
Раздается, раздаваться: как правильно
Данные слова сложны в написании тем, что содержат две вызывающие сомнение гласные: в приставке и корне слова. Что касается инфинитива, можно ошибиться с «ь» в конце. Но написание слов раздается раздаваться не нужно запоминать: все «сомнительные» буквы возможно проверить по правилам.
Написание приставки
И в «раздается», и в «раздаваться» нужна приставка «раз». Объясняется тем, что приставки «рос», «роз» или «рас» в безударном положении в русском языке нет. Это касается «раздаваться». Проверочное слово здесь не требуется: это то правило, которое нужно запомнить.
Но нужно иметь ввиду, что в ударном положении приставки оно может не действовать: «рассыпал», но «россыпи». То же самое в отношении рассматриваемого глагола: «раздавать», но «розданный». Приставка «роз», потому что она находится под ударением («розданный товар»). Такая форма слова не будет определяющей, так как приставки не проверяются ударением.
В случае с сомнением насчет «з» или «с» в приставке, обращаются к другому правилу:
- Приставка оканчивается на «з», если после нее следует звонкая или сонорная согласная.

- Приставка оканчивается на «с», если после нее – глухая, шипящая согласная.
- После приставки следует «д» — звонкая согласная. Значит, правильным будет «раз»: «раздается крик», «раздаваться в тишине».
Написание корня
Корень в обоих словах – «да». Гласная «а» — в безударном положении. С ее правильным написанием можно определиться, подобрав однокоренную словоформу, родственную «раздаваться». Проверочное слово:
- Дать, подать, раздать.
- Подача, сдача, раздача.
- Раздай, сдай, отдай, подай.
Как написать «раздаваться»? Проверочное слово помогает следующим образом:
- «Крик непонятной птицы снова раздается в безмолвном лесу» (раздается — раздать).
- «Отголоски воя сирены еще долго будут раздаваться над застывшим в тревожном ожидании городом» (раздаваться – раздача).
- «Шум вновь раздается, хоть мы уже несколько раз просили их соблюдать тишину» (раздается – раздать).
- «Каждый раз при приближении посторонних будет раздаваться звук сигнализации» (раздаваться – дать).

- «Сегодня на благотворительной ярмарке раздается полевая гречневая каша» (раздается – раздача).
- «Раздаваться в вышине»: поверочное слово «раздать». Так же и для однокоренных слов:
- «Я раздаю (раздача) свои марки друзьям, потому что мне уже скучно их коллекционировать».
- «Какие-то странные шумы раздаются (раздаться) по вечерам возле моего нового дома».
- «Они раздают (раздай) образцы своей продукции всем желающим».
- «Зачем ты раздаешь (отдать) деньги – они обманывают тебя».
- «Сегодня всю ночь раздавались (подайте) раскаты грома, мешая мне уснуть».
Как верно подобрать вариант для «раздается»? Проверочное слово обязательно должно иметь корень «да». По смыслу же однокоренные формы далеко не всегда схожи.
Написание глагольного окончания
Последнее сомнение – специальное глагольное окончание «ть». Как проверить слово «раздаваться»? Тут снова нужно руководствоваться простыми школьными правилами:
- Если проверяемый глагол отвечает на вопросы: «что делает?», «что делают?» (без мягкого знака), то употребляется без «ь».

- Если проверяемый глагол отвечает на вопросы «что сделать?», «что делать?» (с мягким знаком), требуется писать «ь».
Примеры, иллюстрирующие правило:
- Что делать? «Раздаваться в городе посреди дня».
- Что делает? «Ежедневно раздается у соседей».
Проверочные слова в случае с этим глаголом нужны только для корня «да» — «раздай», «подать», «сдача». С написанием приставки и глагольного окончания определяются по правилам.
морфемный разбор слов: предрассветная,раздается,весеннюю,поднимается. — Школьные Знания.com
образуйте краткую форму от приведённых прилагательныхБледный — бледен, бледна; вкусный, молодой, дорогой, весёлый, скромный, хо-аный, страшный, верный … , далёкий, низкий, честныйБедный — бедны, бедны; чистый, яркий, грязный, резкий, милый, новый, близ-кій, пустой, вредный, жалкий, широкий, глупый.помогите пожалуйста
Укажите слово без окончания 1) красный 2) гора 3) быстро 4) сантиметр
озаглавь текст,,,,,,,,,,. ……..……
……..……
составить предложения из междометияой-ой, из-за, еле-еле, ах-ах, из-под, о-о-о, крепко-накрепко
ДАЮ 60Б
Прочитайте и выполните задания
1.Я пошел в лес, видневшийся вдали. 2. Перепрыгнув через небольшой ручеек, я оказался
в тени деревьев. 3. Изре
… дка пролетали птицы, задевая иногда меня крылом. 4. Я присел
на пенек, нагретый солнцем, и стал наблюдать за жизнью леса. 5. Вот рядом я заметил
небольшую кучу муравьев, еще строивших свой дом. 6. На ветке какого-то кустарника
копошилась божья коровка, ловко перелезавшая с листка на листок.
7. Так можно сидеть часами, наблюдая за лесными обитателями.
8. Отдохнув от городской суеты, я направился в наш микрорайон, расположенный рядом
с лесом.
1. Из предложений 1-4 выпишите причастия, определите их разряд и время.
2. Из предложений 6-8 выпишите деепричастия, определив вид и время.
3. Объясните постановку знаков препинания в 1 и 8 предложении: выпишите и
сделайте соответствующий разбор.
4. Из всего текста выпишите все наречия, выделите суффикс.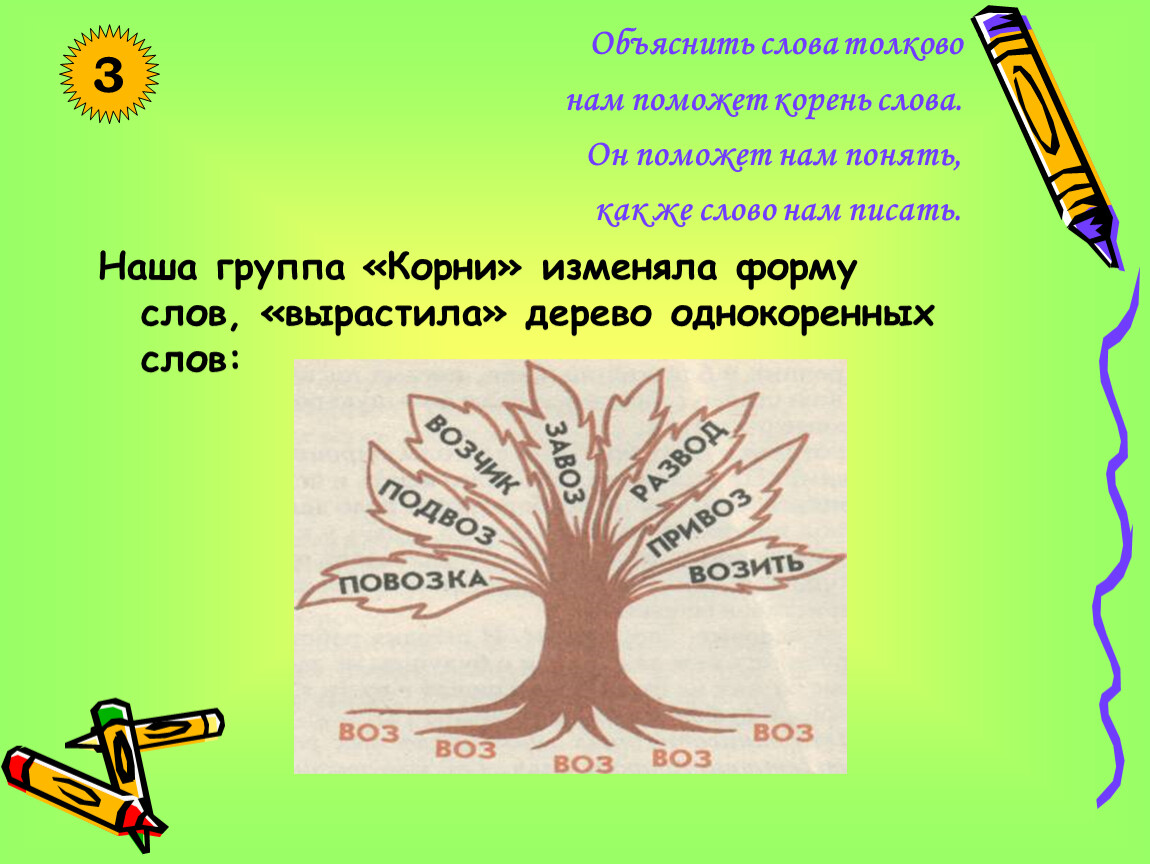
Составить правило НЕ с различными частями речи в виде таблицы. (НЕ с глаголами, НЕ с прилагательными, НЕ с существительными, НЕ с причастиями, НЕ с де … епричастиями, НЕ с местоимениями, НЕ с наречиями). Написать, когда пишется слитно, а когда раздельно
7. Составь и запиши из данных слов предложе-ние, используя предлог. Подчеркни орфограм-мы. Разбери предложение по членам. Выпишисловосочетания с вопро … сами.с ёмочная, под ехала, киностудия, группа.
Помогите пожалуйста распределить слова правильно. Осталась последняя попытка. Только правильно надо. Пожалуйста. Я сделала,но вышло НЕВЕРНО
прочитайте текст и ответьте на вопросы к. симонов жди меня выпишите из текста 2 — 3 повтора и 3 выражения употребить в переносном смысле, объяснив зна … чение выраженийПОЖАЛУЙСТА ПОМОГИТЕ СРОЧНО ДО ЗАВТРА даю 15 баллов
помогите пожалуйста, дам 35 балловУкажите словосочетания, в которых нет ошибок в употреблении числительных.семисот девяноста четырёх метровтремястами … шестидесятью рублямисемистами пятьюдесятью километрамидвести сорок пяти минутболее двухсот пятидесяти двух экземпляровсемьюстами шестьюдесятью днями
Андрей Школьников.
 Корни русского народа и психоисторическая диверсия (Геополитика)
Корни русского народа и психоисторическая диверсия (Геополитика)Периодически раздаются голоса, что России нужно вернуться к традициям предков, принять неоведичество и/или неоязычество. Процент таких людей в нашем обществе невелик, но они есть, и вопрос требует осмысленного ответа. Ведь фактически, речь идет о призыве к самоубийству России, уничтожению традиций и смыслов, ставших основой как Русского мира, так и Красного проекта
Данная статья посвящена рассмотрению обоснованности и серьезности предлагаемого, ведь речь идет ни много, ни мало, а о фактически перечеркивании тысячелетней истории России. Масштаб последствий таких деяний каждый может узреть на примере Европы, где в документах ЕС отсутствует даже упоминание христианства, но есть ссылки на античность, да и Третий Рейх был «чудной» попыткой вернуться к языческой традиции…
В 20-ом веке Россия пережила две психоисторические катастрофы, когда произошел коренной слом взглядов, норм, оценок и установок населения. Наше общество до сих пор не преодолело последствий краха Красного проекта, построенного на марксизме поверх православной русской традиции. Прошло уже более 25 лет, но базис, фундамент норм и взглядов все еще продолжает формироваться, отсеивая самый дикие и маргинальные идеи и течения. Наша система смыслов подобна стабилизированному больному, постепенно идущему на поправку за счет внутренних ресурсов и терапии. Худшее, что мы можем сейчас сделать – вместо покоя после ранения и успешно проведенной операции, отправить пациента на пересадку органов, подспудно уничтожив иммунитет. Собственно, именно это и предлагают сделать всевозможные неоязычники, неоведисты/неоиндуисты и т.д. Даже монархисты с сословным обществом принесут меньший вред России, а это, извините, показатель.
Наше общество до сих пор не преодолело последствий краха Красного проекта, построенного на марксизме поверх православной русской традиции. Прошло уже более 25 лет, но базис, фундамент норм и взглядов все еще продолжает формироваться, отсеивая самый дикие и маргинальные идеи и течения. Наша система смыслов подобна стабилизированному больному, постепенно идущему на поправку за счет внутренних ресурсов и терапии. Худшее, что мы можем сейчас сделать – вместо покоя после ранения и успешно проведенной операции, отправить пациента на пересадку органов, подспудно уничтожив иммунитет. Собственно, именно это и предлагают сделать всевозможные неоязычники, неоведисты/неоиндуисты и т.д. Даже монархисты с сословным обществом принесут меньший вред России, а это, извините, показатель.
В каждом обществе есть 5-10%% населения, что не воспринимают свою традицию и родную культуру, вне зависимости от ее содержания. Нужно воспринимать это как дань изменчивости человеческой природы и не принимать их идеи близко к сердцу.
Именно о таких идеях, а именно предложениях принятия Россией ведической религии и политеизма, скрывающихся за картинкой «а-ля русс» и пойдет речь.
Предки и дальние родственники
Прежде чем говорить о содержании предлагаемых для «возрождения» традиций и смыслов, необходимо сделать небольшой, краткий экскурс в историю, и рассказать о путях племен, что были предками славян и ариев. Ниже прилагается найденная на просторах мировой сети карта миграции гаплогруппы R1a (Y хромосома, передающаяся от отца к сыну без изменений, за исключением ошибок в копировании – в среднем одна на 58 млн. мономеров), основной группы восточных и центральных славян, потомков ариев и др. Автор согласен не со всеми датировками, но эти расхождения в пределах «о малого», что позволяет использовать «как есть», без перерисовывания.
Около 5 тысяч лет назад на Среднерусской равнине и Карпатах жили предки славян и ариев. 4,5 тысячи лет назад предки ариев начали свой долгий путь на восток и юг, распространяясь по Великой Степи от Причерноморья, до Средней Азии и северного Китая. Судя по распространению R1a, произошло покорение большого количества живших там народов, из которых победители брали себе жен и не уничтожали мужчин под корень. За тысячелетия произошло растворение небольшого количества пришельцев и захватчиков с изменением внешности, но Y хромосома не измена и позволяет говорить о происхождении. Патриархальное устройство общества сужает поле для гипотез.
4,5 тысячи лет назад предки ариев начали свой долгий путь на восток и юг, распространяясь по Великой Степи от Причерноморья, до Средней Азии и северного Китая. Судя по распространению R1a, произошло покорение большого количества живших там народов, из которых победители брали себе жен и не уничтожали мужчин под корень. За тысячелетия произошло растворение небольшого количества пришельцев и захватчиков с изменением внешности, но Y хромосома не измена и позволяет говорить о происхождении. Патриархальное устройство общества сужает поле для гипотез.
Миграция гаплогруппы R1a
Климат в те времена был отличен от нашего, так условия жизни в Шумере (5 тыс. лет назад) были благоприятны для земледелия и скотоводства. Для сравнения, посмотрите, как выглядит современный Ирак.
4 тысячи лет назад ариями были основаны города на Южном Урале (синташтинская культура, «Страна городов»), самый известный из них – Аркаим (современное название). Кстати, именно к этому времени относятся первые цивилизации Китая. Тут не нужно путать отдельные поселения и даже изолированные города с цивилизацией – городу Иерехон более 8,5 тыс. лет, но никто не рассматривает его как очаг отдельной цивилизации. Постоянный торговый центр на пересечение путей кочевников в Палестине (ярмарка с укреплениями) и не более.
Тут не нужно путать отдельные поселения и даже изолированные города с цивилизацией – городу Иерехон более 8,5 тыс. лет, но никто не рассматривает его как очаг отдельной цивилизации. Постоянный торговый центр на пересечение путей кочевников в Палестине (ярмарка с укреплениями) и не более.
3,3 — 3,7 тыс. лет назад арии на территории современного Курдистана имели собственную страну — Митанни. Примерно в этот же период другая часть ариев пришла в северную и центральную Индию, покоряя местные народы. На картинке ниже показаны имеющиеся у нас объективные данные, с указанием миграции, климата и религий.
Эволюция религиозных взглядов праславян и ариев
Вот тут и начинается интересное. Находятся люди, которые утверждают, что лежащие в основе неоиндуизма и записанные несколько сот лет назад Веды (сборники гимнов, молитвенных заклинаний и обрядов), что до этого несколько тысяч лет передавались в устной традиции, являются верой праславян.
Вот интересно, а почему неоиндуизм, а не зороастризм? С чего есть мысли, что сами гимны пришли из общего прошлого ариев и славян, а не придуманы по дороге на юг или изменены так, что ничего общего с оригиналом практически не осталось, кроме набора имен и названий, вот много общего у зороастризма и индуизма в сути вероучений? Какие вообще религиозные традиции и смыслы были у праславян в лесах в течение тысячелетий, как ушли арии?
Возникает вопрос по источникам, в частности, какое отношение имеют собранные и записанные уже в Средние века устные сказания к первоначальным? При передаче из уст в уста всегда происходят искажения, меняются языки, социально-экономические связи, термины и т.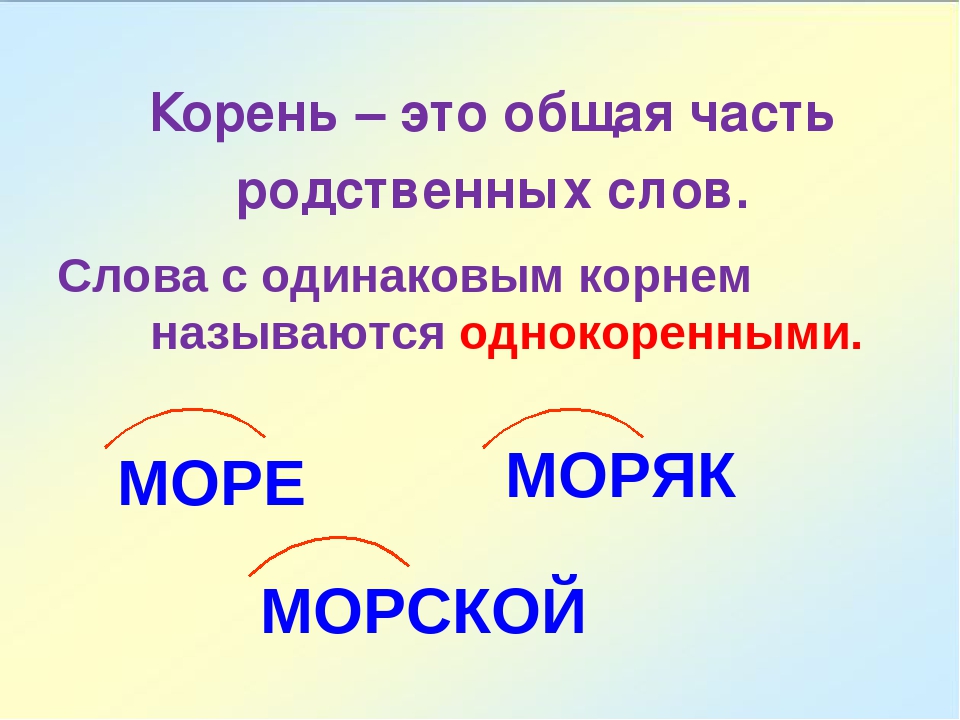 д. Также не стоит забывать, что ведическая религия ариев переродилась в кастовое общество и уже в брахманский период претерпела столько изменений, что сейчас даже не скажешь, что было додумано, а что было изначально.
д. Также не стоит забывать, что ведическая религия ариев переродилась в кастовое общество и уже в брахманский период претерпела столько изменений, что сейчас даже не скажешь, что было додумано, а что было изначально.
Ведь так легко добавить пару абзацев в древний текст, из которых следует, что именно твое понимание истинно верное и правильное… И так каждое поколение. Посмотрите, сколько апокрифов появилось в раннем христианстве за полтора-два века, а тут тысячелетия свободного творчества.
Таким образом, не имея объективных, материальных источников, прошедших все критические научные экспертизы в части датировки и подлинности, говорить о ведической религии/неоиндуизме, как имеющем какое-либо отношение к верованиям праславян нельзя. С тем же успехом, мы можем рассуждать о зороастризме, как исконной религии.
Речь не идет об отказе от истории, но мы должны искать, что было с нашими предками в промежутке от 3 тыс. лет до н.э. (можно и древнее) до 0,5 тыс. лет н.э., а не заполнять пустоту незнания фантазиями, уподобляясь «родине древних укров» или выдавая историю дальних, ушедших на юг родственников за свою.
Возврат России к «традициям предков»
Периодически приходится сталкивается с предложениями сценировать будущее России на базе неоязыческих и ведических принципов, про историческую обоснованность этого написано выше, перейдем к содержательной части. Рассмотрим предлагаемые религиозные системы на предмет и их соответствия текущими психоисторическими смыслами России.
Для сравнения взяты следующие учения:
- ведическая религия – рассмотрены современные толкования, переводы и интерпретации вед, или что за них выдается;
- язычество/политеизм на базе родноверия.
Даже поверхностное изучение постулатов, текстов, ритуалов и т.д. сформировали у автора стойкое понимание искусственности и эклектичности как ведических, так и языческих конструкций, отсутствие связи с реальной историей и профанацией духовной жизни, но другого ничего нет. Да и «возрождать» психоисторические смыслы России предлагают именно на этих или еще более химерических системах.
Сравнение психоисторических смыслов для России
На рисунке показаны параметры обоих учений. Не знаю, как читателей, но автора реально напрягает практически полное несовпадение ведической религии с текущими психоисторическими смыслами. Тут и универсальный космический порядок, которому подчинены и люди, и боги, отсутствие понятий добра и зла, кармические идеи, перерождения и т.д. Правда характерного для индуизма и буддизма негативного отношения к перерождению нет, ну хоть так.
Сколько поколений нужно, чтобы перейти на новые принципы, и главное зачем, что это даст? Отбросить весь пройденный нами за 5 тысяч лет путь, свернув на чужую колею теоретически можно, вот только ради чего?
С язычеством также весело – право сильного (хорошо принимаемое в молодости, пока не пришла старости), недеяние (жизнь в гармонии с природой), поклонение богам и силам природы, где верховное божество подобие античного представление о безличном Космосе, представление о Роке. Собственно, именно в этой конфигурации язычество и потерпело поражения в борьбе с христианством и исламом.
Складывается впечатление, что большую часть взяли из дошедших до нас античных мифов, добавили скандинавской мифологии и харизматичности, да элементов «а-ля русс». Вот и получается после смерти перерождение или переход в светлый Ирий, да другой космический уровень.
В чем опасность языческих идей? Они не самостоятельны и не продуктивны, построены исключительно на отрицании, ненависти. Изначально антихристианские и националистический (против других национальностей), данные смыслы выводят людей из Русского мира. Не даром, значимое количество, так называемых «русских мусульман» проходят по цепочке: язычество/ национализм – антихристианство – салафитский ислам – теракт/ война на стороне боевиков – смерть.
Предложение по принятию Россией «веры предков» можно проиллюстрировать на примере небольшого диалога. Представьте, что у вас есть большой коттедж, который нуждается в текущем ремонте – крышу подлатать, покрасить и т.д. Работа есть, но она не очень значительна и до зимы можно успеть, если не затягивать.
Тут приходит некий чудик и заявляет:
«Чудик: — Трехэтажный коттедж — это плохо, в прошлом веке тут пятистенок стоял, надо все снести и построить наш старый и надежный дом, в традиции, как встарь было.
Вы: — Так ведь зима скоро, по прогнозам самая лютая за последний век, не успеем, помереть можем от холода-то на улице.
Чудик: — Не важно, все вторично, ведь традиции будут учтены.
Вы: — Так там туалета не было нормального и электричества с водой…
Чудик: — Ой, да бросьте, зато какая русская печь, как у предков и речка недалеко!
