
Изучаем царство Растения. Биология, Бактерии. Грибы. Растения (5–6 класс): уроки, тесты, задания.
Вход Вход Регистрация Начало Новости ТОПы Учебные заведения Предметы Проверочные работы Обновления Переменка Поиск по сайту Отправить отзыв- Предметы
- Биология
 org/BreadcrumbList»>
org/BreadcrumbList»>-
Низшие растения — водоросли. Высшие споровые растения
-
Отличительные признаки голосеменных растений
Общая характеристика растений.
 Низшие и высшие растения
Низшие и высшие растенияРастения – это многоклеточные фотосинтезирующие эукариотические организмы.
Все растения входят в отдельное биологическое царство Растения. Вспомним, что в настоящее время большинство учёных выделяют ещё три царства живых организмов: Животные, Грибы и Бактерии. К каждому царству относятся организмы, которые имеют общие особенности внешнего и внутреннего строения.
К растениям относятся мхи, папоротники, хвощи, плауны, голосеменные и покрытосеменные растения. Нередко к растениям относят также водоросли.
По данным Международного союза охраны природы, было описано около 320 тысяч видов растений, из них около 280 тысяч видов покрытосеменных. Однако это число увеличивается, так как постоянно открываются новые виды. Разные растения характерны для лугов, лесов, безводных пустынь и топких болот.
Растения являются объектом
исследования науки ботаники. Ботаника – это комплексная наука,
включающая несколько разделов:
Ботаника – это комплексная наука,
включающая несколько разделов:
· Анатомия растений изучает их внешнее и внутреннее строение.
· Физиология растений изучает процессы их жизнедеятельности.
· Систематика растений – классификацию.
· Геоботаника – распространение растений по материкам и континентам.
· Экология растений – их связь друг с другом и окружающей природой.
Несмотря на огромное разнообразие растений, для них характерны общие признаки. Рассмотрим их.
· Клетки растений имеют плотные клеточные стенки, содержащие целлюлозу.
· В клетках находятся зелёные пластиды – хлоропласты, содержащие зелёный пигмент хлорофилл. Благодаря им большинство растений имеет зелёный цвет. Но есть и исключения. Заразиха (или петров крест) имеет светло-бурую или желтоватую окраску.
· Растения ведут прикреплённый образ жизни.
· Запасное
питательное вещество в клетках – крахмал.
· Растут в течение всей жизни.
Все растения в зависимости от строения делят на две большие группы – низшие растения и высшие растения. Рассмотрим эти группы.
К низшим растениям относятся водоросли. Тело наиболее примитивных водорослей может состоять из одной клетки, например, хлорелла и хламидомонада. К многоклеточным водорослям относятся, например, ульва и анфельция.
Низшие растения не имеют настоящих корней, стеблей и листьев. Их тело называется «слоевище» или «таллом». Органы размножения низших растений устроены просто, и чаще всего они одноклеточные.
В
ходе эволюции низшие растения появились раньше высших. Однако в настоящее время
они широко распространены по всему земному шару. Для низших растений характерна
огромная биомасса и высокая способность к размножению. В водных сообществах они
являются первым звеном цепей питания (их основой). Поэтому они очень важны для
нормального течения энергетических процессов в живой природе.
Ранее учёные относили к растениям бактерии, сине-зелёные водоросли (или цианобактерии), миксомицеты (или слизевики), грибы и лишайники. Однако это отдельные царства и группы в живой природе. Грибы и слизевики, в отличие от растений, питаются готовыми органическими веществами и не осуществляют фотосинтез. Лишайники – это симбиоз гриба и водоросли. Они выделены в отдельную систематическую группу. Бактерии и цианобактерии – прокариоты. Они не имеют оформленного ядра и тоже не могут считаться растениями.
К высшим растениям относятся мхи, хвощи, плауны, папоротники, голосеменные и покрытосеменные растения.
Среди высших растений выделяют две большие группы: споровые и семенные растения. К споровым растениям
Семена
голосеменных растений лежат открыто на чешуевидных листьях, образующих шишку. Эти растения не образуют цветков и плодов.
Эти растения не образуют цветков и плодов.
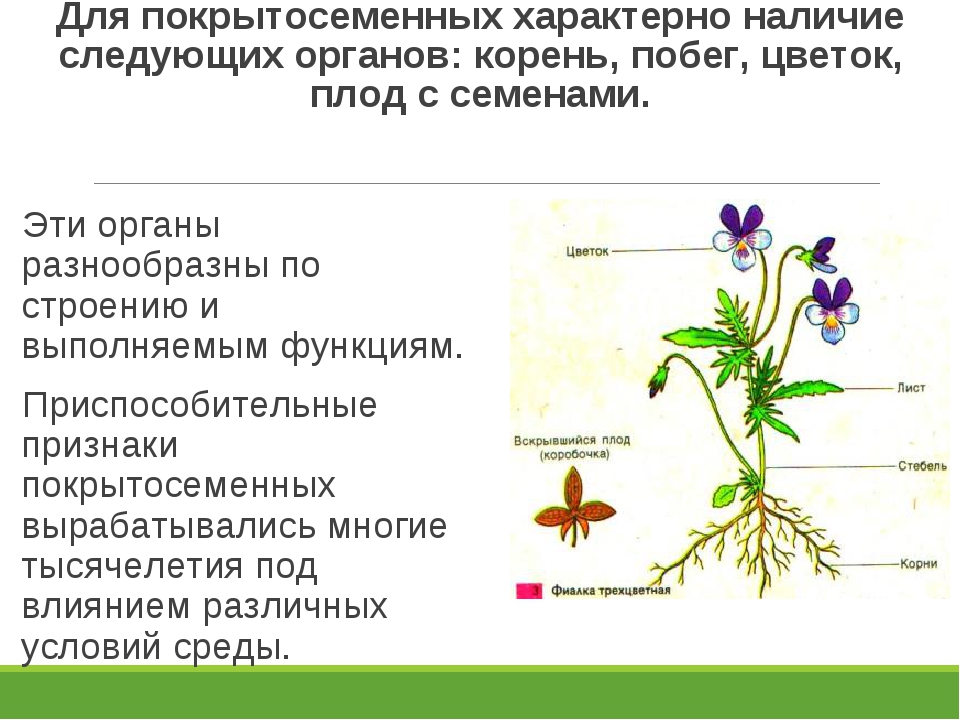
Покрытосеменные – наиболее высокоорганизованные растения. Их главная особенность – наличие органа полового размножения (цветка).
Растения (в первую очередь, покрытосеменные) представлены разными жизненными формами – среди них есть деревья, кустарники и травы. Деревья – это обычно крупные растения с многолетними деревянистыми стволами. Кустарники отличаются от деревьев тем, что имеют не один ствол, а несколько стволиков, отходящих от общего основания. Травы (или травянистые растения) имеют зелёные сочные стебли, они почти всегда ниже деревьев и кустарников. Но есть и исключения. Банан растёт в высоту от 2 до 7 метров. Существуют и крошечные травянистые растения. На поверхности водоёмов живёт ряска, размер каждого растения – несколько миллиметров.
Высшие растения имеют чётко дифференцированные ткани,
которые выполняют специализированные функции. Это является важной отличительной
особенностью высших растений. В теле растений выделяют образовательные,
основные, проводящие, механические и покровные ткани.
В теле растений выделяют образовательные,
основные, проводящие, механические и покровные ткани.
У высших растений из тканей образуются органы – корень, стебель и листья. Исключение составляют мхи. Они имеют стебель и листья, но не имеют корней. В почве они закрепляются с помощью ризоидов.
Выделяют две группы органов. Вегетативные органы (корень, стебель, лист) участвуют в процессах жизнедеятельности и в бесполом размножении.
Высшие растения имеют многоклеточные органы полового и бесполого размножения. Органы полового размножения – гаметангии. В них образуются половые клетки – гаметы. Это отличает половые органы высших растений от низших растений, у которых все клетки способны преобразовываться в гаметы.
Мужские
гаметангии называются «антеридии», женские – «архегонии». Они защищены
оболочкой, образованной бесплодными клетками. В антеридиях образуется большое
количество мелких мужских гамет (сперматозоидов),
которые имеют жгутик и способны передвигаться. Неподвижные мужские гаметы
называют спермиями. В архегониях
развивается одна яйцеклетка. В
отличие от сперматозоидов, яйцеклетки неподвижны, имеют более крупные размеры и
обладают значительным запасом питательных веществ.
В антеридиях образуется большое
количество мелких мужских гамет (сперматозоидов),
которые имеют жгутик и способны передвигаться. Неподвижные мужские гаметы
называют спермиями. В архегониях
развивается одна яйцеклетка. В
отличие от сперматозоидов, яйцеклетки неподвижны, имеют более крупные размеры и
обладают значительным запасом питательных веществ.
Рассмотрим жизненный цикл высших растений на примере папоротниковидных. В жизненном цикле существует две фазы: гаметофит и спорофит. Они закономерно сменяют друг друга. Гаметофит – гаплоидная многоклеточная фаза в жизненном цикле. Гаметофит развивается из спор и образует половые клетки (гаметы), осуществляет половое размножение. Гаплоидные гаметы сливаются и образуют диплоидную зиготу. Она даёт начало зародышу. Необходимо сказать, что у низших растений образовавшиеся в результате деления зиготы клетки сразу же используются для построения слоевища и в последующем мало изменяются.
У
высших растений гаметы всегда образуются в результате митоза. Это принципиально
отличает их от половых клеток животных, которые образуются в результате мейоза.
Фазу гаметофита ещё называют гаметофазой или гаплофазой. Гаметофит бывает однодомным или
двудомным. На однодомном гаметофите происходит одновременное развитие
яйцеклетки и сперматозоидов. У двудомных растений гаметофиты формируют либо
только мужские органы, либо только женские.
Это принципиально
отличает их от половых клеток животных, которые образуются в результате мейоза.
Фазу гаметофита ещё называют гаметофазой или гаплофазой. Гаметофит бывает однодомным или
двудомным. На однодомном гаметофите происходит одновременное развитие
яйцеклетки и сперматозоидов. У двудомных растений гаметофиты формируют либо
только мужские органы, либо только женские.
В ходе эволюции растений происходит постепенная редукция гаметофита. Чем выше уровень организации гаметофита, тем лучше развиты органы полового размножения (гаметангии). У мхов они многочисленны, у папоротниковидных их меньше, у голосеменных они подвергаются редукции, у всех цветковых вовсе не образуются.
Вернёмся к жизненному циклу папоротниковидных. Из зиготы
развивается спорофит. Это диплоидная многоклеточная фаза
в жизненном цикле растений. Так как спорофит образован диплоидными
клетками, то его другое название – диплофаза.
Спорофит не образует гаметы, на нём формируются споры. Он
образован диплоидными клетками, но его споры гаплоидные, так как образуются в
процессе мейоза. Споры растений, грибов, некоторых других организмов
развиваются в особых многоклеточных органах бесполого размножения – спорангиях.
Он
образован диплоидными клетками, но его споры гаплоидные, так как образуются в
процессе мейоза. Споры растений, грибов, некоторых других организмов
развиваются в особых многоклеточных органах бесполого размножения – спорангиях.
Количество
образовавшихся при этом спор зависит от того, какое это растение – равноспоровое или разноспоровое. У
равноспоровых в результате мейоза возникают четыре споры (тетрада).
Они имеют одинаковое строение и размеры. В
зависимости от внешних условий из неотличимых с виду спор развиваются
гаметофиты, на которых формируются как мужские, так и женские половые
органы. У разноспоровых растений
образуется два вида спор: женские мегаспоры и мужские микроспоры. При мейозе
образуется только одна мегаспора, три меньшие клетки погибают. По традиции
мегаспоры называют женскими спорами, но употребление этого термина в прямом
смысле некорректно, так как спорофит представляет собой бесполое поколение.
Следовательно, отдельные особи изначально нельзя делить на мужские и женские.
На формирующихся далее гаметофитах образуются только женские половые органы (архегонии). Также в результате мейоза образуются четыре микроспоры, имеющие более мелкие размеры, чем макроспоры. Микроспоры традиционно считают мужскими спорами, так как на развивающихся из них гаметофитах формируются только мужские половые органы (антеридии).
Вы уже знаете, что гаметофит равноспоровых растений (плаунов, хвощей и части папоротников) хорошо развит. Он способен к фотосинтезу и обеспечивает половые органы органическими веществами. В дальнейшем у разноспоровых растений, к которым относятся голосеменные и покрытосеменные, происходит редукция гаметофита. Часто он не выходит за пределы споры и не осуществляет фотосинтез. Органические вещества, которые необходимы для его развития, изначально запасаются в споре.
В зависимости от преобладания гаплоидной или
диплоидной фазы высшие растения делят на две группы. Первая – растения, у которых преобладает
гаметофит, а спорофит развит слабо (моховидные).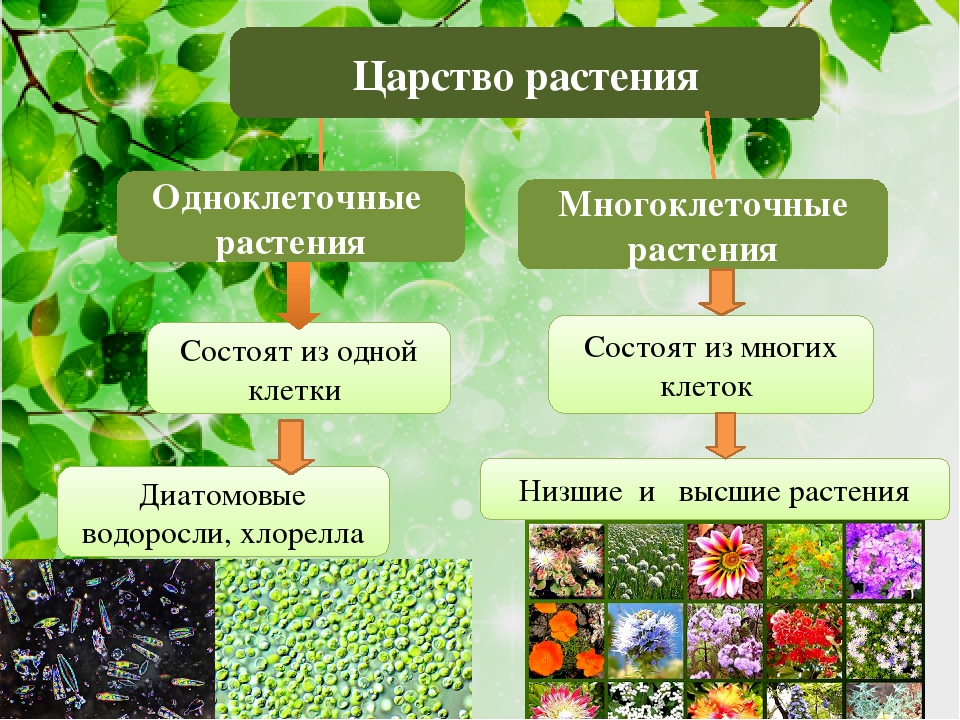 Плауновидные, хвощевидные,
папоротниковидные, голосеменные и покрытосеменные относятся ко второй группе.
