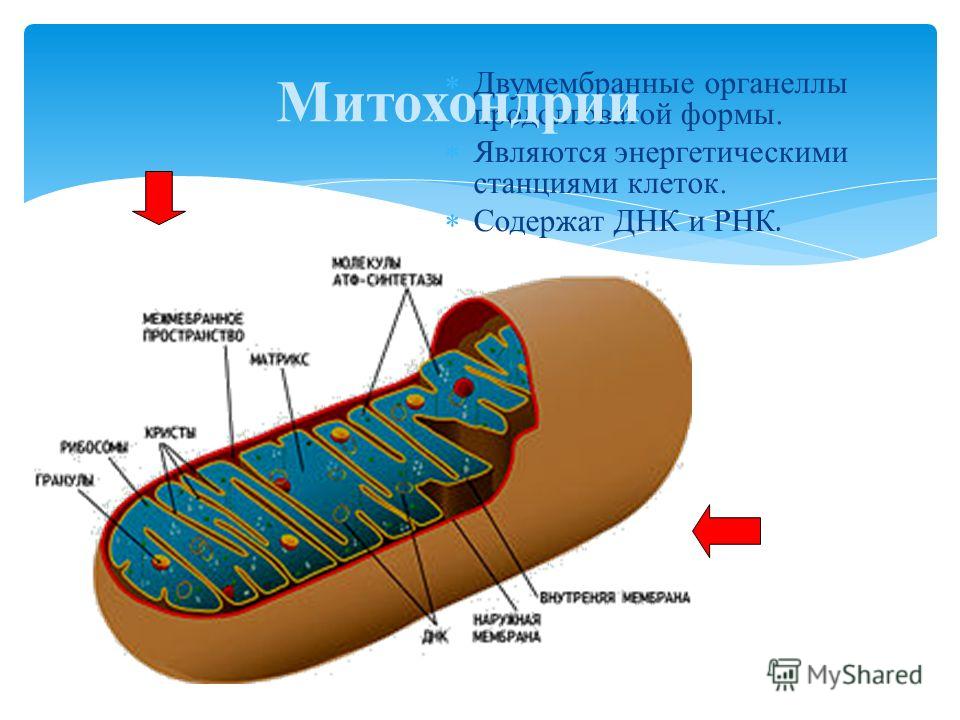 302]
302]
Два флавопротеидных фермента, играющих большую роль в дыхательной цепи, выделены в очищенном состоянии. Это сукцинатдегидрогеназа и НАД-Нг-цитохром-с-редуктаза. Сукцинатдегидрогеназа получена в виде почти гомогенного фермента из митохондрий бычьего сердца. Она содержит одну молекулу ФАД и 4 атома негеминового железа на молекулу фермента. Сукцинатдегидрогеназа катализирует восстановление сукцинатом феназинметасуль-фата и феррицианида метиленовый синий, цитохром с и кислород не восстанавливаются. НАД-Нг-цитохром-с-редуктаза выделена из сердечной мышцы и из частиц, переносящих электроны (см. стр. 225). Как и сукцинатдегидрогеназа, она содержит одну молекулу флавина (точное химическое строение его неизвестно) и 2—4 атома негеминового железа на молекулу фермента. Негеминовое железо у этих двух ферментов, вероятно, играет важную роль в восстановлении цитохрома с. Показано [3], что негеминовое железо в НАД-Нг-Цитохром-с-редуктазе во время катализа претерпевает восстановление и окисление. Недавно из растений были получены растворимые препараты сукцинатдегидрогеназы, а из митохондрий проростков гороха и из початка Arum в частично очищенном виде получена растворимая НАД-Нг-цитохром-с-редуктаза. [c.212]
Недавно из растений были получены растворимые препараты сукцинатдегидрогеназы, а из митохондрий проростков гороха и из початка Arum в частично очищенном виде получена растворимая НАД-Нг-цитохром-с-редуктаза. [c.212]
Фракционирование клеток в солевом растворе осложняется тенденцией гранул образовывать скопления и осаждаться в виде комков, а не в виде отдельных частиц. Это нежелательное явление можно предотвратить, измельчая клетки в 0,88 М растворе сахарозы, в котором митохондрии сохраняют палочковидное строение и способность к суправитальному окрашиванию янусом зеленым В. Однако при указанной концентрации сахарозы среда становится настолько вязкой и плотной, что для осаяедения субклеточных фракций приходится использовать чрезвычайно высокие скорости центрифугирования. Поэтому в современных исследованиях в качестве среды для измельчения клеток используют 0,25 М раствор сахарозы, в котором не происходит агрегации гранул и легко выделяется фракция митохондрий. Последние при этом обладают теми я е биохимическими свойствами, что и митохондрии, получаемые в 0,88 М растворе сахарозы, хотя они уяге не окрашиваются янусом зеленым В и имеют скорее шаровидную, а не удлиненную форму. При разделении путем дифференциального центрифугирования субклеточных фракций из гомогената в 0,25 М растворе сахарозы, полученного в гомогенизаторе Поттера — Эльвейема, удаление ядер и клеточных обломков, включая [c.130]
При разделении путем дифференциального центрифугирования субклеточных фракций из гомогената в 0,25 М растворе сахарозы, полученного в гомогенизаторе Поттера — Эльвейема, удаление ядер и клеточных обломков, включая [c.130]
Еще одно важное различие между эукариотами и прокариотами состоит в том, что эукариотические клетки, кроме организованного ядра, содержат целый рад других ограниченных мембранами внутриклеточных органелл, таких, как митохондрии, эндоплазматический ретикулум и тельца Гольджи. Каждая из этих органелл выполняет специфические функции в метаболизме и жизнедеятельности клетки. На рис. 2-7 показана типичная эукариотическая клетка-клетка печени крысы, имеющая необычайно сложное внутреннее строение с высокой степенью ковлпартментации. Как мы увидим в дальнейшем, эукариотичёским клеткам свойственно более тонкое разделение функций между содержащимися в них многочисленными структурными элементами, каждый из которых играет специфическую роль в жизнедеятельности клетки.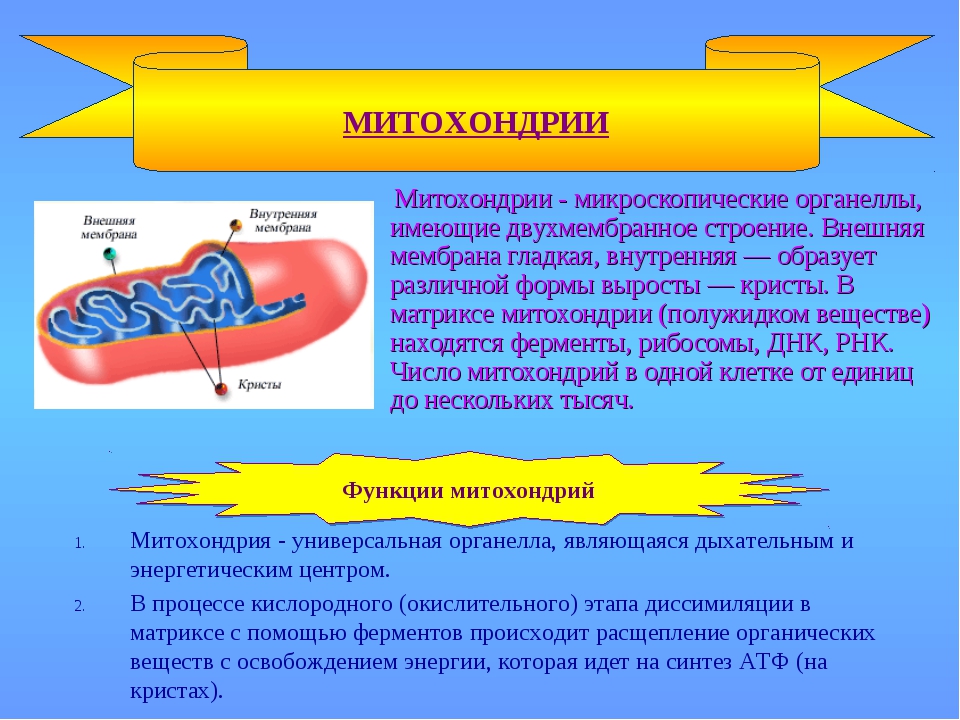 [c.33]
[c.33]
Клеточные ядра, хлоропласты, митохондрии и микросо.мы— это очень сложные структурные образования. В различных клеточных структурах и в основном веществе цитоплазмы главнейшими структурными элементами, которые определяют характер их строения, являются мембраны. Они есть во всех клеточных структурах и определяют многие специфические свойства леточных структур. [c.28]
Значительно подробнее строение клетки было изучено при помощи новейшего метода электронной микроскопии [79—84]. Но алектронпых микрофотографиях (3 и 4) митохондрии (размером 0,5 мк — 5 мк X 0,3—0,7 мк) имеют овальные очертания. Каждая митохондрия имеет наружную и внутреннюю мембраны [c.126]
Какое строение и функции митохондрий. Строение митохондрии
О СЛОЖНОМ ПРОСТЫМ ЯЗЫКОМ.
Тема эта сложная и комплексная, затрагивающая сразу же огромное количество биохимических процессов происходящих в нашем организме. Но давайте все таки попробуем разобраться, что же такое митохондрии и как они работают.
И так, митохондрии это одна из самых важных составляющих живой клетки. Если говорить простым языком то можно сказать, что это энергетическая станция клетки . Их деятельность основана на окисление органических соединений и генерации электрического потенциала (энергии освободившейся при распаде молекулы АТФ) для осуществления мышечного сокращения.
Все мы знаем, что работа нашего организма происходит в строгом соответствии с первым законом термодинамики. Энергия не создается в нашем организме, а лишь превращается. Организм только выбирает форму трансформации энергии, не производя ее, от химической к механической и тепловой. Основным источником всей энергии на планете Земля является Солнце. Приходя к нам в форме света, энергия поглощается хлорофиллом растений, там она возбуждает электрон атома водорода и таким образом дает энергию живой материи.
Своей жизнью мы обязаны энергии маленького электрона.
Работа митохондрии заключается в ступенчатом переносе энергии электрона водорода между атомами металлов, присутствующих в группах белковых комплексов дыхательной цепи (электронно-транспортной цепи белков), где каждый последующий комплекс обладает более высоким сродством к электрону притягивая его, чем предыдущий, до тех пор, пока электрон не соединиться с молекулярным кислородом, обладающим наибольшим сродством к электрону.
Каждый раз при передачи электрона по цепи высвобождается энергия которая аккумулируется в виде электрохимического градиента и затем реализовывается в виде мышечного сокращения и выделения тепла.
Серия окислительных процессов в митохондрии позволяющая перенести энергетический потенциал электрона называется «внутриклеточным дыханием» или часто «дыхательной цепью» , так как электрон по цепочки передается от атома к атому до тех пор пока не достигнет своей конечной цели атома кислорода.
Митохондриям нужен кислород для переноса энергии в процессе окисления.
Митохондрии потребляют до 80% кислорода который мы вдыхаем.
Митохондрия представляет из себя постоянную структуру клетки, расположенную в ее цитоплазме. Размер митохондрии обычно составляет от 0,5 до 1 мкм в диаметре. По форме она имеет зернистую структуру и может занимать до 20% объема клетки. Такая постоянная органическая структура клетки называется органелла
. К органеллам относятся и миофибриллы – сократительные единицы мышечной клетки; и ядро клетки это тоже органелла.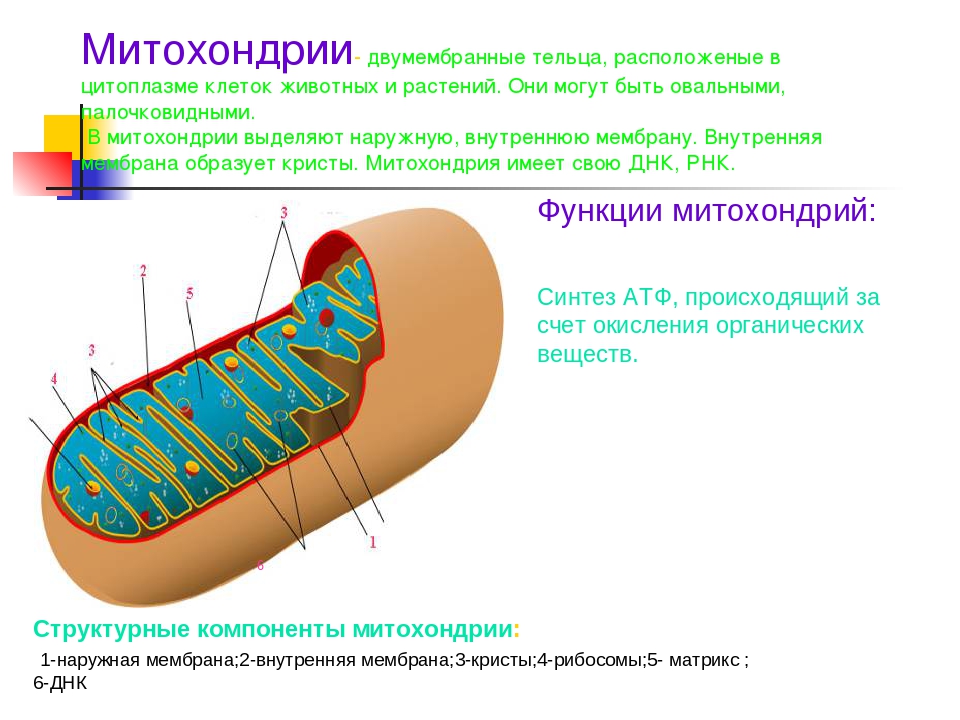 Вообще, любая постоянная структура клетки является органоидом-органеллой.
Вообще, любая постоянная структура клетки является органоидом-органеллой.
Открыл митохондрии и впервые описал немецкий анатом и гистолог Рихард Альтман в 1894 году, а название этой органелле дал другой немецкий гистолог К. Бенд в 1897 году. Но только в 1920 году, опять же немецкий биохимик Отто Вагбург, доказал, что с митохондриями связаны процессы клеточного дыхания.
Существует теория, согласно которой митохондрии появились в результате захвата примитивными клетками, клетками которые сами не могли использовать кислород для генерации энергии, бактерий протогенотов, которые могли это делать. Именно потому, что митохондрия ранее представляла из себя отдельный живой организм она и по сей день обладает собственным ДНК.
Митохондрии ранее представляли из себя самостоятельный живой организм.
В ходе эволюции прогеноты предали множество своих генов сформировавшемуся, благодаря повысившейся энергоэффективности, ядру и перестали быть самостоятельными организмами. Митохондрии присутствуют во всех клетках. Даже в сперматозоиде есть митохондрии. Именно благодаря им приводится в движение хвостик сперматозоида осуществляющий его движение. Но особенно много митахондрий в тех местах, где необходима энергия для любых жизненных процессов. И это конечно прежде всего мышечные клетки.
Даже в сперматозоиде есть митохондрии. Именно благодаря им приводится в движение хвостик сперматозоида осуществляющий его движение. Но особенно много митахондрий в тех местах, где необходима энергия для любых жизненных процессов. И это конечно прежде всего мышечные клетки.
В мышечных клетках митохондрии могут объединяться в группы гигантских разветвленных митохондрий, связанных друг с другом с помощью межмитохондриальных контактов, в которых они создают согласованную работающую кооперативную систему . Пространство в такой зоне имеет повышенную электронную плотность. Новые митохондрии образуются путем простого деления предыдущих органелл. Наиболее «простой» и доступный всем клеткам механизм энергетического обеспечения чаще всего называют общим понятием гликолиз .
Это процесс последовательного разложения глюкозы до пировиноградной кислоты. Если этот процесс происходит без участия молекулярного кислорода
или с недостаточным его присутствием, то он называется анаэробный гликолиз
. При этом глюкоза расщепляется не до конечных продуктов, а до молочной и пировиноградной кислоты которая далее претерпевает дальнейшие превращения в ходе брожения. Поэтому высвобождающейся энергии бывает меньше, но и скорость получения энергии быстрее. В результате анаэробного гликолиза из одной молекулы глюкозы клетка получает 2 молекулы АТФ и 2 молекулы молочной кислоты. Такой «базовый» энергетический процесс может протекать внутри любой клетки без участия митохондрий
.
При этом глюкоза расщепляется не до конечных продуктов, а до молочной и пировиноградной кислоты которая далее претерпевает дальнейшие превращения в ходе брожения. Поэтому высвобождающейся энергии бывает меньше, но и скорость получения энергии быстрее. В результате анаэробного гликолиза из одной молекулы глюкозы клетка получает 2 молекулы АТФ и 2 молекулы молочной кислоты. Такой «базовый» энергетический процесс может протекать внутри любой клетки без участия митохондрий
.
В присутствии молекулярного кислорода внутри митохондрий осуществляется аэробный гликолиз в рамках «дыхательной цепи». Пировиноградная кислота в аэробных условиях вовлекается в цикл трикарбоновых кислот или цикл Кребса . В результате этого многостадийного процесса из одной молекулы глюкозы образуется 36 молекул АТФ. Сравнение энергетического баланса клетки, имеющей развитые митохондрии и клетки, где они не развиты показывает (при достаточном количестве кислорода) различие в полноте использования энергии глюкозы внутри клетки почти в 20 раз!
У человека, волокна скелетных мышц можно условно разделить на три типа исходя из механических и метаболических свойств: — медленные окислительные; — быстрые гликолитические; — быстрые окислительно-гликолитические.
Быстрые мышечные волокна предназначены для выполнения быстрой и тяжелой работы. Для своего сокращения они используют в основном быстрые источники энергии, а именно криатинфосфот и анаэробный гликолиз. Содержание митохондрий в таких типах волокон значительно меньше чем в медленных мышечных волокнах.
Медленные мышечные волокна выполняют медленные сокращения, но способны работать длительное время. В качестве энергии они используют аэробный гликолиз и синтез энергии из жиров. Это дает гораздо больше энергии чем анаэробный гликолиз, но требует в замен больше времени, так как цепочка деградации глюкозы более сложная и требует присутствия кислорода, транспортировка которого к месту преобразования энергии тоже занимает время. Медленные мышечные волокна называют красными из-за миоглобина – белка, ответственный за доставку кислорода внутрь волокна. Медленные мышечные волокна содержат значительное количество митохондрий.
Возникает вопрос, каким образом и с помощью каких упражнений можно развить в мышечных клетках разветвленную сеть митохондрий? Существуют различные теории и методики тренировок и о них в материале по .
Характерные для подавляющего большинства клеток. Основная функция – это окисление органических соединений и производство АТФ-молекул из освободившейся энергии. Маленькая митохондрия является главной энергетической станцией всего организма.
Происхождение митохондрий
Среди ученых на сегодняшний день весьма популярно мнение, что митохондрия появилась в клетке не самостоятельно в ходе эволюции. Скорее всего, это произошло из-за захвата примитивной клеткой, не способной на тот момент самостоятельно использовать кислород, бактерии, которая умела это и, соответственно, была превосходным источником энергии. Подобный симбиоз оказался успешным и закрепился в последующих поколениях. В пользу этой теории указывает наличие в митохондриях собственной ДНК.
Как устроены митохондрии
Митохондрия обладает двумя мембранами: наружной и внутренней. Главная функция наружной мембраны – это отделение органоида от цитоплазмы клетки. Она состоит из билипидного слоя и белков, пронизывающих его, через которые и осуществляется транспорт молекул и ионов, необходимых для работы.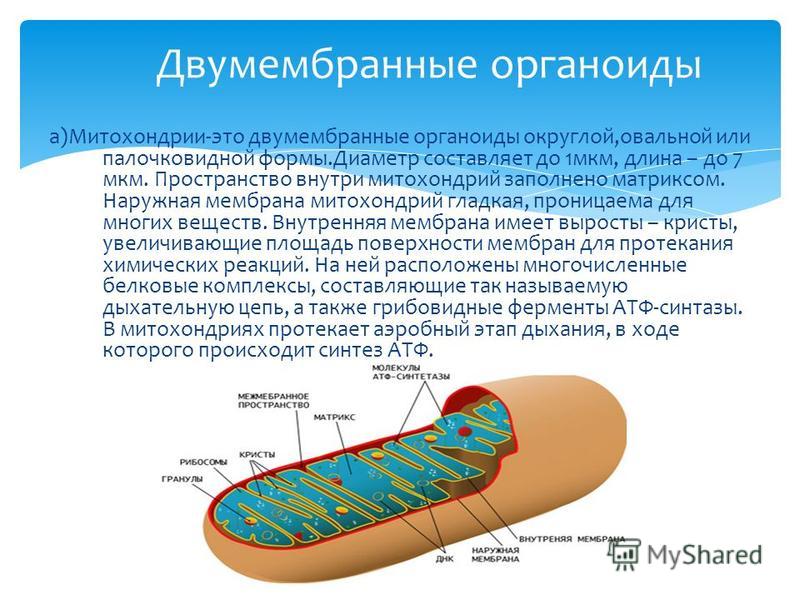 В то время как гладкая, внутренняя образует многочисленные складки – кристы, которые существенно увеличивают ее площадь. Внутренняя мембрана по большей части состоит из белков, среди которых присутствуют ферменты дыхательной цепи, транспортные белки и крупные АТФ-синтетазные комплексы. Именно в этом месте происходит синтез АТФ. Между наружной и внутренней мембраной находится межмембранное пространство с присущими ему ферментами.
В то время как гладкая, внутренняя образует многочисленные складки – кристы, которые существенно увеличивают ее площадь. Внутренняя мембрана по большей части состоит из белков, среди которых присутствуют ферменты дыхательной цепи, транспортные белки и крупные АТФ-синтетазные комплексы. Именно в этом месте происходит синтез АТФ. Между наружной и внутренней мембраной находится межмембранное пространство с присущими ему ферментами.
Внутреннее пространство митохондрий называется матрикс. Здесь расположены ферментные системы окисления жирных кислот и пирувата, ферменты цикла Кребса, а также наследственный материал митохондрий – ДНК, РНК и белоксинтезирующий аппарат.
Для чего нужны митохондрии
Главной функцией митохондрий является синтез универсальной формы химической энергии – АТФ. Также они принимают участие в цикле трикарбоновых кислот, превращая пируват и жирные кислоты в ацетил-CoA, а затем окисляя его. В этом органоиде хранится и передается по наследству митохондриальная ДНК, кодирующая воспроизводство тРНК, рРНК и некоторых белков, необходимых для нормального функционирования митохондрий.
Митохондрия — это спиральная, округлая, вытянутая или разветвленная органелла.
Впервые понятие митохондрия было предложено Бенда в 1897 г. Митохондрии можно обнаружить в живых клетках с помощью фазово-контрастной и интерференционной микроскопии в виде зерен, гранул или нитей. Это довольно подвижные структуры, которые могут перемещаться, сливаться друг с другом, делиться. При окраске специальными методами в погибших клетках при световой микроскопии митохондрии имеют вид мелких зерен (гранул), диффузно распределенных в цитоплазме или концентрирующихся в каких-то определенных ее зонах.
В результате разрушения глюкозы и жиров в присутствии кислорода в митохондриях образуется энергия, а органические вещества превращаются в воду и диоксид углерода. Именно таким образом получают основную энергию, необходимую для жизнедеятельности, животные организмы. Энергия накапливается в аденозинтрифосфате (АТФ), а точнее, в его макроэргических связях. Функция митохондрий тесно связана с окислением органических соединений и использованием освобождающейся при их распаде энергии для синтеза молекул АТФ. Поэтому митохондрии часто называют энергетическими станциями клетки, или органеллами клеточного дыхания. АТФ выполняет функцию поставщика энергии, перенося одну из своих богатых энергией концевых фосфатных групп на другую молекулу, и превращается при этом в АДФ.
Поэтому митохондрии часто называют энергетическими станциями клетки, или органеллами клеточного дыхания. АТФ выполняет функцию поставщика энергии, перенося одну из своих богатых энергией концевых фосфатных групп на другую молекулу, и превращается при этом в АДФ.