Вы: — А есть кто-то, кто печи умеет класть?
Чудик: — Конечно, мастер Великомудр. Он раньше у нас охранником на воротах сидел, звался Васей, за пьянку и воровство выгнали.
Вы: — А он точно знает, как печи класть?
Чудик: — Конечно, он же сам рассказывал, что потомственный печник в двадцать третьем поколении.
Знания ему перешли еще от прадеда, вместе с тайными чертежами»
Ну и кем надо быть, чтобы поддаться на такой развод? Вот собственно такой бред нам и предлагают, говоря о ведах и/или язычестве.
Таким образом, предлагаемые ведической религией и политеизмом смыслы не актуальны и мало общего с реальным положением в России. Идет попытка под видом возрождения полностью изменить смыслы России, т.е. провести психоисторическую диверсию. Попытка внедрения, что неоиндуизма, что язычества приведет к полному слому и национальной катастрофе, масштаб которой превосходит в сумме и 1917 и 1991 годы. Рассказы про древних укров на этом фоне будут восприниматься как мелкие неточности.
Заключение
Психоисторические смыслы создают для человека и этнической системы такую картину мира, в которой у них есть смысл существования. Можно сколько угодно рассуждать о возможном будущем России в рамках ведической религии и язычества, но прошедшие тысячелетия/века, так изменили праславян, славян, русских, что сейчас это равносильно запуску жесткой психоисторической ломки, продолжительностью в несколько поколений и с непонятным результатом.
Фактически, нам предлагается самим сделать ровно то, что в отношении нас хотят сделать враги. При этом им реально без разницы, куда мы будем меняться, главное, чтобы больше внутренних проблем и конфликтов.
На практике, результатом реализации таких предложений станет не столько привнесение новых смыслов, сколько уничтожение существующих, перевод людей и всей этнической системы в промежуточное состояние, когда присутствует максимальная открытость для любых внушений.
Предлагаемые под видом возрождения традиций предков ведическая религия и язычество – суть элемент диверсии и психоисторической борьбы против России, попытка уничтожения всей истории, как православной, так и советской, поскольку Красный проект является продолжением христианских традиций.
P.S. (позднее) Автор с большим уважением относится к Клесову А.А., прочтение его работ и критики на его работы, убедило автора в справедливости и объективности методов именно Клесова А.А., а не его оппонентов.
Проверочное слово к слову «раздаваться»
Безударный гласный «а» в корне глагола «раздаваться» проверят родственные слова «дать», «раздать», «раздача». Приставка раз- пишется согласно орфографическому правилу написания приставок на з/с.
Правописание слова «раздаваться»
В слове «раздаваться» пишется приставка раз-, так как корень слова начинается со звонкого согласного [д]:
раздава́ться — приставка/корень/суффикс/окончание/постфикс.
В приставке раз- в безударном положении пишется буква «а», а под ударением — «о», сравним:
разлива́ть — ро́злив;
разда́ть — ро́зданный.
Проверяем букву «а» в слове «раздаваться»
Если произнести слово «раздава́ться» , то выясним, что в его корне неясно слышится безударный гласный. Чтобы написать букву «а» или «о» в корне рассматриваемого глагола, потребуется проверочное слово. В русском языке безударные гласные проверяют ударением, используя принцип, что в безударном положении пишется та же гласная буква, которая выступает под ударением в том же слоге любой грамматической формы слова или в подобранной родственной лексеме.
Понаблюдаем, как можно проверить безударные гласные в корне следующих слов:
- сиди́т — си́дя;
- ведёт — вёл;
- трава́ — тра́вка;
- гроза́ — гро́зы.
Аналогично подберем проверочное слово к слову «раздава́ться» среди родственных лексем:
- да́ть
- разда́ть
- разда́ча
Проверочное слово «дать», а также «раздать», «раздача» докажут написание буквы «а» в корне слова «раздаваться»
Примеры
Скачать статью: PDFВ любой момент может раздава́ться сигнал к атаке укреплений неприятеля.
Отовсюду стали раздава́ться одобрительные возгласы.
В весеннем лесу с восходом солнца начали раздава́ться щелканье, посвистывание, трели певчих птиц.
В потемневшем небе продолжали раздава́ться оглушительные раскаты грома.
РАЗГОВОР СВИНЬИ С ПРАВДОЮ
М. Е. Салтыков-Щедрин
Е. Салтыков-Щедрин
Свинья, разъевшееся животное; щетина ощерилась и блестит, вследствие беспрерывного обхождения с хлевной жидкостью Правда, особа, которой, по штату, полагается быть вечно юною, но уже изрядно побитая. Прикрыта, по распоряжению начальства, лохмотьями, сквозь которые просвечивает классический полный мундир, то есть нагота. Действие происходит в хлеву.
Свинья (кобенится). Правда ли, сказывают, на небе солнышко светит?
Правда. Правда, свинья.
Свинья. Так ли, полно? Никаких я солнцев, живучи в хлеву, словно не видывала?
Правда. Это оттого, свинья, что когда природа создавала тебя, то, создаваючи, приговаривала: не видать тебе, свинья, солнца красного!
Свинья. Ой ли? (Авторитетно.) А по-моему, так все эти солнцы — одно лжеучение. .. ась?
.. ась?
Правда безмолвствует и сконфуженно поправляет лохмотья.
В публике раздаются голоса: правда твоя, свинья! лжеучения! лжеучения!
Свинья (продолжает кобенится). Правда ли, будто в газетах печатают: свобода-де есть драгоценнейшее достояние человеческих обществ?
Правда. Правда, свинья.
Свинья. А -по-моему, так и без того у нас свободы по горло. Вот я безотлучно в хлеву живу — и го̀рюшка мало! Что мне! Хочу — рылом в корыто уткнусь, хочу — в навозе кувыркаюсь… какой еще свободы нужно! (Авторитетно.) Изменники вы, как я на вас погляжу… ась?
Правда вновь старается прикрыть наготу.
Публика гогочет: правда твоя, свинья! Изменники! изменники! Некоторые из публики требуют, чтоб Правду отвели в участок. Свинья самодовольно хрюкает, сознавая себя на высоте положения.
Свинья. Зачем отводить в участок? Ведь там для проформы подержат, да и опять выпустят. (Ложится в навоз и впадает в сентиментальность.) Ах, нынче и участковые одним языком с фельетонистами говорят! Намеднись я в одной газете вычитала: оттого-де у нас слабо, что законы только для проформы пишутся…
(Ложится в навоз и впадает в сентиментальность.) Ах, нынче и участковые одним языком с фельетонистами говорят! Намеднись я в одной газете вычитала: оттого-де у нас слабо, что законы только для проформы пишутся…
Правда. Так ты и читаешь, свинья?
Свинья. Почитываю. Только понимаю не так, как написано… Как хочу, так и понимаю!.. (К публике.) Так вот что̀, други! в участок мы ее не отправим, а своими средствами… Сыскивать ее станем… сегодня вопросец зададим, а завтра — два… (Задумывается.) Сразу не покончим, а постепенно чавкать будем… (Сопя, подходит к Правде, хватает ее за икру и начинает чавкать.) Вот так!
Правда пожимается от боли; публика грохочет. Раздаются возгласы: ай да свинья! вот так затейница!
Свинья. Что? сладко? Ну, будет с тебя! (Перестает чавкать.) Теперь сказывай: где корень зла?
Правда (растерянно). Корень зла, свинья? корень зла… корень зла… (Решительно и неожиданно для самой себя.) В тебе, свинья!
Корень зла, свинья? корень зла… корень зла… (Решительно и неожиданно для самой себя.) В тебе, свинья!
Свинья (рассердилась). А! так ты вот как поговариваешь! Ну, теперь только держись! Правда ли, сказывала ты: общечеловеческая-де правда против околоточно-участковой не в пример превосходнее?
Правда (стараясь изловчиться). Хотя при известных условиях жизни, невозможно отвергать…
Свинья. Нет, ты хвостом-то не верти! Мы эти момо̀-то слыхивали! Сказывай прямо: точно ли, по мнению твоему, есть какая-то особенная правда, которая против околоточной превосходнее?
Правда. Ах, свинья, как изменнически подло…
Свинья. Ладно; об этом мы после поговорим. (Наступает плотнее и плотнее.) Сказывай дальше. Правда ли, что ты говорила: законы-де одинаково всех должны обеспечивать, потому-де что, в противном случае, человеческое общество превратится в хаотический сброд враждующих элементов. .. Об каких это законах ты говорила? По какому поводу и кому в поучение, сударыня, разглагольствовала? ась?
.. Об каких это законах ты говорила? По какому поводу и кому в поучение, сударыня, разглагольствовала? ась?
Правда. Ах, свинья!
Свинья. Нѐчего мне «свиньей»-то в рыло тыкать. Знаю я и сама, что свинья. Я — Свинья, а ты — Правда… (Хрюканье свиньи звучит иронией.) А ну-тко, свинья, погложи-ка правду! (Начинает чавкать. К публике.) Любо, что ли, молодцы?
Правда корчится от боли.
Публика приходит в неистовство. Слышится со всех сторон: Любо! Нажимай, свинья, нажимай! Гложи ее! чавкай! Ишь ведь, распостылая, еще разговаривать вздумала!
Зри в корень!
Для грамотного письма абсолютно необходимо понимать, как устроено слово, одним из первых правил орфографии, с которым знакомят в школе и которое становится самым используемым на протяжении всей жизни, — безударные гласные В КОРНЕ. Так где же он — корень, как на этот корень «нанизываются» другие части слова, чтобы получились новые слова? Давайте поиграем. …
…
Гнёзда родственных слов в игре сформированы вокруг следующих слов:
ухо, голова, нос, рука, глаз, лёд, снег, вода, земля, дом, дорога, цветок.
Ход игры:
Игра 1. «Домино» ( Количество игроков: от двух и более.)
Играющим сдаются по 5-7 карточек, остальные – резерв («базар»). С помощью жребия или считалки определяется первый, выполняющий ход, который выкладывает любую карточку. Переход хода выполняется по часовой стрелке. Следующий игрок кладёт карточку с подходящим родственным словом. Если подходящей карточки нет, то играющий берёт одну из резерва («базара»). Если и взятую карточку некуда подставить, игрок пропускает ход.
Игра заканчивается, когда один из играющих выложит последнюю карточку – он победитель. Возможно окончание игры «рыбой» — так называется блокировка выкладки, когда на руках еще есть карточки, но доложить их некуда. В таком случае выигрыш принадлежит тому, у кого на руках меньше всех карточек.
Игра 2. «Рядочки» (Количество игроков: от трёх и более)
Раздающий выкладывает горизонтально на столе лицевой стороной карточки на одну меньше, чем игроков. Остальные раздаются между игроками поровну, лишние карточки, если остались, откладываются (они в игре не участвуют). С помощью жребия или считалки определяется первый, выполняющий ход. Игрок вертикально подставляет карточку, с подходящим родственным словом к любой из горизонтально расположенных карточек. Переход хода выполняется по часовой стрелке. В ряду карточки подставляют в любом порядке. Пока не закончен один ряд, нельзя переходить к следующему. Если нет возможности подставить карточку, играющий пропускает ход.
Выигрывает тот, кто первый выложит свои карточки, или тот, у кого меньше всего останется карточек, при варианте «рыбы» (смотри первый вариант игры).
Игра 3. «У кого больше» (Количество игроков: от трёх и более)
Карточки раздаются между игроками поровну. Игроки кладут карточки стопкой лицевой стороной вниз около себя. С помощью жребия или считалки определяется первый, выполняющий ход. Игрок берёт верхнюю карточку из своей стопки и кладет её в центр стола в открытом виде. Следующий по часовой стрелке делает то же самое до тех пор, пока на карточках, наложенных друг на друга, не окажутся родственные слова. В этом случае игрок, последний игрок, забирает себе всю стопку и кладет рядом с собой, не смешивая с другими карточками. Игра продолжается. Когда все карточки, полученные игроками в начале игры, будут выложены, игра считается законченной.
Игроки кладут карточки стопкой лицевой стороной вниз около себя. С помощью жребия или считалки определяется первый, выполняющий ход. Игрок берёт верхнюю карточку из своей стопки и кладет её в центр стола в открытом виде. Следующий по часовой стрелке делает то же самое до тех пор, пока на карточках, наложенных друг на друга, не окажутся родственные слова. В этом случае игрок, последний игрок, забирает себе всю стопку и кладет рядом с собой, не смешивая с другими карточками. Игра продолжается. Когда все карточки, полученные игроками в начале игры, будут выложены, игра считается законченной.
Победителем считается игрок, у кого окажется больше всего «захваченных» карточек.
Варианты дополнительных заданий:
- Ребёнок берёт любую карточку или несколько и составляет предложения со словами, изображенными на них. Предложения могут быть независимыми друг от друга, а могут быть объединены одним сюжетом.
- Ребёнок отбирает карточки с родственными словами и придумывает с этими словами предложения.
.jpg) Предложения могут быть независимыми друг от друга, а могут быть объединены одним сюжетом.
Предложения могут быть независимыми друг от друга, а могут быть объединены одним сюжетом. - Ребёнок берёт любую карточку и придумывает одно предложение с использованием обоих слов.
Полный список слов:
1. Ухо, ушастый, подушка, ушной, наушники, ушанка, лопоухий, длинноухий.
2. Голова, подголовник, головастик, трехголовый, головоломка, головной, заголовок, сорвиголова.
3. Нос, переносица, носовой, курносый, длинноносый, звездонос, носорог, носище.
4. Рука, рукомойник, наручный, безрукавка, рукавицы, вручать, рукопись, рукодельница.
5. Глаз, косоглазый, глазок, одноглазый, синеглазка, глазастый, поглазеть, глазунья.
6. Лёд, льдина, ледоход, ледокол, гололёд, ледяной, подлёдный, леденец.
7. Снег, подснежник, снежинка, снеговик, белоснежный, снегирь, Снегурочка, снежок.
8. Вода, водопой, водяной, водоросли, водолаз, подводный, глубоководный, водопроводчик.
9. Земля, подземный, земляной, землетрясение, земляника, наземный, приземлиться, землекоп.
10. Дом, домохозяйка, домовой, домосед, домовитый, домик, бездомный, домашний.
11. Дорога, велодорожка, бездорожье, железнодорожник, придорожный, подорожник, внедорожник,
дорожный.
12. Цветок, цветовод, цветочный, первоцвет, отцвести, цветочек, цветик-семицветик, соцветие.
Для игры с дошкольниками (упрощенный вариант) можно отобрать 15 карточек. Слова этих карточек являются родственными для 5 слов: ухо, голова, нос, рука, глаз. Эти карточки легко отобрать, правый верхний угл лицевой стороны у них отмечен красными шариками.
РОДСТВЕННЫЕ СЛОВА. КОРЕНЬ. Для детей с ОНР (3) уровня на школьном логопункте.

ТЕМА: РОДСТВЕННЫЕ СЛОВА. КОРЕНЬ.
Коррекционно-образовательные цели:
Уточнить представление о родственных словах; упражнять учащихся в подборе родственных слов; выделять в словах общую часть — корень; обогащать лексический запас; расширять связную речь.
Коррекционно-развивающие цели:
Формировать внимание, наблюдательность, память; учить передавать суть в развернутых высказываниях.
Коррекционно-воспитательные цели:
Воспитывать усидчивость, формировать положительные мотивации к логопедическому занятию.
ХОД ЗАНЯТИЯ
I. Организационный момент
Здравствуйте, ребята. Сегодня к нам пришли гости. Кивком головы и улыбкой поприветствуйте их. А теперь соберите слово из слогов, и вы узнаете, что нас ждет на занятии, (путе-шест-вие: поездка или передвижение пешком по разным странам, местностям). Как вы понимаете это слово? Как называются люди, которые путешествуют?
Вот и мы сегодня будем путешествовать, пойдем в гости к нашему знакомому Г ному. На нашем пути встретятся задания, с которыми мы должны будем справиться. Это позволит продолжить путь. Путь наш будет лежать через лес. За правильные ответы, я буду давать листочки из нашего леса. Этот лес необыкновенный, в нем есть волшебная грамматическая поляна. Послушайте, какие чудеса происходят на ней.
На нашем пути встретятся задания, с которыми мы должны будем справиться. Это позволит продолжить путь. Путь наш будет лежать через лес. За правильные ответы, я буду давать листочки из нашего леса. Этот лес необыкновенный, в нем есть волшебная грамматическая поляна. Послушайте, какие чудеса происходят на ней.
ГРАММАТИЧЕСКАЯ СКАЗКА.
Давно это было. На одной волшебной поляне росли корни. Они обладали удивительной способностью разрастаться. Когда корень разрастался, из него появлялись слова — веточки и называли их однокоренными. А так как ни были дети одного корня-папы то их еще называли родственными. Послушайте стихотворение о таких словах.
Жило-было слово ход
В слове выход, в слове вход,
В слове ходики стучало И в походе в даль шагало.
Какая общая часть у всех этих слов? Назовите её (ход).
Как называется общая часть, от которой образованы эти слова? — Корень.
А что означает слово ход?( движение в каком-либо направлении)
Слово выход? — место, через которое выходят.
Ходики? старинные часы, с гирями.
Поход? — путешествие или прогулка пешком, своим ходом.
Скажите, это слова близкие по смыслу?
В русском языке есть такое правило: все слова, которые имеют общую часть- корень- и близки по смыслу, называются родственными (повторить хором).
Родственные слова близки по смыслу.
В них повторяется общая часть- корень.
Родственные слова также называются -однокоренными.
Вывод: являются ли слова — выход, вход, ходики, поход- родственными? Почему?
II Основная часть.
ЗАДАЧА: нахождение общей части (корня)однокоренными.
На партах у вас лежат карточки, как вы считаете с каким корнем будет первая группа слов? Назовите слова-родственники.
А теперь укажите корень второй группы слов.
(карточки с домиками и грибочками)
Игра — Какие корни убежали?
Детям раздаются карточки с пропущенными корнями, в каждом столбике пропущен один и тот же корень
Определите, какой корень «убежал» из каждого столбика слов, поверните его на место, нарисовав на месте пропуска одну картинку-пиктограмму.
На доске -картинки: глаз, нож, сад, лес, хлеб, гриб, снег, зуб, дом.
Письмо Гнома.
Ребята мы получили от Гнома конверт. Он просит нашей помощи. У него в конверте предложения, которые перепутались. Давайте ему поможем собрать предложения. ( на доске слова — слова: дом, домосед, домовой, домочадец.
Предложения: Гном в лесу построил дом.
У него за печкой жил домовой.
Вечером гном встречался со своим домочадцем.
Они вместе занимались домашними делами.
Домик гнома всегда был открыт для гостей.
Как можно назвать эти слова? Почему?
Правильно, они близки по значению и у них один корень.
Домовой — существо, которое охраняет дом.
Домочадцы — все, кто живет в одном доме, в одной семье.
Ребята, а кто такой домосед? (человек, любящий дом)
III Физминутка.
Ветерок листвой шумит Шу-шу-шу! ( потираем ладони)
В трубах громко он гудит
У-У-У,У-У-У! (хлопают в ладони над головой)
Поднимает снег столбом
Бом-бом-бом (топают ногами)
Дует он везде кругом
Гом-гом-гом, гом-гом-гом. (переступают, повернувшись вокруг себя). Может вызвать бурю он
(переступают, повернувшись вокруг себя). Может вызвать бурю он
Страх-страх-страх, страх (руки в стороны, вращательные движения)
Не удержится и слон
Ах, ах, ах, ах (покачать головой)
IV Группировка родственных слов путём разделения по смыслу каждой из представленных групп (в конверте)
носить носильщик носилки
носки утконос поднос
носище носатый носорог
носишка нос носовой
Разделите слова на две группы, запишите на листочках. Поставьте ударение. Игра «Лестница»
Дети получают карточки с пустой «лестницей.» Необходимо заполнить все клеточки словами родственными слову «гриб».
Игра «Вырасти слова»
Каждому ребенку раздается конверт с рисунком дерева. Ученик должен придумать «корень» и вырастить свое «дерево», написав родственные слова. Рассказать о своем дереве.
V Заключительная часть.
Подведение итогов.
Над чем мы сегодня работали? Чему научились? Какие задания вам понравились?
Пароль на выход: я каждому Я каждому назову слово , а вы должны будете придумать родственное.
Распространение корня — обзор
Тщательный сбор анамнеза и тщательное физикальное обследование являются наиболее важными аспектами обследования пациентов с болезненным состоянием позвоночника. Этот процесс необходим для того, чтобы клиницист мог точно интерпретировать результаты визуализационных исследований в контексте и сформулировать план лечения.
Сбор анамнеза
Как и при любом клиническом опыте, врач интересуется историей болезненного состояния, включая его местонахождение, начало, характер, радиацию, обостряющие и смягчающие факторы, а также сопутствующие симптомы.Очень важно определить, какая область наиболее болезненна; это позволяет клиницисту сформулировать предварительный дифференциальный диагноз и определить, является ли боль осевой, корешковой или исходящей от других структур. Дискогенную боль, возникающую из-за болезненных состояний диска, обычно трудно локализовать. При опросе пациенты часто указывают на общую область боли. Эта боль обычно усиливается при сидении и уменьшается при смене положения. Боль может характеризоваться, но не ограничиваться ноющей или острой болью.В шейном отделе позвоночника верхние шейные диски передают боль в затылочную область, вызывая головные боли. 27 Боль в поясничном отделе позвоночника может относиться к области живота и паховой области. 21
Эта боль обычно усиливается при сидении и уменьшается при смене положения. Боль может характеризоваться, но не ограничиваться ноющей или острой болью.В шейном отделе позвоночника верхние шейные диски передают боль в затылочную область, вызывая головные боли. 27 Боль в поясничном отделе позвоночника может относиться к области живота и паховой области. 21
Боль в шейно-фасеточных суставах обычно, но не явно, усиливается из-за разгибания шеи или бокового сгибания. В поясничной области боль в фасеточных суставах обычно усиливается при стоянии и ходьбе и смягчается при сидении; однако боль в фасеточно-поясничных суставах может возникать и при сгибании вперед.Боль в фасеточных суставах обычно распространяется на фасеточные суставы в параспинальной области и более локализована, чем дискогенная боль. Боль может быть ипсилатеральной или двусторонней.
Доступны специальные схемы, на которых показаны схемы направления к специалистам при боли в шейно-фасеточных суставах (рис. 15-4). Схема направления при боли в шейно-фасеточных суставах изучена и установлена. 31 Чтобы составить карту направлений боли в шейных фасеточных суставах, исследователи стимулировали суставы у здоровых людей и отмечали болезненные места. 26 , 32 , 33 В различных исследованиях были получены очень похожие карты направлений боли в шейном отделе позвоночника. Однако авторам этой главы неизвестны исследования, в которых постоянно воспроизводились карты направлений боли в пояснично-фасеточных суставах.
15-4). Схема направления при боли в шейно-фасеточных суставах изучена и установлена. 31 Чтобы составить карту направлений боли в шейных фасеточных суставах, исследователи стимулировали суставы у здоровых людей и отмечали болезненные места. 26 , 32 , 33 В различных исследованиях были получены очень похожие карты направлений боли в шейном отделе позвоночника. Однако авторам этой главы неизвестны исследования, в которых постоянно воспроизводились карты направлений боли в пояснично-фасеточных суставах.
