
1. Задачи селекции. Методы селекции. Работы Н. И. Вавилова
Все особи такой группы имеют сходные морфологические и физиологические признаки, однотипную реакцию на изменение факторов внешней среды, определённый уровень продуктивности.
1. Искусственный отбор используется для сохранения и размножения особей с желаемой комбинацией признаков. Различают массовый и индивидуальный отбор.
При массовом отборе одновременно отбирают большое число особей с нужным признаком, остальные выбраковывают. Это отбор по фенотипу, он не даёт генетически однородного материала. Повторяется многократно.
При индивидуальном отборе (по генотипу) выделяют одну особь с необходимыми признаками и получают от неё потомство.
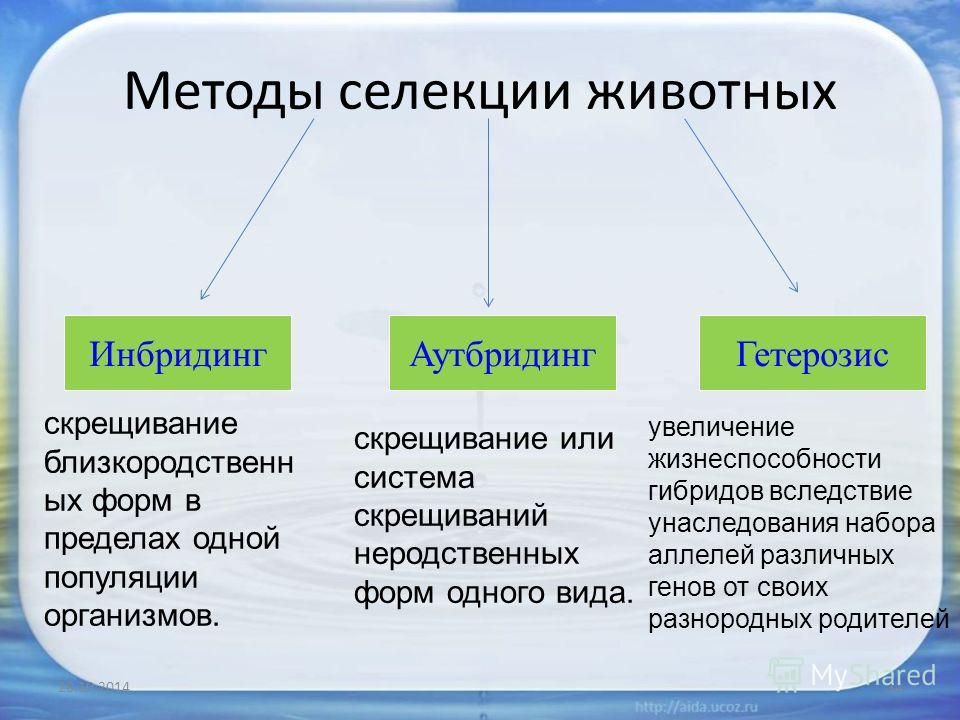
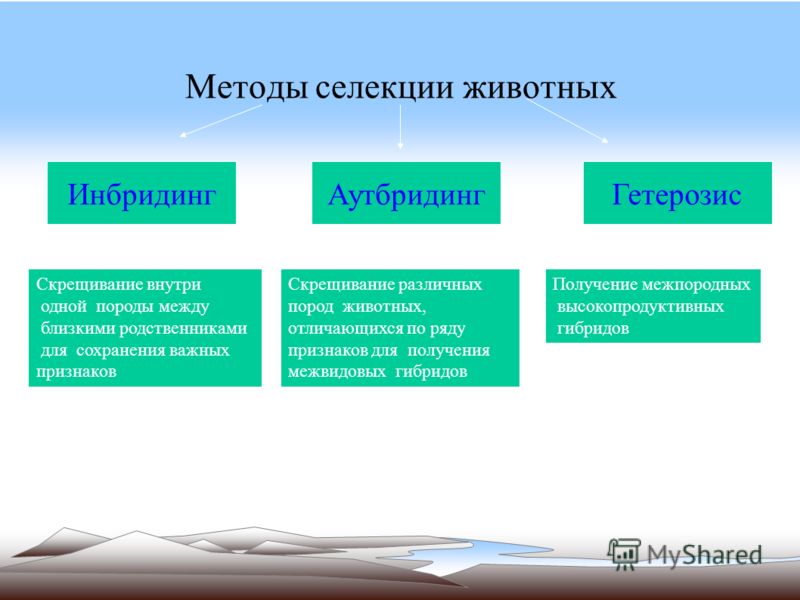
2. В селекционной работе используют следующие методы гибридизации: инбридинг, аутбридинг и отдалённую гибридизацию.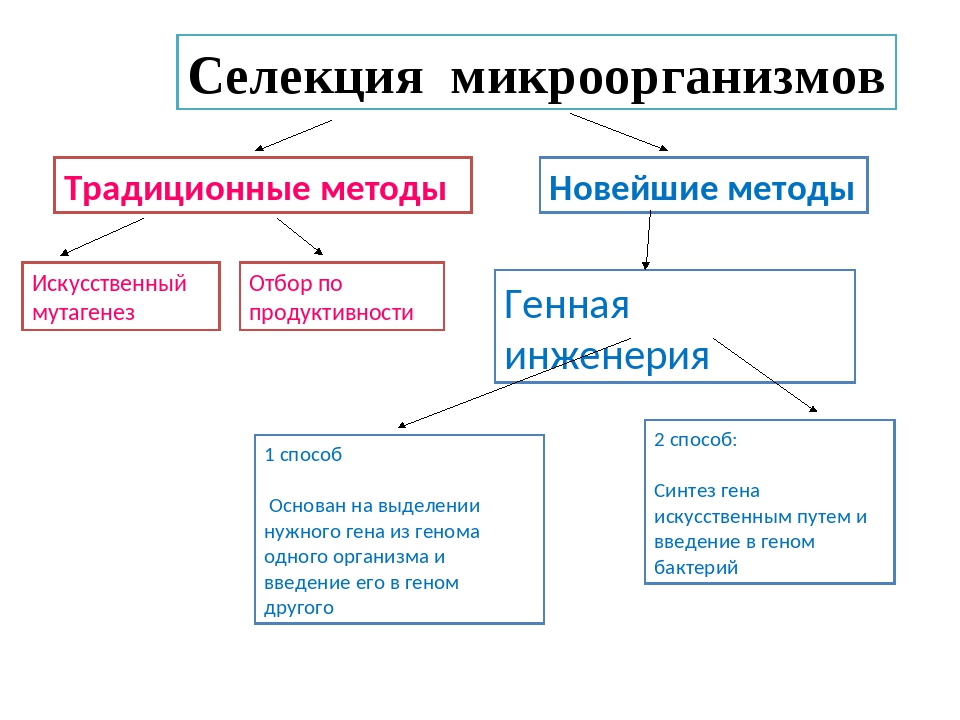
Инбридинг — близкородственное скрещивание.
При инбридинге скрещиваются потомки с родительскими формами или потомки одних и тех же родителей. Этот тип скрещивания применяют для получения чистых линий, т. е. перевода большинства генов в гомозиготное состояние и закрепления ценных признаков. Нежелательным последствием близкородственного скрещивания является инбредная депрессия — снижение продуктивности и жизнеспособности потомства из-за проявления рецессивных мутаций.
Аутбридинг — неродственное (межпородное или межсортовое) скрещивание.
При неродственном скрещивании может наблюдаться эффект гетерозиса (гибридной силы) — повышение жизнеспособности и продуктивности гибридов по сравнению с родительскими формами. Гетерозис проявляется у гибридов первого поколения и обусловлен переходом большинства генов в гетерозиготное состояние. При этом нежелательные рецессивные мутации становятся скрытыми. При половом размножении в следующих поколениях степень гетерозиготности уменьшается и эффект гибридной силы исчезает. Он может сохраняться только при вегетативном размножении.
При этом нежелательные рецессивные мутации становятся скрытыми. При половом размножении в следующих поколениях степень гетерозиготности уменьшается и эффект гибридной силы исчезает. Он может сохраняться только при вегетативном размножении.
Отдалённая гибридизация — скрещивание организмов, относящихся к разным видам и родам.
Осуществляется с трудом, а полученные гибриды бесплодны из-за затруднения конъюгации хромосом разных видов в профазе \(I\) мейоза. Разработаны методы преодоления бесплодия.
3. Искусственный (индуцированный) мутагенез используют для увеличения разнообразия исходного материала. Мутагенез вызывают действием мутагенных факторов, например, рентгеновского облучения. Мутации носят ненаправленный характер, поэтому селекционер отбирает организмы с новыми полезными свойствами.
Геномной мутацией является полиплоидия, т. е. кратное увеличение числа хромосомных наборов. Используется в селекции растений. Полиплоидия позволяет избежать бесплодия межвидовых гибридов. Кроме того, многие полиплоидные формы культурных растений (пшеницы, картофеля, овощных культур) имеют более высокую урожайность, чем родственные диплоидные виды.
е. кратное увеличение числа хромосомных наборов. Используется в селекции растений. Полиплоидия позволяет избежать бесплодия межвидовых гибридов. Кроме того, многие полиплоидные формы культурных растений (пшеницы, картофеля, овощных культур) имеют более высокую урожайность, чем родственные диплоидные виды.
Искусственно полиплоидию вызывают обработкой растений колхицином. Колхицин разрушает нити веретена деления и препятствует расхождению гомологичных хромосом в процессе мейоза.
Для успешной селекционной работы в первую очередь необходим разнообразный исходный материал.
Поиск исходного материала облегчает закон гомологических рядов наследственной изменчивости, открытый Н. И. Вавиловым.
Если известны формы изменчивости одного вида, то можно предположить, что подобные формы будут существовать и у других близкородственных видов.
|
Методы селекции |
Селекция животных |
Селекция растений |
|
Подбор родительских пар |
По хозяйственно ценным признакам и по экстерьеру (совокупности фенотипических признаков) |
По месту их происхождения (географически удаленных) или генетически отдаленных (неродственных) |
|
Гибридизация: а) неродственная (аутбридинг) |
Скрещивание отдаленных пород, отличающихся контрастными признаками, для получения гетерозиготных популяций и проявления гетерозиса. |
Внутривидовое, межвидовое, межродовое скрещивание, ведущее к гетерозису, для получения гетерозиготных популяций, а также высокой продуктивности |
|
б) близкородственная (инбридинг) |
Скрещивание между близкими родственниками для получения гомозиготных (чистых) линий с желательными признаками |
Самоопыление у перекрестноопыляющихся растений путем искусственного воздействия для получения гомозиготных (чистых) линий |
|
Отбор: а) массовый |
Не применяется |
Применяется в отношении перекрестноопыляющихся растений |
|
б) индивидуальный |
Применяется жесткий индивидуальный отбор по хозяйственно ценным признакам, выносливости, экстерьеру |
Применяется в отношении самоопыляющихся растений, выделяются чистые линии — потомство одной самоопыляющейся особи |
|
Метод испытания производителей по потомству |
Используют метод искусственного осеменения от лучших самцов-производителей, качества которых проверяют по многочисленному потомству |
Не применяется |
|
Экспериментальное получение полиплоидов |
Не применяется |
Применяется в генетике и селекции для получения более продуктивных, урожайных форм |
|
Биологически отдаленная гибридизация: а) межвидовая |
Скрещивание представителей разных видов для получения сортов с нужными свойствами |
Вишня Владимирская х черешня Винклера белая = вишня Краса севера (хороший вкус, зимостойкость) |
|
б) межродовая |
Скрещивание представителей разных родов для получения новых форм растений |
Вишня х черемуха = церападус |
|
Географически отдаленная гибридизация |
Скрещивание представителей контрастных природных зон и географически отдаленных регионов с целью привить гибриду нужные качества (вкусовые, устойчивости) |
Груша дикая уссурийская х Бере рояль (Франция) = Бере зимняя Мичурина |
|
Отбор |
Многократный, жесткий: по размерам, форме, зимостойкости, иммунным свойствам, качеству, вкусу, цвету плодов и их лежкости |
Продвинуто на север много сортов яблонь с хорошими вкусовыми качествами и высокой урожайностью |
|
Метод ментора |
Воспитание в гибридном сеянце желательных качеств (усиление доминирования), для чего сеянец прививается на растение-воспитатель, от которого эти качества хотят получить. |
Яблоня Китайка (подвой) х гибрид (Китайка х Кандиль-синап) = Кандиль-синап (морозостойкий) Бельфлер-китайка (гибрид-подвой) х Китайка (привой) = Бельфлер-китайка (лежкий позднеспелый сорт) |
|
Метод посредника |
При отдаленной гибридизации для преодоления нескрещиваемо-сти использование дикого вида в качестве посредника |
Дикий монгольский миндаль х дикий персик Давида = миндаль Посредник Культурный персик х миндаль Посредник = гибридный персик (продвинут на север) |
|
Воздействие условиями среды |
При воспитании молодых гибридов обращалось внимание на метод хранения семян, характер и степень питания, воздействие низкими температурами, бедной питанием почвой, частыми пересадками |
Закаливание гибридного сеянца. |
|
Смешение пыльцы |
Для преодоления межвидовой нескрещиваемости (несовместимости) |
Смешивалась пыльца Материнского растения с пыльцой отцовского, своя пыльцы раздражала рыльце, и оно воспринимало чужую пыльцу |
 Получается бесплодное потомство
Получается бесплодное потомство
 Отбор наиболее выносливых растений
Отбор наиболее выносливых растенийОхарактеризуйте основные методы современной селекции растений. Приведите примеры выведения сортов растений при помощи этих методов.
Охарактеризуйте основные методы современной селекции растений. Приведите примеры выведения сортов растений при помощи этих методов.
Основные методы селекции включают отбор, гибридизацию, полиплоидию и мутагенез. В последнее время в селекции также широко используют новые методы — клеточную и генную инженерию.
1. Отбор. В основе появления селекции как науки находится созданная трудами Ч. Дарвина концепция искусственного отбора. На ранних этапах социальной эволюции человека отбор носил бессознательный характер. Люди сохраняли лучшие растения и животные для получения потомства, а слабые представители уничтожались или употреблялись в пищу. С развитием цивилизации люди стали производить методический отбор, при котором сохранялись особи с желаемыми для человека качествами. Известны два вида искусственного отбора: массовый и индивидуальный. При массовом отборе для получения потомства используют популяцию организмов, обладающих определенными признаками. Сорт, полученный таким способом, генетически неоднороден. При индивидуальном отборе от каждой особи получают отдельное потомство и при последующем само опылении у растений и близкородственном скрещивании получают чистые линии. Чистые линии — группы генетически однородных (гомозиготных) организмов.
Люди сохраняли лучшие растения и животные для получения потомства, а слабые представители уничтожались или употреблялись в пищу. С развитием цивилизации люди стали производить методический отбор, при котором сохранялись особи с желаемыми для человека качествами. Известны два вида искусственного отбора: массовый и индивидуальный. При массовом отборе для получения потомства используют популяцию организмов, обладающих определенными признаками. Сорт, полученный таким способом, генетически неоднороден. При индивидуальном отборе от каждой особи получают отдельное потомство и при последующем само опылении у растений и близкородственном скрещивании получают чистые линии. Чистые линии — группы генетически однородных (гомозиготных) организмов.2. Гибридизация — скрещивание особей. Различают внутривидовую и межвидовую (отдаленную) гибридизацию.
Внутривидовая гибридизация. При близкородственном скрещивании повышается степень гомозиготности организма, начинает проявляться действие многих вредных рецессивных аллелей, находившихся до этого в гетерозиготном со стоянии и подавляющихся доминантными генами Многократное близкородственное скрещивание приводит к резкому ослаблению или вырождению потомков. Однако та кое скрещивание бывает часто необходимо для сохранения каких-либо ценных качеств животных или растений. Например, такое близкородственное спаривание было использонно для получения украинской степной белой породы свиней. При скрещивании особей разных линий удается получить гибриды, превосходящие по своей мощности не только родительские линии, но и сходные формы, из которых эти линии были получены. Гетерозис заключается в повышении мощности гибридов первого поколения по сравнению с родительскими формами. Пример гетерозиса — мул, животное, получившееся при скрещивании осла и лошади.
Однако та кое скрещивание бывает часто необходимо для сохранения каких-либо ценных качеств животных или растений. Например, такое близкородственное спаривание было использонно для получения украинской степной белой породы свиней. При скрещивании особей разных линий удается получить гибриды, превосходящие по своей мощности не только родительские линии, но и сходные формы, из которых эти линии были получены. Гетерозис заключается в повышении мощности гибридов первого поколения по сравнению с родительскими формами. Пример гетерозиса — мул, животное, получившееся при скрещивании осла и лошади.
Межвидовая гибридизация заключается в скрещивании разных видов. Её используют как селекционный метод, позволяющий объединить в гибриде ценные хозяйственные признаки родительских форм. Отдаленная гибридизация, как правило, осуществляется с большим трудом и требует применения специальных методов, так как в процессе эволюции даже у видов одного рода возникли приспособления для предотвращения межвидового скрещивания в природе. Межвидовые гибриды часто оказываются бесплодными вследствие нарушения процессов гаметогенеза. Известны гибриды ржи и пшеницы, редьки и капусты, пшеницы и пырея.
Межвидовые гибриды часто оказываются бесплодными вследствие нарушения процессов гаметогенеза. Известны гибриды ржи и пшеницы, редьки и капусты, пшеницы и пырея.
3. Мутагенез. Искусственный мутагенез позволяет вывести новые сорта растений и породы животных, а также улучшить уже существующие породы и сорта. Мутации возникают при воздействии на организмы различными мутагенами (ультрафиолетовые лучи, ионизирующее излучение, некоторые химические вещества). При воздействии мутагена могут возникнуть мутации как понижающие, так и повышающие жизнеспособность организма. Селекционер отбирает и сохраняет формы с необходимыми свойствами. При помощи этого метода был выведен сорт пшеницы Новосибирская-67, который характеризуется коротким и утолщенным побегом и устойчив к полеганию.
4. Полиплоидия. При выведении новых сортов у растений значительное место отводят полиплоидам. Это связано с тем, что в природе многие виды растений — полиплоиды, среди животных полиплоиды встречаются очень редко. В основе возникновения полиплоидии лежат три причины: удвоение хромосом в неделящихся клетках, слияние соматических клеток или их ядер, нарушение процесса мейоза, при котором получаются гаметы с диплоидным набором хромосом. Мягкая пшеница — природный полиплоид. Искусственно полиплоидию можно вызвать путем обработки организмов колхицином. Колхицин разрушает нити веретена деления и тем самым препятствует расхождению гомологичных хромосом в процессе мейоза. С помощью полиплоидии был получен тритикале (гибрид ржи и пшеницы). Благодаря полиплоидии может восстанавливаться плодовитость гибридов, полученных при отдаленной гибридизации (капустно-редечный гибрид).
В основе возникновения полиплоидии лежат три причины: удвоение хромосом в неделящихся клетках, слияние соматических клеток или их ядер, нарушение процесса мейоза, при котором получаются гаметы с диплоидным набором хромосом. Мягкая пшеница — природный полиплоид. Искусственно полиплоидию можно вызвать путем обработки организмов колхицином. Колхицин разрушает нити веретена деления и тем самым препятствует расхождению гомологичных хромосом в процессе мейоза. С помощью полиплоидии был получен тритикале (гибрид ржи и пшеницы). Благодаря полиплоидии может восстанавливаться плодовитость гибридов, полученных при отдаленной гибридизации (капустно-редечный гибрид).
5. Клеточная инженерия позволяет гибридизировать соматические клетки, культивируемые на искусственных средах вне организма, не только между собой, но и с клетками другого вида. Например, гибридизируют клетки мыши и человека, нормальные и раковые клетки, клетки различных растений. Разработаны методы, с помощью которых из группы клеток растения (культуры ткани), содержащегося в пробирке на искусственной среде, можно восстановить целое растение.
6. Генная инженерия — совокупность методов, позволяющих с помощью изменения генов получить у организмов новые признаки. Методами генной инженерии осуществлен синтез биологически активных веществ и препаратов в измененных клетках микроорганизмов. Получены бактерии, обладающие способностью синтезировать инсулин, гормон роста, интерферон и т.д.
Охарактеризуйте основные методы современной селекции растений. Приведите примеры выведения сортов растений при помощи этих методов.
5 (100%) 1 voteНа этой странице искали :
- охарактеризуйте главные методы селекции
- опишите методы выведения новых сортов и пород
- в чём состоит основной метод выведения новых сортов и пород
- Методы современной селекции
- сорта растений примеры
Сохрани к себе на стену!
Методы современной селекции
Основными методами селекции являются отбор, гибридизация (с использованием гетерозиса и цитоплазматической мужской стерильности), полиплоидия и мутагенез.
Отбор и его творческая роль. В основе селекционного процесса лежит искусственный отбор. В сочетании с генетическими методами он позволяет создавать сорта, породы и штаммы с заранее определенными признаками и свойствами. В селекции различают два основных типа отбора: массовый и индивидуальный.
