
Клетка — СТРОЕНИЕ ТЕЛА ЖИВОТНЫХ
Тип урока: урок общеметодологической направленности.
Используемые технологии: здоровьесбережения, проблемного обучения, развивающего обучения, групповой деятельности, развития критического мышления, интерактивные.
Формируемые УУД: к. — строить речевые высказывания в устной форме; аргументировать свою точку зрения; р. — формулировать цель урока и ставить задачи, необходимые для ее достижения; работать по плану, сверять свои действия с целью и, при необходимости, исправлять ошибки самостоятельно; осуществлять рефлексию своей деятельности; п. — работать с различными источниками информации; строить логические рассуждения, включающие установление причинно-следственных связей; сравнивать и делать выводы; составлять план параграфа; работать с натуральными объектами; л. — формировать и развивать познавательный интерес к изучению биологии, экологическое мышление; выбирать целевые и смысловые установки в своих действиях и поступках по отношению к живой природе.
Планируемые результаты: объяснять значение понятий: клеточная мембрана, цитоплазма, вакуоль, обмен веществ, ядро, хромосомы, органоиды, клеточный центр; сравнивать клетки животных и растений; называть клеточные структуры животной клетки; делать выводы о причинах сходства и различий животной и растительной клеток; устанавливать взаимосвязь строения животной клетки с типом питания; описывать единство живой природы на основе знаний о клеточном строении организмов; характеризовать клетку как открытую биологическую систему.
Оборудование: учебник, магнитная или интерактивная доска, плакаты и таблицы “Растительная клетка”, “Животная клетка”, принадлежности для рисования.
Ход урока
I. Организационный момент
(Учитель приветствует учеников, проверяет готовность к уроку.)
II. Проверка домашнего задания
(Учитель собирает рабочие тетради для проверки.)
III. Работа по теме урока
1. Слово учителя
Сегодня мы рассмотрим строение животной клетки, а также проведем сравнительный анализ животной и растительной клеток.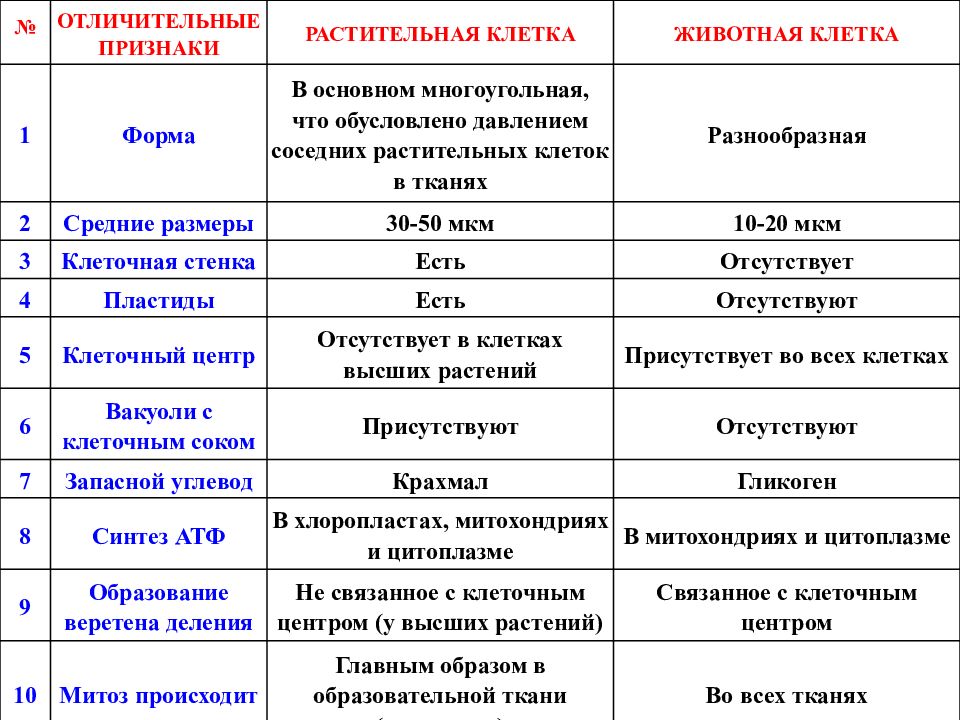 Напомню, что один из признаков живого — это клеточное строение. Животные клетки имеют свое специфическое строение, значит, отличие животных от остальных живых организмов начинается уже на клеточном уровне.
Напомню, что один из признаков живого — это клеточное строение. Животные клетки имеют свое специфическое строение, значит, отличие животных от остальных живых организмов начинается уже на клеточном уровне.
Большинство клеток животных очень маленькие, поэтому изучать их нужно при большом увеличении микроскопа. Формы клеток животных очень разнообразны: они бывают сильно вытянуты в длину или высоту, могут быть веретеновидными или бокаловидными. Размер и форма клеток зависят от того, какую функцию они выполняют в организме.
2. Строение животной клетки
Прошу открыть альбом и нарисовать большую — во весь лист — клетку. Я буду рассказывать о частях клетки, а вы будете их зарисовывать.
(Вариант первый. Учитель рисует на доске строение животной клетки или показывает части клетки на плакате (таблице), а ученики повторяют рисунок в тетради.
Вариант второй. Учитель рассказывает о строении клетки, а ученики изображают в тетради части клетки ассоциативными фигурами разных цветов по своему выбору и записывают основные функции.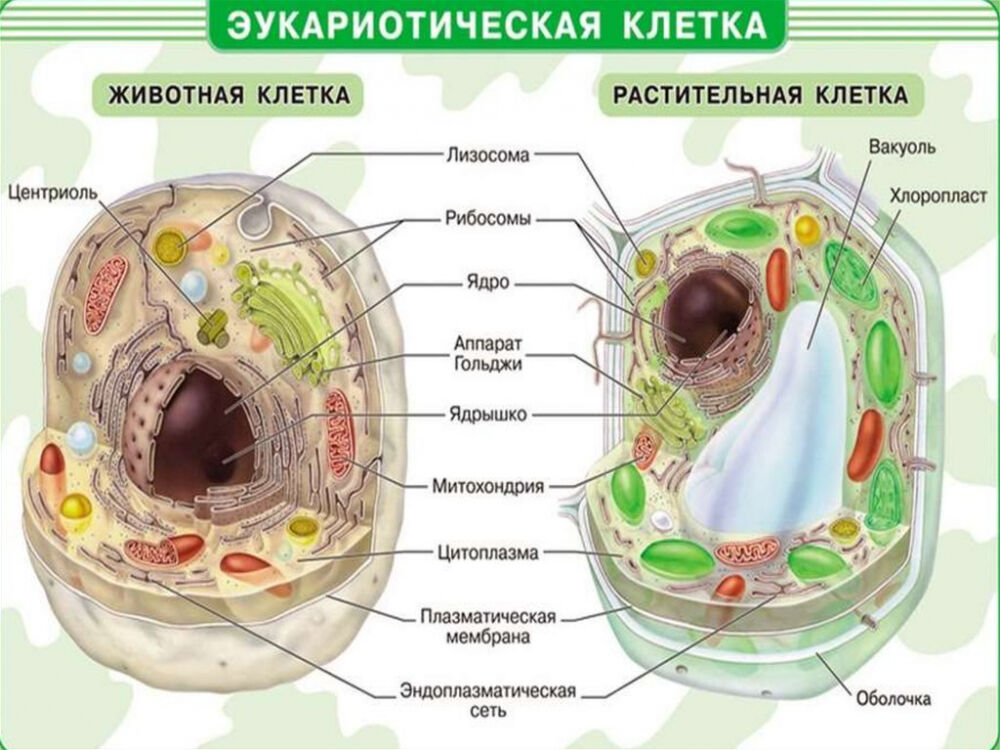 )
)
Каждая клетка снаружи покрыта тонкой и эластичной плазматической мембраной (плазмалеммой). Она отделяет содержимое клетки от внешней среды, а также имеет избирательную проницаемость — пропускает внутрь клетки одни вещества, а другие, наоборот, задерживает. Тем самым обеспечивается обмен веществ между клеткой и окружающей средой.
(Ученики рисуют мембрану простым карандашом и по линии мембраны пишут: “Мембрана отделяет, обмен веществ”.)
Под мембраной находится вязкое, полужидкое содержимое клетки — цитоплазма. В цитоплазме протекают все жизненные процессы клетки, в ней постоянно перемещаются различные вещества и компоненты клетки (органоиды), выполняющие разнообразные функции.
(Цитоплазму на рисунке можно немного закрасить, слегка нажимая на карандаш).
Периодически в цитоплазме образуются пузырьки, наполненные жидкостью, — вакуоли. Они играют важную роль в обмене веществ: в них накапливаются питательные вещества, с их помощью происходит удаление ненужных или вредных веществ из клетки.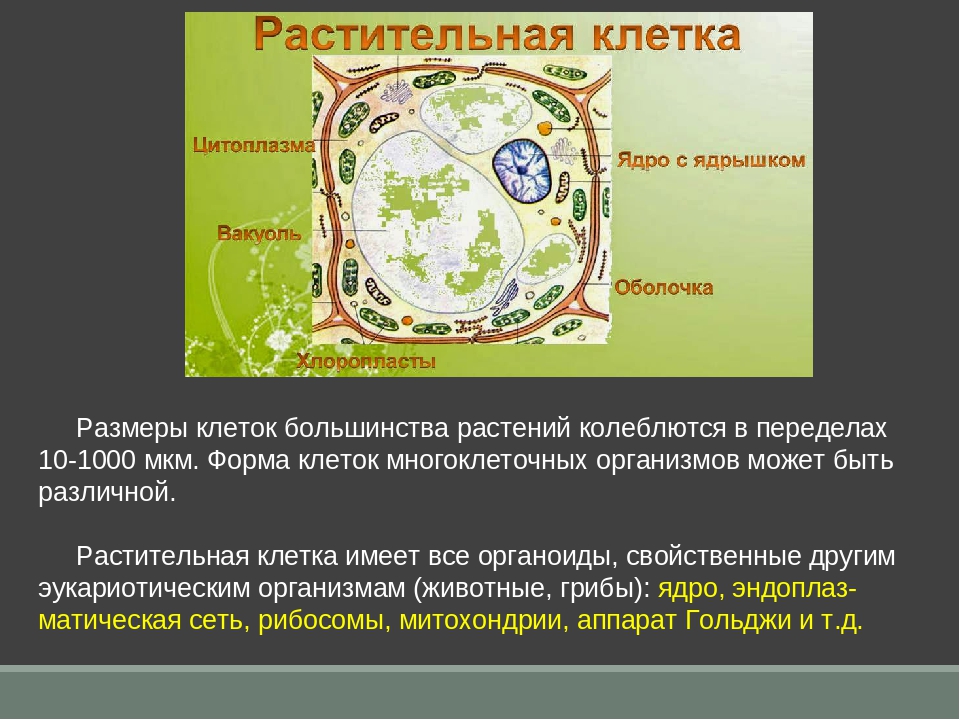 В отличие от клеток растений, где вакуоли занимали почти все пространство внутри клеточной стенки, в клетках животных вакуоли не достигают столь крупных размеров.
В отличие от клеток растений, где вакуоли занимали почти все пространство внутри клеточной стенки, в клетках животных вакуоли не достигают столь крупных размеров.
В цитоплазме хорошо заметно округлое образование — ядро. С помощью ядра регулируются все процессы, происходящие в клетке. Кроме того, ядро принимает самое активное участие в процессе деления клетки.
За обеспечение клетки энергией отвечают продолговатые органоиды — митохондрии. Они есть в клетках и растений, и животных, однако в клетках животных их гораздо больше. Во-первых, животным требуется значительно больше энергии, а во-вторых, в растительной клетке роль митохондрий могут выполнять хлоропласта.
Эндоплазматическая сеть служит для перемещения различных веществ по цитоплазме. Она состоит из полостей и канальцев, образованных мембраной, похожей по строению на плазматическую. По эндоплазматической сети вещества перемещаются значительно быстрее, чем по цитоплазме.
Рибосомы — мелкие органоиды, видные лишь в сильный микроскоп.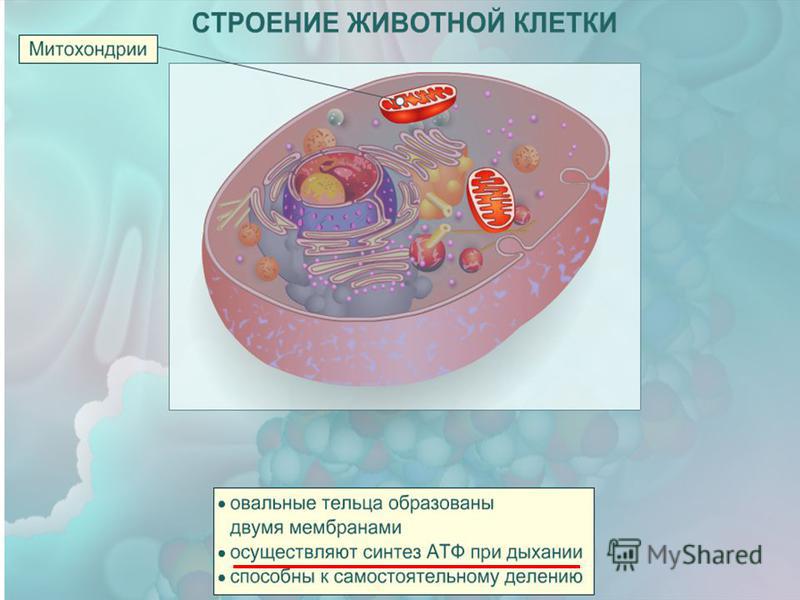 Их очень много как в цитоплазме, так и на эндоплазматической сети. Рибосомы отвечают за сборку молекул белков — одних из наиболее важных органических соединений. Рибосомы есть абсолютно во всех живых клетках.
Их очень много как в цитоплазме, так и на эндоплазматической сети. Рибосомы отвечают за сборку молекул белков — одних из наиболее важных органических соединений. Рибосомы есть абсолютно во всех живых клетках.
Аппарат (комплекс) Гольджи нужен для накопления и хранения разнообразных веществ. Он состоит из нескольких мембранных полостей, в которых накапливаются вещества, при необходимости от них отделяются пузырьки, содержащие те или иные вещества. Пузырьки направляются в цитоплазму для дальнейшего использования различных веществ. Кроме того, в аппарате Гольджи формируются лизосомы — небольшие, округлой формы органоиды с толстой оболочкой, наполненные пищеварительными соками. Лизосомы служат для клеточного пищеварения. В некоторых случаях оболочка лизосом может распадаться, в результате происходит самопереваривание клетки. Это встречается, например, у лягушек, когда у головастиков “пропадает” хвост. На самом деле в клетках хвоста, начиная с кончика, разрушаются лизосомы, и хвост “самопереваривается”.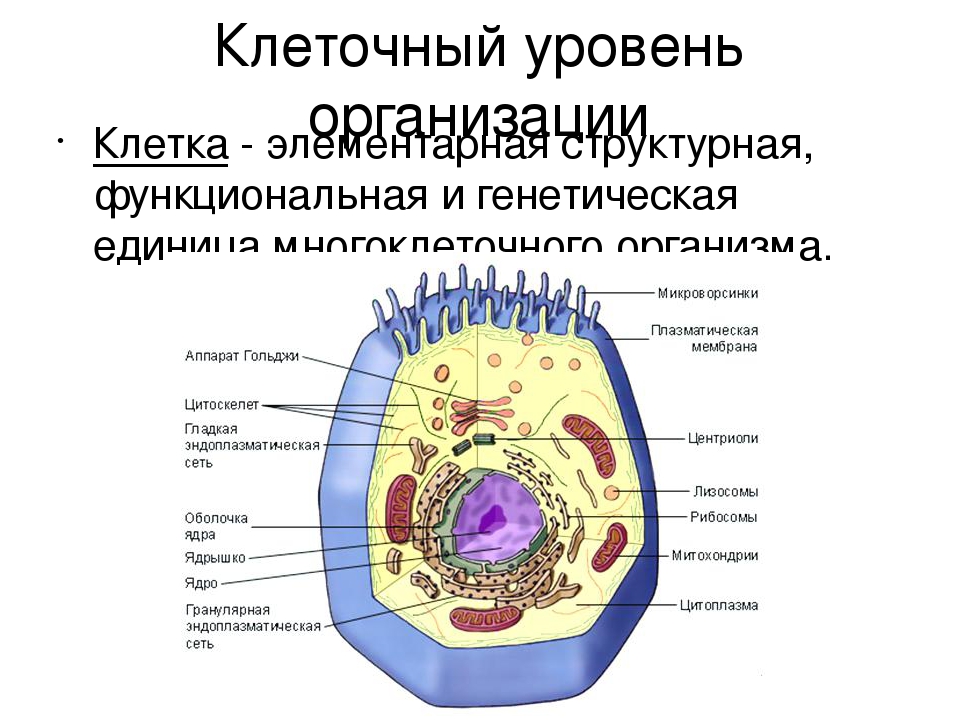
Хлоропластов и других пластид в клетках животных нет. Это одно из основных отличий растительной и животной клеток. Животные питаются гетеротрофно, т. е. получают питательные вещества из внешней среды.
В клетках животных имеются органоиды, которых нет в растительных клетках, — центриоли — два взаимно перпендикулярных цилиндрических тельца. Они составляют клеточный центр, который играет важную роль в процессе деления клетки, обеспечивая равномерное распределение наследственного материала.
Это нужно для того, чтобы дочерняя клетка ничем не отличалась от материнской.
Помимо органоидов, в цитоплазме клетки имеются так называемые включения. Это могут быть капельки жира, кристаллы или зерна. В виде включений в клетке запасаются необходимые вещества.
IV. Рефлексивно-оценочный этап
(Сидящие за одной партой ученики с помощью своих рисунков рассказывают друг другу о строении животной клетки. Учитель слушает беседу учеников, при необходимости поправляя и дополняя.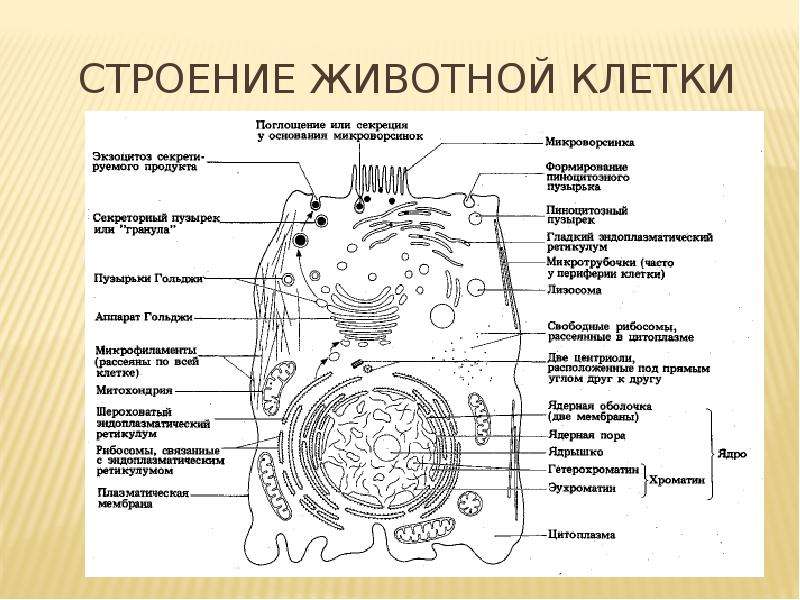
Домашнее задание
1. Прочитать § 6, повторить основные термины.
2. Подготовить сообщение о строении клетки по рисунку и записям в тетради.
Дополнительный материал
А вы знаете, что…
Митохондрии были впервые “приручены” примитивными животными примерно 1.2 млрд лет назад, а сине-зеленые бактерии превратились в первые хлоропласты водорослей 900 млн лет назад.
Размеры одноклеточных животных колеблются в среднем от 5 до 150 мк. Наиболее мелкие среди них — внутриклеточные паразиты (например, жгутиконосцы), а самые крупные представители — колониальные радиолярии — могут достигать 25 см.
Больше всего воды содержат клетки арбуза и огурца (92,1 %), меньше всего — клетки арахиса (5,2%), зубной эмали (3%).
Ученые открыли около 118 химических элементов. 15 составе клетки обнаружено 80 химических элементов; по содержанию их разделяют на три группы: макроэлементы, микроэлементы, ультрамикроэлементы.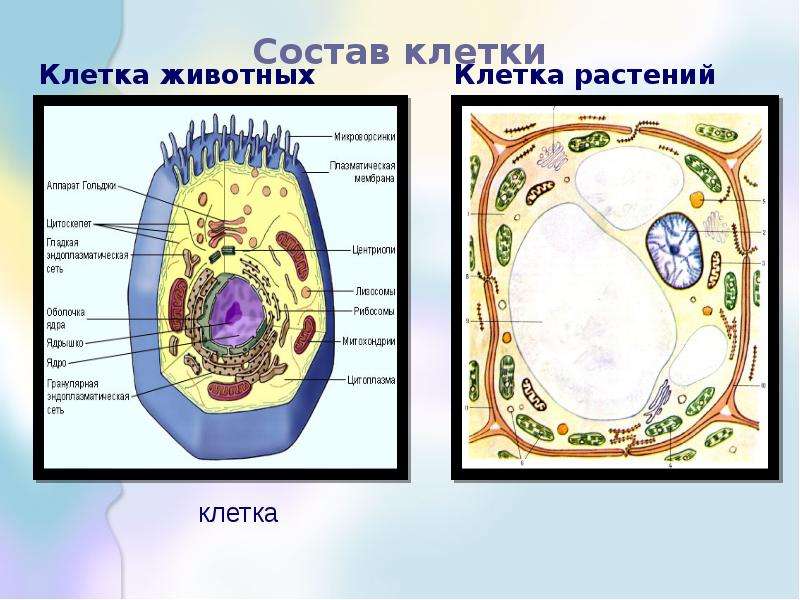
Число клеток в растении достигает астрономических величин (например, лист дерева насчитывает более 100 млн клеток). Человек состоит более чем из 100 трлн клеток.
На долю макроэлементов приходится 99% всей массы клетки. Микроэлементы (бор, кобальт, медь, молибден, цинк, ванадий, йод, бром и др.) составляют от 0.001% до 0,000001% массы клетки. На ультрамикроэлементы (уран, радий, золото, ртуть, бериллий, цезий и др.) приходится 0,000001% массы клетки.
Вода — уникальное вещество, составляющее до 99,7% массы живых организмов. Большинство реакций, протекающих в клетке, могут идти только в водном растворе. Роль воды в клетке определяется ее химическими и структурными свойствами: малыми размерами молекул, их полярностью и способностью соединяться друг с другом водородными связями. Молекулы воды проходят через мембрану клетки в 10 тыс. раз быстрее ионов калия и хлора.