Плауновидные, хвощевидные,
папоротниковидные, голосеменные и покрытосеменные относятся ко второй группе.
У этих растений преобладает спорофит, гаметофит при этом в большей или меньшей степени редуцируется.
Значение растений в природе велико. Они обогащают воздух кислородом и поглощают углекислый газ. Растения служат пищей растительноядным животным, которыми в свою очередь питаются хищники.
Люди тоже питаются растениями и продуктами их переработки. Используют растения как сырьё для различных отраслей промышленности, для приготовления лекарств, как строительный материал и топливо. Горох, фасоль выращивают для получения семян. Яблони, груши, вишни, томаты дают сочные плоды. Морковь, свёклу, петрушку выращивают ради корней. Розы, астры, жасмин, сирень разводят ради красивых цветков. Из плодов пшеницы готовят хлеб, макароны и кондитерские изделия.
4.4.1. Общая характеристика царства Растения
Ботаника изучает строение, жизнедеятельность, закономерности индивидуального и эволюционного развития, распространение и условия обитания растений.
Значение растений в биосфере огромно. Фотосинтезирующие растения являются источником органического вещества и кислорода, оказывают значительное влияние на климат. Жизнь современного человека зависит от использования культурных растений, которых в настоящее время насчитывается около 1500 видов. Человек широко использует природные и культивируемые растения для изготовления лекарственных препаратов или в декоративных целях.
Размеры и строение растений — от 2—3 мкм у одноклеточных водорослей (хлорелла) до сложно устроенных покрытосеменных с высокодифференцированными тканями и органами (эвкалипты до 150 м).
Общие черты растений:
Наличие в составе клеток твердой клеточной оболочки из целлюлозы.
Способ питания — адсорбционный (всасывательный).
Прикрепление к твердому субстрату и ограничение подвижности.
Расселение растений зачатками (спорами, семенами), находящимися в состоянии покоя.
По типу питания растения — автотрофы, запасное питательное вещество — крахмал.
Закономерное чередование двух поколений в жизненном цикле, проходящем от зиготы одного растения до зиготы другого: бесполого диплоидного поколения — спорофита, и полового гаплоидного — гаметофита.
При чередовании поколений гаметофит — растительный организм, на котором формируются гаметы, закономерно сменяется спорофитом: гаплоидные гаметы сливаются диплоидная зигота зародыш
взрослое растение — спорофит (имеет спорангий и производит споры, одинаковые или разные. При образовании спор происходит мейоз, поэтому споры гаплоидные) из спор вырастает гаметофит (гаплоидное поколение, есть половые органы, производящие сперматозоиды и яйцеклетки). Все наземные растения имеют половые органы одного типа: мужские называются антеридии, а женские — архегонии. В мужских органах образуется много сперматозоидов, а в женских — одна крупная неподвижная яйцеклетка.
Гаметофит и спорофит могут быть одинаковыми как по строению, так и по продолжительности жизни. Но у наземных растений оба поколения различны. При этом оба поколения могут быть самостоятельными или одно развивается на другом. Так, у мхов спорофит (коробочка со спорами) — часть одного растения, тело которого является гаметофитом. У семенных растений сильно редуцированный, лишенный хлорофилла гаметофит представлен клетками зародышевого мешка.
Но у наземных растений оба поколения различны. При этом оба поколения могут быть самостоятельными или одно развивается на другом. Так, у мхов спорофит (коробочка со спорами) — часть одного растения, тело которого является гаметофитом. У семенных растений сильно редуцированный, лишенный хлорофилла гаметофит представлен клетками зародышевого мешка.
В процессе эволюции при приспособлении к дефициту воды происходили постепенная редукция (сокращение продолжительности жизненных циклов) полового поколения — гаметофита, упрощение половых органов и увеличение размеров бесполого поколения — спорофита.
Около 600 тыс. видов растений условно делят на:
низшие —тело не расчленено на органы и ткани и называется слоевищем, или талломом. К этой группе относятся водоросли и лишайники.
высшие — имеются органы (корень, стебель и лист), образованные сложно дифференцированными тканями. Зигота высших растений развивается в многоклеточный зародыш. К высшим растениям относятся отделы моховидные, плауновидные, хвощевидные, папоротниковидные, голосеменные и покрытосеменные (цветковые).
К высшим растениям относятся отделы моховидные, плауновидные, хвощевидные, папоротниковидные, голосеменные и покрытосеменные (цветковые).
Растения появились на Земле около 2 млрд лет назад. Первоначально развитие растительных организмов происходило в водной среде, что привело к появлению – водорослей. Затем растения стали осваивать сушу. Этому способствовало возникновение следующих ароморфозов:
– возникновение фотосинтеза;
– возникновение эукариотического строения клеток;
– возникновение мейоза и оплодотворения;
– возникновение многоклеточности и дифференциации клеток с образованием тканей и органов;
– возникновение чередования гаплоидного и диплоидного поколений;
– возникновение семени;
– возникновение цветка.
Эволюция растений шла в направлении от споровых к семенным, от низших к высшим.
Краткая характеристика отделов царства растений
Прежде чем начать знакомство с основными отделами царства растений, рассмотрим их сравнительную характеристику.
I. Низшие растения. Водоросли
Самая древняя группа растений на Земле, объединяющая одно- и многоклеточные организмы. Они дали начало наземным растениям. Тело их называется талломом, или слоевищем. Все они фотосинтезирующие, хотя и имеют различную окраску. Размножаются вегетативно, спорами и половым способом.
Отдел зеленые водоросли
Около 15 000 видов. Одноклеточные, колониальные и многоклеточные растения. Фотосинтезируют. Наиболее характерные представители: хламидомонада, хлорелла, улотрикс, спирогира, ульва.
Отдел диатомовые водоросли
Около 15 000 видов одноклеточных и колониальных растений. Характеризуются наличием на поверхности таллома кремнеземного панциря. Представители — пипуллярия, навикула, цимбелла и др.
Отдел бурые водоросли
Около 1 500 видов исключительно морских растений. Отличаются бурой окраской таллома и четким чередованием поколений. Имеют запасное вещество ламинарии. Основные представители: макроцистис, фукус, ламинария, саргассум.
Отдел красные водоросли, или багрянки
Около 4 000 видов морских обитателей, таллом которых розово-красного цвета. Для них характерно отсутствие жгутиковых стадий в развитии. Они могут жить на очень больших глубинах, что очень важно для поддержания жизни других живых существ в этих условиях.
II. Высшие растения — Высшие споровые растения
Отдел риниеобразные, или риниофиты
Исключительно ископаемые растения, жившие на Земле около 450 млн лет тому назад. Имели очень примитивное строение: не имели листьев, ветвились дихотомически, к почве прикреплялись ризоидами. Все они вымерли. Сюда относились куксопия, риния и др. Дали начало всем остальным растениям.
Отдел псилотообразные, или псилофиты
Известно около 20 видов. Это современные тропические и субтропические растения. Небольших размеров, безлистные с дихотомическим ветвлением. В жизненном цикле преобладает спорофит. Сюда относятся псилоты.
Отдел мохообразные
характеризуется отсутствием корней, настоящих проводящих пучков, мелкими листьями или отсутствием их (слоевище), преобладанием гаметофита в цикле развития. Гаметофит надземный зеленый, несет на себе свободно сидящие антеридии и архегопии. Сперматозоиды двух жгутиковые. Известно около 35 000 видов. Наиболее распространенные — кукушкин лен, маршанция, сфагнум.
Гаметофит надземный зеленый, несет на себе свободно сидящие антеридии и архегопии. Сперматозоиды двух жгутиковые. Известно около 35 000 видов. Наиболее распространенные — кукушкин лен, маршанция, сфагнум.
Отдел ил лунообразные, или ликоподиофиты
Древнейшие вымершие и современные высшие споровые растения, насчитывающие около 1 000 видов. Характеризуются наличием корней, проводящих пучков, мелкими листьями, преобладанием спорофита. Споры у одних видов одинаковые, у других разные. Возникновение разноспоровости — приспособление, которое способствовало выживанию на суше. Наиболее распространены: плаун булавовидный, лепидодендрон, сигиллярия и др.
Отдел хвощеобразные, или эквизетофиты
Известно более 30 000 видов. Сюда относятся многолетние травянистые растения, имеющие развитые корни, проводящие пучки и расположенные мутовками на стебле листья. В жизненном цикле преобладает спорофит. Его тело расчленено на узлы и междоузлья. Листья не развиваются и имеют вид зубчиков.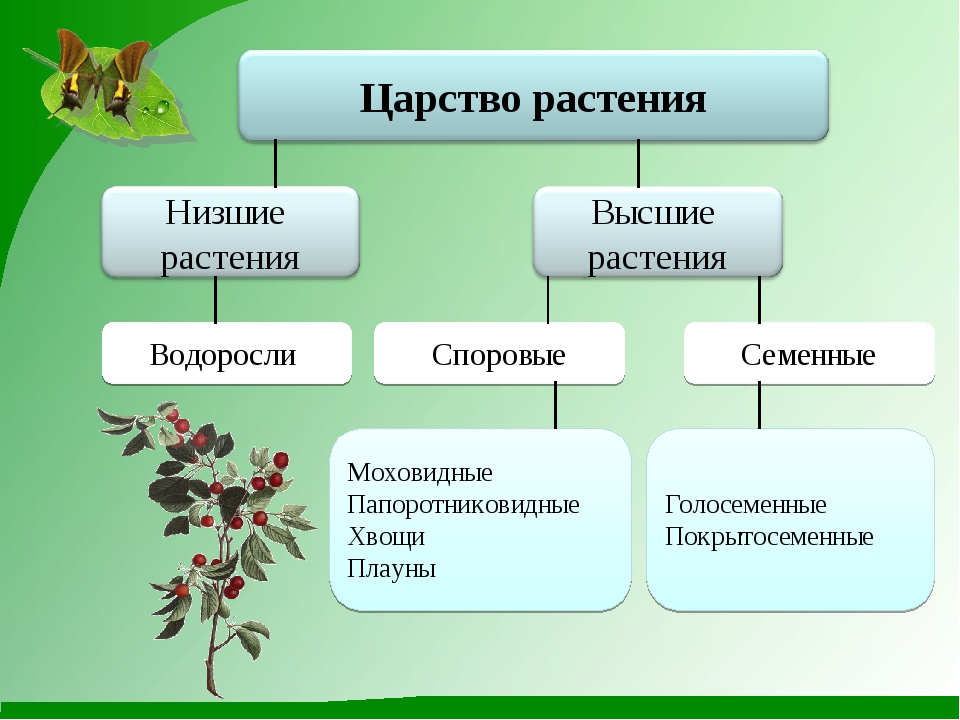 Фотосинтез осуществляется в стебле. Хвощи — равноспоровые растения, но гаметофиты у них обычно раздельнополые. Наиболее распространены: хвощ полевой, хвощ лесной, хвощ луговой, хвощ болотный.
Фотосинтез осуществляется в стебле. Хвощи — равноспоровые растения, но гаметофиты у них обычно раздельнополые. Наиболее распространены: хвощ полевой, хвощ лесной, хвощ луговой, хвощ болотный.
Отдел папоротникообразные, или полиподиофиты
Насчитывает около 10 000 видов. Характеризуется наличием корней, проводящих пучков, крупными листьями, преобладанием спорофита, наличием видов с одинаковыми и разными спорами. Спорангии собраны в сорусы. Гаметофит наземный зеленый. Антеридии и архегонии погружены в ткань гаметофита. Сперматозоиды многожгутиковые. Типичные представители: щитовник мужской, кочедыжник женский, орляк обыкновенный и др.
II. Высшие растения — Высшие семенные растения
Отдел голосеменные, или пинофиты
Насчитывает около 660 видов преимущественно древесных растений. Характеризуется сильно развитой корневой системой, крупными и мелкими листьями, преобладанием спорофита, наличием разных спор (микро и мега).
Голосеменные — это первые семенные растения. Семя имеет больше преимуществ, чем спора. Спора — это одна клетка, из которой формируется гаметофит, а семя — многоклеточное. Оно содержит в себе зародыш (маленький спорофит) и запас питательных веществ для его развития. Голосеменные получили такое название потому, что семена у этих растений образуются не в завязи, а из семяпочек, лежащих открыто (голо) на чешуйках шишки. Голосеменные не образуют, плодов. Типичные представители: ель, сосна, пихта, лиственница, тис, кедр и др.
Семя имеет больше преимуществ, чем спора. Спора — это одна клетка, из которой формируется гаметофит, а семя — многоклеточное. Оно содержит в себе зародыш (маленький спорофит) и запас питательных веществ для его развития. Голосеменные получили такое название потому, что семена у этих растений образуются не в завязи, а из семяпочек, лежащих открыто (голо) на чешуйках шишки. Голосеменные не образуют, плодов. Типичные представители: ель, сосна, пихта, лиственница, тис, кедр и др.