Предполагают, что в эволюции митохондрии были прокариотическими микроорганизмами, которые стали симбиотами в организме древней клетки. В последующем они стали жизненно необходимы, что было связано с увеличением содержания кислорода в атмосфере Земли. С одной стороны, митохондрии удаляли избыток токсичного для клетки кислорода, а с другой — обеспечивали энергией.
Без митохондрий клетка практически не в состоянии использовать кислород как вещество, обеспечивающее поставку энергии, и может восполнять свои энергетические потребности лишь путем анаэробных процессов. Таким образом, кислород — это яд, но яд жизненно важный для клетки, причем избыток кислорода так же вреден, как и его недостаток.
Митохондрии могут изменять свою форму и перемещаться в те области клетки, где потребность в них наиболее высока.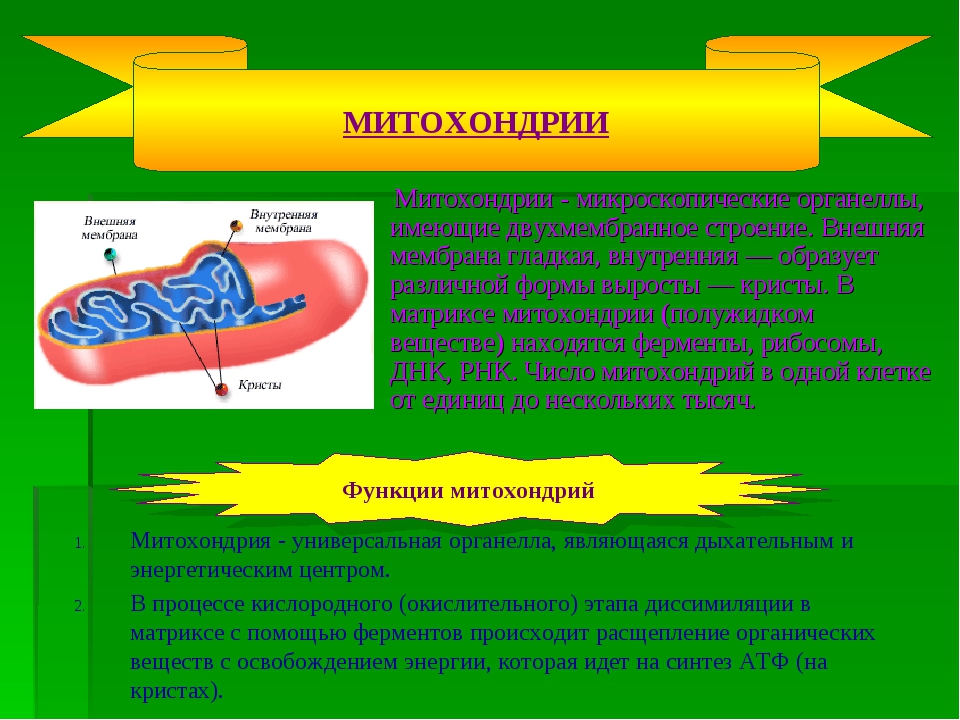 Так, в кардиомиоцитах митохондрии находятся вблизи миофибрилл, в клетках почечных канальцев вблизи базальных впячиваний и т. д. В клетке содержится до тысячи митохондрий, и их количество зависит от активности клетки.
Так, в кардиомиоцитах митохондрии находятся вблизи миофибрилл, в клетках почечных канальцев вблизи базальных впячиваний и т. д. В клетке содержится до тысячи митохондрий, и их количество зависит от активности клетки.
Митохондрии имеют средние поперечные размеры 0,5…3 мкм. В зависимости от размеров выделяют мелкие, средние, крупные и гигантские митохондрии (формируют разветвленную сеть — митохондриальный ретикулум). Размеры и число митохондрий тесно связаны с активностью клетки и ее энергопотреблением. Они крайне изменчивы и в зависимости от активности клетки, содержания кислорода, гормональных влияний могут набухать, изменять число и структуру крист, варьировать в числе, форме и размерах, а также ферментативной активности.
Объемная плотность митохондрий, степень развития их внутренней поверхности и другие показатели зависят от энергетических потребностей клетки. В лимфоцитах имеется всего по несколько митохондрий, а в печеночных клетках их 2…3 тыс.
Митохондрии состоят из матрикса, внутренней мембраны, перимитохондриального пространства и наружной мембраны. Наружная митохондриальная мембрана отделяет органеллу от гиалоплазмы. Обычно она имеет ровные контуры и замкнута так, что представляет собой мембранный мешок.
Наружная митохондриальная мембрана отделяет органеллу от гиалоплазмы. Обычно она имеет ровные контуры и замкнута так, что представляет собой мембранный мешок.
Внешнюю мембрану от внутренней отделяет перимитохондриальное пространство шириной около 10…20 нм. Внутренняя митохондриальная мембрана ограничивает собственно внутреннее содержимое митохондрии — матрикс. Внутренняя мембрана образует многочисленные выпячивания внутрь митохондрий, которые имеют вид плоских гребней, или крист.
По форме кристы могут иметь вид пластинок (трабекулярные) и трубочек (мультивезикулярные на срезе), а направлены они продольно или поперечно по отношению к митохондрии.
Каждая митохондрия заполнена матриксом, который на электронных микрофотографиях выглядит плотнее, чем окружающая цитоплазма. Матрикс митохондрии однородный (гомогенный), иногда мелкозернистый, различной электронной плотности. В нем выявляют тонкие нити толщиной около 2…3 нм и гранулы размером около 15…20 нм. Нити матрикса представляют собой молекулы ДНК, а мелкие гранулы — митохондриальные рибосомы.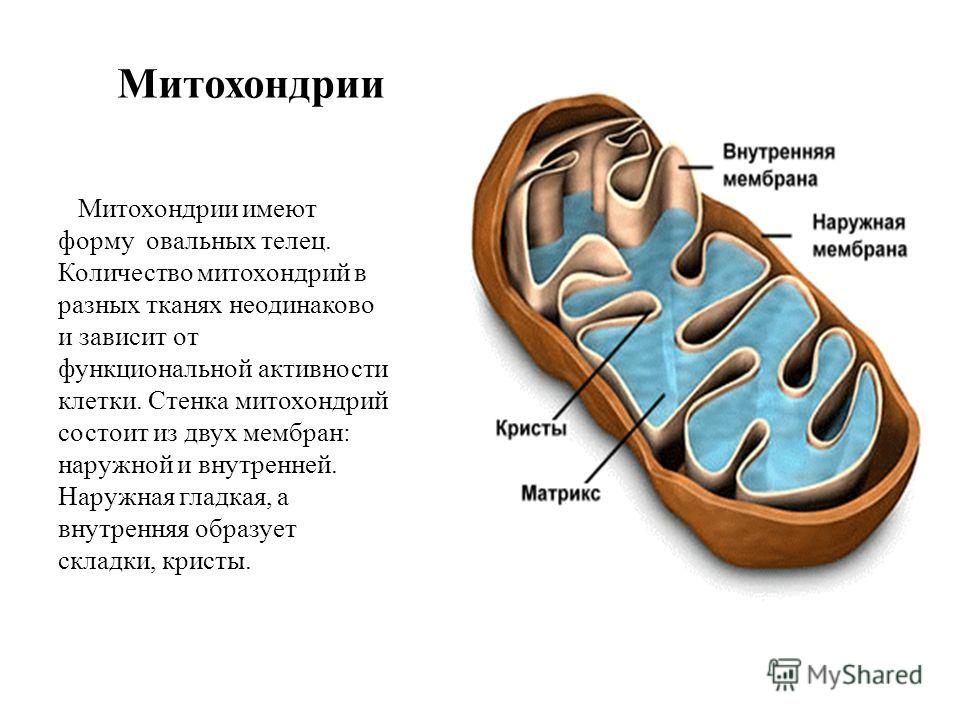 В матриксе содержатся ферменты, одна одноцепочечная, циклическая ДНК, митохондриальные рибосомы, много ионов Са 2+ .
В матриксе содержатся ферменты, одна одноцепочечная, циклическая ДНК, митохондриальные рибосомы, много ионов Са 2+ .
Автономная система белкового синтеза митохондрий представлена молекулами ДНК, свободными от гистонов. ДНК короткая, имеет форму кольца (циклическая) и содержит 37 генов. В отличие от ядерной ДНК в ней практически нет некодирующих последовательностей нуклеотидов. Особенности строения и организации сближают ДНК митохондрий с ДНК бактериальных клеток. На ДНК митохондрий происходит синтез молекул РНК разных типов: информационных, трансфертных (транспортных) и рибосомальных. Информационная РНК митохондрий не подвергается сплайсингу (вырезанию участков, не несущих информационной нагрузки). Малые размеры молекул митохондриальных ДНК не могут определить синтез всех белков митохондрий. Большинство белков митохондрий находится под генетическим контролем клеточного ядра и синтезируется в цитоплазме, так как ДНК митохондрий слабо выражена и может обеспечить образование лишь части ферментов цепи окислительного фосфорилирования. Митохондриальная ДНК кодирует не более десяти белков, которые локализованы в мембранах и представляют собой структурные белки, ответственные за правильную интеграцию отдельных функциональных белковых комплексов митохондриальных мембран. Синтезируются также белки, осуществляющие транспортные функции. Такая система белкового синтеза не обеспечивает всех функций митохондрии, поэтому автономия митохондрий ограниченная и относительная.
Митохондриальная ДНК кодирует не более десяти белков, которые локализованы в мембранах и представляют собой структурные белки, ответственные за правильную интеграцию отдельных функциональных белковых комплексов митохондриальных мембран. Синтезируются также белки, осуществляющие транспортные функции. Такая система белкового синтеза не обеспечивает всех функций митохондрии, поэтому автономия митохондрий ограниченная и относительная.
У млекопитающих митохондрии при оплодотворении передаются лишь через яйцеклетку, а спермий привносит в новый организм ДНК ядра.
В матриксе митохондрий образуются рибосомы, отличающиеся от рибосом цитоплазмы. Они участвуют в синтезе ряда митохондриальных белков, не кодируемых ядром. Митохондриальные рибосомы имеют число седиментации 60 (в отличие от цитоплазматических с числом седиментации 80). Число седиментации — это скорость осаждения при центрифугировании и ультрацентрифугировании. По строению митохондриальные рибосомы близки к рибосомам прокариотических организмов, но меньшего размера и отличаются чувствительностью к определенным антибиотикам (левомицетину, тетрациклину и др. ).
).
Внутренняя мембрана митохондрии обладает высокой степенью избирательности при транспорте веществ. К ее внутренней поверхности прикрепляются тесно прилежащие друг к другу ферменты цепи окислительного фосфорилирования, белки-переносчики электронов, транспортные системы АТФ, АДФ, пируват и др. В результате тесного расположения ферментов на внутренней мембране обеспечивается высокая сопряженность (взаимосвязанность) биохимических процессов, повышающая скорость и эффективность каталитических процессов.
При электронной микроскопии выявляют грибовидные частицы, выступающие в просвет матрикса. Они обладают АТФ-синтетичной (образует АТФ из АДФ) активностью. Транспорт электронов идет по дыхательной цепи, локализованной во внутренней мембране, которая содержит четыре крупных ферментных комплекса (цитохромы). При прохождении электронов по дыхательной цепи ионы водорода откачиваются из матрикса в перимитохондриальное пространство, что обеспечивает формирование протонного градиента (помпы). Энергия этого градиента (различия в концентрации веществ и формирование мембранного потенциала) используется для синтеза АТФ и транспорта метаболитов и неорганических ионов. Содержащиеся на внутренней мембране белки-переносчики транспортируют через нее органические фосфаты, АТФ, АДФ, аминокислоты, жирные кислоты, три — и дикарбоновые кислоты.
Энергия этого градиента (различия в концентрации веществ и формирование мембранного потенциала) используется для синтеза АТФ и транспорта метаболитов и неорганических ионов. Содержащиеся на внутренней мембране белки-переносчики транспортируют через нее органические фосфаты, АТФ, АДФ, аминокислоты, жирные кислоты, три — и дикарбоновые кислоты.
Наружная мембрана митохондрии более проницаема для низкомолекулярных веществ, так как в ней много гидрофильных белковых каналов. На наружной мембране располагаются специфические рецепторные комплексы, через которые белки из матрикса транспортируются в перимитохондриальное пространство.
По своему химическому составу и свойствам наружная мембрана близка к другим внутриклеточным мембранам и плазмолемме. В ней содержатся ферменты, метаболизирующие жиры, активирующие (катализирующие) превращения аминов, аминооксидаза. Если ферменты наружной мембраны сохраняют активность, то это показатель функциональной сохранности митохондрий.
В митохондриях имеются два автономных субкомпартмента.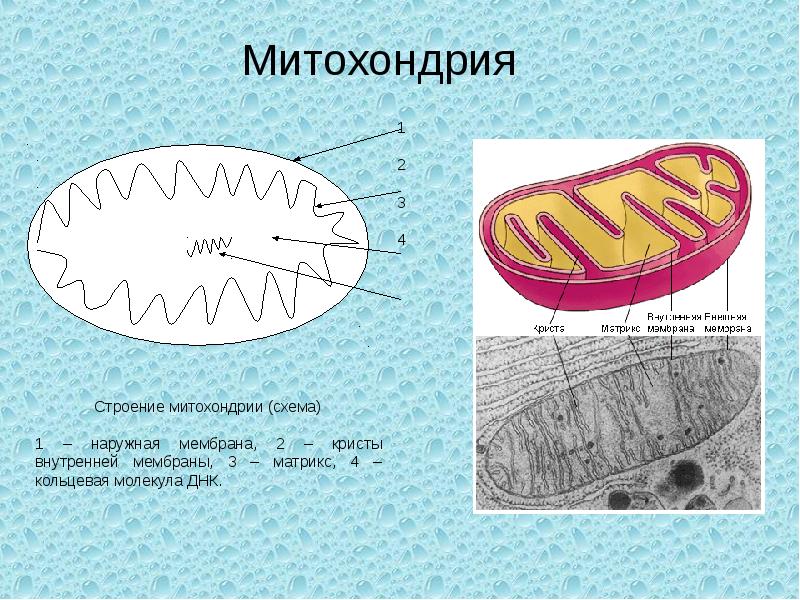 Вели перимитохондриальное пространство, или наружная камера митохондрии (внешний субкомпартмент), формируется за счет проникновения белковых комплексов гиалоплазмы, то внутренний субкомпартмент (матрикс митохондрии) частично образован за счет синтетической активности митохондриальной ДНК. Во внутреннем субкомпартменте (матриксе) содержатся ДНК, РНК и рибосомы. Он отличается высоким уровнем ионов Са 2+ в сравнении с гиалоплазмой. Во внешнем субкомпартменте накапливаются ионы водорода. Ферментативная активность внешнего и внутреннего субкомпартментов, состав белков сильно различаются. Внутренний субкомпартмент имеет более высокую электронную плотность, чем внешний.
Вели перимитохондриальное пространство, или наружная камера митохондрии (внешний субкомпартмент), формируется за счет проникновения белковых комплексов гиалоплазмы, то внутренний субкомпартмент (матрикс митохондрии) частично образован за счет синтетической активности митохондриальной ДНК. Во внутреннем субкомпартменте (матриксе) содержатся ДНК, РНК и рибосомы. Он отличается высоким уровнем ионов Са 2+ в сравнении с гиалоплазмой. Во внешнем субкомпартменте накапливаются ионы водорода. Ферментативная активность внешнего и внутреннего субкомпартментов, состав белков сильно различаются. Внутренний субкомпартмент имеет более высокую электронную плотность, чем внешний.
Специфические маркеры митохондрий — ферменты цитохром-оксидаза и сукцинатдегидрогеназа, выявление которых позволяет количественно охарактеризовать энергетические процессы в митохондриях.
Основная функция митохондрий — синтез АТФ. Вначале в гиалоплазме разрушаются сахара (глюкоза) до молочной и пировиноградной кислот (пирувата) с одновременным синтезом небольшого количества АТФ.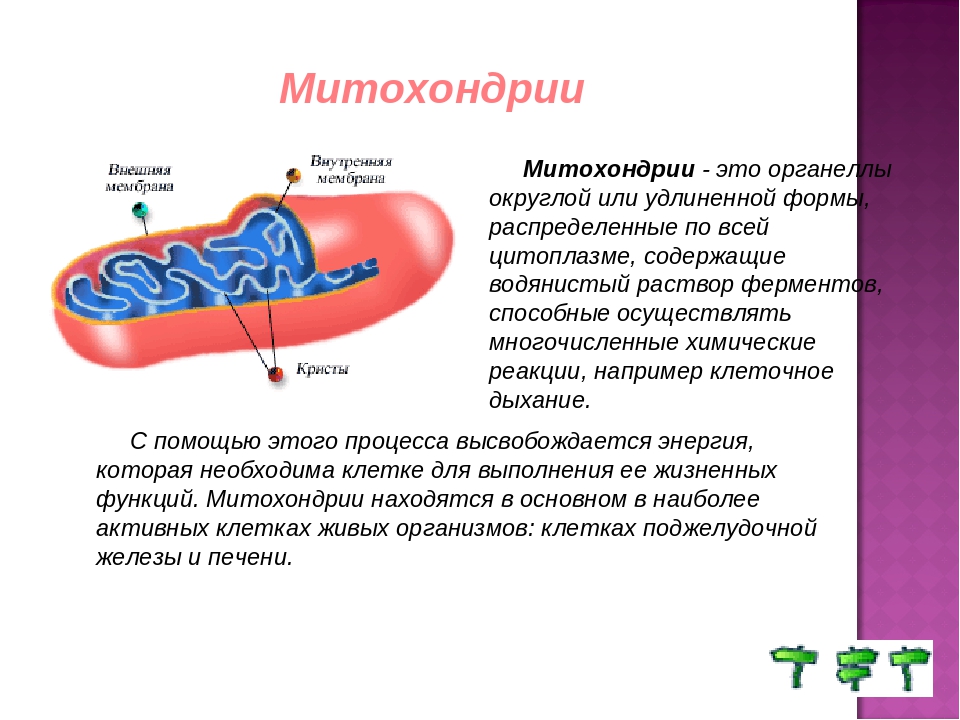 В результате гликолиза одной молекулы глюкозы используется две молекулы АТФ, а образуется четыре. Таким образом, положительный баланс составляют всего две молекулы АТФ. Эти процессы совершаются без кислорода (анаэробный гликолиз).
В результате гликолиза одной молекулы глюкозы используется две молекулы АТФ, а образуется четыре. Таким образом, положительный баланс составляют всего две молекулы АТФ. Эти процессы совершаются без кислорода (анаэробный гликолиз).
Все последующие этапы выработки энергии происходят в процессе аэробного окисления, который обеспечивает синтез большого количества АТФ. При этом органические вещества разрушаются до СO 2 и воды. Окисление сопровождается переносом протонов на их акцепторы. Эти реакции осуществляются с помощью ряда ферментов цикла трикарбоновых кислот, которые находятся в матриксе митохондрии.
В мембраны крист встроены системы переноса электронов и сопряженного с ним фосфорилирования АДФ (окислительное фосфорилирование). При этом происходит перенос электронов от одного белка-акцептора электронов к другому и, наконец, связывание их с кислородом, вследствие чего образуется вода. Одновременно с этим часть энергии, выделяемой при таком окислении в цепи переноса электронов, запасается в виде макроэргической связи при фосфорилировании АДФ, что приводит к образованию большого числа молекул АТФ — основного внутриклеточного энергетического эквивалента.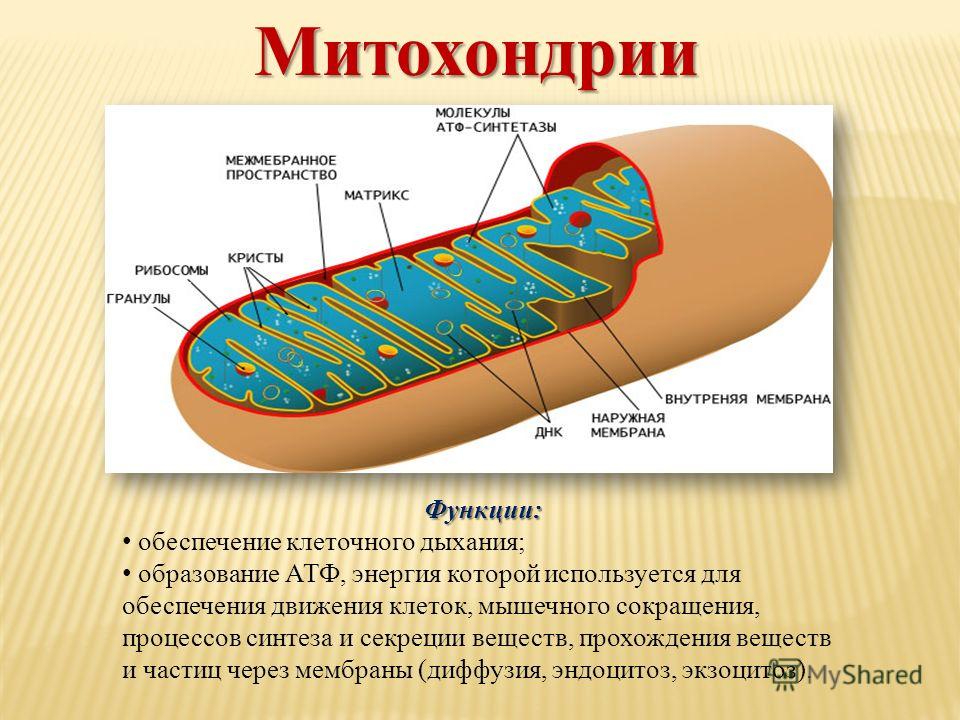 На мембранах крист митохондрий происходит процесс окислительного фосфорилирования с помощью расположенных здесь белков цепи окисления и фермента фосфорилирования АДФ АТФ-синтетазы. В результате окислительного фосфорилирования из одной молекулы глюкозы образуется 36 молекул АТФ.
На мембранах крист митохондрий происходит процесс окислительного фосфорилирования с помощью расположенных здесь белков цепи окисления и фермента фосфорилирования АДФ АТФ-синтетазы. В результате окислительного фосфорилирования из одной молекулы глюкозы образуется 36 молекул АТФ.
К некоторым гормонам и веществам на митохондриях имеются специализированные (аффинные) рецепторы. Трийодтиронин в норме ускоряет синтетическую активность митохондрий. Интерлейкин-1 и высокие концентрации трийодтиронина разобщают цепи окислительного фосфорилирования, вызывают набухание митохондрий, что сопровождается увеличением образования тепловой энергии.
Новые митохондрии образуются путем деления, перетяжкой или почкованием. В последнем случае образуется протомитохондрия, постепенно увеличивающаяся в размерах.