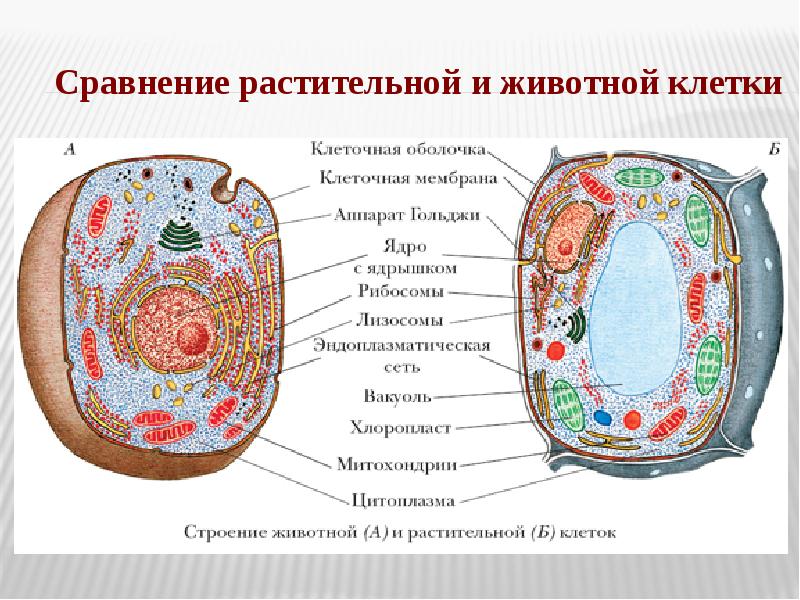
Строение животной и растительной клетки
Клетки животных и растений, как многоклеточных, так и одноклеточных, в принципе сходны по своему строению. Различия в деталях строения клеток связаны с их функциональной специализацией.
Различия в деталях строения клеток связаны с их функциональной специализацией.
Строение животной (слева) и растительной (справа) клеток
Основными элементами всех клеток являются ядро и цитоплазма. Ядро имеет сложное строение, изменяющееся на разных фазах клеточного деления, или цикла. Ядро неделящейся клетки занимает приблизительно 10—20% ее общего объема. Оно состоит из кариоплазмы (нуклеоплазмы), одного или нескольких ядрышек (нуклеол) и ядерной оболочки. Кариоплазма представляет собой ядерный сок, или кариолимфу, в которой находятся нити хроматина, образующие хромосомы.
Обязательными элементами ядра являются хромосомы, имеющие специфическую химическую и морфологическую структуру. Они принимают активное участие в обмене веществ в клетке и имеют прямое отношение к наследственной передаче свойств от одного поколения к другому.
Цитоплазма клетки обнаруживает весьма сложное строение. Введение методики тонких срезов и электронной микроскопии позволило увидеть тонкую структуру основной цитоплазмы.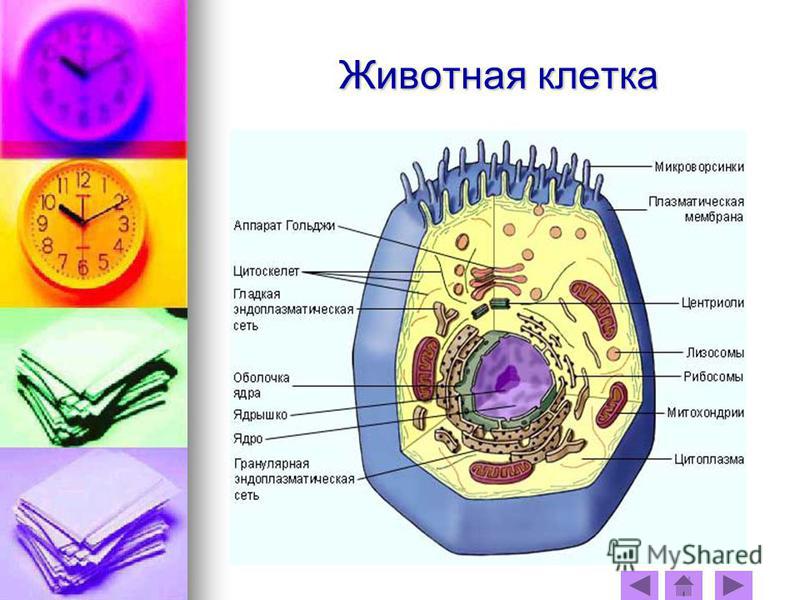
Строение клетки по данным электронной микроскопии
Установлено, что последняя состоит из параллельно расположенных сложных структур, имеющих вид пластинок и канальцев, на поверхности которых располагаются мельчайшие гранулы диаметром 100—120 Å. Эти образования названы эндоплазматическим комплексом. В состав этого комплекса включены различные дифференцированные органоиды: митохондрии, рибосомы, аппарат Гольджи, в клетках животных и низших растений — центросома, животных — лизосомы, у растений — пластиды. Кроме того, цитоплазме обнаруживается целый ряд включений, принимающих участие в обмене веществ клетки: крахмал, капельки жира, кристаллы мочевины и т. д.
Центриоли (клеточный центр) состоит из двух компонентов: триоли и центросферы — особым образом дифференцированного участка цитоплазмы. Центриоли состоят из двух мелких округлых колец. В электронном микроскопе видно, что эти тельца представляют собой систему строго ориентированных трубочек.
Митохондрии в клетках бывают разной формы: палочковидные, нулообразные и др. Полагают, что форма их может изменяться зависимости от функционального состояния клетки. Размеры митохондрии варьируют в значительных пределах: от 0,2 до 2—7 мк. клетках разных тканей они располагаются или равномерно по цитоплазме, или с большей концентрацией в определенных участках. Установлено, что митохондрии принимают участие в окислительных процессах обмена веществ клетки. Митохондрии состоят белков, липидов и нуклеиновых кислот. В них найден ряд ферментов, участвующих в аэробном окислении, а также связанных реакцией фосфорилирования. Полагают, что в митохондриях происходят все реакции цикла Кребса: большая часть освобождаются при этом энергии расходуется на работу клетки.
Полагают, что форма их может изменяться зависимости от функционального состояния клетки. Размеры митохондрии варьируют в значительных пределах: от 0,2 до 2—7 мк. клетках разных тканей они располагаются или равномерно по цитоплазме, или с большей концентрацией в определенных участках. Установлено, что митохондрии принимают участие в окислительных процессах обмена веществ клетки. Митохондрии состоят белков, липидов и нуклеиновых кислот. В них найден ряд ферментов, участвующих в аэробном окислении, а также связанных реакцией фосфорилирования. Полагают, что в митохондриях происходят все реакции цикла Кребса: большая часть освобождаются при этом энергии расходуется на работу клетки.
Строение митохондрий оказалось сложным. Поданным электрон-микроскопических исследований, они представляют собой тельца, суженные гидрофильным золем заключенные в избирательно проницаемую оболочку — мембрану, толщина которой около 80 Å. Митохондрии имеют слоистую структуру в виде системы утренних гребней-кристаллов, толщина которых 180—200 Å. Они отходят от внутренней поверхности мембран, образуя кольцобразные диафрагмы. Предполагается, что митохондрии размножаются путем деления. При делении клетки распределение их по крайним клеткам не подчиняется строгой закономерности, так как % по-видимому, могут быстро размножаться до необходимого клетки количества. По форме, величине и роли в биохимических процессах митохондрии являются характерными для каждого типа ни и вида организма.
Они отходят от внутренней поверхности мембран, образуя кольцобразные диафрагмы. Предполагается, что митохондрии размножаются путем деления. При делении клетки распределение их по крайним клеткам не подчиняется строгой закономерности, так как % по-видимому, могут быстро размножаться до необходимого клетки количества. По форме, величине и роли в биохимических процессах митохондрии являются характерными для каждого типа ни и вида организма.
При биохимических исследованиях цитоплазмы в ней найдены микросомы, которые представляют собой фрагменты мембран с структурой эндоплазматической сети.
В значительном количестве в цитоплазме находятся рибосомы размерам они варьируют от 150 до 350 Å и в световом микроскопе невидимы. Особенностью их является высокое содержание РНК и белков: около 50% всей клеточной РНК находится в рибосомах, что указывает на большое значение последних в деятельности клетки. Установлено, что рибосомы участвуют в синтезе клеточных белков под контролем ядра. Репродукция самих рибосом также контролируется ядром; в отсутствии ядра они теряют способность синтезировать цитоплазматические белки и исчезают.
В цитоплазме имеется также аппарат Гольджи. Он представляет систему гладких мембран и канальцев, располагающихся вокруг ядра или полярно. Предполагают, что этот аппарат обеспечивает выделительную функцию клетки. Тонкое строение его остается еще не выясненным.
Органоидами цитоплазмы являются также лизосомы — литические тела, выполняющие функцию пищеварения внутри клетки. Они открыты пока только в животных клетках. Лизосомы содержат активный сок — ряд ферментов, способных расщеплять белки, нуклеиновые кислоты и полисахариды, поступающие в клетку. В случае если мембрана лизосомы разрывается и ферменты переходят в цитоплазму, то они «переваривают» другие элементы, цитоплазмы и приводят к растворению клетки — «самопоеданию».
Для цитоплазмы растительных клеток характерно присутствие пластид, которые осуществляют фотосинтез, синтез крахмала и пигментов, а также белков, липидов и нуклеиновых кислот. По окраске и выполняемой функции пластиды могут быть разделены на три группы: лейкопласты, хлоропласты и хромопласты. Лейкопласты — бесцветные пластиды, участвующие в синтезе крахмала из сахаров. Хлоропласты представляют белковые тела более плотной консистенции, чем цитоплазма; наряду с белками они содержат много липидов. Белковое тело (строма) хлоропластов несет пигменты, в основном — хлорофилл, чем и объясняется их зеленая окраска, хлоропласты осуществляют фотосинтез. Хромопласты содержат пигменты — каротиноиды (каротин и ксантофилл).
Лейкопласты — бесцветные пластиды, участвующие в синтезе крахмала из сахаров. Хлоропласты представляют белковые тела более плотной консистенции, чем цитоплазма; наряду с белками они содержат много липидов. Белковое тело (строма) хлоропластов несет пигменты, в основном — хлорофилл, чем и объясняется их зеленая окраска, хлоропласты осуществляют фотосинтез. Хромопласты содержат пигменты — каротиноиды (каротин и ксантофилл).
Пластиды размножаются путем прямого деления и, по-видимому, не возникают в клетке заново. До сих пор нам не известен принцип их распределения по дочерним клеткам при делении. Возможно, что строгого механизма, обеспечивающего равное распределение, не существует, так как необходимое число их может быстро восстанавливаться. При бесполом и половом размножении растений через материнскую цитоплазму могут наследоваться признаки, определяемые свойствами пластид.
Здесь мы не будем останавливаться на особенностях изменений отдельных элементов клетки в связи с выполняемыми ими физиологическими функциями, так как это входит в область изучения цитологии, цитохимии, цитофизики и цитофизиологии. Однако следует отметить, что в последнее время исследователи приходят к очень важному выводу в отношении химической характеристики органелл цитоплазмы: ряд из них, такие как митохондрии, пластиды и даже центриоли, имеет собственную ДНК. Какова роль ДНК и каково состояние, в котором она находится, остается пока неясным.
Однако следует отметить, что в последнее время исследователи приходят к очень важному выводу в отношении химической характеристики органелл цитоплазмы: ряд из них, такие как митохондрии, пластиды и даже центриоли, имеет собственную ДНК. Какова роль ДНК и каково состояние, в котором она находится, остается пока неясным.
Мы познакомились с общей структурой клетки лишь для того, чтобы в последующем оценить роль отдельных ее элементов в обеспечении материальной преемственности между поколениями, т. е. в наследственности, ибо все структурные элементы клетки принимают участие в ее сохранении. Следует, однако, иметь в виду, что, хотя наследственность и обеспечивается всей клеткой как единой системой, ядерные структуры, а именно хромосомы, занимают при этом особое место. Хромосомы, в отличие от органелл клетки, представляют собой уникальные структуры, характеризующиеся постоянством качественного и количественного состава. Они не могут взаимозаменять друг друга. Несбалансированность хромосомного набора клетки приводит в конечном счете к ее гибели.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
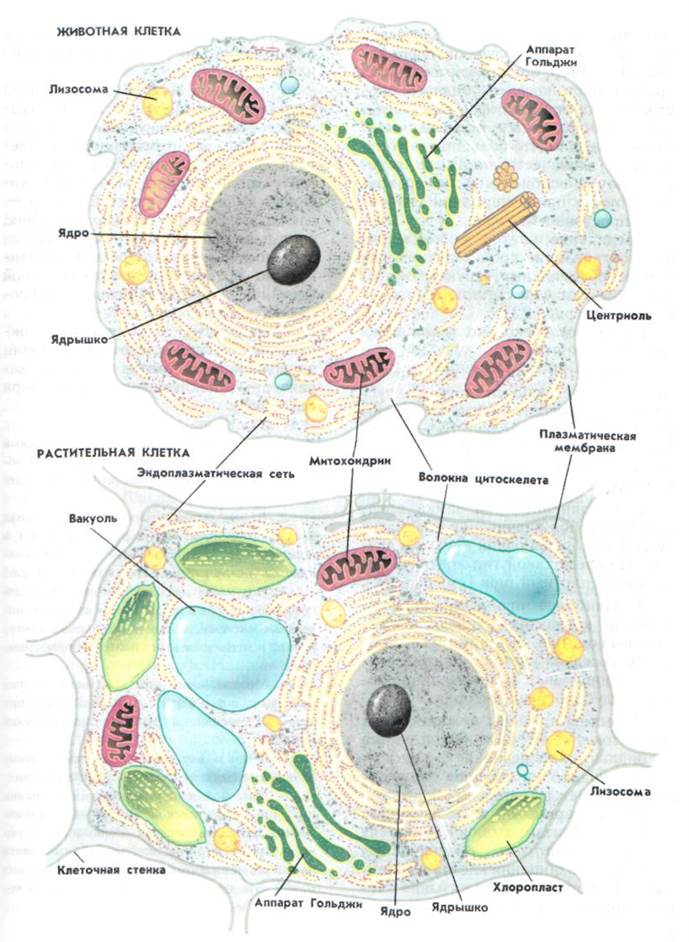
Различия в строении растительной и животной клетки
1. Различия в строении растительной и животной клетки Группа:Б(о)-15к Тағайбек Сабира Қапбар Жұлдыз Әмір Ләззат Батыр Арайлым
2. КЛЕТКА
Клетка – элементарная единица живой системы. Специфические функции в клетке распределены междуорганоидами – внутриклеточными структурами. Несмотря на многообразие форм, клетки разных типов
обладают поразительным сходством в своих главных структурных особенностях.
Клетка представляет собой элементарную живую систему, состоящую из трех основных структурных
элементов – оболочки, цитоплазмы и ядра. Цитоплазма и ядро образуют протоплазму.
Практически все ткани многоклеточных организмов состоят из клеток. С другой стороны, слизевики состоят
из неразделённой перегородками клеточной массы со множеством ядер.
3. Клетка
Мелкие организмы могут состоять всего лишь из сотен клеток.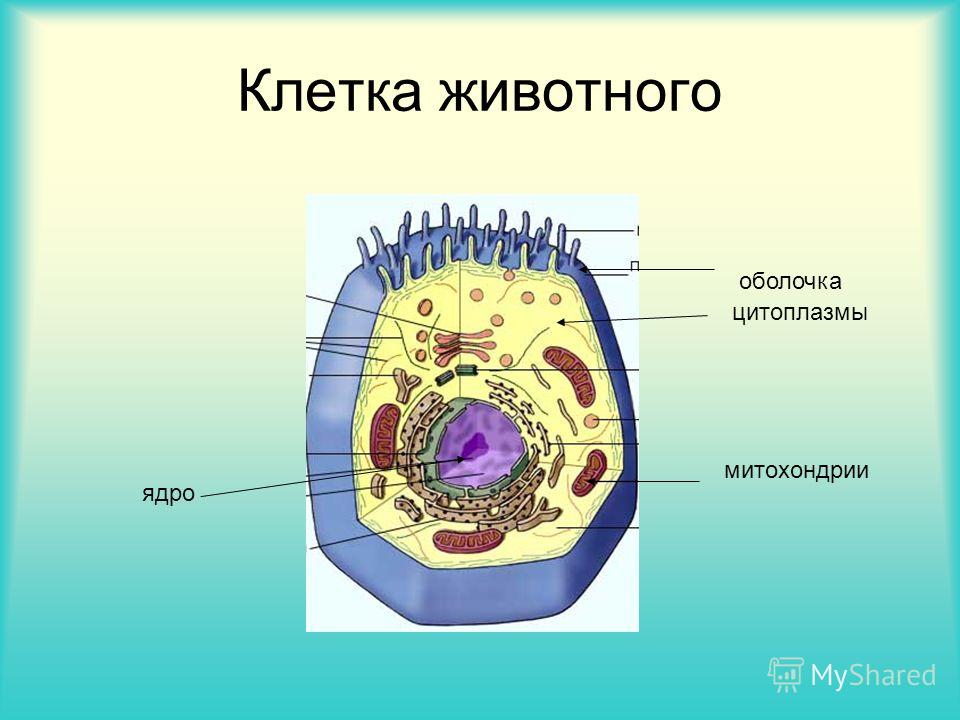 Организм человека
Организм человекавключает в себя 1014 клеток. Самая маленькая из известных сейчас клеток имеет размер
0,2 мкм, самая большая – неоплодотворенное яйцо эпиорниса– весит около 3,5 кг.
Слева истреблённый
несколько веков назад
эпиорнис.
Справа – его яйцо, найденное
на Мадагаскаре
Типичные размеры растительных и животных клеток составляют от 5 до 20 мкм. При этом
между размерами организмов и размерами их клеток прямой зависимости обычно нет.
Для того, чтобы поддерживать в себе необходимую концентрацию веществ, клетка должна
быть физически отделена от своего окружения. Вместе с тем, жизнедеятельность
организма предполагает интенсивный обмен веществ между клетками. Роль барьера
между клетками играет плазматическая мембрана. Внутреннее строение клетки долгое
время было загадкой для ученых; считалось, что мембрана ограничивает протоплазму –
некую жидкость, в которой и происходят все биохимические процессы. Благодаря
электронной микроскопии тайну протоплазмы удалось раскрыть, и сейчас известно, что
внутри клетки имеются цитоплазма, в которой присутствуют различные органоиды, и
генетический материал в виде ДНК, собранный, в основном, в ядре (у эукариот).
4. Строение растительной клетки.
Есть пластиды;Автотрофный тип питания;
Синтез АТФ происходит в
хлоропластах и митохондриях;
Имеется целлюлозная
клеточная стенка;
Крупные вакуоли;
Клеточный центр только у
низших.
5. Строение животной клетки
Пластиды отсутствуют;Гетеротрофный тип питания;
Синтез АТФ происходит в
митохондриях;
Целлюлозная клеточная стенка
отсутствует;
Вакуоли мелкие;
Клеточный центр есть у всех
клеток.
6. Различия в строении растительной и животной клетки.
Растительная клеткаЕсть пластиды;
Автотрофный тип
питания;
Синтез АТФ происходит
в хлоропластах и
митохондриях;
Имеется целлюлозная
клеточная стенка;
Крупные вакуоли;
Клеточный центр только
у низших.
Животная клетка
Пластиды отсутствуют;
Гетеротрофный тип
питания;
Синтез АТФ происходит
в митохондриях;
Целлюлозная клеточная
стенка отсутствует;
Вакуоли мелкие;
Клеточный центр есть у
всех клеток.

7. Общие черты, характерные для животной и растительной клеток
Принципиальное единство строения(поверхностный аппарат клетки,
цитоплазма, ядро.)
Сходство в протекании многих химических
процессов в цитоплазме и ядре.
Единство принципа передачи
наследственной информации при делении
клетки.
Сходное строение мембран.
Единство химического состава.
8. Отличительные признаки растительной и животной клетки
9. ВЫВОД:
1.2.
Принципиальное сходство строения и
химического состава клеток растений и
животных указывает на общность их
происхождения, вероятно от одноклеточных
водных организмов.
Животные и растения далеко отошли друг
от друга в процессе эволюции у них разные
типы питания, различные способы защиты
от неблагоприятных воздействий внешней
среды. Все это отразилось на строении их
клеток.
/home/sites/rulefo/public/documents/main/4klass/5energia/3den.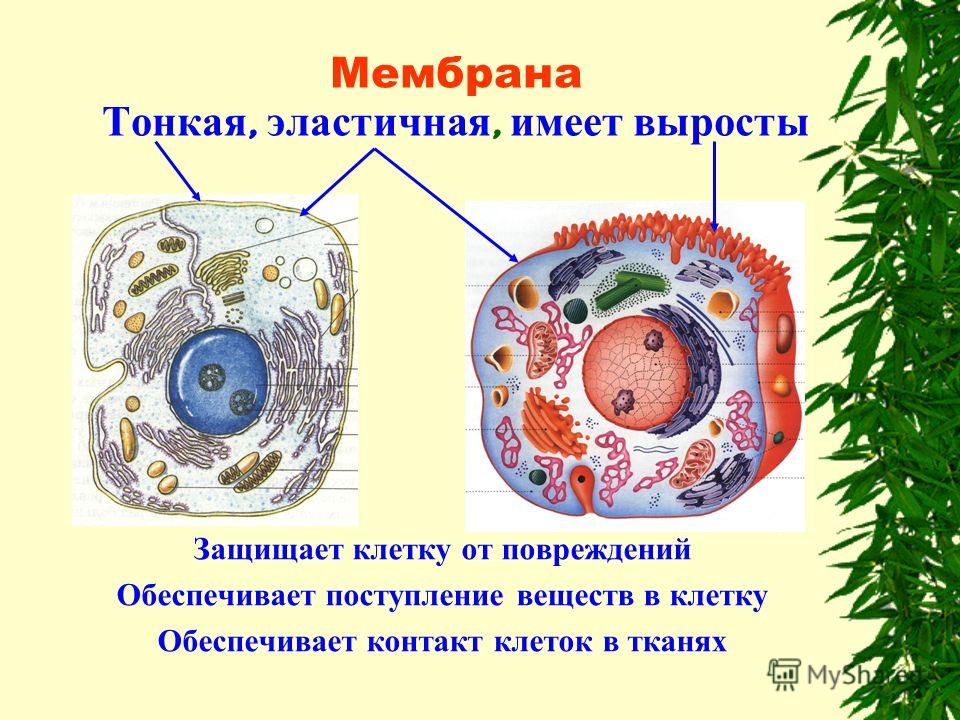 htm
htm
2. Строение клетки.
Позднее, когда микроскопы стали лучше, давали большее увеличение, ученые увидели, что внутри клеток не пустота, а можно увидеть много разных частей.
Центральной частью клетки было ядро, которое под микроскопом выглялит как круглое пятно. Вокруг ядра находится студенистая цитоплазма.
При помощи электронного микроскопа, который дает значительно большее увеличение, были увидены части цитоплазмы — органоиды. Каждый органоид выполняет свою функцию.
Ядро контролирует форму, размер и функционирование клетки, содержит информацию о наследственности.
Клетку окружает тонкая, но прочная оболочка — клеточная мембрана, которая действует как фильтр, пропуская внутрь клетки выборочные вещества и не пропуская другие.