Отдел покрытосеменные (цветковые), или магнолиофиты
Наиболее распространенная группа растений, насчитывающая около 300 000 видов, произрастающих по всему земному шару.
Характеризуются сильно развитой корневой системой, крупными, разнообразными листьями, преобладанием в жизненном цикле спорофита. Размножаются семенами, которые развиваются из семяпочки в завязи пестика в цветке, а из завязи образуется плод. Архегониев нет. Происходит двойное оплодотворение. Покрытосеменные занимают господствующее положение в растительном мире. Прежде всего потому, что у них имеется плод, развивающийся из завязи, внутри которого находятся семена. Процессы цветения, опыления, оплодотворения, образования плодов и семян у покрытосеменных — сложные и многообразные явления, которые в ходе длительной эволюции совершенствовались и продолжают совершенствоваться.
Прежде всего потому, что у них имеется плод, развивающийся из завязи, внутри которого находятся семена. Процессы цветения, опыления, оплодотворения, образования плодов и семян у покрытосеменных — сложные и многообразные явления, которые в ходе длительной эволюции совершенствовались и продолжают совершенствоваться.
Конспекты по биологии
Материалы для подготовки к ЕГЭ по биологии в максимально краткой, доступной и наглядной форме с привлечением методик мнемотехники для быстрого и качественного запоминания.Раздел 1. Биология — наука о жизни
1.1. Биология как наука. Роль биологии
1.2. Признаки и свойства живого
1.3. Основные уровни организации живой природы
Раздел 2. Клетка как биологическая система
2.1. Клеточная теория. Развитие знаний о клетке
2.2. Клетка — единица строения, жизнедеятельности, роста и развития организмов. Сравнительная характеристика клеток растений, животных, бактерий, грибов
2.3. Химическая организация клетки
2. 3.1. Неорганические вещества клетки
3.1. Неорганические вещества клетки
2.3.2. Органические вещества клетки: углеводы, липиды
2.3.3. Органические вещества клетки: белки
2.3.4. Органические вещества клетки: нуклеиновые кислоты
2.4. Строение про— и эукариотической клеток
2.5. Метаболизм
2.5.1. Энергетический и пластический обмен
2.5.2. Диссимиляция
2.5.3. Фотосинтез и хемосинтез
2.6. Биосинтез белка и нуклеиновых кислот. Гены, генетический код
2.7. Клетка — генетическая единица живого. Хромосомы. Жизненный цикл клетки. Митоз. Мейоз.
Раздел 3. Организм как биологическая система
3.1. Разнообразие организмов. Вирусы — неклеточные формы.
3.2. Воспроизведение организмов
3.3. Онтогенез
3.4. Генетика. Основные генетические понятия
3.5. Закономерности наследственности
3.6. Изменчивость признаков у организмов
3.7. Вредное влияние мутагенов на генетический аппарат клетки. Наследствениые болезни человека
3.8. Селекция. Значение генетики для селекции.
Значение генетики для селекции.
3.8.1. Генетика и селекция
3.8.2. Методы работы И.В. Мичурина
3.8.3. Центры происхождения культурных растений
3.9. Биотехнология, клеточная и генная инженерия, клонирование
Раздел 4. Многообразие организмов, их строение и жизнедеятельность
4.1. Систематика. Основные систематические (таксономические) категории
4.2. Царство Бактерии.
4.3. Царство Грибы. Лишайники
4.4. Царство Растения.
4.4.1. Общая характеристика царства Растения
4.4.2. Ткани высших растений
4.4.3. Корень
4.4.4. Побег
4.4.5. Цветок и его функции. Соцветия
4.5. Многообразие растений.
4.5.1. Жизненные циклы отделов растений
4.5.2. Однодольные и двудольные растения
4.5.3. Космическая роль растений
4.6. Царство Животные
4.6.1. Общая характеристика царства Животные
4.6.2. Одноклеточные или Простейшие
4.6.3. Тип Кишечнополостные
4.6.4. Тип плоские черви
4. 6.5. Тип Первичнополостные или Круглые черви
6.5. Тип Первичнополостные или Круглые черви
4.6.6. Тип Кольчатые черви стр.1-5 стр. 6-11
4.6.7. Тип Моллюски
4.6.8. Тип Членистоногие стр.1-10 стр.11-25 стр. 26-35
4.7. Хордовые животные
4.7.1. Общая характеристика типа Хордовых
4.7.2. Надкласс Рыбы
4.7.3. Класс Земноводные
4.7.4. Класс Пресмыкающиеся
4.7.5. Класс Птицы
4.7.6.Класс Млекопитающие
Раздел 5. Человек и его здоровье
5.1. Ткани. Строение и жизнедеятельность органов и систем органов: пищеварения, дыхания, кровообращения, лимфатической системы стр.
5.1.1. Анатомия и физиология человека. Ткани стр.1-7 стр.8-20
5.1.2. Строение и функции пищеварительной системы
5.1.3. Строение и функции дыхательной системы
5.1.4. Строение и функции выделительной системы
5.2. Строение и жизнедеятельность органов и систем органов: опорно-двигательной, покровной, кровообращения, лимфообращения. Размножение и развитие человека
5.2. 1. Строение и функции опорно-двигательной системы
1. Строение и функции опорно-двигательной системы
5.2.2. Кожа, ее строение и функции
5.2.3. Строение и функции системы органов кровообращения и лимфообращения
5.2.4. Размножение и развитие организма человека
5.3. Внутренняя среда организма человека. Иммунитет. Обмен веществ и превращение энергии в организме человека
5.3.1. Внутренняя среда организма. Состав и функции крови. Группы крови. Переливание крови. Иммунитет
5.3.2. Обмен веществ в организме человека
5.4. Нервная и эндокринная системы. Нейрогуморальная регуляция процессов жизнедеятельности организма как основа его целостности, связи со средой
5.4.1. Нервная система. Общий план строения. Функции
5.4.2. Строение и функции центральной нервной системы
5.4.3. Строение и функции вегетативной нервной системы
5.4.4. Эндокринная система. Нейрогуморальная регуляция процессов жизнедеятельности
5.5. Анализаторы. Органы чувств. Высшая нервная деятельность
5.5.1 Органы чувств (анализаторы). Строение и функции органов зрения и слуха
Строение и функции органов зрения и слуха
5.5.2. Высшая нервная деятельность
5.6. Личная и общественная гигиена, здоровый образ жизни. Приемы оказания первой помощи.
Раздел 6. Надорганизменные системы. Эволюция органического мира
6.1. Вид, его критерии и структура. Популяция — структурная единица вида и элементарная единица эволюции. Способы видообразования. Микроэволюция
6.2. Развитие эволюционных идей. Движущие силы, элементарные факторы эволюции. Синтетическая теория эволюции
6.2.1. Развитие эволюционных идей. Значение работ К. Линнея, учения Ж.-Б. Ламарка, эволюционной теории Ч. Дарвина. Взаимосвязь движущих сил эволюции. Элементарные факторы эволюции
6.2.2. Творческая роль естественного отбора. Синтетическая теория эволюции. Исследования С.С.Четверикова. Роль эволюционной теории в формировании современной естественнонаучной картины мира
6.3. Результаты эволюции. Доказательства эволюции живой природы.
6.4. Макроэволюция. Направления и пути эволюции. Биологический прогресс и регресс, ароморфоз, идиоадаптация, дегенерация. Причины биологического прогресса и регресса. Гипотезы возникновения жизни на Земле. Эволюция органического мира. Основные ароморфозы в эволюции растений и животных
Биологический прогресс и регресс, ароморфоз, идиоадаптация, дегенерация. Причины биологического прогресса и регресса. Гипотезы возникновения жизни на Земле. Эволюция органического мира. Основные ароморфозы в эволюции растений и животных
6.5. Происхождение человека. Человек как вид, его место в системе органического мира. Гипотезы происхождения человека. Движущие силы и этапы эволюции человека. Человеческие расы, их генетическое родство. Биосоциальная природа человека
Раздел 7. Экосистемы и присущие им закономерности
7.1. Среды обитания организмов. Факторы среды. Законы оптимума и минимума. Биологические ритмы. Фотопериодизм
7.2. Экосистема, ее компоненты, структура. Цепи и сети питания, их звенья. Правило экологической пирамиды. Структура и динамика численности популяций
7.3. Разнообразие, саморазвитие, смена экосистем. Агроэкосистемы, основные отличия от природных экосистем
7.4. Круговорот веществ и превращения энергии в экосистемах. Биологическое разнообразие, саморегуляция и круговорот веществ — основа устойчивого развития экосистем
7. 5—7.6. Биосфера — глобальная экосистема. Учение В.И. Вернадского
5—7.6. Биосфера — глобальная экосистема. Учение В.И. Вернадского
Царство растений реферат по биологии
Царство растений Классификация растений и само определение последнего термина в ходе развития ботаники постоянно пересматривались. В середине прошлого века все растения было принято делить на тайнобрачные (криптогамы) с «невидимыми» половыми органами и явнобрачные (фанерогамы), у которых репродуктивные структуры хорошо заметны. К тайнобрачным относили папоротники, мхи, водоросли и грибы, т.е. организмы, не образующие семян, а к явнобрачным – семенные виды. Сейчас такая система считается слишком грубой и искусственной. В первые десятилетия 20 в. расширение знаний о родственных связях растений привело к выделению четырех их главных групп: Thallophyta (талломные, слоевцовые, или низшие растения), Bryophyta (моховидные), Pteridophyta (папоротниковидные) и Spermatophyta (семенные). Группа низших растений объединяла бактерии, водоросли и грибы – организмы с талломом, или слоевищем, т. е. телом, не расчлененным на корни, стебли и листья. К моховидным были относены печеночники и листостебельные мхи. У них тоже нет настоящих корней, стеблей и листьев, но от низших они отличаются прежде всего тем, что развитие зародыша из оплодотворенной яйцеклетки происходит у них в особом женском органе на растении, а не в окружающей среде. У папоротниковидных, т.е. папоротников, хвощей, плаунов и близких форм, есть не только настоящие корни, стебли и листья, но и особая проводящая (сосудистая) система, состоящая из специализированных сосудов ксилемы и флоэмы. Однако семян эти растения не образуют. Семенные растения, также обладающие настоящими корнями, стеблями, листьями и сосудистыми тканями, делятся на голосеменные (например, хвойные), не образующие цветков, и покрытосеменные (цветковые). Сегодня и эта система признается неудовлетворительной. Некоторые ботаники стремились усовершенствовать ее, разделив все растения на эмбриофиты (Embryophyta), образующие зародыш внутри родительского организма (моховидные, папоротниковидные, семенные), и лишенные такой особенности низшие растения (бактерии, водоросли, грибы).
е. телом, не расчлененным на корни, стебли и листья. К моховидным были относены печеночники и листостебельные мхи. У них тоже нет настоящих корней, стеблей и листьев, но от низших они отличаются прежде всего тем, что развитие зародыша из оплодотворенной яйцеклетки происходит у них в особом женском органе на растении, а не в окружающей среде. У папоротниковидных, т.е. папоротников, хвощей, плаунов и близких форм, есть не только настоящие корни, стебли и листья, но и особая проводящая (сосудистая) система, состоящая из специализированных сосудов ксилемы и флоэмы. Однако семян эти растения не образуют. Семенные растения, также обладающие настоящими корнями, стеблями, листьями и сосудистыми тканями, делятся на голосеменные (например, хвойные), не образующие цветков, и покрытосеменные (цветковые). Сегодня и эта система признается неудовлетворительной. Некоторые ботаники стремились усовершенствовать ее, разделив все растения на эмбриофиты (Embryophyta), образующие зародыш внутри родительского организма (моховидные, папоротниковидные, семенные), и лишенные такой особенности низшие растения (бактерии, водоросли, грибы). Другие ученые предложили трехчленное деление царства растений – на сосудистые (папоротниковидные и семенные), или Tracheophyta, низшие (бактерии, водоросли, грибы) и моховидные (мхи и печеночники). Однако и эти системы признаны несостоятельными. В настоящее время получила признание система многих царств. Бактерии, водоросли и грибы растениями (т.е. представителями царства растений) больше не признаются. Первых (вместе с сине-зелеными водорослями, которых теперь называют цианобактериями) либо выделяют в одно царство Monera, либо распределяют между царствами Archaebacteria и Eubacteria. Прочие водоросли, включая одноклеточные, а также ряд относительно близких к ним по строению организмов, составляют царство протистов (Protista). Особое царство Fungi образуют грибы. Хотя по ряду признаков они и напоминают растения, многие их типичные представители, как показывает изучение ДНК, вероятно, ближе к животным. То, что в результате остается от царства растений (Plantae), разные специалисты разделяют на 5–18 крупных групп.
Другие ученые предложили трехчленное деление царства растений – на сосудистые (папоротниковидные и семенные), или Tracheophyta, низшие (бактерии, водоросли, грибы) и моховидные (мхи и печеночники). Однако и эти системы признаны несостоятельными. В настоящее время получила признание система многих царств. Бактерии, водоросли и грибы растениями (т.е. представителями царства растений) больше не признаются. Первых (вместе с сине-зелеными водорослями, которых теперь называют цианобактериями) либо выделяют в одно царство Monera, либо распределяют между царствами Archaebacteria и Eubacteria. Прочие водоросли, включая одноклеточные, а также ряд относительно близких к ним по строению организмов, составляют царство протистов (Protista). Особое царство Fungi образуют грибы. Хотя по ряду признаков они и напоминают растения, многие их типичные представители, как показывает изучение ДНК, вероятно, ближе к животным. То, что в результате остается от царства растений (Plantae), разные специалисты разделяют на 5–18 крупных групп. Увеличение их числа отчасти обусловлено вполне правомерным отказом от признания сосудистых растений (Tracheophyta) естественным таксоном: теперь каждая их группа считается самостоятельным отделом. В современной системе отдел – второй по иерархическому рангу таксон после царства (у животных и протистов ему соответствует «тип»). Ранее считалось, что многие из отделов более или менее линейно происходят один от другого и соответствуют последовательным этапам эволюции, но сейчас их рассматривают просто как группы растений, сходных по строению и особенностям размножения форм, не обязательно происходящих от одного предкового вида другого отдела. Таким образом, хотя этот таксономический ранг и отражает существенные различия между организмами, прежнего эволюционного содержания ему уже не приписывают. Перечисленные ниже типы царства протистов традиционно относят к растениям.