Массовый отбор — это выделение группы особей по внешним, фенотипическим признакам без проверки их генотипа. Например, при массовом, или стихийном, отборе из всей популяции кур той или иной породы в хозяйствах оставляют для размножения птиц с яйценоскостью 200—250 яиц, живой массой не менее 1,5 кг, определенной окраски, не проявляющих инстинкты высиживания и т. д. Все остальные куры выбраковываются. При этом потомство каждой курицы и петуха оценивается только по фенотипу. Следовательно, массовый отбор может дать хорошие результаты только при высоком коэффициенте наследуемости ценных признаков, избранных селекционером.
Массовый отбор наиболее эффективен в отношении качественных признаков, контролируемых одним или несколькими генами.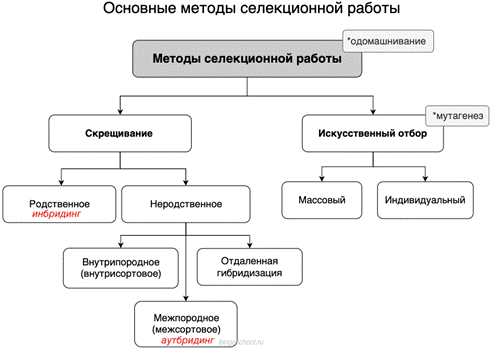 Вместе с тем он редко бывает успешным по полигенным признакам с низким коэффициентом наследования. В этом случае необходимо применять индивидуальный, или методический, отбор.
Вместе с тем он редко бывает успешным по полигенным признакам с низким коэффициентом наследования. В этом случае необходимо применять индивидуальный, или методический, отбор.
При индивидуальном отборе (по генотипу) получают и оценивают потомство каждого отдельного растения или животного в ряду поколений при обязательном контроле наследования интересующих селекционера признаков. На последующих этапах отбора используют только тех особей, которые дали наибольшее число потомков с высокими показателями. В результате появляется возможность оценивать наследственные качества отдельных особей, т. е. способность передавать свойства потомству.
Значение индивидуального отбора особенно велико в тех отраслях сельскохозяйственного производства, где имеется возможность получения от одного организма большого количества потомков. Так, используя искусственное осеменение, от одного быка можно получить до 35000 телят с помощью глубокого замораживания семени, сохраняющегося долгие годы. Поэтому уже теперь во многих странах мира существуют банки спермы животных с ценными генотипами.
Поэтому уже теперь во многих странах мира существуют банки спермы животных с ценными генотипами.
Отбор в селекции отличается наибольшей эффективностью в том случае, если он сочетается с определенными типами скрещиваний.
Методы гибридизации (типы скрещивания) в селекции. Все разнообразие типов скрещиваний сводится к инбридингу и аутбридингу. Инбридинг — это близкородственное (внутрипородное или внутрисортовое), а аутбридинг — неродственное (межпородное или межсортовое) скрещивание.
При инбридинге в качестве исходных форм используются братья и сестры или родители и потомство (отец — дочь, мать — сын, двоюродные братья — сестры и т. д.). Этот тип скрещивания применяют в тех случаях, когда желают перевести большинство генов породы в гомозиготное состояние и, как следствие, закрепить хозяйственно ценные признаки, сохраняющиеся у потомков. Такое скрещивание в определенной степени аналогично самоопылению у растений, которое также приводит к повышению гомозиготности.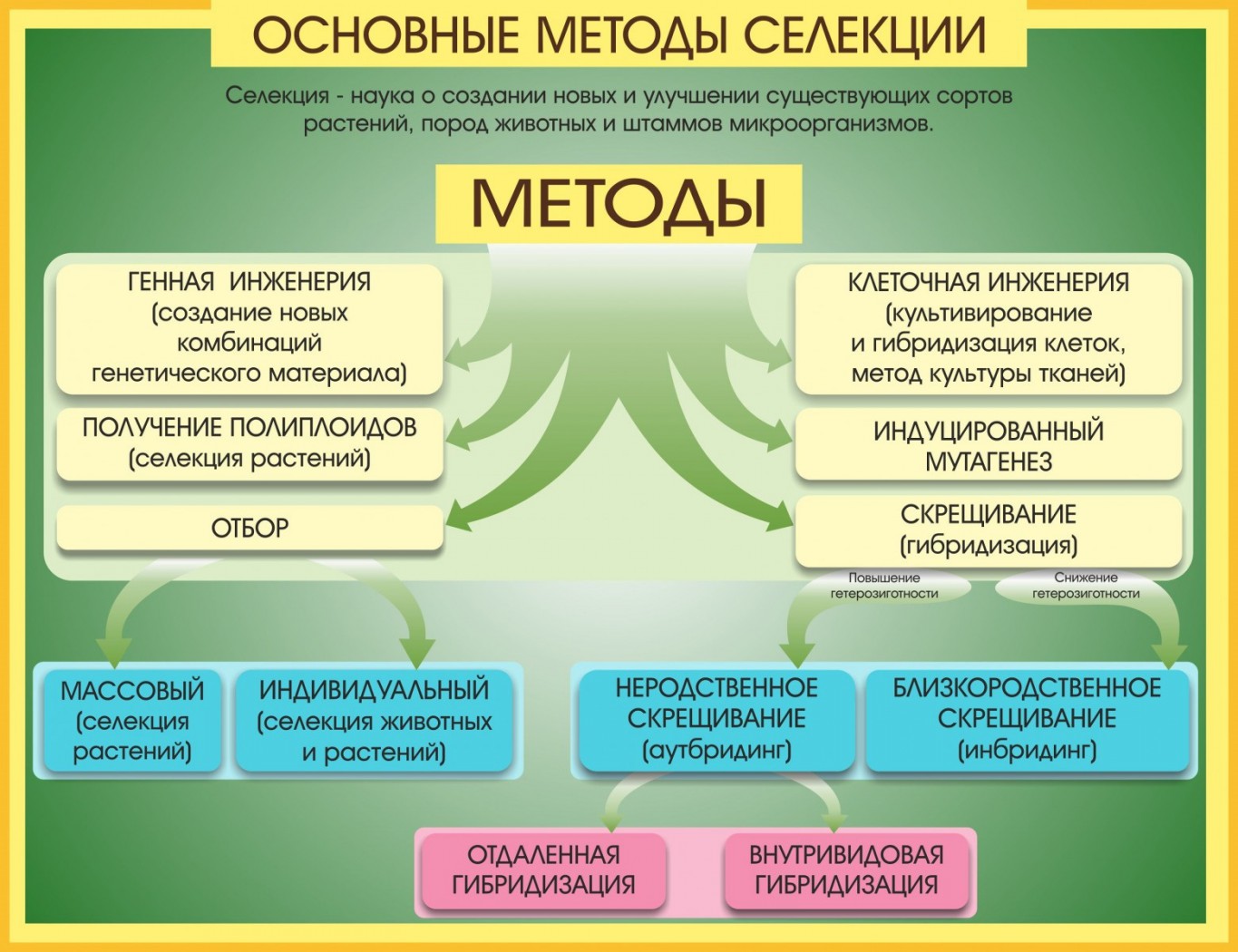
Вместе с тем при инбридинге часто наблюдается ослабление животных, их постепенное вырождение, обусловленное гомозиготизацией рецессивных аллелей. При этом гомозиготизация по генам, контролирующим изучаемый признак, происходит тем быстрее, чем более близкородственные скрещивания используют при инбридинге. Для избежания этого явления необходимо проводить строгий отбор особей, обладающих ценными хозяйственными признаками.
У растений чистые линии также обладают пониженной жизнеспособностью, что, вероятно, связано с переходом в гомозиготное состояние всех рецессивных мутаций, которые в основном являются вредными.
Чистые линии, полученные в результате инбридинга, отличаются не только различными признаками, но и степенью снижения жизнеспособности. Если эти чистые линии скрещивать между собой, то обычно наблюдается эффект гетерозиса.
Неродственное скрещивание между особями одной породы или между особями разных пород (кроссбридинг) животных позволяет поддерживать свойства или улучшать их в ряду следующих поколений гибридов. Аутбридинг повышает уровень гетерозиготности потомства и гетерогенности популяции.
Аутбридинг повышает уровень гетерозиготности потомства и гетерогенности популяции.
Полипюидия и отдаленная гибридизация. При создании новых сортов растений селекционерами широко используется метод автополиплодии, который приводит к увеличению размеров клеток и всего растения вследствие умножения числа наборов хромосом. Кроме того, избыток хромосом повышает их устойчивость к патогенным организмам (вирусам, грибам, бактериям) и ряду других неблагоприятных факторов, например к радиации: при повреждении одной или даже двух гомологичных хромосом остаются неповрежденными другие такие же. Полиплоидные особи жизнеспособнее диплоидных.
Около 80 % современных культурных растений являются полиплоидами. Среди них хлебные злаки, овощные и плодово-ягодные культуры, цитрусовые, технические, лекарственные и декоративные растения, которые гораздо более урожайны, чем исходные диплоидные сорта. Так, триплоидная сахарная свекла отличается от обычной не только большей урожайностью вегетативной массы и более крупными размерами корнеплодов, но и повышенной их сахаристостью, а также устойчивостью к болезням. Однако триплоиды стерильны, поэтому необходимо каждый раз получать гибридные семена от скрещивания диплоидной и тетраплоидной форм. Успешному решению этой проблемы способствовало открытие мужской стерильности свеклы. Стерильность триплоидных гибридов может иметь положительное значение при получении бессемянных плодов, например винограда и арбуза.
Однако триплоиды стерильны, поэтому необходимо каждый раз получать гибридные семена от скрещивания диплоидной и тетраплоидной форм. Успешному решению этой проблемы способствовало открытие мужской стерильности свеклы. Стерильность триплоидных гибридов может иметь положительное значение при получении бессемянных плодов, например винограда и арбуза.
Ценные результаты дает использование в селекции явления аллополиплоидии, основой которого служит метод отдаленной гибридизации, т. е. скрещивания организмов, относящихся к разным видам и даже родам. Например, получены межвидовые полиплоидные гибриды капусты и редьки, ржи и пшеницы. Гибридизация пшеницы (Triticum) и ржи (Secale) позволила получить ряд форм, объединенных общим названием тритикале. Они обладают высокой урожайностью пшеницы и зимостойкостью и неприхотливостью ржи, устойчивостью ко многим болезням, в том числе к линейной ржавчине, являющейся одним из главных факторов, ограничивающих урожайность пшеницы.
На основе гибридизации пшеницы и пырея российским академиком Н.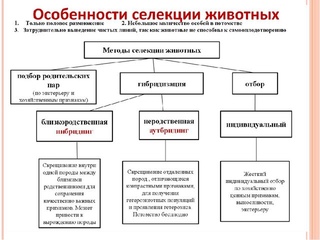 В. Цициным получены пшенично-пырейные гибриды, обладающие высокой урожайностью и устойчивостью к полеганию. Однако отдаленные гибриды, как правило, бесплодны. Это связано с содержанием в геноме различных хромосом, которые в мейозе не конъюгируют. Для восстановления плодовитости у межвидовых гибридов в 1924 г. советский генетик Г. Д. Карпеченко предложил использовать удвоение числа хромосом (полиплоидию) у отдаленных гибридов.
В. Цициным получены пшенично-пырейные гибриды, обладающие высокой урожайностью и устойчивостью к полеганию. Однако отдаленные гибриды, как правило, бесплодны. Это связано с содержанием в геноме различных хромосом, которые в мейозе не конъюгируют. Для восстановления плодовитости у межвидовых гибридов в 1924 г. советский генетик Г. Д. Карпеченко предложил использовать удвоение числа хромосом (полиплоидию) у отдаленных гибридов.
Г. Д. Карпеченко проводил скрещивание редьки и капусты. Число хромосом у этих растений одинаково (2п « 18). Соответственно их гаметы несут по 9 хромосом. Гибрид капусты и редьки имеет 18 хромосом, но он бесплоден, так как хромосомы капусты и редьки в мейозе не конъюгируют, поэтому процесс образования гамет не может протекать нормально. В результате удвоения числа хромосом в бесплодном гибриде оказалось 36 хромосом, слагающихся из двух полных диплоидных наборов редьки и капусты. Это создало нормальные возможности для мейоза; хромосомы капусты и хромосомы редьки конъюгировали между собой. Каждая гамета несла по одному гаплоидному набору редьки и капусты (9 + 9 = 18). В зиготе вновь оказалось 36 хромосом; межвидовой гибрид стал плодовитым. По фенотипу этот новый растительный организм совмещал признаки редьки и капусты, например в строении стручка.
Каждая гамета несла по одному гаплоидному набору редьки и капусты (9 + 9 = 18). В зиготе вновь оказалось 36 хромосом; межвидовой гибрид стал плодовитым. По фенотипу этот новый растительный организм совмещал признаки редьки и капусты, например в строении стручка.
Получение экспериментальным путем полиплоидных животных представляет большую трудность, поэтому такие формы животных — редкость. Так, советскому ученому генетику Б. JI. Астаурову путем межвидовой гибридизации удалось получить полиплоидную форму тутового шелкопряда. На сегодняшний день есть уже полиплоидные рыбы, птицы (например, куры), однако внедрение полиплоидных пород животных в практику сельского хозяйства — дело будущего.
Спонтанный и индуцированный мутагенез. Спонтанные мутанты используются преимущественно в селекции растений. Так, на основе мутанта желтого безалкалоидного люпина получено несколько сортов сладкого люпина, которые выращивают на корм скоту. Люпин, содержащий алкалоиды, для этой цели непригоден, поскольку животные его не едят.
Большое число мутантов известно у плодовых культур, которые используются как новые сорта или в гибридизации с другими формами. Один из наиболее известных спонтанных мутантов кукурузы opaque, отличающийся высоким содержанием аминокислоты лизина в зерне, используется для создания так называемых высоколизиновых гибридов кукурузы.
В последние десятилетия во многих странах мира развернуты работы по получению индуцированных мутантов. Индуцированные рентгеновыми лучами мутанты были выделены у многих злаков (ячменя, пшеницы, ржи и др.). Они отличаются не только повышенной урожайностью, но и укороченным побегом. Такие растения устойчивы к полеганию и имеют заметные преимущества при машинной уборке. Кроме того, короткая и прочная соломина позволяет вести дальнейшую селекцию на увеличение размера колоса и массы семян без опасения, что повышение урожая зерна приведет к полеганию растений.
Особенно успешно индуцированный мутагенез применяют в селекции микроорганизмов.
«Основные методы селекции животных и растений» (10-11 класс)
ДЕПАРТАМЕНТ ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ ТОМСКОЙ ОБЛАСТИ
ОБЛАСТНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ
СРЕДНЕГО ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ
ТОМСКИЙ ИНДУСТРИАЛЬНЫЙ ТЕХНИКУМ
«Основные методы селекция животных и растений»
Методическая разработка
урока по биологии
Разработчик:
Л. А.Акобян
А.Акобян
Томск-2015г
Пояснительная записка к уроку
Урок по теме «Основные методы селекции животных и растений» разработан по естественно-научному направлению для обучающихся учебных заведений системы СПО, специальности «Защита информации» (на примере ОГБОУ СПО «ТомИнТех»). Урок проходит с использованием элементов фронтальной и индивидуальной работы, а так же ИКТ.
Использование данных методов на уроке биологии помогает достигнуть более высокого качества обучения в профессиональном образовании. Это позволяет не только повышать уровень знаний, умений, навыков обучающихся, но и развивать их как личности.
Использование индивидуальной, фронтальной работы позволяет развивать у обучающихся общие компетенции такие как: ОК 4. Осуществлять поиск информации, необходимой для эффективного выполнения задач, ОК 6. Работать в коллективе и команде, эффективно общаться с коллегами, руководством, потребителями и ОК 7.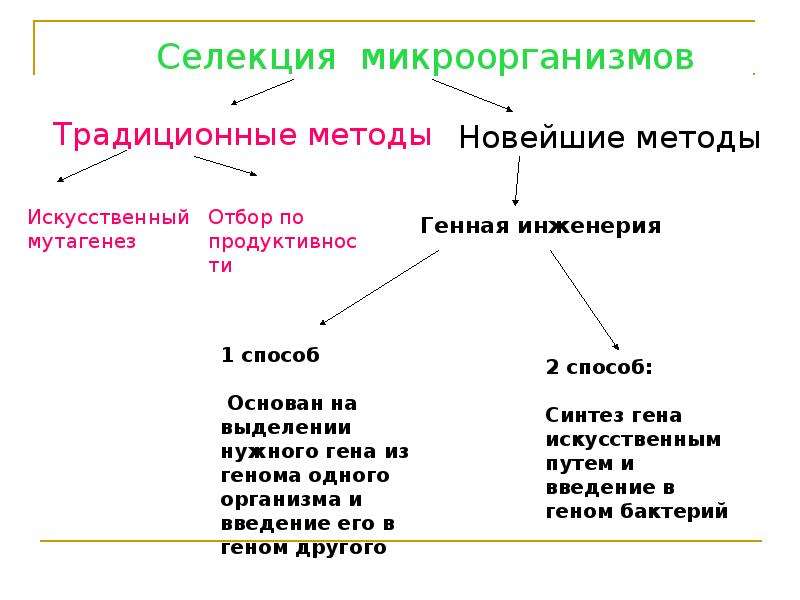 Брать на себя ответственность за работу членов команды (подчиненных), результат выполнения заданий. Развитие данных компетенций входит в перечень требования к результатам освоения программы подготовки специалистов по специальности «Защита информации» и необходимы обучающимся в их бедующей профессиональной деятельности.
Брать на себя ответственность за работу членов команды (подчиненных), результат выполнения заданий. Развитие данных компетенций входит в перечень требования к результатам освоения программы подготовки специалистов по специальности «Защита информации» и необходимы обучающимся в их бедующей профессиональной деятельности.
Особенности урока в том, что:
Совершенствование знаний по теме «Основные методы селекции животных и растений» проводится в интегрированной форме с использованием ИКТ, что способствует стимулированию интереса.
Тип урока: изучения нового материала позволяет показать все основные этапы урока, что демонстрирует профессионализм педагога.
Прослеживается связь с общеобразовательными предметами: география, зоология, ботаника.
Обучающиеся знакомятся с особенностями методов селекции животных и растений, о новых породах и сортах.
 Многие из них узнают в гибридных образцах своих домашних питомцев. Это не только повышает интеллектуальный уровень развития обучающихся, но и понимание значимости предмета биологии.
Многие из них узнают в гибридных образцах своих домашних питомцев. Это не только повышает интеллектуальный уровень развития обучающихся, но и понимание значимости предмета биологии.С некоторыми современными продуктами селекции, ребята сталкиваются в обычной жизни. На прилавках магазинов выставлены товары с надписью « Не содержит ГМО». Проводя аналогию с профессией на которую они обучаются, возникает вопрос о значимости защиты каждого человека о спорных продуктах селекции.
В ходе урока учитывается разные индивидуальные способности обучающихся, поэтому используются разные формы организации учебно-познавательного процесса.
Финалом урока является самостоятельная работа в форме «виртуальной» экскурсии, которая выявляет уровень усвоения и закрепления данного материала каждым обучающимся, а так же решением биологических задач.
Урок состоит из шести этапов.
На первом этапе преподаватель осуществляет проверку готовности обучающихся к уроку, сообщает тему и цели урока, мотивирует их на работу, знакомит с особенностями проведения урока и формой работы.
На втором этапе преподаватель проверяет уровень усвоение пройденного материала путем индивидуальной работы у доски, индивидуальной работы по карточкам и фронтальной работы повторением основных терминов, путем разгадывания кроссворда.
На третьем этапе объяснение нового материала с использованием мини-лекции и заполнением опорной схемы.
Закрепление полученных знаний осуществляется на четвертом этапе. Здесь студентам демонстрируется «виртуальная» экскурсия по гибридным образцам, и ребята должны самостоятельно определить какой из методов селекции был применен. Так же предлагается решить ряд биологических задач.
На пятом этапе рекомендуется провести рефлексию урока с целью осуществления «обратной связи» и сделать выводы об успешности проведённого урока.
На заключительном этапе происходит подведение итогов урока, выдача домашнего задания.
Методика всего урока и каждого отдельного этапа вовлекает обучающихся в работу, активизирует их мыслительную деятельность, развивает познавательную активность.
Технологическая карта урока
Этапы занятия
Цель этапа
Время
Содержание деятельности
Средства
Формы и методы
Преподаватель
Обучающийся
1.Организационный момент
Организовать и настроить обучающихся на работу.
5
Проверяет явку и готовность обучающихся к уроку. Сообщает тему, цели и этапы урока. Реализует мотивационный компонент. Актуализация знаний.
Знакомятся с темой и целью урока. Формой работы.
Эпиграф
Фронтальная.
2.Повторение пройденного материала
Определить насколько успешно был освоен предыдущий материал.