Пациенты с корешковыми симптомами обычно описывают боль преимущественно в конечностях. Боль может начаться как осевая боль (в случае грыжи диска), за которой следует появление боли в конечностях по мере ослабления осевой боли.Важно отметить, что ягодицы и околопаточные области считаются частью конечностей. Боль, преимущественно в этих местах, считается корешковой болью, а не осевой болью. Боль часто описывается как стреляющая, колющая, острая или жгучая, и пациенты часто могут точно локализовать болезненную область. Боль обычно возникает из-за определенного распределения нервных корешков с некоторой вариабельностью. 34 Боль в шейном корешке усиливается из-за разгибания и бокового сгибания шейки матки.Боль может быть уменьшена путем поднятия ипсилатеральной верхней конечности над головой, потому что этот маневр снижает напряжение на нижних нервных корешках (признак облегчения отведения плеча). 35 Доступны карты дерматома, на которых показано распределение корешков спинномозгового нерва (рис. 15-5). Важно отметить, что эти карты не являются точными на 100%, поскольку распределение нервных корешков варьируется от человека к человеку.
Клиническая заметка
Характер дерматомов может варьироваться от человека к человеку.
При грыже межпозвоночного диска и стенозе отверстий поясничная корешковая боль часто усиливается при сидении и уменьшается при ходьбе. В случае стеноза позвоночного канала боль обычно усиливается при ходьбе (разгибание позвоночника) и уменьшается при сидении (нейрогенная хромота). Дифференцировать нейрогенную хромоту от сосудистой хромоты, которая возникает вторично по отношению к сосудистой недостаточности, важно, потому что оба состояния влияют на одну и ту же (пожилую) популяцию. Пациенты с хромотой сосудов чувствуют облегчение от боли, когда они перестают ходить или когда они стоят, а не сидят.Пациенты с нейрогенной хромотой испытывают больше боли при спуске с горы (из-за разгибания позвоночника) и меньше боли при ходьбе с ходунками или тележкой для покупок (из-за сгибания позвоночника). Боль от нейрогенной хромоты следует за распределением нервных корешков, тогда как боль от сосудистой хромоты присутствует в глобальном масштабе в местах, снабжаемых определенными артериями.
Дифференцировать нейрогенную хромоту от сосудистой хромоты, которая возникает вторично по отношению к сосудистой недостаточности, важно, потому что оба состояния влияют на одну и ту же (пожилую) популяцию. Пациенты с хромотой сосудов чувствуют облегчение от боли, когда они перестают ходить или когда они стоят, а не сидят.Пациенты с нейрогенной хромотой испытывают больше боли при спуске с горы (из-за разгибания позвоночника) и меньше боли при ходьбе с ходунками или тележкой для покупок (из-за сгибания позвоночника). Боль от нейрогенной хромоты следует за распределением нервных корешков, тогда как боль от сосудистой хромоты присутствует в глобальном масштабе в местах, снабжаемых определенными артериями.
Боль, направленную из сустава SI, следует подозревать у пациентов, которые описывают боль в пояснице, преимущественно в области крестцовой борозды, с облучением нижних конечностей и паховой области или без него.Боль возникает вторично по отношению к системным состояниям (например, спондилоартропатиям, повторяющимся событиям сдвига) и после травмы или может возникать спонтанно. 36
36
Физикальное обследование
Целенаправленное медицинское обследование, основанное на информации, полученной из истории болезни пациента, устанавливает рабочий дифференциальный диагноз. Некоторые элементы общего физикального обследования важны при оценке пациентов с болезненными состояниями позвоночника. Эти элементы подробно описаны в Таблице 15-1.
Маневры физического обследования позвоночника
Специальные маневры используются для оценки различных состояний и анатомических областей, которые могут вызывать боль у пациента. Эти маневры описаны в Таблице 15-2. Дискогенные провокационные маневры вызывают воспроизводство боли у пациентов с дискогенной болью в пояснице. Признаки натяжения корней в верхних и нижних конечностях воспроизводят корешковую боль. Провокационные маневры SI используются для воспроизведения боли у пациентов с синдромом SI-сустава. Эти тесты не являются ни специфичными, ни чувствительными.(Более подробную информацию об этих и других тестах можно найти в томе 1 этой серии, Ортопедическая физическая оценка. )
)
Лучшим диагностическим методом для оценки боли, исходящей от сустава КП, является внутрисуставная диагностическая инъекция под рентгеноскопическим контролем. с местной анестезией. 37
Сенсорное обследование
Изучены укол, легкое прикосновение, положение и ощущение вибрации, уделяя особое внимание конечности с корешковой болью.Ощущения вибрации и положения уменьшаются при тяжелом стенозе позвоночника, особенно в шейном отделе позвоночника с поражением центрального канала. При радикулопатии уменьшение ощущения укола иглы и легкого прикосновения отмечается в определенном дерматомном распределении.
Изменения чувствительности позвоночника с патологией
- •
Стеноз позвоночника: нарушаются ощущения вибрации и положения
- •
Радикулопатия: ощущения укола иглой и легкого прикосновения затрагиваются в определенных дерматомах
Глубокие сухожилия Рефлексы
Исследование глубоких сухожильных рефлексов полезно при диагностике радикулопатий, особенно при наличии асимметричных рефлексов (Таблица 15-3). Рефлексы глубоких сухожилий могут быть снижены вследствие множества неврологических состояний или в результате возраста. Напротив, глубокие сухожильные рефлексы могут оставаться неизменными при радикулопатиях или даже быть гиперактивными при радикулопатиях, сопровождающихся поражением верхних мотонейронов.
Рефлексы глубоких сухожилий могут быть снижены вследствие множества неврологических состояний или в результате возраста. Напротив, глубокие сухожильные рефлексы могут оставаться неизменными при радикулопатиях или даже быть гиперактивными при радикулопатиях, сопровождающихся поражением верхних мотонейронов.
Ручное мышечное тестирование
Ручное мышечное тестирование проводится на четырех конечностях для определения наличия миотомической слабости. Чтобы обнаружить миотомную слабость, обычно тестируются две разные мышцы, снабжаемые одним и тем же нервным корешком, и два разных периферических нерва.Например, чтобы исследовать корешок L5 нерва на мышечную слабость, исследуют длинный разгибатель большого пальца стопы (L5, глубокий малоберцовый нерв), а также среднюю и малую ягодичные мышцы (L5, верхний ягодичный нерв). Это тестирование исключает различные неврологические состояния, вызывающие слабость, такие как миопатии, травмы периферических нервов и плексопатии.
Диапазон движений позвоночника
Диапазон движений обычно ограничен у пациентов с болями в спине из-за дегенеративных заболеваний позвоночника и защиты мышц вокруг болезненной структуры.
Распределение корней — обзор
Специфические паттерны направления MTrP боли
Давно известно, что боль, передаваемая от MTrP, не соответствует сегментарному паттерну распределения или паттерну распределения нервных корешков (Travell & Bigelow 1946). Несомненно, из-за этого большинство клиницистов, когда сталкиваются с болью, вызванной MTrP, не могут распознать ее как имеющую органическое происхождение (Bonica 1957). Тем не менее, как обсуждалось в главе 5, по крайней мере 50 лет назад Гуд в Великобритании, Келли в Австралии и Трэвелл и ее коллеги в Америке показали в многочисленных богато иллюстрированных отчетах, что каждая отдельная мышца в теле имеет свои собственные специфические особенности. образец обращения к специалисту по боли TrP.
По общему признанию, клиническая картина становится несколько запутанной, когда, как это часто бывает, возникает сложная картина боли из-за поражения более чем одной мышцы. Однако знание всех различных индивидуальных паттернов, каждый из которых необходимо изучать отдельно, снижает шансы упустить из виду миофасциальное происхождение такой боли и дает столь необходимые рекомендации относительно того, какие именно мышцы следует исследовать, чтобы найти TrP. несет ответственность за это.
несет ответственность за это.
В связи с этим следует отметить, что, хотя MTrP очень нежны на ощупь, что несколько удивительно, сами пациенты почти всегда совершенно не замечают своего присутствия и, следовательно, не могут оказать никакой помощи при их поиске. сделал.Также следует помнить, что боль от любого TrP часто ощущается на значительном расстоянии от него. Например, активность TrP в мышце, поднимающей лопатку, вызывает боль, которая ощущается не только у основания шеи и вниз по внутренней границе лопатки, но также может распространяться вверх по направлению к затылку и вниз по внутренней стороне шеи. руку, а иногда и грудную стенку. Это связано с тем, что знание различных конкретных шаблонов обращения к специалистам по лечению боли MTrP настолько важно, что в ходе этой книги каждый из них будет подробно описан.
Моделирование распределения корней, ориентации и плотности длины корней у эвкалипта и оценка связанной с этим эффективности водопользования
Адегбиди Х. Г., Комерфорд Н.Б., Джокела Э.Дж., Баррос Н.Ф. (2004) Развитие корней молодой сосны долевой в сподозолях в Юго-Восточной Джорджии. Soil Sci Soc Am J 68: 596–604
Г., Комерфорд Н.Б., Джокела Э.Дж., Баррос Н.Ф. (2004) Развитие корней молодой сосны долевой в сподозолях в Юго-Восточной Джорджии. Soil Sci Soc Am J 68: 596–604
CAS Статья Google Scholar
Батталья М., Сэндс П., Уайт Д., Маммери Д. (2004) КАБАЛА: связанная углеродная, водная и азотная модель роста леса для поддержки принятия решений по лесоводству.Для Ecol Manag 193: 251–282
Статья Google Scholar
Бернсон Г., Фарнсворт Э., Баззаз Ф. (1995) Распределение внутри и между органами, а также динамика изменений длины корня у двух видов березы. Oecologia 101: 439–447
CAS Статья Google Scholar
Бём В. (1976) Оценка длины корня на естественных профилях почвы на месте. J Agric Camb 87: 365–368
Статья Google Scholar
Bouillet JP, Laclau JP, Arnaud M, Thongo M’Bou A, Saint-André L, Jourdan C (2002) Изменения с возрастом в пространственном распределении корней Eucalyptus clone в Конго. Влияние на поглощение воды и питательных веществ. Для Ecol Manag 171: 43–57
Влияние на поглощение воды и питательных веществ. Для Ecol Manag 171: 43–57
Статья Google Scholar
Box JE (1996) Современные методы исследования корней. В: Waisel Y et al (eds) Корни растений, скрытая половина, 2-е изд. M. Dekker Publishers, Нью-Йорк, стр. 193–237
Google Scholar
Bragg PL, Govi G, Cannel RQ (1983) Сравнение методов, включая угловые и вертикальные миниризотроны, для изучения роста и распределения корней ярового овса.Растительная почва 73: 435–440
Артикул Google Scholar
Брауэрс NC, Мерсер Дж., Лайонс Т., Поот П., Венеклаас Э., Харди Дж. (2012) Климатические и ландшафтные факторы, влияющие на сокращение количества деревьев в средиземноморском экорегионе. Ecol Evol 3: 67–79
Статья Google Scholar
Брауэрс NC, Matusick G, RuthrofK Lyons T, Hardy G (2013) Оценка в ландшафтном масштабе отмирания кроны деревьев после экстремальной засухи и жары в экосистеме средиземноморских эвкалиптовых лесов. Landsc Ecol 28: 69–80
Landsc Ecol 28: 69–80
Статья Google Scholar
Chopart J, Rodrigues SR (2008) Оценка RLD сахарного тростника посредством картирования корней и моделирования ориентации. Почва для растений 313: 101–112. https://doi.org/10.1007/s11104-008-9683-4
CAS Статья Google Scholar
Chopart JL, Siband P (1999) Разработка и проверка модели для описания RLD кукурузы по количеству корней на профилях почвы.Растительная почва 214: 61–74
CAS Статья Google Scholar
Chopart JL, Payet N, Saint Macary H, Vauclin M (2007) Влияет ли на рост корня кукурузы внесение свиного навоза на тропической кислой почве? Корень растения 1: 75–84
Артикул Google Scholar
Дэвис А.С., Джейкобс Д.Ф. (2005) Количественная оценка качества корневой системы саженцев из питомников и взаимосвязи с производительностью высева. Нью-Форест 30: 295–311. https://doi.org/10.1007/s11056-005-7480-y
Нью-Форест 30: 295–311. https://doi.org/10.1007/s11056-005-7480-y
Статья Google Scholar
de Dorlodot S, Forster B, Pagès L, Price A, Tuberosa R, Draye X (2007) Архитектура корневой системы: возможности и ограничения для генетического улучшения сельскохозяйственных культур. Trends Plant Sci 12 (10): 474–481. https://doi.org/10.1016/j.tplants.2007.08.012
CAS Статья PubMed Google Scholar
Дрю М.К., Сакер Л.Р. (1980) Оценка экспресс-метода с использованием кернов почвы для оценки количества и распределения корней сельскохозяйственных культур на поле.Растительная почва 55: 297–305
Артикул Google Scholar
Эль-Лакани М.Х., Мохамед С.Ю. (1993) Характеристики корней четырех видов деревьев под воздействием ирригационных систем. Alex J Agric Res 38: 211–227
Google Scholar
Эскамилла Дж. А., Комерфорд Н. Б., Нири Д. Г. (1991) Сердцевина почвы — метод разлома для оценки распределения корней сосны. Soil Sci Soc Am J 55: 1722–1726
Статья Google Scholar
Fabiao A, Steen E, Madeira M (1987) Корневая масса на плантациях Eucalyptus globulus в Португалии в зависимости от характеристик почвы.Arid Soil Res Rehabil 1: 185–194
Статья Google Scholar
Falkiner RA, Nambiar EKS, Polglase PJ, Theiveyanathan S, Stewart LG (2006) Распространение корней Eucalyptus grandis и Corymbia maculata в деградированных засоленных почвах юго-востока Австралии. Агрофор Syst 67 (3): 279–291. https://doi.org/10.1007/s10457-005-5258-z
Статья Google Scholar
Глобальное почвенное партнерство ФАО (2019).http://www.fao.org/global-soil-partnership/intergovernmental-technical-panel-soils/gsoc17-implementation/internationalnetworkblacksoils/more-on-black-soils/what-is-a-black-soil/en/ . По состоянию на 19 ноября 2019 г.
Fitter AH (1994) Архитектура и распределение биомассы как компоненты пластической реакции корневых систем на неоднородность почвы. В: Caldwell MM, Pearcy RW (eds) Использование неоднородности окружающей среды растениями: экофизиологические процессы над и под землей.Academic, Сан-Диего, стр 305–323
Google Scholar
Грабарник П., Страницы Л., Бенго А. (1998) Геометрические свойства смоделированных корневых систем кукурузы: последствия для плотности длины и плотности пересечения. Растительная почва 200: 157–167
CAS Статья Google Scholar
Хамер Дж. Дж., Венеклаас Э. Дж., Пут П., Мокани К., Рентон М. (2015) Неглубокие градиенты окружающей среды подвергают риску внутренние виды: выводы и выводы из прогнозирования будущего распространения видов эвкалиптов в Юго-Западной Австралии.Austral Ecol. https://doi.org/10.1111/aec.12274
Статья Google Scholar
Hamer JJ, Veneklaas EJ, Renton M, Poot P (2016) Связь между текстурой почвы и корневой архитектурой видов эвкалипта может ограничивать ареалы распространения в условиях будущего климата. Почва растений 403 (1–2): 217–229. https://doi.org/10.1007/s11104-015-2559-5
CAS Статья Google Scholar
Ходж А. (2004) Пластичное растение: реакция корней на неоднородные поставки питательных веществ.New Phytol 162: 9–24
Артикул Google Scholar
Хупер Р.Дж., Сиваситампарам К. (2005) Характеристика повреждений и биотических факторов, связанных с сокращением численности Eucalyptus wandoo на юго-западе Западной Австралии. Can J For Res 35: 2589–2602
Статья Google Scholar
Индийский метеорологический департамент (2013 г.) Крайности Индии
Индийский метеорологический департамент (2015) Климатологический стол Дехрадун 2901–2000
Калкарни HD, Лал П. (1995) Показатели клонов Eucalypthadracha в Индии.В: Конференция CRC-IUFRO по эвкалиптовым плантациям по повышению урожайности и качества волокна, Хобарт, 19–24 февраля 1995 г.
Лал П., Догра А.С., Шарма СК, Чахал GBS (2006) Оценка различных клонов эвкалипта в Пенджабе. Indian For 132 (11): 1383–1390
Google Scholar
Lang ARG, Melhuish FM (1970) Длина и диаметр корней растений в неслучайных популяциях путем анализа плоской поверхности. Биометрия 26: 42–431
Статья Google Scholar
Lopez-Zamora I, Falcão N, Comerford NB, Barros NF (2002) Изотропия корней и оценка метода измерения распределения корней в траншеях почвы.Для Ecol Manag 166: 303–310
Артикул Google Scholar
Линч Дж. (1995) Корневая архитектура и продуктивность растений. Физиология растений 109 (1): 7–13. https://doi.org/10.1104/pp.109.1.7
CAS Статья PubMed PubMed Central Google Scholar
Marriott FHC (1972) Задача Буффона для неслучайного распределения. Биометрия 28: 621–624
Статья Google Scholar
Matusick G, Ruthrof K, Brouwers NC, Dell B, Hardy G (2013) Внезапное обрушение лесного полога, соответствующее сильной засухе и жаре в эвкалиптовом лесу средиземноморского типа на юго-западе Австралии.Eur J For Res 132: 1–14
Статья Google Scholar
Maurice J, Laclau JP, Re DS, Gonçalves JLM, Nouvellon Y, Bouillet JP, Chopart JL (2010) Изотропия тонких корней на плантациях Eucalyptus grandis. К прогнозированию плотности корней на основании количества корней на стенках траншеи. Почва для растений 334 (1): 261–275. https://doi.org/10.1007/s11104-010-0380-8
CAS Статья Google Scholar
Медлин Б.Е., Пеппер Д.А., О’Грэйди А.П., Кейт Х. (2007) Связывание использования воды листьями и деревьями с моделью отдельного дерева.Tree Physiol 27: 1687–1699
Статья Google Scholar
Melhuish CFM, Lang ARJ (1968) Количественные исследования тонких корней в почве. I. Длина и диаметр корней хлопчатника в глинисто-суглинистой почве при анализе поверхностно-отшлифованных блоков почвы, пропитанной смолой. Soil Sci 106: 16–22
Статья Google Scholar
Нордвейк В., Лоусон Дж., Хайриа К., Уилсон Дж. (2015) Распределение корней деревьев и сельскохозяйственных культур: конкуренция и / или взаимодополняемость, Глава 8.В: Ong CK, Black CR, Wilson J (eds) Взаимодействие между деревьями и культурами: агролесоводство в изменяющемся климате, 2-е изд. CAB International, Уоллингфорд, Великобритания, стр. 221–257
Google Scholar
Пьерет А., Латчак К., Чатанвонгса П., Сенгтахуанхунг О., Валентин С. (2007) Взаимодействие между ростом корней, уклоном и отслоением почвы в зависимости от землепользования: тематическое исследование в небольшом горном водосборе Северного Лаоса. Растительная почва 301: 51–64
CAS Статья Google Scholar
Poot P, Lambers H (2003) Реакция роста на заболачивание и осушение проростков древесных Hakea (Proteaceae), происходящих из контрастирующих местообитаний на юго-западе Австралии.Растительная почва 253: 57–70
CAS Статья Google Scholar
Poot P, Veneklaas E (2013) Распространение видов и уменьшение кроны связаны с контрастирующими водными отношениями у четырех распространенных симпатрических видов эвкалиптов на юго-западе Австралии. Почва растений 364: 409–423
CAS Статья Google Scholar
Schiffers K, Tielbörger K, Tietjen B, Jeltsch F (2011) Корневая пластичность сдерживает конкуренцию между растениями: теория соответствует экспериментальным данным.Экология 92: 610–620
Статья Google Scholar
Schmid I, Kazda M (2002) Распространение корней ели европейской в моновидовых и смешанных насаждениях на разных почвах. Для Ecol Manag 159 (1-2): 37–47. https://doi.org/10.1016/S0378-1127(01)00708-3
Статья Google Scholar
Schmid I, Kazda M (2005) Кластерное распределение корней в зрелых древостоях Fagus sylvatica и Picea abies .Oecologia 144: 25–31
Статья Google Scholar
Тардье Ф. (1988) Анализ пространственной изменчивости плотности корней кукурузы. I-Влияние уплотнения колес на пространственное расположение корней. Растительная почва 107: 259–266
Статья Google Scholar
Тейлор Х.М., Бом В. (1976) Использование акриловой пластмассы в качестве окон ризотрона. Agron J 68: 693–694
Артикул Google Scholar
Templeton GF (2011) Двухэтапный подход к преобразованию непрерывных переменных в нормальные: значение и рекомендации для исследования ИБ.In: Communications of the AIS , vol 28, Article 4
Van Noordwijk M (1987) Методы количественной оценки структуры распределения корней и динамики корней в полевых условиях. В: 20-й коллок. Int. Калийный институт, Берн, стр. 247–265
Вепраскас М.Дж., Хойт Г.Д. (1988) Сравнение профилей траншеи и основных методов для оценки распределения корней при исследованиях обработки почвы. Agron J 80: 166–172
Артикул Google Scholar
Xu Z, Zhou G (2008) Ответы плотности устьиц листьев на состояние воды и его связь с фотосинтезом в траве.J Exp Bot 59 (12): 3317–3325. https://doi.org/10.1093/jxb/ern185
CAS Статья PubMed PubMed Central Google Scholar
Xu H, Li Y, Xu G, Zou T (2007) Экофизиологическая реакция и морфологическая адаптация двух центральноазиатских пустынных кустарников к изменению количества летних осадков. Среда растительной клетки 30: 399–409
CAS Статья Google Scholar
Границы | Особенности корней, клубенькование и распределение корней в почве для пяти видов дикой чечевицы и Lens culinaris (Medik.) Выращено в условиях хорошо полива
Введение
Lens culinaris эволюционировала и была одомашнена в Юго-Западной Азии (Ladizinsky, 1993), откуда выращивание чечевицы впоследствии распространилось на подходящие экосистемы на всех континентах. Поскольку чечевица готовится быстро, ее мировое потребление растет быстрее, чем рост населения, а производство быстро растет, особенно в Австралии и на северных равнинах Северной Америки. Поскольку мировое население продолжает расти, а конкуренция за сельскохозяйственные ресурсы усиливается, возникает необходимость в производстве высокоурожайных генотипов чечевицы, которые эффективно используют ресурсы даже в условиях хорошо полива.Эффективное использование воды и питательных веществ станет более важным в будущем по мере усиления конфликта за эти ресурсы (Sauchyn and Kulshreshtha, 2008; Dai, 2011). Прогноз общего мирового использования удобрений (N, P, K) на 2017 год составляет 197 млн. Тонн тысяч тонн, из которых 30 млн. Тонн 1000 тонн азотных удобрений прогнозируются для использования в Северной Америке (Продовольственная и сельскохозяйственная организация Объединенных Наций [ФАО], 2015). Из-за прогнозируемых водных конфликтов, неустойчивого спроса на удобрения в долгосрочной перспективе и выброса парниковых газов в атмосферу необходимо улучшить как улавливание воды, так и биологическую фиксацию азота.Селекция высокоурожайных сортов чечевицы, минимизирующих водопотребление и наиболее эффективное использование азота, будет приобретать все большее значение. Будущие сорта чечевицы должны обладать как эффективной корневой системой для удержания влаги, так и максимальной фиксацией азота. Некоторые сорта чечевицы, выращиваемые в Северной Америке, обладают переменной клубеньковой способностью (Abi-Ghanem et al., 2011). Среди исследованных сортов Lens culinaris , L. culinaris Eston продемонстрировали лучшее клубенькообразование и самую высокую активность клубеньков по сравнению с другими.Возможность использовать этот вариант является частью стратегии по достижению максимального урожая чечевицы. Другая часть этой стратегии — отбор улучшений в корневых характеристиках и архитектуре. Системы корней влияют на поглощение питательных веществ и воды и действуют как поглотители фотоассимилятов и поступления углерода в почву (Gan et al., 2009). Хотя корневые системы различаются от растения к растению в зависимости от окружающей среды и из-за генетической изменчивости, наличие очень тонких корней (диаметр <0,5 мм) и / или тонких корней (диаметр 0.5–2,0 мм) является важным физиологическим компонентом, который определяет большую часть длины корня и площади поверхности корня, доступной для поглощения воды и питательных веществ (Zobel et al., 2007; Zobel and Waisel, 2010). Однако эффективность извлечения воды из почвы зависит от того, есть ли в почве запас воды или водоснабжение обеспечивается сезонными дождями (Tron et al., 2015). В идеале сорта чечевицы должны иметь как глубокую корневую систему, так и корни, которые могут распространяться по большим объемам почвы для захвата поверхностных вод.