Увеличение их числа отчасти обусловлено вполне правомерным отказом от признания сосудистых растений (Tracheophyta) естественным таксоном: теперь каждая их группа считается самостоятельным отделом. В современной системе отдел – второй по иерархическому рангу таксон после царства (у животных и протистов ему соответствует «тип»). Ранее считалось, что многие из отделов более или менее линейно происходят один от другого и соответствуют последовательным этапам эволюции, но сейчас их рассматривают просто как группы растений, сходных по строению и особенностям размножения форм, не обязательно происходящих от одного предкового вида другого отдела. Таким образом, хотя этот таксономический ранг и отражает существенные различия между организмами, прежнего эволюционного содержания ему уже не приписывают. Перечисленные ниже типы царства протистов традиционно относят к растениям.
«Полезные пожиратели». Что будет с нами, если все вирусы исчезнут?
- Рейчел Нюэр
- BBC Future
Автор фото, Science Photo Library
Подпись к фото,Вот так выглядел вирус испанского гриппа, в 1918 году унесшего жизни от 50 до 100 млн человек (по разным оценкам)
Если бы все вирусы вдруг исчезли, мир стал бы совершенно другим — и не факт, что лучше. Что же было бы с нами без вирусов? И что значит «убить победителя»?
Что же было бы с нами без вирусов? И что значит «убить победителя»?
Глядя на пугающие картины пандемии Covid-19, разворачивающиеся, благодаря СМИ и соцсетям, перед глазами всего мира, можно подумать, что вирусы только для того и существуют, чтобы поставить человечество на колени и уморить как можно больше людей.
За прошедшее тысячелетие болезни, ими порождаемые, унесли бесчисленное количество жизней. Некоторые из вирусов убивали значительную часть населения планеты: жертвами эпидемии испанского гриппа в 1918 году стало, по разным оценкам, от 50 до 100 млн человек, еще 200 млн, как считается, умерли от оспы только в XX веке.
И нынешняя пандемия Covid-19 — лишь очередной случай из бесконечной серии нападений смертельных вирусов на человечество.
Большинство из нас сейчас, если бы нам вручили волшебную палочку и предложили ею взмахнуть, чтобы избавиться от всех вирусов на планете, с радостью согласилось бы.
Боюсь, это было бы смертельной ошибкой. Фактически, куда более смертельной, чем любой из самых свирепых вирусов.
«Если бы все вирусы вдруг разом исчезли, мир стал бы прекрасен — примерно на день-полтора. А потом мы бы все умерли, вот и всё, — говорит Тони Голдберг, эпидемиолог из Университета Висконсин-Мэдисон. — Те важнейшие вещи, за которые отвечают вирусы, значительно перевешивают зло от них».
В общем, как говорит Сусана Лопес Шаритон, вирусолог из Национального автономного университета Мексики, «без вирусов нам конец».
Автор фото, Getty Images
Подпись к фото,Некоторые вирусы сберегают здоровье грибам и растениям
Большинство людей даже не догадывается о том, какую роль играют вирусы в жизни на Земле, обращая внимание только на те из них, которые нас убивают.
Почти все вирусологи изучают исключительно патогены, и только недавно несколько ученых решились исследовать вирусы, благодаря которым живы мы и наша планета.
Благодаря этой маленькой группе исследователей мы, возможно, получим более сбалансированный взгляд на мир вирусов. Оказывается, есть среди них и хорошие, причем таких — подавляющее большинство.
Но одно ученые точно знают уже сейчас: без вирусов наша планета, какой мы ее знаем, перестала бы существовать. Да и если бы мы даже задались целью истребить все вирусы на Земле, это практически невозможно.
Но представив, каким был бы мир без вирусов, мы сможем лучше понять, насколько они важны для нашего выживания, и как много нам еще предстоит узнать об этих микроскопических, простейших формах жизни, с которыми всё непросто.
Автор фото, Getty Images
Подпись к фото,Без вирусов наша планета перестала бы существовать
Для начала скажем, что ученым даже неизвестно, сколько всего вирусов существует. Официально классифицированы тысячи, но их — миллионы.
«Нами открыта лишь малая часть, поскольку мы особо не интересовались этим, — говорит Мэрилин Руссинк, вирусный эколог из Университета Пенн Стейт. — Таково предвзятое отношение: науку всегда прежде всего интересовали патогены».
— Таково предвзятое отношение: науку всегда прежде всего интересовали патогены».
Неизвестно ученым и то, какой именно процент всех вирусов опасен для человека. «Если смотреть на большие числа, то статистически процент опасных вирусов приближается к нулю, — говорит Кертис Саттл, вирусолог-эколог из Университета Британской Колумбии. — Почти все существующие вирусы не болезнетворны для нас».
Полезные пожиратели
По крайней мере, нам известно, что фаги (бактериофаги, вирусы, избирательно поражающие бактериальные клетки) — невероятно важны. Их название происходит от греческого «пожираю», и именно этим они и занимаются.
«В мире бактерий они — самые главные хищники, — говорит Голдберг. — Без них нам пришлось бы туго».
Фаги — главный регулятор популяций бактерий в океане, да и, скорее всего, во всех остальных экосистемах нашей планеты. Если бы вирусы вдруг исчезли, некоторые популяции, вероятно, разрослись взрывным образом и подавили бы другие, которые совсем перестали бы расти.
Для океана это стало бы особенно серьезной проблемой, поскольку в нем более 90% всего живого (от общей массы) — микроорганизмы. И эти микробы производят около половины всего кислорода на планете — процесс, который становится возможным, благодаря вирусам.
Автор фото, Getty Images
Подпись к фото,В океане 90% всего живого — микроорганизмы
Эти вирусы каждый день уничтожают примерно 20% всех океанических микробов и около 50% всех океанических бактерий. Этим они обеспечивают достаточно питательных веществ для производящего кислород планктона и тем самым поддерживают жизнь на планете.
«Когда нет смерти, тогда нет и жизни, потому что жизнь полностью зависит от рециркуляции материалов, — подчеркивает Саттл. — Вирусы очень важны для такой утилизации».
Исследователи, изучающие насекомых-вредителей, также обнаружили, что вирусы критически важны для контроля над численностью популяции.
Если некоторые виды начинают слишком разрастаться, «приходит вирус и уничтожает их», говорит Руссинк. Это очень естественный процесс для экосистем.
Это очень естественный процесс для экосистем.
Процесс этот называется «убить победителя» и весьма распространен у многих других видов, в том числе и нашего — пандемии тому доказательство.
«Когда популяция становится чересчур многочисленной, вирусы воспроизводятся необыкновенно быстро и снижают ее объем, освобождая пространство для жизни всего остального», — подчеркивает Саттл.
Если все вирусы вдруг исчезнут, самые конкурентоспособные виды разрастутся в ущерб всем остальным.
«Мы быстро потеряем значительную часть биоразнообразия нашей планеты, — говорит Саттл. — Всё захватят несколько видов, остальные вымрут».
Автор фото, Getty Images
Подпись к фото,По словам экспертов, без вирусов наша планета утратила бы значительную часть биологического разнообразия
Для некоторых организмов вирусы критически важны для выживания или для того, чтобы получить конкурентоспособное преимущество.
Например, ученые предполагают, что вирусы играют важную роль, помогая коровам и другим жвачным животным превращать целлюлозу из травы в сахара, которые метаболизируются и в итоге превращаются в молоко, а также помогают набрать массу тела.
Исследователи считают, что вирусы важны и для поддержания здорового микробиома в организме человека и животных.
«Эти вещи пока еще не до конца исследованы, но мы находим все больше и больше примеров такого тесного взаимодействия с вирусами как важнейшего элемента экосистем», — говорит Саттл.
Руссинк и ее коллеги обнаружили твердое доказательство этому. В одном из исследований они работали с колонией микроскопических грибов, которая сожительствует с определенным видом трав в Йеллоустонском национальном парке (биосферный заповедник в США, знаменитый своим геотермальным ландшафтом и гейзерами — прим. Би-би-си), и обнаружили: вирус, заразивший гриб, позволяет траве более успешно выдерживать геотермальные температуры почвы.
«Когда присутствуют все три элемента — вирус, гриб и трава, тогда травы могут расти на горячей почве, — рассказывает Руссинк. — Один гриб без вируса не способен сделать такое».
Автор фото, Getty Images
Подпись к фото,В Йеллоустонском национальном парке некоторые виды травы стали более устойчивы к высоким температурам — благодаря вирусу
Руссинк и ее коллеги обнаружили, что грибы обычно передают вирусы «по наследству» — из поколения в поколение. И хотя ученым еще не удалось выяснить функцию большинства из этих вирусов, можно заключить, что они чем-то помогают грибам.
И хотя ученым еще не удалось выяснить функцию большинства из этих вирусов, можно заключить, что они чем-то помогают грибам.
«Иначе зачем растениям за них цепляться?» — рассуждает Руссинк.
И если все эти полезные вирусы исчезнут, то травы и другие организмы, в которых они сейчас живут, ослабнут, а возможно и погибнут.
Под защитой вирусов
Инфицирование человеческого организма определенными безвредными вирусами даже помогает отпугивать некоторые патогены.
Вирус GB типа C, распространенный человеческий непатогенный (в отличие от своих дальних родственников вируса Западного Нила и вируса лихорадки денге) увязывается с замедлением развития СПИДа у ВИЧ-инфицированных.
Примерно так же и герпес делает мышей менее подверженными определенным бактериальным инфекциям, в том числе бубонной чумы и листериоза (распространенного типа пищевого отравления).
Конечно, проводить на людях похожие эксперименты с заражением вирусами герпеса, бубонной чумы и листериоза неэтично, авторы исследования предполагают, что и у людей была бы похожая картина.
Автор фото, Science Photo Library
Подпись к фото,Вирус герпеса делает мышей — и, очень возможно, людей — менее подверженными некоторым бактериальным инфекциям
Похоже, что без вирусов и люди, и многие другие виды живых существ были бы более подвержены разным болезням.
Кроме того, вирусы — это одно из самых многообещающих лечебных средств от определенных заболеваний. Фаготерапия (лечение инфекционных больных и бактерионосителей препаратами бактериофага), которую в Советском Союзе начали применять еще с 1920-х годов, использует вирусы для уничтожения бактериальных инфекций.
Сегодня это — быстроразвивающаяся область научного поиска. Не только из-за растущей устойчивости патогенов к антибиотикам, но и потому, что бактериофаги можно точно настраивать на воздействие на определенные виды бактерий — в отличие от антибиотиков, уничтожающих все бактерии без разбора.
«Когда антибиотики ничем не могут помочь, жизни людей спасают вирусы», — подчеркивает Саттл.
Онколитическая вирусная терапия рака, при которой заражаются и уничтожаются исключительно раковые клетки, к тому же менее токсична и более эффективна, чем другие методы лечения онкологии.
Нацеленные на уничтожение вредоносных бактерий или на раковые клетки, терапевтические вирусы действуют как «микроскопические крылатые ракеты, наводящиеся и попадающие точно в цель», отмечает Голдберг.
«Нам нужны такие вирусы, которые выведут нас на новую ступень терапии, терапию нового поколения».
Поскольку вирусы постоянно мутируют и реплицируются (размножаются), они представляют собой огромное хранилище генетических инноваций, которые могут быть использованы другими организмами.
Вирусы внедряются в клетки других существ и захватывают их инструменты размножения.
Если такое случается в клетке зародышевой линии (яйцеклетки и спермы), код вируса может передаваться из поколения в поколение и стать ее постоянной частью.
«Все организмы, которые могут быть заражены вирусами, имеют возможность принять вирусные гены и использовать их в своих интересах, — отмечает Голдберг. — Включение нового ДНК в геном — это основной способ эволюции».
Другими словами, исчезновение всех вирусов отразится на эволюционном потенциале всей жизни на нашей планете. В том числе и homo sapiens.
Вирусные элементы составляют около 8% человеческого генома, а геномы млекопитающих в целом приправлены примерно 100 000 остатками генов, когда-то принадлежавших вирусам.
Код вирусов — это часто неактивная часть ДНК, но иногда он наделяет организм новыми, полезными и даже важными свойствами.
Например, в 2018 году два коллектива исследователей независимо друг от друга сделали удивительное открытие. Ген вирусного происхождения кодирует белок, играющий ключевую роль в формировании долговременной памяти, передавая информацию между клетками нервной системы.
Автор фото, Getty Images
Подпись к фото,Именно древние ретровирусы ответственны за то, что люди способны к живорождению
Есть доказательства того, что мы обязаны своей способностью к живорождению частичке генетического кода, взятой у древних ретровирусов, которыми наши дальние предки заразились более 130 млн лет назад.
Вот что писали авторы того открытия в 2018 году в журнале PLOS Biology: «Очень соблазнительно поспекулировать на тему того, что беременность у людей могла бы протекать совершенно иначе (а то и не существовала бы вообще), если бы наших предков в процессе эволюции не затронули бы многие эпохи ретровирусных пандемий».
Специалисты считают, что такие частички генетического кода можно встретить у всех форм многоклеточной жизни. «Вероятно, они несут множество функций, о которых нам ничего не известно», — подчеркивает Саттл.