В самой цитоплазме тоже находится сложная сетка мембран — эндоплазматическая сеть, которая действет как фильтр и транспортирует вещества. При помощи этой эндоплазматической сети происходит синтез белков. Белки очень важны для клетки, так как регулируют жизнедеятельность клетки.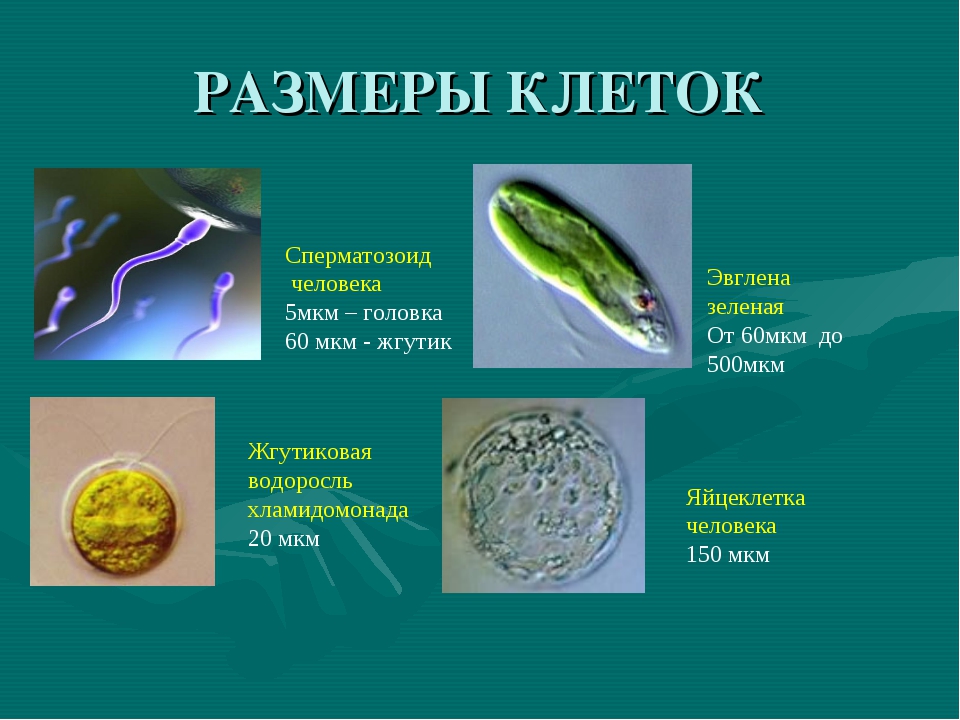
Другой сетчатые аппарат клетки — комплекс Гольджи — состоит из пузырьков, трубочек, цистерн и пластин, участвуют в синтезе компонентов клеточных мембран, секретирует и выводит некоторые вещества на поверность.
Очень важны также находящиеся в цитоплазме митохондрии, которые образуют богатые энергией вещества, необходимые при жизнедеятельности.
Ядро, митохондрии и мембрану можно найти как в растительной. так и в животной клетке. Но в растительной клетке есть еще некоторые составные части, которых нет в животной клетке. Рассматривая растительную клетку под микроскопом, можно увидеть оболочку, которая окружает клетку снаружи. Эта оболочка состоит из целюлозы (одного из углеводов).
Многие растительные клетки содержат заполненные жидкостью пузырьковыми частицами — вакуолями, которые должны держать клетку под напряжением. Вакуоли заполнены клеточным соком. Иногда этот клеточный сок может быть цветным, как, например, у свеклы. Цвет некоторых цветов тоже завсит от клеточного сока, который содержится в клетках лепестков.
Многие растительные клетки содержат еще хролопласт, который находится в цитоплазме. Хролопласт — это овальные органоиды, в состав которых входит хлорофилл, который придает растению зеленый цвет и необходим при фотосинтезе. Хлорофилл связывает энергию белка, которую использует клетка для свеого питания и жизнедеятельности. Хлоропласты хорошо видны и через обычный оптический микроскоп.
В некоторых растительных клетках есть похожие на хлоропласт системы — хромопласты. Они содержат красный или желтый пигмент, который придает цвет цветам и плодам.
1. Строение растительной клетки: оболочка, мембрана, цитоплазма, митохондрии, пластиды и ядро.
Растительные клетки похожи по свеому строению на животные клетки, но горадо больше по размеру. Поэтому при изучении строения клетки рекомендуется использовать растительные клетки.
Для чего нужны клеткам разные части?
Клетку окружает оболочка, которая выполняет защитную функцию. Проникнуть через оболочку можно только по маленьким каналам.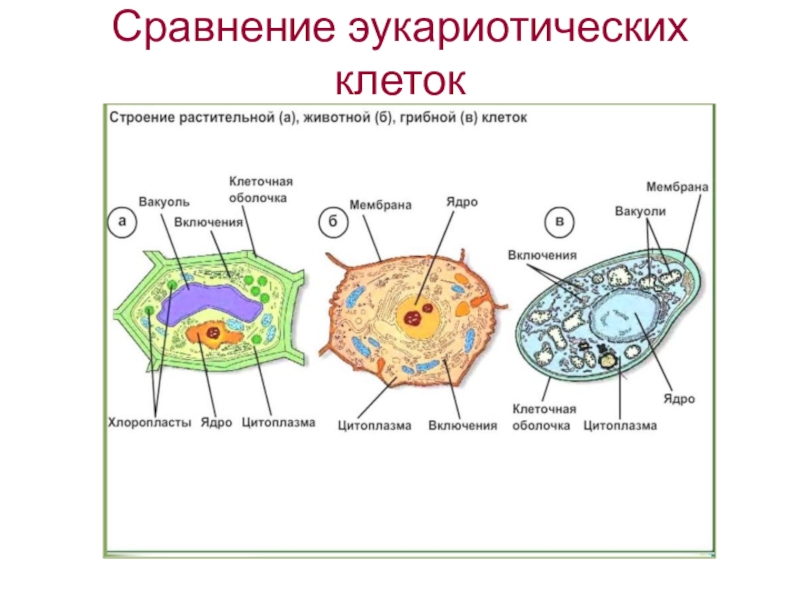
Под оболочкой находится тонкая мембрана. Она выборочно пропускает вещества внутрь и наружу.
Посередине клетки находится ядро, в котором хранится наследственная информация. Ядро руководит жизнедеятельностью клетки.
Клетки не могут расти бесконечно. Ядро содержит наследственную информацию, благодаря которой происходит размножение клетки. Клетка делится на две части, и образуются две совершенно одинаковые клетки.
Энегию клетке дают митохондрии и зеленые солнечные батарейки — пластиды.
В пластидах происходит синтез питательных веществ, митохондрии перерабатывают питательные вещества и высвобождают из них энергию.
Все содержимое клетки находится в полужидкой плазме.
Большую часть растительной клетки заполняет вакуоль, заполненная клеточным соком. Вакуоль держит клетку в напряжении.
Клетка — эта мельчайшая часть живого организма, которая может самостоятельно размножаться, питаться и расти.
Клекти осуществялют все процессы, происходящие в теле.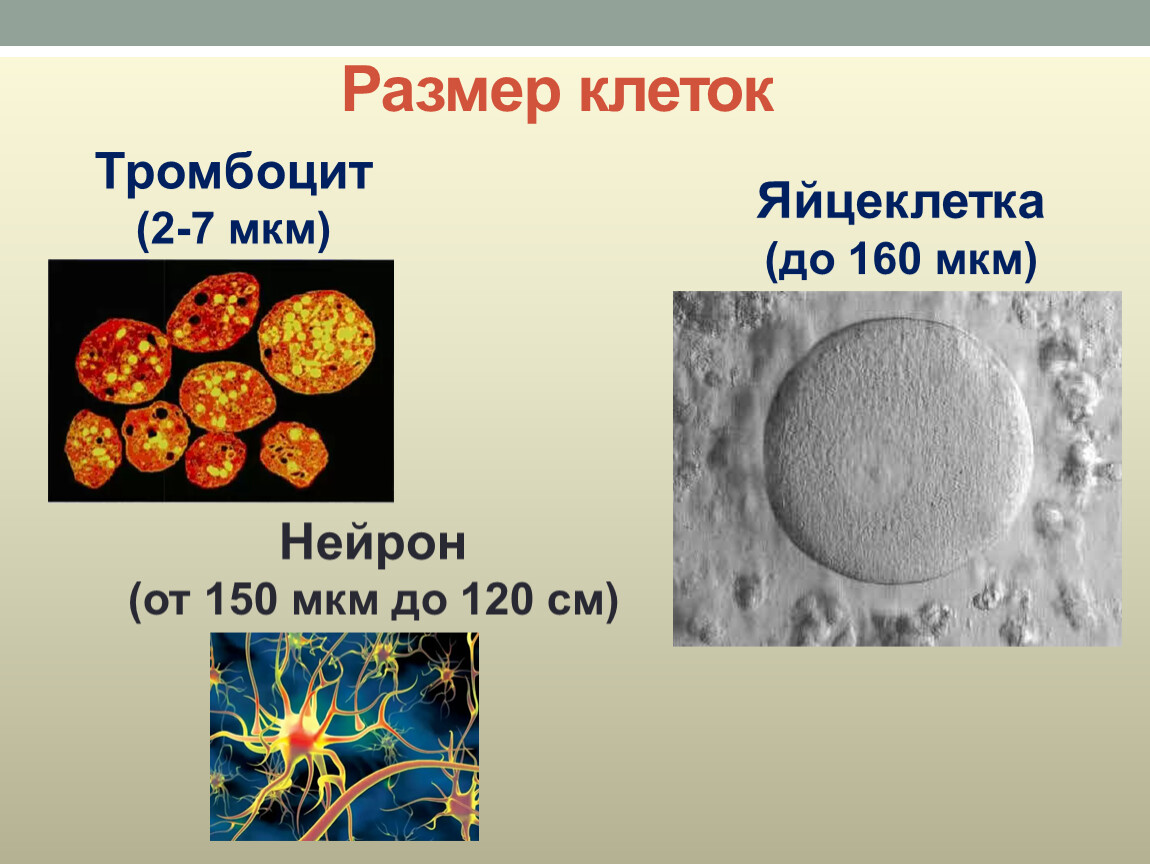
Так как животные клетки очень маленькие, их следует рассматривать с помощью микроскопа, дающего большое увеличение.
У животных клеток отсутствует оболочка, у нних нет больших вакуолей.
Животная клетка в отличие от растительной клетки не может сама синтезировать питательные вещества, в них отсутствуют пластиды.
Сравним: чем отличаются животные клетки от растительных клеток?
| растительная клетка | животная клетка |
| оболочка мембрана цитоплазма ядро митохондрии пластиды вакуоли другие органоиды |
мембрага цитоплазма ядро митохондрии другие органоиды |
Опыты с микроскопом:
микроскоп, стекло-основа, верхнее стекло, игла, впитывающая влагу бумага, нож.
1. Рассматривание лука. Находим части клетки.
Рассматривание лука. Находим части клетки.
Если окрасить лук йодом, то картинка станет еще четче.
2. Рассматривание яблока. Соскрести немного мякоти и капнуть в каплю воды на стекле.
Пластиды в растениях могут быть разного цвета, и у них разные обязанности. Также пластиды дают цвет. Рассматриваем разные пластиды.
3. Хлоропласт в листе водоросли (из аквариума).
4. Хромопласт в мякоти томата.
5. Лейкопласты в чешуе лука. Напомнить детям, что на самом деле лук — это видоизмененный стебель, хотя в нем нет хлоропластов. Почему? Потому что в нем не происходит фотосинтеза из-за того, что р
Органоиды клетки, подготовка к ЕГЭ по биологии
Органоиды (органеллы) клетки — специализированные структуры клетки, выполняющие различные жизненно необходимые функции. Особенно сложно устроены клетки простейших, где одна клетка составляет весь организм и выполняет функции дыхания, выделения, пищеварения и многие другие.
Органоиды клетки подразделяются на:
- Немембранные — рибосомы, клеточный центр, микротрубочки, органоиды движения (жгутики, реснички)
- Одномембранные — ЭПС, комплекс (аппарат) Гольджи, лизосомы и вакуоли
- Двумембранные — ядро, пластиды, митохондрии
Прежде чем говорить об органоидах клетки, без которых невозможна ее жизнедеятельность, необходимо упомянуть о том, без чего вообще не существует клетки — о клеточной мембране. Клеточная мембрана ограничивает клетку от окружающего мира и формирует ее внутреннюю среду.
Клеточная мембрана (оболочка)
Запомните, что в отличие от клеточной стенки, которая есть только у растительных клеток и у клеток грибов (она придает им плотную, жесткую форму) клеточная мембрана есть у всех клеток без исключения! Этот важный момент объясню еще раз 🙂 У клеток животных имеется только клеточная мембрана, а у клеток растений и грибов есть и клеточная стенка, и клеточная мембрана.
Клеточная мембрана представляет собой билипидный слой (лат. bi — двойной + греч. lipos — жир), который пронизывают молекулы белков.
Билипидный слой представлен двумя слоями фосфолипидов. Обратите внимание, что их гидрофобные концы обращены внутрь мембраны, а гидрофильные «головки» смотрят наружу. Билипидный слой насквозь пронизывают интегральные белки, частично — погруженные белки, имеются также поверхностно лежащие белки — периферические.
Белки принимают участие в:
- Поддержании постоянства структуры мембраны
- Рецепции сигналов из окружающей среды (химического раздражения)
- Транспорте веществ через мембрану
- Ускорении (катализе) реакций, которые ассоциированы с мембраной
Интегральные (пронизывающие) белки образуют каналы, по которым молекулы различных веществ могут поступать в клетку или удаляться из нее. «Заякоренные» молекулы олигосахаридов на поверхности клетки образуют гликокаликс, который выполняет рецепторную функцию, участвует в избирательном транспорте веществ через мембрану.
Теперь вы знаете, что гликокаликс — надмембранный комплекс, совокупность клеточных рецепторов, которые нужны клетке для восприятия регуляторных сигналов биологически активных веществ (гормонов, гормоноподобных веществ). Гормон избирателен, специфичен и присоединяется только к своему рецептору: меняется конформация молекулы рецептора и обмен веществ в клетке. Так гормоны регулируют жизнедеятельность клеток.
Вирусы и бактерии не являются исключением: они взаимодействуют только с теми клетками, на которых есть подходящие к ним рецепторы. Так, вирус гриппа поражает преимущественно клетки слизистой верхних дыхательных путей. Однако, если рецепторов нет, то вирус не может проникнуть в клетку, и организм приобретает невосприимчивость к инфекции. Вспомните врожденный иммунитет: именно по причине отсутствия рецепторов человек не восприимчив ко многим болезням животных.
Итак, вернемся к клеточной мембране. Ее можно сравнить со стенами помещения, в котором, вероятно, вы находитесь. Стены дома защищают его от ветра, дождя, снега и прочих факторов внешней среды. Рискну предположить, что в вашем доме есть окна и двери, которые по мере необходимости открываются и закрываются 🙂 Так и клеточная мембрана может сообщать внутреннюю среду клетки с внешней средой: через мембрану вещества поступают в клетку и удаляются из нее.
Подведем итоги. Клеточная мембрана выполняет ряд важнейших функций:
- Разделительная (барьерная) — образует барьер между внешней средой и внутренней средой клетки (цитоплазмой с органоидами)
- Поддержание обмена веществ между внешней средой и цитоплазмой
- Транспортная
- Пассивный — часто идет по градиенту концентрации, без затрат АТФ (энергии). Возможен путем осмоса, простой диффузии или облегченной (с участием белка-переносчика) диффузии.
- Активный
Через мембрану по каналам кислород и питательные вещества поступают в клетку, а продукты жизнедеятельности — мочевина — удаляются из клетки во внешнюю среду.
Тесно связана с обменом веществ, однако здесь мне особенно хочется подчеркнуть варианты транспорта веществ через клетку. Выделяется два вида транспорта:
Внутрь клетки с помощью осмоса поступает вода. Путем простой диффузии в клетку попадают O2, H2O, CO2, мочевина. Облегченная диффузия характерна для транспорта глюкозы, аминокислот.
Активный транспорт чаще происходит против градиента концентрации, в ходе него используются белки-переносчики и энергия АТФ. Ярким примером является натрий-калиевый насос, который накачивает ионы калия внутрь клетки, а ионы натрия выводит наружу. Это происходит против градиента концентрации, поэтому без затрат энергии (АТФ) не обойтись.
Внутрь клетки крупные молекулы попадают путем эндоцитоза (греч. endo — внутрь) двумя путями:
- Фагоцитоз (греч. phago — ем + cytos — клетка) — поглощение твердых пищевых частиц и бактерий фагоцитами
- Пиноцитоз (греч. pino — пью) — поглощение клеткой жидкости, захват жидкости клеточной поверхностью
Фагоцитоз был открыт И.И. Мечниковым, который создал фагоцитарную теорию иммунитета. Это теория гласит, что в основе иммунной системы нашего организма лежит явление фагоцитоза: попавшие в организм бактерии уничтожаются фагоцитами (T-лимфоцитами), которые переваривают их.
В ходе эндоцитоза мембрана сильно прогибается внутрь клетки, ее края смыкаются, захватывая бактерию, пищевые частицы или жидкость внутрь клетки. Образуется везикула (пузырек), который движется к пищеварительной вакуоли или лизосоме, где происходит внутриклеточное пищеварение.
Клетки многих органов, к частности эндокринных желез, которые выделяют в кровь гормоны, транспортируют синтезированные вещества к мембране и удаляют их из клетки с помощью экзоцитоза (от др.-греч. ἔξω — вне, снаружи). Таким образом, процессы экзоцитоза и эндоцитоза противоположны.
Клеточная стенка
Расположена снаружи клеточной мембраны. Присутствует только в клетках бактерий, растений и грибов, у животных отсутствует. Придает клетке определенную форму, направляет ее рост, придавая характерное строение всему организму. Клеточная стенка бактерий состоит из полимера муреина, у грибов — из хитина, у растений — из целлюлозы.
Цитоплазма
Органоиды клетки расположены в цитоплазме, которая состоит из воды, питательных веществ и продуктов обмена. В цитоплазме происходит постоянный ток веществ: поступившие в клетку вещества для расщепления необходимо доставить к органоидам, а побочные продукты — удалить из клетки.
Постоянное движение цитоплазмы поддерживает связь между органоидами клетки и обеспечивает ее целостность.
Прокариоты и эукариоты
Прокариоты (греч. πρό — перед и κάρυον — ядро) или доядерные — одноклеточные организмы, не обладающие в отличие от эукариот оформленным ядром и мембранными органоидами. У прокариот могут обнаруживаться только немембранные органоиды. Их генетический материал представлен в виде кольцевой молекулы ДНК — нуклеоида. К прокариотам относятся бактерии, в их числе цианобактерии (цианобактерий по-другому называют — сине-зеленые водоросли).
Эукариоты (греч. εὖ — хорошо + κάρυον — ядро) или ядерные — домен живых организмов, клетки которых содержат оформленное ядро. Растения, животные, грибы — относятся к эукариотам.
Немембранные органоиды
- Рибосома
- Микротрубочки и микрофиламенты
- Клеточный центр (центросома, от греч. soma — тело)
- Реснички и жгутики
Очень мелкая органелла (около 20 нм), которая была открыта после появления электронного микроскопа. Состоит из двух субъединиц: большой и малой, в состав которых входят белки и рРНК (рибосомальная РНК), синтезируемая в ядрышке.
Запомните ассоциацию: «Рибосома — фабрика белка». Именно здесь в ходе матричного биосинтеза — трансляции, с которой подробнее мы познакомимся в следующих статьях, на базе иРНК (информационной РНК) синтезируется белок — последовательность соединенных аминокислот в заданном иРНК порядке.
Микротрубочки являются внутриклеточными белковыми производными, входящими в состав цитоскелета. Они поддерживают определенную форму клетки, участвуют во внутриклеточном транспорте и процессе деления путем образования нитей веретена деления. Микротрубочки также образуют основу органоидов движения: жгутиков (у бактерий жгутик состоит из сократительного белка — флагеллина) и ресничек.
Микрофиламенты — тонкие длинные нитевидные структуры, состоящие из белка актина. Встречаются во всей цитоплазме, служат для создания тока цитоплазмы, принимают участие в движении клетки, в процессах эндо- и экзоцитоза.
Этот органоид характерен только для животной клетки, в клетках грибов и высших растений отсутствует. Клеточный центр состоит из 9 триплетов микротрубочек (триплет — три соединенных вместе). Участвует в образовании нитей веретена деления, располагается на полюсах клетки.
Это органоиды движения, которые выступают над поверхностью клетки и имеют в основе пучок микротрубочек. Реснички встречаются только в клетках животных, жгутики можно обнаружить у животных, растений и бактерий.
Одномембранные органоиды
- Эндоплазматическая сеть (ЭПС), эндоплазматический ретикулум (лат. reticulum — сеть)
- Комплекс (аппарат) Гольджи
- Лизосома (греч. lisis — растворение + soma — тело)
- Пероксисомы (лат. per — сверх, греч. oxys — кислый и soma — тело)
- Вакуоли
ЭПС представляет собой систему мембран, пронизывающих всю клетку и разделяющих ее на отдельные изолированные части (компартменты). Это крайне важно, так как в разных частях клетки идут реакции, которые могут помешать друг другу, что нарушит процессы жизнедеятельности.
Выделяют гладкую ЭПС и шероховатую ЭПС. Обе они выполняют функцию внутриклеточного транспорта веществ, однако между ними имеются различия. На мембранах гладкой ЭПС происходит синтез липидов, обезвреживаются вредные вещества. Шероховатая ЭПС синтезирует белок, так как имеет на мембранах многочисленные рибосомы (потому и называется шероховатой).
Комплекс Гольджи состоит из трубочек, сети уплощенных канальцев (цистерн) и связанных с ними пузырьков. Располагается вокруг ядра клетки, внешне напоминает стопку блинов. Это — «клеточный склад». В нем запасаются жиры и углеводы, с которыми здесь происходят химические видоизменения.
Модифицированные вещества упаковываются в пузырьки и могут перемещаться к мембране клетки, соединяясь с ней, они изливают свое содержимое во внешнюю среду. Можно догадаться, что комплекс Гольджи хорошо развит в клетках эндокринных желез, которые в большом количестве синтезируют и выделяют в кровь гормоны.
В комплексе Гольджи появляются первичные лизосомы, которые содержат ферменты в неактивном состоянии.
Представляет собой мембранный пузырек, содержащий внутри ферменты (энзимы) — липазы, протеазы, фосфатазы. Лизосому можно ассоциировать с «клеточным желудком».
Лизосома участвует во внутриклеточном пищеварении поступивших в клетку веществ. Сливаясь с фагосомой, первичная лизосома превращается во вторичную, ферменты активируются. После расщепления веществ образуется остаточное тельце — вторичная лизосома с непереваренными остатками, которые удаляются из клетки.
Лизосома может переварить содержимое фагосомы (самое безобидное), переварить часть клетки или всю клетку целиком. В норме у каждой клетки жизненный цикл заканчивается апоптозом — запрограммированным процессом клеточной гибели.
В ходе апоптоза ферменты лизосомы изливаются внутрь клетки, ее содержимое переваривается. Предполагают, что нарушение апоптоза в раковых клетках ведет к бесконтрольному росту опухоли.
Пероксисомы (микротельца) содержат окислительно-восстановительные ферменты, которые разлагают H2O2 (пероксид водорода) на воду и кислород. Если бы пероксид водорода оставался неразрушенными, это приводило бы к серьезным повреждениям клетки.