Корневая система культурной чечевицы ( Lens culinaris Medik) сравнивалась с корневой системой масличных культур, других зернобобовых культур и пшеницы (Gan et al., 2009, 2011; Liu et al., 2010). Характер распределения мелких корней, таких как длина корня, площадь поверхности корня, объем корня и диаметр корня у культурной чечевицы, отличался от такового у других культур и, следовательно, мог играть важную роль в поглощении воды и питательных веществ. Gahoonia et al. (2005) сообщили, что два сорта чечевицы с разной длиной корня дали соответствующие различия в урожайности.Учитывая узкую генетическую базу чечевицы, ее дикие родственники могут быть возможным источником генетических ресурсов. Использование диких родственников чечевицы способствует повышению устойчивости к болезням (Ye et al., 2002; Muehlbauer et al., 2006; Tullu et al., 2006; Podder et al., 2013). Например, Tullu et al. (2006) продемонстрировали, что рекомбинантные инбредные линии, полученные в результате скрещивания восприимчивой линии Lens culinaris Eston и устойчивой линии зародышевой плазмы (PI 320937), обладали устойчивостью к фитофторозу.Podder et al. (2013) путем скрининга образцов культурной и дикой чечевицы продемонстрировали, что более 70% диких образцов обладают устойчивостью к стеблевому ожогу, а многие также обладают устойчивостью к другим болезням, таким как аскохитовый ожог и антракноз. Учитывая большую вариабельность биотических стрессов у видов дикой чечевицы, существует большой потенциал для улучшения как корневой системы, так и способности культурной чечевицы связывать азот. Мало что известно о корневой системе видов дикой чечевицы по сравнению с культурной чечевицей.О нескольких исследованиях, посвященных изучению надземных параметров, таких как урожайность и высота растений, сообщалось о зародышевой плазме дикой чечевицы (Pratap et al., 2014; Singh et al., 2014). Недавнее исследование зародышевой плазмы дикой чечевицы в поле и в помещении показало филогенетические отношения между генотипами в контексте засухоустойчивости, но характеристики корней не были охарактеризованы (Singh et al., 2016). Базовое исследование засухоустойчивости видов дикой чечевицы показало, что существует различие в способах предотвращения засухи между видами и внутри них (Gorim and Vandenberg, 2017).Не существует всестороннего исследования свойств корней дикой чечевицы и характера распределения корней в различных слоях почвы.
Это исследование было сфокусировано на характере распределения корневых признаков и клубеньков на разной глубине почвы у диких и культурных генотипов чечевицы. Мы предположили, что генотипы дикой чечевицы, принадлежащие к определенным генофондам, будут иметь сходные корневые признаки и распределение клубеньков в почве. Наша цель состояла в том, чтобы определить, существуют ли различия в признаках корней и клубеньков между генотипами дикой и культивируемой чечевицы и внутри них, а также между моделями их распределения в разных горизонтах почвы.
Материалы и методы
Грунтовые материалы
Эксперимент проводился в помещении с контролируемой средой в Колледже сельского хозяйства и биоресурсов Университета Саскачевана, Канада (широта 52,133; долгота -106,631). Подробное описание типа почвы, места сбора почвы и внесенных удобрений было аналогично описанию, приведенному Горимом и Ванденбергом (2017). Почву помещали в пробирки диаметром 10 см и длиной 60 см, которые делили на три части по 20 см, соответствующие глубинам горизонтов A, B и C, наблюдаемых в поле.Следовательно, глубина почвы в горизонте А или верхнем слое почвы варьировалась от нуля до 20 см, в горизонте В — от 20 до 40 см, а в горизонте С — от 40 до 60 см. Трубки были скреплены лентой вместе и герметизированы внизу мелкой сеткой, чтобы обеспечить дренаж и предотвратить потерю почвы. Верхняя часть каждой трубы была заполнена 1,8 кг почвы с горизонта А, чтобы оставалось достаточно места для полива. В среднюю и нижнюю части трубы засыпали по 2 кг грунта из горизонтов В и С соответственно.
Растительные материалы
Дикие виды / генотипы чечевицы (сокращение в скобках): L.orientalis ( L. ori. ), IG 72643, L. ori. PI 572376, Lens tomentosus ( L. tom. ) IG 72805, L. odemensis ( L. ode. ) IG 72623, L. lamottei ( L. lam. ) IG 110813 , L. ervoides ( L. erv. ) L-01-827A, L. erv. IG 72815. Lens culinaris ( L. cul .) Eston был единственным культивированным генотипом, включенным в этот эксперимент, поскольку он был успешно скрещен с некоторыми дикими видами.Согласно Wong et al. (2015), L. cul ., L. ori . И L. tom . составляют первичный генофонд; L. ode . и L. lam . находятся во вторичном генофонде, и L. erv . находится в третичном генофонде. Эти три генофонда генетически доступны для интрогрессии признаков посредством межвидовой гибридизации.
Семена генотипов дикой и культивируемой чечевицы скарифицировали, промывали в отбеливателе и затем предварительно проращивали в темной камере при 22 ° C.Через 3 дня отбирали проростки с длиной корешка более 2 см, затем пересаживали в промаркированные пробирки и добавляли к ним 6 г инокулята ризобий ( Rhizobium leguminosarum biovar Viceae штамм 1435; Nodulator XL SCG, Becker Underwood, Canada). поверхность почвы рядом с саженцем в каждой пробирке. Эксперимент представлял собой полностью рандомизированный блочный дизайн с 8 генотипами, двумя периодами сбора урожая (во время заполнения стручков и при полной зрелости) и четырьмя повторами. Количество воды при 100% полевой емкости (FC) для шести случайных незасаженных трубок было определено заранее.Почва в пробирках поддерживалась на уровне 80% FC на протяжении всего эксперимента. Температуру устанавливали на 21 ° C днем / 15 ° C ночью, установленную на 16 часов. Интенсивность света составляла от 308 до 392 мкмольм -2 ⋅с -1 в зависимости от положения трубки и высоты растения. Лампочки в комнате Люминесцентные лампы Т-5 (# 835 Philips, ON, Канада) и светодиодные планки, 730 мм Far Red, (Fluence Bioengineering, Остин, Техас, США). Положения пробирок в каждом блоке были повторно рандомизированы при каждом взвешивании на протяжении всего эксперимента, чтобы минимизировать влияние положения света.
Параметры оценены
Половина растений была собрана для оценки параметров роста через 11 недель после посева (WAS), в то время как другая половина была при 13 WAS, когда были получены зрелые семена. Дни до цветения считались датой, когда более 90% растений имели распустившиеся цветы. При сборе урожая высоту растения измеряли как расстояние от уровня почвы до кончика самого верхнего листа. Подсчитывали количество стручков и семян на растении по достижении зрелости. Биомассу над землей измеряли, помещая все растение в промаркированные бумажные пакеты, а затем сушили в печи при 70 ° C в течение 48 часов перед взвешиванием.Клейкая лента, скреплявшая стыки трех частей трубок, была удалена, затем использовался нож, чтобы разрезать и разделить каждую трубку на соответствующие горизонты A, B и C. Затем почву каждой части осторожно смывали на сите с размером ячеек 0,5 мм, чтобы минимизировать потерю корней. Промытые корни помещали в помеченные пакеты Ziploc, и вручную удаляли мусор и мертвые корни из почвы. Признаки корня измеряли с помощью коммерческого программного обеспечения WinRHIZO TM (Regent Instruments Inc., Канада, 2013) для определения общей длины корня (TRL), общей площади поверхности корня (TRSA), длины корня на единицу объема почвы [называемой плотностью длины корня (RLD)], общего объема корня (TRV), среднего диаметр корня (MRD), объем корня (RV), общее количество кончиков корня (TRT) и общее количество корневых развилок (TRF) в каждом горизонте. WinRHIZO TM также сгенерировал дополнительный вывод, который классифицирует корневые признаки по шести классам диаметров: 0 — 0,5 мм,> 0,5 — 1 мм,> 1 — 1,5 мм,> 1,5 — 2.0 мм,> 2 — 3,5 мм и> 3,5 — 4,5 мм. Долю корней в каждом классе рассчитывали как процент от общего количества для каждой из трех секций корневой трубы.
Перед анализом корней было подсчитано и сфотографировано количество клубеньков в каждом горизонте почвы для каждого вида / генотипа. После анализа WinRHIZO TM корни из каждого горизонта помещали в промаркированные бумажные пакеты, сушили в печи при 70 ° C в течение 48 часов, а затем взвешивали для определения сухой массы корней. Набор растений, которые продолжали расти, помещали в мешки с мелкими ячейками, чтобы предотвратить разрушение, и собирали, когда на каждом растении было высушено более 80% семян.Количество семян, вес семян и вес тысячи зерен определяли для каждого вида / генотипа.
Статистический анализ
Процедура SAS 9.4 PROC GLM (в системе статистического анализа, институт SAS, Кэри, Северная Каролина, США) использовалась для сравнения средних значений между видами / генотипами на заданной глубине почвы для каждого общего признака корня и сухой массы корня. Буквы были присвоены, чтобы показать значимые различия при α = 5%. Наименее значимые различия (LSD) были также получены для сравнения видов / генотипов, принадлежащих к данному классу диаметра на данной глубине почвы с помощью SAS.
Результаты
Характеристики роста линз
над землей ГенотипыНадземные параметры роста как культурной, так и дикой чечевицы были различными, не влияя на их классификацию генофонда (Таблица 1). Между генотипами и между генотипами наблюдалась разная высота растений и сроки цветения. Линза восточная PI 572376 и L. erv . IG 72815 имел самые низкие и самые высокие растения соответственно; остальные генотипы имели аналогичную высоту.Большинство генотипов цветут между 36 и 42 днями. Два L. erv. У генотипов (третичный генофонд) для цветения потребовалось меньше всего дней. Общее количество произведенной биомассы (корень плюс побеги плюс семена) также варьировалось среди генотипов. Наибольшее количество произведено L. cul . Эстон при этом наименьшим был произведен L. erv . L-01-827A. При первом урожае л. Том . IG 72805 имел наибольшее количество стручков, за ним следовало L. erv . L-01-827A, в то время как оба L.ода . IG 72623 и L. erv. У IG 72815 было наименьшее количество стручков (таблица 1). В конце эксперимента количество полученных семян и их масса варьировались между генотипами и их генофондами. Lens culinaris Eston произвел значительно меньше, но более крупных семян по сравнению со всеми генотипами дикой чечевицы (Таблица 1).
ТАБЛИЦА 1. Сравнение наземных фенотипических признаков для Lens culinaris Eston и пяти видов дикой чечевицы, выращенных в условиях полного полива (среднее значение ± стандартная ошибка).
Сравнение производства биомассы дикой и культурной чечевицы
Количество корневой биомассы, произведенной в трех почвенных горизонтах, было различным внутри генотипов и между генотипами (рис. 1). В горизонте А — L. ode. IG 72623 продуцировал значительно больше (α = 5%) биомассы корней по сравнению со всеми другими генотипами. Генотипы, принадлежащие к первичному генофонду, имели сходную биомассу корней, за исключением L. tom. IG 72805, который имел значительно более низкую биомассу. Линза lamottei IG 110813 и L.эрв. L-01-827A, принадлежащие к вторичному и третичному генофонду соответственно, оба имели значительно более низкую (α = 5%) биомассу корней по сравнению со всеми другими генотипами как в горизонтах A, так и в горизонтах B. Lens culinaris Eston, однако, давала значительно более высокую биомассу по сравнению со всеми другими генотипами в горизонте B. Для всех исследованных генотипов наименьшее количество корневой биомассы наблюдалось в горизонте С, и не было значительных различий между генотипами на этой глубине (рис. 1).Для всех видов при сравнении количества образовавшихся корней на побегах было обнаружено, что все генотипы чечевицы выделяли больше своих ресурсов на побеги. Наибольшая вегетативная биомасса (значимая при α = 5%) была произведена у L. ode. IG 72623, хотя это не привело к соответствующему увеличению репродуктивной биомассы. Lens culinaris Eston произвела наибольшее количество совокупной вегетативной и репродуктивной биомассы (рис. 2).
РИСУНОК 1. Сравнение распределения сухой массы корней в трех горизонтах почвы для пяти генотипов дикой чечевицы и Lens culinaris Eston. [Столбец — стандартные ошибки, а буквы — сравнивают сухой вес корней между генотипами для одного и того же горизонта почвы; отсутствие букв означает аналогичную биомассу].
РИСУНОК 2. Сравнение пропорций биомассы корней (R) и побегов (S) и вегетативной биомассы (VB) с репродуктивной биомассой (RB) у пяти видов дикой чечевицы с Lens culinaris Eston.
Сравнение признаков корней дикой и культурной чечевицы
TRL, RLD, TRSA, MRD, TRT и TRF были проанализированы во всех горизонтах почвы, чтобы понять, как различается рост корней между генотипами / видами. Далее мы оценили характер распределения мелких корней в каждом горизонте почвы, чтобы определить, различаются ли черты корней среди генотипов и генофондов. Мы сообщаем результаты определения корневых признаков с точки зрения классификации генофонда и почвенных горизонтов. В горизонте А — L. cul .Eston, L. ori . PI 572376, L. ode . IG 72623 и L. erv . IG 72815 имел значительно высокий (α = 5%) TRL, хотя они представляли три разных генофонда (рис. 3А). Линза восточная IG 72643 и L. tom . IG 72805, оба вида первичного генофонда, имели промежуточный TRL, но он был значительно выше ( P <0,001) по сравнению с таковым у L. lam . IG 110813 и L. erv. L-01-827A (вторичный и третичный генофонд соответственно). Линза culinaris Eston, L. ori . PI 572376 и оба L. erv. Генотипы имели значительно более высокий (α = 5%) процент корней в классе диаметров 0-0,5 мм. При диаметре корня> 1,5 мм процент TRL в L. cul . Eston в большинстве случаев был значительно ниже, чем у дикой чечевицы (рис. 4A). Lens ervoides L-01-827A имеет стабильно высокую долю своего TRL в классе диаметров, 0-0,5 мм.
РИСУНОК 3. Сравнение характеристик корней в трех горизонтах почвы для Lens culinaris Eston и пяти видов дикой чечевицы, выращенных в условиях полного полива. Полоски обозначают стандартную ошибку. Буквы обозначают достоверные различия при α = 5% для генотипов в одном и том же почвенном горизонте. (A) TRL, общая длина корня; (B) TRSA, общая площадь поверхности корня; (C) RLD, плотность длины корня; (D) MRD, средний диаметр корня; (E) TRT, общее количество верхушек корня; (F) TRF, общее количество корневых вилок.
РИСУНОК 4. Процент распределения общей длины корня по шести классам диаметра корня, оцененный в трех горизонтах почвы для семи генотипов дикой чечевицы по сравнению с культурной чечевицей ( Lens culinaris Eston. Линейные столбцы — значения LSD при P < 0,05 для сравнения генотипов в пределах класса диаметров. (А) горизонт А; (В) горизонт В; (В) горизонт С.
TRSA был значительно выше для L.ori . PI 572376, L. ode . IG 72623 и L. erv . IG 72815, хотя они принадлежат к первичному, вторичному и третичному генофондам соответственно. Линза lamottei IG 110813 и L. erv . L-01-827A имел самый низкий TRSA (значимый при α = 5%). Когда пропорции TRSA были классифицированы по классам разного диаметра, наблюдались значительные различия в том, как отдельные генотипы распределяли свои TRSA (Рисунок 5). Lens orientalis PI 572375 содержал значительно более высокую долю TRSA в классе диаметра корня 0-0.5 мм по сравнению со всеми другими генотипами (рис. 5А). Большинство видов имели самую высокую долю TRSA в классе диаметров> 0,5–1,0 мм, но в пределах этого класса TRSA L. cul . Eston был значительно выше (α = 5%), чем у всех видов дикой чечевицы. Lens odemensis IG 72623 имел самую высокую (значительную, α = 5%) долю TRSA в классе диаметров> 2,0–4,5 мм (рис. 5A) по сравнению с другими генотипами. Л. том . IG 72805, имел более высокую долю его TRSA в классе диаметра корня> 2.0–4,5 мм в этом горизонте по сравнению с другими горизонтами почвы. MRD варьировала между генотипами и внутри них во всех почвенных горизонтах, никаких тенденций, которые можно было бы связать с категоризацией генофонда, не наблюдалось (Рисунок 3D). MRD L. cul. Eston и L. erv . L-01-827A были похожи, но значительно ниже (α = 5%) по сравнению со всеми другими генотипами. Lens odemensis IG 72623 имел самый высокий (значимый, α = 5%) MRD.
РИСУНОК 5. Процент распределения общей площади поверхности корня для шести классов диаметра корня в трех горизонтах почвы для семи генотипов дикой чечевицы по сравнению с культурной чечевицей ( Lens culinaris Eston).Линейные столбцы представляют собой значения LSD при P <0,05 для сравнения генотипов в пределах класса диаметра. (А) Горизонт; (Б) Б горизонт; (В) С горизонт.
Для генотипов в первичном генофонде: L. cul . У Eston была самая высокая доля RV в классах диаметра корня <2 мм и наименьшая в классах диаметра корня> 2 мм (Рисунок 6A). Для генотипов во вторичном генофонде — L. ode . IG 72623 имел значительно меньшую (α = 5%) долю корней в классах диаметра корня <2 мм по сравнению с L.lam . IG 110813, но имел более высокую долю RV при классах диаметров корня> 2 мм. Генотипы в третичном генофонде имели сходные RV в большинстве классов диаметров (рис. 6А). TRT также различается между генотипами и внутри генотипов, но четких закономерностей не выявлено из-за высокой вариабельности. Lens lamottei IG 110813 имеет самый низкий TRT (значимый, α = 5%) TRT, в то время как у L. cul . Eston был похож на таковых других видов (рис. 3E). TRF был значительно выше (α = 5%) для L.Куль . Eston, L. ori . PI 572376, L. ode . IG 72623 и L. erv . IG 72815 по сравнению с другими генотипами.
РИСУНОК 6. Распределение объема корней по шести классам диаметра, оцененное в трех горизонтах почвы для пяти генотипов дикой чечевицы в сравнении с культурной чечевицей ( Lens culinaris Eston). Линейные столбцы представляют LSD при P <0,05 для сравнения в пределах данного класса диаметра корня. (А) Горизонт; (Б) Б горизонт; (В) С горизонт.
В горизонте В L. cul. Eston имел значительно более высокий (α = 5%) TRL, чем все другие генотипы, за исключением L. erv . IG 72815. TRL значительно различается среди видов дикой чечевицы. Самый низкий TRL (α = 5%) наблюдался у L. lam . IG 110813 и L. erv . L-01-827A. Lens ervoides L-01-827A имеет высокую долю своего TRL в классе диаметров, 0-0,5 мм по сравнению со всеми другими классами диаметров. С другой стороны, л.ori . IG 72643, который имел значительно низкие доли TRL в классе диаметров 0-0,5 мм и значительно более высокие доли TRL в классах диаметров> 0,5 мм (рис. 4B). Линза culinaris Eston имела повышенную долю корней в классе диаметров, 0,5 — 1,0 мм. При диаметре корня> 1,0 мм, L. ori . IG 72643 имел значительно более высокие (α = 5%) доли TRL по сравнению с L. cul . Eston.
Линза lamottei IG 110813 и L.erv . L-01-827A имел аналогичный, но значительно более низкий (α = 5%) TRSA по сравнению со всеми другими генотипами (рис. 3B). Lens culinaris Eston имела сходную TRSA по сравнению с L. ori. PI 572376 и L. ode . IG 72623. Все три имели значительно более высокий (α = 5%) TRSA, чем другие генотипы. Когда была оценена картина распределения TRSA по классам диаметров, у всех генотипов была самая высокая доля корней в классе диаметров> 0,5 — 1.0 мм. Lens orientalis IG 72643 имел значительно более высокие (α = 5%) пропорции TRSA по сравнению со всеми другими генотипами в классе диаметра корня> 2,0 — 4,5 мм (Рисунок 5B).
Lens orientalis IG 72643, за которым следует L. ode . IG 72623, имел самый высокий MRD (значимый при α = 5%) и оба были L. lam . IG 110813 и L. erv . L-01-827A имел значительно более низкий (α = 5%) MRD, чем другие виды. Остальные виды имели MRD, аналогичную таковой у L.Куль . Эстония (рис. 3D). Аналогичная пропорция RV наблюдалась во всех генотипах при классах диаметра корня> 0,5 — 1,0 мм и> 1,5 — 3,5 мм (Рисунок 6B). Линза восточная IG 72643 имела значительно более низкую (α = 5%) долю ПЖ по сравнению с другими генотипами при классах диаметра корня 0–0,5 и> 1–1,5 мм. Для классов диаметров> 3,5 — 4,5 мм, L. ori . IG 72643 и оба L. erv . у генотипов была значительно более высокая (α = 5%) пропорция RV по сравнению с другими генотипами (Рисунок 6B).
Вся дикая чечевица имела сходный TRT по сравнению с L. cul . Eston (рис. 3E). Однако TRF были аналогичными у L. cul . Eston, L. ori . PI 572376 и L. erv . IG 72815, и они были значительно выше (α = 5%), чем у других генотипов. Линза lamottei IG 110813 и L. erv . L-01-827A имел аналогичный TRF, но он был значительно ниже (α = 5%) по сравнению с таковым у L. cul . Eston.
В горизонте C TRL был аналогичен для генотипов в первичном генофонде, за исключением L.Том . IG 72805, который был значительно ниже (α = 5%), чем остальные. Значения TRL были аналогичны для генотипов во вторичном генофонде и не отличались от таковых для L. cul . Eston. Однако в третичном генофонде L. erv . IG 72815 имел значительно более высокий (α = 5%) TRL по сравнению с L. erv . L-01-827A, также по сравнению со всеми другими генотипами. Хотя распределение пропорций TRL варьировалось между генотипами и внутри них, общая форма распределения по разным классам диаметра корня была сходной (рис. 4C).Большая часть пропорций TRL приходилась на класс диаметров 0–1,0 мм, и не наблюдалось четких закономерностей среди генотипов, принадлежащих к разному генофонду (рис. 4C). При диаметре класса 0 — 0,5 мм л. Том. TRL IG 72805 был аналогичен TRL L. ori PI 572376 и L. erv . L-01-827A, но значительно выше (α = 5%), чем у других видов дикой чечевицы, включая L. cul . Eston. В классе диаметра корня> 0,5 — 1,0 мм — L. cul .У Eston был подобный TRL для всех генотипов, кроме L. tom . IG 72805, который имел значительно более низкий (α = 5%) TRL. Линза culinaris Eston, L. ori. IG 72643 и L. ode . IG 72623 имел аналогичные, но значительно более высокие (α = 5%) доли TRL для класса диаметра корня> 1,0–1,5 мм. Все генотипы дикой чечевицы имели аналогичные пропорции TRL по сравнению с L. cul . Eston в классах диаметра корня> 1,5 мм (рис. 4C).