Ученые только-только начали открывать способы, с помощью которых вирусы помогают поддерживать жизнь. В конечном счете, чем больше мы узнаем о всех вирусах (не только о патогенах, возбудителях болезней), тем лучше мы будем оснащены для того, чтобы использовать определенные вирусы в мирных целях и разработать эффективную защиту от других вирусов, которые могут привести к очередной пандемии.
Более того: изучение богатого вирусного многообразия поможет нам более глубоко понять, как работает наша планета, ее экосистемы и организмы.
По словам Саттла, «нам нужно приложить некоторые усилия, чтобы понять, что происходит и что нас ждет — для нашей же пользы».
Больше статей на подобные темы — на сайте BBC Future.
Введение в биологию растений
Обзор
В биологии растений растения по большей части являются фотосинтезирующими эукариотами, составляющими царство Plantae. В соответствующих средах/средах обитания растения используют воду, световую энергию и углекислый газ (это неорганические вещества) для синтеза сахара/питательных веществ, которые служат пищей для различных животных.
Помимо синтеза собственной пищи, растения в различных средах (водной, наземной и т.д.) играют важную роль в производстве кислорода, что делает их необходимыми для всей жизни на Земле.Хотя биология растений обеспечивает возможность фотосинтеза (объединяющая характеристика), разные типы растений обладают различными физическими и генетическими характеристиками.
Как таковые, они делятся на отдельные группы, которые включают:
· Покрытосеменные — Растения, которые производят цветы и приносят плоды (более 200 000 видов растений относятся к этой категории).
· Голосеменные — Включая семенные сосудистые растения (эти растения не цветут и не приносят плодов).
· Несосудистые растения — Включают печеночники, мхи и роголистники. Большинство несосудистых растений имеют небольшие размеры и характеризуются плохой транспортной системой
· Папоротники и плауновидные — Характеризуются организмами, имеющими сосудистую систему, но не дающими семян и цветков и не древесными по своей природе. .
Новые открытия в биологии растений
Совсем недавно исследования в области биологии растений показали, что растения могут испытывать различные вредные раздражители и «предупреждать/делиться» об этом с другими растениями. На основании микроскопических исследований было обнаружено, что кальций играет важную роль в этом обмене информацией. ..
На основании микроскопических исследований было обнаружено, что кальций играет важную роль в этом обмене информацией. ..
В период с 2016 по 2017 год было обнаружено до 2000 новых видов растений. На основании различных исследований был сделан вывод, что эти растения можно использовать в пищу, в медицине, а также в качестве источника древесины.
Общая анатомия растений
Что касается анатомии/строения растений, то существуют некоторые одноклеточные организмы, которые имеют сходные с растениями характеристики фотосинтеза (например, некоторые зеленые водоросли). Эти организмы различаются по размеру, общей форме/внешнему виду и типам органелл, содержащихся в клеточной стенке.
Однако для большинства растений общая анатомия может состоять из следующих частей в зависимости от растения:
* Корни, стебель и листья известны как вегетативные части растений.
Кроме трех основных частей биологии растений, их анатомия также может включать в себя:
- цветы
- Стел / ветви
- Фрукты
- BUDS
Каждая часть анатомии завода участвует в заданных функциях . В то время как листья содержат хлорофилл и хлоропласты, участвующие в фотосинтезе, стебель обеспечивает поддержку системы побегов, в то время как корни закрепляют все растение и поглощают воду и минералы, необходимые для роста и развития растения.
В то время как листья содержат хлорофилл и хлоропласты, участвующие в фотосинтезе, стебель обеспечивает поддержку системы побегов, в то время как корни закрепляют все растение и поглощают воду и минералы, необходимые для роста и развития растения.
Хотя упомянутые части анатомии растений можно найти у большинства растений, у некоторых может отсутствовать одна или несколько из этих частей в зависимости от их экологии. Эти аспекты растения также могут варьироваться в зависимости от того, где растения находятся.
Например, листья растений, произрастающих в водной среде, имеют минимальную кутикулу и множество пор, позволяющих воде выходить; растения, произрастающие в засушливых/полузасушливых условиях, имеют более толстую кутикулу и меньшее количество пор, что предотвращает потерю растением лишней воды.Как запасающие воду листья, эти листья также могут быть более сочными по сравнению с листьями водных растений.
* Морфология растений отражает экологическую адаптацию.
Внутренние структуры растительной клетки
Структура биологии растительного клеток на основе их ткани:
- 9001 — состоит из ксилома и флоэмы
- Бесполое размножение
- Вегетативное размножение
- Половое размножение
- Луковицы
- Rhizomes
- Rrinks
- Corms
- BRYOPHYLLUM
- Чашечка, состоящая из чашелистиков
- Венчик, содержащий лепестки
- Андроцей, состоящий из тычинок (половых органов мужского пола)
- Pispel органы самки)
- Растения автотрофы . Они производят себе пищу посредством фотосинтеза . Они способны улавливать энергию через зеленый пигмент (хлорофилл) внутри хлоропласта и использовать углекислый газ и воду для производства сахара в качестве пищи и кислорода в качестве побочного продукта.Как автотрофы, растения часто находятся в начале пищевой цепи. Они помечены как производителей . Они служат пищей другим организмам, в том числе животным. Животные, напротив, являются гетеротрофами, и им необходимо потреблять другие организмы для пропитания. Некоторые животные (в частности, травоядных ) зависят исключительно от растений, в то время как другие питаются только мясом или смесью животного или растительного материала. Поскольку растения способны производить себе пищу, они не питаются животными, чтобы расти и выживать.Исключением является группа плотоядных растений ( например, Венерина мухоловка), которые ловят и питаются животными, особенно когда условия менее благоприятны для фотосинтеза.
- Растения эукариоты . Как и у животных, у растений внутри клетки имеется отчетливое, окруженное мембраной ядро. Ядро представляет собой органеллу, содержащую хромосомы, несущие гены. Другими органеллами, взвешенными в цитоплазме растительной клетки, являются аппарат Гольджи, эндоплазматический ретикулум, лизосомы, пероксисомы и пластиды.
- Растения имеют пластиды . Наличие пластид внутри эукариотической клетки свидетельствует о том, что это скорее растение, чем животное. Пластиды бывают разные. Хлоропласты представляют собой пластиды, содержащие хлорофилл ( зеленых пигментов ) и участвующие в фотосинтезе. Хромопласты содержат пигменты помимо зеленого и участвуют в синтезе и хранении пигментов. Системы хлорофилла поглощают световую энергию на определенных длинах волн электромагнитного спектра.Пигменты также отвечают за окраску растительных структур ( например, зеленые листья, красные цветы, желтые плоды).
 Лейкопласты ( например, амилопласты, элайопласты, протеинопласты) представляют собой непигментированных пластид. Их функция в первую очередь для хранения продуктов. Растения запасают пищу в виде сахара, т.е. крахмал.
Лейкопласты ( например, амилопласты, элайопласты, протеинопласты) представляют собой непигментированных пластид. Их функция в первую очередь для хранения продуктов. Растения запасают пищу в виде сахара, т.е. крахмал. - Растения имеют большую вакуоль внутри клетки. Эта цитоплазматическая структура участвует в регуляции тургорного давления.
- Растения имеют жесткие клеточные стенки кроме плазматической мембраны. Клеточная стенка обеспечивает дополнительную структурную поддержку растительной клетке. У растений может не быть скелетной системы, как у животных, но их клеточная стенка состоит в основном из целлюлозного материала, который обеспечивает структурную поддержку.
- Растения имеют характерное клеточное деление, при котором клеточная пластинка (фрагмопласт) разделяет дочерних клеток.
- Растения не так подвижны, как животные.Они не имеют возможности перемещаться из одного места в другое по своему желанию.
 Таким образом, им приходится иметь дело с суровыми условиями, такими как жара. Один из способов, которым они способны противостоять жаре, — это стенки их клеток, которые предотвращают высыхание их тела. Несмотря на это, растения все же демонстрируют движение, но в другой форме. Например, настическое движение иллюстрируется складыванием листочков растения Mimosa pudica при прикосновении и закрыванием листа венериной мухоловки при захвате добычи.Некоторые растения ( например, Betula pendula – береза повислая) даже опускают свои ветки и листья ночью, как будто они «спят» . Другой формой передвижения растений является тропизм. Однако тропизм — это скорее реакция роста на стимул, чем на движение. Например, растения имеют тенденцию расти по направлению к источнику света (фототропизм).
Таким образом, им приходится иметь дело с суровыми условиями, такими как жара. Один из способов, которым они способны противостоять жаре, — это стенки их клеток, которые предотвращают высыхание их тела. Несмотря на это, растения все же демонстрируют движение, но в другой форме. Например, настическое движение иллюстрируется складыванием листочков растения Mimosa pudica при прикосновении и закрыванием листа венериной мухоловки при захвате добычи.Некоторые растения ( например, Betula pendula – береза повислая) даже опускают свои ветки и листья ночью, как будто они «спят» . Другой формой передвижения растений является тропизм. Однако тропизм — это скорее реакция роста на стимул, чем на движение. Например, растения имеют тенденцию расти по направлению к источнику света (фототропизм). - Растения имеют плазмодесмы . В то время как у животных есть клеточные соединения, которые удерживают клетки в животной ткани, у растений есть плазмодесмы, которые действуют как клеточные соединения между растительными клетками.
 Клеточная стенка образует эти цитоплазматические мостики между соседними клетками. Эти «мостики» облегчают связь между клетками и обеспечивают циркуляцию жидкости, тем самым помогая поддерживать тонус растительных клеток.
Клеточная стенка образует эти цитоплазматические мостики между соседними клетками. Эти «мостики» облегчают связь между клетками и обеспечивают циркуляцию жидкости, тем самым помогая поддерживать тонус растительных клеток. - Растения являются многоклеточными , состоящими из множества клеток, организованных в ткани и органы, которые выполняют определенную функцию как единое целое. Органы растений предназначены для закрепления, поддержки и фотосинтеза ( например, корни, стебли, листья, и т. д. )
- Растения способны к неограниченному росту за счет меристематических тканей. Ткань состоит из неопределенных, активно делящихся клеток, которые дают начало дифференцированным тканям, таким как эпидермис, трихомы, феллема и сосудистые ткани.
- Растения не имеют органов чувств , но они могут воспринимать окружающую среду, хотя и по-разному. Растения могут «видеть», «слышать» и «нюхать», несмотря на отсутствие у них глаз, ушей и носа.
 Кажется, что они «чувствуют» и реагируют не так явно, как животные.У растений может не быть нервной системы, как у животных, но у них, по-видимому, есть собственная система, основанная на том, как они реагируют на окружающую среду. Например, Arabidopsis , несмотря на отсутствие глаз, обладает фоторецепторами (по крайней мере, 11 типов), которые помогают растению обнаруживать свет. 1 В другом примере травоядные могут спровоцировать выброс определенных химических веществ на пораженную часть растения. 2 Также было замечено, что растения выделяют защитные химические вещества, отпугивающие травоядных.Было замечено, что помидоры испускают летучие сигналы, чтобы предупредить близлежащие растения о надвигающемся нападении травоядных. 3
Кажется, что они «чувствуют» и реагируют не так явно, как животные.У растений может не быть нервной системы, как у животных, но у них, по-видимому, есть собственная система, основанная на том, как они реагируют на окружающую среду. Например, Arabidopsis , несмотря на отсутствие глаз, обладает фоторецепторами (по крайней мере, 11 типов), которые помогают растению обнаруживать свет. 1 В другом примере травоядные могут спровоцировать выброс определенных химических веществ на пораженную часть растения. 2 Также было замечено, что растения выделяют защитные химические вещества, отпугивающие травоядных.Было замечено, что помидоры испускают летучие сигналы, чтобы предупредить близлежащие растения о надвигающемся нападении травоядных. 3 - Растения размножаются бесполым и половым способами. Бесполое размножение у растений осуществляется путем почкования, фрагментации, деления, спорообразования, вегетативного размножения, апомиксиса, и т.
 д. При половом размножении участвуют мужские и женские гаметы, которые сливаются при оплодотворении. В целом жизненный цикл растений включает смену поколений, т.е.е. чередование фаз спорофита и гаметофита.
д. При половом размножении участвуют мужские и женские гаметы, которые сливаются при оплодотворении. В целом жизненный цикл растений включает смену поколений, т.е.е. чередование фаз спорофита и гаметофита. - Растения «дышат» . Через устьица в растительную клетку поступает углекислый газ из атмосферы. В процессе фотосинтеза углекислый газ превращается в кислород, который растение выделяет как побочный продукт метаболизма в атмосферу через устьица.
- Растения могут не иметь других четко определенных биологических систем, но они производят химические вещества, участвующие в защитных и иммунных функциях растений, а также растительные гормоны, которые действуют как сигнальные молекулы.
- Эмбриональные или меристематические ткани – растительные ткани, состоящие из недифференцированных и митотически активных клеток. Примерами являются апикальная меристема и камбий
- Постоянные ткани – ткани растений, состоящие из дифференцированных клеток. Постоянные ткани можно дополнительно классифицировать на основные (, например, паренхима, колленхима, склеренхима) и сложные (, например, ткани флоэмы и ксилемы)
- Репродуктивные ткани – ткани растений, участвующие в репродукции.Примером являются спорогенные ткани
- мутуализм – напр. растения, дающие нектар пчелам, в то время как пчелы помогают распространять пыльцу растений
- хищничество – e.г. плотоядные растения, ловящие насекомых и мелких животных
- конкуренция – напр. растения, которые конкурируют с другими растениями за среду обитания за доступное пространство и питательные вещества
- комменсализм – напр. плоды растений, которые прикрепляются к шерсти животных для свободного транспорта
- паразитизм – напр. паразитические растения, получающие питательные вещества от своего хозяина, такие как Cuscuta (повилика), которая прикрепляется к дереву акации и производит гаустории, которые поглощают питательные вещества из дерева акации
- Фанерозойский эон » Палеозойская эра » Ордовикский период появились растения).