Вакуоли характерны для растительных клеток, однако встречаются и у животных (у одноклеточных — сократительные вакуоли). У растений вакуоли выполняют другие функции и имеют иное строение: они заполняются клеточным соком, в котором содержится запас питательных веществ. Снаружи вакуоль окружена тонопластом.
Трудно переоценить значение вакуолей в жизнедеятельности растительной клетки. Вакуоли создают осмотическое давление, придают клетке форму.
Примечательно, что по размеру вакуолей можно судить о возрасте клетки: молодые клетки имеют вакуоли небольшого размера, а в старых клетках вакуоли могут настолько увеличиваться, что оттесняют ядро и остальные органоиды на периферию.
Двумембранные органоиды
- Ядро («ядро» по лат. — nucleus, по греч. — karyon)
- Митохондрия
- Пластиды (др.-греч. πλαστός — вылепленный)
- Хлоропласт (греч. chlōros — зелёный)
- Хромопласты (греч. chromos – краска)
- Лейкопласты (др.-греч. λευκός — белый )
Важнейший компонент эукариотической клетки — оформленное ядро, которое у прокариот отсутствует. Внутренняя часть ядра представлена кариоплазмой, в которой расположен хроматин — комплекс ДНК, РНК и белков, и одно или несколько ядрышек.
Ядрышко — место в ядре, где активно идет процесс матричного биосинтеза — транскрипция, с которым мы познакомимся подробнее в следующих статьях. В течение дня, наблюдая за одной и той же клеткой, можно увидеть разное количество ядрышек или не найти ни одного.
Оболочка ядра состоит из двух мембран и пронизана большим количеством ядерных пор, через которые происходит сообщение между кариоплазмой и цитоплазмой. Главными функциями ядра является хранение, защита и передача наследственного материала дочерним клеткам.
Замечу, что хромосомы видны только в момент деления клетки. Хромосомы представляют собой сильно спирализованные молекулы ДНК, связанные с белками.
Я всегда рекомендую ученикам ассоциировать хромосому с мотком ниток: если все нитки обмотать вокруг одной оси, то они становятся мотком и хорошо видны (хромосомы — во время деления, спирализованное ДНК), если же клетка не делится, то нитки размотаны и разбросаны в один слой, хромосом не видно (хроматин — деспирализованное ДНК).
Хромосомы отличаются друг от друга по строению, форме, размерам. Совокупность всех признаков (форма, число, размер) хромосом называется кариотип. Кариотип может быть представлен по-разному: существует кариотип вида, особи, клетки.
Изучая кариотип человека, врач-генетик может обнаружить различные наследственные заболевания, к примеру, синдром Дауна — трисомия по 21-ой паре хромосом (должно быть 2 хромосомы, однако при синдроме Дауна их три).
Органоид палочковидной формы. Митохондрию можно сравнить с «энергетической станцией». Если в цитоплазме происходит анаэробный этап дыхания (бескислородный), то в митохондрии идет более совершенный — аэробный этап (кислородный). В результате кислородного этапа (цикла Кребса) из двух молекул пировиноградной кислоты (образовавшихся из 1 глюкозы) получаются 36 молекул АТФ.
Митохондрия окружена двумя мембранами. Внутренняя ее мембрана образует выпячивания внутрь — кристы, на которых имеется большое скопление окислительных ферментов, участвующих в кислородном этапе дыхания. Внутри митохондрия заполнена матриксом.
Запомните, что особенностью этого органоида является наличие кольцевой молекулы ДНК — нуклеоида, и рибосом. То есть митохондрия обладает собственным генетическим материалом и возможностью синтеза белка, почти как отдельный организм.
В связи с этим, митохондрия считается полуавтономным органоидом. Вероятнее всего, изначально митохондрии были самостоятельными организмами, однако со временем вступили в симбиоз с эукариотами и стали частью клетки.
Митохондрий особенно много в клетках мышц, в том числе — в сердечной мышечной ткани. Эти клетки выполняют активную работу и нуждаются в большом количестве энергии.
Двумембранные органоиды, встречающиеся только в клетках высших растений, водорослей и некоторых простейших. У подавляющего большинства животных пластиды отсутствуют. Подразделяются на три типа:
Получил свое название за счет содержащегося в нем зеленого пигмента — хлорофилла (греч. chloros — зеленый и phyllon — лист). Под двойной мембраной расположены тилакоиды, которые собраны в стопки — граны. Внутреннее пространство между тилакоидами и мембраной называется стромой.
Запомните, что светозависимая (световая) фаза фотосинтеза происходит на мембранах тилакоидов, а темновая (светонезависимая) фаза — в строме хлоропласта за счет цикла Кальвина. Это очень пригодится при изучении фотосинтеза в дальнейшем.
Так же, как и митохондрии, пластиды относятся к полуавтономным органоидам: в них имеется кольцевидная ДНК — нуклеоид, рибосомы.
Пластиды, которые содержат пигменты каратиноиды в различных сочетаниях. Сочетание пигментов обуславливает красную, оранжевую или желтую окраску. Находятся в плодах, листьях, лепестках цветков.
Хромопласты могут развиваться из хлоропластов: во время созревания плодов хлоропласты теряют хлорофилл и крахмал, в них активируется биосинтез каротиноидов.
Не содержат пигментов, образуются в запасающих частях растения (клубни, корневища). В лейкопластах накапливается крахмал, липиды (жиры), пептиды (белки). На свету лейкопласты могут превращаться в хлоропласты и запускать процесс фотосинтеза.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Модель-конструктор к урокам биологии в школе «Сравнение растительной и животной клеток»
Цель
Создать 3D-модель − конструктор животной и растительной клеток для работы на уроках биологии.
Задачи
- Проанализировать и сравнить строение растительной и животной клеток.
- Научиться работать с программой Autodesk Fusion 360.
- Создать модели органоидов клетки и самой клетки.
- Научиться работать с 3D-принтером.
- Провести уроки в 5-х и 6-х классах с готовым наглядным пособием, а также в медицинском классе.
Описание
В процессе работы:
− разработана концепция проекта;
− получен опыт работы в Autodesk Fusion 360;
− сделан эскиз модели, определены размеры;
− клетка смоделирована в программе;
− части клетки напечатаны на 3D-принтере;
− собрана модель;
− проведён пробный урок в 5−6 классах, а также в медицинском классе.
Результат
Авторы провели пробные уроки в 5 и 6 классах. Сначала ребята прошли мини-опросы, на вопросы которых они иногда затруднялись ответить. Но когда авторы стали объяснять всё и показывать с помощью 3D-модели, дети были очень заинтересованы. Они с интересом всё доставали и рассматривали. В результате они смело отвечали на вопросы.
Оснащение и оборудование
- Autodesk Fusion 360
- D-принтер
Перспективы использования результатов работы
Авторы не остановятся на достигнутых результатах. В будущем планируют распечатать большее количество 3D-моделей, чтобы на уроках биологии модель клетки присутствовала на каждой парте. Её использование будет очень удобным, так как модель является переносной за счёт двух крышек, которые будут прочно вставляться в отверстия. А также в клетке имеется два ящика. В них будут помещаться части клетки. Один ящик – для органоидов растительной клетки, а другой – для органоидов животной клетки.
Конспект и презентация урока по биологии на тему «Строение животной клетки», 6 класс
Учитель: Захарова Екатерина Ивановна
Класс: 6
Дата проведения:
Предмет: Биология. Живой организм.
Тема урока: Строение животной клетки
Цель урока: показать особенности строения и функционирования животной клетки
Задачи урока:
Образовательные задачи урока: изучить строение животной клетки и функции органоидов, входящих в состав клетки.
Развивающие задачи урока: развивать навыки самостоятельной работы с различными источниками информации; сравнивать биологические объекты, устанавливать связи, делать выводы; развивать образную память, логическое мышление, речь.
Воспитательные задачи урока: осознанно достигать поставленной цели; воспитывать положительное отношение к совместному труду
Тип урока: комбинированный урок
Приемы обучения: актуализация базовых умений, закрепление учебного материала на основе просмотра презентации, словесные (беседа, объяснение), наглядные (презентация), практические (тест, задания)
Формы работы: общеклассная, парная, индивидуальная
Оборудование: раздаточный материал — тесты, модель животной клетки, компьютер, проектор, презентация “Строение животной клетки”, учебник
Применяемые технологии:
№ п/п | Название современных образовательных технологий, применяемых в УВП | Этапы урока/занятия (мероприятия), на которых технология применяется | |
1 | Здоровьесбережение | Целеполагание, физкультминутка в середине урока | |
2 | ИКТ | На всех этапах (мультимедийная презентация) | |
3 | Личностно-ориентированный подход | Актуализация ранее изученного материала Ответы на вопросы Самостоятельная работа (тест, карточки) | |
4 | Проблемное обучение | Этап мотивации, работа по теме урока, рефлексия | |
5 | Технология развития критического мышления. | Актуализация опорных знаний, усвоение новых знаний, применение изученного материала, подведение итогов | |
Основные этапы урока:
I. Организационный момент — 1 мин.
II. Проверка д/з – 5 мин.
III. Введение в тему – 2 мин.
IV. Изучение нового материала – 20 мин.
V. Закрепление знаний – 8 мин.
VI. Подведение итогов – 2 мин.
VII. Домашнее задание – 1 мин.
VIII Рефлексия – 1 мин.
ХОД УРОКА:
Организационный момент — 1 мин.
Приветствие класса.
Отчет дежурных об отсутствующих.
Готовность к уроку, настроение.
IIроверка домашнего задания – 5 мин.
Несколько учеников работают по индивидуальным карточкам.
Карточка 1
Тестовые задания с выбором одного правильного ответа
1. Кто впервые обнаружил клетку?
а) Роберт Вирхов;
б) Антуан Ван Левенгук;
в) Роберт Гук.
2. В каком году?
а) 1600 г.;
б) 1930 г.;
в) 1665 г.;
3. Клетка снаружи покрыта:
а) цитоплазмой;
б) оболочкой;
в) пластидами.
4. Зеленые пластиды называются:
а) лейкопласты;
б) хлоропласты;
в)хромопласты.
5. Внутренняя среда клетки, где расположены все органоиды, называется:
а) цитоплазма;
б) ядро;
в) вакуоли.
6. Хромосомы находятся в:
а) ядре;
б) цитоплазме;
в) вакуоли.
7. Основная структурная единица организма:
а) корень;
б) орган;
в) клетка.
Учащиеся проверяют работу в парах, меняясь своими тетрадями. Опираясь на демонстрируемый слайд учителем (слайд 2)
Критерий оценки:
1 ошибка – «5» баллов;
2 ошибка – «4» балла;
3 ошибки – «3» балла;
от 4 ошибок – «2» балла.
Карточка 2
Несколько учащихся описывают растительную клетку по карточке
Учитель организует работу в парах: проверка и оценка работ знакомит с критериями оценок и просит оценить работу друг у друга. Опираясь на демонстрируемый слайд учителем (слайд 2)
Введение в тему – 2 мин.
Сегодня нам предстоит изучить очень интересную тему из курса биологии.
Из чего состоят все живые организмы из … (клеток). Правильно.
Так какова же тема сегодняшнего урока? (версии детей)
Учитель записывает проговоренную тему на доске, а дети в тетрадях.
Тема урока «Строение животной клетки» (слайд 3)
Изучение нового материала – 20 мин.
На предыдущем уроке мы изучили строение растительной клетки (слайд 4)
Основные отличия растительной от животной клетки:
1) В растительной клетке присутствует прочная и толстая клеточная стенка из целлюлозы
2) В растительной клетке развита сеть вакуолей, в животной клетке она развита слабо
3) Растительная клетка содержит особые органоиды — пластиды (а именно, хлоропласты, лейкопласты и хромопласты) , а животная клетка их не содержит.
Кто может дать определение органоиду? (органоид – часть клетки, выполняющая определенную функцию).
По строению клетки все живые существа делят на 2 большие группы: доядерные (безъядернвые) и ядерные (слайд 5)
Наиболее просто устроены клетки доядерных (безъядерных) организмов – бактерий, сине-зеленых водорослей (цианобактерий).
Клетки ядерных организмов – грибов, растений и животных – имеют ядро и по сравнению с доядерными организмами обладают более сложным разнообразным строением. Остановимся на строение клетки более подробно (слайд 6)
Любая клетка имеет плазматическую мембрану (от латинского «мембрана» — кожица, пленка). Она защищает внутреннее содержимое клетки от воздействий внешней среды. Выросты и складки на поверхности мембраны способствуют прочному соединению клеток между собой. Мембрана пронизана тончайшими канальцами. По канальцам мембраны осуществляется перенос питательных веществ и продуктов жизнедеятельности клетки
Внутренней средой клетки является цитоплазма, в ней располагаются ядро и многочисленные органоиды. Она состоит из вязкого полужидкого вещества. Цитоплазма связывает между собой органоиды, обеспечивает перемещение различных веществ и является средой, в которой идут многочисленные химические реакции.
Цитоплазма пронизана сетью многочисленных мелких канальцев и полостей, соединенных между собой. Это эндоплазматическая сеть. Она составляет 30—50% объема клетки. Эндоплазматическая сеть связывает все части клетки между собой, участвует в образовании и транспортировке различных органических веществ. Важные функции в клетке выполняет аппарат Гольджи.
Он представляет собой стопку упло щенных мембранных мешочков — полостей, в которых накапливаются и сортируются различные вещества От аппарата Гольджи отделяются мелкие мембранные пузырьки, в которых вещества могут переноситься в любую часть клетки и даже выделяться из клетки.
Лизосомы — одни из самых маленьких органоидов клетки — тоже образу ются в аппарате Гольджи. Эти маленькие пузырьки содержат вещества, которые внутри клетки переваривают пищевые частицы, уничтожают отслужившие органоиды и даже целые клетки. Вспомните, что происходит с хвостом головастика: со временем он как бы растворяется и исчезает — это «работа» лизосом.
Во всех клетках находятся небольшие округлые тельца — рибосомы. Они обеспечивают сборку сложных молекул белков.
В результате расщепления питательных веществ, поступивших в клетку, высвобождается энергия. Эта энергия, необходимая для процессов жизнедеятельности, накапливается в митохондриях. В цитоплазме клеток около ядра находится клеточный центр. В клетках животных и низших растений в его состав входят центриоли. Клеточный центр принимает участие в делении клетки.
Физкультминутка:
Вновь у нас физкультминутка,
Наклонились, ну-ка, ну-ка!
Распрямились, потянулись,
А теперь назад прогнулись.
Хоть зарядка коротка,
Отдохнули мы слегка.
Важнейшая часть клетки — ядро. Обычно оно находится в центре клетки. Однако в клетках растений внутренняя часть обычно занята крупной вакуолью, поэтому ядро располагается около клеточной мембраны. Ядро содержит одно или несколько ядрышек. В ядре хранится наследственная информация о данной клетке и об организме в целом. «Записана» эта информация в молекулах нуклеиновой кислоты, которая входит в состав хромосом (от греческого «хрома» — цвет, «сома» — тело). Хромосомы становятся хорошо заметными во время деления клетки.
Установлено, что все клетки тела животных и высших растений имеют двойной набор хромосом, его принято обозначать 2п. Исключение составляют только половые клетки, в которых набор хромосом одинарный, поэтому его обозначают одной буквой п. Число хромосом для каждого вида организмов постоянно и не зависит от уровня его организации. Так, у человека 46 хромосом, у курицы — 78, у овцы — 54, у шимпанзе — 48, у ржи — 14, у дуба — 24. В клетках хромосомы одинакового строения и размера образуют пары. Хромосомы одной пары называют гомологичными (от греческого «гомология» — соответствие, согласие).
Учитель предлагает самостоятельно поработать с учебником и оформить органоиды животной, их функции в клетки в виде таблицы (слайд 7)
Органоиды | Строение | Функции |
Плазматическая мембрана | ||
Эндоплазматическая сеть (ЭПС) | ||
Рибосомы | ||
Митохондрии | ||
Аппарат Гольджи | ||
Ядро | ||
Лизосомы | ||
Клеточный центр |
V. Закрепление знаний – 8 мин (слайд 8-10)
Сравнение растительной и животной клеток.
Растительная клетка | Органоиды | Животная клетка |
+ отличается | Плазматическая мембрана | + отличается |
+ отличается | оболочка | + отличается |
+ | цитоплазма | + |
+ | эндоплазматическая сеть | + |
+ | аппарат Гольджи | + |
+ | лизосомы | + |
+ | рибосомы | + |
+ | митохондрии | + |
+ | пластиды | нет |
+ | вакуоль | очень мало |
+ не у всех | клеточный центр | + |
+ | Ядро | + |
+ | ядрышки | + |
+ | хромосомы | + |
Проверь свои знания
– Какие клетки изображены на рисунке?
– Какие органоиды изображены?
– Какие функции они выполняют?
VI. Подведение итогов – 2 мин.
Таким образом, все живые организмы состоят из клеток, за исключением доклеточных форм жизни – вирусов. Клетки бывают ядерные и безъядерные. Из безъядерных клеток состоят бактерии. Из ядерных клеток состоят растения, грибы и животные.
VII. Домашнее задание – 1 мин. (слайд 11)
§3, повтор. §1-§3
Творческий уровень:
вылепить из пластилина клетку с ее органоидами
VIII Рефлексия – 1 мин.
Учитель просит учащихся продолжить фразу «сегодня на уроке я:
научился
удивился
понял
узнал
— благодарит учащихся за работу
— объявляет об окончании урока
Список использованной литературы:
Биология. Живой организм. 6 класс: учеб.для общеобразоват.учреждений /Н.И. Сонин. – 4-е изд.,стереотип. – М.:Дрофа, 2010. – 174,[2]с.:ил.
В.Б. Захаров, Е.Т. Захарова «Правильные ответы на вопросы учебника Н.И. Сонина «Биология. Живой организм. 6 класс» -М.: ДРОФА, 2006
Различий между растительными и животными клетками
Элисон Чинкота / Иллюстрация / ThoughtCo
Клетки животных и клетки растений похожи в том, что они оба являются эукариотическими клетками. Эти клетки имеют собственное ядро, в котором находится ДНК и отделено от других клеточных структур ядерной мембраной. Оба этих типа клеток имеют схожие процессы воспроизводства, включая митоз и мейоз. Клетки животных и растений получают энергию, необходимую для роста и поддержания нормальной клеточной функции, в процессе клеточного дыхания.Оба этих типа клеток также содержат клеточные структуры, известные как органеллы, которые специализируются на выполнении функций, необходимых для нормальной работы клеток. Клетки животных и растений имеют некоторые общие компоненты клетки, включая ядро, комплекс Гольджи, эндоплазматический ретикулум, рибосомы, митохондрии, пероксисомы, цитоскелет и клеточную (плазматическую) мембрану. Хотя животные и растительные клетки имеют много общих характеристик, они также различны.
Различия между животными и растительными клетками
Британника / UIG / Getty Images
Размер
Клетки животных обычно меньше клеток растений.Клетки животных имеют длину от 10 до 30 микрометров, а клетки растений — от 10 до 100 микрометров.
Форма
Клетки животных бывают разных размеров и, как правило, имеют круглую или неправильную форму. Клетки растений более похожи по размеру и обычно имеют прямоугольную или кубическую форму.
Накопитель энергии
Клетки животных хранят энергию в виде сложного углеводного гликогена. Клетки растений хранят энергию в виде крахмала.
Белки
Из 20 аминокислот, необходимых для производства белков, только 10 могут вырабатываться естественным путем в клетках животных.Другие так называемые незаменимые аминокислоты необходимо получать с пищей. Растения способны синтезировать все 20 аминокислот.
Дифференциация
В клетках животных только стволовые клетки способны превращаться в клетки других типов. Большинство типов растительных клеток способны к дифференцировке.
Рост
Клетки животных увеличиваются в размерах за счет увеличения числа клеток. Растительные клетки в основном увеличивают размер клетки за счет увеличения. Они растут за счет поглощения большего количества воды центральной вакуолью.
Стенка клетки
Клетки животных не имеют клеточной стенки, но имеют клеточную мембрану. Клетки растений имеют клеточную стенку, состоящую из целлюлозы, а также клеточную мембрану.
центриолей
Клетки животных содержат эти цилиндрические структуры, которые организуют сборку микротрубочек во время деления клеток. Клетки растений обычно не содержат центриолей.
Реснички
Реснички встречаются в клетках животных, но не обычно в клетках растений.Реснички — это микротрубочки, которые помогают перемещению клеток.
Цитокинез
Цитокинез, деление цитоплазмы во время деления клеток, происходит в клетках животных, когда образуется борозда дробления, которая сдавливает клеточную мембрану пополам. При цитокинезе растительной клетки создается клеточная пластинка, которая делит клетку.
Глиоксисомы
Эти структуры не обнаруживаются в клетках животных, но присутствуют в клетках растений. Глиоксисомы помогают расщеплять липиды, особенно в прорастающих семенах, для производства сахара.
Лизосомы
Клетки животных обладают лизосомами, которые содержат ферменты, переваривающие клеточные макромолекулы. Растительные клетки редко содержат лизосомы, поскольку растительная вакуоль справляется с расщеплением молекул.
Пластиды
Клетки животных не имеют пластид. Клетки растений содержат пластиды, такие как хлоропласты, которые необходимы для фотосинтеза.
Клетки животных не имеют плазмодесм. У растительных клеток есть плазмодесмы, которые представляют собой поры между стенками растительных клеток, которые позволяют молекулам и коммуникационным сигналам проходить между отдельными растительными клетками.
Вакуоль
Клетки животных могут иметь множество мелких вакуолей. Растительные клетки имеют большую центральную вакуоль, которая может занимать до 90% объема клетки.
Прокариотические клетки
CNRI / Getty Images
Эукариотические клетки животных и растений также отличаются от прокариотических клеток, таких как бактерии. Прокариоты обычно одноклеточные, а клетки животных и растений — многоклеточные. Эукариотические клетки сложнее и крупнее прокариотических клеток.Клетки животных и растений содержат множество органелл, которых нет в прокариотических клетках. У прокариот нет настоящего ядра, поскольку ДНК не содержится в мембране, а свернута спиралью в области цитоплазмы, называемой нуклеоидом. В то время как животные и растительные клетки размножаются путем митоза или мейоза, прокариоты размножаются чаще всего путем бинарного деления.
Другие эукариотические организмы
МАРЕК МИС / НАУЧНАЯ ФОТОБИБЛИОТЕКА / Getty Images
Клетки растений и животных — не единственные типы эукариотических клеток.Протисты и грибы — два других типа эукариотических организмов. Примеры простейших включают водоросли, эвглены и амебы. Примеры грибов включают грибы, дрожжи и плесень.
Machalek AZ. Внутри камеры. Глава 1: Руководство для владельца ячейки. Национальный институт общих медицинских наук. Проверено 9 августа 2012 г. http://publications.nigms.nih.gov/insidethecell/chapter1.html
Cooper GM. Клетка: молекулярный подход. 2-е издание. Сандерленд (Массачусетс): Sinauer Associates; 2000 г.Молекулярный состав клеток. Доступно по адресу: http://www.ncbi.nlm.nih.gov/books/NBK9879/
Смотри: Что такое метазоа?
Определение размера клеток в клетках животных координирует анаболические скорости роста и развитие клеточного цикла для поддержания однородности размера клеток
Существенных доработок (из обзоров):
1) Введение требует доработки.