Протомитохондрия — мелкая органелла с наружной и внутренней мембранами. Внутренняя мембрана не имеет или содержит слаборазвитые кристы. Органелла характеризуется низким уровнем аэробного фосфорилирования.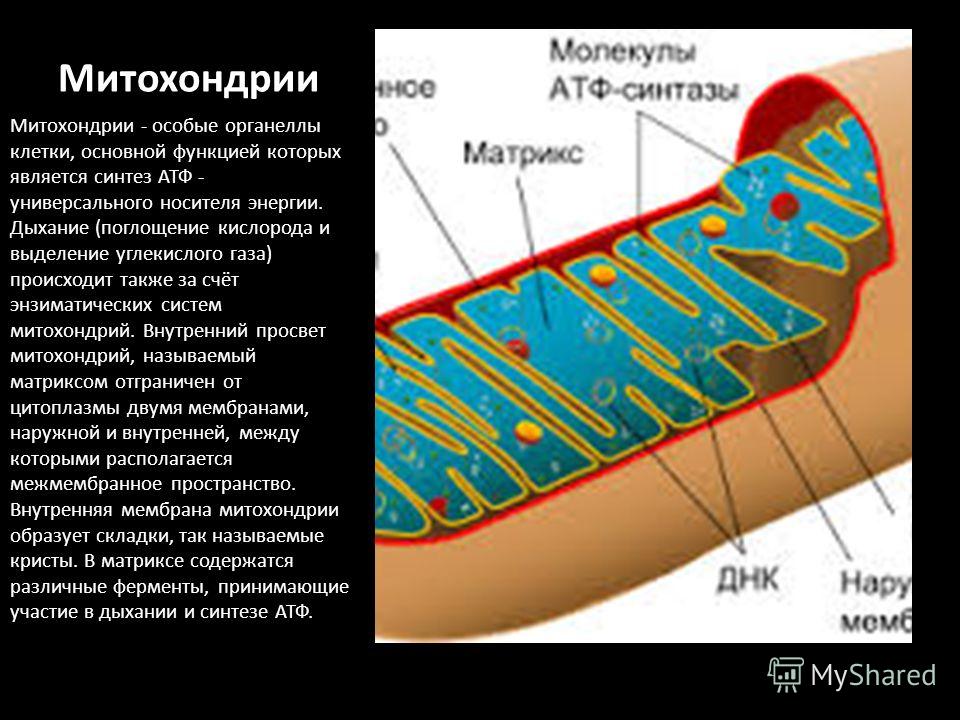 При образовании перетяжки содержимое митохондрии распределяется между двумя новыми довольно крупными органеллами. При любом способе размножения в каждой из вновь образующихся митохондрий имеется собственный геном.
При образовании перетяжки содержимое митохондрии распределяется между двумя новыми довольно крупными органеллами. При любом способе размножения в каждой из вновь образующихся митохондрий имеется собственный геном.
Старые митохондрии разрушаются путем аутолиза (самопереваривания клеткой с помощью лизосом) с образованием аутолизосом. Из аутолизосомы образуется остаточное тельце. При полном переваривании содержимое остаточного тельца, состоящее из низкомолекулярных органических веществ, выводится путем экзоцитоза. При неполном переваривании остатки митохондрий могут накапливаться в клетке в виде слоистых телец или гранул с нипофусцином. В части митохондрий накапливаются нерастворимые соли кальция с образованием кристаллов — кальцинатов. Накопление продуктов дегенерации митохондрий может привести к дистрофии клетки.
Митохондрии (МТ) – одно из самых интересных мне направлений исследований. Объединение митохондрий с другой клеткой в ходе эндосимбиоза около 1,6 млрд лет назад стало основной всех многоклеточных эукариотов со сложной структурой. Предположительно митохондрии произошли от клеток, напоминающих α-протеобактерии.
Предположительно митохондрии произошли от клеток, напоминающих α-протеобактерии.
Лучшее обзорное исследование последнего времени по митохондриях – работа Вернера Кулбрандта «Структура и функция митохондриальных белковых комплексов мембраны ». Если вы знаете английский язык и интересуетесь устройством этих органелл, то настоятельно рекомендую к прочтению. Эта статья так хороша, что может быть смело главой хорошего учебника по молекулярной биологии. Сначала я хотел перевести всю статью, но это бы заняло непростительно много времени и оторвало бы от других дел. Поэтому ограничусь тезисами и картинками. Периодически разбавляя все своими мыслями.
Митохондрия кодирует сама только 13 белков, не смотря на наличие отдельной от клетки ДНК (мтДНК) и всего «производственного» цикла по транскрипции белков. Изолированная митохондрия какое-то время может сохранять композицию и функционировать.
Рисунок 1. Компоненты мембраны митохондриона. Внешняя мембрана отделяет митохондрию от цитоплазмы. Она окружает внутреннюю мембрану, которая отделяет межмембранное пространство от богатого белками центрального матрикса. Внутреннюю мембрану разделяют на внутреннюю пограничную мембрану и кристы. Две эти части непрерывны в местах крепления крист (cristae junction ). Кристы простираются более или менее глубоко в матрикс и являются основным место митохондриального преобразования энергии. Небольшой протоновый градиент в межмембранном пространстве (pH 7,2-7,4) и матрикс (pH 7,9-8,0) приводят к образованию АТФ АТФ-синтазой в мембранах крист.
Она окружает внутреннюю мембрану, которая отделяет межмембранное пространство от богатого белками центрального матрикса. Внутреннюю мембрану разделяют на внутреннюю пограничную мембрану и кристы. Две эти части непрерывны в местах крепления крист (cristae junction ). Кристы простираются более или менее глубоко в матрикс и являются основным место митохондриального преобразования энергии. Небольшой протоновый градиент в межмембранном пространстве (pH 7,2-7,4) и матрикс (pH 7,9-8,0) приводят к образованию АТФ АТФ-синтазой в мембранах крист.
Внешняя мембрана пористая и позволяет веществам из цитоплазмы проходить через нее. Внутренняя мембрана плотная, для ее пересечения нужны транспортные белки [Гилберт Линг обоснованно не согласен], непрерывность барьера позволяет иметь внутренней мембране электрохимический потенциал в -180 mV. У матрикса довольно большой pH (7,9-8). Еще раз углублюсь в Линга. Щелочной (выше 7) pH способствует более развернутой конформации белков . Высокий pH нарушает водородные и солевые связи, делая поляризованные CO и NH доступными молекулам воды, там самым усиливая дипольный момент всей внутриклеточной воды и связывая ее. В этом ключе наличие мембраны нужно не для «удержания» протоплазмы внутри клетки (это делают сами белки при высоком pH), а для наличия потенциала.
Высокий pH нарушает водородные и солевые связи, делая поляризованные CO и NH доступными молекулам воды, там самым усиливая дипольный момент всей внутриклеточной воды и связывая ее. В этом ключе наличие мембраны нужно не для «удержания» протоплазмы внутри клетки (это делают сами белки при высоком pH), а для наличия потенциала.
мтДНК находится в нуклеотидах, которых примерно 1000 на клетку. Белковая плотность матрикса довольна высокая (до 500 мг/мл), что близко к кристаллизованным белкам.
Внутренняя мембрана образует инвагинации, называемые кристами, которые глубоко проникают в матрикс. Кристы определяют третий «отсек» митохондрий – просвет крист (cristae lumen). Мембраны кристы содержат большинство, если не все, полностью «собранные» комплексы цепи переноса электронов и АТФ-синтазы. Просвет кристы содержит большое количество маленького растворимого белкового переносчика электронов (цитохром с). Митохондриальные кристы, таким образом, основное место биологической конверсии энергии во всех не фотосинтетических эукариотах.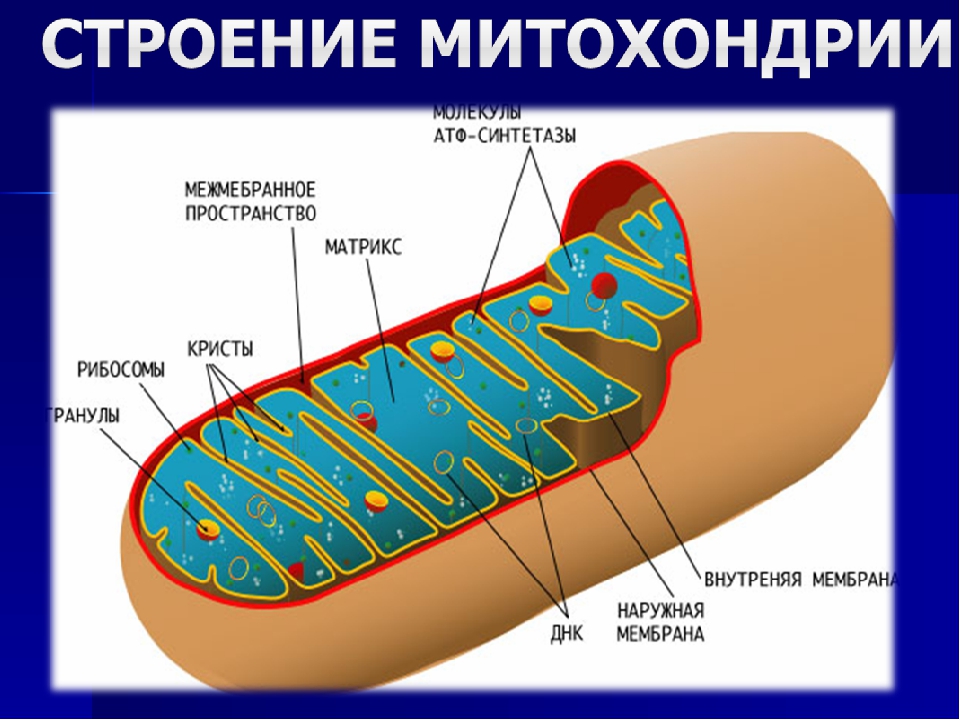
С кристами тоже много всего интересного. Оптические свойства кристы влияют на распространение и генерацию света в тканях. Я даже встречал идеи о том, что поверхность крист подобна (предположение) поверхностям топологических изоляторов (подразумевалась суперпроводимость без диссипации заряда).
Рисунок 2. Мембранные белковые комплексы дыхательной цепи. Комплекс I (NADH / убихинон оксидоредуктаза, синий), Комплекс II (сукцинат дегидрогеназа, розовы), Комплекс III (цитохром С редуктаза, оранжевый), Комплекс IV (цитохром С оксидаза, зеленый) и митохондриальная АТФ синтаза (известная как комплекс V , бежевая) работают вместе во время окислительного фосфорилирования, чтобы клетки могли использовать энергию. Комплексы I , III , IV выкачивают протоны вдоль мембраны кристы, создавая протоновый градиент, стимулирующий синтез АТФ.
Теперь немного внимания на комплекс II. Вы помните, что жир (кето) делает упор в метаболизме на FADh3 и комплекс II. Они восстанавливают пару CoQ, в какой-то момент окисленного CoQ не хватает для транспортировки электронов на комплекс III и образует обратный поток электронов на комплекс I с образованием супероксида. При долгом HFLC-питании комплекс I будет обратимо разрушен, при этом это нормальная физиологическая оптимизация.
Вы помните, что жир (кето) делает упор в метаболизме на FADh3 и комплекс II. Они восстанавливают пару CoQ, в какой-то момент окисленного CoQ не хватает для транспортировки электронов на комплекс III и образует обратный поток электронов на комплекс I с образованием супероксида. При долгом HFLC-питании комплекс I будет обратимо разрушен, при этом это нормальная физиологическая оптимизация.
Еще прошу вас заметить, что комплекс II не выкачивает протоны. Что у нас рассеивает протоновый градиент, нарушает фосфорилирование и стимулирует сжигание жиров на тепло? Правильно, стресс холода. Термогенез связан с метаболизмом через комплекс, который не выкачивает протоны, тем самым не давая дополнительных протонов для АТФ-синтазы. Можно только удивляться как замечательно у нас продуман организм.
Крепления крист и MICOSМеста крепления крист (cristae junctions) – маленькие круглы отверстия примерно 25 нм диаметром. В митохондриях всех организмов есть система MICOS (mitochondria contact site and cristae to outer membrane), сборка из пяти мембранных и одного растворимого белков, прикрепляющих кристы к наружной мембране.
В клетках с повышенной потребностью в энергии, такие как скелетные и сердечные мышцы, кристы плотно заполняют большую часть объема митохондрии. В тканях с меньшими потребностями в энергии, таких как печень и почки, кристы находятся не так плотно по отношению друг к другу. Остается больше места в матрице для биосинтетических ферментов.
Рисунок 3. Томографический объем митохондрии сердца мыши. А) Трехмерный объем митохондрии сердца мыши, снятый cryo-ET. Наружная мембрана (серая) окутывает внутреннюю мембрану (светло-синяя). Внутренняя мембрана плотно наполнена кристами б) Томографический срез объема. Плотно заполненный матрикс, содержащий большую часть митохондриальных белков, выглядит темным на электронном микроскопе. В то время как межмембранное пространство и просветы крист выглядят светлыми из-за низкой концентрации белков.
Димеры АТФ синтазыМитохондриальная F1-F0 АТФ синтаза является самым заметным белковым комплексом кристы. АТФ синтаза – это древняя наномашина, которая использует электрохимический протновых градиент вокруг внутренней мембраны для создания АФТ посредством вращательного катализа
.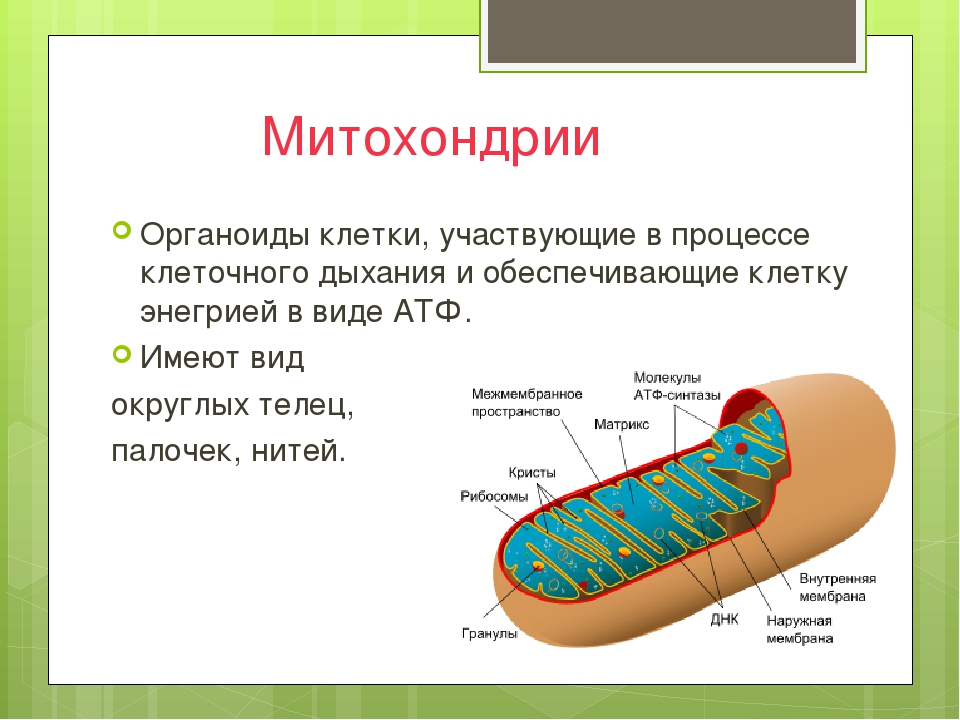 Протоны, двигающиеся через F0 комплекс мембраны, вращают ротор из 8 (у млекопитающих) или 10 (у дрожжей) с-узлов. Центральный стебель передает крутящий момент c-ротора каталитической головке F1, где АТФ образуется из АДФ и фосфата через последовательность конформационных изменения. Периферийный стебель предотвращает непродуктивное вращение Головы F1 против комплекса F0.
Протоны, двигающиеся через F0 комплекс мембраны, вращают ротор из 8 (у млекопитающих) или 10 (у дрожжей) с-узлов. Центральный стебель передает крутящий момент c-ротора каталитической головке F1, где АТФ образуется из АДФ и фосфата через последовательность конформационных изменения. Периферийный стебель предотвращает непродуктивное вращение Головы F1 против комплекса F0.
Многие годы считалось, что АТФ синтаза случайным образом располагается на внутренней мембране. Но оказалось, что АТФ синтаза располагается двойными рядами . Причем линейные ряды АТФ синтазы – фундаментальный атрибут всех живых митохондрий.
Рисунок 4. Двойные ряды АТФ синтазы у семи разных видов.
Ряды АТФ синтазы располагаются в основном вдоль хребтов крист. Димеры изгибают липидный бислой и как следствие само-организуются в ряды. Когда у митохондрий дрожжей выбивали узлы e и g АФТ синтазы, то штамм рос на 60% медленней диких собратьев, и потенциал мембран их митохондрий был снижен вдвое.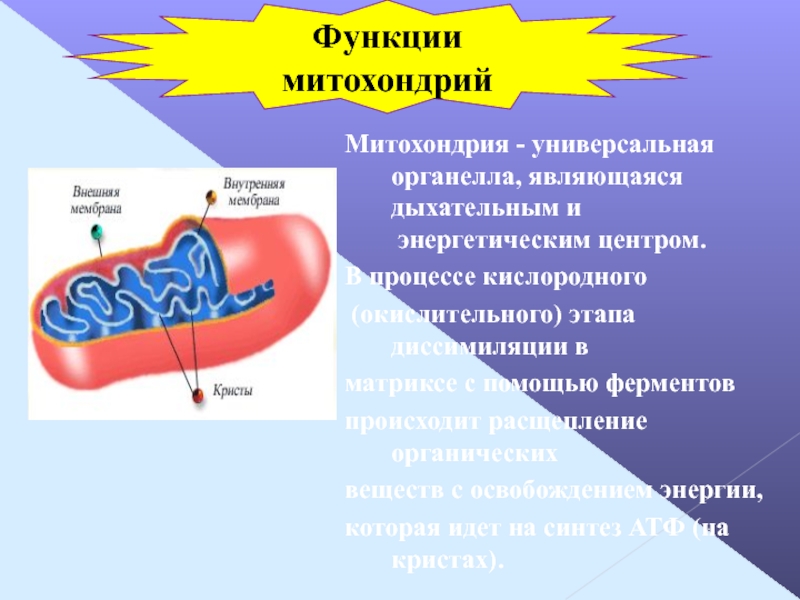 У АФТ синтазы прокариотов недостает нескольких узлов, связанных с димерами, ряды димеров не были найдены у бактерий и архей. Кристы и ряды димеров АФТ синтазы, таким образом, являются адаптацией к большим энергетическим потребностям организма.
У АФТ синтазы прокариотов недостает нескольких узлов, связанных с димерами, ряды димеров не были найдены у бактерий и архей. Кристы и ряды димеров АФТ синтазы, таким образом, являются адаптацией к большим энергетическим потребностям организма.
Рисунок 5. Структура димера АТФ синтазы митохондрии polymella sp. Вид сбоку на V-образный димер АТФ синтазы.
Комплексы и суперкомплексы дыхательной цепиПротоновый градиент вокруг внутренней мембраны создается тремя крупными мембранными комплексами, известными как комплекс I, комплекс III и комплекс IV (см. рисунок 2). Комплекс I кормится электронами из NADH, высвобождаемая при передаче электрона энергия выкачивает четыре протона. Комплекс III получает электрон от восстановленного хинола и передает его носителю электронов (цитохрому с), выкачивая в процессе один протон. Комплекс IV получает электрон из цитохрома с и передает его молекулярному кислороду, выкачивая 4 протона за каждую молекулу кислорода, превращенную в воду.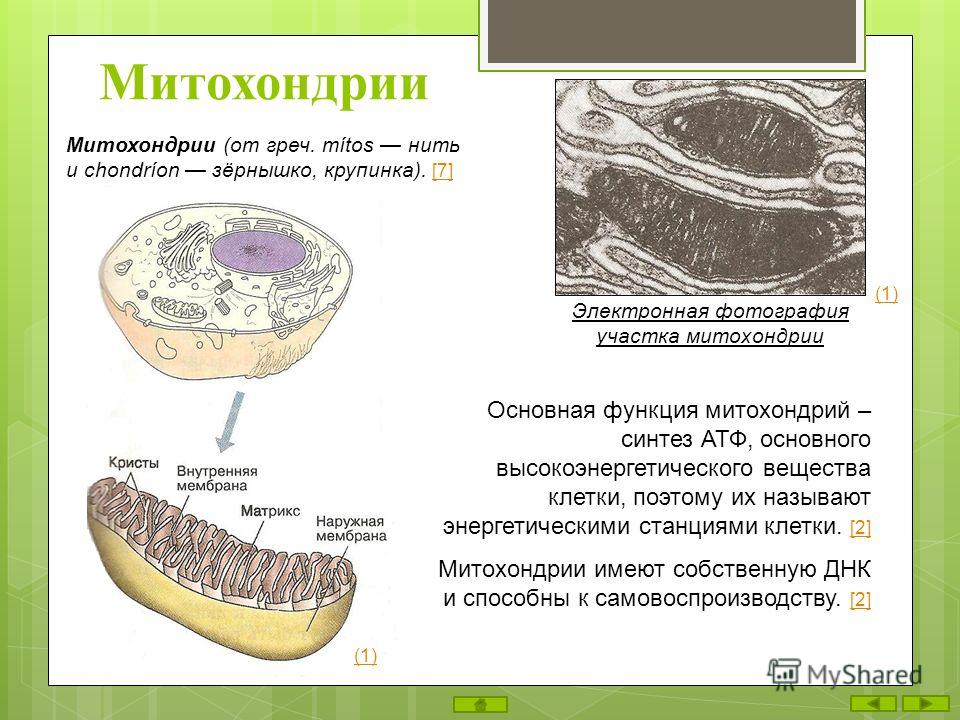 Комплекс II не выкачивает протоны, напрямую передавая электроны хинолу. Как перенос электронов из NADH в хинол связан с транслокацией протонов пока не ясно. Комплекс I – крупнее III и IV вместе взятых.
Комплекс II не выкачивает протоны, напрямую передавая электроны хинолу. Как перенос электронов из NADH в хинол связан с транслокацией протонов пока не ясно. Комплекс I – крупнее III и IV вместе взятых.
Рисунок 6. Комплекс I митохондрии коровьего сердца. Матриксная часть содержит ряд из восьми железно-серных (Fe-S) кластеров, которые направляют электроны из NADH в хинол на пересечении матрикса и мембраны. Мембранная часть состоит из 78 лопастей, включая выкачивающие протоны молекулы.
Комплексы I, III и IV соединяются в суперкомплексы или респирасомы. У пекарских дрожжей (saccharomyces cerevisiae ) нет комплекса I, их суперкомплексы состоят из III и IV. Роль суперкомплексов пока еще не ясна. Предполагают, что это делает транспорт электронов более эффективным, но прямых доказательств этому пока нет.
Рисунок 7. Суперкомплекс митохондрии коровьего сердца. Обратите внимание на дистанцию между комплексами I и III, который надо проделать хинолу. Стрелки – движения электрона в суперкомплексе.
Стрелки – движения электрона в суперкомплексе.
Основным белком просвета кристы является цитохром с, который переносит электрон из комплекса III в комплекс IV. Если цитохром с высвобождается в цитоплазму клетки, то вызывает апоптоз .
Рисунок 8. Ряды димеров АТФ синтазы задают форму кристам. У хребта кристы АФТ синтаза (желтый) образует слив для протонов (красный), протоновые насосы электронной цепи (зеленый) находятся по обоим сторона рядов димеров. Направляя протоны от источника к АТФ синтазе, кристы работают как протоновые направляющие, позволяющие эффективное производство АТФ. Красные стрелки показывают направление потока протонов.
Реорганизация мембраны во время старенияСтарение – фундаментальный и плохо понимаемый процесс всех эукариотов. Исследовали старение митохондрий на грибах Podospora anserina , которые живут всего 18 дней. В нормальной митохондрии кристы проникают глубоко в матрикс. Для этого нужны ряды димеров АТФ синтазы и MICOS комплекс у мест крепления крист. С возрастом кристы начинают все ближе подходить к поверхности мембраны, димеры АФТ синтазы превщаются в мономеры, и все заканчивается высвобождением цитохрома с и клеточной смертью.
С возрастом кристы начинают все ближе подходить к поверхности мембраны, димеры АФТ синтазы превщаются в мономеры, и все заканчивается высвобождением цитохрома с и клеточной смертью.
Транспорт электронов создает супероксид в комплексах I и III. Это побочный продукт метаболизма. Одновременно необходимый и смертельно опасный. Во время старения деление (fission) начинает превалировать над сращением (fussion). Это не дает поврежденным митохондриям «спастись» путем сращения и ускоряет неизбежное.
Рисунок 9. Изменения морфологии внутренней мембраны и димеров АТФ синтазы во время старения митохондрии.