- Фанерозойский эон » Палеозойская эра » Девонский период : В девонский период ( 415 миллионов лет до 360 миллионов лет назад ) примитивные растения, деревья и кустарниковые леса доминировали на суше и предоставили новые места обитания наземным животным . Ранний семенной папоротник Elkinsia дал семена, особенно в поздний девонский период.
- Фанерозойский эон » Мезозойская эра : Эта эра длилась с 252 миллионов до 66 миллионов лет назад .В триасе (примерно 200 млн лет назад ) появляются цветковые растения.
- Фанерозойский эон » Кайнозойская эра : Эта эра, называемая «новой жизнью», является самой последней геологической эрой, которая охватывает период с 66 миллионов лет назад до наших дней . В эту эпоху, примерно 90 526 40 миллионов лет назад, 90 527 года, появились травы. Эти растения и многие другие группы растений развили новый механизм метаболизма, чтобы выжить в условиях низкого содержания CO 2 и засушливых условиях тропиков.

- хлорофитами
- Charophyta
- печёночные мхи (печеночники)
- Anthocerotophyta (роголистники)
- мохообразные (мхи)
- плауновидные (плауны)
- Pteridophyta (папоротники, ветряные папоротники и хвосты)
- CycadoPhyta (Cycads)
- Гинкгофита (Гинкго)
- Pinophyta (Confers)
- GNETOPHYTA (GNETOPHYTES)
- GNETOPHYTA (GNETOPHYTES)
- Magnoliophyta (цветущие растения)
- Палеоботаника – изучение ископаемых растений
- Альгология – изучение водорослей
- Микология – изучение грибов
- Бриология – изучение мхов, печеночников и роголистников
- Птеридология – изучение папоротников
- Палинология – изучение пыльцевых зерен и спор
- Домен: EUKARYOTA
- (рассеивается): Archaeplastida
- Королевство: Plantae Copeland, 1956
- Gabbatiss, J. (2017, 1 января). Растения могут видеть, слышать и обонять – и реагировать.Получено с http://www.bbc.com/earth/story/20170109-plants-can-see-hear-and-smell-and-respond
- Растение подает сигнал «SOS» для защиты растений, когда ему причиняют вред — Биология Блог и словарь онлайн. (2018, 22 сентября). Получено с https://www.biologyonline.com/plant-sets-off-sos-for-plant-defense
- Mescher, MC, & De Moraes, CM (2014). Передайте боеприпасы. Природа , 510 (7504), 221–222. https://doi.org/10.1038/510221a
- Бренчли, Р., Spannagl, M., Pfeifer, M., Barker, G.L.A., D’Amore, R., Allen, A.M., Hall, N., et al. . (2012). Анализ генома мягкой пшеницы с использованием полногеномного секвенирования дробовика. Природа , 491 (7426), 705–710. https://doi.org/10.1038/nature11650
- Находящиеся в критическом состоянии (CR). (2016). Получено с http://cmsdocs.s3.amazonaws.
 com/summarystats/2016-2_Summary_Stats_Page_Documents/2016_2_RL_Stats_Table_2.pdf
com/summarystats/2016-2_Summary_Stats_Page_Documents/2016_2_RL_Stats_Table_2.pdf - Перепись морской жизни. (2011, 24 августа).Сколько видов на Земле? По новой оценке, около 8,7 млн. человек. ScienceDaily . Получено с http://www.sciencedaily.com/releases/2011/08/110823180459.htm
- CHEM 101DL Основные понятия химии ИЛИ
- CHEM 110DL Награды за химию: основные понятия в контексте ИЛИ
- CHEM 21 Общая химия Кредит
- CHEM 101DL Основные понятия химии ИЛИ
- CHEM 110DL Награды за химию: основные понятия в контексте ИЛИ
- CHEM 21 Общая химия Кредит
- CHEM201DL Органическая химия
- MATH 111L Лабораторный расчет I ИЛИ
- МАТЕМАТИКА 121 Введение в исчисление I ИЛИ
- МАТЕМАТИКА 21 Вводное исчисление I ИЛИ
- ОБА MATH 105L/106L Лабораторные расчеты и функции I и II
- ОБА MATH 105L/106L Лабораторные расчеты и функции I и II ИЛИ
- MATH 111L Лабораторный анализ I (ИЛИ Math 21 или 112L или 122) ИЛИ
- STA 102 Вводная биостатистика ИЛИ
- БИОЛОГИЯ 304 (204) Анализ биологических данных (не может быть засчитан дважды как факультативный) ИЛИ
- STA 101 или выше Анализ данных/статистический вывод
- MATH 112L Лабораторный расчет II ИЛИ
- МАТЕМАТИКА 122 Введение в исчисление II ИЛИ
- МАТЕМАТИКА 22 Введение в исчисление II ИЛИ
- STA 102 Биостатистика ИЛИ STA 101 или выше ИЛИ
- БИОЛОГИЯ 304 Анализ биологических данных (не может также считаться факультативным)
- ФИЗИКА 141L Общая физика I ИЛИ
- ФИЗИКА 151L Введение в механику ИЛИ
- ФИЗИКА 161L Введение в экспериментальную физику I ИЛИ
- ФИЗИКА 25 (кредит AP)
- БИОЛОГИЯ 201L Молекулярная биология (необходимое условие: Chem 101D или эквивалент)
- БИОЛОГИЯ 202L Генетика и эволюция
- БИОЛОГИЯ 203L Молекулярная биология, генетика и эволюция (необходимые условия: Биология AP 5 и Chem 101D или эквивалент)
- 1 курс «Разнообразие организмов», выбранный из следующего:
- БИОЛОГИЯ 344S Разнообразие растений: полевой подход
- БИОЛОГИЯ 341L Растительные сообщества Северной Каролины
- БИОЛОГИЯ 342L Систематика и эволюция растений
- БИОЛОГИЯ 347L Растения и люди
- 1 курс — Биологическая структура и функции
- 1 курс — Экология
- 3 курса, выбранных из следующих (нельзя удвоить курс, взятый для соблюдения вышеуказанного требования разнообразия организмов):
- БИОЛОГИЯ 207 Организм Эволюция
- БИОЛОГИЯ 228 Пища и топливо для растущего населения: основы роста и производства растений
- БИОЛОГИЯ 262 Люди, растения и загрязнение: введение в городскую среду
- БИОЛОГИЯ 280LA Основы тропической биологии
- БИОЛОГИЯ 284A Экосистемы и разнообразие Южной Африки
- БИОЛОГИЯ 287A Биоразнообразие Аляски
- БИОЛОГИЯ 288A Биогеография в австралийском контексте
- БИОЛОГИЯ 341L Растительные сообщества Северной Каролины
- БИОЛОГИЯ 342L Систематика и эволюция растений
- БИОЛОГИЯ 344S Разнообразие растений: полевой подход
- БИОЛОГИЯ 347L Растения и люди
- БИОЛОГИЯ 361LS Полевая экология
- БИОЛОГИЯ 490S Семинар по специальным темам (с одобрения регионального консультанта)
- БИОЛОГИЯ 540L Микология
- БИОЛОГИЯ 547L Энтомология
- БИОЛОГИЯ 556L Систематическая биология
- БИОЛОГИЯ 557L Микробная экология и эволюция
- БИОЛОГИЯ 566S Литература по экофизиологии растений
- БИОЛОГИЯ 588S Макроэволюция
- БИОЛОГИЯ 293 / 493 Исследование Независимое исследование, проводимое с преподавателем в области концентрации или с одобрения консультанта области концентрации
- 1 курс, указанный или включенный в перекрестный список по биологии для уровня 200 или выше (или утвержденный альтернативный курс по выбору) при использовании Bio 201L/202L ИЛИ 2 курса, указанный или включенный в список для биологии уровня 200 или выше ( или утвержденный альтернативный курс по выбору) при использовании Bio 203L
· Дермальная ткань — состоит из различных клеток на внешнем поверхности растения, включая эпидермальные клетки, замыкающие клетки, а также волосковые клетки, расположенные на листьях стебля растения.
·
· Наземная ткань — состоит из трех типов клеток, которые включают в себя:
o Паренхимы клетки — участвующие в хранении и фотосинтез
o клетки главня — клетки, которые обеспечивают поддержку
o склерехима клетки — В основном мертвые клетки, поддерживающие данную ткань
Репродукция
Для растений способ размножения зависит от типа растения или условий окружающей среды. Одни растения размножаются путем клеточного деления родительской клетки, другие требуют участия мужских и женских органов.
Одни растения размножаются путем клеточного деления родительской клетки, другие требуют участия мужских и женских органов.
Способы размножения включают:
Бесполое размножение
При этом способе размножения потомство производится одним родителем (игровые/половые клетки не участвуют).Хотя это основной способ размножения большинства одноклеточных растительноподобных организмов, некоторые многоклеточные растения также размножаются этим методом.
Существует несколько типов бесполого размножения у растений, в том числе:
· Деление — Тип размножения, при котором клетка делится с образованием нескольких клеток, напоминающих родительскую клетку. Множественное деление, наблюдаемое у некоторых водорослей , происходит в неблагоприятных условиях.
· Почкование — Форма бесполого размножения, при которой отросток, образовавшийся на одной стороне родительского организма, в конечном итоге отламывается, образуя новый организм. У растений этот метод включает прививку почки (одного растения) на стебель другого растения, где он может развиться, чтобы сформировать новую особь.
У растений этот метод включает прививку почки (одного растения) на стебель другого растения, где он может развиться, чтобы сформировать новую особь.
· Фрагментация — Форма бесполого размножения, наблюдаемая у таких растений, как спирогира (нитчатые зеленые водоросли).Во время фрагментации отколовшиеся фрагменты организма развиваются, образуя новый организм. Этот способ размножения распространен среди многоклеточных растений, которые после созревания распадаются на несколько частей.
· Спороношение — Спорообразование является средством размножения, которое происходит у низших животных (нецветковых растений), таких как папоротники. Здесь споры, образующиеся в результате мейоза, попадают в окружающую среду, где в конечном итоге начинают прорастать, образуя новую особь в благоприятных условиях.
Вегетативное размножение
Вегетативное размножение — это форма бесполого размножения, при которой из вегетативных частей растения (корней, стебля и т. д.) развивается новая особь (растение). Таким образом, гаметы/половые клетки не участвуют. Здесь стоит отметить, что некоторые растения, использующие этот способ размножения, также способны размножаться половым путем.
д.) развивается новая особь (растение). Таким образом, гаметы/половые клетки не участвуют. Здесь стоит отметить, что некоторые растения, использующие этот способ размножения, также способны размножаться половым путем.
Некоторые из растений, которые используют этот режим воспроизведения, включают в себя:
Учитывая, что вегетативное размножение/размножение включает выращивание новых растений из частей более старого растения (корни, стебли, почки и т. д.), некоторые из методов, используемых для размножения, включают: Присоединение части побега (привоя) растения к нижней части (стеблю или корню) того же или другого вида растения (подвоя) для продолжения роста привоя.
· Окулировка — Аналогично прививке, но включает использование пазушной почки. Здесь пазушная почка присоединяется к подвою для дальнейшего роста. Как и при прививке, окулировка, как правило, бывает успешной в тех случаях, когда две части тесно связаны (например, растения апельсина и лимона).
Как и при прививке, окулировка, как правило, бывает успешной в тех случаях, когда две части тесно связаны (например, растения апельсина и лимона).
· Воздушные отводки (Markotting ) — Метод, используемый для стимуляции корнеобразования. Здесь раненый (срезанный) участок растения оборачивают влажным мхом сфагнумом.Затем мох покрывают черной бумагой или алюминиевой фольгой, чтобы предотвратить солнечные ожоги и сохранить влагу. Растению требуется от нескольких недель до нескольких месяцев, чтобы укорениться.
· Черенки — Другой метод, который используется для получения новых растений из частей данного растения. Здесь небольшая часть получается из растения и выращивается для получения нового растения. Это может включать использование таких частей, как часть стебля или листа растения (с почкой), для выращивания нового растения при выращивании в почве.Этот метод был использован для успешного выращивания таких растений, как виноград и бугенвиллия.
Половое размножение
По сути, половое размножение — это форма размножения, которая включает образование зиготы после слияния мужских и женских половых клеток, известных как гаметы.
У цветковых растений цветки служат репродуктивными единицами растения.
Цветки состоят из следующих частей:
У растений половое размножение начинается после опыления.Во время опыления мужские гаметы в пыльце того же цветка или другого цветка (одного и того же вида) переносятся на рыльце, которое является женской частью цветка.
Здесь ветер, насекомые, птицы и даже капли воды могут играть роль в переносе пыльцы и поэтому называются агентами переноса. Насекомых и птиц привлекают яркие цвета или сладкий запах.
* При самоопылении пыльца растения опыляет цветок того же растения. С другой стороны, при перекрестном опылении пыльца растения опыляет цветок другого растения того же вида.
С другой стороны, при перекрестном опылении пыльца растения опыляет цветок другого растения того же вида.
После опыления пыльцевое зерно образует тонкую трубку, которая спускается по столбику к завязи. В то же время клетка, находящаяся в пыльцевом зерне, подвергается делению с образованием двух мужских клеток (гамет), которые проходят через пыльцевую трубку и достигают женской гаметы в семяпочке завязи.
Оплодотворение происходит, когда одна из мужских гамет сливается с женской гаметой в яйцеклетке, таким образом объединяя гаплоидные хромосомы двух.В результате образуется оплодотворенная яйцеклетка (зигота), содержащая полную пару хромосом (диплоидная). Другая мужская гамета сливается с вторичным ядром, образуя триплоидное ядро эндосперма. Это, в свою очередь, развивается в эндосперм, который является источником питания для зародыша.
* Зародыш развивается, чтобы произвести семя (двудольное или однодольное), которое может развиться, чтобы сформировать новое растение, когда условия благоприятны.
* В зависимости от растения в плодах могут находиться семена.