10
1.Проверяет знания студентов у доски по карте.
2.Раздает карточки с заданием «Центры происхождения культурных растений».
3.Проверяет терминологию пройденного материала, используя кроссворд. Задает вопросы.
1.Слушают друг друга, дополняют материал.
2.Выполняют задание по карточкам, проверяют друг у друга.
3. Разгадывают кроссворд.
— Политическая карта мира.
— Карточки с заданием «Центры происхождения культурных растений»
-Кроссворд (презентация)
1.Индивидуальная у доски
2.Индивидуальная по карточкам
3.Фронтальная
3. Усвоения новых знаний
Сформировать конкретные представления об изучаемых понятиях. Выявить сущность и главное в изучаемом.
15
Рассказывает о методах селекции. Просит конспектировать основные моменты.
Слушают материал и заполняют опорную таблицу по теме.
-таблица «Основные методы селекции растений и животных»
— Презентация
1. Мини-лекция
Мини-лекция
2.Самостоятельная работа с конспектом.
4. Закрепление полученных знаний
Организовать деятельность учащихся по применению знаний, используя индивидуальную работу.
10
1. Предлагает слово одному из студентов, который провел исследовательскую работу по аутбридингу.
2. Зачитывает биологические задачи.
1.Студенты самостоятельно заполнят таблицу и определяют, какой из методов аутбридинга был применен для получения данного гибрида.
2. Внимательно слушают задачи и определяют метод гибридизации.
-презентация «Виртуальная экскурсия»
— карточка с заданием №1 «Прослушайте презентацию «Виртуальная» экскурсия. Заполните таблицу».
— карточка с заданием №2 «Решите задачу».
1.Интегрированный
2.Индивидуальный
5. Рефлексия
Уметь анализировать деятельность.
3
Зачитывает вопросы со слайда. При необходимости задает наводящие вопросы.
При необходимости задает наводящие вопросы.
Анализируют свою деятельность, выявляют причины возникающих затруднений и пути их преодоления.
— презентация
Фронтальная беседа
5. Домашнее задание.
Сориентировать учащихся на возможные затруднения, на правильное выполнение задания.
2
Выдает домашнее задание.
Записывают основные моменты домашнего задания.
— презентация
Сообщение
Тема: Основные методы селекции растений и животных
Как приятно знать, что ты что-то узнал.
Мольер.
Цель: познакомится с методами, применяемыми в селекции растений и животных.
Задачи:
Тип урока: урок изучения нового материала.
Вид урока: Уроки совершенствования знаний, умений и навыков.
Демонстрационное оборудование: мультимедийный проектор, презентация PowerPoint с изображениями животных, распечатанные карточки с заданиями, опорная таблица по теме урока.
План урока:
I.Организационный момент — 5 мин.
II. Этап проверки домашнего задания – 10 мин.
III. Объяснение нового материала — 15 мин.
IV. Закрепление полученных знаний – 10 мин.
V. Рефлексия – 3 мин.
VI Домашнее задание – 2 мин.
ХОД УРОКА
I. Организационный момент 2 мин (Тема, отсутствующие) слайд 1.приложение 4.
– Здравствуйте ребята, сегодня на уроке нам нужно изучить очень объемный материал. Поэтому давайте максимально сконцентрируемся на уроке и будем работать быстро. Итак, давайте вспомним к изучению какой темы мы приступили на прошлом уроке? (Селекция)
– Сегодня на уроке мы с вами должны:
– повторить основные понятия темы «Селекция»;
– углубить материал по данной теме;
– а также вы должны показать мне какие вы трудолюбивые, добрые и внимательные.
– Вы готовы это сделать? (Да)
II. Этап проверки домашнего задания 10 мин
–слайд 2. Итак, скажите пожалуйста, какая основная задача селекции как науки? (Выведение новых и совершенствование старых сортов растений, пород животных, штаммов микроорганизмов.)
Итак, скажите пожалуйста, какая основная задача селекции как науки? (Выведение новых и совершенствование старых сортов растений, пород животных, штаммов микроорганизмов.)
– Хорошо. А скажите, какая наука является теоретической базой селекции? (Генетика)
– А теперь подумайте, почему именно генетические знания играют большую роль в развитии селекции?(Потому что основой успеха селекционной работы в значительной степени является генетическое разнообразие исходного материала. В своей работе селекционеры стараются использовать все многообразие диких и культурных растений.)
слайд 3.Правильно. На необходимость использовать в селекции растений все видовое многообразие флоры нашей планеты указывал еще академик Николай Иванович Вавилов – выдающийся генетик и селекционер. Именно Николаем Ивановичем были выделены 8 центров происхождения культурных центров растений.
Слайд 4. Давайте вспомним, какие это центры. Преподаватель вызывает одного студента к доске, и он по карте, (приложение 1), показывает все центры происхождения культурных растений.
Слайд 5 Итак, мы с вами вспомнили центры происхождения, а теперь давайте вспомним, родиной каких растений является тот или иной центр. Обратите внимание, на слайде перечислены культурные растения, такое же задание и на карточках на ваших столах (приложение 2). Ваша задача записать на карточках центры их происхождения. (Слайд 5)
Учащиеся заполняют карточки.
– Время вышло. Поменяйтесь карточками. На след. Слайде 6 красным цветом обозначены правильные ответы. Отметьте знаком «+» правильные ответы, знаком «–» – неверные.
– Кто сколько ошибок допустил?
Учащиеся называют количество ошибок.
– Для того чтобы дальше продолжать изучение темы нам нужно повторить основные термины. Мы будем с вами разгадывать кроссворд. У меня к вам просьба, не выкрикивайте ответы с места, а поднимайте руки, если знаете правильный ответ т.к. самые активные получать за урок оценки. (Слайды 7-15)
Вопросы:
В. Наука о методах создания новых и улучшение существующих пород животных, сортов растений, штаммов микроорганизмов с полезным для человека свойствам (селекция)
Г.
 Так называется популяция растений искусственно созданная человеком? (Сорт)
Так называется популяция растений искусственно созданная человеком? (Сорт)Г. Как называется метод при котором проводят различные скрещивания организмов? (Гибридизация)
По горизонтали:
В. В основе этого метода, который используется до сих пор лежит концепция разработанная еще Ч.Дарвиным.(отбор)
В. Так называется популяция животных искусственно созданная человеком?
В. Так называется популяция микроорганизмов искусственно созданная человеком?
– Таким образом, какие 2 основных метода выделяют в селекции? (Отбор и гибридизация)
– Какие объекты изучают с помощью отбора и гибридизации? (Животных, растений, микроорганизмы)
– А вам известны методы селекции этих живых организмов? (Нет)
– Именно сегодня на уроке мы и познакомимся с особенностями селекции животных и растений, методами, применяемыми в их селекции, а о селекции микроорганизмов поговорим на следующем уроке.
– Откройте тетради и запишите тему сегодняшнего урока – «Основные методы селекция животных и растений» (Слайд 16)
Слайд 17. Проговорить цель урока.
Объяснение нового материала.
Слайд 18 Ребята! У вас на столах лежат опорные таблицы «Современные методы селекции» приложение 3. По ходу моей лекции вы должны заполнять пустые ячейки. Это таблица останется у вас, и вы сможете ее вклеить в тетрадь.
Итак, разгадывая кроссворд мы с вами определили два основных метода селекции: это гибридизация и отбор. Теперь более подробнее поговорим о каждом методе.
Примитивная селекция растений возникла одновременно с земледелием. Начав возделывать растения, человек стал отбирать, сохранять и размножать лучшие из них. Многие культурные растения возделывались примерно за 10 тысяч лет до нашей эры. Древние селекционеры создали прекрасные сорта плодовых растений, винограда, многие сорта пшеницы, породы домашних животных. Им были известны некоторые современные селекционные приемы. Например, искусственное опыление финиковой пальмы применяли в Египте и Месопотамии за несколько веков до н.э.
Им были известны некоторые современные селекционные приемы. Например, искусственное опыление финиковой пальмы применяли в Египте и Месопотамии за несколько веков до н.э.
Слайд 19.Самый первый и самый древний метод селекции – искусственный отбор. Различают две формы: массовый и индивидуальный. Массовый отбор – это выделение группа особей по внешним признакам. Например, при массовом отборе из всей популяции кур в хозяйстве оставляют для размножения птиц с яйценоскостью 200-250 яиц в год, живой массой не менее 1,5 кг, определенной окраски, не проявляющих инстинкта высиживания и т.д. все остальные куры выбраковываются. Достоинства: простота, экономичность, недостатки – невозможность индивидуальной оценки по потомству.
Слайд 20. Значение индивидуального отбора велико в том случаи, когда от одного организма можно получить большое количество потомков. Применяя искусственное осеменение от одного быка можно получить до 35000 телят.
Слайд 21. Так же различают два основных метода гибридизации: инбридинг и аутбридинг.
Так же различают два основных метода гибридизации: инбридинг и аутбридинг.
Инбридинг – родственная гибридизация. Скрещивание между братьями и сестрами или между родителями и потомством ведет к гомозиготности, т.е. к чистым линиям, и часто сопровождается ослаблением животных, уменьшению их устойчивости к неблагоприятным условиям, снижению плодовитости, а чтобы разнообразить генотип, насыщать его различными аллелями, Тем не менее инбридинг применяют с целью закрепления в породе характерных хозяйственно ценных признаков.
Слайд 22-25Аутбридинг – неродственная гибридизация. Это скрещивание сопровождается строгим отбором, что позволяет усилить и поддерживать ценные качества породы.
Сочетание родственной и неродственной гибридизации широко применяется селекционерами для выведения новых пород животных.
Слайд 26. Теперь давайте сверим таблицу «методы селекции» с тем, что получилось у вас. Исправьте ошибки если они есть.
Слайд 27. Привести примеры современной селекции.
Закрепление полученных знаний
Слайд 28. Ребята! Сейчас мы с вами прослушаем сообщение, которое приготовила нам Роза Махмудова. Она изучила большое количество материала по результатам аутбридинга, ваша задача внимательно слушать и заполнить таблицу с заданием №1. (приложение 4)
Слайд 29. Виртуальная экскурсии приложение 5.
Сегодня мы попробуем провести «виртуальную» экскурсию среди некоторых пород домашних животных растений, а заодно и вспомним основные методы селекции животных.
Слайд 30. Проверим заполненную таблицу. Подсчитайте количество баллов.
Слайд 31. Перед вами приложение 4 задание №2 «Решение биологических задач»:
В 1760-е годы английский селекционер Р. Бейкуэлл сформулировал два правила селекции крупного рогатого скота: «Скрещивай лучшее с лучшим» и «Подобное рождает подобное». Трудами этого специалиста Англия во многом обязана своим лидирующим положением в племенном животноводстве.
 О каких методах селекции идёт речь в данных высказываниях?
О каких методах селекции идёт речь в данных высказываниях?Знаменитый русский селекционер И.В. Мичурин вывел более 300 сортов плодовых и ягодных культур, на выведение же одного сорта требуется не менее 20 лет. Поэтому жизнь этого учёного – беспримерный подвиг, пример колоссального трудолюбия и патриотизма. Мало кому известно, что Мичурин занимался и селекцией цветов – роз, лилий. Голландцы предлагали большие деньги за лилию фиалкоцветную. Не продал… А каков метод получения этого растения, излюбленный мичуринский метод?
Всеми любимый виноград сорта кишмиш не имеет семечек, обладает раннеспелостью и приятным вкусом. Сорт Кишмиш Чёрный и сорт Кишмиш Белый овальный обладают хромосомным набором 4 п. Как называются такие растения, как можно получить растения с удвоенным набором хромосом?
Слайд 32. Проверим заполненную таблицу. Подсчитайте количество баллов. Поставьте оценку. Ребята, с последних парт передайте мне все карточки, я их проверю еще раз и выставлю оценку за урок.
Слайд 33. Ответьте на вопросы:
V. Рефлексия.
1. Что нового вы узнали на уроке?
2. Как вы считаете, какой из методов более эффективный?
3. Какие новые гибриды вы могли бы предложить?
4. Как вы считаете, достигли ли мы цели урока?
VI. Домашнее задание (Слайд 34)
– Подготовить сообщение об истории создания породы ваших домашних питомцев;
— Повторить основные понятия генетики: ген, гетерозигота, гетерозис, фенотип, экстерьер, сорт, штамм, биотехнология и т. д.
Слайд 35. Урок хочется закончить анализом знаменитого мичуринского высказывания «Мы не можем ждать милостей от природы, взять у неё – наша задача» Но мало кто знает продолжение этой фразы: «Но к природе надо относиться бережно и по возможности сохранять её в первозданном виде». В наш сегодняшний век – век манипулирования генетическим материалом, актуальнее вторая часть мичуринского завета, поэтому перед селекционерами сейчас важнейшими являются и нравственные задачи.
Таким образом, сегодня на уроке, повторили основные понятия темы «Селекция», познакомились с методами селекции растений и методами селекции животных
Слайд 36. Спасибо за урок. До свидания.
Список литературы:
Биология в таблицах и схемах. Для школьников и абитуриентов. Изд. 2-е. СПб, ООО «Виктория плюс», 2012.-128стр.
С.Г. Мамонтов Биология для поступающих в вузы, М. Высшая школа, 1992.
Новый справочник школьника. 5-11 класс. Универсальное пособие. Т.2.-СПб.: ИД «ВЕСЬ», 2002 год.
Научно-теоретический и методический журнал Министерства образования Российской Федерации. Биология в школе, 1993 год 1 издание.
Популярный энциклопедический иллюстрированный словарь — ЕВРОПЕДИЯ. Олма медиагрупп. Олма-пресс 2004.
Энциклопедический справочник школьника. Естественные науки. Москва 2002.
Приложение 1. Карта с изображением центров происхождения культурных растений
Приложение 2.
Фамилия _____________
Центры происхождения культурных растений
Рис, сахарный тростник, цитрусовые, баклажаны и др. (50% культурных растений).__________________________________________________________________________
Соя, просо, гречиха, плодовые и овощные культуры – слива, вишня и др. (20% культурных растений). __________________________________________________________________________
Пшеница, рожь, бобовые культуры, лен, конопля, репа, чеснок, виноград и др. (14% культурных растений). _______________________________________________________________
Капуста, сахарная свекла, маслины, клевер (11% культурных растений).__________________________________________________________________________
Твердая пшеница, ячмень, кофейное дерево, бананы, сорго. ___________________________________________________________________________________
Кукуруза, какао, тыква, табак, хлопчатник.
 ________________________________________
________________________________________Картофель, ананас, хинное дерево. ______________________________________________
Название центров: Южноазиатский тропический, Юго-западноазиатский, Центральноамериканский Среднеземноморский, Абессинский, Восточноазиатский, Южноамериканский
.
Приложение 3. Опорная таблица «Современные методы селекции»
Приложение 4.
Группа______ ФИ____________
Задание 1. Прослушайте презентацию «Виртуальная» экскурсия. Заполните таблицу.
Гибриды
Метод
Баллы
1.
Ландарас
2.
Тигролев
3.
Ларка
4.
Австралийский шелковистый терьер
5.
Левопард
6.
Культурная слива
7.
Британская короткошерстная
8.
Лигр
9.
Сильванер
10.
Хайнак
Итого:
Задание 2. Решите задачу.
Задача
Ответ
Балл
1.
В 1760-е годы английский селекционер Р. Бейкуэлл сформулировал два правила селекции крупного рогатого скота: «Скрещивай лучшее с лучшим» и «Подобное рождает подобное». Трудами этого специалиста Англия во многом обязана своим лидирующим положением в племенном животноводстве. О каких методах селекции идёт речь в данных высказываниях?
2.
Знаменитый русский селекционер И.В. Мичурин вывел более 300 сортов плодовых и ягодных культур, на выведение же одного сорта требуется не менее 20 лет. Поэтому жизнь этого учёного – беспримерный подвиг, пример колоссального трудолюбия и патриотизма. Мало кому известно, что Мичурин занимался и селекцией цветов – роз, лилий. Голландцы предлагали большие деньги за лилию фиалкоцветную. Не продал… А каков метод получения этого растения, излюбленный мичуринский метод?
Поэтому жизнь этого учёного – беспримерный подвиг, пример колоссального трудолюбия и патриотизма. Мало кому известно, что Мичурин занимался и селекцией цветов – роз, лилий. Голландцы предлагали большие деньги за лилию фиалкоцветную. Не продал… А каков метод получения этого растения, излюбленный мичуринский метод?
3.
Всеми любимый виноград сорта кишмиш не имеет семечек, обладает раннеспелостью и приятным вкусом. Сорт Кишмиш Чёрный и сорт Кишмиш Белый овальный обладают хромосомным набором 4 п. Как называются такие растения, как можно получить растения с удвоенным набором хромосом?
Итого:
Итого: Критерии оценки: 12-13 баллов – отлично, 11-10 баллов – хорошо; 8-9 баллов – удовлетворительно; менее 7 – неудовлетворительно
Приложение 4. Презентация к уроку.
Слайд 1.
Слайд 5.
Слайд 2.
Слайд 6.
Слайд 3.
Слайд 7.
Слайд 4.
Слайд 8.
Слайд 9.
Слайд 13.
Слайд 10.
Слайд 14.
Слайд 11.
Слайд 15.
Слайд 12.
Слайд 16.
Слайд 17.
Слайд 21.
Слайд 18.
Слайд 22.
Слайд 19.
Слайд 23.
Слайд 20.
Слайд 24.
Слайд 25.
Слайд 29.
Слайд 26.
Слайд 30.
Слайд 27.
Слайд 31.
Слайд 28.
Слайд 32.
Слайд 33.
Слайд 34.
Слайд 35.
Слайд 36.
Приложение 5. Презентация «Виртуальная экскурсия».
Слайд 1.
Слайд 5.
Слайд 2.
Слайд 6.
Слайд 3.
Слайд 7.
Слайд 4.
Слайд 8.
Слайд 9.
Слайд 13.
Слайд 10.
Слайд 11.
Слайд 12.
Современные генетические методы в селекции растений Текст научной статьи по специальности «Сельское хозяйство, лесное хозяйство, рыбное хозяйство»
Александр Кильчевский Елена Сычёва
директор Института генетики и цитологии НАН Беларуси, член-корреспондент
ученый секретарь Института генетики и цитологии НАН Беларуси, кандидат биологических наук
Современные генетические методы в селекции растений
Крылатые слова академика Н.И. Вавилова о том, что селекцию можно рассматривать как науку, как искусство и как отрасль сельскохозяйственного производства, сохраняют свою актуальность [1]. Генетика в свою очередь является теоретическим фундаментом селекции как науки. в последнее время арсенал методов селекции значительно пополнился за счет прорывных достижений в области генетики, геномики и биотехнологии, определив основные приоритеты современной селекции растений и расширив пути их достижения (табл. 1).
Расширение спектра генетической изменчивости
важнейшую роль в селекции играет анализ существующей генетической изменчивости, а также создание новой — в результате мутагенеза, рекомбино-генеза и трансгенеза. генетические ресурсы в мире хранятся в более чем 1,5 тыс. генбанков, где сосредоточено около 6 млн. образцов растений. Поиск доноров и источников хозяйственно ценных признаков, их анализ, формирование рабочих коллекций — важнейшая задача селекционера. Если ценные признаки у исходных образцов сцеплены с нежелательными, создать новые группы сцепления можно с помощью рекомби-
ногенеза. весьма часто одного цикла гибридизации бывает недостаточно для того, чтобы добиться оптимального сочетания желаемых генов. Этой цели служит периодический отбор, включающий несколько стадий. Такого рода селекция напоминает выпаривание раствора соли в сосуде. Технологии ее проведения разработаны за рубежом и усовершенствованы в Институте генетики и цитологии для отдельных перекрестноопыляющихся (кукуруза) и самоопыляющихся культур (томат) [2].
новые возможности для расширения спектра генетической изменчивости и создания принципиально новых генотипов растений открывает генетическая инженерия. Трансгенные технологии позволяют получать формы растений, толерантные к гербицидам, передавать устойчивость к насекомым и вирусам, улучшать качественные характеристики. Многие из этих задач сложно или практически невозможно решить с помощью традиционной селекции.
В Беларуси сорта трансгенных растений пока не выращиваются. Однако исследования в этой области проводятся достаточно динамично по целому ряду направлений на базе академических институтов — генетики и цитологии, биофизики
и клеточной инженерии, — НПЦ по картофелеводству и плодоовощеводству, а также центрального ботанического сада и БГУ. Получены первичные трансгенные растения (рис. 1), представляющие значительный научный и практический интерес [3]. В рамках Государственной программы «Инновационные биотехнологии» ведутся работы по созданию
¡,с
Рис. 1. Трансгенные растения картофеля, устойчивые к насекомым, пока еще в пробирке: А — контрольное растение картофеля сорта Скарб на селективной среде; Б — контрольное растение картофеля сорта Скарб на среде без антибиотика; В — трансгенное растение картофеля сорта Скарб на селективной среде
СЕЛЕКЦИЯ КУЛЬТУРНЫХ РАСТЕНИЙ
трансгенных форм картофеля с устойчивостью к колорадскому жуку, вирусам, грибным и бактериальным болезням; высокоурожайного клевера; клюквы с повышенной резистентностью к патогенам и измененным вкусом плодов. В планах — получение генно-модифици-рованных рапса и льна. Всего к 2012 г. планируется получить 9 первичных трансгенных линий растений с заданными свойствами по 5 сельскохозяйственным культурам.