Опочивенная линза IG 72805 и L.erv . IG 72815 имел значительно (α = 5%) более низкий и более высокий t TRSA, соответственно, по сравнению со всеми другими генотипами чечевицы (рис. 3B). У корня диаметром 0 — 0,5 мм, L. ori . PI 572375 имел сходный TRSA по сравнению с L. tom . IG 72805, L. lam . IG 110813 и L. erv . L-01-827A, и эта доля была значительно выше (α = 5%), чем у остальных генотипов (рис. 5C). Для класса диаметра корня> 0,5 — 1,0 мм все дикие генотипы имели сходные пропорции TRSA по сравнению с L.Куль . Eston. Линза восточная PI 572376 и L. lam . IG 110813 имел значительно более низкие (α = 5%) пропорции TRSA в классе диаметра корня> 1,0–1,5 мм по сравнению с другими генотипами. Lens odemensis IG 72623 имел значительно более высокие (α = 5%) пропорции TRSA в> 1,5–2,0 мм по сравнению с другими генотипами (рис. 5C).
MRD большинства диких генотипов аналогичен MRD L. cul . Eston кроме L.ori PI 572376 и L. tom . IG 72805 (Рисунок 3D). В горизонте C высокая доля RV во всех генотипах приходилась на классы диаметра корня, 0,5 — 3,5 мм (Рисунок 6C). Хотя L. ode . IG 72623 имел более высокую долю RV в классе диаметра корня при диаметре 0-0,5 мм, это не сильно отличалось от такового у L. cul . Eston. Для всех классов диаметров не было значительных различий в TRT и TRF между любой из дикой чечевицы и L.Куль . Eston (Рисунки 3E, F).
Клубеньки дикой чечевицы по сравнению с
L. culinarisВсе генотипы чечевицы были инокулированы одним и тем же штаммом ризобий, но почва, использованная для этого эксперимента, не была стерилизована. В горизонте А L. lam. IG 110813 и L. cul. Eston были генотипами с наименьшим и наибольшим средним числом клубеньков, соответственно, и они значительно отличались (α = 5%) от таковых для других видов дикой чечевицы.В горизонте В L. erv. L-01-827A имел самое низкое (α = 5%) среднее количество узелков. Всего L. cul . В Эстонии было самое высокое среднее количество конкреций по горизонтам A и B. В горизонте С конкреции не образовывались. Между повторениями был обнаружен широкий диапазон количества клубеньков, поэтому общее, среднее значение и диапазон общего количества клубеньков для комбинированных горизонтов A и B представлены по генотипу (Таблица 2). Все оцениваемые генотипы имели больше узелков в горизонте A по сравнению с горизонтом B.Среди генотипов дикой чечевицы — L. tom . IG 72805, L. erv . IG 72815 и L. ode . IG 72623 имел самое высокое среднее количество клубеньков, которые значительно отличались (α = 5%) только от таковых для L. lam . IG 110813 и L. erv . L-01-827A.
ТАБЛИЦА 2. Диапазон и среднее количество клубеньков в горизонтах почвы A и B для пяти видов дикой чечевицы по сравнению с Lens culinaris Eston (Среднее ± SE).
В большинстве случаев форма узелков, образованных каждым генотипом в этом эксперименте, была уникальной. Lens culinaris Eston образовали длинные одиночные узелки с закругленными концами, похожие по форме на узелки, наблюдаемые у L. tom , IG 72805 (рис. 7A, F). Клубеньки L. ori . PI 572376 сформировал прикрепления к корням как единое основание, на котором развилось несколько почек, образуя форму кластера (рис. 7B). Lens orientalis Узелки IG 72643 были прикреплены к корню большой ладонной поверхностью с разветвлениями, напоминающими пальцы (Рисунок 7E). Lens odemensis IG 72623 сформировал скопление скоплений узелков с пальцеобразными ветвями (рис. 7C). Lens lamottei IG 110813 имел одиночные длинные узелки с зеленой основой и интенсивным розовым цветом на концах (рис. 7D). Lens ervoides L-01-827A имел большие узелки, которые прикреплялись узким основанием к корню, но разветвлялись в виде пальцевидных зачатков (рис. 7G), а у L. erv . IG 72815 имел продолговатые узелки с зелеными основаниями, которые появлялись либо поодиночке, либо парами (Рисунок 7H).
РИСУНОК 7. Вариации формы клубеньков, обнаруженные в корневой системе Lens culinaris Eston и семи генотипов дикой чечевицы, инокулированных штаммами Rhizobium leguminarum ( Rhizobium leguminosarum biovar vice Беккер Андервуд, Канада). (A) L. cul . Eston (B) L. orientalis PI 572376 (C) L. odemensis IG 72623 (D) L. lamottei IG 110813 (E) L. orientalis IG 726415 (F) L. tomentosus IG 72805 (G) L. ervoides L-01-827A (H) L. ervoides IG 72815.
Обсуждение
Обширные различия между генотипами дикой чечевицы и L.Куль. Eston наблюдались по большинству оцениваемых параметров. Высота растений была одинаковой у всех генотипов, кроме L. ori , кроме . PI 572376, у которого были значительно более короткие растения. Несмотря на то, что он короткий, он считается одним из генотипов с хорошо развитой корневой системой, простирающейся до горизонта C. Это может указывать на то, что высота растения не является надежным показателем глубины укоренения и, следовательно, не является предиктором засухоустойчивости для генотипов дикой чечевицы, тем более что большинство видов дикой чечевицы описаны как низкорослые по привычке роста.
Большинство генотипов зацвело через 36-42 дня после посева, за исключением 2 видов L. ervoides , которые зацвели значительно раньше ( L. erv . L-01-827A) или позже ( L. erv . IG 72815 ), чем другие. Это может отражать возможное взаимодействие гена с окружающей средой в центрах происхождения, где эти генотипы развились (Ladizinsky, 1993), или специфические реакции на качество освещения в помещении (Yuan et al., 2017).
Генотипы дикой чечевицы дали множество легких семян, которые были небольшими по сравнению с L.Куль. Eston (см. Таблицу 1). Количество стручков не было напрямую связано с общим количеством произведенных семян, учитывая их неопределенный характер роста, что привело к непрерывному образованию цветов, стручков и семян. Мелкие семена могут быть адаптивной стратегией, которая обеспечивает распространение семян в дикой природе, но потенциально может снизить урожай, если предполагается интрогрессия в крупнозернистые семена L. culinaris . Три диких генотипа ( L. ori . PI 572376, L. ode IG 72623 и L.erv . IG 72815) имели хорошо развитую корневую систему, но лишь небольшая часть их биомассы была преобразована в урожай семян (рис. 2). Эффект был еще больше в условиях дефицита влаги (данные не показаны). Однако более многочисленные и мелкие семена могут быть полезны для добавления генетического разнообразия для увеличения урожайности в программах селекции в Юго-Восточной Азии, где предпочтительны мелкосеменные Lens culinaris .
Корневая система культурной чечевицы, а также других бобовых культур, выращиваемых в полевых условиях в Саскачеване, оказалась неглубокой: 77-85% биомассы корней было обнаружено в первых 40 см почвы (Gan et al., 2009). Мы обнаружили, что более 90% корневой биомассы приходится на верхние 40 см почвы, что превышает количество, зарегистрированное в поле. Это могло произойти из-за ограничений, накладываемых сторонами трубок на распространение корня во время роста, заставляя боковые корни расти вниз. Наличие хорошо развитой глубокой корневой системы, которая проникает в более глубокие слои почвы, важно, поскольку большинство сельскохозяйственных систем подвержены периодам засухи. Биомасса корней была распределена по-разному в трех почвах в зависимости от изученного генотипа (Рисунок 1).Это означает, что вариабельность корневой биомассы может быть использована для дальнейшего улучшения культивируемой чечевицы.
Определенные особенности корня могут также иметь потенциальное влияние на будущее генетическое улучшение чечевицы. Оценка доли распределения корней по разным горизонтам почвы (см. Рис. 4–6) показала, что в горизонте А аналогичные TRL и RLD наблюдались для L. cul . Eston и дикие генотипы, такие как L. ori . PI 572376, L. ode . IG 72623 и L. erv .IG 72815, каждый из которых представляет отдельный генофонд. Однако при рассмотрении пропорций корней, распределенных по разным классам диаметра корней, сходства не наблюдалось (рисунки 3A, C). В горизонте C большинство диких генотипов имели более высокие доли TRL при диаметре корня <0,5 мм (т.е. более тонкие корни) по сравнению с L. cul . Эстон подразумевает, что они смогли более эффективно использовать как воду, так и питательные вещества, учитывая, что основные функции тонких корней — это поглощение воды и питательных веществ (Liu et al., 2010). TRL с диаметром корня> 2,0 мм были обнаружены у двух видов дикой чечевицы ( L. ori . IG 72643 и L. ode. IG 72623) как в горизонтах A, так и в горизонтах B, что может означать, что они более эффективны в длинных условиях. дистанционная транспортировка воды и питательных веществ к побегам. Эти генотипы могут быть кандидатами для использования в стратегиях разведения, где требуется одновременная продуктивность как при дефиците влаги, так и при дефиците питательных веществ. Корни диаметром от 0 до 2,0 мм, как известно, обладают высокой пластичностью и способностью изменять свой рост и развитие, чтобы приспособиться к изменяющимся условиям окружающей среды, например, у риса в ответ на засуху (Wang et al., 2009). Это говорит о том, что существует широкий простор для пластичности как у диких, так и у культивируемых видов чечевицы, поскольку большая часть их TRL попадает в этот диапазон во всех трех почвенных горизонтах. Это может иметь значение для селекции в условиях засухи.
Классификация генофонда диких и культивируемых видов чечевицы по-разному влияла на свойства корней. Например, L. ori . IG 72643 и L. tom . IG 72805 (первичный генофонд), который имел сходную биомассу корней, TRL и RLD, также имел аналогичное распределение TRL в горизонте A по всем классам диаметров.Этого не произошло в горизонте B, где находится L. ori . IG 72643 по сравнению с L. tom . IG 72805, имел значительно более низкие доли TRL в классе диаметров 0-0,5 мм, что подразумевает снижение потенциального поглощения воды и питательных веществ. Кроме того, L. lam . IG 110813 и L. erv . L-01-827A (вторичный и третичный генофонды соответственно) имели сходную биомассу корней, TRL и RLD в горизонтах A и B, но их разделение TRL на различные классы диаметра корня было различным. Lens ervoides L-01-827A имеет большую часть своего TRL в классах диаметров <1 мм, в то время как L. lam . IG 110813 имел более высокие доли TRL при классах диаметров> 1 мм. Наличие очень тонких корней во всех горизонтах почвы, что наблюдается у L. erv . L-01-827A может быть причиной его раннего цветения, поскольку он способен впитывать больше влаги, а при отсутствии более толстых корней для хранения этой влаги направляет избыток влаги в урожай. Эту стратегию можно использовать, когда раннее созревание представляет интерес в программах разведения.Более толстые корни (> 2 мм в диаметре) с соответствующими более высокими пропорциями RV и TRSA имеют преимущества в том, что более толстые корни могут проникать в твердые поверхности почвы. Это позволяет им более эффективно добывать воду и питательные вещества (Clarkson, 1985; Gu et al., 2017). Эти признаки наблюдались как у L. ori . IG 72643 и L. ode . IG 72623, что делает их главными кандидатами на интрогрессию при разведении в средах, подверженных засухе и нехватке питательных веществ.
Корни большего диаметра позволяют растениям проникать в плотную почву, тем самым способствуя пористости почвы (Goss, 1991), а более крупные поры способствуют формированию микробных сообществ почвы (Lavelle et al., 2004). Диаметр менее двух мм считается точкой отсечения, при которой корни классифицируются как «мелкие», особенно у других культур с большим диаметром корня (Merrill et al., 2005; Zobel, 2005). Лю и др. (2010) предположили, что существует необходимость в создании другой шкалы категорий диаметра для зернобобовых и, в частности, для чечевицы, учитывая, что большинство корней чечевицы относятся к этому классу диаметра.Предположение о том, что эти корни функционируют так же, как тонкие корни злаков, не подтверждено никакими научными оценками. Однако шкала диаметров, представленная в настоящем эксперименте, отличалась от шкалы, описанной в оценках полей чечевицы Liu et al. (2010). В этих экспериментах, которые проводились в закрытом помещении на ортическом темно-коричневом черноземе, почве, аналогичной почве, использованной Liu et al., Были более высокие доли TRL и TRSA при классах диаметра корня> 2 мм для культурной чечевицы.(2010). Следовательно, различия в пропорциях распределения корневых признаков могли быть результатом разных условий роста чечевицы и того факта, что оценивался разный, гораздо более широкий генотипический диапазон зародышевой плазмы чечевицы. Тем не менее, шкала диаметра корня должна быть установлена для каждой конкретной культуры, включая чечевицу, чтобы сделать значимые выводы относительно классов диаметра корня.
Другие важные характеристики корня включают объем корня (RV) и общую площадь поверхности корня (TRSA).RV является зеркалом размера корневой системы, ключевой характеристики растений, которые поглощают воду и питательные вещества из почвы (Fageria, 2004). RV — это произведение длины корня и диаметра корня, в то время как TRSA включает диаметр корня. Диаметр корня влияет на дальние пути транспортировки воды и питательных веществ к тканям растений и составляет большую часть TRSA, ответственного за обмен между корнем и почвой (Waisel and Eshel, 2002; Gu et al., 2017). В горизонте А — L. cul .Eston и L. erv . L-01-827A имел наименьшую MRD, тогда как оба — L. ori. IG 72643 и L. ode . IG 72623 имел значительно (α = 5%) более высокий MRD по сравнению с L. cul . Eston в горизонтах A и B. Таким образом, согласно литературным данным (Waisel, Eshel, 2002; Gan et al., 2011), как L. ori. IG 72643 и L. ode . Корни IG 72623 с более толстым диаметром могут способствовать транспортировке и хранению воды и питательных веществ на большие расстояния по сравнению с другими видами дикой чечевицы и L.Куль . Eston. Может быть ошибочным рассматривать только MRD во время оценок, учитывая, что он не указывает, какие классы диаметра корня присутствуют или их распределение в каждом генотипе и горизонте почвы. Важно знать пропорции корней в разных классах диаметров, чтобы сделать вывод о потенциальной роли и функции корней. Например, в горизонтах А и С L. ori . PI 572376 имеет наибольшее количество TRSA в классе диаметров 0-0,5 мм (рис. 4). Эта значительно более высокая (α = 5%) пропорция TRSA, возможно, позволила ему усвоить больше питательных веществ и воды, что привело к получению большого количества семян, которые он произвел.Было бы полезно понять реакцию, которую этот вид проявляет в условиях засухи.
Объем корня также играет роль в усвоении питательных веществ. Предыдущие исследования в этой области, проведенные Gan et al. (2011) сообщили, что L. culinaris «CDC Glamis» накапливают RV около 35 мм 3 см -3 в поле при позднем цветении. Наши результаты в помещении показали, что L. cul . Эстон накапливал более высокие ПП (55,4 ± 7,3 мм 3 см -3 ), а также имел различную долю ПП в трех горизонтах почвы.Они сообщили, что в чечевице 59,0, 20,8 и 16,2 процента RV было накоплено на глубинах почвы 0-20, 20-40 и 40-60 см соответственно, а остаток — в более глубоких слоях. Мы обнаружили 47,6, 42,9 и 9,5 процента RV на этих глубинах почвы. Повышенная доля RV в нашем исследовании в горизонте B могла быть результатом разрыхления поверхности почвы во время заполнения трубы, что могло обеспечить больший доступ корней в более глубокие слои почвы.
Высокая изменчивость наблюдалась в общем количестве кончиков корней, которые в основном были обнаружены в классе диаметра корня ≤ 1 мм (данные не показаны).Это могло быть результатом воздействия промывки корней после разделения трех горизонтов почвы. Возможно, это могло привести к увеличению количества корневых кончиков, которые были неотличимы от базового количества реальных кончиков, обнаруженных программой анализа корня WinRHIZO TM . Поскольку внесенная изменчивость не могла быть оценена, различия в общем количестве кончиков корней были представлены в качестве индикатора, поскольку реальные значения были затенены. Тем не менее, количество кончиков корней дает представление о том, как корни проникают в слои почвы и разрастаются в них.Приведенные значения могут служить руководством в будущих исследованиях сенсорной способности корней и синтеза регуляторов роста растений. То же самое относится и к объяснению наблюдаемой изменчивости общего количества вилок.
Корни также служат местом фиксации азота. Количество азота, зафиксированного в бобовых, представляет собой комбинацию того, какое количество азота может удалить растение из почвы и сколько зафиксировано ризобиями в их клубеньках. Сообщается, что чечевица предпочитает биологическую азотфиксацию (Zakeri et al., 2012). В этом исследовании почва была обеднена азотом, и его не применяли. Таким образом, предполагается, что почти весь азот в образующемся сухом веществе является результатом активности конкреций. Мы обнаружили, что количество клубеньков, образовавшихся на корнях дикой и культурной чечевицы в горизонтах A и B, сильно варьировало. Эта изменчивость могла быть результатом клубеньков с местными бактериями из почвы, а также бактериями из внесенного посевного материала или и тем, и другим. Lens culinaris В Эстонии было гораздо большее количество узелков по сравнению с тем, которое сообщалось Abi-Ghanem et al.(2011). Возможной причиной этого могут быть различия в используемых штаммах ризобий в сочетании с короткой продолжительностью эксперимента. Lens culinaris В Эстонии было наибольшее количество клубеньков, но когда его общая биомасса (вегетативная и репродуктивная) была разделена на общее количество клубеньков, получилось L. cul . Eston не так эффективно фиксировал азот по сравнению с L. ori . IG 72643, L. lam . IG 110813 и L. erv .L-01-827A. Это было дополнительно продемонстрировано относительно низкими значениями SPAD культурной чечевицы по сравнению с генотипами дикой чечевицы (данные не показаны). Следовательно, существует потребность в дальнейшем улучшении азотфиксирующей способности культурной чечевицы, и использование диких видов чечевицы может обеспечить полезные генетические вариации в этом отношении. У разных генотипов клубеньки разной формы, а иногда на одном и том же растении наблюдались две разные формы (рис. 7). Это поднимает несколько вопросов, например, могут ли оптимальные штаммы ризобий быть видоспецифичными или специфичными для генотипа, учитывая, что наша почва не была стерилизована.Более того, некоторые клубеньки в генотипах дикой чечевицы были временными. Почему они были временными и какие штаммы ризобий (из почвы или инокулята) сформировали эти клубеньки, требует дальнейшего изучения; может случиться так, что азотфиксирующий метаболизм клубеньков с переменным цветом может быть различным. Следовательно, необходимо провести глубокие исследования биологии клубеньков у этих видов чечевицы, чтобы определить, в какой степени наблюдаемые вариации зависели от взаимодействия штамм × генотип.Может существовать потенциал для улучшения биологической азотфиксации культурной чечевицы посредством межвидовой гибридизации с L. culinaris , которая была успешно разработана для четырех диких видов, о которых здесь сообщается.
Заключение
Выбор генотипов для этого исследования производился на основе их способности к гибридизации с Lens culinaris. В генотипах дикой чечевицы существует широкий спектр фенотипического и генетического разнообразия почти по всем признакам, относящимся к наземным и подземным фенотипам.Это разнообразие может быть дополнительно изучено и использовано как средство повышения устойчивости культурной чечевицы к широкому спектру биотических и абиотических стрессов. Межвидовые гибриды доступны для скрещивания между Lens culinaris и четырьмя дикими видами L. orientalis, L. odemensis, L. tomentosus и L. ervoides . Классификация генофонда, основанная на геномном анализе, не объясняет характер вариаций, обнаруженных в фенотипах корней в этом исследовании. Большое разнообразие корневых признаков наблюдалось даже у небольшого спектра генотипов дикой чечевицы, а также наблюдались значительные различия в корневых признаках между генотипами, принадлежащими к одним и тем же диким видам ( Lens ervoides и L.orientalis). Значительно более высокие значения для всех признаков корня во всех 3 горизонтах почвы были обнаружены у трех генотипов ( L. ori . PI 572376, L. ode . IG 72623 и L. erv . IG 72815) по сравнению с L . кул. Eston. Было замечено, что как у культурной, так и у дикой чечевицы большая часть их TRL приходилась на классы диаметра менее 2 мм, диапазон поглощения относился к «тонким корням». Однако, что это означает в контексте потребностей зернобобовых культур. подлежит дальнейшему определению, поскольку все их корни не могут быть тонкими корнями, как это определено для других культур, особенно для корневой системы злаков.Хотя корневая биомасса и характеристики корней, такие как TRL, TRSA и RV, имеют важное значение, дальнейшее понимание корневых систем может быть получено только путем изучения моделей распределения корней на различной глубине почвы. Кроме того, дикие генотипы с глубокой корневой системой выделяли свои ресурсы в основном на производство биомассы, а не на семена, что означает, что, когда межвидовая гибридизация и интрогрессия станут частью долгосрочной селекционной стратегии чечевицы, необходимо будет разработать соответствующие стратегии отбора для одновременного отбора урожайность и корневые признаки.Среди генотипов дикой чечевицы существует широкий диапазон количества и формы клубеньков, что требует дальнейших исследований в контексте азотфиксации и потенциального урожая.
Авторские взносы
LG: Ответственный за дизайн, выполнение, анализ экспериментов и подготовку рукописи с участием руководителя. Утвердил представленный окончательный вариант и проверил правильность всех аспектов этой работы. АВ: Разработка исходных концепций экспериментальных работ в рамках грантовых заявок, принимаемых различными агентствами, финансирующими исследования.Под руководством LG и отредактированной рукописи для публикации.
Финансирование
Это исследование получило финансовую поддержку от Канадской программы кафедры промышленных исследований NSERC (#IRCPG 395994-09), производителей зернобобовых в Саскачеване (#BRE 1202) и Глобального фонда разнообразия сельскохозяйственных культур.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Большое спасибо бригаде полевой лаборатории Центра развития сельскохозяйственных культур за их вклад во время сбора почвы и промывки корней. Это исследование финансировалось: Программой промышленных исследований NSERC по генетическому усовершенствованию чечевицы, зернобобовыми из Саскачевана, Университетом Саскачевана и Глобальным фондом разнообразия сельскохозяйственных культур.