(из обзора № 1): во введении не описываются предложенные и экспериментально подтвержденные модели «калибратора» для почкующихся и делящихся дрожжей.Их следует объяснить, поскольку они являются лучшими примерами контроля размера в литературе. Вдобавок я бы не согласился с утверждением авторов, что эта область считает, что «рост и прогрессирование клеточного цикла не скоординированы». Напротив, есть масса доказательств того, что рост клеток регулирует развитие клеточного цикла. Перепишите вступление, чтобы лучше отразить литературу и верования в этой области.
(из обзора № 2): авторы могли бы лучше сформулировать проблему контроля размера, возможно, начав с описания наблюдений Зеттерберга и др.кратко. Я не согласен с утверждением, что общее мнение состоит в том, что в клетках животных не существует контроля размера. Доказательства в пользу контроля размеров, восходящие к Зеттербергу, убедительны, и обзор, процитированный об обратном, является необычным мнением, которое никоим образом не репрезентативно, и поэтому я не думаю, что уместно цитировать из этот обзор. Сосредоточение внимания на общем вопросе активного контроля размера в некоторой степени отвлекает от ключевого вывода о том, что существует добросовестный механизм компенсации, зависящий от размера, в отличие от простого порогового значения.В этой заметке авторы не упоминают все точки ограничения, которые могут показаться логичным кандидатом на первую из их компенсационных фаз, учитывая время и эффект ингибитора CDK4 / 6. Точку R следует упомянуть во введении и обсуждении.
Следуя предложению обзоров, мы полностью переписали Введение. В новом «Введении» мы рассматриваем литературу о размере клеток как в клетках животных, так и в дрожжах. Другие темы, затронутые в новом Введении, включают различие между измерителем размера модели и сумматором модели , модель определения размера титрования / разбавления (Whi5, Cln3), геометрические модели измерения размера (Cdr2 / Pom1) и модель Зеттерберга. оригинальные публикации.
2) Несмотря на значимость существенных результатов, работа представлена не очень хорошо. Во введении нет точного или исчерпывающего описания предыдущей работы, раздел «Результаты» наполнен отвлекающими отрывками из обсуждения, которые не являются результатами, а описания методов, анализа и результатов в некоторых точках непонятны и трудны для понимания и оценки. Чтобы убедить рецензентов в обоснованности выводов и доступности их для потенциальных читателей, статья нуждается в серьезной доработке.
В обновленной версии мы тщательно отредактировали раздел «Результаты» для большей ясности. Кроме того, некоторые разделы были удалены из раздела «Результаты» и после значительного повторного редактирования были перенесены в раздел «Обсуждение».
3) Корреляция размера ядра и длины G1 (рис. 1C) довольно слабая и не очень сильно поддерживает вывод авторов. Более того, имеется мало доказательств, подтверждающих предположение авторов о том, что зависимость размера ядра от размера клетки является фиксированной.Пожалуйста, предоставьте такие доказательства, если это возможно, чтобы подтвердить обоснованность использования измерения размера ядер.
Мы решаем эту проблему с помощью четырех разных подходов:
1) Мы предоставляем новые измерения для подтверждения использования ядерной площади в качестве показателя размера ячейки. Эти новые измерения включены в рисунок 2 и рисунок 2 — дополнение к рисунку 2 отредактированной рукописи. Эти новые измерения обсуждаются в разделе «Результаты» исправленной рукописи, в разделе, который начинается с предложения «Известно, что у дрожжей ядро непрерывно растет на всех стадиях клеточного цикла и коррелирует с размером клетки (Jorgensen et al., 2007). Чтобы проверить, так ли обстоит дело и с нашей экспериментальной системой, мы измерили корреляцию между размером ядра и размером клетки […] »
Вкратце, чтобы показать, что площадь ядра является надежным показателем размера клетки, мы представляем следующие измерения:
A) Мы показываем, что корреляция размера клетки и размера ядра очень значительна (рис. 2A).
B) Мы показываем, что и размер клетки, и размер ядра неуклонно увеличиваются по мере того, как клетки продвигаются в клеточном цикле (Рисунок 2B-2C и Рисунок 2 — рисунок Приложение 2).Кроме того, эти цифры показывают, что увеличение размера ядра и увеличение размера клетки линейно коррелируют. Рисунок 2C также показывает, что разрешение измерений площади ядра является достаточным, чтобы различать различия в средних размерах, которые возникают в результате менее чем трех часов роста клеток (~ 15% длины клеточного цикла).
C) На рис. 2 — приложение 2, мы показываем, что рост ядер не ограничивается S-фазой и является непрерывным на протяжении всего клеточного цикла. Фактически, чтобы показать, что рост размера ядра не отражает репликацию ДНК, мы показываем, что размер ядра неуклонно увеличивается в клетках, которые задерживаются афидиколином (ингибитором репликации ДНК).
2) Мы подтвердили, что, хотя корреляция между размером ядра и длиной G1 является слабой, она является значимой и высокой воспроизводимостью . Измерения корреляции размера ядра с длиной G1 были независимо повторены по крайней мере шесть раз с очень согласованными результатами. Фактически, повторение того же измерения также встречается в Liu et al., Рукописи, которая была отправлена вместе с этой и получила очень положительные отзывы в eLife .
3) В новое Обсуждение мы добавили раздел, в котором конкретно рассматриваются причины, объясняющие, почему эта корреляция не является сильной.Этот новый раздел начинается с предложения «Хотя корреляция длины клеточного цикла и размера ядра является статистически значимой (p <6,7x10 -8 ) и воспроизводимой (N> 4), остается вопрос, почему эта корреляция мала в величина […] ”
4) Цель измерения корреляции длины G1 и площади ядра — поддержать утверждение о том, что меньшие клетки проводят более длительные периоды времени в G1. С этой целью стоит отметить, что корреляция размера ядра и длины G1 является лишь одним из нескольких доказательств, которые мы предоставляем в этом направлении.Фактически, за исключением рисунка 4, все остальные рисунки в рукописи содержат свидетельства в этом направлении. В частности, на рисунке 1D мы показываем разницу в среднем размере ячеек G1 по сравнению с ячейками S фазы, которые не объясняются возрастом. Также на Рисунках 3 и 6-10 мы показываем, что длина G1 увеличивается из-за фармакологических и генетических нарушений, которые уменьшают размер клетки.
4) Опыты с лекарствами (рисунки 5, 6) очень плохо представлены и объяснены. Я обнаружил, что не могу интерпретировать эти графики и хочу видеть необработанные данные этих экспериментов.Почему за каждое условие выставляется несколько баллов? Где цвет на Рисунке 5D? Каковы единицы «средней скорости роста» на рисунках 5C-F и 6A? Как можно рассчитать продолжительность клеточного цикла для клетки в трех разных точках времени в течение одного клеточного цикла (5E, F)? Как графики подтверждают выводы в тексте? Предлагаю авторам пересмотреть представление этих данных. Однако, даже если он станет более понятным, результаты, полученные с препаратами, следует рассматривать как скорее предварительные, чем окончательные.Всесторонний анализ с использованием одного хорошо охарактеризованного, специфического ингибитора клеточного цикла и одного ингибитора роста может быть более убедительным.
На Рисунке 6 отредактированной рукописи мы показываем новые измерения из «всестороннего анализа с одним хорошо охарактеризованным ингибитором клеточного цикла и одним ингибитором роста». В частности, мы провели измерения размера клеток и количества клеток с течением времени в популяциях, которые лечились SNS032 (хорошо изученным ингибитором клеточного цикла) и рапамицином (хорошо изученным ингибитором роста).Мы используем эти измерения, чтобы продемонстрировать согласованность скорости роста и размера клеток. В частности, мы используем эти два препарата, чтобы показать, что нарушения скорости роста приводят к компенсаторному увеличению длины клеточного цикла и, наоборот, что нарушения длины клеточного цикла приводят к компенсаторным изменениям скорости роста.
Кроме того, для большей ясности в этом эксперименте мы добавили новые рисунки (рисунки 7,8), которые представляют необработанные данные. Наконец, мы изменили текст, чтобы значительно повысить ясность описания этого эксперимента.
5) Компенсация размера, которая представлена для различных клеточных циклов и ингибиторов роста, убедительна, и тот факт, что ингибитор Cdk4 / 6 и ингибитор Hsp90, по-видимому, отменяют компенсацию, поразителен. Компенсация клеточного цикла подтверждается генетически сверхэкспрессией циклина E, но было бы полезно проверить действие ингибитора Cdk4 / 6 и генетически, т.е. не справляется ли мутантная линия сайта фосфорилирования Rb с компенсацией? Это обеспечило бы убедительную связь между контролем размера и одной из наиболее часто мутирующих осей при раке человека.См. Ниже дополнительную информацию о руб.
6) Что касается эксперимента с циклином Е, я обеспокоен тем, что эксперимент, похоже, был проведен путем трансфекции в массе. У этого есть два недостатка, а именно то, что клетки будут подвергаться стрессу в результате процедуры трансфекции, и что уровни белка могут значительно варьироваться от клетки к клетке. Какова была эффективность трансфекции и как долго клетки могли восстанавливаться? Лучшим экспериментом было бы проанализировать 2 или 3 стабильных клона, которые экспрессируют циклин E под индуцибельным промотором, поэтому, возможно, авторы могли бы это рассмотреть.Как предложено выше, подобный эксперимент может быть проведен параллельно с нефосфорилируемым Rb или сверхэкспрессируемым CDK4 / 6 с предсказанием, что гомеостаз размера будет нарушен.
На рисунке 10 в измененной рукописи показаны измерения трех различных стабильных клеточных линий, которые мы создали в соответствии с этими комментариями рецензента:
1) Клеточная линия с индуцируемой доксициклином экспрессией циклина E
2) Клеточная линия с индуцируемой доксициклином экспрессией циклина D
3) Клеточная линия с индуцируемой доксициклином экспрессией p27
Как и ожидалось, сверхэкспрессия циклина E и циклина D, зависящая от доксициклина, приводит к увеличению скорости деления клеток (более короткие клеточные циклы), в то время как сверхэкспрессия p27 приводит к более медленным скоростям деления клеток (более длинным клеточным циклам).В полном соответствии с фармакологическими измерениями уменьшение длины клеточного цикла, вызванное сверхэкспрессией циклина E, компенсируется повышенной скоростью роста, так что размер клеток остается неизменным. Напротив, когда длина клеточного цикла укорачивается из-за сверхэкспрессии циклина D, размер клеток значительно уменьшается.
7) Сила рукописи состоит в том, что аналогичные результаты были получены на сильно злокачественных клетках HeLa и нормальных эпителиальных клетках RPE1 (которые, как я полагаю, не были иммортализованы теломеразой — это правда?).Примечательно, что клетки HeLa сохраняют контроль над размером, несмотря на доказательства широко распространенной анеуплоидии, вариации числа копий, хромотрипсиса и нарушения клеточного цикла и путей регуляции роста (PMID: 23550136). Это заслуживает комментария, особенно в отношении специфических эффектов инсерции HPV18, которые должны были нарушить ось Rb (см. Выше) и инактивировать p53. Было бы некоторым дополнительным утешением с точки зрения универсальности механизма компенсации, если бы авторы могли предоставить подтверждающие данные для третьей клеточной линии, например Rb-линии.Меня озадачивает очевидная неповрежденная компенсация в клетках HeLa, но ее нарушение ингибитором CDK4 / 6. Нокауты членов семейства Rb приводят к тому, что клетки становятся маленькими, и можно было ожидать, что они будут разрушены для одной из точек компенсации. Прокомментируйте, пожалуйста, эти моменты.
В оригинальной рукописи выводы были сделаны на основе измерений на двух клеточных линиях, hTERT-иммортализованных клетках Rpe1 и клетках HeLa. К отредактированной рукописи мы добавили измерения из трех дополнительных клеточных линий: двух клеточных линий, лишенных активности Rb (Rb-нулевые клетки SAOS2 и SV40-иммортализованные клетки 16HBE), и одну клеточную линию с интактной передачей сигналов Rb (U2OS).Измерения на всех клеточных линиях показаны на Рисунке 9 исправленной рукописи. Необработанные данные измерений на новых клеточных линиях показаны в дополнениях к Рисунку 9.
В соответствии с тем, что было показано в оригинальной рукописи, координация скорости роста и длины клеточного цикла наблюдается также в линиях клеток, в которых отсутствует активность Rb. В то время как рисунки 8 и 10 показывают, что генетические и химические нарушения циклина D / Cdk4 вызывают значительные изменения в размере клеток, рисунок 9 показывает, что Rb не является необходимым для координации скорости роста и длины клеточного цикла.В Обсуждении мы посвятили раздел рассмотрению значения этих результатов. Мы предполагаем, что ось Rb / CDK4 может играть роль в определении размера цели, но не в определении размера, и предлагаем эту возможность в качестве основания для будущих исследований.
Дополнительное примечание к следующему комментарию:
«Меня озадачивает очевидная неповрежденная компенсация в клетках HeLa, но ее нарушение ингибитором Cdk4 / 6. Нокауты членов семейства Rb приводят к тому, что клетки становятся маленькими, и, возможно, ожидалось, что они будут разрушены для одной из точек компенсации.Прокомментируйте, пожалуйста, эти моменты ».
В первоначальном исследовании, хотя координация скорости роста и длины клеточного цикла наблюдалась как в Rpe1, так и в HeLa, ее нарушение при ингибировании Cdk4 / 6 пальбоциклибом наблюдалось только в Rpe1. Клетки HeLa были нечувствительны к пальбоциклибу, как и ожидалось в Rb-неактивной клеточной линии. Это было неясно в исходной рукописи и четко прояснено в исправленной рукописи, где результаты обеих клеточных линий представлены бок о бок на Рисунке 9 (вместе с результатами дополнительных клеточных линий, обсуждаемых выше).
8) Ключевой недостающей частью информации является стадия клеточного цикла, на которой дисперсия размеров активно уменьшается (например, на рис. 3G, H, J). Авторы, кажется, намеренно неясны в этом вопросе, который следует прояснить — когда возникают эффекты компенсации размера? Пожалуйста, четко укажите это.
Анализ значения Gcv последовательно определяет два периода уменьшения дисперсии размера ячеек. Один из этих двух периодов совпадает по времени со средним временем перехода G1 / S, а другой совпадает по времени с поздней S-фазой / G2.Как следует из вопроса рецензента, эти данные свидетельствуют о том, что процесс, вызывающий снижение дисперсии, регулируется компонентами часов клеточного цикла. Однако мы сочли преждевременным делать такой вывод в нашей рукописи. Связывание этих падений в вариации с любым из молекулярных процессов, которые определяют клеточный цикл (например, деградация циклина или репликация ДНК), пока невозможно при нынешнем дизайне экспериментов по ряду причин. Во-первых, отметим, что наблюдаемое уменьшение дисперсии размеров ячеек является конечным продуктом процесса, который произошел раньше.В частности, именно кумулятивный эффект регулирования скорости роста в зависимости от размера приведет к возможному уменьшению дисперсии размера клеток, поэтому мы не хотим переоценивать точное время падения дисперсии до того, как будут проведены дальнейшие эксперименты.
Более того, возможно, что наблюдаемые периоды активной коррекции размера клеток на самом деле не запускаются конкретными событиями клеточного цикла, происходящими в эти периоды, а вместо этого совпадают с ними из-за мешающего фактора.Например, падение дисперсии размера клеток (и соответствующее падение корреляции размера / скорости роста, наблюдаемое на траекториях роста отдельных клеток), которое совпадает со средним временем перехода в S-фазу, может отражать механизм коррекции размера, который активируется при то время. Однако также возможно, что этот корректирующий механизм фактически запускается клетками, достигающими определенного размера (например, клетки, превышающие определенный размер, замедляют свой рост), и что, поскольку большинство клеток рождаются маленькими, средняя клетка попадает только в диапазон этот корректирующий механизм ближе к концу G1.Поскольку прогрессирование клеточного цикла и размер клеток сильно коррелированы, трудно определить, запускается ли наблюдаемая нами регуляция скорости роста непосредственно вступлением в S-фазу.
Тот факт, что мы наблюдаем два различных периода регуляции скорости роста в зависимости от размера во время клеточного цикла, является значительным и указывает на зависимое от клеточного цикла изменение режима регуляции размера. (Даже если регулирование напрямую запускается размером, как в приведенном выше примере, наличие двух провалов может указывать на то, что «целевой размер» увеличивается для клеток на более поздних этапах клеточного цикла.) Однако мы не можем предположить, что молекулярные события, ответственные за это изменение, точно совпадают с наблюдаемыми провалами в дисперсии размеров ячеек.
В настоящее время мы разрабатываем несколько подходов, чтобы связать снижение дисперсии размера клеток с конкретными регуляторами клеточного цикла. Мы расширили анализы, описанные в этой рукописи, и оптимизировали их для использования в сочетании с высокопроизводительными фильтрами возмущений. Однако, поскольку нам потребуется гораздо больше времени, прежде чем мы сможем получить значимые результаты, мы надеемся решить этот вопрос в следующей публикации.
[Примечание редакции: до принятия были запрошены дополнительные исправления, как описано ниже.]
Рецензент № 1:
Как и в исходном документе, в этой статье рассматривается, как размер клетки контролируется в клетках культуры ткани. Это очень базовый вопрос, который обсуждается в течение многих лет, с довольно незначительным прогрессом в достижении консенсуса в понимании лежащих в основе механизмов. Здесь проблема решается логически, с использованием высокоточных одноклеточных методов.В статье делается вывод, что размер клеток находится под активным контролем в течение как минимум двух фаз цикла деления в нескольких типах клеток. Кроме того, авт. Показывают, что ингибирование либо Cdk4, либо Hsp90 может нарушать механизмы, которые снижают вариабельность размеров, давая либо очень большие клетки, либо сильно вариабельные клетки, которые дефектны в связи скорости роста и размера с делением клеток. Это важные выводы. Редакция этой интересной статьи заметно улучшена. Он короче, лаконичнее и легче для понимания.Представлено много новых измерений, дополнительно подтверждающих выводы авторов. Тем не менее, документ может быть более четким и кратким, и прогресс в механистическом понимании управления размером по-прежнему остается скромным. Документ по-прежнему представляет собой характеристику процесса, показывая, что размер клеток активно регулируется во время роста, и даются лишь поверхностные подсказки относительно того, как это происходит механически. Более того, эти ключи, а именно то, что ингибитор Cdk4 / 6 Palbociclib и ингибитор Hsp90 Radicocol нарушают регуляцию размера, и не прослеживаются дальше, чем раньше.Как минимум, я ожидаю, что результаты генетической проверки этих хитовых лекарств будут опубликованы в статье. Это важно для подтверждения выводов. В частности, авт. Д. Оценить контроль размера в клетках, лишенных функции генов Cdk4 / 6 и Hsp90, используя нокауты генов с помощью siRNA или делеции. Кроме того, я бы посоветовал авторам и дальше стремиться к более короткой и удобочитаемой рукописи.
Рефери поднимает три очка:
A) Стиль письма и его ясность можно улучшить.
Мы принимаем критику и улучшили ясность отредактированной рукописи.
B) Для исследования будет полезна генетическая проверка влияния CDK4 / 6 и HSP90 на размер клеток.
Мы оценили контроль размера в клетках, лишенных cdk4 / 6, с помощью siRNA, но оставили валидацию HSP90 для будущей работы. В оригинальной рукописи мы представили доказательства, свидетельствующие о влиянии активности CDK4 / 6 на размер клеток. В нашей оригинальной рукописи доказательства связи активности CDK 4/6 с размером клеток получены из двух источников; фармакологическое ингибирование активности CDK 4/6 и генетическое активация активности CDK 4/6.Фармакологическое ингибирование CDK 4/6 привело к заметному увеличению размера клеток. Чтобы дополнительно подтвердить влияние CDK 4/6 на рост клеток, мы провели второй эксперимент, в котором активность CDK4 была активирована, а не химически ингибирована. Наша гипотеза заключалась в том, что, поскольку ингибирование CDK4 / 6 приводит к более крупным клеткам, активация CDK4 / 6 должна приводить к меньшему размеру клеток. Для активации CDK4 / 6 мы использовали линию клеток RPE1 с доксициклин-зависимой экспрессией активатора CDK 4/6, циклина D.Как мы и ожидали, сверхэкспрессия циклина D приводила к значительному уменьшению размера клеток. В этом заключительном раунде комментариев рецензента рефери предложил окончательный контроль: нокаутировать CDK 4/6 и проверить, воспроизводят ли результаты увеличенный размер клеток, наблюдаемый при химическом ингибировании активности CDK. В соответствии с предложением рецензента мы использовали миРНК для подавления CDK4, CDK6 или обоих. Во всех случаях нокдаун CDK 4/6 приводил к увеличению размера клеток (Рисунок 8 — рисунок в приложении 1), повторяя результат, полученный с химическим ингибитором, пальбоциклибом.В заключение, теперь у нас есть три линии доказательств, свидетельствующих о причастности активности CDK 4/6 к фенотипу увеличенного размера клеток. Во-первых, фармакологическое ингибирование CDK 4/6 приводит к увеличению размера клеток. Во-вторых, генетическая активация CDK 4/6 со сверхэкспрессией циклина D приводит к уменьшению размера клеток. В-третьих, генетическое ингибирование CDK 4/6 с помощью siRNA приводит к увеличению размера клеток. Мы полагаем, что это существенно обосновывает влияние CDK 4/6 на размер клеток. Вопрос, который все еще остается открытым: как CDK 4/6 влияет на размер клетки? Поскольку роль CDK 4/6 в регулировании размера не является основной темой в нашем исследовании, мы решили оставить этот вопрос для более позднего исследования.
C) Рефери правильно утверждает, что прогресс в механистическом понимании контроля размеров невелик.
Согласны, но с оговоркой. Настоящая статья является частью более крупного исследования. Механизмы регуляции длины G1 описаны в прилагаемой рукописи (Liu et al.). Кроме того, в более общем плане механизмы можно искать только после описания явления. В этой статье мы описываем явления, которые еще не были продемонстрированы.
https: // doi.org / 10.7554 / eLife.26957.040От чего зависит размер ячейки? | BMC Biology
Раппе М.С., Коннон С.А., Верджин К.Л., Джованнони С.Дж.: Выращивание повсеместно распространенной клады морского бактериопланктона SAR11. Природа. 2002, 418: 630-633. 10.1038 / природа00917.
PubMed Google Scholar
Schulz HN, Jorgensen BB: Большие бактерии. Annu Rev Microbiol. 2001, 55: 105-137. 10.1146 / annurev.micro.55.1.105.
PubMed CAS Google Scholar
Bailey JV, Salman V, Rouse GW, Schulz-Vogt HN, Levin LA, Orphan VJ: Диморфизм в экотипах крупнейших известных бактерий, обитающих в метановых выходах. Исме Дж. 2011, 5: 1926-1935. 10.1038 / ismej.2011.66.
PubMed CAS PubMed Central Google Scholar
Менделл Дж. Э., Клементс К. Д., Чоат Дж. Х., Ангерт Э. Р.: Экстремальная полиплоидия у крупной бактерии. Proc Nat Acad Sci USA. 2008, 105: 6730-6734. 10.1073 / pnas.0707522105.