Как видите, полей для будущих исследований очень много. Я предполагаю стык физики и биологии, где физики будут пытаются объяснить почему такая структура более энергетически эффективна. Тем более публикации по квантовой биологии
 Вопросы для самоподготовки к освоению данной темы:
Вопросы для самоподготовки к освоению данной темы: Микроскопический анализ постоянного препарата «Лизосомы»
Микроскопический анализ постоянного препарата «Лизосомы»Митохондрии — это структуры палочковидной или овальной формы (греч. mitos — нить, chondros — гранула). Они обнаружены во всех животных клетках (исключая зрелые эритроциты): у высших растений, у водорослей и простейших. Отсутствуют они только у прокариот бактерий.
Эти органеллы впервые были обнаружены и описаны в конце прошлого столетия Альтманом. Несколько позже эти структуры были названы митохондриями. В 1948 г. Хогебум указал на значение митохондрий как центра клеточного дыхания, а в 1949 г. Кеннеди и Ленинджер установили, что в митохондриях протекает цикл окислительного фосфорилирования. Так было доказано, что митохондрии служат местом генерирования энергии.
Митохондрии видны в обычном световом микроскопе при специальных методах окраски. В фазово — контрастном микроскопе и в «темном поле» их можно наблюдать в живых клетках.
Строение, размеры, форма митохондрий очень вариабельны. Это зависит в первую очередь от функционального состояния клеток. Например, установлено, что в мотонейронах мух, летающих непрерывно 2 часа, проявляется огромное количество шаровидных митохондрий, а у мух со склеенными крыльями число митохондрий значительно меньше и они имеют палочковидную форму (Л. Б. Левинсон). По форме они могут быть нитевидными, палочковидными, округлыми и гантелеобразными даже в пределах одной клетки.
Митохондрии локализованы в клетке, как правило, либо в тех участках, где расходуется энергия, либо около скоплений субстрата (например, липидных капель), если таковые имеются.
Строгая ориентация митохондрий обнаруживается вдоль жгутиков сперматозоидов, в поперечно-полосатой мышечной ткани, где они располагаются вдоль миофибрилл, в эпителии почечных канальцев локализуются во впячиваниях базальной мембраны и т.д.
Количество митохондрий в клетках имеет органные особенности, например, в клетках печени крыс содержится от 100 до 2500 митохондрий, а в клетках собирательных канальцев почки — 300, в сперматозоидах различных видов животных от 20 до 72, у гигантской амебы Chaos chaos их число достигает 500 000. Размеры митохондрий колеблются от 1 до 10 мкм.
Ультрамикроскопическое строение митохондрий однотипно, независимо от их формы и размера. Они покрыты двумя липопротеидными мембранами: наружной и внутренней. Между ними располагается межмембранное пространство.
Впячивания внутренней мембраны, которые вдаются в тело митохондрий, называются кристами . Расположение крист в митохондриях может быть поперечным и продольным. По форме кристы могут быть простыми и разветвленными. Иногда они образует сложную сеть. В некоторых клетках, например, в клетках клубочковой зоны надпочечника кристы имеют вид трубочек. Количество крист прямо пропорционально интенсивности окислительных процессов, протекающих в митохондриях. Например, в митохондриях кардиомиоцитов их в несколько раз больше, чем в митохондриях гепацитов. Пространство, ограниченное внутренней мембраной, составляет внутреннюю камеру митохондрий. В нем между кристами находится митохондриальный матрикс — относительно электронно плотное вещество.
Белки внутренней мембраны синтезируются миторибосомами, а белки внешней мембраны — циторибосомами.
«Наружная мембрана митохондрий по многим показателям сходна с мембранами ЭПС. Она бедна окислительными ферментами. Немного их и в мембранном пространстве. Зато внутренняя мембрана и митохондриальный матрикс буквально насыщены ими. Так, в матриксе митохондрий сосредоточены ферменты цикла Кребса и окисления жирных кислот. Во внутренней мембране локализована цепь переноса электронов, ферменты фосфорилирования (образования АТФ из АДФ), многочисленные транспортные системы.
Кроме белка и липидов, в состав мембран митохондрий входит РНК, ДНК, последняя обладает генетической специфичностью, и по своим физико-химическим свойствам отличается от ядерной ДНК.
При электронно-микроскопических исследованиях обнаружено, что поверхность наружной мембраны покрыта мелкими шаровидными элементарными частицами. Внутренняя мембрана и кристы содержат подобные элементарные частицы на «ножках», так называемые грибовидные тельца. Они -состоят из трех частей: головки сферической формы (диаметр 90-100 А°), ножки цилиндрической формы, длиной 5 нм и шириной 3-4 нм, основания, имеющего размеры 4 на 11 нм. Головки грибовидных телец связаны с фосфорилированием, затем обнаружено, что головки содержат фермент, обладающий АТФ-идной активностью.
В межмембранном пространстве находится вещество, обладающее более низкой электронной плотностью, чем матрикс. Оно обеспечивает сообщение между мембранами и поставляет для ферментов, находящихся в обеих мембранах, вспомогательные катализаторы-коферменты.
В настоящее время известно, что наружная мембрана митохондрий хорошо проницаема для веществ, имеющих низкий молекулярный вес, в частности, белковых соединений. Внутренняя мембрана митохондрий обладает избирательной проницаемостью. Она практически непроницаема для анионов (Cl -1 , Br -1 , SO 4 -2 , HCO 3 -1 , катионов Sn +2 , Mg +2 , ряда cахаров и большинства аминокислот, тогда как Са 2+ , Мп 2+ , фосфат, многокарбоновые кислоты легко проникают через нее. Имеются данные о наличии во внутренней мембране нескольких переносчиков, специфических к отдельным группам проникающих анионов и катионов. Активный транспорт веществ через мембраны осуществляется благодаря использованию энергии АТФ-азной системы или электрического потенциала, генерируемого на мембране в результате работы дыхательной цепи. Даже АТФ, синтезированная в митохондриях, может выйти с помощью переносчика (сопряженный транспорт).
Матрикс митохондрий представлен мелкозернистым электронно-плотным веществом. В нем располагаются миторибосомы, фибриллярные структуры, состоящие из молекул ДНК и гранул, имеющих диаметр более 200А ◦ образованные солями: Ca 3 (PO 4) , Ba 3 (PO 4) 2 , Mg 3 (PO 4) . Полагают, что гранулы служат резервуаром ионов Са +2 и Мg +2 . Их количество увеличивается при изменении проницаемости митохондриальных мембран.
Присутствие в митохондриях ДНК обеспечивает участие митохондрий в синтезе РНК и специфических белков, а также указывает на существование цитоплазматической наследственности. Каждая митохондрия содержит в зависимости от размера одну или несколько молекул ДНК (от 2 до 10). Молекулярный вес митохондриальной ДНК около (30-40)*10 6 у простейших, дрожжей, грибов. У высших животных около (9–10) *10 6.
Длина ее у дрожжей примерно равна 5 мкм, у растений — 30 мкм. Объем генетической информации, заключенный в митохондриальной ДНК, невелик: он состоит из 15-75 тыс. пар оснований, которые могут кодировать в среднем 25-125 белковых цепей с молекулярным весом около 40000.
Митохондриальная ДНК отличается от ядерной ДНК рядом особенностей: более высокой скоростью синтеза (в 5-7 раз), она более устойчива к действию ДНК-азы, представляет собой двухкольцевую молекулу, содержит больше гуанина и цитозина, денатурируется при более высокой температуре и легче восстанавливается. Однако не все митохондриальные белки синтезируются митохондриальной системой. Так, синтез цитохрома С и других ферментов обеспечивается информацией, содержащейся в ядре. В матриксе митохондрий локализованы, витамины А, В 2 , В 12 , К, Е, а также гликоген.
Функция митохондрий заключается в образовании энергии, необходимой для жизнедеятельности клеток. Источником энергии в клетке могут служить различные соединения: белки, жиры, углеводы. Однако единственным субстратом, который немедленно включается в энергетические процессы, является глюкоза.
Биологические процессы, в результате которых в митохондриях образуется энергия, можно подразделить на 3 группы: I группа — окислительные реакции, включающие две фазы: анаэробную (гликолиз) и аэробную. II группа — дефосфорилирование, расщепление АТФ и высвобождение энергии. III группа — фосфорилирование, сопряженное с процессом окисления.
Процесс окисления глюкозы вначале происходит без участия кислорода (анаэробным или гликолитическим путем) до пировиноградной или молочной кислоты.
Однако при этом энергии выделяется лишь небольшое количество. В дальнейшем эти кислоты вовлекаются в процессы окисления, которые протекают с участием кислорода, т. е. являются аэробными. В результате процесса окисления пировиноградной и молочной кислоты, названной циклом Кребса, образуется углекислый газ, вода и большое количество энергии.
Образующаяся энергия не выделяется в виде тепла, что привело бы к перегреванию клеток и гибели всего организма, а аккумулируется в удобной для хранения и транспорта форме в виде аденозинтрифосфорной кислоты (АТФ). Синтез АТФ происходит из АДФ и фосфорной кислоты и вследствие этого называется фосфорилированием .
В здоровых клетках фосфорилирование сопряжено с окислением. При заболеваниях сопряженность может разобщаться, поэтому субстрат окисляется, а фосфорилирование не происходит, и окисление переходит в тепло, а содержание АТФ в клетках снижается. В результате повышается температура и падает функциональная активность клеток.
Итак, основная функция митохондрий заключается в выработке практически всей энергии клетки и происходит синтез компонентов, необходимых для деятельности самого органоида, ферментов «дыхательного ансамбля», фосфолипидов и белков.
Еще одной стороной деятельности митохондрий является их участие в специфических синтезах, например, в синтезе стероидных гормонов и отдельных липидов. В ооцитах разных животных образуются скопления желтка в митохондриях, при этом они утрачивают свою основную систему. Отработавшие митохондрии могут накапливать также продукты экскреции.
В некоторых случаях (печень, почки) митохондрии способны аккумулировать вредные вещества и яды, попадающие в клетку, изолируя их от основной цитоплазмы и частично блокируя вредное действие этих веществ. Таким образом, митохондрии способны брать на себя функции других органоидов клетки, когда это требуется для полноценного обеспечения того или иного процесса в норме или в экстремальных условиях.
Биогенез митохондрий. Митохондрии представляют собой обновляющиеся структуры с довольно кратким жизненным циклом (в клетках печени крысы, например, период полужизни митохондрий охватывает около 10 дней). Митохондрии образуются в результате роста и деления предшествующих митохондрий. Деление их может происходить тремя способами: перетяжкой, отпочковыванием небольших участков и возникновением дочерних митохондрий внутри материнской. Делению (репродукции) митохондрий предшествует репродукция собственной генетической системы — митохондриальной ДНК.
Итак, согласно взглядам большинства исследователей, образование митохондрий происходит преимущественно путем саморепродукции их de novo.
Митохондрии строение
Цитоплазмой (протоплазмой) называется все содержимое клетки, за исключением ядра и оболочки. Термин «цитоплазма» был предложен в 1882 г. Э. Страсбургером. По своему значению он более точно указывает на то, что речь идет именно о плазме клетки, а не обо всем содержимом клетки — протопласте, или протоплазме в ее широком понимании. В молодой растительной клетке цитоплазма занимает большую часть ее объема. В эмбриональных клетках растений и животных цитоплазма отличается слабо развитой системой внутриклеточных мембран, почти полностью состоит из гиалоплазмы (основного матрикса) и рибосом. В процессе эволюции клетки возникли внутриклеточные мембраны, а также некоторые клеточные органеллы, например митохондрии, пластиды и центри-оли, составляющие большую часть цитоплазмы (рис. 9).[ …]
Строение хлоропластов несколько напоминает строение митохондрий, Их тело (матрикс, или строма) ограничено двумя белково-липидными мембранами, толщиной 7 нм каждая. Наружный слой, соприкасающийся с цитоплазмой, обычно имеет ровные контуры и не образует выпячиваний или складок, внутренний, как и у других пластид, образует складчатые выпячивания внутрь стромы.[ …]
Митохондрии — энергетический центр клетки. Под световым микроскопом они выглядят как мелкие плотные зернышки, но под электронным микроскопом видно их сложное складчатое внутреннее строение. Митохондрии состоят из белков и фосфолипидов. Все известные функции митохондрий связаны с активностью ферментов, в свою очередь связанной с окислительным обменом веществ. Эта активность проявляется благодаря образованию аденозин-трифосфата (АТФ) — носителя энергии.[ …]
Строение хвоста спермиев показано на рис. 24. В нем нет «грубых» фибрилл и митохондрий.[ …]
Митохондрии — мелкие тельца округлой или продолговатой формы, размером 0,5 — 1,5 мк, т. е. величиной с бактерию. Число их в клетке обычно велико, порядка 100—3000. Бывают, однако, клетки и с малым количеством митохондрий. Так, в спермии морской водоросли фукуса содержится всего 4 митохондрии, а в одноклеточной водоросли микромо-нас — одна. Митохондрии видны иод световым микроскопом, однако их тонкое строение можно изучать лишь с помощью электронного микроскопа (табл. 10, схема строения — рис. 31). Митохондрии — это образования, построенные из липопротеиновых мембран, погруженных в основное вещество — матрикс. Оболочка митохондрии образована двумя мембранами, между которыми имеется промежуток.[ …]
Вверху и в середине — вид продольного среза через митохондрию (вверху — митохондрия из эмбриональной клетки кончика корян; в середине — из клетки взрослого листа элодеи). В я и о у — трехмерная схема, на которой часть митохондрии срезана, что позволяет видеть ее внутреннее строение. 1 — наружная мембрана; г — внутренняя мембрана; з — кристы; 4 — матрикс.[ …]
Электронная микрофотография среза нерва пятого членика лапки таракана приведена на рис. 1 [15]. Его строение, вероятно, подобно строению аналогичного нерва саранчи [1]. Хойл [18] показал, что вокруг нерва лапки саранчи присутствует мембрана трахеолы. Однако Аш-хурст [1] недавно доказал ее отсутствие у саранчи. К настоящему времени не найдено анатомических различий, позволяющих объяснить разницу в проникновении иона калия в нервную систему саранчи и таракана. Может быть, существует различие в действии, регулирующем перенос иона, который, по-видимому, происходит в богатом митохондрией слое клеток невроглии, лежащем ниже оболочки соединительной ткани. Энзимное действие (обычно связанное с митохондрией) в оболочке нерва может привести к пслупнтоксикации насекомого, но только в том случае, если повреждаемые ткани всех насекомых одинаково защищены протиз проникновения иона.[ …]
Нуклеиновые кислоты очень высокополимерны, их раствор, так же как и раствор белка, имеет коллоидальный характер. Полимерные молекулы нуклеиновых кислот состоят из различных по строению и свойствам фрагментов, расположение и состав которых в молекуле нуклеиновой кислоты могут быть различными. В настоящее время доказано, что нуклеиновые кислоты необходимы для осуществления в клетках процессов роста, формообразовательных процессов, построения внутриклеточных структур (ядра, пластид, митохондрий, гранул). В работах с растительными объектами показано, что молодые растущие ткани всегда содержат большее количество нуклеиновых кислот. Изучены многие структурные особенности нуклеиновых кислот растений.[ …]
Все содержимое клетки, за исключением ядра и клеточной стенки, называется цитоплазмой. В жидкой, бесструктурной фазе цитоплазмы (матриксе) находятся рибосомы, мембранные системы, митохондрии, пластиды и другие структуры, а также запасные питательные вещества. Цитоплазма обладает чрезвычайно сложной, тонкой структурой (слоистая, гранулярная). С помощью электронного микроскопа раскрыты многие интересные детали строения клетки.[ …]
Итак, ядерная мембрана представляет собой полый мешок, отделяющий содержимое ядра от цитоплазмы, и состоит из двух слоев: внешний слой ограничивает перинуклеарное пространство снаружи, т. е. со стороны цитоплазмы, внутренний—изнутри, т. е. со стороны ядра. Из всех внутриклеточных мембранных компонентов подобным строением мембран обладают ядро, митохондрии и пластиды. Морфологическое строение каждого слоя такое же, как и внутренних мембран цитоплазмы. Отличительная особенность ядерной оболочки — наличие в пей пор — округлых перфораций, образующихся в местах слияния внешней и внутренней ядерных мембран (рис. 35).[ …]
Исключительное совершенствование отмечается в организации половых клеток, особенно мужских. У большинства беспозвоночных половые клетки обладают структурными компонентами, характерными для одноклеточных простейших и соматических клеток многоклеточных. Имеются центриоли, митохондрии и жгутик с аксоне-мой. У организмов, приспособившихся к жизни на суше, после развития внутреннего оплодотворения сперматозоиды развились в вытянутые структуры, а в их хвосте появились дополнительные цитоскелетные элементы. У высших наземных организмов развиваются разные формы подвижности сперматозоидов и разные типы акросом, что зависит от строения яйцевых оболочек.[ …]
Бактерии и сине-зеленые водоросли (циа-неи) — две филогенетически родственные группы — резко отличаются от всех остальных живых существ (в том числе от грибов) отсутствием истинного ядра и тем, что ДНК лежит в их клетке свободно, погруженная в так называемую нуклеоплазму, которая не отделена от цитоплазмы ядерной мембраной. У них нет также митохондрий и сложных жгутиков. Жгутики у них (когда они имеются) устроены проще и имеют принципиально иное строение, чем у остальных организмов; их клеточная стенка состоит из гетерополимерного вещества му-реина, которое не было обнаружено ни у одной другой группы организмов. Эти организмы называют прокариотами (Ргосагуо-1а — доядерные). У всех остальных организмов, как одноклеточных, так и многоклеточных, имеется настоящее ядро, окруженное ядерной мембраной и тем самым резко отграниченное от цитоплазмы. Такие организмы называют эукариотами (Еисагуо1а — ядерные). Кроме ясно дифференцированного ядра и цитоплазмы, у них имеются также митохондрии, а у многих также пластиды и сложные жгутики. Постепенно стало выясняться, что различия между прокариотами и эукариотами гораздо более глубокие и фундаментальные, чем, например, различия между высшими животными и высшими растениями (те и другие—эукариоты).[ …]
Крайне малые размеры клеток являются характерной, но не главной особенностью бактерий. Все бактерии представлены особым типом клеток, лишенных истинного ядра, окруженного ядерной мембраной. Аналогом ядра у бактерий является нуклеоид — ДНК-содержащая плазма, не отграниченная от цитоплазмы мембраной. Кроме того, для бактериальных клеток характерны отсутствие митохондрий, хлоропла-стов, а также особое строение и состав мембранных структур и клеточных стенок. Организмы, в клетках которых отсутствует истинное ядро, называются прокариотами (доядер-ными) или протоцитами (т. е. организмами с примитивной организацией клеток).[ …]
С другой стороны, имеются сведения о чрезвычайно раннем развитии и, видимо, постоянном наличии интерстициальных компонентов типа клеток Лейдига в семенниках у форели (Oota, Yamamoto, 1966). Они обладают хорошо развитым эндоплазматическим ретикулумом, внутренние мембраны митохондриев имеют характерную трубчатую или пузырчатую структуру.[ …]
В соответствии с другой гипотезой считают, что эукариотическая клетка произошла от прокариотической клетки, содержавшей несколько геномов, прикрепленных к клеточной мембране. В результате инвагинаций клеточной мембраны образовывались мезо-сомы, способные первоночально к фотосинтезу. Однако в дальнейшем произошла специализация этих органелл, в результате чего одна из них, утратив дыхательную и фотосинтетическую функцию, развилась в ядро, другие, наоборот, развив эти функции, стали митохондриями у животных и пластидами у растений. В пользу этой гипотезы свидетельствуют данные о двойном строении мембран ядра, митохондрий и пластид.[ …]
У растительноядных рыб различают три основных качественных состояния икры, которые наблюдаются в период разведения. Это икра недостаточно зрелая, продуцируемая самками обычно в начале сезона разведения или поздносозревающими особями, икра, постовуляторно перезревшая в результате ее запоздалого получения после овуляции, и икра, перезревшая в фолликулярной оболочке при затянувшемся сезоне разведения или содержании самок в воде с повышенной температурой. Каждое из этих состояний характеризуется комплексом особенностей строения компонентов клетки, различимых с помощью как электронного, так и светового микроскопа. Для недостаточно зрелой икры характерны более мелкие гранулы желтка, узкий слой периферической цитоплазмы, малое число и меньшие размеры лежащих в нем кортикальных альвеол. При постовуляторном перезревании происходит перемещение крупных гранул желтка в анимальную часть ооцита, слияние их в большие глыбки, между которыми располагается цитоплазма. Кортикальные альвеолы уплощаются, становясь на срезах вытянутыми параллельно оболочке ооцита. Существенно нарушается ультраструктура ооцитов — теряется четкость мембранных структур, митохондрии приобретают большую электронную плотность, кристы в них становятся менее выраженными или нечеткими. Везикулы эндоплазматической сети и митохондрии теряют четкие очертания, их мембраны становятся более извилистыми. В отдельных участках цитоплазмы мембраны органелл и сами органеллы могут исчезнуть. Существенно меняется и структура оболочки. При перезревании икры в фолликуле деструктивные изменения способного еще овулировать ооцита близки к тем, которые наблюдаются при постовуляторном перезревании [10].[ …]
Основным прибором цитологических исследований является световой микроскоп, до сих пор не утративший своего значения при изучении клетки. Существуют самые разнообразные модели световых микроскопов. Для каждого способа микроскопирова-ния необходимы свои методы приготовления препаратов. При изучении клетки под световым микроскопом многие ее структурные компоненты остаются незамеченными. Кроме того, при этом методе исследования живую клетку приходится обычно фиксировать (умерщвлять), дифференцированно ее окрашивать для выделения отдельных структур, что позволяет получить постоянные препараты хорошего качества, на которых отчетливо видно строение растительной клетки. Однако в некоторых случаях фиксирующие агенты (спирт, кислоты, формалин, соли металлов) и красители могут исказить истинную картину клеточной структуры, заменив ее артефактами (структуры, созданные фиксирующим веществом). В этом случае наряду с постоянными препаратами следует параллельно изучать живые клетки. Последние чаще всего окрашивают: нейтральными красителями — цитоплазму, янусом зеленым — митохондрии, метиленовым синим — комплекс Гольджи. Используют и некоторые другие красители, сравнительно легко проникающие в живые клеткй.[ …]
Строение митохондрии рисунок. Что такое митохондрии? Их строение и функции
Митохондрия (с греческого μίτος (митос) – нить и χονδρίον (хондрион) – гранула) клеточная – двумембранный органоид, содержит свой собственный генетический материал, митохондриальную . Они встречаются как сферические или трубчатые клеточные структуры у почти всех эукариотов, но не у прокариотов.
Митохондрии – это органеллы, которые регенерируют высокоэнергетическую молекулу аденозинтрифосфата через дыхательную цепь. В дополнение к этому окислительному фосфорилированию они выполняют другие важные задачи, например, участвуют в образовании кластеров железа и серы . Строение и функции таких органоидов подробно рассмотрены ниже.
Вконтакте
Общие сведения
Особенно много находится митохондрий в с высоким энергопотреблением. К ним относятся мышечные, нервные, сенсорные клетки и ооциты. В клеточных структурах сердечной мышцы объемная доля этих органоидов достигает 36 %. Они имеют диаметр около 0.5-1.5 мкм и разнообразные формы, от сфер до сложных нитей. Их число корректируется с учетом энергетических потребностей клетки.
Эукариотические клетки, которые теряют свои митохондрии, не могут их восстановить . Существуют также эукариоты без них, например, некоторые простейшие. Количество данных органоидов на клеточную единицу обычно составляет от 1000 до 2000 при объемной доле в 25 %. Но эти значения могут сильно варьироваться в зависимости от типа клеточной структуры и организма. В зрелой клетке спермы их около четырех-пяти, в зрелой яйцеклетке – несколько сотен тысяч.
Митохондрии передаются через плазму яйцеклетки только от матери, что стало причиной исследования материнских линий. В настоящее время установлено, что также через сперму некоторые мужские органоиды импортируются в плазму оплодотворенной яйцеклетки (зиготы). Вероятно, они будут устранены довольно быстро. Однако есть несколько случаев, когда врачи смогли доказать, что митохондрии ребенка были отцовской линии. Заболевания, вызванные мутациями в митохондриальных генах, наследуются только от матери.