* У нецветковых растений, таких как мхи, мужские и женские структуры образуются во время развития растений для полового размножения.
Фотосинтез — это процесс, посредством которого растения используют различные неорганические материалы для синтеза пищи.
Некоторые из частей, которые участвуют в фотосинтезе, включают:
· Листья — Листья содержат хлоропласты в слое мезофилла, где происходит фотосинтез.Листья также имеют поры (устьица) на своей поверхности, через которые углекислый газ поглощается из атмосферы.
· Сосудистая система — Вода от корней транспортируется к листьям (и другим частям растения), где она используется для фотосинтеза.
· Корни — Корни растений играют важную роль в поглощении воды из почвы. Затем вода транспортируется по сосудистой системе к месту фотосинтеза.
* Пигмент хлорофилла в хлоропластах (в клетках листьев) захватывает световую энергию, используемую для фотосинтеза.
Из-за различий в экологических и климатических условиях растения в разных экологиях адаптировались к различным типам фотосинтеза, которые включают:
C3 фотосинтез растения (около 83% всех растений).Во время первой фазы фотосинтеза углекислый газ реагирует с RuBP (рибулозо-1,5-бифосфатом), двумя молекулами 3-фосфоглицериновой кислоты, которая проходит цикл Кальвина с образованием сахаров и энергии.
* C3 представляет собой трехуглеродное соединение 3-фосфоглицериновой кислоты
* Было показано, что в жарких и сухих условиях процесс фотосинтеза C3 является очень расточительным из-за процесса, известного как фотодыхание. .
Фотосинтез С4
У растений С4 светозависимые реакции протекают в клетках мезофилла, тогда как цикл Кальвина протекает в особых клетках, известных как клетки оболочки пучка. На первом этапе (в клетках мезофилла) углекислый газ фиксируется фосфоенолпируватом с образованием малата (4-углеродного). Затем малат транспортируется к клеткам оболочки пучка, где он расщепляется на углекислый газ и 3-углеродное соединение.
На первом этапе (в клетках мезофилла) углекислый газ фиксируется фосфоенолпируватом с образованием малата (4-углеродного). Затем малат транспортируется к клеткам оболочки пучка, где он расщепляется на углекислый газ и 3-углеродное соединение.
* Одним из самых больших преимуществ этого процесса фотосинтеза является то, что для цикла Кальвина требуется большое количество углекислого газа. Это, в дополнение к отсутствию кислорода, означает, что процесс не страдает от эффектов фотодыхания
* Недостаток этого процесса заключается в том, что в жарких и сухих условиях Rubisco имеет тенденцию потреблять больше кислорода, что приводит к фотодыхание.
Фотосинтез метаболизма крассуловой кислоты (CAM)
Около 10 процентов растений используют фотосинтез CAM. Эти растения обычно встречаются в очень жарких и сухих условиях и включают в себя ананас, орхидеи и кактусы.
По сравнению с фотосинтезом C4, где световые реакции являются отдельными, процессы фотосинтеза CAM разделены на разные периоды времени. Например, ночью растения открывают устьица, что позволяет листьям поглощать углекислый газ.
Например, ночью растения открывают устьица, что позволяет листьям поглощать углекислый газ.
Затем газ фиксируется ФЕП-карбоксилазой с образованием оксалоацетата, который, в свою очередь, превращается в малат и накапливается в вакуолях. В течение дня устьица остаются закрытыми. Хотя углекислый газ не поступает из атмосферы, органические кислоты (например, малат), хранящиеся в вакуолях, высвобождаются и расщепляются с выделением углекислого газа, что соответствует циклу Кальвина.
* Эта форма фотосинтеза полезна для растений, произрастающих в очень жарких и засушливых районах, поскольку устьица остаются закрытыми в течение дня, что предотвращает потерю ими воды.
Вклад микроскопии в понимание растительной клетки и биологии
После ряда усовершенствований ученые смогли идентифицировать отдельные клетки, что привело к выводу, что все сущее состоит из клеток (клеточная теория).
В биологии растений микроскопия позволила идентифицировать различные типы клеток и, таким образом, определить их роль в развитии растений. Кроме того, микроскопия позволила исключить некоторые организмы (например, грибы) из царства растений.
Кроме того, микроскопия позволила исключить некоторые организмы (например, грибы) из царства растений.
Таким образом, это позволило определить характеристики организмов, подходящих для царства, и исключить те, которые не соответствуют этим характеристикам. Итак, в настоящее время некоторые другие царства описываются в соответствии с их особыми характеристиками.
Некоторые другие вклады микроскопии в биологию растений включают:
· Идентификация одноклеточных растительноподобных организмов – сделала возможным дифференцировать растения от растительноподобных организмов, таких как зеленые водоросли.
· Идентификация различных типов фотосинтеза.
· Изучение характеристик хлоропластов (например, содержание пигмента, способность двигаться в ответ на раздражители и т. д.).
· В настоящее время передовая микроскопия используется для определения способов повышения урожайности за счет управления различными процессами растений.
Другие биологии растений связанные статьи:
Структура листьев под микроскопом
Passipyntesis
Page на Plastids
Узнайте больше о хлоропластах
мезофиллированные клетки
Meristem клетки
Bucdding
Спорация
Трансгенные растения
Микроразмножение
Что такое эпифиты?
Понимание фотосинтезирующих бактерий
Каковы различия между растительной клеткой и животной клеткой?
Возврат с главной страницы биологии растений на главную страницу MicroscopeMaster
Ссылки
Кингсли Р.Стерн, Джеймс Э. Бидлак и Шелли Х. Янски. (2008). Введение в биологию растений. Издание одиннадцатое.
Лики Р.Р.Б. (2014). Физиология вегетативного размножения.
Юань Л. и Сундаресан В. (2015). Спорообразование у растений: бесспоровидное и др. NCBI.
Ссылки
https://www. bbc.com/news/science-environment-39897119
bbc.com/news/science-environment-39897119
Биология растений – обзор
Резюме
Биотехнология охватывает три аспекта биологии древесных растений: (1) культура in vitro, (2) использование иммунологических методов для количественного определения метаболитов растений и (3) генная инженерия, т.е. , перенос генов между организмами без нормального полового размножения.Быстрое развитие в этих областях сделало возможным производство и массовое размножение новых генотипов важных лесных и садовых сельскохозяйственных культур.
Процедуры биотехнологии во многом зависят от понимания молекулярной биологии репликации, транскрипции и трансляции ДНК. В связи с клеточным делением репликация ДНК происходит с помощью ДНК-полимераз одновременно во многих местах генома, где была раскручена хромосомная ДНК. Экспрессия генов начинается с транскрипции, синтеза информационной РНК (мРНК) в серии реакций с участием РНК-полимераз и различных регуляторных молекул.Этот процесс обеспечивает транскрипцию специфических генов, тем самым обеспечивая правильную координацию экспрессии генов в ходе развития. Белковые продукты возникают в результате трансляции мРНК на рибосомах в цитоплазме.
Белковые продукты возникают в результате трансляции мРНК на рибосомах в цитоплазме.
В процессе генетической трансформации новые гены стабильно внедряются в геном клеток-мишеней. Существует множество методов для достижения трансформации, в том числе связанные с биологическими векторами и физическими методами введения путем прямого воздействия ДНК на клетки или протопласты и ускорения небольших снарядов, покрытых ДНК, в ткани растений.В наиболее частом биологическом методе трансформации растений используются виды Agrobacterium , которые несут плазмиду, содержащую участок ДНК, который легко интегрируется в геном растения. Совместное культивирование бактерии с растительными клетками или протопластами приводит к включению интересующего гена вместе с (1) последовательностями ДНК, которые направляют его транскрипцию, (2) репортерным геном, который подтверждает наличие желаемого гена после элиминации бактерии из культура и (3) ген, придающий устойчивость к последующим химическим веществам, так что нетрансформированные клетки могут быть уничтожены. Прямому переносу сходных компонентов ДНК способствуют такие обработки, как воздействие на протопласты полиэтиленгликолем и импульсными электрическими полями (электропорация). Баллистические методы введения ДНК могут быть достигнуты с тканями, если клетки в меристематических областях могут быть трансформированы.
Прямому переносу сходных компонентов ДНК способствуют такие обработки, как воздействие на протопласты полиэтиленгликолем и импульсными электрическими полями (электропорация). Баллистические методы введения ДНК могут быть достигнуты с тканями, если клетки в меристематических областях могут быть трансформированы.
Доступно несколько методов подтверждения переноса и экспрессии генов. Наличие специфического участка ДНК в геноме растительной клетки может быть подтверждено саузерн-гибридизацией, при которой растительная ДНК расщепляется на фрагменты специфическими рестрикционными ферментами с последующей гибридизацией фрагментов с радиоактивно мечеными последовательностями ДНК гена интерес (зонды).Транскрипцию генов можно подтвердить с помощью Нозерн-гибридизации, процедуры, аналогичной Саузерн-гибридизационному анализу, при котором РНК расщепляется по определенным последовательностям ферментами и исследуется известными радиоактивно меченными последовательностями нуклеиновых кислот, которые соответствуют интересующей мРНК.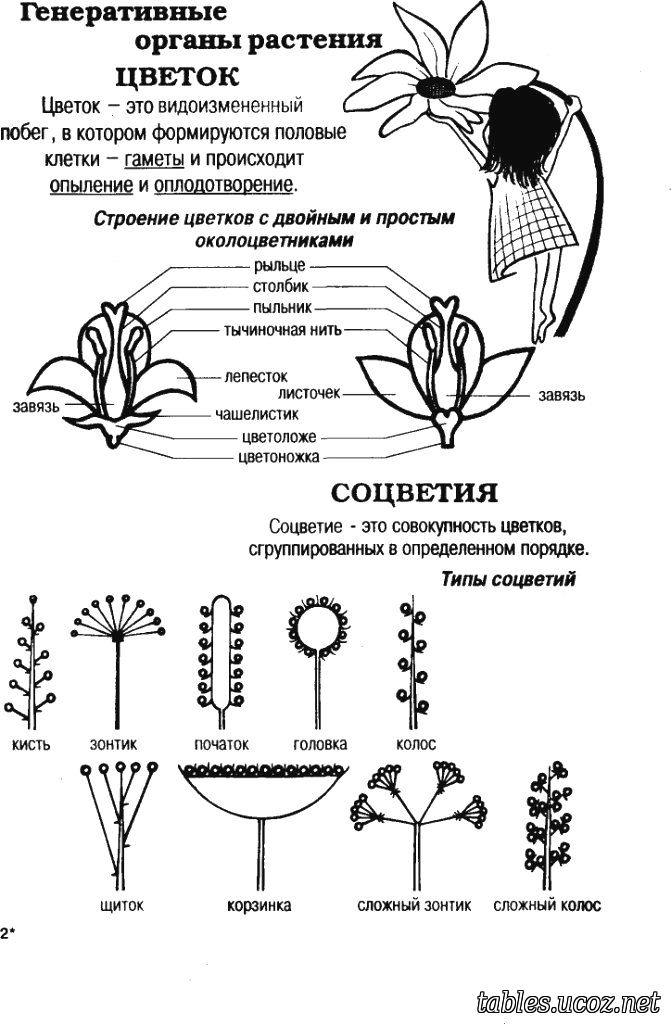
Белковые продукты часто выявляются и количественно определяются с помощью анализов на основе антител. Антитела вырабатываются у животных в ответ на введение в организм чужеродных макромолекул. Эти антитела могут быть выделены и очищены, и они будут связываться с метаболитами растений, из которых они были получены.Связь антител с ферментами, катализирующими цветообразующие реакции, позволяет проводить простые колориметрические измерения концентрации.
Методы молекулярной биологии значительно упростили генетический анализ. Генетическая изменчивость на уровне ДНК может быть обнаружена с помощью анализа полиморфизма длин рестрикционных фрагментов (ПДРФ), в котором растительная ДНК расщепляется ферментами рестрикции по определенным последовательностям оснований, а фрагменты разделяются с помощью электрофореза. Вариации последовательности оснований между разными генотипами приводят к тому, что фрагменты имеют разную длину и, следовательно, перемещаются в геле на разные расстояния. Количество растительной ДНК, необходимой для анализа, было значительно уменьшено за счет разработки полимеразной цепной реакции (ПЦР), в которой используется циклическая репликация in vitro последовательностей специфических рестрикционных фрагментов ДНК для значительного увеличения количества фрагментов выделенной ДНК. Другой вариант этого метода, процедура случайной амплификации полиморфной ДНК (RAPD), использует праймеры случайной последовательности для прямой репликации коротких фрагментов ДНК.
Количество растительной ДНК, необходимой для анализа, было значительно уменьшено за счет разработки полимеразной цепной реакции (ПЦР), в которой используется циклическая репликация in vitro последовательностей специфических рестрикционных фрагментов ДНК для значительного увеличения количества фрагментов выделенной ДНК. Другой вариант этого метода, процедура случайной амплификации полиморфной ДНК (RAPD), использует праймеры случайной последовательности для прямой репликации коротких фрагментов ДНК.
Генная инженерия часто проводится на клетках и протопластах, культивируемых in vitro, которые необходимо регенерировать до целых растений.Кроме того, часто желательно массовое размножение новых генотипов, созданных традиционными методами селекции и селекции. Следовательно, были предприняты значительные усилия, направленные на разработку средств вегетативного размножения растительных материалов для распространения. Такие методы могут быть связаны с традиционными методами, такими как укоренение черенков или методы культивирования in vitro. Многие древесные растения, например ивы и тополя, легко размножаются укорененными черенками. В других случаях укоренение получить не так легко; тем не менее, был достигнут достаточный прогресс в укоренении важных деревьев, таких как сосна монтерейская и европейская ель, так что коммерческая доступность превосходных генотипов этих видов из черенков стала реальностью.