PubMed CAS PubMed Central Google Scholar
Молодой К.Д .: Селективная ценность формы бактерий. Microbiol Mol Biol Rev.2006, 70: 660-703. 10.1128 / MMBR.00001-06.
PubMed PubMed Central Google Scholar
Совет по космическим исследованиям Национального исследовательского совета: Пределы размеров очень мелких микроорганизмов: Материалы семинара. 1999, Вашингтон, округ Колумбия: National Academic Press
Google Scholar
Beveridge TJ: Бактериальная поверхность: общие соображения по дизайну и функциям. Может J Microbiol. 1988, 34: 363-372. 10.1139 / m88-067.
PubMed CAS Google Scholar
Wagner JK, Setayeshgar S, Sharon LA, Reilly JP, Brun YV: роль поглощения питательных веществ для расширения оболочки бактериальных клеток. Proc Natl Acad Sci USA. 2006, 103: 11772-11777. 10.1073 / pnas.0602047103.
PubMed CAS PubMed Central Google Scholar
Каланетра К.М., Джойе С.Б., Сансери Н.Р., Нельсон, округ Колумбия: Новые вакуолизированные серные бактерии из Мексиканского залива воспроизводятся посредством восстановительного деления в трех измерениях. Environ Microbiol. 2005, 7: 1451-1460. 10.1111 / j.1462-2920.2005.00832.x.
PubMed CAS Google Scholar
Молодой К.Д .: Бактериальная форма. Mol Microbiol. 2003, 49: 571-580.
PubMed CAS Google Scholar
Молодой К.Д .: Форма бактерий: двумерные вопросы и возможности. Annu Rev Microbiol. 2010, 64: 223-240. 10.1146 / annurev.micro.112408.134102.
PubMed CAS PubMed Central Google Scholar
den Blaauwen T, de Pedro MA, Nguyen-Disteche M, Ayala JA: Морфогенез палочковидных саккулусов. FEMS Microbiol Rev.2008, 32: 321-344. 10.1111 / j.1574-6976.2007.00090.x.
PubMed CAS Google Scholar
Марголин В .: Формирование бактериальной клетки. Curr Biol. 2009, 19: R812-R822. 10.1016 / j.cub.2009.06.033.
PubMed CAS PubMed Central Google Scholar
Осмис Н., Кун Дж. Р., Якобс-Вагнер К. Бактериальный цитоскелет: промежуточная филаментоподобная функция в форме клетки. Клетка. 2003, 115: 705-713. 10.1016 / S0092-8674 (03) 00935-8.
PubMed CAS Google Scholar
Багчи С., Томениус Х., Белова Л. М., Аусмис Н.: Промежуточные филаментоподобные белки у бактерий и функция цитоскелета у Streptomyces . Mol Microbiol. 2008, 70: 1037-1050.
PubMed CAS PubMed Central Google Scholar
Ромберг Л., Левин П.А.: Динамика сборки белка деления бактериальных клеток FtsZ: на грани стабильности. Annu Rev Microbiol. 2003, 57: 125-154. 10.1146 / аннурьев.micro.57.012903.074300.
PubMed CAS Google Scholar
Аддиналл С.Г., Голландия B: предок тубулина, FtsZ, рисовальщик, разработчик и движущая сила бактериального цитокинеза. J Mol Biol. 2002, 318: 219-236. 10.1016 / S0022-2836 (02) 00024-4.
PubMed CAS Google Scholar
Грауман П.Л .: Элементы цитоскелета у бактерий. Annu Rev Microbiol.2007, 61: 589-618. 10.1146 / annurev.micro.61.080706.093236.
PubMed CAS Google Scholar
Лоу Дж., Ван ден Энт Ф., Амос Л.А.: Молекулы бактериального цитоскелета. Annu Rev Biophys Biomol Struct. 2004, 33: 177-198. 10.1146 / annurev.biophys.33.110502.132647.
PubMed Google Scholar
Ингерсон-Махар М., Гитаи З .: Растущая семья: расширяющаяся вселенная бактериального цитоскелета.FEMS Microbiol Rev.2012, 36: 256-266. 10.1111 / j.1574-6976.2011.00316.x.
PubMed CAS PubMed Central Google Scholar
Барри Р.М., Гитаи З .: Самособирающиеся ферменты и происхождение цитоскелета. Curr Opin Microbiol. 2011, 14: 704-711. 10.1016 / j.mib.2011.09.015.
PubMed CAS PubMed Central Google Scholar
Ротфилд Л., Тагбалаут А., Ши Ю.Л .: Пространственный контроль размещения бактериального деления на участке.Nat Rev Microbiol. 2005, 3: 959-968. 10.1038 / nrmicro1290.
PubMed CAS Google Scholar
Lutkenhaus J: Минимальные колебания у бактерий. Adv Exp Med Biol. 2008, 641: 49-61.
PubMed CAS Google Scholar
Wu LJ, Errington J: Окклюзия нуклеоидов и деление бактериальных клеток. Nat Rev Microbiol. 2012, 10: 8-12.
CAS Google Scholar
Чиен А.С., Хилл Н.С., Левин П.А.: Контроль размера клеток у бактерий. Curr Biol. 2012, 22: R340-349. 10.1016 / j.cub.2012.02.032.
PubMed CAS PubMed Central Google Scholar
Weart RB, Lee AH, Chien AC, Haeusser DP, Hill NS, Levin PA: метаболический датчик, определяющий размер клеток у бактерий. Клетка. 2007, 130: 335-347. 10.1016 / j.cell.2007.05.043.
PubMed CAS PubMed Central Google Scholar
van Teeffelen S, Wang S, Furchtgott L, Huang KC, Wingreen NS, Shaevitz JW, Gitai Z: бактериальный актин MreB вращается, и вращение зависит от сборки клеточной стенки. Proc Natl Acad Sci USA. 2011, 108: 15822-15827. 10.1073 / pnas.1108999108.
PubMed CAS PubMed Central Google Scholar
Гарнер ЕС, Бернард Р., Ван В., Чжуанг X, Руднер Д.З., Митчисон Т. Сопряженные круговые движения аппарата синтеза клеточной стенки и филаментов MreB в B.subtilis . Наука. 2011, 333: 222-225. 10.1126 / science.1203285.
PubMed CAS PubMed Central Google Scholar
Домингес-Эскобар Дж., Частанет А., Кревенна А.Х., Фромион В., Ведлих-Зольднер Р., Карбаллидо-Лопес Р.: Процессивное движение MreB-ассоциированных биосинтетических комплексов клеточной стенки у бактерий. Наука. 2011, 333: 225-228. 10.1126 / science.1203466.
PubMed CAS Google Scholar
Conlon I, Raff M: Различия в том, как клетки млекопитающих и клетки дрожжей координируют рост клеток и развитие клеточного цикла. J Biol. 2003, 2: 7-10.1186 / 1475-4924-2-7.
PubMed PubMed Central Google Scholar
Turner JJ, Ewald JC, Skotheim JM: Контроль размера клеток в дрожжах. Curr Biol. 2012, 22: R350-359. 10.1016 / j.cub.2012.02.041.
PubMed CAS PubMed Central Google Scholar
Mitchison Creanor: Индукционная синхронность в делящихся дрожжах Schizosaccharomyces pombe. Exp Cell Res. 1971, 67: 368-374. 10.1016 / 0014-4827 (71)
PubMed CAS Google Scholar
Джонстон, Прингл, Хартвелл: Координация роста с делением клеток у дрожжей saccharomyces cerevisiae. Exp Cell Res. 1977, 105: 79-98. 10.1016 / 0014-4827 (77)
-9.PubMed Google Scholar
Эллиотт С., Маклафлин С. Скорость синтеза макромолекул через клеточный цикл дрожжей Saccharomyces cerevisiae. Proc Natl Acad Sci USA. 1978, 75: 4384-4388. 10.1073 / pnas.75.9.4384.
PubMed CAS PubMed Central Google Scholar
Годин М., Дельгадо Ф.Ф., Сон С., Гровер У.Х., Брайан А.К., Цур А., Йоргенсен П., Пайер К., Гроссман А.Д., Киршнер М.В., Маналис С.Р .: Использование плавучей массы для измерения роста отдельных клеток.Нат методы. 2010, 7: 387-390. 10.1038 / nmeth.1452.
PubMed CAS PubMed Central Google Scholar
Креанор Дж., Митчисон Дж. М.: Паттерны синтеза белка во время клеточного цикла делящихся дрожжей Schizosaccharomyces pombe. J Cell Sci. 1982, 58: 263-285.
PubMed CAS Google Scholar
Nurse P, Thuraux P, Nasmyth K: Генетический контроль цикла деления клеток у делящихся дрожжей Schizosaccharomyces pombe.Molec Gen Genet. 1976, 146: 167-178. 10.1007 / BF00268085.
PubMed CAS Google Scholar
Фантес П., медсестра П.: Контроль размера клеток при делении у делящихся дрожжей с помощью контроля размера с модулированным ростом при ядерном делении. Exp Cell Res. 1977, 107: 377-386. 10.1016 / 0014-4827 (77)
PubMed CAS Google Scholar
Медсестра П: Генетический контроль размера клеток при делении клеток у дрожжей.Природа. 1975, 256: 547-551. 10.1038 / 256547a0.
PubMed CAS Google Scholar
Nurse P, Thuraux P: контролирует время репликации ДНК во время клеточного цикла делящихся дрожжей. Exp Cell Res. 1977, 107: 365-375. 10.1016 / 0014-4827 (77)
-5.PubMed CAS Google Scholar
Hartwell LH: клеточный цикл Saccharomyces cerevisiae. Bacteriol Rev.1974, 38: 164-198.
PubMed CAS PubMed Central Google Scholar
Медсестра P: Универсальный механизм управления, регулирующий начало M-фазы. Природа. 1990, 344: 503-508. 10.1038 / 344503a0.
PubMed CAS Google Scholar
Рассел П., медсестра П.: cdc25 + действует как индуктор в митотическом контроле делящихся дрожжей. Клетка. 1986, 45: 145-153.10.1016 / 0092-8674 (86)
-5.PubMed CAS Google Scholar
Рассел П., Медсестра П.: Отрицательная регуляция митоза с помощью wee1 +, гена, кодирующего гомолог протеинкиназы. Клетка. 1987, 49: 559-567. 10.1016 / 0092-8674 (87)
PubMed CAS Google Scholar
Гулд Л., медсестра П. Фосфорилирование тирозина протеинкиназы cdc2 + делящихся дрожжей регулирует вступление в митоз.Природа. 1989, 342: 39-45. 10.1038 / 342039a0.
PubMed CAS Google Scholar
Лундгрен К., Уолворт Н., Бухер Р., Дембски М., Киршнер М., Бич Д.: mik1 и wee1 взаимодействуют в ингибирующем фосфорилировании тирозина cdc2. Клетка. 1991, 64: 1111-1122. 10.1016 / 0092-8674 (91)
-2.PubMed CAS Google Scholar
Moseley JB, Mayeux A, Paoletti A, Nurse P: Пространственный градиент координирует размер клетки и митотический вход в делящихся дрожжах.Природа. 2009, 459: 857-860. 10.1038 / природа08074.
PubMed CAS Google Scholar
Мартин С.Г., Бертело-Грожан М: Полярные градиенты киназы Pom1 семейства DYRK связывают длину клетки с клеточным циклом. Природа. 2009, 459: 852-856. 10.1038 / природа08054.
PubMed CAS Google Scholar
Hachet O, Berthelot-Grosjean M, Kokkoris K, Vincenzetti V, Moosbrugger J, Martin SG: цикл фосфорилирования формирует градиенты киназы Pom1 семейства DYRK на плазматической мембране.Клетка. 2011, 145: 1116-1128. 10.1016 / j.cell.2011.05.014.
PubMed CAS Google Scholar
Кудрез Д., медсестра П.: Управление клеточным циклом с минимальной сетью управления CDK. Природа. 2010, 468: 1074-1079. 10.1038 / природа09543.
PubMed CAS Google Scholar
Futcher B: Циклины и структура цикла дрожжевых клеток. Дрожжи. 1996, 12: 1635-1646.10.1002 / (SICI) 1097-0061 (199612) 12:16 <1635 :: AID-YEA83> 3.0.CO; 2-O.
PubMed CAS Google Scholar
Rupes I: Проверка размера клеток дрожжей. Тенденции Genet. 2002, 18: 479-485. 10.1016 / S0168-9525 (02) 02745-2.
PubMed CAS Google Scholar
Nash R, Tokiwa G, Sukhuijt A, Erickson K, Futcher B: Ген WHI1 + Saccharomyces cerevisiae связывает деление клетки с размером клетки и является гомологом циклина.EMBO J. 1988, 7: 4335-4346.
PubMed CAS PubMed Central Google Scholar
Перекрестный FR: DAF1, мутантный ген, влияющий на контроль размера, арест феромона и кинетику клеточного цикла Saccharomyces cerevisiae. Mol Cell Biol. 1988, 8: 4675-4684.
PubMed CAS PubMed Central Google Scholar
Тайерс М., Токива Г., Футчер Б. Сравнение циклинов G1 Saccharomyces cerevisiae: Cln3 может быть активатором выше по течению для Cln1, Cln2 и других циклинов.EMBO J. 1993, 12: 1955-1968.
PubMed CAS PubMed Central Google Scholar
Тайерс М., Токивал Г., Нэш Р., Футчер Б. Киназный комплекс Cln3 — Cdc28 S.cerevisiae регулируется протеолизом и фосфорилированием. EMBO J. 1992, 11: 1773-1784.
PubMed CAS PubMed Central Google Scholar
Полименис М., Шмидт Э.В.: Связь деления клеток с ростом клеток с помощью трансляционного контроля циклина CLN3 G1 в дрожжах.Genes Dev. 1997, 11: 2522-2531. 10.1101 / gad.11.19.2522.
PubMed CAS PubMed Central Google Scholar
de Bruin RA, McDonald WH, Kalashnikova TI, Yates J, Wittenberg C: Cln3 активирует G1-специфическую транскрипцию посредством фосфорилирования репрессора Whi5, связанного с SBF. Клетка. 2004, 117: 887-898. 10.1016 / j.cell.2004.05.025.
PubMed CAS Google Scholar
Костанцо М., Нисикава Дж. Л., Тан X, Миллман Дж. С., Шуб О., Брейткройц К., Дьюар Д., Рупес I, Эндрюс Б., Тайерс М.: активность CDK противодействует Whi5, ингибитору транскрипции G1 / S в дрожжах. Клетка. 2004, 117: 899-913.
PubMed CAS Google Scholar
Нейман Ф.Р., медсестра П.: Контроль размера ядер у делящихся дрожжей. J Cell Biol. 2007, 179: 593-600. 10.1083 / jcb.200708054.
PubMed CAS PubMed Central Google Scholar
Йоргенсен П., Эджингтон Н. П., Шнайдер Б. Л., Рупес I, Тайерс М., Футчер Б. Размер ядра увеличивается по мере роста дрожжевых клеток. Mol Biol Cell. 2007, 18: 3523-3532. 10.1091 / mbc.E06-10-0973.
PubMed CAS PubMed Central Google Scholar
Медсестра П: Генетический контроль объема клеток. Эволюция размера генома. Отредактировано: Cavalier-Smith T. 1985, Chichester: Wiley, 185-196.63.
Google Scholar
Грегори TR: Совпадение, коэволюция или причинно-следственная связь? Содержание ДНК, размер клеток и загадка значения C. Биол Рев Камб Филос Соц. 2001, 76: 65-101. 10.1017 / S1464793100005595.
PubMed CAS Google Scholar
Мортимер РК: Радиобиологические и генетические исследования полиплоидного ряда (от гаплоидного до гексаплоидного) Saccharomyces cerevisiae. Radiation Res. 1958, 9: 312-326. 10.2307 / 3570795.
PubMed CAS Google Scholar
Fantes PA, Grant WD, Pritchard RH, Sudbery PE, Wheals AE: Регулирование размера клеток и контроль митоза. J Theor Biol. 1975, 50: 213-244. 10.1016 / 0022-5193 (75)
-Х.
PubMed CAS Google Scholar
Donachie WD: Взаимосвязь между размером клетки и временем инициации репликации ДНК. Природа. 1968, 219: 1077-1079. 10.1038 / 21
a0.
PubMed CAS Google Scholar
Wang H, Carey LB, Cai Y, Wijnen H, Futcher B: Привлечение циклина Cln3 к промоторам контролирует вход в клеточный цикл через гистондеацетилазу и другие мишени. PLoS Biol. 2009, 7: e1000189-10.1371 / journal.pbio.1000189.
PubMed PubMed Central Google Scholar
Хара Y, Кимура A: Зависимый от размера клетки контроль размеров органелл во время развития. Результаты Пробл Ячейки Различаются. 2011, 53: 93-108. 10.1007 / 978-3-642-19065-0_5.
PubMed CAS Google Scholar
Hara Y, Kimura A: Зависимое от размера клетки удлинение веретена у ранних эмбрионов Caenorhabditis elegans. Curr Biol. 2009, 19: 1549-1554. 10.1016 / j.cub.2009.07.050.
PubMed CAS Google Scholar
Карвалью А., Десаи А., Оегема К. Структурная память в сократительном кольце делает продолжительность цитокинеза независимой от размера клетки.Клетка. 2009, 137: 926-937. 10.1016 / j.cell.2009.03.021.
PubMed CAS Google Scholar
Greenan G, Brangwynne CP, Jaensch S, Gharakhani J, Julicher F, Hyman AA: Размер центросомы определяет длину митотического веретена у эмбрионов Caenorhabditis elegans. Curr Biol. 2010, 20: 353-358. 10.1016 / j.cub.2009.12.050.
PubMed CAS Google Scholar
Siller KH, Doe CQ: Ориентация веретена во время асимметричного деления клеток.Nat Cell Biol. 2009, 11: 365-374. 10.1038 / ncb0409-365.
PubMed CAS Google Scholar
Kellogg DR, Moritz M, Alberts BM: Центросома и клеточная организация. Анну Рев Биохим. 1994, 63: 639-674. 10.1146 / annurev.bi.63.070194.003231.
PubMed CAS Google Scholar
Grill SW, Hyman AA: Позиционирование шпинделя кортикальными тянущими силами.Dev Cell. 2005, 8: 461-465. 10.1016 / j.devcel.2005.03.014.
PubMed CAS Google Scholar
Минк Н., Берджесс Д., Чанг Ф .: Влияние геометрии ячейки на позиционирование в плоскости деления. Клетка. 2011, 144: 414-426. 10.1016 / j.cell.2011.01.016.
PubMed CAS PubMed Central Google Scholar
Strome S, Wood WB: Генерация асимметрии и сегрегации гранул зародышевой линии у ранних C.elegans эмбрионы. Клетка. 1983, 35: 15-25. 10.1016 / 0092-8674 (83)
-9.
PubMed CAS Google Scholar
Reinsch S, Gönczy P: Механизмы ядерного позиционирования. J Cell Sci. 1998, 111: 2283-2295.
PubMed CAS Google Scholar
Хамагучи М.С., Хирамото Y: Анализ роли астральных лучей в пронуклеарной миграции в яйцах песочного доллара с помощью метода колцемид-УФ.Разница в росте разработчиков. 1986, 28: 143-156. 10.1111 / j.1440-169X.1986.00143.x.
Google Scholar
Holy TE, Dogterom M, Yurke B, Leibler S: Сборка и размещение звездочек микротрубочек в камерах микротрубочки. Proc Natl Acad Sci USA. 1997, 94: 6228-6231. 10.1073 / pnas.94.12.6228.
PubMed CAS PubMed Central Google Scholar
Тран П.Т., Марш Л., Дой В., Иноуэ С., Чанг Ф .: Механизм ядерного позиционирования у делящихся дрожжей, основанный на толкании микротрубочек.J Cell Biol. 2001, 153: 397-411. 10.1083 / jcb.153.2.397.
PubMed CAS PubMed Central Google Scholar
Kimura A, Onami S: Компьютерное моделирование и обработка изображений показывают, что тянущая сила, зависящая от длины, является основным механизмом миграции пронуклеусов самцов C. elegans. Dev Cell. 2005, 8: 765-775. 10.1016 / j.devcel.2005.03.007.
PubMed CAS Google Scholar
Vallee RB, Stehman SA: Как динеин помогает клетке найти свой центр: сервомеханическая модель. Trends Cell Biol. 2005, 15: 288-294. 10.1016 / j.tcb.2005.04.005.
PubMed CAS Google Scholar
Догтером М., Керсемакерс Дж. В., Ромет-Лемонн Г., Янсон М. Е.: Генерация силы динамическими микротрубочками. Curr Opin Cell Biol. 2005, 17: 67-74. 10.1016 / j.ceb.2004.12.011.
PubMed CAS Google Scholar
Кимура А., Онами С. Моделирование сил, опосредованных микротрубочками, и позиционирования центросом у эмбрионов Caenorhabditis elegans. Методы Cell Biol. 2010, 97: 437-453.
PubMed CAS Google Scholar
Чжу Дж., Бураков А., Родионов В., Могильнер А. Обнаружение клеточного центра путем баланса вытягивания динеина и миозина и выталкивания микротрубочек: вычислительное исследование. Mol Biol Cell. 2010, 21: 4418-4427. 10.1091 / mbc.E10-07-0627.
PubMed CAS PubMed Central Google Scholar
Кимура К., Кимура А. Внутриклеточные органеллы опосредуют силу притяжения цитоплазмы для центросомного центрирования у ранних эмбрионов Caenorhabditis elegans. Proc Natl Acad Sci USA. 2011, 108: 137-142. 10.1073 / pnas.1013275108.
PubMed CAS PubMed Central Google Scholar
Кимура К., Кимура А: Новый механизм силы, зависящей от длины микротрубочек, которая притягивает центросомы к центру клетки. Биоархитектура. 2011, 1: 74-79.10.4161 / bioa.1.2.15549.
PubMed PubMed Central Google Scholar
Laan L, Pavin N, Husson J, Romet-Lemonne G, van Duijn M, Lopez MP, Vale RD, Julicher F, Reck-Peterson SL, Dogterom M: кортикальный динеин контролирует динамику микротрубочек для создания тянущих сил это положение звездочек микротрубочек. Клетка. 2012, 148: 502-514. 10.1016 / j.cell.2012.01.007.
PubMed CAS PubMed Central Google Scholar
Wühr M, Tan ES, Parker SK, Detrich HW, Mitchison TJ: Модель для определения плоскости дробления у ранних эмбрионов амфибий и рыб. Curr Biol. 2010, 20: 2040-2045. 10.1016 / j.cub.2010.10.024.
PubMed PubMed Central Google Scholar
Митчисон Т, Киршнер М: Динамическая нестабильность роста микротрубочек. Природа. 1984, 312: 237-242. 10.1038 / 312237a0.
PubMed CAS Google Scholar
Ховард Дж: Механика моторных белков и цитоскелета. 2001, Массачусетс: Sinauer Associates
Google Scholar
Уокер Р.А., О’Брайен Е.Т., Прайер Н.К., Собойро М.Ф., Воттер В.А., Эриксон Х.П., Салмон ED: динамическая нестабильность отдельных микротрубочек, проанализированная с помощью световой микроскопии: константы скорости и частоты переходов. J Cell Biol. 1988, 107: 1437-1448. 10.1083 / jcb.107.4.1437.