Успехи в этом направлении ощутимы, но говорить об отечественных трансгенных сортах еще рано, и пройдет не один год экспериментов, испытаний и анализа, прежде чем они дойдут до промышленного производства. Такой взвешенный, продуманный подход вполне обоснован, принимая во внимание новизну и недостаточный опыт использования достижений в этой области. Исключение возможных биологических рисков при применении ГМО в долгосрочной перспективе требует проведения широкомасштабных исследований. С этой целью в Институте генетики и цитологии реализуются проекты по оценке биобезопасности трансгенных растений для здоровья человека, животных и окружающей среды и созданию полигона для их испытания.
Повышение эффективности отбора
Методы отбора генотипов, применяемые в селекции, весьма разнообразны и зависят от биологии размножения растения, особенностей наследования признака, гибридного поколения, условий среды и других факторов. Важнейшее условие эффективного отбора—точная идентификация генотипа по фенотипу. В случае, когда речь идет о качественных признаках, за наследование которых «отвечают» 1—2 гена, отбор относительно прост. однако большинство хозяйственно ценных признаков являются количественными, и формирование их зависит от среды. В этом случае значе-
ние генотипа можно определить только как среднее значение всех фенотипов в изменяющихся условиях. Для повышения эффективности отбора по количественным признакам применяется разработанная в Институте генетики и цитологии экологическая организация селекционного процесса [2].
Существенно повышают эффективность отбора и селекционного процесса современные технологии молекулярного маркирования, с помощью которых можно выявлять доноров агрономически важных признаков, маркировать гены устойчивости к болезням и другим биотическим и абиотическим факторам, идентифицировать сорта и др. В настоящее время в нашем институте мар-кер-ассоциированная селекция ведется более чем по 30 генам устойчивости и качества у растений (рис. 2). Преимущества отбора с использованием ДНК-маркеров по сравнению с традиционными методами налицо: возможность
идентифицировать необходимые гены на ранних стадиях развития организма еще до начала их экспрессии, быстро и эффективно тестировать большое количество селекционного материала значительно сокращает занимаемые площади и снижает трудозатраты.
на основе ДнК-маркеров разработана система идентификации и паспортизации сортов растений для проверки соответствия новых сортов критериям ооС-теста (отличимость, однородность и стабильность), решения спорных вопросов об их чистоте, авторстве и т.д. Система адаптирована для 8 основных сельскохозяйственных культур, возделываемых в стране: пшеницы, ячменя, картофеля, льна, томата, сои, подсолнечника, яблони. Ведутся работы на сахарной свекле. С целью освоения ДНК-паспортизации утверждены методические рекомендации и предложена схема государственного контроля закупки семян с использованием ДНК-паспортов (рис. 3).
со
Рис. 2. Результаты ДНК-маркирования сортов картофеля по устойчивости к цистообразующей нематоде (1, 4—6, 8—10 — сорта с геном h2, определяющим устойчивость к патотипу Ro1 цистообразующей картофельной нематоды)
Таблица 1. Основные приоритеты современной селекции растений
Приоритеты Пути достижения
I. Расширение спектра генетической изменчивости Создание и использование генбанков; мутагенез; рекомбиногенез; трансгенез
II. Повышение эффективности отбора Селекция с помощью молекулярных маркеров; экологическая организация селекционного процесса
III. Повышение информативности селекционного процесса Генетико-статистические методы; информационные технологии, компьютеризация селекции
IV. Сокращение сроков создания сортов и гибридов Использование фитотронов; биотехнологические методы (гаплоидия, клональное микроразмножение ценных генотипов и др.)
С целью обобщения пятидесятилетнего труда белорусских ученых по разработке генетически обоснованных методов селекции Институтом генетики и цитологии HAH Беларуси реализуется на практике амбициозная инициатива по изданию не имеющей аналогов в СНГ четырехтомной коллективной монографии «Генетические основы селекции растений». На настоящий момент опубликованы два тома: «Общая генетика растений» и «Частная генетика растений». Готовится к изданию третий — об использовании в селекции растений методов клеточной инженерии.
Совершенно очевидно, что ДНК-паспортизация выводит сортоиспытание и семеноводство на принципиально новый уровень. Ее внедрение позволит повысить качество селекционно-семеноводческого процесса в Беларуси и эффективность контроля за создаваемыми в республике сортами, улучшить систему патентования новых, усилить контроль за кондиционностью семян, закупаемых за рубежом, исключить возможность фальсификации и связанных с этим экономических потерь.
Повышение информативности селекционного процесса
Важнейшим инструментом повышения эффективности селекционного процесса является информатизация. На стыке ряда наук — биометрии, генетики, селекции — сформировались генети-ко-статистические методы, в развитие которых Институт генетики и цитологии внес значительный вклад. Разработаны и усовершенствованы способы оценки комбинационной способности в селекции на гетерозис, адаптивной способности — на повышение адаптивного потенциала, пакеты прикладных про-
грамм для статистического анализа результатов селекционных и генетических экспериментов РИШОН и АБСТАТ. Их использование существенно облегчает труд ученых. Создание автоматизированного рабочего места (АРМ) селекционера — программного комплекса на базе современных информационных технологий — ближайшая задача. Она будет решаться специалистами института совместно с коллегами из НПЦ НАН Беларуси по земледелию.
Сокращение сроков создания сортов и гибридов
Интенсификация растениеводства привела к быстрой сортосмене. Продолжительность жизни сорта постоянно сокращается, и селекционер должен мгновенно отвечать на вызовы времени, подключая все возможности для ускорения селекционного процесса. Среди методов, позволяющих сократить сроки создания сорта — гетерозисная и гамет-ная селекция, использование культуры in vitro, применение фитотронов и молекулярных маркеров, селекция на основе гаплоидов.
Селекция на гетерозис позволяет объединить в гибридном генотипе комплекс желаемых генов, что не всегда удается при выведении константных сортов. Институт генетики и цитологии традиционно лидирует среди научных учреждений стран СНГ по вопросам методологии гетерозисной селекции растений. С использованием разработанных подходов только за 2004— 2009 гг. при участии института в Госсортоиспытание переданы 13 гибридов (9 — томата, 2 — подсолнечника, по одному — перца и капусты). Всего за историю существования института создано более 30 сортов, районированных в Беларуси и за ее пределами.
Селекция на основе гаплоидов позволяет быстро гомозиготизировать гибридный материал, добиться его выравнен-ности, что дает возможность сократить длительность селекционного процесса на 3—4 года и уменьшить затраты на выведение новых сортов. Такие технологии разработаны в институте для сахарной свеклы, картофеля, тритикале, льна-долгунца [4].
Важный резерв ускорения селекции и повышения ее эффективности — гамет-ное направление исследований. В Институте генетики и цитологии совместно с БГСХА разработан метод гаметофит-но-спорофитной селекции томата на холодостойкость, в БГу — метод гаме-тофитной селекции люпина и томата на устойчивость к фузариозу [5].
В целом следует отметить, что традиционная селекция за последние годы существенно дополнена методами биотехнологии, генетики, экологии, информатики. Значит ли это, что традиционный отбор будет заменен селекцией в пробирках и фитотронах? Конечно же, нет. однако применение современных генетико-биотехнологических подходов создает принципиально новые возможности. Свидетельство тому — широкое применение гетерозисных гибридов в защищенном грунте и полевых условиях, расширение посевов трансгенных культур, новых культур (тритикале и др.), полученных при отделенной гибридизации. успех селекции сегодня — это результат творческого союза селекционеров с генетиками, биотехнологами, фитопатологами, физиологами, экологами, математиками и представителями других специальностей.
Литература
1. Теоретические основы селекции растений. Т. 1. Общая селекция растений / Под общ. ред. Вавилова Н.И. — М., Л., 1935.
2. Генетические основы селекции растений. В 4 т. Т. 1. Общая генетика растений / Под науч. ред. Кильчевского А.В., Хотылевой Л.В. — М., 2008.
3. Исаенко Е.В., Шахбазов А.В., Картель Н.А. Генетическая модификация картофеля: достижения и перспективы // Вес. Нац. акад. навук Беларуа Сер. бiял. навук. №3, 2007. С. 110—115.
4. Генетические основы селекции растений. В 4 т. Т. 2. Частная генетика растений / Под науч. ред. Кильчевского А.В., Хотылевой Л.В. — Мн., 2010.
5. Кильчевский А.В., Лещина Н.Ю., Пугачева И.Г. Результаты селекции томата на холодостойкость и продуктивность с использованием гаметофит-но-спорофитного отбора // Овощеводство: Сб. науч. тр. Вып. 14. — Мн., 2008. С. 166—175.
Инновации в селекции
Петр ВАСЬКО, заместитель генерального директора по науке НПЦ НАН Беларуси по земледелию, кандидат биологических наук, доцент
Фестулолиум — новый перспективный вид многолетней злаковой травы, полученный путем межродового скрещивания овсяницы луговой или овсяницы тростниковой и райграса пастбищного или многоукосного с использованием экспериментальной полиплоидии. Нами разработаны новые сортообраз-цы культуры, которые характеризуются высоким содержанием белка — 18— 22%, низким содержанием клетчатки — 16—18% и концентрацией обменной энергии 11—11,8 МДж/кг сухого вещества при пастбищном использовании. При этом сбор белка с 1 га составил 1,4—1,8 т. Это самое лучшее сочетание качеств, ни у одной другой культуры его нет. Фактически в период пастбищной спелости, когда высота травостоя составляет 20—25 см, по уровню обменной энергии фестулолиум приравнивается к зерну кукурузы. Важно и то, что включение фестулолиума в клеверо-райграсовые пастбищные травосмеси позволяет формировать травостои с 7 циклами стравливания вместо 5—6.
Ядвига ПИЛЮК, руководитель лаборатории крестоцветных культур НПЦ НАН Беларуси по земледелию, кандидат сельскохозяйственных наук
Зимостойкие сорта озимого рапса, разработанные в нашей лаборатории, даже в сложнейших условиях этого года — при малом количестве снега и сильных морозах — перезимовали в Беларуси на 70%. Прежде в столь неблагоприятных условиях посеянное почти полностью погибало. Работы по созданию стрессоустойчивых сортов озимого рапса можно признать успешными. В последние годы сорта селек-
ции НПЦ НАН Беларуси по земледелию занимают более 95% от посевной площади рапса, новые внедряются очень быстро. Сейчас активно создаются новые гибриды, и отечественные хозяйства проявляют к ним большой интерес.
Нынешние успехи были бы невозможны без опоры на фундамент, созданный белорусскими исследователями за предыдущие два с половиной десятка лет научной работы, а также без активного участия в разработках молодых ученых — они составляют две трети сотрудников лаборатории.
Леонид ШИМАНСКИй, директор Полесского института растениеводства
В НПЦ по земледелию НАН Беларуси созданы гибриды кукурузы зернового направления, которые позволяют осуществить импортозамещение завозимых сортов западной селекции. Урожайность наших гибридов в условиях государственного сортоиспытания колеблется от 120 до 160 ц зерна на 1 га и до 300 ц сухого вещества. Это на 30—50% выше, чем у стандартных сортов. Кроме того, удалось получить уникальную холодостойкость и устойчивость к временной засухе, повышенное содержание сухого вещества, хорошую скороспелость. В условиях Беларуси это чрезвычайно важно. Надеемся, что уже в следующем году такая кукуруза будет выращиваться на полях не только в режиме испытаний. Достичь высоких показателей помогло то, что мы перестроили нашу селекционную практику. Исходим из параметров идеального сорта и подбираем родительские компоненты для создания гибридов интенсивного типа. Исходный материал, самоопыленные линии и т.п. — все белорусское. По этой разработке проводим процедуру патентования.
Подготовил Владимир ЛЕБЕДЕВ
Достижения и основные направления современной селекции
За последнее столетие селекционеры добились поразительных успехов. Урожайность зерновых повысилась в 10 раз. В развитых странах получают до 100 ц/га пшеницы, риса, кукурузы. Новые сорта картофеля дают почти 1 000 ц/га — это в четыре раза выше урожая прежних сортов. Успехи наблюдаются и в селекции других культур.
Путем гибридизации географически отдаленных форм и отбора академик П. П. Лукьяненко получил высокопродуктивные сорта кубанской пшеницы «безостая 1», «аврора», «кавказ». Академик В. Н. Ремесло вывел замечательные морозоустойчивые сорта озимой пшеницы «мироновская 808», «юбилейная 50», «харьковская 63». В разных регионах России (в Сибири, Поволжье) и за рубежом широко используются сорта яровой пшеницы, полученные А. П. Шехурдиным и В. Н. Мамонтовой: «саратовская 29», «саратовская 36», «саратовская 210». Саратовские сорта занимают более половины посевных площадей яровой пшеницы. «Саратовская 29» обладает прекрасными технологическими свойствами и служит стандартом хлебопекарных качеств.
Академик В. С. Пустовойт на Кубани получил сорт подсолнечника, содержащий в семенах до 50—52 % масла.
Серьезная проблема связана с сохранением культурных форм: возделывание лишь отдельных сортов резко сокращает генофонд, снижает приспосабливаемость. При изменении климата или по другим причинам сорт может исчезнуть. При селекции высокомасличных сортов подсолнечника на Кубани оказались отобранными особи с тенденцией к позднему созреванию. Эта тенденция стала развиваться, подсолнечник созревал все позже и, наконец, перестал вызревать до дождей, начал гнить на полях. Восстановить культурные сорта оказалось делом не легким: к тому времени сорта В. С. Пустовойта сменили по всему миру все другие сорта подсолнечника.
Значительный вклад в селекцию новых пород животных внес отечественный селекционер М. Ф. Иванов. Им была выведена одна из самых продуктивных в мире пород шерстно-мясных тонкорунных овец — «асканийский рамбулье», высокопродуктивная порода свиней «украинская степная белая», мясомолочная «костромская» порода коров. Для получения «асканийского рамбулье» были скрещены лучшие представители украинских мериносов с «американскими рамбулье». В результате девятилетней селекционной работы по скрещиванию привезенного из Англии выдающегося производителя «крупной белой» породы с лучшими местными породами была получена порода «украинская степная белая», которая по весу, скороспелости, плодовитости и качеству продукции не уступает «крупной белой», но прекрасно переносит местные условия.
Гибридизация с дикими видами придает культурным формам устойчивость к условиям среды и невосприимчивость к болезням. Гибрид тонкорунных и грубошерстных овец с диким бараном архаром — архаромеринос — может использовать высокогорные пастбища, недоступные обычным овцам. Проведена гибридизация яка с крупным рогатым скотом. В результате успешного применения гетерозиса выводят бройлерных цыплят. Межродовый гибрид белуги со стерлядью — бестер — неприхотлив и может выращиваться в непроточных водоемах.
Селекция микроорганизмов направлена на создание генетических линий (штаммов), обеспечивающих максимальную производительность полезных веществ. Продукты жизнедеятельности бактерий и одноклеточных эукариот (водорослей, дрожжей и плесневых грибов) находят применение в различных областях промышленности и медицины. На деятельности микроорганизмов основано брожение теста, получение большинства молочных продуктов, квасов, виноделие, пивоварение, квашение капусты, кормовых добавок, а также производство лекарств и биологически активных соединений.
С целью увеличения эффективности селекции диапазон наследственной изменчивости исходных организмов иногда удается расширить с помощью мутагенеза. У бактерий набор хромосом гаплоидный, поэтому каждая мутация проявляется в фенотипе уже в первом поколении, облегчая отбор. Большая скорость размножения позволяет быстро получить значительное потомство. Полученные штаммы подвергают многократному отбору с пересевом на питательные среды и контролем на образование требуемого продукта.
Использование данной технологии позволяет получать штаммы значительно более продуктивные, чем природные формы. Так, получены плесневые грибы, продуцирующие в тысячи раз больше антибиотика, чем исходные формы. Новые штаммы микроорганизмов синтезируют в необходимых для человечества количествах витамины В1, В12, которые неспособны вырабатывать организмы животных и человека.
Опорные точки:
Значительные успехи достигнуты в области районирования культурных растений в различных климатических условиях.
Размножение гибридных — мичуринских сортов осуществляется вегетативным путем.
В условиях непрерывного роста населения актуальным является выведение новых высокопродуктивных пород животных и сортов растений.
Проверь себя:
- Приведите примеры достижений отечественных селекционеров в области растениеводства.
- Расскажите о методах селекционной работы И.В.Мичурина.
- Привидите примеры достижений селекционеров нашей страны в области животноводства.
Произошла ошибка при настройке пользовательского файла cookie
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в cookie-файлах может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
границ | Треугольник современной селекции растений: оптимизация использования данных геномики, феномики и окружающей среды
Введение
Продолжающийся рост генетического прироста демонстрирует успех установленных государственных и частных программ селекции растений. Тем не менее, за последние два десятилетия было разработано все больше современных технологий, которые теперь ожидают эффективной интеграции в традиционные системы разведения. Эта интеграция предлагает привлекательные преимущества, но вместе с тем сопряжена с проблемами внесения изменений в существующие и действующие системы, недавним примером которых является селекция риса (Collard et al., 2019). Новые доступные технологии, быстрый цикл геномики (Crossa et al., 2017), фенотипирование с высокой пропускной способностью (HTP, phenomics) (Montesinos-López et al., 2017) и исторические описания взаимосвязи с окружающей средой (enviromics) (Costa-Neto et al. , 2020a, b; Resende et al., 2020; Rogers et al., 2021) имеют решающее значение для улучшения традиционных схем разведения и увеличения генетической выгоды. Интеграция этих новых технологий в повседневную селекционную систему поддержит поставку сортов с высокими урожаями в условиях ожидаемых неблагоприятных экологических условий в будущем, вызванных изменением климата и, как следствие, увеличением числа биотических и абиотических стрессов.Здесь мы кратко описываем использование этих технологий и их реализацию для обеспечения экономичных и экономящих время подходов к селекции растений. Мы также даем обзор взаимосвязей между этими методами. Наконец, мы представляем себе будущие перспективы для реализации более взаимосвязанного селекционного подхода, который использует преимущества так называемого современного треугольника селекции растений: интеграция геномики, феномены и окружающей среды.
Почему геномика для улучшения селекции?
Одно из самых популярных применений геномики в селекции — это предсказание селекционной ценности.Геномный отбор (GS) сокращает время цикла, увеличивает точность расчетных значений селекции и повышает точность отбора. Например, для кукурузы эффективность GS была доказана для двух родительских популяций (Massman et al., 2013; Beyene et al., 2015; Vivek et al., 2017), а также для многоплодных популяций. родительские популяции (Zhang et al., 2017). Его использование также было задокументировано у видов с долгим периодом генерации, таких как деревья (Grattapaglia et al., 2018) и молочное скотоводство, где сокращение цикла размножения увеличило реакцию на селекцию по сравнению с системой тестирования потомства (García -Ruiz et al., 2016).
Геномная селекция была применена ко многим культурам, включая пшеницу, нут, маниоку и рис (Roorkiwal et al., 2016; Crossa et al., 2017; Wolfe et al., 2017; Huang et al., 2019), а также количество программ, которые переходят от «обычных» к GS, растет. Результаты по пшенице показывают, что геномные прогнозы, использованные на ранних этапах селекционного цикла, привели к значительному повышению производительности в последующих поколениях (Bonnett et al., 2021 this issue).
Определение основных основных родителей для селекции с помощью геномной селекции
В геномной селекции оптимизация состава обучающей выборки является важной темой, потому что обучающая и тестовая выборки должны быть генетически связаны таким образом, чтобы генетическое разнообразие, присутствующее в тестовой выборке, могло быть охвачено и отражено разнообразием в обучающей выборке. .Программы разведения должны начать формирование первоначальных базовых основных родителей (обучающих популяций), которые представляют генетическое разнообразие, обнаруженное в текущем потомстве, и в максимально возможной степени соответствуют тестируемой (ым) популяции (ям) (Hickey et al., 2012). Эти основополагающие родители должны быть экстенсивно фенотипированы в различных целевых популяциях окружающей среды и генотипированы с помощью систем маркеров высокой плотности. Эти обучающие наборы основных родителей смогут создать модель с высокой точностью для текущих тщательно отобранных потомков (Zhang et al., 2017).
Почему подробные феномены и использование анализа нескольких признаков для улучшения селекции?