Интересно! Популярный научный термин «энергетическая станция клетки» был придуман в 1957 году Филиппом Сикевицем.
Схема строения митохондрии
Рассмотрим особенности строения этих важных структур. Они образованы в результате сочетания нескольких элементов. Оболочка этих органоидов складывается из внешней и внутренней мембраны, они в свою очередь состоят из фосфолипидных бислоев и белков. Обе оболочки отличаются по своим свойствам. Между ними расположено пять различных отсеков: наружная мембрана, межмембранное пространство (промежуток между двумя мембранами), внутренняя, криста и матрикс (пространство внутри внутренней мембраны), в целом – внутренние структуры органоида.
На иллюстрациях в учебниках митохондрия преимущественно выглядит как отдельная бобовидная органелла. Так ли это на самом деле? Нет, они образуют трубчатую митохондриальную сеть , которая может проходить и изменять всю клеточную единицу. Митохондрии в клетке способны сочетаться (путем слияния) и повторно делиться (делением).
Обратите внимание! В дрожжах за одну минуту совершается около двух митохондриальных слияний. Поэтому невозможно точное определение текущей численности митохондрий в клетках.
Внешняя мембрана
Наружная оболочка окружает всю органеллу и включает в себя каналы белковых комплексов, что позволяют обмен молекулами и ионами между митохондрией и цитозолем. Крупные молекулы не могут пройти через мембрану .
Внешняя, которая охватывает всю органеллу и не свернута, имеет весовое отношение фосфолипида к белку 1:1 и, таким образом, похожа на эукариотическую плазматическую мембрану. Она содержит множество интегральных белков, поринов. Порины образуют каналы, которые обеспечивают свободную диффузию молекул с массой до 5000 дальтон через оболочку. Более крупные белки могут вторгаться, когда сигнальная последовательность на N-конце связывается с большой субъединицей белка транслоксазы, из которой они затем активно перемещаются по мембранной оболочке.
Если трещины возникают во внешней оболочке, белки из межмембранного пространства могут выходить в цитозоль, что может привести к гибели клетки . Наружная мембрана может сливаться с оболочкой эндоплазматического ретикулума, а затем формировать структуру под названием MAM (ER, ассоциированную с митохондрией). Это важно для обмена сигналами между ER и митохондрией, что также необходимо для переноса .
Межмембранное пространство
Участок представляет собой промежуток посреди внешней и внутренней мембраны. Поскольку внешняя обеспечивает свободное проникновение малых молекул, их концентрация, таких как ионы и сахар, в межмембранном пространстве идентична концентрациям в цитозоле. Однако для больших белков требуется передача специфической сигнальной последовательности, так что состав белков различается между межмембранным пространством и цитозолем. Таким образом, белок, который удерживается в межмембранном промежутке, является цитохромом.
Внутренняя мембрана
Внутренняя митохондриальная мембрана содержит белки с четырьмя видами функций:
- Белки – проводят реакции оксидации респираторной цепочки.
- Аденозинтрифосфатсинтаза, которая производит в матрице АТФ.
- Специфические транспортные белки, которые регулируют проход метаболитов между матрицей и цитоплазмой.
- Системы импорта белков.
Внутренняя имеет, в частности, двойной фосфолипид, кардиолипин, замещенный четырьмя жирными кислотами. Кардиолипин обычно характерен для митохондриальных мембран и бактериальных плазматических мембран. В организме человека он в основном присутствует в областях с высокой метаболической активностью или высокой энергетической активностью, таких как сократительные кардиомиоциты, в миокарде.
Внимание! Внутренняя мембрана содержит более 150 различных полипептидов, около 1/8 всех митохондриальных белков. В результате концентрация липидов ниже, чем у внешнего бислоя, и его проницаемость ниже.
Разделяется на многочисленные кристы, они расширяют внешнюю область внутренней митохондриальной оболочки, поднимая ее способность вырабатывать АТФ.
В типичной митохондрии печени, например, внешняя область, в частности кристы, примерно в пять раз превышает площадь наружной мембраны. Энергетические станции клеток, которые имеют более высокие потребности в АТФ, например, мышечные клетки, содержат больше крист, чем типичная митохондрия печени.
Внутренняя оболочка охватывает матрикс, внутреннюю жидкость митохондрии. Он соответствует цитозолю бактерий и содержит митохондриальную ДНК, ферменты цитратного цикла и их собственные митохондриальные рибосомы, которые отличаются от рибосом в цитозоле (но также и от бактерий). Межмембранное пространство содержит ферменты, которые могут фосфорилировать нуклеотиды под потреблением АТФ.
Функции
- Важные пути деградации: цитратный цикл, для которого пируват вводится из цитозоля в матрикс. Затем пируват декарбоксилируют пируватдегидрогеназой до ацетилкофермента А. Другим источником ацетилкофермента А является деградация жирных кислот (β-окисление), которая происходит в клетках животных в митохондриях, но в растительных – только в глиоксисомах и пероксисомах. С этой целью ацилкофермент А переносят из цитозоля путем связывания с карнитином через внутреннюю митохондриальную мембрану и превращают в ацетилкофермента А. Из него большинство восстановительных эквивалентов в цикле Кребса (также известный как цикл Кребса или цикл трикарбоновой кислоты), которые затем превращаются в АТФ в окислительной цепи.
- Окислительная цепь. Установлен электрохимический градиент между межмембранным пространством и митохондриальным матриксом, который служит для получения АТФ с помощью АТФ-синтазы, с помощью процессов переноса электронов и накопления протонов. Электроны и протоны, необходимые для создания градиента, получают путем окислительной деградации из питательных веществ (например, глюкозы), поглощаемых организмом. Первоначально гликолиз происходит в цитоплазме.
- Апоптоз (запрограммированная гибель клеток)
- Хранение кальция: благодаря способности абсорбировать ионы кальция и затем высвобождать их, митохондрии вмешиваются в гомеостаз клетки.
- Синтез кластеров железа-серы, требуемый, среди прочего, многими ферментами дыхательной цепи. Эта функция теперь считается существенной функцией митохондрий, т.е. как это причина, по которой почти все клетки полагаются на энергетические станции для выживания.
Матрикс
Это пространство, включенное во внутреннюю митохондриальную мембрану. Содержит около двух третей общего белка. Играет решающую роль в производстве АТФ с помощью синтазы АТФ, включенной во внутреннюю мембрану. Содержит высококонцентрированную смесь сотен различных ферментов (главным образом, участвующих в деградации жирных кислот и пирувата), митохондриально-специфических рибосом, передаточной РНК и нескольких копий ДНК митохондриального генома.
Данные органоиды имеют свой собственный геном, а также ферментативное оборудование, необходимое для осуществления собственного биосинтеза белка .
Митохондрия Что такое Митохондрия и её функции
Строение и функционирование митохондрий
Вывод
Таким образом, митохондриями называются клеточные электростанции, которые производят энергию и занимают ведущее место в жизни и выживаемости отдельной клетки в частности и живого организма в целом. Митохондрии – это неотъемлемая часть живой клетки, в том числе растительной, которые до конца еще не изучены. Особенно много митохондрий в тех клетках, которым требуется больше энергии.
МИТОХОНДРИИ (mitochondria ; греч, mitos нить + chondrion зернышко) — органоиды, присутствующие в цитоплазме клеток животных и растительных организмов. М. принимают участие в процессах дыхания и окислительного фосфорилирования, продуцируют энергию, необходимую для функционирования клетки, представляя, таким образом, ее «силовые станции».
Термин «митохондрии» был предложен в 1894 г. Бендой (С. Benda). В середине 30-х гг. 20 в. удалось впервые выделить М. из клеток печени, что позволило исследовать эти структуры биохим, методами. В 1948 г. Хогебумом (G. Hogeboom) были получены окончательные доказательства того, что М. действительно являются центрами клеточного дыхания. Значительные успехи в изучении этих органоидов были сделаны в 60-70 гг. в связи с применением методов электронной микроскопии и молекулярной биологии.
Форма М. варьирует от почти круглых до сильно вытянутых, имеющих вид нитей (рис. 1), Размер их колеблется от 0,1 до 7 мкм. Количество М. в клетке зависит от типа ткани и функционального состояния организма. Так, в сперматозоидах число М. невелико — ок. 20 (на клетку), в клетках эпителия почечных канальцев млекопитающих их содержится до 300 в каждой, а у гигантской амебы (Chaos chaos) обнаружено 500 000 митохондрий, В одной клетке печени крысы ок. 3000 М., однако в процессе голодания животного число М. может сократиться до 700. Обычно М. распределяются в цитоплазме достаточно равномерно, однако в клетках нек-рых тканей М. могут быть постоянно локализованы в участках, особенно нуждающихся в энергии. Напр., в скелетной мышце М. часто находятся в контакте с контрактильными участками миофибрилл, образуя правильные трехмерные структуры. В сперматозоидах М. образуют спиральный футляр вокруг осевой нити хвоста, что, вероятно, связано с возможностью использовать энергию АТФ, синтезируемую в М., для движений хвоста. В аксонах М. концентрируются вблизи синаптических окончаний, где происходит процесс передачи нервных импульсов, сопровождающийся энергозатратой. В клетках эпителия почечных канальцев М. связаны с выпячиваниями базальной клеточной мембраны. Это вызвано необходимостью постоянного и интенсивного снабжения энергией процесса активного переноса воды и растворенных в ней веществ, протекающего в почках.
Электронно-микроскопически установлено, что М. содержит две мембраны — наружную и внутреннюю. Толщина каждой мембраны ок. 6 нм, расстояние между ними — 6-8 нм. Наружная мембрана гладкая, внутренняя образует сложные выросты (кристы), вдающиеся в полость митохондрии (рис. 2). Внутреннее пространство М. носит название матрикса. Мембраны представляют собой пленку из компактно уложенных молекул белков и липидов, в то время как матрикс подобен гелю и содержит в своем составе растворимые белки, фосфаты и другие хим. соединения. Обычно матрикс выглядит гомогенным, лишь в нек-рых случаях в нем можно обнаружить тонкие нити, трубочки и гранулы, содержащие ионы кальция и магния.
Из особенностей строения внутренней мембраны необходимо отметить наличие в ней сферических частиц ок. 8-10 нм в поперечнике, сидящих на короткой ножке и иногда выступающих в матрикс. Эти частицы были открыты в 1962 г. Фернандес-Мораном (H. Fernandez-Moran). Они состоят из белка, обладающего АТФ-азной активностью, получившего обозначение F1. Белок прикрепляется к внутренней мембране только со стороны, обращенной к матриксу. Частицы F1 располагаются на расстоянии 10 нм друг от друга, а в каждой М. содержится 10 4 -10 5 , таких частиц.
В кристах и внутренних мембранах М. содержится большинство дыхательных ферментов (см.), дыхательные ферменты организованы в компактные ансамбли, распределенные с правильными промежутками в кристах М. на расстоянии 20 нм друг от друга.
М. почти всех типов клеток животных и растений построены по единому принципу, однако возможны отклонения в деталях. Так, кристы могут располагаться не только поперек длинной оси органоида, но и продольно, напр, в М. синаптической зоны аксона. В ряде случаев кристы могут ветвиться. В М. простейших организмов, нек-рых насекомых и в клетках клубочковой зоны надпочечников кристы имеют форму трубочек. Число крист различно; так, в М. клеток печени и половых клеток крист очень мало и они короткие, в то время как матрикс обилен; в М. мышечных клеток кристы многочисленны, а матрикса мало. Существует мнение, что число крист коррелирует с окислительной активностью М.
Во внутренней мембране М. осуществляются параллельно три процесса: окисление субстрата цикла Кребса (см. Трикарбоновых кислот цикл), перенос освободившихся при этом электронов и накопление энергии путем образования макроэргических связей аденозинтрифосфата (см. Аденозинфосфорные кислоты). Основной функцией М. является сопряжение синтеза АТФ (из АДФ и неорганического фосфора) и аэробного процесса окисления (см. Окисление биологическое). Накопленная в молекулах АТФ энергия может трансформироваться в механическую (в мышцах), электрическую (нервная система), осмотическую (почки) и т. д. Процессы аэробного дыхания (см. Окисление биологическое) и сопряженного с ним окислительного фосфорилирования (см.) являются основными функциями М. Кроме того, в наружной мембране М. может происходить окисление жирных к-т, фосфолипидов и нек-рых других соединений.
В 1963 г. Насс и Насс (М. Nass, S. Nass) установили, что в М. содержится ДНК (одна или несколько молекул). Все исследованные до сих пор митохондриальные ДНК из животных клеток состоят из ковалентно замкнутых колец диам. ок. 5 нм. У растений митохондриальная ДНК значительно длиннее и не всегда имеет форму кольца. Митохондриальная ДНК во многих отношениях отличается от ядерной. Репликация ДНК происходит при помощи обычного механизма, однако не совпадает во времени с репликацией ядерной ДНК. Количество генетической информации, заключенной в молекуле митохондриальной ДНК, по-видимому, недостаточно для кодирования всех белков и ферментов, содержащихся в М. Митохондриальные гены кодируют в основном структурные белки мембран и белки, участвующие в морфогенезе митохондрий. М. имеют свои транспортные РНК и синтетазы, содержат все компоненты, необходимые для синтеза белка; их рибосомы меньше цитоплазматических и более похожи на рибосомы бактерий.
Продолжительность жизни М. сравнительно невелика. Так, время обновления половины количества М. составляет для печени 9,6-10,2 сут., для почки — 12,4 сут. Пополнение популяции М. происходит, как правило, из предсуществующих (материнских) М. путем их деления или почкования.
Давно высказывалось предположение, что в процессе эволюции М. возникли, вероятно, путем эндосимбиоза примитивных ядросодержащих клеток с бактериоподобными организмами. Имеется большое число доказательств этому: наличие собственной ДНК, более сходной с ДНК бактерий, чем с ДНК ядра клетки; присутствие в М. рибосом; синтез ДНК-зависимой РНК; чувствительность митохондриальных белков к антибактериальному препарату — хлорамфениколу; сходство с бактериями в реализации дыхательной цепи; морфол., биохим, и физиол, различия между внутренней и наружной мембраной. Согласно симбиотической теории клетка-хозяин рассматривается как анаэробный организм, источником энергии для к-рого является гликолиз (протекающий в цитоплазме). В «симбионте» же реализуется цикл Кребса и дыхательная цепь; он способен к дыханию и окислительному фосфорилированию (см.).
М. являются весьма лабильными внутриклеточными органоидами, раньше других реагирующими на возникновение каких-либо патол, состояний. Возможны изменения числа М. в клетке (вернее, в их популяциях) или изменения их структуры. Напр., при голодании, действии ионизирующего облучения число М. уменьшается. Структурные изменения обычно состоят в набухании всего органоида, просветлении матрикса, разрушении крист, нарушении целостности наружной мембраны.
Набухание сопровождается значительным изменением объема М. В частности, при ишемии миокарда объем М. увеличивается в 10 раз и более. Различают два типа набухания: в одном случае оно связано с изменением осмотического давления внутри клетки, в других случаях — с изменениями клеточного дыхания, сопряженного с ферментативными реакциями и первичными функциональными расстройствами, вызывающими изменения водного обмена. Помимо набухания, может происходить вакуолизация М.
Независимо от причин, вызывающих патол, состояние (гипоксия, гиперфункция, интоксикация), изменения М. довольно стереотипны и неспецифичны.
Наблюдаются такие изменения структуры и функции М., к-рые, по-видимому, становились причиной возникновения болезни. В 1962 г. Луфт (R. Luft) описал случай «митохондриальной болезни». Больному с резко повышенной интенсивностью обмена веществ (при нормальной функции щитовидной железы) была сделана пункция скелетной мышцы и найдено повышенное число М., а также нарушение структуры крист. Дефектные митохондрии в клетках печени наблюдались и при выраженном тиреотоксикозе. Виноград (J. Vinograd) с сотр. (с 1937 по 1969) обнаружил, что у больных с определенными формами лейкемии митохондриальные ДНК из лейкоцитов заметно отличались от нормальных. Они представляли собой открытые кольца или группы сцепленных колец. Частота этих аномальных форм снижалась в результате химиотерапии.
Библиография: Гаузе Г. Г. Митохондриальная ДНК, М., 1977,библиогр.; Д e P о-бертис Э., Новинский В. и С а э с Ф. Биология клетки, пер. с англ., М., 1973; Озернюк Н. Д. Рост и воспроизведение митохондрий, М., 1978, библиогр.; Поликар А. и Бесси М. Элементы патологии клетки, пер. с франц., М., 1970; РудинД. и Уилки Д. Биогенез митохондрий, пер. с англ., М., 1970, библиогр.; Серов В. В. и Пауков В. С. Ультраструктурная патология, М., 1975; С э д ж e р Р. Цитоплазматические гены и органеллы, пер. с англ., М., 1975.
Т. А. Залетаева.
(от греч. mitos — нить, chondrion — зернышко, soma — тельце) представляют собой гранулярные или нитевидные органоиды ( рис. 1, а). Митохондрии можно наблюдать в живых клетках, так как они обладают достаточно высокой плотностью. В таких клетках митохондрии могут двигаться, перемещаться, сливаться друг с другом. Особенно хорошо митохондрии выявляются на препаратах, окрашенных различными способами. Размеры митохондрий непостоянны у разных видов, так же изменчива их форма. Все же у большинства клеток толщина этих структур относительно постоянна (около 0,5 мкм), но длина колеблется, достигая у нитчатых форм 7-60 мкм.
Митохондрии независимо от их величины и формы имеют универсальное строение, их ультраструктура однообразна. Митохондрии ограничены двумя мембранами ( рис. 1, б), у них четыре субкомпартмента: митохондриальный матрикс , внутренняя мембрана , мембранное пространство и внешняя мембрана , обращенная к цитозолю. Внешняя мембрана отделяет ее от остальной цитоплазмы. Толщина внешней мембраны около 7 нм, она не связана ни с какими другими мембранами цитоплазмы и замкнута сама на себя, так что представляет собой мембранный мешок. Наружную мембрану от внутренней отделяет межмембранное пространство шириной около 10-20 нм. Внутренняя мембрана (толщиной около 7 нм) ограничивает собственно внутреннее содержимое митохондрии, ее матрикс , или митоплазму . Характерной чертой внутренних мембран митохондрий является их способность образовывать многочисленные выпячивания (складки) внутрь митохондрий. Такие выпячивания ( кристы , рис. 27) чаще всего имеют вид плоских гребней. Митохондрии осуществляют синтез АТФ, происходящий в результате процессов окисления органических субстратов и фосфорилирования АДФ.
Митохондрии специализируются на синтезе АТФ путем транспорта электронов и окислительного фосфорилирования. (рис 21-1). Хотя они имеют свою собственную ДНК и аппарат белкового синтеза, большинство их белков кодируется клеточной ДНК и поступает из цитозоля. Более того, каждый поступивший в органеллу белок должен достичь определенного субкомпартмента, в котором он функционирует.
Митохондрии — это «энергетические станции» эукариотических клеток. В кристы встроены ферменты, участвующие в преобразовании энергии питательных веществ, поступающих в клетку извне, в энергию молекул АТФ. АТФ — «универсальная валюта», которой клетки расплачиваются за все свои энергетические расходы. Складчатость внутренней мембраны увеличивает поверхность, на которой размещаются ферменты, синтезирующие АТФ. Количество крист в митохондрии и количество самих митохондрий в клетке тем больше, чем больше энергетических трат осуществляет данная клетка. В летательных мышцах насекомых каждая клетка содержит несколько тысяч митохондрий. Меняется их количество и в процессе индивидуального развития (онтогенеза): в молодых эмбриональных клетках они более многочисленны, чем в клетках стареющих. Обычно митохондрии скапливаются вблизи тех участков цитоплазмы, где возникает потребность в АТФ, образующейся в митохондриях.
Расстояние между мембранами в кристе составляет около 10-20 нм. У простейших, одноклеточных водорослей в некоторых клетках растений и животных выросты внутренней мембраны имеют вид трубочек диаметром около 50 нм. Это так называемые трубчатые кристы.
Митохондриальный матрикс гомогенен и имеет более плотную консистенцию, чем окружающая митохондрию гиалоплазма. В матриксе выявляются тонкие нити ДНК и РНК, а также митохондриальные рибосомы, на которых синтезируются некоторые митохондриальные белки. С помощью электронного микроскопа на внутренней мембране и кристах со стороны матрикса можно увидеть грибовидные образования — АТФ-сомы. Это ферменты, образующие молекулы АТФ. Их может быть до 400 на 1 мкм.
Немногие белки, которые кодируются собственным геномом митохондрий, расположены в основном во внутренней мембране. Они обычно образуют субъединицы белковых комплексов, другие компоненты которых кодируются ядерными генами и поступают из цитозоля. Образование таких гибридных агрегатов требует сбалансирования синтеза этих двух типов субъединиц; каким образом координируется синтез белка на рибосомах разных типов, разделенных двумя мембранами, остается загадкой.
Обычно митохондрии располагаются в местах, где необходима энергия для любых жизненных процессов. Возник вопрос, каким образом транспортируется в клетке энергия — путем ли диффузии АТФ и нет ли в клетках структур, исполняющих роль электрических проводников, которые могли бы энергетически объединять отдаленные друг от друга участки клетки. Гипотеза заключается в том, что разность потенциалов в определенной области мембраны митохондрий передается вдоль нее и превращается в работу в другой области той же мембраны [ Скулачев В.П., 1989 ].
Как представлялось, подходящими кандидатами на эту же роль могли быть мембраны самих митохондрий. Кроме того, исследователей интересовали взаимодействие в клетке множественных митохондрий друг с другом, работа всего ансамбля митохондрий, всего хондриома — совокупности всех митохондрий.
Митохондрии характерны за малым исключением для всех эукариотических клеток как аутотрофных (фотосинтезирующие растения), так и гетеротрофных (животные, грибы) организмов. Их основная функция связана с окислением органических соединений и использованием освобождающейся при распаде этих соединений энергии в синтезе молекул АТФ. Поэтому митохондрии часто называют энергетическими станциями клетки.
Митохондрии — органеллы энергообеспечения метаболических процесов в клетке. Размеры их варьируют от 0,5 до 5-7 мкм, количество в клетке составляет от 50 до 1000 и более. В гиалоплазме митохондрии распределены обычно диффузно, однако в специализированных клетках сосредоточены в тех участках, где имеется наибольшая потребность в энергии. Например, в мышечных клетках и симпластах большие количества митохондрий сосредоточены вдоль рабочих элементов — сократительных фибрилл. В клетках, функции которых сопряжены с особо высокими энергозатратами, митохондрии образуют множественные контакты, объединяясь в сеть, или кластеры (кардиомиоциты и симпласты скелетной мышечной ткани). В клетке митохондрии выполняют функцию дыхания. Клеточное дыхание — это последовательность реакций, с помощью которых клетка использует энергию связей органических молекул для синтеза макроэргических соединений типа АТФ. Образующиеся внутри митохондрии молекулы АТФ переносятся наружу, обмениваясь на молекулы АДФ, находящиеся вне митохондрии. В живой клетке митохондрии могут передвигаться с помощью элементов цитоскелета. На ультрамикроскопическом уровне стенка митохондрии состоит из двух мембран — наружной и внутренней. Наружная мембрана имеет относительно ровную поверхность, внутренняя — образует направленные в центр складки, или кристы. Между наружной и внутренней мембранами возникает неширокое (около 15 нм) пространство, которое называется наружной камерой митохондрии; внутренняя мембрана ограничивает внутреннюю камеру. Содержимое наружной и внутренней камер митохондрии различно, и так же, как и сами мембраны, существенно отличается не только по рельефу поверхности, но и по ряду биохимических и функциональных признаков. Наружная мембрана по химическому составу и свойствам близка к другим внутриклеточным мембранам и плазмолемме.