PubMed CAS Google Scholar
Cassimeris L, Pryer NK, Salmon ED: Наблюдения в реальном времени за динамической нестабильностью микротрубочек в живых клетках. J Cell Biol. 1988, 107: 2223-2231. 10.1083 / jcb.107.6.2223.
PubMed CAS Google Scholar
Soltys BJ, Borisy GG: Полимеризация тубулина in vivo: прямые доказательства сборки на концах микротрубочек и из центросом. J Cell Biol. 1985, 100: 1682-1689. 10.1083 / jcb.100.5.1682.
PubMed CAS Google Scholar
Альбертсон Д.Г.: Формирование первого веретена дробления у эмбрионов нематод. Dev Biol. 1984, 101: 61-72. 10.1016 / 0012-1606 (84)-9.
PubMed CAS Google Scholar
Gönczy P, Bellanger JM, Kirkham M, Pozniakowski A, Baumer K, Phillips JB, Hyman AA: zyg-8, ген, необходимый для позиционирования веретена у C. elegans, кодирует родственную даблкортину киназу, которая способствует сборка микротрубочек. Dev Cell. 2001, 1: 363-375.10.1016 / S1534-5807 (01) 00046-6.
PubMed Google Scholar
Wühr M, Dumont S, Groen AC, Needleman DJ, Mitchison TJ: Как клетка миллиметрового размера находит свой центр ?. Клеточный цикл. 2009, 8: 1115-1121. 10.4161 / cc.8.8.8150.
PubMed PubMed Central Google Scholar
Догтером М., Юрке Б. Динамика микротрубочек и расположение центров организации микротрубочек.Phys Rev Lett. 1998, 81: 485-488. 10.1103 / PhysRevLett.81.485.
CAS Google Scholar
Файвре-Москаленко С., Догтером М: Динамика астры микротрубочек в микрокамерах: роль катастроф. Proc Natl Acad Sci USA. 2002, 99: 16788-16793. 10.1073 / pnas.252407099.
PubMed CAS PubMed Central Google Scholar
Neurohr G, Naegeli A, Titos I, Theler D, Greber B, Diez J, Gabaldon T, Mendoza M, Barral Y: линейка на основе средней зоны регулирует уплотнение хромосом в соответствии с длиной веретена анафазы.Наука. 2011, 332: 465-468. 10.1126 / science.1201578.
PubMed CAS Google Scholar
Ladouceur AM, Ranjan R, Maddox PS: Размер клетки: хромосомы подвергаются ударам правителя средней зоны. Curr Biol. 2011, 21: R388-390. 10.1016 / j.cub.2011.04.009.
PubMed CAS Google Scholar
Grill SW, Gönczy P, Stelzer EH, Hyman AA: Полярность контролирует силы, управляющие асимметричным расположением веретена у эмбриона Caenorhabditis elegans.Природа. 2001, 409: 630-633. 10.1038 / 35054572.
PubMed CAS Google Scholar
Могилнер А., Уоллман Р., Чивелекоглу-Шоли Г., Шоли Дж .: Моделирование митоза. Trends Cell Biol. 2006, 16: 88-96. 10.1016 / j.tcb.2005.12.007.
PubMed CAS Google Scholar
Уилсон Э.Б.: Клетка в развитии и наследственности. 1934, Нью-Йорк: MacMillan
Google Scholar
Hämmerling J: нуклео-цитоплазматические взаимодействия в Acetabularia и других клетках. Annu Rev Plant Physiol. 1963, 14: 65-92. 10.1146 / annurev.pp.14.060163.000433.
Google Scholar
Мац М.В., Франк Т.М., Маршалл, штат Нью-Джерси, Виддер Э.А., Йонсен С.: Гигантский глубоководный протист оставляет следы, похожие на билатерианские. Curr Biol. 2008, 18: 1849-1854. 10.1016 / j.cub.2008.10.028.
PubMed CAS Google Scholar
Джейкобс В.П .: Каулерпа. Sci Am. 1994, 271: 100-105.
Google Scholar
Арнольд З.М.: Наблюдения за половым поколением Gromia oviformis Dujardin. J Protozool. 1966, 13: 23-27.
PubMed CAS Google Scholar
Кроули Дж .: Некоторые наблюдения за тонкой структурой гамет и зигот Acetabularia. Planta. 1966, 69: 365-376.10.1007 / BF00392287.
PubMed CAS Google Scholar
Гольдштейн М., Морралл С. Гаметогенез и оплодотворение у каулерпы. Ann N Y Acad Sci. 1970, 175: 660-672. 10.1111 / j.1749-6632.1970.tb45183.x.
Google Scholar
Смирнов А., Насонова Е., Берни С., Фарни Дж., Боливар И., Павловски Дж .: Молекулярная филогения и классификация лобозных амеб.Протист. 2005, 156: 129-142. 10.1016 / j.protis.2005.06.002.
PubMed CAS Google Scholar
Роджерсон А., Паттерсон Д.Д.: Голые рамикристатные амебы (Gymnamoebae). Иллюстрированный путеводитель по простейшим. 2002, Лоуренс, Канзас: Общество протозоологов, 1023-1053. 2
Google Scholar
Chalkley H: Наблюдение за митозом в живой клетке Amoeba proteus.Наука. 1934, 80: 208-209. 10.1126 / science.80.2070.208.
PubMed CAS Google Scholar
Кудо R: Pelomyxa carolinensis Wilson. II. Ядерное деление и плазмотомия. J Morphol. 1947, 80: 93-143. 10.1002 / jmor.1050800105.
PubMed CAS Google Scholar
Кудо R: Pelomyxa carolinensis Wilson. III. Дальнейшие наблюдения по поводу плазмотомии.J Morphol. 1949, 85: 163-176. 10.1002 / jmor.1050850107.
PubMed CAS Google Scholar
Roeder AH, Cunha A, Ohno CK, Meyerowitz EM: Клеточный цикл регулирует тип клеток в чашелистике Arabidopsis. Разработка. 2012, 139: 4416-4427. 10.1242 / dev.082925.
PubMed CAS Google Scholar
Morgan TH: Регенерация пропорциональных структур в Stentor.Biol Bull. 1901, 2: 311-328. 10.2307 / 1535709.
Google Scholar
Tartar V: Биология Stentor. 1961, Оксфорд: Pergamon Press
Google Scholar
Tartar V: Реакция Stentor coeruleus на гомопластическую трансплантацию. J Exp Zool. 1954, 127: 511-575. 10.1002 / jez.1401270306.
Google Scholar
Tartar V: Восстановление измельченного Stentor coeruleus. J Exp Zool. 1960, 144: 187-207. 10.1002 / jez.1401440208.
PubMed CAS Google Scholar
Uhlig G: Entwicklungsphysiologische Untersuchungen zur Morphogenese von Stentor coeruleus Ehrbg. Арка Протистентенк. 1960, 105: 1-109.
Google Scholar
Tartar V: Морфогенез в гомополярном тандемном трансплантате Stentor coeruleus.J Exp Zool. 1964, 156: 243-251. 10.1002 / jez.1401560302.
PubMed CAS Google Scholar
Элинсон Р.П. Оплодотворение и водное развитие пуэрториканской наземной лягушки Eleutherodactylus coqui. J Morphol. 1987, 193: 217-224. 10.1002 / jmor.1051930208.
Google Scholar
Collazo A, Keller R: Раннее развитие Ensatina eschscholtzii: земноводное с большим желточным яйцом.EvoDevo. 2010, 1: 6-10.1186 / 2041-9139-1-6.
PubMed PubMed Central Google Scholar
Элинсон Р.П., дель Пино Е.М.: Разнообразие развития земноводных. Wiley Interdiscip Rev Membr Transp Signal. 2012, 1: 345-369.
CAS Google Scholar
Саммерс К., Си МакКеон К., Хейинг Х .: Эволюция родительской заботы и размер яиц: сравнительный анализ у лягушек.Proc Biol Sci. 2006, 273: 687-692. 10.1098 / rspb.2005.3368.
PubMed PubMed Central Google Scholar
дель Пино Е.М., Лоор-Вела S: Модель раннего дробления сумчатой лягушки Gastrotheca riobambae. Разработка. 1990, 110: 781-789.
PubMed CAS Google Scholar
Gould SJ: Ограничение развития в Cerion, с комментариями по определению и интерпретации ограничения в эволюции.Эволюция. 1989, 43: 516-539. 10.2307 / 2409056.
Google Scholar
Jenkins R: Тонкая структура деления у инфузорий простейших I. Микроядерный митоз у Blepharisma. J Cell Biol. 1967, 34: 463-481. 10.1083 / jcb.34.2.463.
PubMed CAS PubMed Central Google Scholar
Tartar V: Микрохирургические эксперименты по цитокинезу в Stentor coeruleus.J Exp Zool. 1968, 167: 21-35. 10.1002 / jez.1401670103.
PubMed CAS Google Scholar
Гилберт С: Биология развития. 2006, Сандерленд, Массачусетс: Sinauer Associates, 8
Google Scholar
Su TT, O’Farrell PH: Контроль размера: пролиферация клеток не равна росту. Curr Biol. 1998, 8: R687-689. 10.1016 / S0960-9822 (98) 70436-1.
PubMed CAS PubMed Central Google Scholar
Kozma SC, Thomas G: Регулирование размера клеток при росте, развитии и заболеваниях человека: PI3K, PKB и S6K. Биологические исследования. 2002, 24: 65-71. 10.1002 / bies.10031.
PubMed CAS Google Scholar
Fankhauser G: Поддержание нормальной структуры гетероплоидных личинок саламандры путем компенсации изменений в размере клеток путем корректировки количества и формы клеток. J Exp Zool. 1945, 100: 445-455. 10.1002 / jez.1401000310.
PubMed CAS Google Scholar
Nieuwkoop PD, Faber J: Нормальный стол Xenopus laevis (Daudin). 1994, Нью-Йорк: Гарленд, 3
Google Scholar
Данильчик М.В., Бедрик С.Д., Браун Е.Е., Луч К. Микротрубочки борозды и локализованный экзоцитоз при расщеплении эмбрионов Xenopus laevis. J Cell Sci. 2003, 116: 273-283. 10.1242 / jcs.00217.
PubMed CAS Google Scholar
Wuhr M, Chen Y, Dumont S, Groen AC, Needleman DJ, Salic A, Mitchison TJ: Доказательства верхнего предела длины митотического веретена. Curr Biol. 2008, 18: 1256-1261. 10.1016 / j.cub.2008.07.092.
PubMed PubMed Central Google Scholar
Ван П., Хайден С., Масуи Ю.: Переход клеточного цикла бластомера от независимого от размера клетки к зависимому от размера контролю на стадии средней бластулы у Xenopus laevis. J Exp Zool. 2000, 287: 128-144.10.1002 / 1097-010X (20000701) 287: 2 <128 :: AID-JEZ3> 3.0.CO; 2-G.
PubMed CAS Google Scholar
Ньюпорт Дж., Киршнер М: Главный переход в развитии ранних эмбрионов Xenopus: II. Контроль начала транскрипции. Клетка. 1982, 30: 687-696. 10.1016 / 0092-8674 (82)
PubMed CAS Google Scholar
Ньюпорт Дж., Киршнер М: Главный переход в развитии ранних эмбрионов Xenopus: I.характеристика и сроки клеточных изменений на стадии средней бластулы. Клетка. 1982, 30: 675-686. 10.1016 / 0092-8674 (82)
-0.PubMed CAS Google Scholar
Леви Д.Л., Хилд Р.: Размер ядра регулируется импортином альфа и Ntf2 в Xenopus. Клетка. 2010, 143: 288-298. 10.1016 / j.cell.2010.09.012.
PubMed CAS PubMed Central Google Scholar
Brown KS, Blower MD, Maresca TJ, Grammer TC, Harland RM, Heald R: экстракты яиц Xenopus tropicalis дают представление о масштабировании митотического веретена. J Cell Biol. 2007, 176: 765-770. 10.1083 / jcb.200610043.
PubMed CAS PubMed Central Google Scholar
Woolner S, O’Brien LL, Wiese C, Bement WM: филаменты миозина-10 и актина необходимы для функции митотического веретена. J Cell Biol. 2008, 182: 77-88. 10.1083 / jcb.200804062.
PubMed CAS PubMed Central Google Scholar
Loughlin R, Wilbur JD, McNally FJ, Nedelec FJ, Heald R: Катанин вносит вклад в межвидовое масштабирование длины веретена у Xenopus. Клетка. 2011, 147: 1397-1407. 10.1016 / j.cell.2011.11.014.
PubMed CAS PubMed Central Google Scholar
Lecuit T, Lenne PF: Механика клеточной поверхности и контроль формы клеток, структуры тканей и морфогенеза.Nat Rev Mol Cell Biol. 2007, 8: 633-644.
PubMed CAS Google Scholar
Келлер Р: Биология развития. Физическая биология возвращается к морфогенезу. Наука. 2012, 338: 201-203. 10.1126 / science.1230718.
PubMed CAS Google Scholar
Fine LG, Norman J: Клеточные события при почечной гипертрофии. Annu Rev Physiol. 1989, 51: 19-32.10.1146 / annurev.ph.51.030189.000315.
PubMed CAS Google Scholar
Лиенкамп С., Лю К., Карнер С., Кэрролл Т., Ронненбергер О., Уоллингфорд Дж. Б., Уолц Г.: Почечные канальцы удлиняются с использованием нового способа конвергентного расширения, зависящего от полярности плоских клеток. Нат Жене. 2012.
Google Scholar
Karner CM, Chirumamilla R, Aoki S, Igarashi P, Wallingford JB, Carroll TJ: передача сигналов Wnt9b регулирует плоскую полярность клеток и морфогенез почечных канальцев.Нат Жене. 2009, 41: 793-799. 10,1038 / нг.400.
PubMed CAS PubMed Central Google Scholar
Happe H, van der Wal AM, Leonhard WN, Kunnen SJ, Breuning MH, de Heer E, Peters DJ: Измененная передача сигналов Hippo при поликистозе почек. J Pathol. 2011, 224: 133-142. 10.1002 / путь.2856.
PubMed CAS Google Scholar
Макита Р., Учидзима Ю., Нишияма К., Амано Т., Чен К., Такеучи Т., Митани А., Нагасе Т., Ятоми И., Абуратани Н., Накагава О., Смолл Е. В., Кобо-Старк П., Игараси П., Мураками М., Томинага Дж., Сато Т., Асано Т., Курихара Ю., Курихара Х .: Множественные кисты почек, дефекты концентрации мочи и эмфизематозные изменения легких у мышей, лишенных ТАЗ.Am J Physiol Renal Physiol. 2008, 294: F542-553. 10.1152 / айпренал.00201.2007.
PubMed CAS Google Scholar
Chen JK, Chen J, Neilson EG, Harris RC: Роль млекопитающих-мишеней передачи сигналов рапамицина в компенсаторной почечной гипертрофии. J Am Soc Nephrol. 2005, 16: 1384-1391. 10.1681 / ASN.2004100894.
PubMed CAS Google Scholar
Chen JK, Chen J, Thomas G, Kozma SC, Harris RC: Нокаут киназы 1 S6 ингибирует гипертрофию почек, вызванную унефрэктомией или диабетом.Am J Physiol Renal Physiol. 2009, 297: F585-593. 10.1152 / айпренал.00186.2009.
PubMed CAS PubMed Central Google Scholar
Прайс Х, Воробей А., Науман А.Ф.: Корреляция между объемом ядра, объемом клетки и содержанием ДНК в меристематических клетках травянистых покрытосеменных растений. Cell Mol Life Sci. 1973, 29: 1028-1029. 10.1007 / BF01930444.
CAS Google Scholar
Haber A, Foard D: Анатомический анализ роста пшеницы без деления клеток. Am J Bot. 1961, 4: 438-446.
Google Scholar
Haber AH, Carrier WL, Foard DE: Метаболические исследования гамма-облученной пшеницы, выращиваемой без деления клеток. Am J Bot. 1961, 48: 431-438. 10.2307 / 2439444.
CAS Google Scholar
Haber AH, Foard DE: Дальнейшие исследования гамма-облученной пшеницы и их значение для использования митотического ингибирования для исследований развития.Am J Bot. 1964, 51: 151-159. 10.2307 / 2440099.
CAS Google Scholar
Фоард Д.Е., Хабер А.Х., Фишман Т.Н.: Инициирование зачатков боковых корней без завершения митоза и без цитокинеза в однорядном перицикле. Am J Bot. 1965, 52: 580-590. 10.2307 / 2440119.
Google Scholar
Келлихер Т., Уолбот В. Возникновение и формирование паттерна пяти типов клеток локулы пыльника Zea mays.Dev Biol. 2011, 350: 32-49. 10.1016 / j.ydbio.2010.11.005.
PubMed CAS PubMed Central Google Scholar
Tsukaya H: Форма и размер органа: урок изучения морфогенеза листьев. Curr Opin Plant Biol. 2003, 6: 57-62. 10.1016 / S1369526602000055.
PubMed Google Scholar
Asl LK, Dhondt S, Boudolf V, Beemster GT, Beeckman T, Inze D, Govaerts W., De Veylder L: модельный анализ эпидермальных клеток листьев арабидопсиса выявляет различные паттерны деления и разрастания тротуаров и ограждений. клетки.Plant Physiol. 2011, 156: 2172-2183. 10.1104 / pp.111.181180.
PubMed CAS PubMed Central Google Scholar
Roeder AHK, Chickarmane V, Cunha A, Obara B, Manjunath BS, Meyerowitz EM: Изменчивость в контроле клеточного деления лежит в основе формирования эпидермального паттерна чашелистика у Arabidopsis thaliana. PLoS Biol. 2010, 8: e1000367-10.1371 / journal.pbio.1000367.
PubMed PubMed Central Google Scholar
Девитт В., Риу-Кхамличи С., Скофилд С., Хили Дж. М., Жакмар А., Килби Н. Дж., Мюррей Дж. А.: изменение распределения клеточного цикла, гиперплазия и ингибирование дифференцировки арабидопсиса, вызванное циклином D-типа CYCD3. Растительная клетка. 2003, 15: 79-92. 10.1105 / tpc.004838.
PubMed CAS PubMed Central Google Scholar
Menges M, Samland AK, Planchais S, Murray JA: Циклин CYCD3; 1 D-типа является лимитирующим для перехода из G1 в S-фазу у Arabidopsis.Растительная клетка. 2006, 18: 893-906. 10.1105 / tpc.105.039636.
PubMed CAS PubMed Central Google Scholar
Hemerly A, Engler Jde A, Bergounioux C, Van Montagu M, Engler G, Inze D, Ferreira P: Доминантно отрицательные мутанты киназы Cdc2 отделяют деление клеток от итеративного развития растений. EMBO J. 1995, 14: 3925-3936.
PubMed CAS PubMed Central Google Scholar
Бемис С.М., Торий К.У .: Автономия клеточной пролиферации и программ развития во время морфогенеза надземных органов Arabidopsis. Dev Biol. 2007, 304: 367-381. 10.1016 / j.ydbio.2006.12.049.
PubMed CAS Google Scholar
Eloy NB, de Freitas Lima M, Van Damme D, Vanhaeren H, Gonzalez N, De Milde L, Hemerly AS, Beemster GT, Inze D, Ferreira PC: субъединица APC / C 10 играет важную роль в пролиферации клеток во время развития листа.Плант Дж. 2011, 68: 351-363. 10.1111 / j.1365-313X.2011.04691.x.
PubMed CAS Google Scholar
Rojas CA, Eloy NB, Lima Mde F, Rodrigues RL, Franco LO, Himanen K, Beemster GT, Hemerly AS, Ferreira PC: Сверхэкспрессия субъединицы CDC27a комплекса, стимулирующего анафазу Arabidopsis, увеличивает скорость роста и размер органов. Завод Мол Биол. 2009, 71: 307-318. 10.1007 / s11103-009-9525-7.
PubMed CAS Google Scholar
Уилсон Э.Б.: Клетка в развитии и наследственности. 1928, Нью-Йорк: Macmillan, 3
Google Scholar
Йоргенсен П., Тайерс М.: Как клетки координируют рост и деление. Curr Biol. 2004, 14: R1014-R1027. 10.1016 / j.cub.2004.11.027.
PubMed CAS Google Scholar
Сугимото-Ширасу К., Робертс К.: «Большое дело»: эндоредупликация и контроль размера клеток в растениях.Curr Opin Plant Biol. 2003, 6: 544-553. 10.1016 / j.pbi.2003.09.009.
PubMed CAS Google Scholar
Melaragno JE, Mehrotra B, Coleman AW: Взаимосвязь между эндополиплоидией и размером клеток в эпидермальной ткани Arabidopsis. Растительная клетка. 1993, 5: 1661-1668.
PubMed PubMed Central Google Scholar
Qi R, John PC: Экспрессия геномного AtCYCD2; 1 у Arabidopsis вызывает деление клеток при меньших размерах клеток: последствия для контроля роста растений.Plant Physiol. 2007, 144: 1587-1597. 10.1104 / pp.107.096834.
PubMed CAS PubMed Central Google Scholar
Де Вейлдер Л., Бекман Т., Бимстер Г.Т., Кролс Л., Террас Ф., Ландриу И., ван дер Шуерен Е., Маес С., Наудтс М., Инзе Д. Функциональный анализ ингибиторов циклин-зависимых киназ арабидопсиса. Растительная клетка. 2001, 13: 1653-1668.
PubMed CAS PubMed Central Google Scholar
Verkest A, Manes CL, Vercruysse S, Maes S, Van Der Schueren E, Beeckman T, Genschik P, Kuiper M, Inze D, De Veylder L: ингибитор циклин-зависимой киназы KRP2 контролирует начало цикла эндоредупликации во время арабидопсиса. развитие листа через ингибирование митотических CDKA; 1 киназных комплексов. Растительная клетка. 2005, 17: 1723-1736. 10.1105 / tpc.105.032383.
PubMed CAS PubMed Central Google Scholar
Черчман М.Л., Браун М.Л., Като Н., Кирик В., Хюльскамп М., Инзе Д., Де Вейлдер Л., Уокер Дж. Д., Чжэн З., Оппенгеймер Д. Г., Гвин Т., Дж. Черчман, Ларкин Дж. К.: SIAMESE, завод -специфический регулятор клеточного цикла, контролирующий начало эндорепликации у Arabidopsis thaliana.Растительная клетка. 2006, 18: 3145-3157. 10.1105 / tpc.106.044834.
PubMed CAS PubMed Central Google Scholar
Walker JD, Oppenheimer DG, Concienne J, Larkin JC: SIAMESE, ген, контролирующий клеточный цикл эндоредупликации в трихомах Arabidopsis thaliana. Разработка. 2000, 127: 3931-3940.
PubMed CAS Google Scholar
Castellano MM, del Pozo JC, Ramirez-Parra E, Brown S, Gutierrez C: Экспрессия и стабильность CDC6 Arabidopsis связаны с эндорепликацией.Растительная клетка. 2001, 13: 2671-2686.
PubMed CAS PubMed Central Google Scholar
Horvath BM, Magyar Z, Zhang Y, Hamburger AW, Bako L, Visser RG, Bachem CW, Bogre L: EBP1 регулирует размер органов через рост и пролиферацию клеток в растениях. EMBO J. 2006, 25: 4909-4920. 10.1038 / sj.emboj.7601362.