Наиболее важным ограничением для определения точных фенотипов было время и затраты, необходимые для измерения признаков в полевых условиях. Полевые феномены нацелены на изучение всех фенотипов растений в различных условиях окружающей среды. Современные методы феномены позволяют использовать гиперспектральные / мультиспектральные камеры для получения сотен точек данных по отражательной способности в дискретных узких полосах во многих средах и на многих этапах развития сельскохозяйственных культур.Теперь технологию фенотипирования можно использовать для быстрого и точного получения данных об агрономических признаках на основе достижений в технологиях фенотипирования растений (Atkinson et al., 2018). Таким образом, основная цель фенотипа с высокой пропускной способностью (HTP) — снизить стоимость данных для каждого участка и повысить точность прогнозов на ранних этапах вегетации сельскохозяйственных культур с использованием высоконаследственных вторичных фенотипов, тесно связанных с селекцией. фенотипы. Стоимость обработки данных HTP можно минимизировать, используя программное обеспечение с открытым исходным кодом, такое как FieldImageR (Matias et al., 2020).
Имеются свидетельства того, что анализ нескольких признаков повышает точность прогнозов, если в процессе моделирования учитываются генетические и остаточные корреляции. Новые геномные модели, которые принимают во внимание множественные признаки и множественные среды, наряду с взаимодействиями признак × среда, признак × генотип и признак × генотип × среда, предлагают огромный потенциал для использования корреляций между различными переменными и для дифференциации между ними. эффекты.Интеграция текущих моделей с несколькими признаками GBLUP с моделями, которые учитывают информацию об окружающей среде с условиями двух- и трехстороннего взаимодействия, обеспечивает мощную, унифицированную модель прогнозирования всего генома.
Байесовская модель с несколькими признаками и множеством сред (BMTME) (Montesinos-López et al., 2016, 2019a) позволяет создавать общие ковариационные матрицы для признаков и сред, которые лучше отражают корреляции между признаками и средами. Эта унифицированная модель может быть реализована для выбора генотипов с признаками, измеренными в одной среде, и для прогнозирования в других, непроверенных средах.Его также можно применять для прогнозирования характеристик, которые сложно или дорого измерить во всех средах.
Крайне важно получить большие и взаимодействующие наборы данных по феноменам на основе полевого фенотипирования. Это следует использовать для характеристики основных основных родителей в различных средах и включения их в визуальные данные, собранные в различных средах. Эти данные, наряду с информацией о происхождении и геноме, могут быть использованы для соответствия байесовским линейным смешанным моделям для вычисления BLUP генетических значений материала в обучающей выборке.Программы разведения должны собирать данные по множеству признаков в мультисреде, используемой для основных основных родителей, и использовать возможные корреляции между признаками, которые в конечном итоге повысят точность прогнозов. Геномика и феномена основных основных родителей с множеством признаков важны для использования вместе с данными по окружающей среде.
Почему Enviromics должна улучшать испытания в разных средах для селекции растений с помощью геномики?
Фенотипические вариации, наблюдаемые в различных средах, являются продуктом генетических и экологических вариаций.Таким образом, окружающая среда выступает в качестве центрального узкого места для применения современных инструментов прогнозирования с помощью геномики, особенно для использования в нескольких средах. Новые подходы объединяют данные полевых испытаний с последовательностями ДНК с использованием различных источников окружающей среды, таких как линейные и нелинейные модели реакции-нормы (например, Jarquín et al., 2014; Morais-Júnior et al., 2018; Millet et al., 2019 ; Monteverde et al., 2019; Costa-Neto et al., 2020a), результаты модели роста сельскохозяйственных культур (CGM) (Heslot et al., 2014; Rincent et al., 2017, 2019), CGM интегрирована с GS (Cooper et al., 2016; Messina et al., 2018; Robert et al., 2020) и историческими метеорологическими записями для прогнозирования сортов в ближайшие годы (de los Campos et al. , 2020).
Например, стратегия, предложенная de los Campos et al. (2020) оценивает закономерности генома × среда (G × E), извлеченные из полевых испытаний, и прогнозирует ожидаемую производительность сорта в окружающей среде, но также оценивает ожидаемое распределение производительности сорта по другим возможным погодным условиям, учитывая при этом неопределенность модели. параметры.Это новый метод анализа испытаний в различных средах, который может ускорить оценку адаптивности и стабильности урожая зерна.
Еще одним недавним примером является подход, который может повысить разрешение в прогнозировании нескольких сред для обеспечения стабильности за счет использования преимуществ крупномасштабной среды с различными методами ядра (Costa-Neto et al., 2020a). Связь полевых испытаний с окружающей средой может быть сформирована с помощью линейных ковариаций (как было предложено Jarquín et al., 2014) и нелинейных методов (гауссово ядро, глубокое обучение и глубокое ядро) (Cuevas et al., 2016, 2017, 2018, 2019; Монтесинос-Лопес и др., 2018a, b, 2019b, c). Использование нелинейных ядер привело к повышению точности при прогнозировании новых генотипов при известных условиях, но в основном при прогнозировании новых условий окружающей среды (непроверенные среды). Этот подход был расширен, чтобы учесть несколько структур окружающей среды на разных стадиях развития сельскохозяйственных культур (Коста-Нето и др., 2020b). Для последнего авторы наблюдали повышенную способность объяснять G × E с точки зрения генотип-специфичных норм реакции для ключевых факторов окружающей среды или ключевых стадий развития.Эта повышенная способность объяснять G × E была важна для достижения более высокой точности по сравнению с моделями без экологической информации.
В недавней исследовательской статье Rogers et al. (2021) подчеркнули важность включения данных об окружающей среде с высокой пропускной способностью в модели геномного прогнозирования, чтобы выполнять прогнозы в новых средах, характеризующихся такими же характеристиками окружающей среды. Автор пришел к выводу, что, помимо других факторов, взаимодействия G × E и ковариаты окружающей среды должны быть включены в модели прогнозирования для повышения точности прогнозов.
Взаимосвязь в современном растениеводстве
Прогресс в модернизации статистических и количественных генетических моделей для анализа селекции растений в испытаниях с несколькими средами стал более очевидным по мере увеличения доступности информации о геномике, феномене и окружающей среде (см., Среди прочего, Vargas et al., 1998; Crossa et al., 2010; Burgueño et al., 2012; Heslot et al., 2014; Jarquín et al., 2014; Montesinos-López et al., 2017; Millet et al., 2019; Costa-Neto et al. al., 2020a; де лос Кампос и др., 2020; Роберт и др., 2020). Таким образом, мы видим, что все элементы, описанные выше, предлагают явный потенциал для ускорения генетических достижений в селекции растений. Однако для достижения больших возможностей требуется эффективная интеграция на основе данных, особенно с точки зрения повышения точности прогнозирования. Некоторые из основных связей между геномикой, феноменом и окружающей средой описаны ниже, а их потенциальные воздействия кратко изложены на Рисунке 1.
Рисунок 1 .Современный селекционный треугольник включает в себя геномику, феноменику и экологию. Связи между каждым из этих элементов могут быть полезны для ускорения генетических достижений.
Связывание геномики и феномики
Связывание массивных наборов данных геномики и феномены сопряжено со сложностями, требующими статистических моделей для работы с очень большим количеством коррелированных предикторов. Монтесинос-Лопес и др. (2017) предложили связать геномику и феномику с байесовскими моделями функциональной регрессии, которые учитывают все доступные полосы отражения (250 полос или длины волны), геномную или родословную информацию, основные эффекты линий и окружающей среды, а также эффекты взаимодействия.Они заметили, что модели с условиями взаимодействия длина волны × среда были наиболее точными для прогнозирования производительности в трех различных средах и в различные моменты времени развития сельскохозяйственных культур. Модели функциональной регрессии скупы и эффективны с точки зрения вычислений, поскольку математические базисные функции позволяют выбрать только 21 бета-коэффициент (а не использовать все 250). Недавно Lopez-Cruz et al. (2020) предложили метод прогнозирования генетических достоинств сортов на основе многомерных данных ПВТ путем интеграции многомерных регрессий в стандартную методологию селекционного индекса.
Связывание данных с несколькими признаками и данными из разных сред
Данные с несколькими признаками и несколькими средами (MTME) используют преимущества крупномасштабных корреляций между различными признаками, оцениваемыми в различных средах, для обучения точным моделям GS. Из-за этого использование GS в данных MTME является многообещающим подходом для уменьшения усилий по полевому фенотипированию. Например, Ибба и др. (2020) оценили эффективность прогноза по 13 качественным признакам пшеницы с использованием двух моделей с несколькими признаками и пяти наборов данных, основанных на полевых оценках в течение двух последовательных лет.На второй год (тестирование) линии были спрогнозированы с использованием информации о качестве, полученной в первый год (обучение). Для большинства качественных признаков они обнаружили умеренную или высокую точность прогнозов, что позволяет предположить, что использование GS на более ранних стадиях может быть рекомендовано. В целом результаты показывают, что байесовская модель MTME помогает уловить корреляцию между признаками и корреляцию между годами, тем самым повышая точность прогнозов. Наконец, мы видим перспективы моделирования норм реакции на основе MTME с участием других омиков, таких как феномен и окружающая среда.Последние могут улучшить анализ MTME с точки зрения создания большего количества биологических моделей роста, развития и компонентов урожая сельскохозяйственных культур (например, Robert et al., 2020).
Interplay Genomics and Enviromics
С 1960-х годов несколько исследователей предложили использовать информацию об окружающей среде для объяснения различий между сортами из-за взаимодействий G × E (например, Perkins and Jinks, 1968; Freeman and Perkins, 1971; Wood, 1976; Vargas et al., 1998; Crossa et al., 1999). Использование геномики с окружающей средой является основой для прогнозирования сортов в различных условиях выращивания (например,г., Яркин и др., 2014; Мессина и др., 2018; Millet et al., 2019), который полезен для прогнозирования глобального потепления.
Тем не менее, усилия по внедрению экологических ковариат в модели геномной селекции обычно сосредотачиваются на нескольких экологических ковариатах, таких как температура, осадки и солнечное излучение, определенных на конкретных стадиях развития культуры. С использованием крупномасштабных данных экологического типирования можно разработать глобальную сеть экологического типирования полевых испытаний для обучения моделей GS и выполнения «экологической сборки» для прогнозирования большего числа условий выращивания на основе исторических данных о климате и почве (R пакет EnvRtype, Коста-Нето и др., 2020b). Кроме того, ведутся исследования по изучению модели Enviromic + Genomic prediction (E-GP) для связи вариаций генотип-фенотип, а также для объяснения фенотипических вариаций в разных средах. В качестве инструмента прогнозирования разведения E-GP может способствовать изучению структур G × E, в котором в качестве исследовательского инструмента E-GP может способствовать оптимизации экспериментальных сетей полевых испытаний и приводить к более эффективным обучающим сетам для GS (например, Rincent et al., 2017). Кроме того, на ранних этапах отбора геномика и окружающая среда могут использоваться для разработки оптимизированных испытаний фенотипирования и прогнозирования селекционной ценности кандидата для отбора (Morais-Júnior et al., 2018) или одиночного кросс-гибридного предсказания (Costa-Neto et al., 2020a).
Посредством сборки в окружающей среде можно установить взаимосвязь между полевыми испытаниями и, таким образом, использовать только наиболее репрезентативный набор экспериментов для обучения моделей GS. Другой перспективой E-GP является использование крупномасштабных данных об окружающей среде в обучающих моделях, включающих нормы реакции, специфичные для генотипа (например, Ly et al., 2018; Millet et al., 2019), и фенотипические ландшафты, реализуемые геномикой с ростом урожая. модели (CGM) (e.г., Мессина и др., 2018; Bustos-Korts et al., 2019; Роберт и др., 2020). Возможное использование основанных на изображениях ответов, связанных с основными экологическими стрессами, такими как жара и стресс из-за засухи, также может способствовать внедрению геномных платформ для прогнозных целей и позволяет лучше представить взаимодействие растений и окружающей среды.
Перспективы на будущее
Для решения хорошо задокументированных проблем продовольственной безопасности и безопасности питания настоятельно необходимо использовать новые технологии для ускорения прогресса селекции растений.Эти методы могут быть включены в обычные программы фенотипической селекции или помочь в изменении устоявшихся схем фенотипической селекции, чтобы обеспечить постепенный переход к перспективе, в большей степени ориентированной на данные. Преимущества феномена и окружающей среды вместе в эталонных геномных конвейерах предлагают потенциал для обеспечения большего повышения точности и эффективности племенных конвейеров, когда мы отбираем лучше адаптированные генотипы рентабельным способом, а также в более короткие сроки. Геномический, феноменальный, многосторонний и экологический анализ взаимосвязаны, и их использование может быть оптимизировано в зависимости от ресурсов и структуры программы.Вместе они предлагают путь для обычного фенотипического разведения, чтобы представить разнообразный набор возможностей для ускорения генетических достижений.
Авторские взносы
JC подготовил первые проекты заключения, RF-N, OM-L, GC-N, SD и AM-L прочитали и исправили первую версию. AB подготовил несколько обзоров документов и работал с JC над окончательной версией. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
взносов за открытый доступ поступило от Фонда Билла и Мелинды Гейтс.Мы признательны за финансовую поддержку, предоставленную Фондом Билла и Мелинды Гейтс [INV-003439 BMGF / FCDO Accelerating Genetic Gains in кукуруза и пшеница для повышения уровня жизни (AG2MW)], а также проектами USAID [поправка. № 9 MTO 069033, USAID-CIMMYT Wheat / AGGMW, AGG-Maize Supplementary Project, AGG (Стрессоустойчивая кукуруза для Африки)], на основе которого были получены данные CIMMYT, проанализированные в этом исследовании. Мы также благодарны за финансовую поддержку, оказываемую фондами сбора за исследования сельскохозяйственных продуктов (F.F.J.) и Фонд исследований сельскохозяйственных соглашений (J.A.) в Норвегии через грант NFR 267806, а также CIMMYT CRP (кукуруза и пшеница).
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Список литературы
Аткинсон, Дж. А., Джексон, Р. Дж., Бентли, А. Р., Обер, Э., и Уэллс, Д. М. (2018). Полевое фенотипирование будущего. Annu. Завод Rev. Интернет. DOI: 10.1002 / 9781119312994.apr0651
CrossRef Полный текст | Google Scholar
Бейене, Ю., Семагн, К., Муго, С., Тарекегне, А., Бабу, Р., Мейзель, Б. и др. (2015). Генетический выигрыш в урожайности зерна за счет геномной селекции в восьми двух родительских популяциях кукурузы в условиях засухи. Crop Sci. 55, 154–163. DOI: 10.2135 / cropci2014.07.0460
CrossRef Полный текст | Google Scholar
Burgueño, J., de los Campos, G., Вейгель, К., Кросса, Дж. (2012). Геномное предсказание племенной ценности при моделировании взаимодействия генотипа X с окружающей средой с использованием родословных и плотных молекулярных маркеров. Crop Sci. 52, 707–719. DOI: 10.2135 / cropci2011.06.0299
CrossRef Полный текст | Google Scholar
Bustos-Korts, D., Malosetti, M., Chenu, K., Chapman, S., Boer, M. P., Zheng, B., et al. (2019). От QTL к адаптационным ландшафтам: использование моделей от генотипа к фенотипу для характеристики G × E с течением времени. Фронт. Plant Sci. 10, 1–23. DOI: 10.3389 / fpls.2019.01540
CrossRef Полный текст | Google Scholar
Коллард, Б.С.Й., Грегорио, Г.Б., Томсон, М.Дж., Ислам, М.Р., Вергара, Г.В., Лаборте, А.Г. и др. (2019). Преобразование селекции риса: перепроектирование орошаемого селекционного трубопровода в Международном научно-исследовательском институте риса (IRRI). Crop Breed Genet. Геном 1: e1
. DOI: 10.20900 / cbgg201CrossRef Полный текст | Google Scholar
Купер, М., Technow, F., Messina, C., Gho, C., and Radu Totir, L. (2016). Использование моделей роста сельскохозяйственных культур с прогнозированием всего генома: применение в мультиэкологических испытаниях кукурузы. Crop Sci. 56, 2141–2156. DOI: 10.2135 / cropci2015.08.0512
CrossRef Полный текст | Google Scholar
Коста-Нето, Г., Фриче-Нето, Р., и Кросса, Дж. (2020a). Нелинейные ядра, данные о доминировании и типировании окружающей среды повышают точность прогнозов на основе генома в испытаниях с несколькими средами. Наследственность (Edinb). 126, 92–106. DOI: 10.1038 / s41437-020-00353-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Коста-Нето, Г., Галли, Г., Фанелли, Х., Кросса, Дж., И Фриче-Нето, Р. (2020b). EnvRtype: программное обеспечение для взаимодействия экологической и количественной геномики в сельском хозяйстве. bioRxiv [препринт]. DOI: 10.1101 / 2020.10.14.339705
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Crossa, J., Pérez-Rodríguez, P., Cuevas, J., Montesinos-López, O.A., Jarquín, D., de Los Campos, G., et al. (2017). Геномная селекция в селекции растений: методы, модели, перспективы. Trends Plant Sci . 22, 961–975. DOI: 10.1016 / j.tplants.2017.08.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кросса, Дж., Варгас, М., и Джоши, А. К. (2010). Линейные, билинейные и линейно-билинейные фиксированные и смешанные модели для анализа взаимодействия генотипа x с окружающей средой в селекции растений и агрономии. Банка. J. Plant Sci. 90, 561–574. DOI: 10.4141 / CJPS10003
CrossRef Полный текст | Google Scholar
Crossa, J., Vargas, M., Van Eeuwijk, F.A., Jiang, C., Edmeades, G.O., and Hoisington, D. (1999). Интерпретация взаимодействия генотипа x окружающей среды у тропической кукурузы с использованием связанных молекулярных маркеров и переменных окружающей среды. Теор. Прил. Genet. 99, 611–625. DOI: 10.1007 / s001220051276
CrossRef Полный текст | Google Scholar
Куэвас, Дж., Кросса, Дж., Монтесинос-Лопес, О.А., Бургеньо, Дж., Перес-Родригес, П., и де лос Кампос, Г. (2017). Байесовское геномное предсказание с помощью моделей ядра среды генотип x. G3: Гены | Геномы | Генетика 7, 41–53. DOI: 10.1534 / g3.116.035584
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Куэвас, Дж., Кросса, Дж., Соберанис, В., Перес-Элизальде, С., Перес-Родригес, П., де лос Кампос, Г. и др. (2016). Геномное предсказание ядерных регрессионных моделей взаимодействия генотипа x с окружающей средой. Геном растений 9, 1–20. DOI: 10.3835 / plantgenome2016.03.0024
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Куэвас, Дж., Гранато, И., Фриче-Нето, Р., Монтесинос-Лопес, О. А., Бургеньо, Дж., Бандейра и Соуза, М. и др. (2018). Модели ядра прогнозирования с поддержкой генома со случайными перехватами для испытаний в нескольких средах. G3: Гены | Геномы | Генетика 8, 1347–1365. DOI: 10.1534 / g3.117.300454
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Куэвас, Дж., Монтесинос-Лопес, О.А., Хулиана, П., Гусман, К., Перес-Родригес, П., Гонсалес-Бусио, Дж. И др. (2019). Глубокое ядро для геномного прогнозирования и прогнозирования в ближней инфракрасной области в испытаниях разведения в нескольких средах. G3: Гены | Геномы | Генетика 9, 2913–2924. DOI: 10.1534 / g3.119.400493
CrossRef Полный текст | Google Scholar
де лос Кампос, Г., Перес-Родригес, П., Богард, М., Гуаш, Д., и Кросса, Дж. (2020). Платформа моделирования на основе данных для прогнозирования характеристик сортов в неопределенных погодных условиях. Нат. Commun. 11: 4876. DOI: 10.1038 / s41467-020-18480-y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фримен, Г. Х., и Перкинс, Дж. М. (1971). Экологические и генотип-экологические компоненты изменчивости: Viii Отношения между генотипами, выращенными в различных средах, и меры этих сред. Наследственность (Edinb). 27, 15–23. DOI: 10.1038 / HDY.1971.67
CrossRef Полный текст | Google Scholar
Гарсия-Руис, А., Коул, Дж. Б., ВанРаден, П. М., Вигганс, Г. Р., Руис-лопес, Ф. Дж., И Ван Тасселл, К. П. (2016). Изменения дифференциалов генетического отбора и интервалов между поколениями у молочного скота американской голштинской породы в результате геномного отбора. Proc. Natl. Акад. Sci. США 113, E3395 – E4004. DOI: 10.1073 / pnas.151
13
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Граттапалья Д., Сильва-Джуниор О. Б., Ресенде Р. Т., Каппа Э. П., Мюллер Б. С. Ф. и Тан Б.(2018) Количественная генетика и геномика объединяются для ускорения селекции лесных деревьев. Фронт. Завод Sci . 871, 1–10. DOI: 10.3389 / fpls.2018.01693.