Ее характеризует высокая проницаемость, благодаря наличию гидрофильных белковых каналов. Эта мембрана имеет в своем составе рецепторные комплексы, распознающие и связывающие вещества, поступающие в митохондрию. Ферментный спектр наружной мембраны небогат: это ферменты метаболизма жирных кислот, фосфолипидов, липидов и др. Главной функцией наружной мембраны митохондрии является отграничение органеллы от гиалоплазмы и транспорт необходимых для осуществления клеточного дыхания субстратов. Внутренняя мембрана митохондрий в большинстве клеток тканей различных органов формирует кристы в виде пластин (ламеллярные кристы), что значительно увеличивает площадь поверхности внутренней мембраны. В последней 20-25 % всех белковых молекул составляют ферменты дыхательной цепи и окислительного фосфорилирования. В эндокринных клетках надпочечников и половых желез митохондрии участвуют в синтезе стероидных гормонов. В этих клетках митохондрии имеют кристы в виде трубочек (тубул), упорядоченно расположенных в определенном направлении. Поэтому кристы митохондрий в стероидпродуцирующих клетках названных органов именуются тубулярными. Матрикс митохондрии, или содержимое внутренней камеры, представляет собой гелеобразную структуру, содержащую около 50 % белков. Осмиофильные тельца, описанные при электронной микроскопии, — это резервы кальция. Матрикс содержит ферменты цикла лимонной кислоты, катализирующие окисление жирных кислот, синтез рибосом, ферменты, участвующие в синтезе РНК и ДНК. Общее число ферментов превышает 40. Помимо ферментов, матрикс митохондрии содержит митохондриальную ДНК (митДНК) и митохондриальные рибосомы. Молекула митДНК имеет кольцевидную форму. Возможности внутримитохондриального белкового синтеза ограничены — здесь синтезируются транспортные белки митохондриальных мембран и некоторые ферментные белки, участвующие в фосфорилировании АДФ. Все остальные белки митохондрии кодируются ядерной ДНК, и их синтез осуществляется в гиалоплазме, и в дальнейшем они транспортируются в митохондрию. Жизненный цикл митохондрий в клетке короткий, поэтому природа наделила их двойственной системой воспроизводства — помимо деления материнской митохондрии, возможно образование нескольких дочерних органелл путем почкования.
Строение и функции митохондрий представляют собой довольно сложный вопрос. Наличие органеллы характерно почти для всех ядерных организмов – как для автотрофов (растений, способных к фотосинтезу), так и для гетеротрофов, которыми являются почти все животные, некоторые растения и грибы.
Главное предназначение митохондрий – окисление органических веществ и последующее использование освободившейся в результате этого процесса энергии. По этой причине органеллы имеют также и второе (неофициальное) название – энергетические станции клетки. Иногда их называют «пластидами катаболизма».
Что такое митохондрии
Термин имеет греческое происхождение. В переводе это слово означает «нить» (mitos), «зернышко» (chondrion). Митохондрии являются постоянными органоидами, которые имеют огромное значение для нормального функционирования клеток и делают возможным существование всего организма в целом.
«Станции» имеют специфическую внутреннюю структуру, которая изменяется в зависимости от функционального состояния митохондрии. Их форма может быть двух видов – овальная или продолговатая. Последняя нередко имеет ветвящийся вид. Число органоидов в одной клетке колеблется от 150 до 1500.
Особый случай – половые клетки. В сперматозоидах присутствует всего лишь одна спиральная органелла, в то время как женских гаметах содержится в сотни тысяч больше митохондрий. В клетке органоиды не зафиксированы в одном месте, а могут передвигаться по цитоплазме, совмещаться друг с другом. Их размер составляет 0,5 мкм, длина может достигать 60 мкм, в то время как минимальный показатель – 7 мкм.
Определить размер одной «энергетической станции» – непростая задача. Дело в том, что при рассмотрении в электронный микроскоп на срез попадает только часть органеллы. Случается так, что спиральная митохондрия имеет несколько сечений, которые можно принять за отдельные, самостоятельные структуры.
Только объемное изображение позволит выяснить точное клеточное строение и понять, идет речь о 2-5 отдельных органоидах или же об одной, имеющей сложную форму митохондрии.
Особенности строения
Оболочка митохондрии состоит из двух слоев: наружного и внутреннего. Последний включает в себя различные выросты и складки, которые имеют листовидную и трубчатую форму.
Каждая мембрана имеет особенный химический состав, определенное количество тех или иных ферментов и конкретное предназначение. Наружную оболочку от внутренней отделяет межмембранное пространство толщиной 10-20 нм.
Весьма наглядно выглядит строение органеллы на рисунке с подписями.
Схема строения митохондрии
Посмотрев на схему строения, можно сделать следующее описание. Вязкое пространство внутри митохондрии называется матриксом. Его состав создает благоприятную среду для протекания в ней необходимых химических процессов. В его составе присутствуют микроскопические гранулы, которые содействуют реакциям и биохимическим процессам (например, накапливают ионы гликогена и других веществ).
В матриксе находятся ДНК, коферменты, рибосомы , т-РНК, неорганические ионы. На поверхности внутреннего слоя оболочки располагаются АТФ-синтаза и цитохромы. Ферменты способствуют таким процессам, как цикл Кребса (ЦТК), окислительное фосфорилирование и т. д.
Таким образом, главная задача органоида выполняется как матриксом, так и внутренней стороной оболочки.
Функции митохондрий
Предназначение «энергетических станций» можно охарактеризовать двумя основными задачами:
- выработка энергии: в них осуществляются окислительные процессы с последующим выделением молекул АТФ;
- хранение генетической информации;
- участие в синтезе гормонов, аминокислот и других структур.
Процесс окисления и выработки энергии проходят в несколько стадий:
Схематичный рисунок синтеза АТФ
Стоит отметить: в результате цикла Кребса (цикл лимонной кислоты) не образуются молекулы АТФ, происходит окисление молекул и выделение углекислого газа. Это промежуточный этап между гликолизом и электронтранспортной цепью.
Таблица «Функции и строение митохондрий»
От чего зависит число митохондрий в клетке
Превалирующее число органоидов скапливается рядом с теми участками клетки, где возникает необходимость в энергетических ресурсах. В частности, большое количество органелл собирается в зоне нахождения миофибрилл, которые являются частью мышечных клеток, обеспечивающих их сокращение.
В мужских половых клетках структуры локализуются вокруг оси жгутика – предполагается, что потребность в АТФ обусловлена постоянным движением хвоста гаметы. Точно так же выглядит расположение митохондрий у простейших, которые для передвижения используют специальные реснички – органеллы скапливаются под мембраной у их основания.
Что касается нервных клеток, то локализация митохондрий наблюдается вблизи синапсов, через которые передаются сигналы нервной системы. В клетках, синтезирующих белки, органеллы скапливаются в зонах эргастоплазмы – они поставляют энергию, которая обеспечивает данный процесс.
Кто открыл митохондрии
Свое название клеточная структура обрела в 1897-1898 годах благодаря К. Бренду. Связь процессов клеточного дыхания с митохондриями сумел доказать Отто Вагбург в 1920 году.
Заключение
Митохондрии являются важнейшей составляющей живой клетки, выступая в роли энергетической станции, которая производит молекулы АТФ, обеспечивая тем самым процессы клеточной жизнедеятельности.
Работа митохондрий основана на окислении органических соединений, в результате чего происходит генерация энергетического потенциала.
Митохондрия: определение, функция и структура — видео и стенограмма урока
Митохондрии и другие органеллы
Митохондрии похожи на другие органеллы в том, что они покрыты собственными мембранами. Они отличаются тем, что их мембраны являются результатом свободно плавающих рибосом, находящихся в окружающем цитозоле, а также рибосом, содержащихся внутри митохондрий. Цитозоль — водорастворимый компонент цитоплазмы, расположенный внутри клеточной мембраны. Рибосомы — это крошечные органеллы, состоящие из рибонуклеиновой кислоты (РНК), и они играют важную роль в синтезе белка.
Митохондрии также содержат небольшое количество дезоксирибонуклеиновой кислоты — или ДНК, как ее более известно, — которая является учебным материалом, используемым для программирования белков, вырабатываемых рибосомами митохондрий. Митохондрии считаются полуавтономными, потому что они могут расти и воспроизводиться самостоятельно без указаний ядра.
Размер, форма, структура митохондрий
Взгляните на диаграмму ниже:
Вы смотрите на поперечное сечение митохондрии. Обратите внимание на складки, или cristae , на внутренней мембране.
В зависимости от метаболической активности клетки в одной клетке могут быть сотни или даже тысячи митохондрий. Каждый из них имеет длину от одного до десяти микрометров и меньше самого короткого человеческого волоса, длина которого составляет примерно 17 микрометров.Хотя реальная форма митохондрии может измениться, она похожа на фасоль.
Внешняя сторона митохондрии покрыта гладкой мембраной, а внутренняя мембрана имеет многочисленные складки, называемые кристами. Эти складки позволяют метаболической активности, такой как клеточное дыхание, происходить на большей площади. У этой области есть конкретное название митохондриальный матрикс , где много разных ферментов и рибосом выполняют свою работу.
Резюме урока
Давайте рассмотрим то, что мы здесь узнали. Митохондрии — это органеллы, которые представляют собой специализированные структуры, обнаруженные внутри почти всех эукариотических клеток, и считаются двигателем клетки. Эукариотические клетки характеризуются своим ядром и мембраносвязанными органеллами. Их исчисляются сотнями или даже тысячами, и они необходимы для клеточного дыхания , которое происходит, когда кислород расщепляет молекулы пищи и генерирует энергию, или АТФ.
Митохондрии автономны в своей собственной мембране и могут расти и воспроизводиться независимо от самой клетки.Они также содержат небольшое количество ДНК , учебного материала, используемого для программирования белков, происходящих из митохондриальных рибосом. Помните также, что рибосом представляют собой крошечные органеллы, состоящие из рибонуклеиновой кислоты (РНК), и они играют важную роль в синтезе белка и содержатся внутри клеточной мембраны вместе с цитозолем , который является водорастворимым компонентом цитоплазмы. Митохондрии — ключ к выживанию всего живого, включая нас самих.Без их помощи мы не смогли бы расщеплять пищу и создавать энергию.
Структура митохондрий
Цитоплазма почти всех эукариотических клеток содержат митохондрии, хотя есть как минимум одно исключение, Протист Chaos (Pelomyxa) carolinensis . Они есть особенно много в клетках и частях клеток, которые связаны с активными процессами. Например, у жгутиковых простейших или в сперме млекопитающих митохондрии сосредоточены вокруг основание жгутика или жгутика.В сердечной мышце, митохондриях окружают сократительные элементы. Летающая мышца колибри является одним из самых богатых известных источников митохондрий. Таким образом, только по их распространению можно было бы заподозрить, что они вовлечены в производство энергии. Многоклеточные организмы вероятно, не могло бы существовать без митохондрий. Невозможность удалить электроны из система и накопление конечных продуктов метаболизма ограничивают полезность анаэробного метаболизма.Через окислительное фосфорирование митохондрии эффективно используют молекулы питательных веществ. Они являются причиной того, что нам вообще нужен кислород.
Двухмембранная митохондрия можно условно охарактеризовать как большой помятый мешок упакованы в меньший, неморщенный пакет. В две мембраны создают отдельные отсеки внутри органеллы, и сами очень разные по структуре и по функциям.
Наружная мембрана относительно простой фосфолипидный бислой, содержащий белковые структуры, называемые поринами, которые сделать его проницаемым для молекул размером около 10 килодальтон или меньше (размер мельчайших белков).Ионы, молекулы питательных веществ, АТФ, АДФ и т. д. могут проходить через внешняя мембрана с легкостью.
Внутренняя мембрана свободно проницаема только для кислорода, углекислого газа, и вода. Его структура очень сложна, в том числе все комплексы электронного транспорта система, комплекс синтетазы АТФ, и транспортные белки. Морщины, или складки, организованы в ламиллы (слои), называется кристами (единственное число: Криста).Кристы значительно увеличивают общий площадь внутренней мембраны. Чем больше площадь поверхности дает место многим другим из вышеперечисленных структуры, чем если бы внутренняя мембрана имела форму как внешняя мембрана.
Мембраны создают два отсеки. Межмембранное пространство, как подразумевается, это область между внутренней и внешней мембранами. Он играет важную роль в основной функции митохондрий, что является окислительным фосфорилированием.
Матрица содержит ферменты, отвечающие за лимонную реакции кислотного цикла. Матрица также содержит растворенный кислород, вода, углекислый газ, пригодный для повторного использования промежуточные звенья, которые служат энергетическими шаттлами, и многое другое (см. «другое функции «). Распространение — очень медленный процесс. Из-за складок крист нет части матрица находится далеко от внутренней мембраны.Следовательно компоненты матрицы могут диффундировать к внутренней мембране комплексов и транспортных белков в относительно мало времени.
На электронных микрофотографиях обнаружено трехмерная структура митохондрий. Однако, поскольку микрофотографии сами по себе двумерны, их интерпретация может вводить в заблуждение. Тексты часто покажите изображение «типичной» митохондрии в виде эллипсоид размером с бактерию (возможно, 0.5 на 1 мкм). Однако они сильно различаются по форме и размеру. Электрон микрофотографии редко показывают такие вариации, потому что это двухмерные изображения.
Изолированные митохондрии, например, из гомогенизированная мышечная ткань, имеет округлый вид на электронных микрофотографиях, подразумевая, что митохондрии представляют собой сферические органеллы.
Митохондрии in situ может быть свободным в цитоплазме или упакованным среди прочего жесткие структуры, такие как миофибриллы ткани сердечной мышцы.В таких клетках, как мышцы, ясно, что митохондрии не сферические, и часто даже не эллипсоидные. В некоторых тканях митохондрии почти нитевидные, что характерно для что двумерные микрофотографии могут не выявить.
Плоский разрез прорезает один или несколько частей органеллы, составляющие единый органелл, по-видимому, больше одной. В изображение, которое мы видим круглой или эллипсоидальной органеллы может скрыть истинную природу митохондрии.
Структура, функция и эволюция аппарата деления митохондрий
Митохондрии происходят из свободноживущих α-протеобактерий, которые были поглощены эукариотическими клетками-хозяевами в процессе эндосимбиоза и, следовательно, имеют свою собственную ДНК, которая организована с использованием основных белков для образуют ядра органелл (нуклеоиды). Митохондрии делятся и разделяются между дочерними клетками во время пролиферации клеток. Их деление можно разделить на два основных этапа: деление митохондриальных ядер и деление матрикса (так называемое митохондриальное деление или митохондриокинез).В этом обзоре мы сначала сосредоточимся на цитогенетических отношениях между делением митохондриального ядра и митохондриокинезом. Митохондриокинез возникает после деления митохондриального ядра, аналогично бактериальному цитокинезу. Затем мы описываем тонкую структуру и динамику делительного кольца митохондрий (MD-кольца) как основной морфологический фон митохондриокинеза. Исследования с помощью электронной микроскопии сначала идентифицировали небольшое электронно-плотное кольцо MD в цитоплазме в местах сужения митохондрий в слизистой плесени Physarum polycephalum , а затем два больших кольца MD (с внешней цитоплазматической и внутренней сторонами матрикса) в красных водорослях. Cyanidioschyzon merolae. Теперь кольца MD обнаружены у всех эукариот. В третьем разделе мы описываем отношения между кольцом MD и кольцом FtsZ, происходящим от предковых бактерий. Помимо GTPase, FtsZ, митохондрии потеряли большую часть белков, необходимых для бактериального цитокинеза, вследствие эндосимбиоза. Белок FtsZ образует электронно-прозрачное кольцо (кольцо FtsZ или Z) в матрице внутри внутреннего кольца MD. В четвертом разделе мы описываем динамическую ассоциацию между внешним MD-кольцом и кольцом, состоящим из эукариот-специфичного динамина GTPase.Недавние исследования показали, что эукариот-специфичные динамины GTPase образуют электронно-прозрачное кольцо между внешней мембраной и кольцом MD. Таким образом, считается, что митохондриокинез контролируется аппаратом деления митохондрий (MD), включающим динамическое трио, а именно FtsZ, MD и кольца динамина, которые состоят из химеры колец от бактерий и эукариот в примитивных организмах. Поскольку гены MD-кольца и динаминовых колец не обнаруживаются в прокариотическом геноме, геномы хозяина могут создавать эти кольца, чтобы активно контролировать деление митохондрий.В пятой части мы сосредоточимся на динамических изменениях в формировании и разборке колец FtsZ, MD и Dynamin. Кольца FtsZ перевариваются во время более позднего периода деления митохондрий, а затем, наконец, аппараты MD и динаминовые кольца отщепляются от дочерних митохондрий, подтверждая идею о том, что геномы хозяина несут ответственность за окончательный контроль деления митохондрий. Мы обсуждаем эволюцию от оригинальных аппаратов для деления везикул (ВД) до аппаратов ВД, включая классические динаминовые кольца и аппараты МД.Вероятно, что аппараты MD с участием динамического трио эволюционировали в аппарат деления пластид (PD) у Bikonta, тогда как у Opisthokonta аппарат MD был упрощен во время эволюции и, возможно, разветвился в аппарат слияния митохондрий. Наконец, мы описываем возможность интактной изоляции больших аппаратов MD / PD, идентификацию всех их белков и связанных с ними генов с использованием информации о геноме C. merolae и анализа TOF-MS. Эти результаты помогут выяснить универсальный механизм и эволюцию аппаратов MD, PD и VD.
4.3D: Митохондрии — Biology LibreTexts
Митохондрии — это органеллы, которые отвечают за выработку аденозинтрифосфата (АТФ), основной молекулы, переносящей энергию.
Задачи обучения
- Объясните роль митохондрий.
Ключевые моменты
- Митохондрии содержат собственные рибосомы и ДНК; В сочетании с их двойной мембраной эти особенности предполагают, что когда-то они могли быть свободноживущими прокариотами, которые были поглощены более крупной клеткой.
- Митохондрии играют важную роль в клеточном дыхании за счет производства АТФ с использованием химической энергии, содержащейся в глюкозе и других питательных веществах.
- Митохондрии также ответственны за образование кластеров железа и серы, которые являются важными кофакторами многих ферментов.
Ключевые термины
- альфа-протеобактерии : таксономический класс внутри филума Proteobacteria — фототропные протеобактерии.
- аденозинтрифосфат : многофункциональный нуклеозидтрифосфат, используемый в клетках в качестве кофермента, часто называемый «молекулярной единицей энергетической валюты» при внутриклеточной передаче энергии
- кофактор : неорганическая молекула, необходимая для функционирования фермента
Одним из основных отличий прокариот от эукариот является наличие митохондрий.Митохондрии — это двухмембранные органеллы, содержащие собственные рибосомы и ДНК. Каждая мембрана представляет собой бислой фосфолипидов, залитый белками. Эукариотические клетки могут содержать от одной до нескольких тысяч митохондрий, в зависимости от уровня потребления энергии клеткой. Каждая митохондрия имеет длину от 1 до 10 микрометров (или больше) и существует в клетке в виде органеллы, которая может быть яйцевидной, червеобразной или сложно разветвленной.
Структура митохондрий
Большинство митохондрий окружены двумя мембранами, что могло бы произойти, когда один мембраносвязанный организм был поглощен вакуолью другим мембраносвязанным организмом.Внутренняя мембрана митохондрий обширна и включает значительные складки, называемые кристами, которые напоминают текстурированную внешнюю поверхность альфа-протеобактерий. Матрикс и внутренняя мембрана богаты ферментами, необходимыми для аэробного дыхания.
Рисунок \ (\ PageIndex {1} \): Структура митохондрий : На этой электронной микрофотографии показана митохондрия в просвечивающем электронном микроскопе. Эта органелла имеет внешнюю и внутреннюю мембраны. Внутренняя мембрана содержит складки, называемые кристами, которые увеличивают площадь ее поверхности.Пространство между двумя мембранами называется межмембранным пространством, а пространство внутри внутренней мембраны называется митохондриальным матриксом. Синтез АТФ происходит на внутренней мембране.Митохондрии имеют собственную (обычно) кольцевую хромосому ДНК, которая стабилизируется за счет прикрепления к внутренней мембране и несет гены, аналогичные генам, экспрессируемым альфа-протеобактериями. Митохондрии также имеют особые рибосомы и передающие РНК, которые напоминают эти компоненты у прокариот. Все эти особенности подтверждают гипотезу о том, что митохондрии когда-то были свободноживущими прокариотами.
Функция митохондрий
Митохондрии часто называют «электростанциями» или «энергетическими фабриками» клетки, потому что они отвечают за выработку аденозинтрифосфата (АТФ), основной молекулы, несущей энергию в клетке. АТФ представляет собой кратковременную запасенную энергию клетки. Клеточное дыхание — это процесс производства АТФ с использованием химической энергии, содержащейся в глюкозе и других питательных веществах. В митохондриях этот процесс использует кислород и производит углекислый газ в качестве побочного продукта.Фактически, углекислый газ, который вы выдыхаете при каждом вдохе, возникает в результате клеточных реакций, которые производят углекислый газ в качестве побочного продукта.
Важно отметить, что в мышечных клетках очень высокая концентрация митохондрий, производящих АТФ. Вашим мышечным клеткам нужно много энергии, чтобы ваше тело двигалось. Когда ваши клетки не получают достаточно кислорода, они не производят много АТФ. Вместо этого небольшое количество АТФ, которое они производят в отсутствие кислорода, сопровождается образованием молочной кислоты.
Помимо аэробной генерации АТФ, митохондрии выполняют несколько других метаболических функций. Одна из этих функций — генерировать кластеры железа и серы, которые являются важными кофакторами многих ферментов. Такие функции часто связаны с уменьшением происходящих из митохондрий органелл анаэробных эукариот.
Происхождение митохондрий
Есть две гипотезы о происхождении митохондрий: эндосимбиотическая и аутогенная, но наиболее признанной в настоящее время является эндосимбиоз.Эндосимбиотическая гипотеза предполагает, что митохондрии изначально были прокариотическими клетками, способными реализовывать окислительные механизмы. Эти прокариотические клетки могли быть поглощены эукариотом и стали эндосимбионтами, живущими внутри эукариота.
молекулярных выражений клеточная биология: митохондрии
Митохондрии
Митохондрии представляют собой палочковидные органеллы, которые можно рассматривать как генераторы энергии в клетке, преобразующие кислород и питательные вещества в аденозинтрифосфат ( ATP ).АТФ — это химическая энергетическая «валюта» клетки, которая обеспечивает метаболическую активность клетки. Этот процесс называется аэробным дыханием и является причиной того, что животные дышат кислородом. Без митохондрий (единичных, митохондрий) высшие животные, вероятно, не существовали бы, потому что их клетки могли бы получать энергию только от анаэробного дыхания (в отсутствие кислорода), процесса, гораздо менее эффективного, чем аэробное дыхание. Фактически, митохондрии позволяют клеткам производить в 15 раз больше АТФ, чем они могли бы в противном случае, а сложные животные, такие как люди, нуждаются в большом количестве энергии, чтобы выжить.
Количество митохондрий, присутствующих в клетке, зависит от метаболических потребностей этой клетки и может варьироваться от одной большой митохондрии до тысяч органелл. Митохондрии, которые встречаются почти у всех эукариот, включая растения, животных, грибы и протистов, достаточно велики, чтобы их можно было наблюдать с помощью светового микроскопа, и были впервые обнаружены в 1800-х годах. Название органелл было придумано, чтобы отразить то, как они выглядели для первых ученых, которые их наблюдали, происходящее от греческих слов, обозначающих «нить» и «гранула».«В течение многих лет после своего открытия считалось, что митохондрии передают наследственную информацию. Современное понимание функции митохондрий было разработано только в середине 1950-х годов, когда был разработан метод выделения неповрежденных органелл.