Список литературы
Аби-Ганем Р., Карпентер-Боггс Л. и Смит Дж. Л. (2011). Влияние сортов на азотфиксацию гороха и чечевицы. Biol. Fertil. Почвы 47, 115–120. DOI: 10.1007 / s00374-010-0492-6
CrossRef Полный текст | Google Scholar
Кларксон, Д. Т. (1985). Факторы, влияющие на усвоение растениями минеральных питательных веществ. Annu. Rev. Plant Physiol. 36, 77–115. DOI: 10.1146 / annurev.pp.36.060185.000453
CrossRef Полный текст | Google Scholar
Фагерия, Н. К. (2004). Влияние сухого вещества и длины корней на рост пяти полевых культур при различных уровнях содержания цинка и меди в почве. J. Plant Nutr. 27, 1517–1523. DOI: 10.1081 / PLN-200025995
CrossRef Полный текст | Google Scholar
Гахония, Т. С., Али, О., Саркер, А., Рахман, М. М., и Эрскин, В. (2005). Свойства корней, усвоение питательных веществ, урожайность зерна в разных местах и соотношение цена / качество для двух сортов чечевицы ( Lens culinaris , Medikus.). Почва растений 272, 153–161. DOI: 10.1007 / s11104-004-4573-x
CrossRef Полный текст | Google Scholar
Ган, Ю., Лю Л., Катфорт Х., Ван Х. и Форд Г. (2011). Профили вертикального распределения и временные закономерности роста корней выбранных масличных культур, зернобобовых культур и яровой пшеницы. Crop Pasture Sci. 62, 457–466. DOI: 10.1071 / CP10406
CrossRef Полный текст | Google Scholar
Ган Ю. Т., Кэмпбелл К. А., Янзен Х. Х., Лемке Р., Лю Л. П., Баснят П. и др. (2009). Корневая масса масличных и зернобобовых культур: рост и распределение в почвенном профиле. Банка. J. Plant Sci. 89, 883–893. DOI: 10.4141 / CJPS08154
CrossRef Полный текст | Google Scholar
Горим, Л. Ю., Ванденберг, А. (2017). Оценка видов дикой чечевицы как генетических ресурсов для улучшения засухоустойчивости культивируемой чечевицы. Фронт. Plant Sci. 8: 1129. DOI: 10.3389 / fpls.2017.01129
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Госс, М. Дж. (1991). «Последствия активности корней на почве», в Рост корней растений: экологическая перспектива , изд.Д. Аткинсон (Лондон: Научные публикации Блэквелла), 171–186.
Google Scholar
Гу, Д., Чжэнь, Ф., Ханнауэй, Д. Б., Чжу, Ю., Лю, Л., Цао, В. и др. (2017). Количественная классификация длины и диаметра корня риса ( Oryza sativa L.) с использованием анализа изображений. PLOS ONE 12: e0169968. DOI: 10.1371 / journal.pone.0169968
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ладизинский, Г. (1993). Одомашнивание чечевицы: от качества доказательств и аргументов. Экон. Бот. 47, 60–64. DOI: 10.1007 / BF02862206
CrossRef Полный текст | Google Scholar
Лавель П., Руланд К., Бине Ф., Диуф М. и Керсанте А. (2004). «Регулирование микробной активности корнями и почвенными беспозвоночными», в Регулирование микробной активности , ред. Ф. Бускот и А. Варма (Берлин: Springer).
Лю Л., Ган Ю., Бюкерт Р., Рис, К. В., и Варкентин, Т. (2010). Прекрасное распределение корней у масличных и зернобобовых культур. Crop Sci. 50, 222–226. DOI: 10.2135 / cropci2009.03.0156
CrossRef Полный текст | Google Scholar
Меррилл, С. Д., Танака, Д. Л., и Хэнсон, Дж. Д. (2005). Сравнение миниризотронов с неподвижными стенками и миниризотронами с герметичными стенками для измерения роста тонких корней у восьми видов сельскохозяйственных культур. Agron. J. 97, 1367–1373. DOI: 10.2134 / agronj2004.0209
CrossRef Полный текст | Google Scholar
Мюльбауэр, Ф. Дж., Чо, С., Саркер, А., Макфи, К. Э., Койн, К.Дж., Раджеш, П. Н. и др. (2006). Применение биотехнологии в селекции чечевицы для устойчивости к биотическим и абиотическим стрессам. Euphytica 147, 149–165. DOI: 10.1007 / s10681-006-7108-0
CrossRef Полный текст | Google Scholar
Поддер Р., Банница С. и Ванденберг А. (2013). Скрининг зародышевой плазмы дикой и культивируемой чечевицы на устойчивость к Stemphylium blight. Plant Genet. Ресурс. 11, 26–35. DOI: 10.1017 / S1479262112000329
CrossRef Полный текст | Google Scholar
Пратап, А., Кумар, Дж., И Кумар, С. (2014). Оценка диких генотипов чечевицы по агроморфологическим признакам. Legume Res. 37, 11–18. DOI: 10.5958 / j.0976-0571.37.1.002
CrossRef Полный текст | Google Scholar
Саучин Д., Кульшрешта С. (2008). «Прерии», в От воздействий к адаптации: Канада в изменяющемся климате 2007 , ред. Д. С. Леммен, Ф. Дж. Уоррен, Дж. Лакруа и Э. Буш (Оттава, Онтарио: Правительство Канады), 275–328.
Google Scholar
Сингх, Д., Сингх, К.К., Томар, Р.С.С., Таунк, Дж., Сингх, Р., Маурья, С. и др. (2016). Молекулярный ассортимент Lens видов с различной адаптацией к условиям засухи с использованием SSR-маркеров. PLOS ONE 11: e0147213. DOI: 10.1371 / journal.pone.0147213
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сингх М., Бишт И. С., Камар С., Датта М., Бансал К. К., Карале М. и др. (2014). Глобальная одичавшая ежегодная коллекция Lens : потенциальный ресурс для расширения генетической базы чечевицы и повышения урожайности. PLOS ONE 9: e107781. DOI: 10.1371 / JOURNAL.PONE.0107781
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Трон, С., Боднер, Г., Лайо, Ф., Ридольфи, Ф., и Лейтнер, Д. (2015). Может ли разнообразие корневой архитектуры объяснить эффективность использования воды растениями? Модельное исследование. Ecol. Модель. 312, 200–210. DOI: 10.1016 / j.ecolmodel.2015.05.028
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Туллу, А., Таран, Б., Брейткрейц, К., Buchwaldt, L., Banniza, S., Warkentin, T. D., et al. (2006). Локус количественного признака устойчивости к фитофторозу [ Ascochyta lentis ] картирован близко к гену устойчивости к антракнозу [ Colletotrichum truncatum ] у чечевицы. Банка. J. Plant Pathol. 28, 558–595. DOI: 10.1080 / 07060660609507337
CrossRef Полный текст | Google Scholar
Вайсель, Ю., и Эшель, А. (2002). «Функциональное разнообразие различных компонентов одной корневой системы», в Plant Roots, The Hidden Half , ред. Y.Вайзел, А. Эшел и У. Кафкафи (Нью-Йорк, штат Нью-Йорк: Марсель Деккер), 157–174.
Google Scholar
Ван Х., Сиопонгко Дж., Уэйд Л. Дж. И Ямаути А. (2009). Фрактальный анализ корневой системы растений риса в ответ на стресс от засухи. Environ. Exp. Бот. 65, 338–344. DOI: 10.1016 / j.envexpbot.2008.10.002
CrossRef Полный текст | Google Scholar
Wong, M. M. L., Gujaria-Verma, N., Ramsay, L., Yuan, H. Y., Caron, C., Diapari, M., et al. (2015).Классификация и характеристика видов в пределах рода Lens с использованием генотипирования путем секвенирования (GBS). PLOS ONE 10: e0122025. DOI: 10.1371 / journal.pone.0122025
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Е. Г., Макнейл Д. Л. и Хилл Г. Д. (2002). Селекция на устойчивость к аскохитовой болезни чечевицы. Растение породы. 121, 185–191. DOI: 10.1046 / j.1439-0523.2002.00705.x
CrossRef Полный текст | Google Scholar
Юань, H.Ю., Саха, С., Ванденберг, А., Кирстин, Э., Бетт, К. Э. (2017). Реакция цветения и роста культурной чечевицы и зародышевой плазмы дикой Lens на различия в соотношении красного и дальнего красного и фотосинтетически активную радиацию. Фронт. Plant Sci. 8: 386. DOI: 10.3389 / fpls.2017.00386
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Закери, Х., Лафонд, Г. П., Шенау, Дж. Дж., Пахлавани, М. Х., Ванденберг, А., Мэй, В. Ф. и др. (2012).Продуктивность чечевицы в зависимости от погоды, продолжительности нулевой обработки почвы и азота в Саскачеване. Agron. J. 104, 1501–1509. DOI: 10.2134 / agronj2011.0339
CrossRef Полный текст | Google Scholar
Зобель, Р. У. (2005). «Третичные корневые системы» в Корни и управление почвой: взаимодействие между корнями и почвой. Агрономические монографии 48 , ред. Р. В. Зобель и С. Ф. Райт (Мэдисон, Висконсин: ASA, CSSA и SSSA), 35–56.
Зобель, Р. У., Кинрейд, Т. Б.и Балигар В.С. (2007). Диаметр тонких корней может изменяться в ответ на изменение концентрации питательных веществ. Почва растений 297, 243–254. DOI: 10.1007 / s11104-007-9341-2
CrossRef Полный текст | Google Scholar
Зобель, Р. У., и Вайзел, Ю. (2010). Архитектурная систематика корневой системы растений: основа для корневой номенклатуры. Завод Биосист. 144, 507–512. DOI: 10.1080 / 11263501003764483
CrossRef Полный текст | Google Scholar
Перейти к основному содержанию ПоискПоиск
- Где угодно
Поиск Поиск
Расширенный поиск- Войти | регистр
- Подписка / продление
- Учреждения
- Индивидуальные подписки
- Индивидуальные продления
- Библиотекари 9134 9134
- Выплаты и полные заказы Чикагский пакет
- Полный цикл и охват содержимого
- Файлы KBART и RSS-каналы
- Разрешения и перепечатки
- Инициатива развивающихся стран Чикаго
- Даты отправки и претензии
- Часто задаваемые вопросы библиотекарей
- заказы, тарифы
- и платежи
- О нас
- Публикуйте у нас
- Недавно приобретенные
- журналов 9010 tners
- Подпишитесь на уведомления eTOC
- Пресс-релизы
- СМИ
- Книги издательства Чикагского университета
- Распределительный центр в Чикаго
- Чикагский университет
- Положения и условия
- Заявление об этике публикаций
- Уведомление о конфиденциальности
- Доступность Chicago Journals
- Доступность университета
- Следуйте за нами на facebook
- Следуйте за нами в Twitter
- Свяжитесь с нами
- Запросы СМИ и рекламы
- Открытый доступ в Чикаго
- Следуйте за нами на facebook
- Следуйте за нами в Twitter
Звуковой детектор корней для выявления грубого корня дерева
Линч, Дж. П. Крутой, дешевый и глубокий: идеотип для оптимизации поглощения воды и азота корневой системой кукурузы. Ann. Бот 112 , 347–357 (2013).
CAS PubMed PubMed Central Статья Google Scholar
Sorgonà, A., Abenavoli, M. R. & Cacco, G. Сравнительное исследование двух подвоев цитрусовых: влияние нитратов на морфотопологию корня и чистое поглощение нитратов. Почва растений 270 , 257–267 (2005).
Артикул CAS Google Scholar
Dupuy, L. X., Fourcaud, T., Lac, P. & Stokes, A. Общая трехмерная конечно-элементная модель крепления деревьев, объединяющая механику почвы и реальную архитектуру корневой системы. Am. J Bot. 94 , 1506–1514 (2007).
PubMed Статья Google Scholar
Куттс, М. П., Нильсен, К. К. Н. и Николл Б.C. Развитие симметрии, жесткости и закрепления в структурной корневой системе хвойных пород. Почва растений 217 , 1–15 (1999).
Артикул Google Scholar
Дюпюи, Л. X., Фурко, Т. и Стокс, А. Численное исследование влияния типа почвы и структуры корней на крепление деревьев. Почва растений. 278 , 119–134 (2005).
CAS Статья Google Scholar
Фурко Т., Джи, Дж. Н., Чжан, З. К. и Стокс, А. Понимание влияния морфологии корня на механизмы опрокидывания: подход к моделированию. Ann. Бот. 101 , 1267–1280 (2008).
PubMed Статья Google Scholar
Лундстрем, Т., Йонас, Т., Стокли, В. и Амманн, В. Закрепление зрелых хвойных пород: крутящий момент сопротивления, геометрия корня-почвенной пластины и ориентация роста корней. Tree Physiol. 27 , 1217–1227 (2007).
PubMed Статья Google Scholar
Николл, Б. К., Гардинер, Б. А. и Пис, А. Дж. Улучшение крепления, обеспечиваемое адаптацией лесных деревьев к ветровому стрессу. Лесное хозяйство. 81 , 389–398 (2008).
Артикул Google Scholar
Tamasi, E. et al. .Влияние ветровой нагрузки на развитие и архитектуру корневой системы у сеянцев дуба ( Quercus robur L.). Деревья 19 , 374–384 (2005).
ADS Статья Google Scholar
МГЭИК. Изменение климата 2013: основы физических наук. Вклад рабочей группы I в пятый оценочный доклад Межправительственной группы экспертов по изменению климата. (Издательство Кембриджского университета, 2013 г.).
IPCC. Изменение климата 2014: воздействия, адаптация и уязвимость. Часть А: глобальные и отраслевые аспекты. Вклад рабочей группы II в пятый оценочный доклад Межправительственной группы экспертов по изменению климата. (Издательство Кембриджского университета, 2014).
МГЭИК. Изменение климата 2014: сводный отчет. Вклад рабочих групп I, II и III в пятый оценочный доклад Межправительственной группы экспертов по изменению климата. 151 (Женева, 2014 г.).
Ганди, К. и др. . Катастрофический ураган и меры по сокращению расхода топлива изменяют сообщества жужелиц (Coleoptera: Carabidae) в суббореальных лесах Северной Америки. Для. Ecol. Manag 256 , 1104–1123 (2008).
Артикул Google Scholar
Маккарти, Дж. К. и др. . Прогнозирование образования пятен и деградации упавших деревьев после урагана в лучевом лесу сосны. Для.Ecol. Manag 260 , 1456–1466 (2010).
Артикул Google Scholar
Терхо, М. и Халаксела, А. М. Характеристики разложения опасных деревьев тилиа, бетула и асер, вырубленных городскими лесопользователями в районе Хельсинки. Лесное хозяйство. 81 , 151–159 (2008).
Артикул Google Scholar
Cermak, J., Hruska, J., Martinkova, M. & Prax, A. Корневые системы городских деревьев и их выживаемость возле домов проанализированы с использованием георадаров и методов сокодвижения. Почва растений 219 , 103–116 (2000).
CAS Статья Google Scholar
Данжон, Ф., Синокет, Х., Годин, К., Колин, Ф. и Дрексхаге, М. Характеристика структурной корневой архитектуры дерева с использованием трехмерной оцифровки и программного обеспечения AMAPmod. Почва растений 211 , 241–258 (1999).
CAS Статья Google Scholar
Ди Иорио, А., Лассер, Б., Сиппа, Г. С. и Чиатанте, Д. Архитектура корневой системы деревьев Quercus pubescens , растущих на разных уклонах. Ann. Бот. 95 , 351–361 (2005).
PubMed Статья Google Scholar
Ватанабэ, Т. и др. . Морфогенез риса и архитектура растений: измерение, спецификация и реконструкция структурного развития с помощью трехмерного архитектурного моделирования. Энн Бот 95 , 1131–1143 (2005).
PubMed PubMed Central Статья Google Scholar
Di Iorio, A., Lasserre, B., Scippa, G. S. & Chiatante, D. Структура вторичного утолщения в корневой системе Quercus cerris . Tree Physiol. 27 , 407–412 (2007).
PubMed Статья Google Scholar
Вальдес-Родригес, О.А., Санчес-Санчес, О., Перес-Васкес, А., Каплан, Дж. С. и Данжон, Ф. Jatropha curcas L. Структура корней и рост в различных почвах. Sci. Мир. J. 2013 , 827295, https://doi.org/10.1155/2013/827295 (2013).
Артикул Google Scholar
Di Iorio, A., Lasserre, B., Petrozzi, L., Scippa, GS & Chiatante, D. Адаптивный продольный рост боковых корней первого порядка древесных пород ( Spartium junceum ) к склону и различным почвенным условиям — рост поверхностных корней вверх. Environ Exp Bot 63 , 207–215 (2008).
Артикул Google Scholar
Ву, Дж., Пагес, Л., Ву, К., Янг, Б. и Го, Ю. Трехмерная архитектура осевых корней кукурузы, выращиваемой в полевых условиях. Почва растений. 387 , 363–377 (2015).
CAS Статья Google Scholar
Соргона А., Прото А. Р., Абенаволи Л. и Ди Иорио А. Пространственное распределение биомассы грубых корней и углерода в оливковом саду с высокой плотностью посадки: влияние механических методов сбора урожая. Деревья 32 , 919–931 (2018).
Артикул CAS Google Scholar
Годин, К., Костес, Э. и Синокет, Х. Моделирование архитектуры растений: виртуальные растения и сложные системы в Архитектура растений и ее манипуляции (изд. Тернбулл, C.G.N.) 238–287 (Blackwell, 2005).
van der Heijden, G.W.A.M., de Visser, P.H.B. & Heuvelink, E. Измерения для функционально-структурных моделей растений в Функционально-структурное моделирование растений в растениеводстве (ред. Вос, Дж., Марселис, Л.Ф.М., де Виссер, П.Х. Б., Струик, П.К., Эверс, Дж. Б.) 13–25 (Springer Berlin, 2007).
Mairhofer, S. et al . RooTrak: автоматическое восстановление трехмерной структуры корней растений в почве по изображениям рентгеновской микрокомпьютерной томографии с использованием визуального отслеживания. Plant Physiol. 158 , 561–569 (2012).
CAS PubMed Статья Google Scholar
Гао, В., Шлютер, С., Блазер, С. Р. Г. А., Цзяньбо Шен, Дж.& Веттерлейн, Д. Метод на основе формы для автоматической и быстрой сегментации корней в почве по изображениям рентгеновской компьютерной томографии: Rootine. Почва растений. 441 , 643–655 (2019).
CAS Статья Google Scholar
Maenhout, P. et al . Полуавтоматическая сегментация и визуализация сложных ненарушенных корневых систем с помощью рентгеновской μCT. Soil & Tillage Res 192 , 59–65 (2019).
Артикул Google Scholar
Корона-Лопес, Д. Д. Дж., Соммер, С., Рольфе, С. А., Подд, Ф. и Грив, Б. Д. Электроимпедансная томография как инструмент фенотипирования корней растений. Растительные методы. 15 , 49, https://doi.org/10.1186/s13007-019-0438-4 (2019).
Артикул PubMed PubMed Central Google Scholar
Росси Р. и др. . Томография электрического сопротивления как неразрушающий метод картирования корневой биомассы в саду. Eur. J. Soil Sci 62 (2), 206–215 (2011).
Артикул Google Scholar
Zhou, X.C. И Ло, X.W. Достижения в методах неразрушающего измерения и трехмерной визуализации корней растений на основе машинного зрения в Труды 2-й Международной конференции по биомедицинской инженерии и информатике .1–5. (Тяньцзинь, BMEI’09, 2009).
Jahnke, S. et al . Комбинированная МРТ-ПЭТ анализирует динамические изменения в структурах и функциях растений. Завод J 59 , 634–644 (2009).
CAS PubMed Статья Google Scholar
Hagrey, S. A. Геофизические изображения неоднородности корневой зоны, ствола и влажности. J. Exp. Бот 58 , 839–854 (2007).
Артикул CAS Google Scholar
Фанг, С., Ян, X. и Ляо, Х. Трехмерная реконструкция и динамическое моделирование корневой архитектуры in situ и ее применение для исследования фосфора сельскохозяйственных культур. Завод J 60 , 1096–1108 (2009).
CAS PubMed Статья Google Scholar
Дои, К. Диагностическая визуализация за последние 50 лет: исследования и разработки в области науки и технологий медицинской визуализации. Phys. Med. Биол. 51 , 5–27 (2006).
Артикул Google Scholar
Бутнор, Дж. Р., Дулиттл, Дж. А., Кресс, Л., Коэн, С. и Джонсен, К. Х. Использование георадара для изучения корней деревьев на юго-востоке США. Tree Physiol. 21 , 1269–1278 (2001).
CAS PubMed Статья Google Scholar
Крайнюков, А.& Лякса, И. Обнаружение корней деревьев в городской черте с помощью георадаров. Транспорт и телекоммуникации 17 (4), 362–370 (2016).
Артикул Google Scholar
Ринн, Ф. Оценка корневой пластинки дерева зондированием. Западный лесовод. Осень 2016 , 26–29 (2016).
Google Scholar
Буза, А. К. и Дивос, Р. Оценка устойчивости корней неразрушающими методами. Acta Silv. Lign. Hung 12 , 125–164 (2016).
Артикул Google Scholar
Montagnoli, A. et al. . Текущие изменения в архитектуре корневой системы Pinus ponderosa, растущей на наклонном участке, выявленном с помощью анализа годичных колец. Дендрохронология 58 , 125650 (2019).
Артикул Google Scholar
Ринн, Ф. Основные основы томографии Sonic Tree. J Soc Comm Arb 19 , 8–10 (2014).
Google Scholar
Ринн Ф. Центральные дефекты ультразвуковой томографии. Западный лесовод. Весна 2015 , 38–41 (2015).
Google Scholar
Фишер, Н.И. Статистический анализ циркулярных данных (Cambridge University Press, 1993).
Мардиа, К.В. И Джапп, П. Направленная статистика (Wiley, 2000).
Ковач В.Л. Ориана для окон, вер. 2.01 (Kovach Computing Services, 1994).
Альберс, Б. Анализ распространения звуковых волн в частично насыщенных грунтах с помощью макроскопической линейной пороупругой модели. Пер. Пористая среда. 80 , 173–192 (2009).
CAS Статья Google Scholar
Вей, К. и Муралитаран, К. К. Континуальная теория пористых сред, насыщенных несколькими несмешивающимися жидкостями: I. Линейная пороупругость. Внутр. J Eng. Sci. 40 , 1807–1833 (2002).
MathSciNet МАТЕМАТИКА Статья Google Scholar
Эльзе М. Л., О’Брайен В. Д. и Дармоди Р. Г. Измерение затухания и скорости звука в почвах. Почвоведение. Soc. Являюсь. J 66 , 788–796 (2002).
ADS CAS Статья Google Scholar
Буза, А. К. и Гёнц, Б. Сравнение деревьев и методы неразрушающего контроля. Wood Research. 60 , 45–58 (2015).
Google Scholar
Харрис П. и Эндрюс М. Новые технологии оценки качества древесины для обработки. 3 rd Симпозиум по качеству древесины в Роторуа (1999).
Легг, М. и Брэдли, С. Измерение жесткости стоящих деревьев и срубленных бревен с помощью акустики: обзор. J Acoust Soc Am 139 , 588–604 (2016).
ADS PubMed Статья Google Scholar
Куттс, М. П. Процессы развития в корневых системах деревьев. Банка. J. For. Res 17 , 761–767 (1987).
Артикул Google Scholar
Ахим, А. и Николл, Б. С. Моделирование закрепления деревьев с мелкими корнями. Лесное хозяйство. 82 , 273–284 (2009).
Артикул Google Scholar
Лундстрем, Т., Йонас, Т., Штёкли, В. и Амманн, В. Закрепление зрелых хвойных пород: крутящий момент сопротивления, геометрия корня-почвенная пластина и ориентация роста корней. Tree Physiol. 27 , 1217–1227 (2007).