PubMed CAS PubMed Central Google Scholar
Kurepa J, Smalle JA: Структура, функция и регуляция протеасом растений. Биохимия. 2008, 90: 324-335. 10.1016 / j.biochi.2007.07.019.
PubMed CAS Google Scholar
Kurepa J, Wang S, Li Y, Zaitlin D, Pierce AJ, Smalle JA: Потеря функции протеасомы 26S приводит к увеличению размера клеток и снижению количества клеток в органах побегов Arabidopsis. Plant Physiol. 2009, 150: 178-189. 10.1104 / pp.109.135970.
PubMed CAS PubMed Central Google Scholar
Sonoda Y, Sako K, Maki Y, Yamazaki N, Yamamoto H, Ikeda A, Yamaguchi J: Регулирование размера органа листа с помощью субъединицы протеасомы RPT2a 19S Arabidopsis. Плант Дж. 2009, 60: 68-78. 10.1111 / j.1365-313X.2009.03932.x.
PubMed CAS Google Scholar
Депрост Д., Яо Л., Сормани Р., Моро М., Летерре Дж., Николай М., Беду М., Робалья С., Мейер С. Киназа TOR арабидопсиса связывает рост растений, урожайность, устойчивость к стрессу и трансляцию мРНК.EMBO Rep.2007, 8: 864-870. 10.1038 / sj.embor.7401043.
PubMed CAS PubMed Central Google Scholar
Hay N, Sonenberg N: до и после mTOR. Genes Dev. 2004, 18: 1926-1945. 10.1101 / gad.1212704.
PubMed CAS Google Scholar
Wullschleger S, Loewith R, Hall MN: передача сигналов TOR в росте и метаболизме. Клетка. 2006, 124: 471-484.10.1016 / j.cell.2006.01.016.
PubMed CAS Google Scholar
Ren M, Qiu S, Venglat P, Xiang D, Feng L, Selvaraj G, Datla R: Мишень рапамицина регулирует развитие и экспрессию рибосомной РНК через киназный домен в Arabidopsis. Plant Physiol. 2011, 155: 1367-1382. 10.1104 / pp.110.169045.
PubMed CAS PubMed Central Google Scholar
Cosgrove DJ: Рост клеточной стенки растений. Nat Rev Mol Cell Biol. 2005, 6: 850-861. 10.1038 / nrm1746.
PubMed CAS Google Scholar
Косгроув Д. Д. Растяжимость стенок: ее природа, измерение и связь с ростом растительных клеток. Новый Фитол. 1993, 124: 1-23. 10.1111 / j.1469-8137.1993.tb03795.x.
PubMed CAS Google Scholar
Cosgrove DJ: Разрыхление стенок растительных клеток экспансинами. Природа. 2000, 407: 321-326. 10.1038 / 35030000.
PubMed CAS Google Scholar
Маккуин-Мейсон С., Дурачко Д.М., Косгроув Д.Д.: Два эндогенных белка, которые вызывают расширение клеточной стенки у растений. Растительная клетка. 1992, 4: 1425-1433.
PubMed CAS PubMed Central Google Scholar
Cho HT, Cosgrove DJ: Измененная экспрессия экспансина модулирует рост листьев и опадание цветоножек у Arabidopsis thaliana. Proc Natl Acad Sci USA. 2000, 97: 9783-9788. 10.1073 / pnas.160276997.
PubMed CAS PubMed Central Google Scholar
Paredez AR, Somerville CR, Ehrhardt DW: Визуализация целлюлозосинтазы демонстрирует функциональную ассоциацию с микротрубочками. Наука. 2006, 312: 1491-1495. 10.1126 / наука.1126551.
PubMed CAS Google Scholar
Амброуз Дж. К., Шоджи Т., Котцер А. М., Пигин Дж. А., Уэйстней Г.О .: Ген CLASP арабидопсиса кодирует связанный с микротрубочками белок, участвующий в размножении и делении клеток. Растительная клетка. 2007, 19: 2763-2775. 10.1105 / tpc.107.053777.
PubMed CAS PubMed Central Google Scholar
Перемыслов В.В., Прохневский А.И., Доля В.В.: Миозины класса XI необходимы для развития, размножения клеток и организации F-актина у Arabidopsis.Растительная клетка. 2010, 22: 1883-1897. 10.1105 / tpc.110.076315.
PubMed CAS PubMed Central Google Scholar
Bringmann M, Li E, Sampathkumar A, Kocabek T, Hauser MT, Persson S: Взаимодействие POM-POM2 / целлюлозосинтазы1 необходимо для функциональной ассоциации синтазы целлюлозы и микротрубочек у Arabidopsis. Растительная клетка. 2012, 24: 163-177. 10.1105 / tpc.111.093575.
PubMed CAS PubMed Central Google Scholar
Эльснер Дж., Михальски М., Квятковска Д.: Пространственно-временные вариации роста эпидермальных клеток листа: количественный анализ растений дикого типа Arabidopsis thaliana и растений с тройным мутантом cyclinD3. Энн Бот. 2012, 109: 897-910. 10.1093 / aob / mcs005.
PubMed CAS PubMed Central Google Scholar
Haber AH: Несущественность параллельных делений клеток для степени поляризации роста листа .1. Исследования с радиационно-индуцированным митотическим ингибированием.Am J Bot. 1962, 49: 583-589. 10.2307 / 2439715.
Google Scholar
Kim JH, Choi D, Kende H: Семейство предполагаемых факторов транскрипции AtGRF участвует в росте листьев и семядолей у Arabidopsis. Плант Дж. 2003, 36: 94-104. 10.1046 / j.1365-313X.2003.01862.x.
PubMed CAS Google Scholar
Ли Б.Х., Ко Дж. Х., Ли С., Ли Й, Пак Дж. Х., Ким Дж. Х .: Семейство генов ВЗАИМОДЕЙСТВУЮЩИХ с GRF ФАКТОРАМИ Arabidopsis выполняет перекрывающуюся функцию в определении размера органов, а также множественных свойств развития.Plant Physiol. 2009, 151: 655-668. 10.1104 / pp.109.141838.
PubMed CAS PubMed Central Google Scholar
Horiguchi G, Kim GT, Tsukaya H: Фактор транскрипции AtGRF5 и коактиватор транскрипции AN3 регулируют пролиферацию клеток в зачатках листьев Arabidopsis thaliana. Плант Дж. 2005, 43: 68-78. 10.1111 / j.1365-313X.2005.02429.x.
PubMed CAS Google Scholar
Kim JH, Kende H: Коактиватор транскрипции, AtGIF1, участвует в регуляции роста и морфологии листьев у Arabidopsis. Proc Natl Acad Sci USA. 2004, 101: 13374-13379. 10.1073 / pnas.0405450101.
PubMed CAS PubMed Central Google Scholar
Roeder AH, Tarr PT, Tobin C, Zhang X, Chickarmane V, Cunha A, Meyerowitz EM: Вычислительная морфодинамика растений: интеграция развития в пространстве и времени.Nat Rev Mol Cell Biol. 2011, 12: 265-273. 10.1038 / nrm3079.
PubMed CAS PubMed Central Google Scholar
Dinneny JR, Yadegari R, Fischer RL, Yanofsky MF, Weigel D: Роль JAGGED в формировании боковых органов. Разработка. 2004, 131: 1101-1110. 10.1242 / dev.00949.
PubMed CAS Google Scholar
Оно СК, Редди Г.В., Хейслер М.Г., Мейеровиц Е.М.: Ген Arabidopsis JAGGED кодирует белок цинкового пальца, который способствует развитию ткани листа.Разработка. 2004, 131: 1111-1122. 10.1242 / dev.00991.
PubMed CAS Google Scholar
Schiessl K, Kausika S, Southam P, Bush M, Sablowski R: JAGGED контролирует анизотропию роста и координацию между размером клетки и клеточным циклом во время органогенеза растений. Curr Biol. 2012, 22: 1739-1746. 10.1016 / j.cub.2012.07.020.
PubMed CAS PubMed Central Google Scholar
Roeder AH: Когда и где делятся клетки растений: взгляд на компьютерное моделирование. Curr Opin Plant Biol. 2012, 15: 638-644. 10.1016 / j.pbi.2012.08.002.
PubMed Google Scholar
Траас Дж, Хулскэмп М., Гендро Э, Хофте Х: Эндоредупликация и развитие: правило без разделения ?. Curr Opin Plant Biol. 1998, 1: 498-503. 10.1016 / S1369-5266 (98) 80042-3.
PubMed CAS Google Scholar
Завод vs.Клетки животных | Biology Dictionary
Клетки — это фундаментальные единицы, из которых состоит все живое, включая растения и животных. Все клетки растений и животных являются эукариотическими, поэтому у них есть несколько общих клеточных процессов и органелл. Однако есть также ключевые различия между растительными и животными клетками с точки зрения их размера, формы и клеточных структур, которые они содержат.
В чем сходство и различие между растительными и животными клетками?
Растительные клетки | Клетки животных |
| Содержат хлоропласты | Не содержат хлоропластов |
| Клеточная стенка | Без клеточной стенки |
| Содержит одну большую вакуоль | Может содержать несколько мелких вакуолей |
| 10-30 мкм | 10 — 100 мкм |
| Круглая неправильная форма | Прямоугольная или коробчатая |
Завод vs.Размер животной клетки
Растительные клетки часто больше, чем клетки животных. В то время как нормальный диапазон размеров животной клетки составляет от 10 до 30 микрометров (мкм), растительные клетки могут иметь размер от 10 до 100 мкм.
Форма клеток растений и животных
Клетки животных имеют тенденцию быть круглыми с неправильной формой. Это отличается от растительных клеток, которые имеют фиксированную прямоугольную или коробчатую форму.
Растительные и животные клетки имеют разную форму
Хранение энергии на заводе vs.Клетки животных
Клетки растений и животных накапливают энергию, но используют для этого разные молекулы. Клетки животных хранят энергию в виде молекул гликогена молекул, тогда как клетки растений хранят свою энергию в крахмале .
Структуры клеток растений и животных
Клетки растений и животных содержат многие из одних и тех же органелл, но некоторые структуры встречаются только в клетках растений. Другие встречаются как в растительных, так и в животных клетках, но их функции немного отличаются.
Стенка клетки
Растительные клетки окружены прочной структурой на основе целлюлозы, называемой клеточной стенкой. Клеточная стенка находится за пределами клеточной мембраны растительных клеток, но отсутствует в клетках животных.
Хлоропласты
Хлоропласты являются местом фотосинтеза, который использует энергию солнца для преобразования углекислого газа и воды в глюкозу. Так растения получают пищу, поэтому хлоропласты — обычная черта растительных клеток. Однако хлоропласты никогда не обнаруживаются в клетках животных, поскольку животные получают пищу за счет еды, а не за счет фотосинтеза.
Хлоропласты находятся в клетках растений, но не в клетках животных
Лизосомы
Лизосомы — это маленькие сферические органеллы, наполненные пищеварительными ферментами, которые используются для расщепления и переработки нежелательных материалов. Лизосомы также используются для уничтожения вторгающихся патогенов. Они распространены в клетках животных, но редко в клетках растений, поскольку прочная клеточная стенка растений помогает удерживать нежелательных захватчиков.
Вакуоли
И растения, и животные содержат вакуоли, но их структура сильно различается.Клетка животного может содержать несколько небольших вакуолей, которые обычно используются для хранения продуктов жизнедеятельности. Напротив, вакуоль растительной клетки очень велика и может занимать до 90% объема клетки. Он используется для хранения различных веществ (включая воду, сахар, белки, соли и т. Д.) И помогает поддерживать тургорное давление клетки.
Плазмодесматы
Плазмодесмы — это небольшие каналы или поры между растительными клетками, которые напрямую соединяют одну клетку с другой.Клетки растений могут использовать плазмодесмы для обмена молекулами друг с другом. Плазмодесмы встречаются только в клетках растений и водорослей; эквивалент животной клетки называется щелевым соединением .
Плазмодесматы — это каналы, соединяющие одну растительную клетку с другой.
Сходства между растительными и животными клетками
Оба являются эукариотами
Клетки растений и животных являются типами эукариотических клеток, что означает, что они оба содержат истинное ядро, а также другие мембраносвязанные органеллы.
Оба респира
Все клетки нуждаются в энергии для роста и функционирования, и клетки животных и растений получают эту энергию от клеточного дыхания. Этот метаболический процесс происходит в митохондриях клеток растений и животных и включает расщепление глюкозы с высвобождением энергии.
Клетки растений и животных содержат митохондрии для дыхания
Органеллы, обнаруженные в клетках растений и животных
Клетки растений и животных содержат многие из одних и тех же клеточных структур, включая ядро, митохондрии, рибосомы, эндоплазматический ретикулум, аппарат Гольджи, пероксисомы, цитоплазму и клеточную мембрану.
сравнений размеров бактерий, амеб, клеток животных и растений | Образование
Клетки — это основные единицы жизни. Все живые организмы состоят из одной или нескольких клеток. Бактерии состоят из отдельных клеток и намного меньше клеток растений и животных. Клетки растений и животных бывают самых разных размеров и форм, в зависимости от их функции. Большинство клеток растений и животных составляют органы и определенные ткани и складываются вместе, как кусочки пазла. Почти все клетки слишком малы, чтобы их можно было увидеть человеческим глазом, но вы можете наблюдать их под световым микроскопом.
Бактериальные клетки
Бактериальные клетки очень маленькие — примерно в 10 раз меньше, чем большинство клеток растений и животных. Большинство бактериальных клеток имеют размер от 0,2 до 10 микрон или микрометров (от 0,0000079 до 0,00039 дюйма). Обычные бактерии Escherichia coli, или кишечная палочка, представляют собой палочковидные бактерии размером 1 микрон на 2 микрона. Меньшие клетки имеют большую площадь поверхности по сравнению с объемом клетки, в то время как более крупные клетки имеют меньшее отношение площади поверхности к объему. Одна из причин, по которой бактериальные клетки настолько малы, заключается в том, что им требуется большая площадь поверхности по сравнению с объемом клетки, чтобы принимать питательные вещества.Бактерии накапливают питательные вещества из окружающей среды путем диффузии.
Амеба
Амебы — одноклеточные организмы, обитающие в озерах, прудах, ручьях, сточных водах, загрязненной воде и кишечниках животных. Некоторые амебы вызывают заболевания у людей и животных. Обыкновенная амеба, Amoeba proteus, обитает в пресноводных прудах и озерах, содержащих много разлагающихся органических веществ. Размер амебы протей колеблется от 250 до 750 микрон. Американское общество микробиологии заявляет, что размер амебы частично зависит от наличия пищи в данном месте.
Клетки животных
Большинство клеток животных имеют размер от 10 до 100 микрон. Размер частично зависит от типа клетки и ее функции. Эритроциты, которым не нужно делиться и воспроизводиться, имеют диаметр всего около 8 микрон, в то время как многие мышечные и нервные клетки тонкие, веретенообразные и чрезвычайно длинные. Например, нервная клетка жирафа может достигать двух метров (около 6 футов). Яйца — еще один пример необычно больших одиночных клеток. Яйцеклетка страуса имеет длину примерно 6 дюймов; яйцеклетка человека составляет 100 микрон.Яичные клетки служат для хранения питательных веществ и не нуждаются в активном обмене веществ.
Растительные клетки
Растительные клетки сравнимы с клетками животных по размеру: от 10 до 100 микрон; однако большее количество растительных клеток находится в верхнем конце этого диапазона. Хотя и животные, и растительные клетки имеют мягкую гибкую мембрану, мембраны большинства растительных клеток покрыты жесткими угловатыми клеточными стенками. Стенки клеток состоят из сахарных полимеров, таких как пектин и целлюлоза. Клеточная стенка клеток паренхимы, которых много у молодых растений и обычно содержат хлорофилл, тонкая.Клетки, обеспечивающие структуру, гибкую опору и проводящие воду, имеют веретенообразную форму, имеют удлиненную форму и толстые стенки и могут выполнять свою функцию даже после смерти. Примерами являются клетки деревьев, которые составляют ксилему и проводят воду от корней к надземным частям растений.
Ссылки
Ресурсы
Биография писателя
Мари-Луиза Блю, живущая в Коннектикуте, ведет колонку о местном садоводстве и публиковалась в журналах «Органическое садоводство» и «Back Home».»Блю имеет докторскую степень в области биологических наук в Государственном университете Нью-Йорка в Стоуни-Брук и писал научные статьи почти 20 лет, прежде чем в 2004 году начал писать статьи о садоводстве.
О молекулярных механизмах, регулирующих гомеостаз размера клеток животных
Основные моменты
- •
Размер клетки имеет решающее значение для физиологии клетки, поскольку он устанавливает геометрический масштаб клеточных органелл и способствует определению биосинтетической способности клетки.
- •
Гомеостатический контроль размера клеток достигается за счет сочетания размера клеток, скорости роста клеток и развития клеточного цикла.
- •
В клетках животных контроль размера клеток в основном происходит в G1 и проявляется как обратная корреляция между размером клетки при рождении и величиной роста в фазе G1.
- •
Контроль размера G1 может возникнуть из-за активации p38 в малых клетках и разбавления ингибитора клеточного цикла RB по мере роста клеток.
- •
Визуализация отдельных клеток, геномные и протеомные исследования, изучающие динамику активности и концентраций ключевых регуляторов клеточного цикла, могут определить молекулярные механизмы, лежащие в основе контроля размера клеток.
Размер клетки имеет фундаментальное значение для физиологии клетки, потому что он определяет масштаб внутриклеточной геометрии, органелл и биосинтетических процессов. В клетках животных гомеостаз размера контролируется двумя феноменологически различными механизмами.Во-первых, прогрессирование клеточного цикла в зависимости от размера гарантирует, что меньшие клетки задерживают развитие клеточного цикла, чтобы накапливать больше биомассы, чем более крупные клетки, до клеточного деления. Во-вторых, рост клеток в зависимости от размера гарантирует, что более крупные и мелкие клетки растут медленнее на единицу массы, чем клетки с более оптимальным размером. В этом десятилетии произошел значительный прогресс в одноклеточных технологиях, установивших различные явления контроля размера клеток в клетках животных. Здесь мы рассматриваем этот недавний прогресс и предлагаем дальнейшие пути определения лежащих в основе молекулярных механизмов.
Ключевые слова
размер клетки
скорость роста клеток
клеточный цикл
разведение белка
Стохастический классификатор G1 / S
Рекомендуемые статьиЦитирующие статьи (0)
Полный текст© 2020 Elsevier Ltd. Все права защищены.
Рекомендуемые статьи
Ссылки на статьи
Клетки животных и их формы — Science Learning Hub
Клетки — это строительные блоки жизни — все живые организмы состоят из них.В учебниках часто приводится единственный «типичный» пример растительной или животной клетки, но в действительности формы клеток могут сильно различаться. В частности, клетки животных бывают самых разных форм и размеров. Формы растительных клеток, как правило, очень похожи друг на друга из-за их жесткой клеточной стенки.
Мы можем многое узнать о том, что делает клетка, глядя на ее форму и размер, и микроскопы — идеальный инструмент для этого.
Формы для задачи
Ячейки имеют разную форму, потому что они выполняют разные функции.Каждый тип клеток играет свою роль, помогая нашему телу работать должным образом, а их форма помогает им эффективно выполнять эти функции. Все перечисленные ниже типы ячеек имеют необычную форму, которая важна для их функции.
Нейроны — это клетки головного мозга и нервной системы. Их работа — передавать электрические сообщения от мозга к остальным частям тела и обратно (почти как электрический провод), поэтому они представляют собой очень длинные и тонкие клетки. Им также необходимо соединяться с другими нейронами для формирования сетей связи, поэтому у них много длинных ветвей.Вы можете узнать больше о нейронах в другом месте на нашем сайте.
Фоторецепторные клетки (палочки и колбочки) — это клетки глаза, которые улавливают свет. На самом деле это очень специализированная форма нейрона. Фоторецепторы должны собирать свет с максимальной эффективностью, поэтому у них есть специальный выступ из клетки (называемый внешним сегментом), который заполнен молекулами, поглощающими свет. У стержней, которые особенно хорошо обнаруживают свет, выступы больше. Внешний сегмент теперь известен как сильно модифицированный вид первичной реснички, недавно обнаруженной органеллы.Вы можете прочитать больше о первичной ресничке в другом месте в этом контексте.
Иммунные клетки — это клетки, которые реагируют на заражение организма (например, бактерией). Чтобы выполнять свою работу, они должны уметь менять форму. Например, лимфоцитам может потребоваться пройти через ткани тела, чтобы добраться до места инфекции, поэтому они изменяют свою форму, чтобы протиснуться мимо плотно упакованных клеток ткани. Некоторые иммунные клетки (например, нейтрофилы) поглощают бактерии и вирусы, поэтому им необходимо изменить свою форму, чтобы «проглотить» их.Вы можете узнать больше о различных типах иммунных клеток в разделе «Борьба с инфекциями».
Микроскопы на клетках
Практически все наше понимание формы клеток основано на годах накопленных микроскопических экспериментов. Нет другого инструмента, который позволяет нам напрямую смотреть на форму клетки. Используя световую микроскопию, ученые смогли увидеть живые клетки, чтобы увидеть, как их формы меняются с течением времени. Они также смогли наблюдать клеточные процессы, которые включают изменения формы, такие как митоз
. Из-за конструкции электронных микроскопов живые клетки не могут выжить в суровых условиях внутри микроскопа и, следовательно, не могут быть просмотрены напрямую.Однако электронные микроскопы могут дать информацию с высоким разрешением о форме отдельных клеток, которые были подготовлены для просмотра, включая небольшие участки клетки, которые имеют определенную форму, такие как первичные реснички и микроворсинки.
Клетки в 3D
Клетки — это трехмерные объекты сложной формы, но изображения, полученные с помощью большинства микроскопов, являются двухмерными. Это затрудняет понимание общей формы клеток и того, как они взаимодействуют друг с другом.Однако теперь несколько микроскопических методов позволяют создавать трехмерные модели клеток или их частей. Это делается путем сбора нескольких двумерных изображений в цифровом виде, а затем их объединения с помощью компьютерных программ.
Доктор Ребекка Кэмпбелл и доцент Тони Пул — два ученых из Университета Отаго, которые используют изображения с микроскопа для создания трехмерных моделей изучаемых клеток. Ребекка использует несколько изображений с конфокального лазерного сканирующего микроскопа для построения трехмерных изображений целых нейронов, а Тони строит трехмерную модель первичной реснички.Вы можете узнать больше о работе Ребекки и Тони в другом месте в этом контексте.
Природа науки
Научные эксперименты часто обнаруживают неожиданную информацию, которая может привести к новым гипотезам и теориям. Ученые 1600-х годов были удивлены, увидев крошечные «строительные блоки», когда рассматривали ткани под оптическими микроскопами. Их наблюдения в конечном итоге привели к развитию клеточной теории — идеи, что клетка является основной единицей жизни.
Полезные ссылки
Посмотрите это короткое видео, чтобы увидеть, как иммунная клетка (нейтрофил) меняет свою форму, поскольку она следует за бактерией по крови и в конечном итоге поглощает ее.