Сложная структура митохондрии очень важна для функционирования органеллы (см. Рисунок 1). Две специализированные мембраны окружают каждую митохондрию, присутствующую в клетке, разделяя органеллу на узкое межмембранное пространство и гораздо более крупную внутреннюю матрицу , каждая из которых содержит узкоспециализированные белки.Наружная мембрана митохондрии содержит множество каналов, образованных белком порином , и действует как сито, отфильтровывая слишком большие молекулы. Точно так же внутренняя мембрана, которая сильно извилистая, так что образуется большое количество складок, называемых кристами , также позволяет проходить через нее только определенным молекулам и является гораздо более избирательной, чем внешняя мембрана. Чтобы убедиться, что в нее попадают только те материалы, которые необходимы для матрицы, внутренняя мембрана использует группу транспортных белков, которые будут транспортировать только правильные молекулы.Вместе различные компартменты митохондрии могут работать в гармонии, генерируя АТФ в сложном многоступенчатом процессе.
Митохондрии, как правило, представляют собой продолговатые органеллы, размер которых колеблется от 1 до 10 микрометров в длину, и их количество напрямую коррелирует с уровнем метаболической активности клетки. Однако органеллы довольно гибкие, и покадровые исследования живых клеток показали, что митохондрии быстро меняют форму и почти постоянно перемещаются в клетке.Движения органелл, по-видимому, каким-то образом связаны с микротрубочками, присутствующими в клетке, и, вероятно, транспортируются по сети с моторными белками. Следовательно, митохондрии могут быть организованы в длинные перемещающиеся цепи, плотно упакованы в относительно стабильные группы или появляться во многих других образованиях в зависимости от конкретных потребностей клетки и характеристик ее сети микротрубочек.
На рисунке 2 представлено цифровое изображение митохондриальной сети, обнаруженной в ткани яичников родственника горного козла, известного как гималайский тар, которое видно в флуоресцентном оптическом микроскопе.Обширная переплетенная сеть помечена синтетическим красителем MitoTracker Red (красная флуоресценция), который локализуется в дышащих митохондриях живых клеток в культуре. Редкие двойные ядра в этой клетке были окрашены синим красителем (голубая флуоресценция), чтобы обозначить их централизованное расположение по отношению к митохондриальной сети. Флуоресцентная микроскопия — важный инструмент, который ученые используют для изучения структуры и функций внутренних клеточных органелл.
Митохондрия отличается от большинства других органелл, потому что она имеет собственную кольцевую ДНК (подобную ДНК прокариот) и воспроизводится независимо от клетки, в которой она находится; очевидный случай эндосимбиоза .Ученые предполагают, что миллионы лет назад маленькие свободноживущие прокариоты были поглощены, но не потреблены более крупными прокариотами, возможно, потому, что они были способны противостоять пищеварительным ферментам организма-хозяина. Два организма со временем развили симбиотические отношения: более крупный организм обеспечивает меньший организм достаточным количеством питательных веществ, а меньший организм — молекулами АТФ более крупному. В конце концов, согласно этой точке зрения, более крупный организм превратился в эукариотическую клетку, а меньший организм — в митохондрию.
Митохондриальная ДНК локализована в матриксе, который также содержит множество ферментов, а также рибосомы для синтеза белка. Многие важные метаболические этапы клеточного дыхания катализируются ферментами, которые способны диффундировать через митохондриальный матрикс. Другие белки, участвующие в дыхании, включая фермент, вырабатывающий АТФ, встроены во внутреннюю мембрану митохондрий. Вздутие крист резко увеличивает площадь поверхности, доступную для размещения ферментов, ответственных за клеточное дыхание.
Митохондрии похожи на хлоропласты растений в том, что обе органеллы способны вырабатывать энергию и метаболиты, которые требуются клетке-хозяину. Как обсуждалось выше, митохондрии являются местами дыхания и генерируют химическую энергию в форме АТФ, метаболизируя сахара, жиры и другие химические топлива с помощью молекулярного кислорода. Хлоропласты, напротив, обнаруживаются только в растениях и водорослях и являются основными участками фотосинтеза. Эти органеллы работают по-другому, преобразовывая энергию солнца в биосинтез необходимых органических питательных веществ с использованием углекислого газа и воды.Подобно митохондриям, хлоропласты также содержат собственную ДНК и могут независимо расти и воспроизводиться внутри клетки.
У большинства видов животных митохондрии, по-видимому, в основном наследуются по материнской линии, хотя некоторые недавние данные свидетельствуют о том, что в редких случаях митохондрии могут также передаваться по отцовской линии. Обычно в своем хвосте сперматозоид несет митохондрии в качестве источника энергии на долгом пути к яйцеклетке. Когда сперма прикрепляется к яйцеклетке во время оплодотворения, хвост отваливается.Следовательно, единственные митохондрии, которые обычно получает новый организм, происходят из яйцеклетки, предоставленной его матерью. Следовательно, в отличие от ядерной ДНК, митохондриальная ДНК не перетасовывается в каждом поколении, поэтому предполагается, что она изменяется более медленными темпами, что полезно для изучения эволюции человека. Митохондриальная ДНК также используется в судебной медицине в качестве инструмента для идентификации трупов или частей тела и причастна к ряду генетических заболеваний, таких как болезнь Альцгеймера и диабет.
НАЗАД К СТРУКТУРЕ КЛЕТКИ ЖИВОТНЫХ
НАЗАД К СТРУКТУРЕ ЯЧЕЙКИ
Вопросы или комментарии? Отправить нам письмо.
© 1995-2021, автор — Майкл В. Дэвидсон и Государственный университет Флориды. Все права защищены. Никакие изображения, графика, программное обеспечение, сценарии или апплеты не могут быть воспроизведены или использованы каким-либо образом без разрешения правообладателей. Использование этого веб-сайта означает, что вы соглашаетесь со всеми юридическими положениями и условиями, изложенными владельцами.
Этот веб-сайт обслуживается нашим
Команда разработчиков графики и веб-программирования
в сотрудничестве с оптической микроскопией в Национальной лаборатории сильного магнитного поля
.
Последнее изменение: пятница, 13 ноября 2015 г., 13:18
Счетчик доступа с 1 октября 2000 г .: 2196999
Микроскопы предоставил:
Митохондрии — определение, свойства, структура, функции
Главная »Клеточная биология» Митохондрии — определение, свойства, структура, функции
Свойства митохондрий- Митохондрии — органелла клетки, имеющая нитевидную и зернистую структуру.
- Он присутствует в высших растениях, животных и некоторых микроорганизмах.
- Он отсутствует у бактерий, но обнаруживается в водорослях, простейших и грибах.
- Для осуществления энергетического обмена митохондрии имеют липопротеиновый каркас.
- Он состоит из различных ферментов и коферментов.
- Митохондрии также состоят из специфической ДНК и рибосом.
- Рибосомы участвуют в синтезе белка, тогда как специфическая ДНК участвует в цитоплазматическом наследовании.
- Митохондрии были впервые обнаружены Колликером в 1850 году.
- В поперечно-полосатых мышцах наблюдалась зернистая структура.
- Позже, в 1888 году, он изолировал их от мускулов насекомых.
- В 1882 году Флемминг назвал его fila.
- Раньше митохондриям давали разные названия, такие как
- Фуксинофильные гранулы
- парабазальные тела
- плазмосомы
- пластосомы
- fila
- вермикулы
- биобластов
- хондриосомы
- В цитоплазме митохондрии распределены равномерно.
- Но также обнаружено, что их распространение ограничено во многих клетках.
- В зависимости от функции клетки митохондрии распределяются соответствующим образом.
- Распределение митохондрий зависит от вида и типа клеток.
- Но некоторые клетки могут содержать большое количество митохондрий, например:
- 50,000 в Хаос хаос
- От 140 000 до 150 000 яиц морских ежей
- 300000 в ооцитах земноводных.
- Обнаружено, что в клетках печени крысы присутствует всего от 500 до 1600 митохондрий.
- По сравнению с животной клеткой у зеленых растений меньше митохондрий.
- Это потому, что в зеленых растениях присутствует хлоропласт, который выполняет функцию митохондрий.
- В животной клетке саркосомы представляют собой митохондрии, присутствующие в мышечной клетке миокарда. Они многочисленны и крупны.
- Форма митохондрий может быть нитчатой или зернистой.
- В зависимости от физиологического состояния клеток их форма может переходить из одной формы в другую.
- Значит, формы могут быть булавы, ракетки, везикулярной, кольцевой или круглой формы.
- В крысах или первичных сперматоцитах митохондрии гранулярны.
- В клетках печени митохондрии имеют булавовидную форму.
- Митохондрии могут сливаться и разделять клетку, что вызывает изменение формы.
- В течение дня митохондрии могут сливаться в сетчатую структуру в некоторых клетках эвгленоидов.
- Затем он отделяется или распадается в темноте.
- В случае дрожжей в зависимости от различных условий культивирования происходят изменения.
- Размер митохондрий составляет от 0,5 до 2,0 мкм, поэтому их нельзя четко увидеть под световым микроскопом.
- Встречается также иногда длина достигает 7 мкм.
Структура митохондрий
Митохондрии состоят из митохондриальной мембраны и митохондриальной камеры.
1. Митохондриальная мембранаСостоит из двух мембран. Их:
а. Наружная мембрана- Это гладкая мембрана.
- Состоит из 40% липидов и 60% белков .
- Из-за наличия пор или поринов он проницаемый.
- Он состоит из 20% липидов и 80% белков.
- Это избирательно проницаемая мембрана.
- Так как мембрана загнута внутрь и есть кристы, ее называют шероховатой мембраной.
- Кристы — многочисленные пальцевидные выросты.
- В каждой кристе присутствуют частицы, похожие на теннисную ракетку.
- Частицы ранее назывались субъединицами внутренней мембраны, частицами F0-F1, элементарными частицами или оксисомами.
- В 1963 году Парсонс назвал его частицами переноса электронов (ETP).
- В каждой митохондрии присутствует около 104-105 частиц.
- Основание, стебель и голова присутствуют в каждой элементарной частице.
- Основание и голова также называются частицами F0 и F1 соответственно.
- Стебель действует как звено, соединяющее основание и голову.
- Основание состоит из гидрофобных белков и встроено в липидные молекулы мембраны.
- Голова состоит из пяти типов полипептида.
- АТФаза или АТФ-синтетаза — это присутствующий в нем фермент.
- АДФ помогает в образовании АТФ.
- Аналогичным образом образуется неорганический фосфат. Это связано с окислительным фосфорилированием.
- Стебель также состоит из факторов связи, которые связывают дыхательную цепь с элементарной частицей.
В митохондриях присутствуют две камеры, то есть внешняя и внутренняя камеры.
а. Наружная камера (перимитохондриальное пространство)- Между внешней и внутренней мембранами митохондрий существует пространство между ними, известное как перимитохондриальное пространство.
- В жидкости, содержащейся в ней, также присутствует небольшое количество ферментов.
- Находится во внутренней части внутренней мембраны.
- В нем присутствует полужидкая матрица, которая состоит из:
- Вода
- Минералы
- Белковые частицы
- Рибосомы 70-х годов
- РНК
- Круговая ДНК
- Ферменты
- Митохондрии накапливают и выделяют энергию в форме АТФ (аденозинтрифосфат).Это происходит за счет окисления углеводов, белков и жиров. В дальнейшем он будет использоваться в различных метаболических процессах. Итак, митохондрии известны как электростанция клетки или аккумуляторные батареи клетки.
- Митохондрии помогают в образовании гема гемоглобина.
- Во время клеточного дыхания митохондрии образуют различные промежуточные продукты. Они используются для синтеза цитохромов, хлорофилла, ферредоксина, стероидов, алкалоидов, пиримидинов и др.
- Кальций может накапливаться и высвобождаться митохондриями.
- Помогает в образовании аминокислот.
- В матрице митохондрий можно синтезировать несколько жирных кислот.
- В процессе оогенеза они помогают формированию желтка.
- В процессе сперматогенеза они помогают формированию средней части сперматозоидов.
- В процессе наследования по материнской линии признаки передаются митохондриями напрямую от матери к потомству.
- Митохондрии также присутствуют в клетке печени. Они помогают детоксикации аммиака с помощью своих ферментов.
- Митохондрии являются местом генерации тепла, известного как термогенез.
- Иногда может произойти ненормальная гибель клетки. Это может быть связано с нарушением работы митохондрий. Это может повлиять на функцию органа.
- Он помогает в образовании некоторых частей гормона тестостерона и эстрогена.
- Шакья, М., Mehata, K.R., Gautam, M.K., Pokhrel, K.R., and Khanal, K. (2020) «Принципы биологии», Asmita Books Publisher and Distributors Ltd, Бхотахити, Непал
- Верма, П. С., Агравал, В. К. (2006). Клеточная биология, генетика, молекулярная биология, эволюция и экология (1-е изд.). С.Чэнд энд Компани Лтд.
- https://kullabs.com/class-11/biology11/cell-organelles/mitochondria
- https://alevelbiology.co.uk/notes/mitochondria-structure-and-functions/
- https: // www.biologydiscussion.com/cell/plant-cell/structure-of-a-plant-cell-with-diagram/68806 — 12%
- https://biologyteach.com/power-house-of-the-cell/- 11%
- https://www.sciencedirect.com/topics/agricultural-and-biological-sciences/pavement-cells- 4%
- https://www.sciencefacts.net/mitochondria.html- 2%
- https://biologywise.com/mitochondria-structure-functions- 1%
- https://www.sciencedirect.com/science/article/pii/S0005272810000058- 1%
Bid поддерживает структуру и функцию митохондриальных крист, а также защищает от сердечных заболеваний в рамках интегративного исследования геномики
[Примечание редакции: ответы автора на первый раунд рецензирования приводятся ниже.]
Рецензент № 1:
На Рисунке 1 авторы упоминают метаболический стресс, но тестируют только весьма экстремальные клеточные стрессы, такие как синдром отмены сыворотки. Чтобы лучше поддержать утверждения, рекомендуется более физиологический стресс, такой как перевод клеток в галактозу вместо глюкозной среды, чтобы управлять Ox / Phos. Различия на рисунке 1I с носителем и дигитонином между клетками с Bid и без него могут быть статистически значимыми, но не убедительными.
Поскольку основное внимание в рукописи уделяется метаболизму сердца, который мы характеризуем с помощью специфических субстратов респираторного комплекса (теперь с увеличением числа мышей, рис. 6E и F), мы удалили эти данные из рукописи.
Мы согласны с тем, что изъятие сыворотки действительно является экстремальным клеточным стрессом. Мы обнаружили, что миелоидные клетки-предшественники очень метаболически пластичны (обзор в Hsu and Qu, (2013)) и могут использовать несколько источников углерода, включая галактозу, в присутствии 10% сыворотки. Также было показано, что галактоза не обязательно управляет метаболизмом митохондрий (Elkalaf et al., 2013), и ее влияние на метаболизм митохондрий зависит от типа клеток, и, таким образом, метаболический статус миелоидных клеток больше не имеет отношения к измененной рукописи.
Что касается анализа мобилизации цитохрома с, мы согласны с тем, что этот пункт, хотя и свидетельствует о дефекте крист, не является важным наблюдением и был удален из этой версии рукописи, чтобы сохранить точные результаты.
Следует отметить, что мы не ожидали большой разницы при использовании дигитонина. Для высвобождения значительного количества цитохрома с необходимо использовать апоптотический tBid. (Скоррано и др., 2002).
Интересны данные EM на Рисунке 2A.Было бы важно подтвердить это, исследуя фиксированные и взятые у мышей клетки в ткани напрямую, без культивирования. Есть ли что-то особенное в миелоидных клетках-предшественниках или Bid регулирует морфологию крист в других типах клеток? В частности, авторы должны исследовать сердце (с лечением адреналином и без него), так как этому в рукописи будет уделено особое внимание.
Теперь мы проанализировали митохондрии сердца с помощью ЭМ у мышей WT и Bid — / — в присутствии и в отсутствие адреналина.Мы включили эти новые данные в рисунки 4A и B. Мы сделали несколько важных наблюдений.
Наблюдается явная потеря организации митохондрий в сердечной ткани Bid — / — . В то время как митохондрии в сердечной ткани дикого типа локализованы в областях между линейными кардиомиоцитами, митохондрии в сердечной ткани Bid — / — обнаруживаются разбросанными по всей ткани прерывистым и дезорганизованным образом.
Во-вторых, существует разительная разница в морфологии крист как до, так и после введения адреналина (18-часовое воздействие при 0.5 мг / кг). Мы оценили это, измерив ширину крист. Мы обнаружили, что в необработанной ткани митохондрии Bid — / — обычно менее электронно-плотные и имеют более округлую форму. После обработки митохондрии Bid — / — становятся менее электронно-плотными и поврежденными. Митохондрии WT, однако, имеют пластинчатую форму и не претерпевают значительных изменений при лечении адреналином. Мы предоставили сильные
доказательств роли Bid в морфологии крист как в MPC, так и в тканях, взятых непосредственно у мышей WT и Bid — / — .
Странно, что экспрессия Puma и Bim снижена в ячейках с нулевым Bid (Рисунок 2 — приложение к рисунку 2C). Это показывает, что плейотропные эффекты в этих клетках могут давать фенотипы, косвенно или далекие от экспрессии Bid как таковой.
Что касается других белков, содержащих только Bh4, мы согласны с тем, что интригует то, что наблюдается снижение экспрессии Bim, Puma и Bad в миелоидных предшественниках Bid — / — . Исследование кардиомиоцитов левого желудочка на наличие этих белков также выявляет снижение экспрессии Bad и Bim (Puma не наблюдалась ни у мышей WT, ни у мышей Bid — / — ).Эти кляксы были добавлены к Фигуре 2 — добавлению к рисунку 2C.
Хотя потеря этих белков может иметь плейотропные эффекты, выходящие за рамки данной рукописи, эти данные демонстрируют, что аномалии крист, наблюдаемые в клетках и тканях Bid — / — , не являются следствием компенсаторной сверхэкспрессии других белков, содержащих только Bh4. Кроме того, мы можем восстановить нормальную морфологию митохондрий, добавив обратно неапоптотический Bid к клеткам Bid — / — . Наконец, теперь у нас есть новые генетические доказательства роли экспрессии BID и сердечных заболеваний.Пациенты, сгруппированные по экспрессии генов, показывают, что пациенты, у которых экспрессия BID находится в пределах 5% самого низкого уровня, обогащены по сердечным фенотипам, включая инфаркт миокарда, в среднем более чем в 4 раза, чем пациенты с ИМ в синтетической производной Университета Вандербильта.
Дополнение к рисунку 1D неадекватно. Требуется большее увеличение. Необходимо показать несколько изображений ячеек. Cytosolic Bid несомненно в некоторой степени перекрывается с митохондриями. FRAP для удаления цитозольного пула Bid может выявить более убедительную митохондриальную локализацию оставшегося Bid.Рисунок 3A лучше и может заменить рисунок в дополнении 1D. Как Bid связывает митохондрии? Обязателен ли МТЧ3? Если это объясняет локализацию, то какое это имеет значение, учитывая данные PrediXcan, показывающие отсутствие связи с MTCH 2 (но влияние BID) на рисунке 6B?
Мы использовали биохимический подход, чтобы окончательно продемонстрировать локализацию полноразмерных митохондрий Bid. На рисунке 3 мы теперь включаем следующие результаты:
Выделение неочищенных митохондрий и митохондрий, очищенных перколлом, из печени (для удаления легкого загрязнения мембран), выявляет локализацию полноразмерного Bid в отсутствие апоптоза (рис. 3B).
Обработка изолированных митохондрий печени протеиназой K (PK)подтверждает достоверность субмитохондриальной локализации Bid независимым методом. Эти результаты также показывают Bid в митохондриях даже после обработки протеиназой K в условиях, в которых мы наблюдаем деградацию Bak (рис. 3C).
Используя технику субмитохондриального фракционирования, которая использует преимущество чувствительности митохондрий к гипотоническому стрессу, мы можем изолировать внешнюю мембрану (OMM) от митопласта (внутренней мембраны и матрикса).Мы демонстрируем Bid как в OMM, так и во фракции митопластов, что согласуется с нашими результатами PK. Таким образом, мы показываем гомеостатическую локализацию Bid в митохондриях и находим пул Bid в матриксе, который может играть роль в структуре крист.
Что касается MTCh3, хотя есть литература, подтверждающая роль MTCh3 в качестве док-платформы для апоптотического tBid (Zaltsman et al., 2010), еще предстоит определить, является ли полноразмерный Bid (или фосфорилированный мутант Bid S61AS78A (AA) , который, как полагают, локализован в митохондриях) ассоциирует с MTCh3.Кроме того, известно, что Bid также ассоциируется с митохондриальным липидом кардиолипином в качестве стыковочной платформы (Kim et al., 2004). Мы предполагаем, что способность Бида стабилизировать кристы частично зависит от его матричной локализации, и утверждаем, что MTCh3 не важен для этого процесса.
В литературе мы обнаружили, что исследования MTCh3, проведенные лабораторией Гросса, демонстрируют, что модели мышей Bid AA и MTCh3 F / F Vav1-Cre + демонстрируют потерю гемопоэтических стволовых клеток (покой HSC), и, исходя из этих данных, они пришли к выводу, что при расширении обе модели будут иметь сопоставимые изменения в митохондриях (Марьянович и др., 2015). Хотя это возможно, нет прямых доказательств того, что структурные изменения, наблюдаемые в митохондриях, опосредованы взаимодействием Bid с MTCh3. Действительно, нет данных, демонстрирующих неапоптотическую функцию взаимодействия Bid / MTCh3. Следовательно, никаких структурно-функциональных исследований не проводилось, и, таким образом, минимальный связывающий домен для взаимодействия между Bid и MTCh3 неизвестен.
Наконец, как отметил автор обзора, мы не находим генетических доказательств роли MTCh3 как на уровне наследственности, так и какой-либо связи с экспрессией и инфарктом миокарда.Однако мы обнаруживаем связь между повышенной экспрессией гена MTCh3 и повышенным ИМТ, что подтверждается данными, ранее найденными в литературе (Bar-Lev et al., 2016).
Принимая во внимание эти наблюдения, а также наш новый результат, показывающий Bid в матрице (а также связанный с MCl-1, обсуждаемый ниже), мы не уделяли внимания обсуждению MTCh3 в рукописи.
Как Bid регулирует морфологию крист, остается неясным и ограничивает значимость этого исследования.Авторы находят снижение активности Комплекса I в сердечной ткани Bid KO и димеры более низкой АТФ-синтазы, которые могут объяснить изменение морфологии крист. Но в равной степени возможно и то, что изменение морфологии крист может опосредовать изменение Комплекса V. Таким образом, показана только корреляция, а механизм не раскрыт.
В свете наших новых данных, предполагающих, что Bid может локализоваться в митохондриальном матриксе, мы проверили, может ли Bid взаимодействовать с белком BCl-2 MCl-1. Было продемонстрировано, что MCl-1 локализуется в митохондриальном матриксе и влияет на структуру крист, а также на сердечную функцию, включая кардиомиопатию (Wang et al., 2012 и Thomas et al., 2013).
Мы представляем данные, демонстрирующие коиммунопреципитацию WT-Bid (FlagHA tagged-Bid) с WT, внешней мембраной (MCl-1 OM ) и матриксной локализацией (MCl-1 Matrix ) форм MCl-1. Это подтверждает потенциальную роль Bid в митохондриальном матриксе через MCl-1 (рис. 9a). Взаимодействие Бида с MCl-1 Matrix особенно интересно, поскольку другой белок, содержащий только Bh4, Bim, не взаимодействует с матричным MCl-1 (Perciavalle et al., 2012). Мы предполагаем, что потеря Bid может косвенно влиять на функцию, субмитохондриальную локализацию или белок-белковые взаимодействия MCl-1, влияя на структуру крист.