PubMed Статья Google Scholar
Характеристики роста корней и пространственного распределения для проростков, выращенных в субстрате и пересаженного хлопка
Abstract
В данном исследовании рассматривается пересадка саженцев хлопка, выращенных в искусственный субстрат, в связи с возросшим в последнее время интересом к подходам, экономящим труд при посадке хлопка.Способы выращивания рассады хлопчатника в питомниках влияют на рост корней. Однако основные функциональные реакции роста корней на вариации в методах пересадки рассады хлопчатника изучены недостаточно. Мы оценили реакцию корней хлопчатника ( Gossypium hirsutum L.) на различные методы посадки путем проведения полевых экспериментов с хлопком в 2012 и 2013 годах. Был использован однофакторный случайный блочный дизайн с тремя повторностями и тремя различными схемами посадки хлопка (субстрат для рассады. пересаженный хлопок (SSTC), рассада пересаженного хлопка (ScSTC) и хлопок прямого посева (DSC).Были определены распределения и дисперсии плотности площади корня (RAD) и плотности длины корня (RLD) на разных этапах выращивания хлопка и нескольких компонентов урожая. В целом наблюдались следующие результаты: 1) RAD и RLD были наибольшими вблизи растений (горизонтальное расстояние 0 см), но были ниже на W20 и W40 см в отсутствие пленочного мульчирования, чем на E20 и E40 см с пленочным мульчированием. . 2) Корни были ограничены небольшой глубиной (20-40 см), и глубина корня SSTC и DSC была больше глубины корня ScSTC.3) Сильный рост корней наблюдался в SSTC на этапах цветения хлопчатника и завязывания коробочек. Кроме того, ранний рост корней произошел в ScSTC, а сильный рост корней произошел на всех стадиях роста хлопка в DSC. 4) У растений SSTC было больше боковых корней с более высокой биомассой корней (RB), чем у ScSTC, что привело к более высоким урожаям хлопка. Однако раннее начало роста корней в ScSTC привело к увеличению урожайности семян хлопчатника до заморозков (PFSC). Эти результаты могут быть использованы для вывода о том, как корни хлопка распределяются в почве и улавливают питательные вещества.
Образец цитирования: Zhi X, Han Y, Li Y, Wang G, Feng L, Yang B, et al. (2017) Характеристики роста корней и пространственного распределения для сеянцев, выращенных в субстрате и пересаженного хлопка. PLoS ONE 12 (12): e01. https://doi.org/10.1371/journal.pone.01
Редактор: Дебджани Сихи, Центр экологических наук Мэрилендского университета, США
Поступила: 2 мая 2017 г .; Одобрена: 24 ноября 2017 г .; Опубликован: 22 декабря 2017 г.
Авторские права: © 2017 Zhi et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Все соответствующие данные находятся в документе и его файлах с вспомогательной информацией.
Финансирование: Это исследование было поддержано Индустрией научных исследований национального общественного благосостояния (201203057-01) для XZ и Национальной программой поддержки ключевых технологий (2014BAD03B02-2).
Конкурирующие интересы: Авторы заявили, что никаких конкурирующих интересов не существует.
Введение
Хлопок ( Gossypium hirsutum L.) — одна из важнейших волокнистых культур в мире. Китай является крупнейшим производителем и потребителем хлопка [1–4] и имеет три основных региона выращивания хлопка. В долинах реки Хуанхэ и Янцзы большая часть хлопка перемежается с зерном, что приводит к лучшему улавливанию солнечного света, множественным индексам урожая и экономическим выгодам [5–8].
Конкуренция промежуточных посевов эстафетных растений за воду, питательные вещества и солнечный свет является интенсивной, особенно между хлопком и пшеницей ( Triticumaestivum Linn .), Когда хлопок высевается до сбора урожая пшеницы [9]. Эта конкуренция задерживает рост хлопка и снижает урожайность хлопка. Но пересадка саженцев хлопка является эффективным методом предотвращения или смягчения негативных последствий повторного пересева на обе культуры [10–13] и может устранить холодовой стресс в начале сезона перед пересадкой [12, 14–15].Более того, колонные почвенные блоки, изготовленные из почвы и органических удобрений, обычно используются в рассадниках [3, 16], что может снизить затраты на выращивание саженцев хлопка и снизить заболеваемость болезнями или насекомыми-вредителями, а также улучшить укоренение хлопка [17]. Однако пересадка рассады, которая является интенсивной практикой выращивания хлопка, требует много труда, особенно для изготовления кубиков почвы для посадки рассады в питомнике [18–19]. Таким образом, хотя пересадка саженцев полезна, производители сталкиваются с серьезными проблемами из-за нехватки рабочей силы, вызванной быстрой урбанизацией и перемещением рабочей силы из сельской местности в города [20–21].Чтобы решить проблему ограниченности сельскохозяйственных рабочих в Китае, необходимы высокоэффективные и упрощенные методы пересадки рассады.
Пересадка саженцев с вермикулитовых субстратов, которые широко используются в производстве саженцев сельскохозяйственных культур [22–26], может снизить объем и интенсивность ручного труда, необходимого за счет устранения необходимости делать кубики почвы для облегчения роста промышленных саженцев хлопка и механизированной пересадки [10 , 19]. Кроме того, местные фермеры использовали субстратную пересадку рассады для выращивания рассады хлопка в питомниках.Однако характеристики саженцев хлопка, пересаженных с субстрата и с использованием традиционных методов, и хлопка, посеянного непосредственно в поле, еще не задокументированы.
Корни являются важным связующим звеном между почвой и растениями [27–28]. Кроме того, корневые системы выполняют важные физиологические и биологические функции для роста и урожайности сельскохозяйственных культур [29–30]. Урожайность семенного хлопка и качество ворса зависят от состояния здоровья корней хлопчатника, более того, рост и развитие корня хлопчатника в значительной степени зависят от условий почвы, например текстуры и структуры почвы, как показано в Ahmadi et al.обнаружили, что большая часть корней картофеля (сорт Folva) накапливается в поверхностных слоях крупного песка по сравнению с другими типами почв, однако более высокая плотность корней обнаруживается в супеси, чем в супеси и крупном песке [31–32]. Таким образом, при изучении продуктивности рассады хлопка с использованием различных методов выращивания рассады важно понимать развитие и распространение корней. Поскольку они, как правило, скрыты, анализ корневой системы растений может быть более сложной задачей, чем надземные компоненты растений; тем не менее, распределение корней растений можно точно получить, смыв почву с корней [33].Кроме того, исследования плотности длины корня (RLD) и плотности корневой зоны (RAD) предпочтительнее исследований плотности массы корня для определения распределения корней и характеристики корневых систем, поскольку RLD и RAD более значимы с научной точки зрения для оценки функции корней растений в этом Поглощение воды и питательных веществ корнями зависит от общей площади почвы, с которой корни контактируют [34–38]. Однако мало что известно о том, чем отличаются RLD и RAD при разных методах пересадки рассады.
Основная цель этого исследования заключалась в изучении различий в пространственном распределении корней между непосредственно посеянным хлопком, саженцами хлопчатника, пересаженными из субстрата, и саженцами хлопчатника, пересаженными из кубиков почвы. Кроме того, мы стремимся описать рост корней хлопчатника и двумерное распределение корней хлопчатника на разных стадиях роста, а также определить влияние пересадки рассады на семена хлопчатника и другие компоненты урожая.
Материалы и методы
Экспериментальная площадка
Полевой эксперимент проводился в 2012 и 2013 годах на экспериментальной ферме Института исследования хлопка Китайской академии сельскохозяйственных наук в Аньяне, Хэнань, Китай (36 ° 06 ‘северной широты, 114 ° 21’ восточной долготы).Почва на полевом участке имеет структуру суглинка с общим содержанием азота, фосфора и калия 820,0, 30,6 и 285,5 мг / кг -1 , соответственно. Метеорологическая информация получена со станции на экспериментальной площадке. Средние температуры с апреля по октябрь составляли 22,3 ° C в 2012 году и 21,6 ° C в 2013 году; продолжительность солнечного сияния составила 1092 ч в 2012 г. и 1157 ч в 2013 г .; активные накопленные температуры (≥15 ° C) составляли 4338 ° C в 2012 году и 3998 ° C в 2013 году; общее количество осадков составило 408 мм в 2012 году и 480 мм в 2013 году.
Лечение, экспериментальный дизайн и управление.
В этом исследовании однофакторный эксперимент с тремя методами посадки субстратом рассады пересаженного хлопка (SSTC), пересаженного хлопка из почвенного куба (ScSTC) и прямого посева хлопка (DSC), организованных в полностью рандомизированном блочном дизайне с тремя повторами для хлопка. . Для всех обработок использовали устойчивый к трансгенным насекомым Bt ( Bacillus thuringiensis ) хлопок ( Gossipium hirsutium L.) сорт CRI46 был посажен при плотности 45000 растений -1 га. Участок имел ширину 6,4 м и длину 8 м, состоял из 8 рядов хлопчатника с междурядьями 80 см. Чтобы повысить воспроизводимость наших результатов, лабораторные протоколы были доступны в Интернете по адресу: dx.doi.org/10.17504/protocols.io.js4cngw
Субстрат, используемый в системе субстрата для пересадки рассады, имеет национальный патентный сертификат и был предоставлен Институтом исследований хлопка Китайской академии сельскохозяйственных наук.Запатентованный субстрат содержит вермикулит, органические удобрения и минералы, безопасные для окружающей среды, человека и домашнего скота. Кроме того, наблюдались лучшие характеристики аэрации и водоудерживающая способность. В соответствии с местной агрономической практикой 29–30 марта в оба года по два семени хлопчатника были посеяны в каждую полость лотков, содержащих 100 полостей (60 см * 33 см * 5 см), и помещены в теплицы с минимальной температурой 6 ° C и самая высокая температура 20 ° C, что примерно на 5 ° C выше, чем температура наружного воздуха, чтобы обеспечить благоприятные условия для прорастания и роста проростков.Согласно плану экспериментов в оба года, саженцы были пересажены 30 апреля в ямы глубиной 10 см, вырытые с помощью ручной машины, изготовленной вручную.
В системе пересадки рассады почва-кубик, для выращивания рассады, столбчатые почвенные блоки (диаметром 4–6 см и высотой 8–12 см), состоящие из почвы и удобрений (N: 15%, P 2 O 5 : 15%, K 2 O: 15%) с формами соответствующего размера, были подготовлены в середине марта перед посадкой. Затем засеянные кубики почвы аккуратно поместили на грядки с рассадой (глубиной 10 см и шириной 2 м) в хижине, напоминающей оранжерею.По одному семени высевали в каждый блок сразу после полива 29–30 марта в оба года. Блоки с проростками пересаживали в ямки глубиной 10 см вручную на опытных участках 30 апреля согласно плану эксперимента.
Хлопок был посажен 17 апреля в системах обычного посева и прямого посева с пластиковым мульчированием. Семена хлопчатника были засыпаны влажной почвой во всех системах и мульчированы широкой пластиковой пленкой (толщиной 6 мкм и шириной 120 см), которая удерживалась на месте, закапывая края почвы в два ряда.После появления всходов в пленке делали отверстия, чтобы проростки могли прорасти. Все участки прореживали до желаемой густоты растений, оставляя наиболее сильнорослые растения на стадии 2 настоящих листьев.
В обоих опытах каждый год землю вспахивали и орошали ранней весной перед посевом. На всех участках было внесено 225 кг га -1 N, 150 кг га -1 P 2 O 5 и 225 кг га -1 K 2 O. Все удобрения были равномерно распределены по почве и внесены в верхние 20 см почвы перед посевом.Приблизительно 45 мм дополнительной воды (на полив) было добавлено путем затопления борозд и в соответствии с рекомендациями местных агрономов. Были использованы другие стратегии управления полями, основанные на местных агрономических практиках. Прежде чем использовать пластиковую мульчу и пестициды для борьбы с насекомыми и болезнями, боролись с сорняками вручную.
Сбор данных
Исследования корневого распределения.
Образцы корней собирали один раз перед каждым поливом (9 раз) в течение каждого сезона.Устройство для керна почвы (диаметром 7 см, высотой 10 см) [34, 39–40] использовалось для получения образцов, чтобы минимизировать горизонтальное и вертикальное повреждение корней. Для изучения горизонтального распределения корней корневые стержни (образцы почвы, содержащие корни) собирали в двух направлениях (восточном и западном) от области ствола (0 см) растений на расстоянии 20 и 40 см, в которых пленка мульчировалась. был применен в восточной части (E20 и E40), но без покрытия в западной части (W20 и W40) (рис. 1). Точка, наиболее удаленная к востоку от ствола (E40 см), была расположена в центре пленки, а точка, наиболее удаленная к западу от ствола (W40 см), была расположена в центре обнаженной почвы.Для изучения вертикального распределения корней шнек вставляли на глубину 120 см от поверхности почвы. Полученный керн почвы был разделен на 12 блоков размером 0–10 см, 10–20 см, 20–30 см, 30–40 см, 40–50 см, 50–60 см, 60–70 см, 70–80 см, 80–90 см, 90–100 см, 100–110 см и 110–120 см в обоих направлениях отбора проб, и почва была осторожно удалена с помощью ручных инструментов из каждого блока на глубину 10 см. Одновременно каждую сердцевину корня глубиной 10 см помещали в сито с круглой сеткой диаметром 0,05 см и промывали проточной водой для удаления частиц почвы с корней.Собранные образцы корней сканировали с помощью сканера (Phantom 9800X, MiCROTEK, Шанхай, Китай) и анализировали с помощью программного обеспечения DT-SCAN (Delta-T Devices Ltd, Великобритания) для определения длины корня (RL), среднего диаметра корня и поверхности корня. площадь (РА). Затем измеряли плотность длины корня (RLD, длина корня на объем почвы, мм / см 3 ) и плотность поверхности корня (RAD, площадь поверхности корня на объем почвы, мм 2 / см 3 ). определяется как характеристики пространственного распределения.
Рис. 1. Схематическое изображение мест отбора проб корней в 2012 и 2013 годах.
Примечание: открытая поверхность в левой части W20 см и W40 см, но мульчирование пленкой в правой части E20 см и E40 см, как показано ниже.
https://doi.org/10.1371/journal.pone.01.g001
Корневая биомасса.
Во время выборки были исключены не менее двух краевых рядов, чтобы избежать граничных эффектов. После измерения площади листьев корни отбирали в зависимости от стадии их развития каждые 15 дней с 15 мая по 17 сентября в 2012 и 2013 годах, затем сушили в печи при 80 ° C до постоянного веса, а затем взвешивали.
Доходность и ее компоненты.
Семенной хлопок (SC) на каждом участке собирали вручную три раза каждые десять дней с 15 октября 2012 г. и 10 октября 2013 г. и до 20 октября, чтобы получить предварительный заморозок семян хлопчатника (PFSC). Выход ворса (LY) и образование коробочек с трещинами определяли после очистки от хлопка. Компоненты урожая включали общее количество коробочек (NB), вес коробочки (BW) и процент ворса (LP: вес ворса / вес семян хлопка), очищенных с помощью лабораторного джина (MPSY-100A), которые были определены путем случайного сбора 50 открывать коробочки и взвешивать их после сушки при каждом сборе урожая.
Статистика
Вариации между местоположениями и годами выборки корней считались случайными, данные RLD и RAD за два года были объединены и интерполированы с использованием Кригинга [41] для определения двумерных распределений корней на разных этапах роста выборки. Тем временем с помощью Surfer (Golden Software Inc., США) была сделана контурная карта, чтобы точно описать развитие корней и распространение в результате различных методов посадки.На всех стадиях роста RAD и RLD (горизонтальный и вертикальный) усреднялись, чтобы отдельно наблюдать изменение корней в обоих направлениях. Чтобы смоделировать динамику RL и RA на хлопковых полях, мульчированных пленкой, и исследовать взаимосвязь между RL, RA и периодом роста хлопка (CGP), был проведен нелинейный регрессионный анализ (кубический полином) с использованием CGP в качестве независимой переменной. Модель логистической регрессии использовалась для сравнения RB и CGP для различных методов посадки.Дисперсионный анализ (ANOVA) проводили с использованием PROC GLM (версия 9.2, SAS Institute Ltd., США), при которой система трансплантации имела фиксированный эффект. Множественное тестирование Дункана с уровнем вероятности 0,05 использовалось для сравнения парных средних. Данные были проверены и преобразованы в соответствии с требованиями ANOVA (нормальное распределение, однородность дисперсии). Данные урожайности и компонентов урожайности были статистически проанализированы с использованием однофакторного дисперсионного анализа с уровнем значимости 0,05, с методом посева в качестве независимой переменной и с использованием теста множественного сравнения Дункана для разделения средних значений.Для печати использовались OriginPro 8 (OriginLab) и Adobe Illustrator CS5 (Adobe).
Результаты
Пространственное распределение корней хлопчатника
Горизонтальное распределение корневой системы хлопчатника.
Наблюдения за горизонтальным распределением корней на всех стадиях роста представлены на рис. 2. RAD и RLD изменились в горизонтальном направлении, перпендикулярном рядам растений в трех вариантах обработки. В целом, RAD и RLD уменьшались от стебля растения к западу и востоку, с более высокими RAD и RLD на востоке (E20 см и E40 см), чем на западе (W20 см и W40 см).Самые большие RAD и RLD в горизонтальном направлении были обнаружены на высоте 0 см (где были стержневые корни), что составляло 24,77 и 24,11% от общего числа корней, соответственно. Следующие по величине RAD и RLD наблюдались на E20 см для пленочного мульчирования со значениями приблизительно 23,76 и 23,58% соответственно. Кроме того, RAD и RLD 21,30 и 21,72%, 18,88 и 19,23%, а также 11,28 и 11,35% приходятся на W20 см (голая почва), E40 см (мульчированная пленка) и W40 см (голая почва) соответственно. Самые низкие значения при ширине 40 см были связаны с отсутствием пленочной мульчи, которая могла бы повысить температуру почвы и способствовать дальнейшему росту корней.
Рис. 2. Горизонтальные распределения RAD и RLD при посадочных системах DSC, ScSTC и SSTC.
Примечание: A и B представляют собой горизонтальные распределения RAD и RLD для различных обработок, соответственно.
https://doi.org/10.1371/journal.pone.01.g002
Вертикальное корневое распределение.
На всех стадиях роста наблюдаемое вертикальное распределение корней представлено на рис. 3. Хотя вертикальный образец распределения корней был однородным, наблюдались значительные различия между RAD и RLD SSTC, ScSTC и DSC в зависимости от глубины почвы.RAD и RLD в трех вариантах обработки уменьшались с глубиной почвы с 20 см до 120 см, и большинство RAD (0,68 мм 2 / см 3 ) и RLD (2,66 мм / см 3 ) наблюдались на глубине 20 см в SSTC. Кроме того, 70% корней были расположены на глубине 0–40 см, при этом самые высокие значения RAD (77,36%) и RLD (77,62%) приходились на DSC, за которыми следовали ScSTC (74,03% RAD и 76,25% RLD) и SSTC (70,75% RAD и 73,24% RLD) соответственно. Однако 13–20% корней образовались на глубине 40–80 см, при которой RAD (20.51%) и RLD (19,26%) были больше в SSTC, чем в ScSTC (20,61% RAD и 18,30% RLD) и DSC (13,82% RAD и 13,61% RLD). Тем не менее, менее 10% корней были расположены на глубине 80–120 см, со значениями RAD и RLD 8,82% и 8,77%, 8,74% и 7,50%, а также 5,36% и 5,45% для DSC, SSTC и ScSTC. , соответственно.
Рис. 3. Вертикальные распределения RAD и RLD при посадочных системах DSC, ScSTC и SSTC.
Примечание: A и B представляют собой горизонтальные распределения RAD и RLD для различных обработок, соответственно.
https://doi.org/10.1371/journal.pone.01.g003
Пространственное распределение плотности длины корней (RLD) хлопка, пересаженного из субстрата (SSTC) на стадии прорастания хлопка, стадии коробочки и стадии раскрытия коробочки.
RLD были наибольшими на глубине 0–30 см, когда стержневые корни интенсивно росли (рис. 4A), и были ниже на глубине 40–60 см через 46 дней после пересадки (стадия проростков). Кроме того, RLD был наибольшим на 0 см в горизонтальном направлении возле стержневых корней хлопка.Однако RLD был больше на E20 см, чем на W20 см из-за мульчирующей пленки.
Рис. 4. Пространственное распределение RLD при SSTC на стадии прорастания хлопчатника, стадии коробочки и стадии раскрытия коробочки.
Примечание: A, B и C представляют стадию всхода, стадию коробочки и стадию раскрытия коробочки, соответственно.
https://doi.org/10.1371/journal.pone.01.g004
Корни достигли максимального бокового распространения через 91 день после пересадки (стадия коробочки, Рис. 4B), и многие корни хлопчатника были продвинуты на глубину 0–50 см глубины.Однако немногие корни выросли на глубину 60 см. Поскольку рост боковых корней был сильным, RLD были максимальными на E20 см и W20 см, что указывало на то, что корневые системы были сконцентрированы в почве. Так как мульчирующая пленка повысила температуру почвы, RLD была значительно выше на востоке (E20 см и E40 см), чем на западе (W20 см и W40 см).
Подобные вариации RLD наблюдались через 125 дней после пересадки (стадия раскрытия коробочки, рис. 4C). Однако от стержневого корня плотность боковых корней резко снизилась.Кроме того, на западе было обнаружено меньше корней, чем на востоке, что объяснялось интенсивным ростом корней хлопчатника под пленкой из мульчи на стадии прорастания.
Пространственное распределение плотности длины корней (RLD) для пересаженного хлопчатника (ScSTC) на стадии проростков хлопчатника, стадии коробочки и стадии раскрытия коробочки.
Различия в RLD в ScSTC соответствовали таковым в SSTC. Максимальные RLD в основном были распределены в поверхностном слое (0–30 см) через 46 дней после пересадки (стадия проростков).Однако ниже 30 см количество корней резко уменьшалось. Тем не менее, корни проникли на глубину 60 см и значительно увеличились на глубине 30–50 см через 91 день после пересадки (стадия коробочки). До 125 дней после пересадки (стадия раскрытия коробочки) RLD был наибольшим на глубине 0–50 см, но был ниже, чем RLD на 91 день после пересадки (рис. 5).
Рис. 5. Пространственное распределение RLD под ScSTC на стадии прорастания хлопчатника, стадии коробочки и стадии раскрытия коробочки.
Примечание: A, B и C представляют стадию прорастания хлопчатника, стадию коробочки и стадию раскрытия коробочки, соответственно.
https://doi.org/10.1371/journal.pone.01.g005
Пространственное распределение хлопка прямого посева (DSC) на стадии прорастания хлопка, стадии коробочки и стадии раскрытия коробочки.
Аналогичная изменяющаяся тенденция RLD под DSC, SSTC и ScSTC на всех стадиях выращивания была постоянной; однако стержневые корни SSTC и ScSTC сохраняли обильный рост только 15 июня (стадия всходов) и 30 июля (стадия коробочки).Кроме того, стержневые корни хлопка в DSC достигли глубины до 100 см на стадии раскрытия коробочки (рис. 6). На стадии проростков большинство корней располагалось на глубине 0–30 см, а некоторые корни — на глубине 40–60 см. RLD был наибольшим на глубине 0–20 см в точке 0 см, что соответствовало SSTC и ScSTC.
Рис. 6. Пространственное распределение RLD при ДСК на стадии прорастания хлопчатника, стадии коробочки и стадии раскрытия коробочки.
Примечание: A, B и C представляют стадию прорастания хлопчатника, стадию коробочки и стадию раскрытия коробочки, соответственно.
https://doi.org/10.1371/journal.pone.01.g006
Вариации пространственной плотности длины корня (RLD) при различных обработках.