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Heslot, N., Akdemir, D., Sorrels, M. E., and Jannink, J.-L. (2014). Интеграция ковариат окружающей среды и моделирования культур в структуру геномной селекции для прогнозирования генотипа по взаимодействиям с окружающей средой. Теор. Прил. Genet. 127, 463–480. DOI: 10.1007 / s00122-013-2231-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хики, Дж. М., Кросса, Дж., Бабу, Р., и де лос Кампос, Г. (2012). Факторы, влияющие на точность вменения генотипа в популяциях из нескольких селекционных программ кукурузы. Crop Sci. 52, 654–663. DOI: 10.2135 / cropci2011.07.0358
CrossRef Полный текст | Google Scholar
Хуанг, М., Балимпония, Э. Г., Мгонья, Э. М., Макхейл, Л. К., Лузи-Кихупи, А., Го-Лян Ван, Г.-L., Et al. (2019). Использование геномной селекции в селекции риса ( Oryza sativa L.) на устойчивость к взрывам риса ( Magnaporthe oryzae ). Мол. Порода . 39: 114. DOI: 10.1007 / s11032-019-1023-2
CrossRef Полный текст | Google Scholar
Ибба, М. И., Кросса, Дж., Монтесинос-Лопес, О. А., Монтесинос-Лопес, А., Джулиана, П., Гусман, К. и др. (2020). Прогнозирование нескольких показателей качества пшеницы на основе генома на несколько лет. Геном растений 1:14.DOI: 10.1002 / tpg2.20034
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Jarquín, D., Crossa, J., Lacaze, X., Du Cheyron, P., Daucour, J., Lorgeou, J., et al. (2014). Модель нормы реакции для геномного отбора с использованием многомерных геномных и экологических данных. Теор. Прил. Genet. 127, 595–607. DOI: 10.1007 / s00122-013-2243-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лопес-Крус, М., Олсон, Э., Ровере, Г., Crossa, J., Dreisigacker, S., Mondal, S, et al. (2020). Регуляризованные индексы выбора для прогнозирования племенной ценности с использованием данных гиперспектрального изображения. Sci. Реп . 10. DOI: 10.1038 / s41598-020-65011-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ly, D., Huet, S., Gauffreteau, A., Rincent, R., Touzy, G., Mini, A., et al. (2018). Полногеномное прогнозирование норм реакции на стресс окружающей среды у мягкой пшеницы ( Triticum aestivum L.) методом случайной геномной регрессии. F. Crop. Res. 216, 32–41. DOI: 10.1016 / j.fcr.2017.08.020
CrossRef Полный текст | Google Scholar
Massman, J. M., Jung, H.-J. Г., Бернардо Р. (2013). Полногеномный отбор в сравнении с периодическим отбором с использованием маркеров для улучшения урожайности зерна и улучшения качества соломы на целлюлозный этанол кукурузы. Crop Sci. 53, 58–66. DOI: 10.2135 / cropci2012.02.0112
CrossRef Полный текст | Google Scholar
Матиас, Ф. И., Караза-Хартер, М.В., Эндельман Дж. Б. (2020). FIELDimageR: пакет R для анализа ортофотопланов, полученных в ходе сельскохозяйственных полевых испытаний. Plant Phenome J. 3, 1–6. DOI: 10.1002 / ppj2.20005
CrossRef Полный текст | Google Scholar
Мессина, К. Д., Технов, Ф., Танг, Т., Тотир, Р., Гхо, К., и Купер, М. (2018). Использование биологических представлений и изменений окружающей среды для улучшения фенотипического прогнозирования: интеграция моделей роста сельскохозяйственных культур (CGM) с прогнозированием всего генома (WGP). Eur.J. Agron. 100, 151–162. DOI: 10.1016 / j.eja.2018.01.007
CrossRef Полный текст | Google Scholar
Миллет, Э. Дж., Круиджер, В., Купель-Ледру, А., Альварес Прадо, С., Кабрера-Боске, Л., Лакуб, С., и др. (2019). Геномное прогнозирование урожайности кукурузы в условиях окружающей среды Европы. Нат. Genet. 51, 952–956. DOI: 10.1038 / s41588-019-0414-у
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Монтесинос-Лопес, А., Монтесинос-Лопес, О.A., Cuevas, J., Mata-López, W. A., Burgueño, J., Mondal, S., et al. (2017). Геномные байесовские функциональные регрессионные модели с взаимодействиями для прогнозирования урожайности зерна пшеницы с использованием данных гиперспектрального изображения. Растительные методы 13:62. DOI: 10.1186 / s13007-017-0212-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Монтесинос-Лопес, А., Монтесинос-Лопес, О. А., Джанола, Д., Кросса, Дж., И Эрнандес-Суарес, К. М. (2018a). Геномное прогнозирование свойств растений в разных средах с использованием глубоких учеников с плотной архитектурой. G3: Гены | Геномы | Генетика 8, 3813–3828. DOI: 10.1534 / g3.118.200740
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Монтесинос-Лопес, М. А., Монтесинос-Лопес, А., Луна-Васкес, Х. Ф., Толедо, Ф. Х., Паулино Перес-Родригес, П., Лиллемо, М., и др. (2019a). Пакет R для байесовского анализа данных с несколькими средами и несколькими признаками для прогнозирования на основе генома. G3. Гены | Геномы | Генетика 9, 1355–1369. DOI: 10,1534 / g3.119.400126
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Монтесинос-Лопес, О.А., Мартин-Вальехо, Дж., Кросса, Дж., Джанола, Д., Эрнандес-Суарес, К., Монтесинос-Лопес, А. и др. (2019b). Сравнительный анализ между глубоким обучением, опорной векторной машиной и байесовским порогом наилучшего линейного несмещенного прогноза для прогнозирования порядковых признаков в селекции растений. G3: Гены | Геномы | Генетика 9, 601–618. DOI: 10.1534 / g3.118.200998
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Монтесинос-Лопес, О.А., Мартин-Вальехо, Дж., Кросса, Дж., Джанола, Д., Эрнандес-Суарес, К. М., Монтесинос-Лопес, А. и др. (2019c). Новая модель прогнозирования на основе генома с глубоким обучением для множественных признаков с бинарными, порядковыми и непрерывными фенотипами. G3: Гены | Геномы | 9, 1545–1556. DOI: 10.1534 / g3.119.300585
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Монтесинос-Лопес, О. А., Монтесинос-Лопес, А., Кросса, Дж., Толедо, Ф. Х., Перес-Эрнандес, О., Эскридж, К. М. и др.(2016). Геномная байесовская модель с несколькими признаками и средой. G3 Гены | Геномы | Генетика 6, 2725–2744. DOI: 10.1534 / g3.116.032359
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Монтесинос-Лопес, О. А., Монтесинос-Лопес, А., Джанола, Д., Кросса, Дж., И Эрнандес-Суарес, К. М. (2018b). Моделирование глубокого обучения с несколькими признаками и средами для прогнозирования растений на основе генома. G3: Гены | Геномы | Генетика 8, 3829–3840. DOI: 10.1534 / g3.118.200728
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Монтеверде, Э., Гутьеррес, Л., Бланко, П., Перес де Вида, Ф., Росас, Дж. Э, Боннекарре, В., и др. (2019). Интеграция молекулярных маркеров и ковариат окружающей среды для интерпретации генотипа по взаимодействию с окружающей средой у риса ( Oryza sativa L.), выращенного в субтропических регионах. G3 Гены | Геномы | Генетика 9, 1519–1531. DOI: 10.1534 / g3.119.400064
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Morais-Júnior, O.П., Дуарте, Дж. Б., Бресегелло, Ф., Коэльо, А. С. Г. и Магальяйнс, А. М. младший (2018). Одноступенчатые модели нормы реакции для геномного прогнозирования в испытаниях повторяющегося отбора в различных средах. Crop Sci. 58, 592–607. DOI: 10.2135 / cropci2017.06.0366
CrossRef Полный текст | Google Scholar
Перкинс, Дж. М., и Джинкс, Дж. Л. (1968). Экологический и генотип-средовой компоненты изменчивости. 3. Множественные линии и кресты. Наследственность (Edinb). 23, 339–356.DOI: 10.1038 / HDY.1968.48
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ресенде, Р. Т., Пьефо, Х. П., Роза, Г. Дж. М., Сильва-Жуниор, О. Б., Сильва, Ф. Ф. Э., де Ресенде, М. Д. В. и др. (2020). Энвиромика в селекции: приложения и перспективы селекции с помощью энвиротипов. Теор. Прил. Genet. 134, 95–112. DOI: 10.1007 / s00122-020-03684-z
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ринсент, Р., Кун, Э., Монада, Х., Ури, Ф. X., Руссе, М., Аллард, В. и др. (2017). Оптимизация испытаний в нескольких средах для геномной селекции на основе моделей сельскохозяйственных культур. Теор. Прил. Genet. 130, 1735–1752. DOI: 10.1007 / s00122-017-2922-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Rincent, R., Malosetti, M., Ababaei, B., Touzy, G., Mini, A., Bogard, M., et al. (2019). Использование ковариат стресса модели роста сельскохозяйственных культур и разложения AMMI для лучшего прогнозирования взаимодействия генотипа с окружающей средой. Теор. Прил. Genet. 132, 3399–3411. DOI: 10.1007 / s00122-019-03432-y
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Роберт П., Ле Гуи Дж. И Ренсан Р. (2020). Сочетание моделирования роста сельскохозяйственных культур с прогнозированием на основе признаков улучшило прогнозирование генотипа по взаимодействиям с окружающей средой. Фронт. Plant Sci. 11, 1–11. DOI: 10.3389 / fpls.2020.00827
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Роджерс, А.R., Dunne, J.C., Romay, M.C., Bohn, M., Buckler, E.S., Ciampitti, I.A., et al. (2021 г.). Важность доминирования и взаимодействия между генотипом и окружающей средой в изменении урожайности зерна в крупномасштабном государственном кооперативном кукурузе. G3: Гены | Геномы | Генетика 1: jkaa050.doi: 10.1093 / g3journal / jkaa050
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Руркивал М., Ратхор А., Дас Р. Р., Сингх М. К., Джайн А., Сринивасан С. и др. (2016). Модели прогнозирования на основе генома признаков, связанных с урожайностью, у нута. Фронт. Завод Sci . 7: 1666. DOI: 10.3389 / fpls.2016.01666
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Варгас М., Кросса Дж., Сэйр К., Рейнольдс М., Рамирес М. Э. и Талбот М. (1998). Интерпретация взаимодействия генотипа X с окружающей средой с использованием частичной регрессии наименьших квадратов. Crop Sci. 38, 679–689. DOI: 10.2135 / cropci1998.0011183X003800030010x
CrossRef Полный текст | Google Scholar
Вивек, Б.С., Кришна, Г.K., Vengadessan, V., Babu, R., Zaidi, P.H., Kha, L.Q., et al. (2017). Использование геномных оценочных значений селекционной ценности приводит к быстрому генетическому улучшению засухоустойчивости кукурузы ( Zea mays L.). Геном растений 10, 1–8, DOI: 10.3835 / plantgenome2016.07.0070
CrossRef Полный текст | Google Scholar
Вулф, М. Д., Дель Карпио, Д. П., Алаби, О., Эзенвака, Л. К., Икеогу, Ю. Н., Кайондо, И. С. и др. (2017). Перспективы геномной селекции в селекции маниоки. Геном растений 10:15. DOI: 10.3835 / plantgenome2017.03.0015
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжан, X., Перес-Родригес, П., Бургеньо, Дж., Олсен М., Баклер, Э., Атлин, Г., и Прасанна, Б. М. (2017) Быстрая циклическая геномная селекция в многоплодной популяции тропической кукурузы. G3: Ген / Геном / Генет 7, 2315–2326. DOI: 10.1534 / g3.117.043141.
CrossRef Полный текст | Google Scholar
(PDF) Краткое описание методов селекции растений для студентов
Международный журнал сельского хозяйства и агробизнеса
ISSN: 2391-3991, том 3, выпуск 2, стр. 156 — 203
Замбрут.com. Дата публикации: 13 мая 2019 г.
Degefa, I. 2019. Методы селекции: кратко о …………
6.2. Цветение и способы опыления
В процессе цветения первое раскрытие цветка известно как цветение. Его детали
различаются от растения к другому, и на него сильно влияют факторы окружающей среды, такие как влажность и температура
. Знание цветков желательно для успешных скрещиваний и во многом определяет способ опыления.Опыление означает перенос пыльцевых зерен с пыльников на рыльца.
Когда пыльца с пыльника попадает на рыльце того же растения, происходит самоопыление (автогамия), но
, когда пыльца передается на рыльце другого растения, происходит перекрестное опыление (аллогамия).
6.2.1. Самоопыление (инбридинг)
Многие виды культурных растений размножаются путем самоопыления. Считается, что
этих видов произошли от перекрестно опыляемых предков; у них есть цветки-гермафродиты, и многие из них перекрестно опыляются
до 5%.Обычно самоопыляемые культуры включают: пшеницу, рис, ячмень, овес, теф, горох, арахис,
нут, вигну, сою, чечевицу, льняное семя, кунжут, помидоры, картофель, салат, персик и т. Д. Генетические последствия
при самоопылении наблюдается повышенная гомозиготность. Существуют различные механизмы, которые способствуют самоопылению
, как показано ниже:
Клистогамия: цветок не раскрывается, и это обеспечивает полное самоопыление, поскольку чужеродная пыльца
не может достичь стигмы закрытого цветка (например,г., пшеница, ячмень, овес).
Хасмогай: цветки раскрываются, но только после опыления (например, пшеница, ячмень, рис, овес).
Пыльники плотно окружают рыльце, и это положение обеспечивает самоопыление (например, помидор). Цветочные органы
(два чашелистика) скрывают или окружают тычинки и рыльца (например, некоторые бобовые, такие как горох, соя
и нут). Клеймо становится восприимчивым и удлиняется по тычинному столбу
, обеспечивая самоопыление.
6.2.2. Перекрестное опыление (аутбридинг)
При перекрестном опылении им способствуют ветер (анемофилия), вода (гидрофилия) или насекомые
(энтомофилия). Такие перекрестно опыляемые культуры могут иметь 5-10% самоопылений. Обычно перекрестно опыляемые культуры
включают: кукурузу, рожь, клещевину, нугу, подсолнечник, люцерну, клевер, сахарный тростник, яблоко, авокадо, банан,
винограда, финиковую пальму, папайю, капусту, морковь, лук, перец, тыкву, сладкий картофель, чеснок, кориандр,
манго и др.Генетическим следствием перекрестного опыления является то, что оно сохраняет и способствует гетерозиготности
в популяции, а также демонстрирует легкую и тяжелую депрессию инбридинга и значительное количество гетерозисов на
. Открытоопыляемые сорта, гибриды и синтетические сорта являются целью селекционера
без снижения гетерозиготности до значительного уровня. Механизмы, которые способствуют перекрестному опылению:
:
Однополость (диклиния): тычиночный или пестичный цветок; он бывает двух типов.
а. Однообразие: тычиночные или пестичные цветки встречаются на одном и том же растении
b. Размножение: тычиночные и пестичные цветки присутствуют на разных растениях
2. Дихогамия: тычинки и пестики цветков-гермафродитов созревают в разное время
a. Протогиния: пестики созревают раньше тычинок (например, сорго)
b. Протандрия: тычинки созревают раньше, чем пестики (кукуруза, сахарная свекла)
3. Стигма покрыта восковой пленкой, которую пчелы часто разрушают (напр.g., люцерна)
4. Самонесовместимость: пыльца не может опылить или оплодотворить яйцо того же растения. Обычно
делятся на две группы:
a. Гетероморфные системы (различная морфология, например, штифт и барабан)
б. Гомоморфные системы (те же структуры, но контролируемые генетикой)
5. Мужское бесплодие: отсутствие функциональных или нефункциональных пыльцевых зерен в цветках гермафродитов.
Мужское бесплодие подразделяется на три группы: генетическая, цитоплазматическая и цитоплазматическая.
Один рецессивный ген регулирует генетическое мужское бесплодие, которое возникает спонтанно или может быть вызвано
искусственно.Линия с мужской стерильностью может поддерживаться путем скрещивания ее с гетерозиготными мужскими фертильными растениями
. Генетическая мужская стерильность используется при производстве гибридных семян. С другой стороны, цитоплазматическая мужская стерильность
определяется цитоплазмой. Поскольку цитоплазма зиготы происходит в основном из клетки яйца
, потомство таких растений с мужской стерильностью всегда будет иметь мужскую стерильность. Он известен в таких культурах, как
кукурузы, пшеницы, табака, хлопка, сорго и подсолнечника. Цитоплазматическая мужская стерильность
может быть передана и сохранена в данном сорте с использованием этого сорта в качестве опылителя в шести-семи последовательных поколениях
программы обратного скрещивания.Используется для получения гибридных семян декоративных растений
Вы здесь: Сельское хозяйство> Трансгенные растения> Защита растений> Методы селекции растений 2.1 Обычные методыСелекция растений определяется как выявление и выбор желаемых признаков. в растениях и объединение их в одно отдельное растение. С 1900 г. Законы генетики Менделя обеспечили научную основу для растений. разведение.Поскольку все признаки растения контролируются генами, расположенными на хромосом, обычная селекция растений может рассматриваться как манипуляции комбинацией хромосом. В общем, есть три основных способа манипулирования хромосомой растения комбинация. Во-первых, растения данной популяции, показывающие желаемое признаки могут быть выбраны и использованы для дальнейшего разведения и выращивания, процесс, называемый (чисто линейным) отбором. Во-вторых, найдены желаемые черты в разных линиях растений можно комбинировать вместе для получения растений которые проявляют обе эти черты одновременно, метод, называемый гибридизация.Гетерозис, явление повышенной силы, — это получены гибридизацией инбредных линий. В-третьих, полиплоидия (повышенная количество наборов хромосом) может способствовать улучшению урожая. Наконец, новая генетическая изменчивость может быть привнесена спонтанно. или искусственно вызванные мутации. [2]
Успех в использовании традиционных принципов селекции растений и сельскохозяйственных техники достигли своего пика, когда линии высокоурожайной пшеницы и риса возделывались в 1960-е гг. Удвоение и утроение производительности этих важных культур в Азии сигнализировали о сельскохозяйственной революции в развивающиеся страны. Этот прорыв в производстве продуктов питания был называется «Зеленая революция», чтобы описать социальную, экономическую и питательное влияние новых высокоурожайных сортов пшеницы и риса.Норман Борлоуг был награжден Благородной премией в 1970 году за вклад в селекцию новых высокоурожайных сортов зерновых культур. Тем не мение, эти штаммы сильно зависели от удобрений, орошения и агрохимикаты и требовали энергоемких инвестиций. Это привело к деградации и потере почв и других неблагоприятных экологических проблемы во всем мире. По этим причинам «Зеленая революция» хвалили и осуждали. [2] 2.2 Биотехнологические методыБиотехнология — это дисциплина, связанная с использованием живых организмов. организмы или их продукты. В этом широком смысле также традиционный сельское хозяйство можно рассматривать как форму биотехнологии. Европейский Федерация биотехнологий определяет биотехнологию как «интегрированный использование биохимии, микробиологии и инженерных наук в целях добиться технологического (промышленного) применения возможностей микроорганизмы, культивируемые тканевые клетки и их части ».В последнее время лет биотехнология быстро развивалась как практическое средство для ускорение успеха в селекции растений и экономическое улучшение важные культуры. Наиболее важные методы, используемые для достижения этих цели описаны ниже. Методы генной инженерии, которые являются частью биотехнологии, более подробно будут рассмотрены в следующая глава.