Мы также показываем, что мутировавший M148T Bid (наша потеря функции SNP, выявленная у пациентов с BioVU) снижает взаимодействие с MCl-1 Matrix по сравнению как с WT, так и с Bid D59A (рис. 9B). Это соответствует потере опосредованной Bid структуры крист в присутствии Bid M148T (новые данные), обеспечивая дополнительные доказательства роли спирали 6 Bid в регуляции структуры крист через ассоциацию с MCl-1.
Спасает ли M148T Bid морфологию крист в Bid KO-клетках? ПДК или сердце?
Мы показываем генетически, что в когорте BioVU статус носителя Bid SNP M148T увеличивает ассоциацию с MI. Когда эта точечная мутация производится и сочетается с Bh4, мутировавшим Bid (спасательный мутант), мы не можем восстановить дыхание MPC.
В соответствии с этим открытием, новые данные EM от MPC (рис. 8E-G) показывают, что этот двойной мутант приводит к значительному уменьшению количества крист по сравнению с Bid , только Bh4 (в Bid — / — клетках), показывая, что Bid M148T приводит к потере митохондриальной структуры в дополнение к ранее показанной функции.Это также подтверждается нашими выводами для MCl-1.
В целом данных много, и митохондрии Bid KO кажутся отличными от WT. Но как это происходит, и значение изменений непонятно. Также неясно, влияют ли морфология митохондрий и биохимические изменения на результаты популяционных исследований. Главный урок заключается в том, что Bid, по-видимому, имеет функции, выходящие за рамки регуляции апоптоза, но что это такое, неизвестно.
Теперь мы предоставили дополнительные данные для рассмотрения потенциального биологического механизма роли полноразмерного Bid в митохондриях.Кроме того, теперь мы можем более прочно связать мышиную модель с генетикой человека как с дополнительными экспериментальными данными, так и с новым генетическим подходом.
Мы расширили наш анализ эхокардиограммы мышей WT и Bid — / — . Теперь у нас есть дополнительные данные на исходном уровне (без лечения), через 18 часов после приема адреналина и в новой временной точке восстановления (через 5 дней после лечения). Эти данные показывают снижение фракции выброса, а также увеличение конечного диастолического и конечного систолического объемов — параметров, которые наблюдаются в условиях инфаркта миокарда и напрямую связаны с плохим прогнозом пациента после инфаркта миокарда (Di Bella et al., 2013 и White et al., 1987) (рис. 4C-G).
Мы также оценили фиброз в нашей модели. После повреждения миокарда, такого как при инфаркте миокарда, миокард подвергается ремоделированию для восстановления повреждений. Это ремоделирование проявляется в виде фиброза, который можно использовать как показатель повреждения миокарда. (Талман и др., 2016). Используя окрашивание трихромом, мы определили степень фиброза как через 18 часов, так и через точки восстановления, используя объективный количественный алгоритм (см. Раздел «Материалы и методы»).Мы обнаружили повышенное фиброзное повреждение у мышей Bid — / — по сравнению с мышами WT, что напрямую связывает наши результаты с наблюдениями за пациентами (рис. 5).
Теперь мы находим Bid в митохондриальном матриксе и обнаруживаем связь между Bid и формой митохондриального матрикса MCl-1, другого члена семейства BCl-2, который, как сообщается, локализуется в митохондриальном матриксе и влияет на структуру крист. Мы обнаружили, что эта ассоциация частично зависит от интактной спирали 6 Bid, таким образом подразумевая связь с MCl-1 Matrix в роли Bid в регулировании структуры крист.
Мы использовали генетический подход, чтобы определить, существует ли связь между 41 геном, который, как известно, участвует в поддержании структуры крист и инфаркте миокарда. Анализ этих генов с помощью PrediXcan выявил снижение экспрессии двух генов, связанных с ИМ; MCL-1 и MTX1 (метаксин-1), митохондриальный переносчик, который, как известно, связан с участком контакта митохондрий и системой организации крист (MICOS). Таким образом, мы задействовали MCl-1 как взаимодействующего партнера с Bid, а также как связанный с ИМ на генетическом уровне.Мы также включили структуру митохондриальных крист в восприимчивость к ИМ.
Мы также воспользовались синтетической производной BioVU, которая содержит более 2,8 миллионов деидентифицированных историй болезни. Мы составили таблицу непредвиденных обстоятельств, чтобы определить относительный риск ИМ с кардиомиопатией, поскольку потеря MCl-1 для сердца приводит к кардиомиопатии (Wang et al., 2012 и Thomas et al., 2013). Мы находим очень значимую оценку относительного риска для этой ассоциации по сравнению с известными факторами риска ИМ, такими как гипертония (рис. 9Dd.Таким образом, можно предположить, что сердечное заболевание митохондриальной этиологии является значительным фактором риска ИМ.
Наконец, как упоминалось выше, мы также существенно усилили связь между экспрессией BID и данными от клеток и мышей. В частности, мы идентифицировали пациентов в BioVU с наименьшими 5% экспрессии BID, максимально имитируя фенотип генетического нокаута. Мы обнаружили, что ИМ у этих людей в> 4 раза выше, чем у пациентов с остальной частью синтетического производного Вандербильта.Эти новые данные существенно усиливают связь между данными о мышах и людях.
Рецензент № 2:
Эта рукопись предоставляет интересные данные, показывающие, что полноразмерный Bid модулирует митохондриальные кристы и дыхание в гомеостатических условиях независимо от расщепления каспазой-8 (D59A) и ее Bh4-домена. Дополнительные данные с использованием генного подхода, примененного к биобанку, подтвержденные в независимом крупномасштабном GWAS, показали, что снижение экспрессии гена BID коррелирует с инфарктом миокарда (ИМ).Статус носителя с несинонимичными вариациями, включая M148T, в мембранно-связывающем домене Bid также ассоциируется с предрасположенностью к ИМ.
1) Во введении упоминается, что существует связь между выражением Bid и MI. Может ли быть, что MI приводит к потере Bid, а не BID, регулирующий MI?
Подход, который мы использовали с PrediXcan, напрямую решает эту серьезную проблему. PrediXcan анализирует влияние изменений зародышевой линии, таких как полиморфизм, на экспрессию генов.Мы исключаем как воздействие окружающей среды, так и влияние самой болезни на экспрессию гена. Эти условные значения генетически детерминированной экспрессии основаны на эталонном транскриптоме, таком как проект GenotypeTissue Expression (GTEx), который генотипирует доноров ткани и связывает это с генотипированием с экспрессией гена (мРНК). Мы обнаружили, что когда PrediXcan применяется к BioVU, а также к когортам с множественной репликацией, снижение генетического компонента экспрессии гена BID связано с ИМ, что позволяет предположить, что направление идет от BID к MI.
2) В подразделе «Bid — / — клетки имеют дефект крист, который можно исправить с помощью Bid с мутацией Bh4 или D59» упоминается, что субклеточное фракционирование показывает, что Bid находится в митохондриальной фракции. Были проведены контроли для проверки наличия загрязнений от других фракций. Были ли митохондрии обработаны протеазой, чтобы определить, был ли Bid прикреплен к митохондриям, а не в митохондриальном матриксе. Это важные элементы управления.
Мы значительно улучшили биохимический анализ Bid в митохондриях.На обновленном Рисунке 3 мы приводим несколько линий дополнительных доказательств.
Сюда входят:
Как сырые, так и очищенные перколлом митохондрии, выделенные из печени мыши (без легкого загрязнения мембран), выявляют присутствие полноразмерного Bid (рис. 3B).
Обработка митохондрий протеиназой К, показывающая присутствие Bid во фракции, обработанной протеазой, в условиях, в которых Bak удаляется (разлагается) (рис. 3C).
Субмитохондриальное фракционирование с использованием осмотического метода сжатия-набухания, отделяющего внешнюю мембрану (OMM) от митопласта (внутренней мембраны и матрикса).Мы обнаруживаем, что фракция Bid присутствует во фракции, обогащенной митопластами.
Наконец, мы совместно иммунопреципитировали FlagHA-tagged Bid с WT, внешней митохондриальной мембраной и матриксными формами члена семейства BCl-2 MCl-1. Mcl1, как было установлено, играет роль в структуре митохондриальных крист (Perciavalle et al., 2012) и сердечной дисфункции (Wang et al., 2012 и Thomas et al., 2013). Кроме того, мы обнаружили, что Bid M148T уменьшил взаимодействие с MCl-1 Matrix по сравнению с WT или Bid D59A .
Это дополнительное свидетельство предполагает, что действительно существует часть полноразмерного Bid, которая присутствует не только на внешней митохондриальной мембране, но и в матриксе митохондрий. Бид может ассоциироваться с матричной формой MCl-1 и участвует в регуляции митохондриальных крист.
3) Сделан вывод, что острое лечение адреналином (30 мкг) приводит к увеличению LVIDd и LVID. Было бы полезно указать график изменений.Значения n также низкие для этих исследований.
Хотя мы действительно наблюдаем увеличение LVIDd / s при острой дозе 30 мкг адреналина, мы не видим функции снижения других сердечных параметров, вероятно, поскольку этому повреждению требуется время, чтобы проявиться. Мы согласны с тем, что эти данные можно улучшить с помощью большего количества повторов. Чтобы наилучшим образом укрепить эти результаты, мы сосредоточились на получении дополнительных повторов с измерениями, сделанными на исходном уровне, через 18 часов после адреналина (в дозе 0,5 мг / кг), а также через 5 дней после адреналина для оценки восстановления.
Теперь мы добавили значительно больше мышей для полного анализа перед адреналином WT n = 12 и Bid — / — n = 12), в 18-часовой временной точке (WT n = 12 и Bid — / — n = 11), а время восстановления — через 5 дней (WT n = 5 и Bid — / — n = 6). Через 18 часов после эпитаксии мы видим увеличение LVIDd, а также LVID, а также снижение EF и тенденцию к снижению FS (p = 0,068) для мышей Bid — / — . Кроме того, мы наблюдаем увеличение конечного диастолического и конечного систолического объемов.Эти параметры напрямую связаны с плохим прогнозом пациента после инфаркта миокарда (Di Bella et al., 2013 и White et al., 1987) (рис. 4C-G).
4) Представлены данные, показывающие, что через 18 часов после введения адреналина в дозе 0,5 мг / кг наблюдается увеличение LVIDd и LVID. Это также привело к снижению FS и EF. Сохраняется ли это снижение EF и FS или оно восстанавливается? Требуется более подробная характеристика.
Интересно, что через 5 дней, момент времени восстановления, мы наблюдаем, что мыши Bid — / — функционально возвращаются к базовому параметру и не отличаются от мышей WT.Однако нам было интересно узнать, если бы это было похоже на фиброзное повреждение, наблюдаемое у пациентов после инфаркта миокарда (Talman et al., 2016), мы бы наблюдали усиление фиброза в сердцах Bid — / — после введения адреналина. Используя окрашивание трихромом, мы измерили степень фиброза как через 18 часов, так и через 5 дней, используя количественный алгоритм. Мы обнаружили повышенное фиброзное повреждение у мышей Bid — / — по сравнению с мышами WT в оба момента времени, что напрямую связывает наши результаты с наблюдениями за пациентами с ИМ (рис. 5).
5) Описание анализа PrediXcan и некоторых других биоинформатических анализов немного длинное и может быть сокращено.
Мы ценим возможность более краткого описания генетического анализа. Мы рассмотрели эту проблему и существенно изменили этот раздел, чтобы подчеркнуть наши наиболее важные выводы. Мы объединили анализ на уровне экзома в раздел сайт-направленного мутагенеза, чтобы сделать раздел генетики более кратким и подчеркнуть функциональный компонент валидации наших исследований.Далее мы объединили анализ другого семейства BCl-2 с первичным анализом PrediXcan, чтобы подчеркнуть уникальную роль Bid. Дополнительные сведения о семействе BCl-2 теперь представлены в новом разделе дополнительной информации.
6) N для суперкомплексных данных низкие (n = 3). Я не считаю данные на рис. 3F убедительными, особенно с учетом низкого значения n. Кроме того, рисунок 2 — рисунок в приложении 2 не показывает каких-либо существенных различий ни по одному из сложных видов деятельности. Кроме того, в подразделе «Комплекс I, содержащий суперкомплексы, проявляется пониженная активность в митохондриях сердца Bid — / -», говорится о значительном снижении активности комплекса I внутри SC (рис. 3F).Однако на рис. 3F рассматривается плотность полосы частот, а не активность. Активность измеряется на дополнительном рисунке, но не показывает изменений в активности.
Что касается количества прогонов для этого анализа, в частности CI, мы должны уточнить, что представленные данные являются репрезентативными для 4 независимых экспериментов. Однако мы представляем количественный анализ из одного репрезентативного эксперимента (митохондрии, выделенные от 3 мышей WT и 3 Bid — / — мышей). Поскольку на ферментативную активность могут влиять небольшие изменения температуры и времени, в течение которых фермент подвергается воздействию субстрата, наш анализ проводится на митохондриях, прогоняемых на одном геле, для точности и точности.
Все данные, первоначально представленные на Рисунке 3, а также на Рисунке 2, в приложении 2 к рисунку измеряют ферментативную активность. Мы ценим непонятную оригинальную маркировку. Чтобы решить эту проблему, мы уточнили маркировку.
На нашем исходном рисунке 3 мы представили результаты активности в геле (IGA) для респираторного комплекса I и АТФ-синтазы (Комплекс V) из изолированных митохондрий сердца. Каждый тестируемый комплекс имеет окрашенный кумасси гель рядом с соответствующим анализом активности.Анализы активности основаны на колориметрическом изменении, которое происходит, когда активный фермент в геле инкубируется в присутствии его соответствующего субстрата (НАДН для CI и АТФ для CV и цитохрома c для CIV). Затем мы измеряем плотность полосы как показатель активности: чем активнее фермент, тем сильнее изменение цвета и, следовательно, тем плотнее полоса. Во всех анализах активности в геле ферментативная активность измеряется путем определения плотности полос.
Мы также сосредоточили наши результаты на активности АТФ-синтазы.Данные об активности в геле теперь поддерживаются дополнительными повторами анализа синтеза АТФ в реальном времени из ткани сердца, и эти данные теперь представлены вместе на рисунке 6. Анализ активности в геле для CI и CIV теперь представлен на рисунке 6 — добавление к рисунку 1.
Рецензент № 3:
Salsbury-Ruf et al. Сообщают, что потеря Bid приводит к повышенной гибели клеток (вопреки известной роли Bid), дефектам митохондриальных крист и снижению дыхания на клеточном уровне. У мышей с нокаутом они обнаруживают нарушение функции сердца при введении лекарств.Они находят доказательства на человеческом уровне, что более низкие генетически обусловленные уровни BID связаны с учащением инфаркта миокарда, используя данные своего биобанка (BioVU). Наконец, они обнаружили, что избыточные вариации в кодирующей области BID также связаны с инфарктом миокарда.
Результаты на клетках и мышах кажутся убедительными. Я оставлю обзор экспериментальной установки и реализации другим рецензентам, более знакомым с методами и модельными системами. Читая, я не обнаружил никаких очевидных статистических проблем с анализом этого первого раздела.
Связь между BID и риском инфаркта миокарда в Biovu, основанном на электронных медицинских картах Вандербильта, также убедительна. Однако обоснование выбора 9 салфеток из 50 в GTEx было менее ясным. На рисунках показаны значения p, но ткань четко не указана. Каждая модель ткани дает разные значения p. Обоснование выбора тканей должно быть объяснено, а цифры должны четко обозначать, какой тканевый результат представлен.
Теперь мы реорганизовали представление результатов нашего открытия и репликации, чтобы сосредоточиться на ассоциации в наиболее важной ткани, сердце (для полноты, результаты в тканях вне сердца были перемещены в новый раздел дополнительной информации).
Наши оригинальные анализы проводились в 9 тканях. Важно отметить, что существует довольно много общей генетической архитектуры экспрессии генов между тканями (GTEx Consortium, 2017), и поэтому возможно, что даже если «причинные» или «патогенные» ткани или типы клеток недоступны, мы могли бы использовать эту общую архитектуру для выявления генетических ассоциаций с заболеванием (Gamazon et al., 2018). И, конечно же, не все ткани могут быть тканями, вызывающими интересующее нас заболевание.Например, неясно, имеют ли некоторые репродуктивные ткани в GTEx отношение к ИМ. Наконец, не все ткани имеют модели прогнозирования с достаточно высоким качеством из-за размера выборки для тканей и / или из-за неоднородности размеров генетических эффектов. Однако мы наблюдаем связь с ИМ в сердечной ткани.
Анализ наследуемости выполняется для предварительной фильтрации исследуемых генов. Однако при анализе используются модели прогнозов из разных тканей.Последнее решение могло бы быть оправданным, если бы модели прогнозирования давали перекрестно подтвержденную значимость, что свидетельствует о генетической основе признака экспрессии гена. В этом случае предварительная фильтрация генов по наследственности в разных тканях будет ненужной.
Мы благодарим рецензента за возможность уточнить этот анализ. Предоставленные нами оценки наследуемости (из DGN) согласуются с условным условием R2 из моделей прогнозирования (в ткани сердца GTEx).В этом случае предварительная фильтрация, основанная на наследуемости или качестве вменения, даст те же результаты. Мы решили сообщить о наследуемости в DGN из-за гораздо большего размера выборки (n = 922). Мы пояснили это в тексте следующим образом: «Более того, в соответствии с оценками наследуемости в DGN, экспрессия MTCh3 в сердце не может быть хорошо вменена с использованием генетической вариации из цис-области гена […]».
Приятно видеть, что BID было связано, по крайней мере, с умеренным значением в сердечной ткани в КАРДИОГРАММЕ.Но как насчет других тканей? Были ли они незначительными?
Мы считаем, что сердце является наиболее важной тканью из всех исследуемых тканей GTEx. Ср
изменили представление, чтобы выделить эту ткань. Мы включили другие ткани для полноты картины. Мы подчеркиваем, что у нас есть значительные предварительные доказательства роли BID в сердечных заболеваниях из наших функциональных исследований (на мышах и клетках), которые теперь выделены в рабочем процессе (рис. 1). Мы не пытаемся беспристрастно исследовать геном в поисках генов, связанных с заболеванием.Скорее, мы воспроизводим в исследованиях на людях то, что мы в значительной степени наблюдаем у мышей и клеток. Кроме того, наше исследование пытается выяснить механизм роли BID в сердечных заболеваниях человека и в регуляции структуры крист.
Намного больший размер выборки в исследовании CARDIOGRAM не привел к повышению значимости, что кажется немного тревожным. Аргумент об однородности этнической принадлежности в BioVU vs CARDIOGRAM звучит несколько правдоподобно, но не до конца убедительно. Образцы GTEx не так однородны, как подразумевается в тексте.
Мы считаем, что неоднородность этнической принадлежности может существенно повлиять на результаты. Фактически, доля пациентов неевропейского происхождения в
Набор данныхCARDIOGRAM (23%) существенно выше, чем доля в GTEx (10%). Мы также подчеркиваем, что наше исследование не было объективным поиском гена, связанного с инфарктом миокарда, однако оно было мотивировано нашими обширными функциональными исследованиями Bid на мышах, которые уже продемонстрировали вызванный стрессом фиброз, дисфункцию миокарда и структурные нарушения митохондрий.
Приятно видеть тиражирование данных британского биобанка. Однако почему на рисунке 7 C показаны только 3 ткани (кожа, большеберцовый нерв, мозг-мозжечок)? Были ли они выбраны за то, что они значительны?
Мы изменили представление наших результатов. Теперь мы сосредоточимся на соответствующей ткани — сердце. Для полноты мы представляем результаты для тканей, не относящихся к сердцу, и переместили эти результаты в дополнительную информацию и на дополнительный рисунок 7 — приложение к рисунку 3C.
Уточнение этих моментов необходимо, чтобы прояснить, что не было никакого отбора из представленных тканей и ассоциаций.
См. Выше. Поскольку теперь мы сосредоточили наши результаты на наиболее подходящей ткани, сердце, теперь ясно, что дополнительные не относящиеся к сердцу ткани были включены только для полноты. Примечательно, что наши выводы не зависят от включения этих несердечных тканей.
[Примечание редакции: ответы автора на повторную рецензию приводятся ниже.]
Рукопись была улучшена, но остаются некоторые проблемы, которые необходимо решить перед принятием, как указано ниже:
Рецензент № 2:
Раздел результатов
Я считаю предложение о возможности интеграции результатов в клиническое применение неясным и расплывчатым.
Мы понимаем, что это предложение было неясным в отношении потенциальных клинических применений наших результатов. Нашим намерением было предложить идею о том, что генетический подход, который мы использовали здесь, в частности PrediXcan, можно было бы использовать как способ клинической проверки предрасположенности к болезням.Анализ PrediXcan основан на эталонном транскриптоме, который можно применить к биобанку или набору данных GWAS. Этот транскриптом основан на SNP от здоровых людей, и эти SNP имеют высокую частоту аллелей в общей популяции. Таким образом, вероятно, что аллели риска в этих SNP будут часто обнаруживаться у пациентов. Если конкретная комбинация SNP приводит к изменениям экспрессии гена, связанного с заболеванием, то влияние на риск заболевания можно оценить, и, следовательно, в некоторых случаях уход за пациентом может отражать этот потенциальный риск.
Тем не менее, мы чувствуем, что это утверждение может быть достигнуто, поскольку проспективное исследование, проверяющее эту идею, может стать следующим подходящим шагом в интеграции этого типа анализа в клинику. Мы удалили это предложение из Введения в свете этого и понимаем, что этот тип широкого применения еще не определен.
Подраздел «Анализ PrediXcan выявляет снижение экспрессии BID, связанное с инфарктом миокарда». Почему наследственность BECN1 согласуется с известной защитной ролью аутофагии и сердечно-сосудистых заболеваний?
В рукописи больше нет этой связи.Мы удалили это из подраздела «Анализ PrediXcan выявляет снижение экспрессии BID, связанное с инфарктом миокарда» в текущей версии.
Наш анализ наследуемости показал, что в дополнение к BID, BECN1 был в значительной степени наследуемым и, таким образом, как BID, также находился под генетическим контролем. Первоначально мы заявили, что это было интересно, поскольку это «соответствует защитной роли аутофагии и сердечно-сосудистых заболеваний». Мы полностью согласны с рецензентом в том, что одна только наследуемость не связывает эти два наблюдения.
Нашей первоначальной целью было передать идею о существовании ранее известной связи между ролью Beclin1 в аутофагии и защитой от сердечно-сосудистых заболеваний. Это является уникальным для Beclin1 среди белков, оцениваемых в нашем вторичном анализе, поскольку ранее не было известно роли (насколько нам известно) этих белков в защите от сердечных заболеваний или инфаркта миокарда. Однако мы согласны с тем, что наследуемость не отражает эту роль для Beclin1.
Фактически, анализ BECN1 с помощью PrediXcan (Рисунок 7 — приложение к рисунку 7A) показывает связь с фенотипами сердца.Мы обнаружили, что значительно повышенная генетически обусловленная экспрессия связана с сердечной недостаточностью, тогда как снижение экспрессии связано с сердечным шунтом и первичной кардиомиопатией. Таким образом, ассоциация BECN1 с сердечной недостаточностью, вероятно, обусловлена генетической изменчивостью.
Подраздел «Проверка в BioVU и CARDIoGRAMplusC4D GWAS» Я не нахожу ткань сердца, как упоминалось здесь: «В отдельном наборе образцов BioVU (см. Раздел« Материалы и методы »и рисунок 7 — приложение к рисунку 6c и d) мы наблюдали значительную корреляция (p = 0.002) между снижением генетически детерминированной экспрессии BID в сердечной ткани и инфарктом миокарда ».
Мы ценим возможность уточнить наши генетические данные, представленные на рисунке 7 — рисунок в приложении 6C и D. Мы обнаружили значительную корреляцию со снижением экспрессии BID (p = 0,002) с ИМ в аорте. Мы разъяснили это в Подразделе «Валидация в BioVU и CARDIoGRAMplusC4D GWAS» настоящего проекта, указав, что ассоциация была обнаружена в аорте.
https: // doi.org / 10.7554 / eLife.40907.043 .