Пространственные распределения RLD в SSTC, ScSTC и DSC были разделены на шесть равных частей (1–1, 1–2, 2–1, 2–2, 3–1 и 3–2), которые находились в в вертикальном направлении снизу вверх и в горизонтальном направлении слева направо соответственно (рис. 4, 5 и 6). Средние значения RLD трех обработок на этапах выращивания хлопка в различных точках (1–1, 1–2, 2–1, 2–2, 3–1 и 3–2) показаны на рис. 7.Аналогичная тенденция пространственного распределения RLD была обнаружена для SSTC, ScSTC и DSC, что указывает на то, что самый высокий RLD имел место в части 1-2. Кроме того, более высокие RLD наблюдались в 1–2, чем в 1–1, по причине мульчирования пленки в 1–2, аналогично, RLD выше на 2–2 и 3–2 по сравнению с 2–1 и 3– 1 соответственно. В 1–1 и 1–2 RLD под SSTC и ScSTC были значительно выше, чем RLD под DSC; однако между SSTC и ScSTC не наблюдалось значительной разницы в RLD.RLD DSC были выше, чем у ScSTC и SSTC в 2–1, 2–2, 3–1 и 3–2, что указывает на то, что корни DSC проникают глубже, чем корни SSTC и ScSTC. Кроме того, RLD SSTC были выше, чем RLD ScSTC в 2–1, 2–2 и 3–2.
Динамика площади поверхности корня (RA) и длины корня (RL)
Кубические функции для распределений RA и RL в SSTC, ScSTC и DSC, полученные с помощью регрессионного анализа, показаны в таблице 1. Результаты моделирования RA и RL хорошо согласуются с результатами измерений, как показано на рисунке 8.Измеренные и смоделированные RA и RL непрерывно увеличивались и резко снижались, соответственно, в середине июля при всех обработках, а максимальные моделируемые RA и RL в SSTC наблюдались через 91–94 дня после пересадки (1–4 августа) и составляли 10370 мм. 2 завод -1 и 38486 мм завод -1 соответственно. В ScSTC максимальное RA (7794 мм 2 растение -1 ) и RL (31272 мм растение -1 ) произошло через 85–87 дней после пересадки (25–27 июля).Однако самое высокое RA (11102 мм, 2, , растение -1 ) и RL (46942 мм, растение -1 ) в DSC произошло примерно через 83 дня после посадки (15 июля).
Исследования корневой биомассы (RB)
В дополнение к общим RAD и RLD в различных обработках, RB также измеряли на различных стадиях роста хлопчатника (Таблица 2). Динамика продукции RB при всех обработках следовала типичной сигмовидной кривой с лаг-фазой, за которой следовали экспоненциальная фаза и стационарная фаза.Накопление RB значительно различалось между обработками из-за продолжительности экспоненциального накопления RB (Таблица 2). SSTC и DSC обычно имели более длительную продолжительность экспоненциального накопления, чем ScSTC (таблица 2). Однако существенной разницы в максимальной скорости экспоненциального накопления не наблюдалось. В ЮЮТК максимальная скорость накопления RB приходилась на 20–28 июня. Кроме того, значение RB было максимальным 4–9 июля и уменьшалось 19 июля, что указывало на то, что продолжительность экспоненциального накопления RB была увеличена примерно до 23–29 дней.В ScSTC максимальная скорость накопления RB приходилась на четыре дня, а в SSTC максимальная скорость накопления RB наблюдалась за 15–30 дней. Более того, в DSC максимальная скорость накопления RB приходилась на 13–20 июня, что на 6 дней раньше, чем в ScSTC.
Доходность и ее составляющие
SSTC привел к увеличению выхода SC на 4,0% по сравнению с ScSTC и DSC в 2012 г. (Таблица 3). Существенные различия в урожайности СК наблюдались в 2013 г., у SSTC — 8.Выходы SC на 0% и 5,2% выше, чем у ScSTC и DSC, соответственно. Хотя существенной разницы в LY не было, LY были на 1,7% и 1,1% больше в SSTC, чем в ScSTC и DSC в 2012 году, то же самое было обнаружено в 2013 году (на 7,2% и 2,3% больше). И никаких значительных различий в BW и LP между обработками в 2012 и 2013 годах не наблюдалось. Кроме того, PFSC в ScSTC был на 1,3% больше, чем в SSTC и DSC в 2012 году, и на 3,9% и 0,4% выше, чем PFSC в SSTC и DSC в 2013.
Обсуждение
Пространственное распределение корней хлопчатника
Корни любого растения играют ключевую роль в жизненном цикле растений [42].Хлопок имеет глубокую корневую систему, которая помогает поглощать воду и питательные вещества из более глубоких слоев почвы. Знание закономерностей распределения корней трудосберегающих методов пересадки рассады может быть использовано для управления питательными веществами и орошением при выращивании хлопка.
Влияние SSTC на развитие корней хлопчатника можно лучше понять, описав их пространственное распределение [30]. Максимальное горизонтальное распространение корней в SSTC составляло 0-20 см от хлопкового ряда. Такая картина в основном объясняется уменьшением испарения почвы и повышением температуры почвы из-за мульчирования полиэтиленовой пленкой после пересадки [3, 43].Кроме того, в соответствии с предыдущими исследованиями [34, 44–45], RAD и RLD во всех вариантах лечения были больше на E20 см, чем на W20 см в этом исследовании. Кроме того, количество корней хлопчатника под пленочной мульчей (E20 см и E40 см) увеличилось на 8–12% по сравнению с корнями голой почвы (W20 см и W40 см).
Различия RAD и RLD в вертикальном направлении были значительными на всех исследованных глубинах. Уменьшение RLD с глубиной можно объяснить тем, что рост корней начинается в верхнем слое почвы, таким образом, корни растут в верхнем слое почвы дольше, чем в более глубоких слоях почвы.Следовательно, RB в поверхностных слоях будет больше до того, как корни разрастутся на большую глубину [46]. В этом исследовании кривые RAD и RLD были типичными унимодальными сортами, при этом большинство корней сосредоточено на глубинах 20-40 глубин, как сообщалось ранее [42, 47-49]. Кроме того, максимальная глубина укоренения составляла 20–40 для всех обработок, а глубина укоренения в DSC была примерно на 10 см больше, чем в SSTC и ScSTC, возможно потому, что пересадка нарушает рост корней [16]. Однако в этом исследовании RAD и RLD значительно различались между SSTC и DSC как в горизонтальном, так и в вертикальном направлениях.
Развитие корней на стадиях роста хлопчатника
Физическое расположение корней сильно влияет на способность растений улавливать питательные вещества и водные ресурсы, следовательно, по нему можно определить, будет ли хлопок процветать или погибнуть. Следовательно, рост корня хлопчатника был инициирован на стадии проростков хлопчатника, за которым последовало интенсивное развитие на стадии хлопковой коробочки и более медленное развитие на стадии раскрытия хлопковой коробочки [10]. Предыдущие исследования показали, что RLD равны 0.2–0,3 см / см 3 подходят для выращивания хлопка [50]. В этом исследовании максимальное значение RLD на одно растение составляло 2,66 мм / см 3 на глубине 10–20 см. SSTC предоставил довольно высокий RLD, который удовлетворял потребности в выращивании хлопка. Более того, максимальная скорость роста корней в SSTC произошла через 91–94 дня после пересадки (1–4 августа), а максимальная скорость роста корней в ScSTC произошла через 83 дня после пересадки. Эти результаты показали, что раннее развитие корня происходит в ScSTC, что может объяснить более высокий PFSC в ScSTC, чем в SSTC.Однако, согласно рис. 8, RAD и RLD в ScSTC были ниже, чем RAD и RLD в SSTC после стадии возведения хлопка в квадрат. Более того, как RAD, так и RLD в SSTC были самыми высокими после стадии вскрытия хлопковых коробочек. В целом, сильное развитие корней в SSTC произошло на ключевых этапах выращивания хлопка, что имеет большое значение, хотя рост корней начался поздно.
Корневая биомасса (RB) и урожай
Точно так же наблюдалось раннее начало роста корней в ScSTC, и RB изменялся на разных стадиях роста.Конечный сухой вес корней хлопчатника в основном контролировался продолжительностью экспоненциального накопления RB [51]. Это исследование показало, что SC SSTC составлял 4,0% (2012 г.) и 9,0% (2013 г.). Кроме того, рост боковых корней определялся текущей массой корней или RLD и доступностью воды в почве на заданной глубине [46], что также могло объяснить большее количество боковых корней в SSTC (Рис. 4; Рис. 5). Большее количество боковых корней может быть результатом более рыхлой текстуры субстрата в SSTC, чем в ScSTC, потому что уплотненные почвы обычно создают более сильные механические ограничения, чем рыхлые почвы, и перенос кислорода и CO 2 к корням растений и от них, соответственно, уменьшается. в компактных почвах [52].Более того, в предыдущих исследованиях сообщалось, что корни без ветвей обычно имеют более короткий срок жизни, чем корни с ветвями [53].
Увеличение RL и RLD также улучшило закрепление корней растений [52]. Культурные практики, основанные на создании корневых систем, которые продлевают период цветения или ускоряют рост и развитие растений, например ранняя посадка хлопка, вероятно, увеличивают урожайность сельскохозяйственных культур [54]. Пересадку саженцев во избежание переохлаждения в начале сезона можно осуществить, используя хижины, похожие на теплицы.Более того, в предыдущих исследованиях сообщалось, что период цветения был продлен примерно на одну неделю, и что пиковое цветение произошло на 5 дней раньше у пересаженного хлопка, чем у DSC [3]. Несколько исследований показали, что повышение урожайности может быть связано с изменениями в архитектуре корня [55]. Кроме того, результаты этого исследования показали, что SSTC значительно увеличил урожайность SC в 2012 и 2013 годах. Таким образом, среда была подходящей для стадий роста хлопка, и высокие урожаи SC сохранялись в SSTC, что можно объяснить увеличением количества NB. и большая ширина полосы в SSTC, чем в ScSTC и DSC.
Выводы
Данные двух сезонов полевых экспериментов были использованы для исследования распределения RAD и RLD в результате различных методов пересадки рассады. По горизонтали RAD и RLD хлопка были наибольшими на 0 см и были больше ближе к растениям хлопка и под мульчирующей пленкой. По вертикали RAD и RLD в различных вариантах лечения уменьшались с глубиной и увеличивались со временем. SSTC привел к более высоким RAD и RLD в горизонтальном и вертикальном направлениях, за которыми следовали DSC и ScSTC.
Во-первых, самый сильный рост корней произошел во время цветения хлопчатника и стадии коробочки в SSTC, с ранним началом роста корней в ScSTC и сильным ростом корней на всех стадиях роста хлопка в DSC. Во-вторых, более длительное экспоненциальное увеличение накопления RB привело к более высокой урожайности SC. В-третьих, самый высокий уровень PFSC наблюдался в ScSTC в 2012 и 2013 годах, эти улучшения в основном были приписаны паттерну раннего начала роста корней в ScSTC.
Понимание динамики корней и изменений урожайности хлопка в результате различных методов посадки полезно для содействия использованию питомников для выращивания рассады хлопка для улучшения продуктивности растений и решения растущей проблемы нехватки рабочей силы.В этом исследовании был достигнут прогресс в недооценке преимуществ пересадки саженцев хлопка на урожайность и трудосбережение.
Благодарности
Это исследование было поддержано Индустрией национального общественного благосостояния (201203057–01) и Национальной программой поддержки ключевых технологий (2014BAD03B02-2). Мы высоко ценим работу техников экспериментальной станции Института исследования хлопка Китайской академии сельскохозяйственных наук.
Список литературы
- 1. ФАО ООН. FAOSTAT, Сельскохозяйственные культуры. 2013. Доступно в Интернете по адресу: http://www.fao.org/faostat/en/#data/QC
- 2. Дай Дж.Л., Донг Х.З. Технологии интенсивного выращивания хлопка в Китае: достижения, проблемы и меры противодействия. Полевые культуры Res. 2014; 155: 99–110.
- 3. Dong HZ, Li WJ, Tang W, Li ZH, Zhang DM. Повышение роста, развития растений и урожайность волокна трансгенного хлопка Bt за счет интеграции пластикового мульчирования и пересадки рассады.Ind. Crops Prod. 2007; 26: 298–306.
- 4. Кабал Б., Лароя Г. Технологическое развитие и производство хлопка в Индии и Китае. Curr. Sci. 2001; 80: 925–932.
- 5. Родриго В.Х., Стирлинг К.М., Теклехайманот З., Нугавела А. Пересечение бананов для улучшения фракционного улавливания и эффективности использования радиации незрелых каучуковых плантаций. Полевые культуры Res. 2001; 69: 237–249.
- 6. Цубо М., Уокер С. Модель перехвата и использования излучения межкорпусным пологом кукурузы и бобов.Agric. Для. Meteorol. 2002; 110: 203–215.
- 7. Авал М.А., Коши Х., Икеда Т. Улавливание и использование радиации в межкорпусных пологах кукурузы / арахиса. Agric. Для. Meteorol. 2006; 139: 74–83.
- 8. Zhang LZ, Werf WVD, Bastiaans L, Zhang S, Li B, Spiertz JHJ. Легкий перехват и использование в промежуточных посевах пшеницы и хлопка. Урожай поля. Res. 2008; 107: 29–42.
- 9. Смит CW, Варвил JJ. Двойной урожай хлопка и пшеницы. Агрон. J.1982; 74: 862–865.
- 10. Институт исследования хлопка Китайской академии сельскохозяйственных наук. Выращивание хлопка в Китае. 1 ст. изд. Шанхай: Шанхайская научно-техническая пресса; 2013. (на китайском языке)
- 11. Dong HZ, Li WJ, Tang W, Li ZH, Zhang DM. Повышение урожайности и доходов с помощью системы пересадки рассады для производства гибридных семян Bt-хлопка. J. Agron. Crop Sci. 2005; 191: 116–124.
- 12. Карве А.Д. Высокий урожай богарного хлопка за счет пересадки.Curr. Sci. 2003; 85: 122–123.
- 13. Шен ЦЙ. Изучение и борьба с болезнью хлопчатника. Пекин: Science Press; 1992 г.
- 14. Кояма А., Цуюдзаки С. Механизм воздействия со стороны осоковых и пушицовых кочанов при закладке рассады на заболоченном торфянике. Завод Ecol. 2012; 213: 1729–1737.
- 15. Чжан З., Ли Х., Ван Х., Нан Дж. Использование SNP для пересадки саженцев хлопка для увеличения роста и урожайности растений. Int. J. Plant Prod.2012; 6: 173–184.
- 16. Мао С.К., Ли Ю.К., Хань Ю.С., Ван Г.П., Ли Ю.Б., Ван XH. Предварительные наблюдения за морфологическими параметрами корневой системы хлопчатника корневищного пересадочного ( Gossypium hirsutum L.). Cotton Sci. 2008; 20: 76–78.
- 17. EI-Sahrigi AF, Kamel AS, EI-Khatib SI. Исследование по механизации пересадки хлопчатника. Египет J. Agric. Res. 2001; 79: 740–756.
- 18. Дун Х.З., Чжан Д.М., Тан В., Ли В.Дж., Ли Чж. Влияние системы посадки, густоты растений и удаления цветов на урожайность и качество гибридных семян хлопка.Полевые культуры Res. 2005; 93: 74–84.
- 19. Mao SC, Han YC, Wang GP, Li YB, Dong CW, Feng L и др. Расширить технику промышленного выращивания рассады и механизированной пересадки, ускорить развитие современного хлопководства. Китай Хлопок. 2012; 39: 1–5.
- 20. Гу С. Два типа миграции населения в Китае: сравнительное исследование. Подбородок. J. Popul. Sci. 1992; 4: 75–84. pmid: 12286127
- 21. Окадзаки Ю. Население Китая, рождаемость и урбанизация.Jinko Mondai Kenkyu. 1985; 1–18.
- 22. Акпо Э., Стомф Т.Дж., Коссу Д.К., Оморе А.О., Струик П.С. Влияние практик управления питомниками на морфологические признаки качества саженцев деревьев при посадке: пример масличной пальмы (Elaeis guineensis Jacq.). Для. Ecol. Управлять. 2014; 324: 28–36.
- 23. Caldeira MVW, Peroni L, Gomes DR, Delarmelina WM, Trazzi PA. Различные пропорции биотвердых веществ осадка сточных вод в составе субстратов для производства саженцев тимбо (Ateleia glazioveana Baill).Sci For. 2012; 40: 15–22.
- 24. da Luz PB, Paiva PDD, Tavares AR. Влияние внекорневых и субстратных удобрений на рост и развитие проростков женской пальмы. J. Plant Nutr. 2008; 31: 1313–1320.
- 25. Гарсия В.А., Модоло В.А., Лагоа АММА, Номура ЭС, Саес Лос-Анджелес. Характеристики остатков песка в качестве составного субстрата для выращивания саженцев персиковой пальмы (Bactris Gasipaes Kunth). Преподобный Арвор. 2011; 35: 595–604.
- 26. Санчес-дель-Кастильо F, Морено-Перес ED, Моралес-Маза A, Пена-Ломели A, Колинас-Леон MT.Плотность заселения и объем субстрата на проростках томатов (Lycopersicum licopersicon Mill.). Agrociencia-Mexico. 2012; 46: 255–266.
- 27. МакМайкл Б.Л., Остерхейс DM, Зак JC, Бейрути, Калифорния. Рост развития корневых систем. В: McD Stewart, Oosterhuis Heitholt и Mauney. Физиология хлопка. Springer; 2010.
- 28. Си Б., Ван И, Цзя Л. М., Блумберг М. Ли Г. Д., Ди Н. Характеристики тонкой корневой системы и водопоглощения триплоидной плантации Populus tomentosa на Северо-Китайской равнине: значение для управления поливной водой.Agric. Управление водными ресурсами. 2013; 117: 83–92.
- 29. Лян З.С., Ян Й., Шен К., Чжоу Дж., Ян Л. Ферментативная активность почвы и рост риса и ячменя под влиянием органических удобрений в антропогенной почве. Геодермия. 2003; 115: 149–160.
- 30. Ян Ч., Чай Ц., Хуанг ГБ. Распределение корней и реакция урожайности при пересадке пшеницы / кукурузы на чередующийся полив в засушливых районах северо-западного Китая. Plant Soil Environ. 2010; 56: 253–262.
- 31. Ахмади С.Х., Плауборг Ф., Андерсен М.Н., Сепасхах А.Р., Дженсен С.Р., Хансен С.Влияние стратегий орошения и почвы на картофель, выращиваемый в поле: распределение корней. Agric. Управление водными ресурсами. 2011; 98: 1280–1290.
- 32. Декстер А.Р. Физическое качество почвы. Часть I. Теория, влияние текстуры, плотности и органического вещества почвы, а также влияние на рост корней. Геодермия. 2004; 120: 201–214.
- 33. Адику С.Г.К., Озье-Лафонтен Х., Баязет Т. Модели роста корней и водопоглощения смеси кукурузы и вигны, выращенной в тепличных условиях. Почва растений.2001; 235: 85–94.
- 34. Нин С.Р., Ши Дж.К., Цзо Кью, Ван С., Бен-Гал А. Обобщение распределения плотности длины корней хлопка при капельном орошении с пленочной мульчированием. Полевые культуры Res. 2015; 177: 125–136.
- 35. Мин В., Го Х. Дж., Чжоу Г. В., Чжан В., Ма LJ, Е Дж и др. Распространение корней и рост хлопчатника при капельном орошении соленой водой. Исследования полевых культур. 2014; 169: 1–10.
- 36. Бешарат С., Наземи А.Х., Садраддини А.А.Параметрическое моделирование плотности длины корней и водопоглощения корней в ненасыщенной почве. Турок. J. Agric. Для. 2010; 34: 439–449.
- 37. Ню Дж.Ф., Пэн Ю.Ф., Ли СиДжей. Различие изменений длины корней на репродуктивной стадии у растений кукурузы, выращиваемых в поле и кварцевом песке. Питание растений и почвоведение. 2010; 173: 306–314.
- 38. Зубайди А., Макдональд Г. К., Холламби Г. Дж. Рост побегов, рост корней и урожай хлебной и твердой пшеницы в Южной Австралии.Aust. J. Exp. Agric. 1999; 39: 709–720.
- 39. Hou TY, Wang SH, Liang SX, Jiang WX, Luo DD, Huang DH. Эффективность скрининга сывороточного 1,3-бета-D-глюкана у пациентов с инвазивными грибковыми заболеваниями: метаанализ проспективных когортных исследований. Plos One. 2015; 10. doi: ARTN e0131602
- 40. Чжан В., Аханбике П., Ван Б.Дж., Сюй В.Л., Ли Л.Х., Кристи П. и др. Распределение корней и взаимодействие в системе агролесоводства мармелад / пшеница. Агрофор. Syst. 2013; 87: 929–939.
- 41. Оливер М.А., Вебстер Р. Кригинг: метод интерполяции для географических информационных систем. Int. J. Geog. Инф. Syst. 1990; 4: 313–332.
- 42. Сомкувар Р.Г., Тавар П.Б., Бондаж Д.Д., Навале С. Длина корня, масса корня и распределение сухого вещества в различных частях виноградной лозы Томпсон без косточек, привитой на разные подвои в тяжелой почве Махараштры. Турок. J. Agric. Для. 2012; 36: 543–552.
- 43. Чжоу Л.М., Ли FM, Цзинь С.Л., Сон Ю.Дж.Как два гребня и борозда, мульчированная пластиковой пленкой, влияют на влажность почвы, температуру почвы и урожай кукурузы на полузасушливом Лессовом плато в Китае. Полевые культуры Res. 2009; 113: 41–47.
- 44. МакМайкл Б.Л., Апчерч Д.Р., Берк Дж. Дж. Прогноз плотности корней хлопка на основе температуры почвы. Environ. Exp. Бот. 1996; 36: 303–312.
- 45. Zhao CY, Yan YY, Yimamu Y, Li JY, Zhao ZM, Wu LS. Влияние влажности почвы на плотность корней хлопчатника и урожайность при капельном орошении пластиковой мульчей на сельскохозяйственных угодьях оазиса Аксу.J. Засушливые земли. 2010; 2: 243–249.
- 46. Адику СГК, Брэддок Р.Д., Роуз CW. Моделирование влияния изменения влажности почвы на динамику роста корней однолетних культур. Почва растений. 1996; 185: 125–135.
- 47. Ло Х., Чжан Х., Хань Х., Ху Й., Чжан И., Чжан В. Влияние накопления воды в более глубоких слоях почвы на рост, урожайность и продуктивность хлопка (Gossypium Hirsutuml.) В засушливых районах северо-западного Китая. Ирриг. Осушать. 2014; 63: 59–70.
- 48. Морано Л., Кливер В. М..Распределение корней трехцветного подвоя виноградной лозы, привитого к Каберне Совиньон, выращенному на гравийно-глинистой почве в Оквилле, Калифорния. Являюсь. J. Enol. Витич. 1994; 45: 345–348.
- 49. Стивенс Р.М., Николас ПР. Длина корней и массовая плотность сортов Vitis-vinifera L Muscat Gordo Blanco и Shiraz (том 22, стр. 381, 1994). N. Z. J. Crop Hortic. Sci. 1995; 23: 237–237.
- 50. Kramer PJ. Водные отношения растений. Пекин: Science Press; 1989.
- 51.Тан Ф.Й., Сяо В.Дж. Накопление и разделение сухого вещества на различные фракции коробочек хлопка. Exp. Agric. 2013; 49: 543–555.
- 52. Анил С. Системы земледелия. J. Crop Prod. 2003; 8: 15–16.
- 53. Фусседер А. Продолжительность жизни и активность первичного корня кукурузы. Почва растений. 1987; 101: 257–265.
- 54. Петтигрю В.Т. Повышение урожайности благодаря системе производства хлопка раннего посева. Агрон. J. 2002; 94: 997–1003.
- 55. Ло ХХ, Тао XP, Ху YY, Чжан Ю.Л., Чжан В.Ф. Реакция роста корней хлопчатника и урожайность на ограничение корней при различных водных и азотных режимах.