2.3 Генная инженерияГенная инженерия — это термин, используемый для направленного манипулирования гены, т. е. перенос генов между организмами или изменения в последовательность гена. С этой областью тесно связаны методы которые используют гены или определенные последовательности для идентификации признаков и другие аналитические цели. В селекции растений самое важное и уже широко распространенный метод такого рода — Restriction Fragment Полиморфизм длины (RFLP).Основы этой техники: описано ниже. Принципы генной инженерии растений будут описано в следующей главе. Для тех, кто не знаком с принципы генетики, глоссарий в конце этого отчета объясняет наиболее важные термины. 2.3.1 Полиморфизм длины рестрикционного фрагмента (RFLP)Методы традиционного разведения требуют очень много времени. Сделав скрещивания, также большое количество нежелательных генов вводится в геном растения.Нежелательные гены нужно «отсортировать» обратным переходом. Использование рестрикционного полиморфизма длины фрагмента значительно облегчает обычное разведение растений, потому что можно прогрессировать в программе разведения намного быстрее, с меньшими популяций и не полагаясь исключительно на тестирование желаемого фенотип. RFLP использует эндонуклеазы рестрикции. Это ферменты, которые распознавать и вырезать определенные нуклеотидные последовательности в ДНК.Например, последовательность GAATTC разрезается эндонуклеазой EcoR1. После лечения генома растения, рестриктирующего эндонуклеазы, ДНК растения нарезать кусочками разной длины, в зависимости от количества сайты узнавания на ДНК. Эти фрагменты можно разделить в соответствии с их размером с помощью гель-электрофореза и производятся видны в виде полос на геле при гибридизации фрагментов ДНК растений с радиоактивно меченными или флуоресцентными ДНК-зондами.Поскольку два генома не идентичны даже в пределах данного вида из-за мутаций, количество сайты рестрикции и, следовательно, длина и количество фрагментов ДНК различаются, что приводит к различному рисунку полос на электрофорезе гель. Эта изменчивость была названа длиной рестрикционного фрагмента. полиморфизм (ПДРФ). Чем ближе два организма связаны между собой, тем больше узор из полос перекрывается. Если сайт рестрикции находится рядом с или даже в пределах важного гена наличие определенной группы коррелирует с конкретным признаком растения, например.грамм. болезнь сопротивление. Глядя на рисунок полос, заводчики могут определить лица, унаследовавшие гены устойчивости, и устойчивые растения можно отбирать для дальнейшего разведения. Использование этой техники не только значительно ускорит прогресс в селекции растений, но и также облегчит идентификацию генов устойчивости, тем самым открытие новых возможностей в селекции растений (сравните также главу 3.1.3.1). [4] 2.3.2 Перенос геновВ традиционной селекции набор доступных генов и черты, которые они код для ограничен из-за сексуальной несовместимости с другими строками рассматриваемой культуре и их диким сородичам. Это ограничение можно преодолеть с помощью методов генной инженерии, которые в принципе позволяют привнести ценные черты, закодированные конкретными гены любого организма (других растений, бактерий, грибов, животных, вирусов) в геном любого растения.Первые эксперименты по переносу генов с заводами произошла в начале 1980-х. Обычно трансгены вставлен в ядерный геном растительной клетки. Недавно это было стало возможным вводить гены в геном хлоропластов и другие пластиды (маленькие органеллы растительных клеток, обладающие отдельный геном). Преимущества этого метода обсуждаются в глава 3.1.1.1. Трансгенные растения были получены с использованием Agrobacterium-опосредованного переноса ДНК. и прямой перенос ДНК, последний включает такие методы, как бомбардировка частицами, электропорация и полиэтиленгликоль проницаемость.Большинство растений трансформировано с использованием Трансформация, опосредованная Agrobacterium. [5] 2.3.2.1 Перенос генов, опосредованный AgrobacteriumМетод, опосредованный Agrobacterium, включает естественный перенос генов. система, являющаяся резидентом бактериальных патогенов растений этого рода Agrobacterium. В природе Agrobacterium tumefaciens и Agrobacterium rhizogenes — возбудители венечного галла и волосистой корневые заболевания соответственно.Полезность Agrobacterium как гена система передачи была впервые признана, когда было продемонстрировано, что эти болезни растений на самом деле возникли в результате передачи и интеграция генов бактерий в геном растения. Оба вида Agrobacterium несут большую плазмиду (маленькую кольцевую ДНК молекула) называется Ti в A. tumefaciens и Ri в A. rhizogenes. Сегмент этой плазмиды, обозначенной T- (для переноса) ДНК, передается посредством этот организм на отдельные растительные клетки, обычно внутри раненых ткань (см. рисунок 2.1). Сегмент Т-ДНК проникает в растительную клетку ядро и случайным образом интегрируется в геном, где стабильно включены и унаследованы, как и любой другой ген растения, в предсказуемой, доминирующая менделевская мода. Экспрессия естественных генов на Т-ДНК приводит к синтезу продуктов генов, которые направляют наблюдаемые морфологические изменения, т.е. е. опухоль или волосатый корень формирование. В генной инженерии: индуцирующие опухоль гены в Т-ДНК, которые Причина болезни растения удаляется и заменяется чужеродными генами.Эти гены затем стабильно интегрируются в геном растения. после заражения измененным штаммом Agrobacterium, как и природная Т-ДНК. Поскольку все гены, вызывающие опухоль, удалены, Перенос гена не вызывает никаких симптомов заболевания. Самое важное шаги в переносе гена, опосредованном Agrobacterium, показаны на рисунке. 2.1.Этот надежный метод переноса генов хорошо подходит для растений. которые восприимчивы к заражению Agrobacterium.К несчастью, многие виды, особенно экономически важные бобовые и однодольные, такие как злаки, не реагируют положительно на Опосредованная агробактериями трансформация. Для этих растений следующие должны применяться методы прямого захвата ДНК. [5] 2.3.2.2 Бомбардировка частицамиЭтот метод, также называемый биолистической трансформацией (от биологической баллистика) включает нанесение биологически активной ДНК на небольшие частицы вольфрама или золота (диаметром 1-5 мкм) и ускоряющие их в ткань растения с большой скоростью.Частицы проникают в растение клеточная стенка и поселяются внутри клетки, где ДНК высвобождается, что приводит к трансформации отдельной растительной клетки в эксплант. Этот метод обычно менее эффективен, чем Опосредованная агробактериями трансформация, но тем не менее особенно полезен для некоторых видов растений, особенно для зерновых посевы. Введение ДНК в организованные морфогенные ткани таких как семена, зародыши или меристемы, позволили успешно преобразование и регенерация риса, пшеницы, сои и кукурузы, тем самым демонстрируя огромный потенциал этого метода. [5] 2.3.2.3 Электропорация и прямое проникновение ДНК в протопластыЭлектропорация — это процесс, при котором очень короткие электрические импульсы используется для обратимой проницаемости липидных бислоев мембран растительных клеток. Электрический разряд обеспечивает диффузию макромолекул, таких как как ДНК через непроницаемую в противном случае плазматическую мембрану. Поскольку клеточная стенка растений не позволит эффективно распространять многие трансгены. конструкции, протопласты (клетки без клеточных стенок) должны быть подготовлены.Это требование представляет собой серьезное препятствие для многих приложений, так как протоколы, позволяющие полностью восстановить протопласты. для многих видов растений не существует. протопластами растений также может стимулироваться фосфатом или соосаждение кальция и полиэтиленгликоля (ПЭГ). Однако эти все методы страдают тем недостатком, что они используют протопласты в качестве реципиент-хозяин, который часто не может быть регенерирован в целые растения. [5] 2.3.3 Экспрессия трансгенаВ большинстве случаев введение гена в геном растения только оказывают влияние на растение, если трансген экспрессируется, т.е. е. транскрибируется в мРНК и транслируется в белок. Промоутер — это последовательность нуклеиновых кислот, в которой РНК-полимераза (сложный фермент синтезирует транскрипт мРНК) прикрепляется к матрице ДНК. В природа промотора определяет (вместе с другими регулирующими экспрессию элементов), при каких условиях и с какой интенсивностью ген будут расшифрованы.Промотор гена 35S мозаики цветной капусты вирус очень часто используется в генной инженерии растений. Этот промотор обеспечивает высокий уровень экспрессии экзогенных генов в большинстве типы клеток практически всех протестированных видов. Как это часто бывает выгодно экспрессировать трансген только в определенных тканях или количествах или в определенное время ряд других промоторов доступны, например промоторы, индуцирующие экспрессию генов после ранения или только во время созревания плодов. Используемые в настоящее время методы переноса генов приводят к случайной интеграции чужеродной ДНК по всему геному реципиентных клеток. Сайт вставка может иметь сильное влияние на уровни экспрессии экзогенного гена, что приводит к различным уровням экспрессии введенный ген, даже если использовалась такая же конструкция промотор / ген. Точный механизм этого явления до конца не изучен. [5] 2.3.4 Выбор и регенерация растенийВ эксперименте по трансформации доля трансформированных клеток составляет обычно мало по сравнению с количеством ячеек, которые остаются неизменными. Чтобы выбрать только те ячейки, которые действительно содержат новые гены, гены, кодирующие желаемый признак, сливаются с геном что позволяет производить отбор трансформированных клеток, так называемых маркерных генов. Экспрессия маркерного гена позволяет трансгенным клеткам расти в присутствии селективного агента, обычно антибиотика или гербицидом, а клетки без маркерного гена погибают.Один из многих обычно используемые маркерные гены — это бактериальный аминогликозид-3 ‘ ген фосфотрансферазы (APH (3 ‘) II), также называемый неомицином фосфотрансфераза 11 (NPTII). Этот ген кодирует фермент, который инактивирует антибиотики канамицин, неомицин и G418 за счет фосфорилирование. Помимо НПТИИ, существует ряд других антибиотиков. гены устойчивости использовались в качестве селективных маркеров, например гигромицин ген фосфотрансферазы, придающий устойчивость к гигромицину. Другая группа селективных маркеров — это гены толерантности к гербицидам. Гербицид толерантность была получена путем включения и выражения гена, который обезвреживает гербицид аналогичным образом как продукты гена устойчивости к антибиотикам или ген, который экспрессирует продукт, который действует как гербицид, но не влияет на него гербицидом. Толерантность к гербицидам может служить не только признаком полезен для селекции при разработке трансгенных растений, но также имеет некоторый коммерческий интерес.Трансгенная устойчивость к гербицидам поэтому растения являются одними из первых культур, подходящих к рынку вступление. Трансформация протопластов, клеток и тканей растений обычно только полезно, если их можно регенерировать в целые растения. Ставки регенерации сильно различаются не только у разных видов, но также между сортами одного вида. Как упоминалось в capter 2.2, во многих случаях регенерация целых растений из клеток невозможна. возможно или очень сложно.Помимо возможности ввести ген в геном вида растений, регенерация интактных, плодородных растения из трансформированных клеток или тканей — самый ограничивающий шаг в развитии трансгенных растений. [5] |
Технологии редактирования генома и их потенциальное применение в селекции садовых культур
Xu Y. Молекулярная селекция растений . CABI, 2010.
Google Scholar
Диркс Р., Ван Дун К., Де Сну С.Б., Ван Ден Берг М., Леливелт К.Л., Воерманс В. и др.Обратное разведение: новый подход к селекции, основанный на инженерном мейозе. Завод Биотехнология J 2009; 7 : 837–845.
CAS PubMed PubMed Central Google Scholar
Лю И, Ян Х, Саканиши А. Ультразвук: механический перенос генов в клетки растений путем сонопорации. Biotechnol Adv 2006; 24 : 1–16.
CAS PubMed Google Scholar
van Nocker S, Gardiner SE.Селекция лучших сортов быстрее: применение новых технологий для быстрого внедрения лучших садовых культур деревьев. Hor Res 2014; 1 14022.
Google Scholar
Longhi S, Giongo L, Buti M, Surbanovski N, Viola R, Velasco R et al. Молекулярная генетика и геномика Rosoideae: состояние дел и перспективы на будущее. Hor Res 2014; 1 .
Peng J, Richards DE, Hartley NM, Murphy GP, Devos KM, Flintham JE, et al.Гены «зеленой революции» кодируют мутантные модуляторы ответа гиббереллина. Nature 1999; 400 : 256–2825.
CAS Google Scholar
Ban Y, Honda C, Hatsuyama Y, Igarashi M, Bessho H, Moriguchi T. Выделение и функциональный анализ гена фактора транскрипции MYB, который является ключевым регулятором развития красной окраски кожи яблока. Physiol растительных клеток 2007; 48 : 958–970.
CAS Google Scholar
Alleweldt G, Possingham JV. Прогресс в селекции винограда. Theor Appl Genet 1988; 75 : 669–673.
Google Scholar
Парри М.А., Мэджвик П.Дж., Байон К., Тиралл К., Эрнандес-Лопес А., Баудо М. и др. Открытие мутаций для улучшения урожая. J Exp Bot 2009; 60 : 2817–2825.
CAS PubMed Google Scholar
Шу QY. Превращение селекции с мутациями растений в новую эру: селекция с молекулярными мутациями. В индуцированных мутациях растений в эпоху геномики . Рим: ФАО, 2009: 425–427.
Google Scholar
Hansen G, Wright MS. Последние достижения в трансформации растений. Trends Plant Sci 1999; 4 : 226–231.
CAS PubMed Google Scholar
Джеймс К., Краттигер А.Ф. Глобальный обзор полевых испытаний и коммерциализации трансгенных растений: 1986–1995 гг. ISAAA Briefs 1996; (1).
Брюнинг Дж., Лион Дж. Корпус томата FLAVR SAVR. Calif Agr 2000; 54 : 6–7.
Google Scholar
Кларк Д., Кли Х, Дандекар А.Несмотря на выгоды, коммерциализация трансгенных садовых культур отстает. Calif Agr 2004; 58 : 89–98.
Google Scholar
Азад М.А., Амин Л, Сидик Н.М. Генная технология для борьбы с болезнями, вызванными вирусом кольцевой пятнистости папайи. The Scientific World J 2014; 2014 : 768038.
Google Scholar
Луо К.М., Дуань Х., Чжао Д.Г., Чжэн XL, Дэн В., Чен YQ и др.«GM-ген-deletor»: слитые последовательности узнавания loxP-FRT резко повышают эффективность рекомбиназы FLP или CRE при удалении трансгена из пыльцы и семян растений табака. Завод Биотехнология J 2007; 5 : 263–274.
CAS PubMed Google Scholar
Дин Дж., Дуань Х., Дэн З.Н., Чжао Д.Г., Йи ГДж, Ли Ю. Молекулярные стратегии решения проблем с потоком генов и их потенциальное применение в трансгенных растениях, устойчивых к абиотическому стрессу. Crit Rev Plant Sci 2014; 33 : 190–204.
CAS Google Scholar
Кэрролл Д. Геномная инженерия с целевыми нуклеазами. Annu Rev Biochem 2014; 83 : 409–439.
CAS PubMed Google Scholar
Gaj T, Gersbach CA, Barbas CF, 3rd. Методы геномной инженерии на основе ZFN, TALEN и CRISPR / Cas. Trends Biotechnol 2013; 31 : 397–405.
CAS PubMed PubMed Central Google Scholar
Wyman C, Kanaar R. Восстановление двухцепочечных разрывов ДНК: все хорошо, что хорошо кончается. Annu Rev Genet 2006; 40 : 363–383.
CAS PubMed Google Scholar
Pabo CO, Peisach E, Grant RA. Дизайн и выбор новых белков цинковых пальцев Cys2His2. Annu Rev Biochem 2001; 70 : 313–340.
CAS PubMed Google Scholar
Cathomen T, Joung JK. Нуклеазы «цинковые пальцы»: появляется следующее поколение. Mol Ther 2008; 16 : 1200–1207.
CAS PubMed Google Scholar
Павлетич Н.П., Pabo CO. Распознавание цинкового пальца-ДНК: кристаллическая структура комплекса Zif268-ДНК в точке 2.1 A. Science 1991; 252 : 809–817.
CAS PubMed Google Scholar
Ким Ю.Г., Ча Дж., Чандрасегаран С. Гибридные рестрикционные ферменты: слияния «цинковые пальцы» с доменом расщепления Fok I. Proc Natl Acad Sci USA 1996; 93 : 1156–1160.
CAS PubMed Google Scholar
Perez EE, Wang J, Miller JC, Jouvenot Y, Kim KA, Liu O et al.Установление устойчивости к ВИЧ-1 в CD4 + Т-клетках путем редактирования генома с помощью нуклеаз цинкового пальца. Nat Biotechnol 2008; 26 : 808–816.
CAS PubMed PubMed Central Google Scholar
Дойон Й., Маккаммон Дж. М., Миллер Дж. К., Фараджи Ф., Нго С., Катибах Г. Е. и др. Наследственное целевое разрушение генов у рыбок данио с использованием разработанных нуклеаз типа «цинковые пальцы». Nat Biotechnol 2008; 26 : 702–708.
CAS PubMed PubMed Central Google Scholar
Мэн Х, Нойес М.Б., Чжу Л.Дж., Лоусон Н.Д., Вулф С.А. Нацеленная инактивация генов у рыбок данио с помощью сконструированных нуклеаз типа «цинковые пальцы». Nat Biotechnol 2008; 26 : 695–701.
CAS PubMed PubMed Central Google Scholar
Osakabe K, Osakabe Y, Toki S. Сайт-направленный мутагенез Arabidopsis с использованием специально разработанных нуклеаз цинковых пальцев. Proc Natl Acad Sci USA 2010; 107 : 12034–12039.
CAS PubMed Google Scholar
Townsend JA, Wright DA, Winfrey RJ, Fu F, Maeder ML, Joung JK et al. Высокочастотная модификация генов растений с использованием сконструированных нуклеаз типа «цинковые пальцы». Nature 2009; 459 : 442–445.
CAS PubMed PubMed Central Google Scholar
Шукла В.К., Дойон Ю., Миллер Дж. К., ДеКелвер Р. К., Моеле Е. А., Уорден С. Е. и др.Точная модификация генома у сельскохозяйственных культур Zea mays с помощью нуклеаз типа «цинковые пальцы». Nature 2009; 459 : 437–441.
CAS PubMed Google Scholar
Hsu PD, Zhang F. Рассечение нейронной функции с использованием целевых технологий геномной инженерии. ACS Chem Neurosci 2012; 3 : 603–610.
CAS PubMed PubMed Central Google Scholar
Joung JK, Sander JD.TALENs: широко применяемая технология для целевого редактирования генома. Обзоры природы Mol Cell Biol 2013; 14 : 49–55.
CAS Google Scholar
Boch J, Bonas U. Эффекторы семейства Xanthomonas AvrBs3 типа III: открытие и функции. Annu Rev Phytopathol 2010; 48 : 419–436.
CAS PubMed Google Scholar
Богданове А.Ю., Шорнак С, Лахайе Т.Эффекторы TAL: поиск генов растений для болезней и защиты. Curr Opin Plant Biol 2010; 13 : 394–401.
CAS PubMed Google Scholar
Bonas U, Stall RE, Staskawicz B. Генетическая и структурная характеристика гена авирулентности avrBs3 из Xanthomonas campestris pv. vesicatoria. Mol Gen Genet 1989; 218 : 127–136.
CAS PubMed Google Scholar
Москва MJ, Богданов AJ.Простой шифр управляет распознаванием ДНК эффекторами TAL. Наука 2009; 326 : 1501.
CAS PubMed Google Scholar
Boch J, Scholze H, Schornack S, Landgraf A, Hahn S, Kay S. et al. Нарушение кода специфичности связывания ДНК эффекторов TAL-типа III. Наука 2009; 326 : 1509–1512.
CAS PubMed Google Scholar
Mak AN, Bradley P, Cernadas RA, Bogdanove AJ, Stoddard BL.Кристаллическая структура эффектора TAL PthXo1 связана с его ДНК-мишенью. Наука 2012; 335 : 716–719.
CAS PubMed PubMed Central Google Scholar
Дэн Д., Ян Ц., Пан Х, Махфуз М., Ван Дж., Чжу Дж. К. и др. Структурная основа для последовательного распознавания ДНК эффекторами TAL. Наука 2012; 335 : 720–723.
CAS PubMed PubMed Central Google Scholar
Сандер Дж. Д., Кейд Л., Хайтер С., Рейон Д., Петерсон Р. Т., Джунг Дж. К. и др.Направленное разрушение генов в соматических клетках рыбок данио с использованием сконструированных TALEN. Nat Biotechnol 2011; 29 : 697–698.
CAS PubMed PubMed Central Google Scholar
Хуан П., Сяо А, Чжоу М., Чжу З., Линь С., Чжан Б. Наследственное нацеливание на гены у рыбок данио с использованием индивидуальных TALEN. Nat Biotechnol 2011; 29 : 699–700.
PubMed PubMed Central Google Scholar
Tesson L, Usal C, Menoret S, Leung E, Niles BJ, Remy S. et al.Нокаутные крысы, полученные в результате микроинъекции эмбрионов TALEN. Nat Biotechnol 2011; 29 : 695–696.
CAS PubMed Google Scholar
Миллер Дж.С., Тан С., Цяо Дж., Барлоу К.А., Ван Дж., Ся Д.Ф. и др. Архитектура нуклеазы TALE для эффективного редактирования генома. Nat Biotechnol 2011; 29 : 143–148.
CAS PubMed Google Scholar
Хоккемейер Д., Ван Х., Киани С., Лай С.С., Гао К., Кэссиди Дж. П. и др.Генетическая инженерия плюрипотентных клеток человека с использованием нуклеаз TALE. Nat Biotechnol 2011; 29 : 731–734.
CAS PubMed PubMed Central Google Scholar
Ли Т, Лю Б., Сполдинг М. Х., Weeks DP, Ян Б. Высокоэффективное редактирование генов на основе TALEN дает устойчивый к болезням рис. Nat Biotechnol 2012; 30 : 390–392.
CAS PubMed Google Scholar
Ван И, Ченг Х, Шань Ц., Чжан И, Лю Дж, Гао Ц и др.Одновременное редактирование трех гомеоаллелей в гексаплоидной мягкой пшенице обеспечивает наследственную устойчивость к мучнистой росе. Nat Biotechnol 2014; 32 : 947–951.
CAS PubMed PubMed Central Google Scholar
Кристиан М., Чермак Т., Дойл Э.Л., Шмидт С., Чжан Ф., Хаммель А. и др. Нацеливание на двухцепочечные разрывы ДНК с помощью эффекторных нуклеаз TAL. Genetics 2010; 186 : 757–761.
CAS PubMed PubMed Central Google Scholar
Cermak T, Doyle EL, Christian M, Wang L, Zhang Y, Schmidt C et al. Эффективный дизайн и сборка пользовательских TALEN и других эффекторных конструкций на основе TAL для нацеливания на ДНК. Nucleic Acids Res 2011: gkr218.
Google Scholar
Li L, Piatek MJ, Atef A, Piatek A, Wibowo A, Fang X et al.Быстрое и высокоэффективное создание регуляторов транскрипции и нуклеаз на основе TALE для модификации генома. Завод Мол Биол 2012; 78 : 407–416.
CAS PubMed PubMed Central Google Scholar
Савай С., Охьяма К., Ясумото С., Секи Х., Сакума Т., Ямамото Т. и др. Редуктаза 2 боковой цепи стерола является ключевым ферментом в биосинтезе холестерина, обычного предшественника токсичных стероидных гликоалкалоидов в картофеле. Растительная клетка 2014; 26 : 3763–3774.
CAS PubMed PubMed Central Google Scholar
Лор В.С., Старкер К.Г., Войтас Д.Ф., Вайс Д., Ольшевский Н.Е. Направленный мутагенез гена PROCERA томата с использованием эффекторных нуклеаз, подобных активатору транскрипции. Plant Physiol 2014; 166 : 1288–1291.
PubMed PubMed Central Google Scholar
Йинек М., Чилински К., Фонфара I, Хауэр М., Дудна Ю.А., Шарпантье Э.Программируемая эндонуклеаза ДНК, управляемая двойной РНК, для адаптивного бактериального иммунитета. Наука 2012; 337 : 816–821.
CAS PubMed PubMed Central Google Scholar
Ishino Y, Shinagawa H, Makino K, Amemura M, Nakata A. Нуклеотидная последовательность гена iap, ответственного за превращение изофермента щелочной фосфатазы в Escherichia coli, и идентификация продукта гена. J Bacteriol 1987; 169 : 5429–5433.
CAS PubMed PubMed Central Google Scholar
Wiedenheft B, Sternberg SH, Doudna JA. Генетические системы сайленсинга, управляемые РНК, у бактерий и архей. Природа 2012; 482 : 331–338.
CAS PubMed PubMed Central Google Scholar
Pourcel C, Salvignol G, Vergnaud G. Элементы CRISPR у Yersinia pestis приобретают новые повторы за счет преимущественного поглощения ДНК бактериофага и предоставляют дополнительные инструменты для эволюционных исследований. Микробиология 2005; 151 : 653–663.
CAS PubMed Google Scholar
Болотин А, Куинквис Б, Сорокин А, Эрлих С.Д. Сгруппированные в регулярные промежутки короткие повторы палиндрома (CRISPR) имеют спейсеры внехромосомного происхождения. Микробиология 2005; 151 : 2551–2561.
CAS PubMed Google Scholar
Mojica FJ, Diez-Villasenor C, Garcia-Martinez J, Soria E.Промежуточные последовательности регулярно расположенных прокариотических повторов происходят от чужеродных генетических элементов. J Mol Evol 2005; 60 : 174–182.
CAS PubMed Google Scholar
Deveau H, Garneau JE, Moineau S. Система CRISPR / Cas и ее роль во взаимодействиях фагов и бактерий. Annu Rev Microbiol 2010; 64 : 475–493.
CAS PubMed Google Scholar
Уолш Р.М., Хочедлингер К.Вариант системы CRISPR-Cas9 добавляет гибкости в геномную инженерию. Proc Natl Acad Sci USA 2013; 110 : 15514–15515.
CAS PubMed Google Scholar
Мали П., Ян Л., Эсвелт К.М., Аач Дж., Гуэль М., ДиКарло Дж. Э. и др. РНК-управляемая инженерия генома человека с помощью Cas9. Наука 2013; 339 : 823–826.
CAS PubMed PubMed Central Google Scholar
Fu Y, Foden JA, Khayter C, Maeder ML, Reyon D, Joung JK et al.Высокочастотный мутагенез вне мишени, индуцированный нуклеазами CRISPR-Cas в клетках человека. Nat Biotechnol 2013; 31 : 822–826.
CAS PubMed PubMed Central Google Scholar
Hsu PD, Scott DA, Weinstein JA, Ran FA, Konermann S, Agarwala V et al. Специфичность нацеливания на ДНК РНК-управляемых нуклеаз Cas9. Nat Biotechnol 2013; 31 : 827–832.
CAS PubMed PubMed Central Google Scholar
Pattanayak V, Lin S, Guilinger JP, Ma E, Doudna JA, Liu DR.Высокопроизводительное профилирование расщепления ДНК вне мишени выявляет РНК-запрограммированную специфичность нуклеазы Cas9. Nat Biotechnol 2013; 31 : 839–843.
CAS PubMed PubMed Central Google Scholar
Hou Z, Zhang Y, Propson NE, Howden SE, Chu L-F, Sontheimer EJ et al. Эффективная геномная инженерия плюрипотентных стволовых клеток человека с использованием Cas9 из Neisseria meningitidis. Proc Natl Acad Sci USA 2013; 110 : 15644–15649.
CAS PubMed Google Scholar
Cho SW, Kim S, Kim Y, Kweon J, Kim HS, Bae S. et al. Анализ нецелевых эффектов CRISPR / Cas-производных РНК-управляемых эндонуклеаз и никаз. Genome Res 2014; 24 : 132–141.
CAS PubMed PubMed Central Google Scholar
Fu YF, Sander JD, Reyon D, Cascio VM, Joung JK.Повышение специфичности нуклеазы CRISPR-Cas с помощью усеченных направляющих РНК. Nat Biotechnol 2014; 32 : 279–284.
CAS PubMed PubMed Central Google Scholar
Чанг Н., Сун К., Гао Л., Чжу Д., Сюй Х, Чжу Х и др. Редактирование генома с помощью РНК-управляемой нуклеазы Cas9 в эмбрионах рыбок данио. Cell Res 2013; 23 : 465–472.
CAS PubMed PubMed Central Google Scholar
Hwang WY, Fu Y, Reyon D, Maeder ML, Tsai SQ, Sander JD et al.Эффективное редактирование генома рыбок данио с помощью системы CRISPR-Cas. Nat Biotechnol 2013; 31 : 227–229.
CAS PubMed PubMed Central Google Scholar
Шен Б., Чжан Дж, Ву Х, Ван Дж, Ма К., Ли З и др. Создание генно-модифицированных мышей посредством нацеливания на гены, опосредованные Cas9 / РНК. Cell Res 2013; 23 : 720–723.
CAS PubMed PubMed Central Google Scholar
Tsai SQ, Wyvekens N, Khayter C, Foden JA, Thapar V, Reyon D. et al.Димерные CRISPR РНК-управляемые нуклеазы Fok I для высокоспецифичного редактирования генома. Nat Biotechnol 2014; 32 : 569–576.
CAS PubMed PubMed Central Google Scholar
Guilinger JP, Thompson DB, Liu DR. Слияние каталитически неактивной Cas9 с нуклеазой Fok I повышает специфичность модификации генома. Nat Biotechnol 2014; 32 : 577–582.
CAS PubMed PubMed Central Google Scholar
Li JF, Norville JE, Aach J, McCormack M, Zhang D, Bush J et al. Мультиплексное и гомологичное редактирование генома, опосредованное рекомбинацией, у Arabidopsis и Nicotiana benthamiana с использованием направляющей РНК и Cas9. Nat Biotechnol 2013; 31 : 688–691.
CAS PubMed PubMed Central Google Scholar
Feng Z, Mao Y, Xu N, Zhang B, Wei P, Yang DL et al.Анализ нескольких поколений показывает наследование, специфичность и паттерны модификаций генов, вызванных CRISPR / Cas у Arabidopsis. Proc Natl Acad Sci USA 2014; 111 : 4632–4637.
CAS PubMed Google Scholar
Shan Q, Wang Y, Li J, Zhang Y, Chen K, Liang Z et al. Направленная модификация генома сельскохозяйственных культур с помощью системы CRISPR-Cas. Nat Biotechnol 2013; 31 : 686–688.
CAS PubMed Google Scholar
Мяо Дж., Го Д., Чжан Дж., Хуанг Кью, Цинь Г, Чжан Х и др. Направленный мутагенез риса с использованием системы CRISPR-Cas. Cell Res 2013; 23 : 1233–1236.
CAS PubMed PubMed Central Google Scholar
Се К., Ян Ю. РНК-управляемое редактирование генома растений с использованием системы CRISPR-Cas. Завод Мол 2013; 6 : 1975–1983.
CAS PubMed Google Scholar
Цзя Х.Г., Ван Н. Целевое редактирование генома сладкого апельсина с использованием Cas9 / sgRNA. PloS one 2014; 9 : e93806.
PubMed PubMed Central Google Scholar
Ван Х, Ян Х, Шивалила С.С., Давлати М.М., Ченг А.В., Чжан Ф. и др. Одношаговое создание мышей, несущих мутации в нескольких генах, с помощью CRISPR / Cas-опосредованной геномной инженерии. Cell 2013; 153 : 910–918.
CAS PubMed PubMed Central Google Scholar
Nagamangala Kanchiswamy C, Sargent DJ, Velasco R, Maffei ME, Malnoy M. Ждем генетически отредактированных плодовых культур. Trends Biotechnol 2015; 33 : 62–64.
CAS PubMed Google Scholar
Яник Дж.Селекция садовых растений: достижения прошлого, направления на будущее. Proc. ЕСТЬ на Hort. в Азиатско-Тихоокеанском регионе 2004; 694 : 61–65.
Google Scholar
Сирам Н.П., Адамс Л.С., Харди М.Л., Хебер Д. Общий экстракт клюквы в сравнении с его фитохимическими составляющими: антипролиферативный и синергетический эффекты против линий опухолевых клеток человека. J AgrFood Chem 2004; 52 : 2512–2517.
CAS Google Scholar
Джозеф Дж. А., Шукит-Хейл Б., Денисова Н. А., Белински Д., Мартин А., Макьюен Дж. Дж. И др. Устранение возрастного снижения передачи нейронных сигналов, когнитивных и двигательных нарушений поведения с помощью пищевых добавок из черники, шпината или клубники. J Neurosci 1999; 19 : 8114–8121.
CAS PubMed Google Scholar
Renaud S, de Lorgeril M.Вино, алкоголь, тромбоциты и французский парадокс ишемической болезни сердца. Lancet 1992; 339 : 1523–1526.
CAS PubMed Google Scholar
Hou DX, Fujii M, Terahara N, Yoshimoto M. Молекулярные механизмы химиопрофилактического действия антоцианидинов. Journal Biomed Biotechnol 2004; 5 : 321–325.
Google Scholar
Allan AC, Hellens RP, Laing WA.Факторы транскрипции MYB, которые окрашивают наши плоды. Trends Plant Sci 2008; 13 : 99–102.
CAS PubMed Google Scholar
Альберт Н.В., Дэвис К.М., Льюис Д.Х., Чжан Х., Монтефиори М., Брендолиз С. и др. Консервативная сеть активаторов и репрессоров транскрипции регулирует пигментацию антоцианов у эвдикотов. Растительная клетка 2014; 26 : 962–980.
CAS PubMed PubMed Central Google Scholar
Холлендер, Калифорния, Дардик С.Молекулярные основы архитектуры дерева покрытосеменных. Новый Фитол 2015; 206 : 541–556.
PubMed Google Scholar
Gomez-Roldan V, Fermas S, Brewer PB, Puech-Pagès V, Dun EA, Pillot J-P et al. Стриголактон подавляет ветвление побегов. Nature 2008; 455 : 189–194.
CAS PubMed Google Scholar
Умехара М., Ханада А., Йошида С., Акияма К., Арите Т., Такеда-Камия Н. и др.Подавление ветвления побегов новыми терпеноидными гормонами растений. Nature 2008; 455 : 195–200.
CAS PubMed Google Scholar
Рейд М.С., Ву М-Дж. Этилен и старение цветков. Регулятор роста растений 1992; 11 : 37–43.
CAS Google Scholar
Savin KW, Baudinette SC, Graham MW, Michael MZ, Nugent GD, Lu C-Y et al.Антисмысловая РНК АСС-оксидазы задерживает старение лепестков гвоздики. HortScience 1995; 30 : 970–972.
CAS Google Scholar
Сибуя К., Барри К.Г., Сиарди Дж.А., Лукас Х.М., Андервуд Б.А., Нуризаде С. и др. Центральная роль PhEIN2 в ответах на этилен во время развития растений петунии. Plant Physiol 2004; 136 : 2900–2912.
CAS PubMed PubMed Central Google Scholar
Голдберг Н.П.Мучнистая роса. Сборник болезней перца APS Press: St Paul 2003: 19–20.
Google Scholar
Buschges R, Hollricher K, Panstruga R, Simons G, Wolter M, Frijters A et al. Ген Mlo ячменя: новый элемент контроля устойчивости растений к патогенам. Cell 1997; 88 : 695–705.
CAS PubMed Google Scholar
Devoto A, Hartmann HA, Piffanelli P, Elliott C, Simmons C, Taramino G et al.Молекулярная филогения и эволюция растительного семи трансмембранного семейства MLO. J Mol Evol 2003; 56 : 77–88.
CAS PubMed Google Scholar
Pavan S, Schiavulli A, Appiano M, Marcotrigiano AR, Cillo F, Visser RGF et al. Устойчивость к мучнистой росе гороха er1 связана с мутациями потери функции в гомологичном локусе MLO. Theor Appl Genet 2011; 123 : 1425–1431.
PubMed PubMed Central Google Scholar
Jiwan D, Roalson EH, Main D, Dhingra A. Антисмысловая экспрессия гена локуса O (PpMlo1) устойчивости к мучнистой росе персика придает межвидовую устойчивость к мучнистой росе у Fragaria × ananassa. Transgenic Res 2013; 22 : 1119–1131.
CAS PubMed Google Scholar
Bolger ME, Weisshaar B, Scholz U, Stein N, Usadel B, Mayer KFX.Секвенирование генома растений — приложения для улучшения сельскохозяйственных культур. Curr Opin Biotech 2014; 26 : 31–37.
CAS PubMed Google Scholar
Zhang S, Chen W, Xin L, Gao Z, Hou Y, Yu X et al. Геномные варианты генов, связанных с тремя признаками садоводства2 яблони, выявленные при повторном секвенировании генома. Horticulture Res 2014; 1 : 14045.
Google Scholar
Мур Ф. Э., Рейон Д., Сандер Дж. Д., Мартинез С. А., Блэкберн Дж. С., Хайтер С. и др.Улучшенный соматический мутагенез у рыбок данио с использованием эффекторных нуклеаз, подобных активаторам транскрипции (TALEN). PLoS One 2012; 7 : e37877.
CAS PubMed PubMed Central Google Scholar
Быстрая селекция и замена разновидностей имеют решающее значение для адаптации систем земледелия в развивающихся странах к изменению климата
Основные моменты
- •
Непрерывный оборот разновидностей сельскохозяйственных культур имеет решающее значение для адаптации к изменению климата.
- •
Фермеры, использующие коммерческие системы земледелия с умеренным климатом, с быстрой сменой сортов, лучше всего защищены.
- •
Селекционным программам необходим беспрепятственный доступ к элитным сортам из регионов, которые сейчас сталкиваются с их будущим климатом.
- •
Быстрые и эффективные циклы размножения необходимы для адаптации к изменению климата в реальном времени.
- •
Фермеры в развивающихся странах подвергаются риску из-за медленных циклов размножения и смены сортов.
Реферат
Селекция растений — ключевой механизм адаптации систем земледелия к изменению климата. Многие дискуссии о селекции в условиях изменения климата сосредоточены на генах, сильно влияющих на устойчивость к жаре и засухе, но фенология и устойчивость к стрессу очень полигенны. Таким образом, адаптация будет в основном результатом постоянной корректировки частот аллелей во многих локусах посредством быстрого цикла селекции, которая обеспечивает постоянный поток постепенно улучшаемых сортов.Это потребует доступа к элитной зародышевой плазме из других регионов, сокращенных циклов размножения и систем тестирования в нескольких местах, которые адекватно отбирают целевую популяцию окружающей среды. Целью систем селекции и семеноводства, обслуживающих мелких фермеров, должно быть обеспечение того, чтобы они использовали сорта, выведенные за последние 10 лет. Быстрый сортовой оборот должен поддерживаться активным распространением новых сортов и активным изъятием устаревших. Коммерческие семеноводческие системы в регионах с умеренным климатом достигают этого за счет конкурентных рынков семян, но в развивающихся странах большинство культур не обслуживается конкурентоспособными коммерческими семеноводческими системами, и многие сорта появились в конце Зеленой революции (конец 1970-х годов, когда второе поколение современных сортов риса и пшеницы).Эти устаревшие сорта были выведены в климате, отличном от сегодняшнего, что подвергало фермеров риску. Чтобы снизить этот риск, необходима усиленная селекционная система с более свободным международным обменом элитными сортами, короткими циклами селекции, высокой интенсивностью селекции, широкомасштабным фенотипированием и точной селекцией, поддерживаемой геномными технологиями. Правительствам необходимо стимулировать системы выпуска и распространения сортов для постоянной замены устаревших сортов.
Ключевые слова
Адаптация к изменению климата
Быстрое селекционное производство
Замена сортов
Обмен зародышевой плазмы
Генетические выгоды
Семенные системы
Рекомендуемые статьи Цитирующие статьи (0)
© 2017 Bill & Melinda Gates Foundation.Опубликовано Elsevier B.V.
Рекомендуемые статьи
Цитирующие статьи
Инновации в селекции растений — Международная федерация семян
Около
«Инновации в селекции растений» — это термин, который мы используем для описания постоянно развивающихся идей и практик, которые расширяют сферу селекции растений. Сегодняшние инновации в селекции растений разрабатываются с использованием передовой науки и технологий, включая клеточную биологию, генное картирование и селекцию с помощью маркеров.
Новые методы селекции (NBT) и методы точной селекции (PBT) относятся к инструментам и методам, используемым для более точного и быстрого выведения новых сортов. Эти методы отражают научные открытия того времени. Например, в двадцатом веке возросшее понимание физиологии растений и молекулярной биологии ускорило разработку новых сортов, которые, вероятно, не были бы получены селекционерами, использующими только традиционный отбор. Инновации XXI века основываются на этих знаниях для создания сортов в ответ на экологические, сельскохозяйственные и социальные проблемы нашего времени.Инновации в селекции растений не могут и не заменят традиционные методы, они просто увеличивают набор инструментов, доступных селекционерам растений.
Благодаря инновациям мы можем производить улучшенные сорта, которые сохраняют и потенциально повышают урожайность и лучше приспособлены к борьбе с болезнями и последствиями изменения климата, такими как засуха или наводнения, поддерживая устойчивое сельское хозяйство и продовольственную безопасность.
Преимущества
«Семя — это жизнь» — это девиз Международной федерации семян.Наше видение — мир, в котором семена высочайшего качества доступны всем. Наша миссия — продвигать селекцию растений и инновации в семеноводстве. Мы знаем, что выращивание более качественных семян в конечном итоге улучшает жизнь.
- Устойчивый и повышенный урожай сельскохозяйственных культур: Семена, устойчивые к вредителям и болезням и способные противостоять последствиям изменения климата, позволяют фермерам собирать более обильные и надежные урожаи. Семена, адаптированные к различным средам и условиям выращивания, означают, что мы можем производить больше урожая с одной и той же земли.
- Улучшенное питание: Благодаря инновациям в растениеводстве фермеры могут выращивать сорта с высоким содержанием белка — недостаточное количество белка в рационе является существенным фактором недостаточного питания.
- Лучшее качество продуктов питания: Благодаря инновациям в области растениеводства продукты питания дольше остаются свежими. Многие виды фруктов и овощей можно легче транспортировать и хранить, что продлевает срок их хранения и сокращает количество пищевых отходов.
Постановление
Решения глобальных проблем в области регулирования селекции растений можно найти только при совместной работе всех заинтересованных сторон.По мере роста мировой торговли семенами растет потребность в согласованных критериях, определяющих, как регулируются продукты с использованием новейших методов селекции растений.
Страны имеют ряд различных систем для оценки и регулирования новых продуктов, поступающих на рынок, что может привести к возникновению торговых барьеров и снижению конкурентоспособности. Чтобы облегчить торговлю и перемещение семян по всему миру, нам нужно начать с равных условий.
Ключевой проблемой для селекционеров во всем мире является обеспечение единообразия критериев, используемых для регулирования продуктов, разработанных с помощью новейших методов селекции растений.Четкая государственная политика необходима для обеспечения того, чтобы использование этих методов разведения не застопорилось на стадии исследований и разработок.