Многие древесные растения, например ивы и тополя, легко размножаются укорененными черенками. В других случаях укоренение получить не так легко; тем не менее, был достигнут достаточный прогресс в укоренении важных деревьев, таких как сосна монтерейская и европейская ель, так что коммерческая доступность превосходных генотипов этих видов из черенков стала реальностью.
Микроразмножение может быть осуществлено путем культивирования недифференцированных тканей каллуса и последующей индукции органов путем манипулирования культуральной средой и концентрациями регуляторов роста растений. В качестве альтернативы черенки можно получить непосредственно путем укоренения микрочеренков, полученных из культуры побегов дифференцированных тканей, таких как побеги, почки или листья. Соматические эмбрионы также были получены с помощью соответствующих манипуляций с культуральными средами, чтобы они напоминали режим развития in situ. Неограниченное производство проэмбриоидов может поддерживаться, а полученные проэмбрионы могут созревать благодаря воздействию АБК и низкому осмотическому потенциалу.
Микроразмножение также может быть достигнуто путем выделения протопластов, культивирования и регенерации. Протопласты выделяют путем воздействия на клетки или ткани ферментов, расщепляющих клеточную стенку. После этого можно манипулировать свободными протопластами для слияния (иногда с образованием настоящих соматических гибридов) или генетически трансформировать. Вскоре формируются клеточные стенки протопластов, и при последующем культивировании протопластов можно получить генетически однородные колонии.
Потенциальные улучшения растений, которые могут быть получены с помощью генной инженерии, многочисленны, включая повышенную устойчивость к гербицидам, большую устойчивость к насекомым, грибкам и вирусным вредителям, преждевременное цветение, задержку деградации плодов и старения цветков, изменение окраски цветочных культур и повышенная устойчивость к загрязнениям. Экспрессия признаков у трансформированных растений не всегда стабильна, так как транскрипция введенных генов может быть подавлена. Кроме того, поскольку для достижения трансформации растений обычно требуется культура in vitro , эти методы могут быть недоступны для многих неподатливых древесных растений.Необходимо разработать соответствующие меры контроля для предотвращения перемещения чужеродных генов в естественные популяции растений.
Кроме того, поскольку для достижения трансформации растений обычно требуется культура in vitro , эти методы могут быть недоступны для многих неподатливых древесных растений.Необходимо разработать соответствующие меры контроля для предотвращения перемещения чужеродных генов в естественные популяции растений.
Многие древесные виды, в том числе важные виды покрытосеменных и голосеменных растений, подвергались микроразмножению. Однако микроразмножение многих древесных растений, хотя и весьма полезное, представляет собой сложную задачу. Культивирование растений in vitro в значительной степени является эмпирической задачей, и утомительный поиск подходящих сред и протоколов сопровождает культивирование почти каждого нового растительного материала.Регенерация растений также часто затруднена. Хотя с точки зрения селекции высших генотипов желательно получать культуры из зрелых растений, микроразмножение зрелых растений часто оказывается довольно трудным. Кроме того, затраты на микроразмножение все еще высоки по сравнению с традиционным размножением семенами.
Определения растений и примеры — Биологический онлайн-словарь
существительное, растение
Любой фотосинтезирующий эукариотический организм с жесткой клеточной стенкой.
Определение растений
( ботаника ) Любой из эукариотических организмов биологического царства Plantae, характеризующийся фотосинтезом и жесткой клеточной стенкой. Этимология : от латинского planta («росток, побег, срез»).
Характеристики растений
Растение относится к любому из эукариот, принадлежащих биологическому царству Plantae. Растения, в самом строгом смысле, являются эмбриофитами, к которым относятся сосудистые растения, печеночники, роголистники и мхи.В некоторых менее строгих источниках зеленые водоросли рассматриваются как растения. Зеленые водоросли состоят из одноклеточных и многоклеточных видов, которые имеют хлоропласты и клеточную стенку. Основные характеристики, перечисленные ниже, относятся к эмбриофитам. Они следующие:
Они следующие:
Тело растения
Эмбриофиты, как правило, имеют две основные системы органов: (1) побеговую систему и (2) корневую систему. Система побегов включает части тела, расположенные в верхней части растения, тогда как корневая система состоит из частей тела, находящихся в нижней части.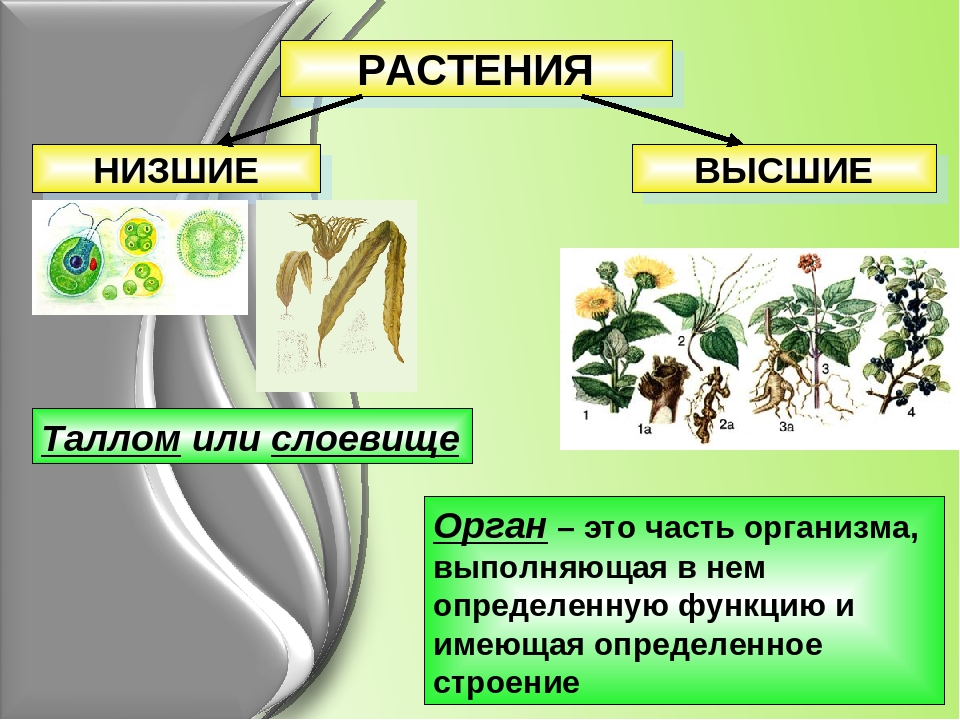 Система побегов может включать органы растения, такие как стебли, ветви, листья, цветы и плоды. Их часто можно найти над землей. Корневая система включает корни, клубни и корневища.Их часто находят под землей.
Система побегов может включать органы растения, такие как стебли, ветви, листья, цветы и плоды. Их часто можно найти над землей. Корневая система включает корни, клубни и корневища.Их часто находят под землей.
Ткани растений:
Клетки растений эукариотические, т.е. с хорошо выраженным ядром. Ядро содержит хромосомы, несущие гены. Помимо ядра, другими органеллами являются эндоплазматический ретикулум, аппарат Гольджи, митохондрии, лизосомы и пластиды. Пластиды можно классифицировать на основе пигментов: хлоропласты (с хлорофиллом, зеленым пигментом), хромопласты (с пигментами, кроме зеленого) и лейкопласты (бесцветные пластиды).Крупная структура внутри растительной клетки — вакуоль. Он отвечает за регуляцию тургорного давления.
Пластиды можно классифицировать на основе пигментов: хлоропласты (с хлорофиллом, зеленым пигментом), хромопласты (с пигментами, кроме зеленого) и лейкопласты (бесцветные пластиды).Крупная структура внутри растительной клетки — вакуоль. Он отвечает за регуляцию тургорного давления.
Плазматическая мембрана окружает цитоплазму, в которой находятся эти органеллы. Помимо плазматической мембраны, клетка имеет дополнительный слой, называемый клеточной стенкой. Однако клеточная стенка характерна не только для эмбриофитов. Другие организмы, такие как грибы, водоросли и некоторые бактерии, имеют клеточные стенки. Клеточная стенка эмбриофитов состоит из первичной и вторичной клеточных стенок.Первичная клеточная стенка содержит целлюлозу, гемицеллюлозы и пектин. Вторичная клеточная стенка представляет собой более толстый слой. Он богат лигнином, который укрепляет и делает стену водонепроницаемой. Клеточная стенка выполняет множество важных функций, и одна из них заключается в сопротивлении осмотическому давлению.
Когда растительную клетку помещают в гипертонический раствор, вода проникает в клетку и вызывает ее набухание. Наличие клеточной стенки предотвращает разрыв клетки при чрезмерном осмосе. И наоборот, когда растительную клетку помещают в гипотонический раствор, вода диффундирует из клетки, и тургорное давление теряется, в результате чего клетка становится дряблой.Дальнейшая потеря воды приведет к плазмолизу и, наконец, к циторизу , полному разрушению клеточной стенки.
Помимо осморегуляции, к основным физиологическим процессам, которые осуществляют растения, относятся фотосинтез, дыхание, транспирация, тропизмы, настические движения, фотопериодизм, циркадные ритмы, прорастание семян и покой.
Геномика растений
Растения имеют большой геном. Среди геномов растений, которые были секвенированы, геном пшеницы Triticum asestivum является самым большим, с его примерно 94 000 генов. 4
Жизненный цикл растений
Жизненный цикл растений состоит из двух поколений: поколение гаметофита и поколение спорофита . Фаза чередования диплоидных и гаплоидных форм называется чередованием поколений . Это также наблюдается у некоторых водорослей, таких как Archaeplastida и Heterokontophyta. У водорослей с чередованием поколений спорофит и гаметофит являются самостоятельными организмами.
Фаза чередования диплоидных и гаплоидных форм называется чередованием поколений . Это также наблюдается у некоторых водорослей, таких как Archaeplastida и Heterokontophyta. У водорослей с чередованием поколений спорофит и гаметофит являются самостоятельными организмами.
У эмбриофитов поколение гаметофитов – это поколение, в котором фаза начинается с гаплоидной споры (n). Спора претерпевает серию митотических делений, в результате чего образуется гаметофит. Гаметофит — гаплоидная многоклеточная форма растения. У него будет только один набор хромосом. Фаза гаметофита — это половая фаза в жизненном цикле, поэтому у растения разовьются половые органы, производящие гаметы, которые также являются гаплоидными. Гаметы, участвующие в оплодотворении, позже войдут в поколение спорофита, характеризующееся диплоидной формой растения после объединения гамет.
У трахеофитов ( сосудистых растений ) спорофит находится в многоклеточной форме и в доминирующей фазе. Таким образом, спорофит представляет собой основное растение, которое мы видим. И наоборот, у мохообразных (90 526, например, 90 527 мхов и печеночников) гаметофит является доминирующим и, следовательно, является основным растением, которое мы воспринимаем.
Таким образом, спорофит представляет собой основное растение, которое мы видим. И наоборот, у мохообразных (90 526, например, 90 527 мхов и печеночников) гаметофит является доминирующим и, следовательно, является основным растением, которое мы воспринимаем.
Как правило, жизненные стадии трахеофитов начинаются с семени, которое развивается в отросток, когда условия благоприятны для роста. Привой растет за счет образования листьев и роста стеблей и ветвей.Он развивается во взрослое растение, которое в конечном итоге дает цветы. Цветки несут половые клетки, такие как сперматозоиды в пыльцевом зерне и яйцеклетки в семязачатках завязи. В результате объединения половых клеток внутри семени образуется зигота. Однодомные растения несут обе половые клетки, тогда как двудомные растения несут только один тип половых клеток.
Растения также могут размножаться бесполым путем. Они делают это, не вовлекая гаметы. При бесполом размножении новые растения возникают путем почкования, фрагментации, деления, спорообразования, вегетативного размножения и апомиксиса.
Старение растений относится к процессу старения растений. Например, пожелтение листьев происходит в результате деградации хлорофилла, в результате чего остаются только каротиноиды во время старения листьев. Однако некоторые растения могут продолжать формировать новые листья, например лиственные растения.
Экология растений
Поскольку растения способны к фотосинтезу, им не нужно охотиться или питаться животными для получения пищи (за исключением плотоядных растений). Они могут производить себе пищу, используя энергию света, углекислый газ из атмосферы и молекулы воды.Тем не менее, одним из источников углекислого газа являются отходы , которые животные выдыхают при дыхании. Взамен они выделяют кислород как побочный продукт фотосинтеза. Кислород имеет решающее значение для выживания аэробных организмов, включая животных.
Растения получают другие жизненно важные питательные вещества из минералов, растворенных в почве. Они поглощают их через свои корни. Некоторые из макроэлементов, которые они получают из почвы, — это кальций, магний, азот, фосфор, калий и сера.Что касается микроэлементов, то растения усваивают бор, хлорид, медь, железо, марганец, молибден. Таким образом, мертвые части или целые растения приводят к их разложению и возвращению на Землю необходимых минералов и соединений.
Некоторые из макроэлементов, которые они получают из почвы, — это кальций, магний, азот, фосфор, калий и сера.Что касается микроэлементов, то растения усваивают бор, хлорид, медь, железо, марганец, молибден. Таким образом, мертвые части или целые растения приводят к их разложению и возвращению на Землю необходимых минералов и соединений.
Из-за чувства независимости их часто помещают в начало пищевой цепи. Они являются основными производителями в экосистеме. Таким образом, вымирание видов растений может оказать серьезное воздействие на экосистему. Международный союз охраны природы (МСОП) Красный список видов, находящихся под угрозой исчезновения , система оценки статуса сохранения видов во всем мире, использовала систему маркировки видов на основе риска исчезновения.Соответственно, виды можно разделить на категории: «недостаточно данных», «вызывающие наименьшее беспокойство», «находящиеся под угрозой исчезновения», «уязвимые», «находящиеся под угрозой исчезновения», «находящиеся под угрозой исчезновения», «исчезнувшие в регионе», «исчезнувшие в дикой природе» и «вымерший». В 2016 году МСОП сообщил, что 2493 растения находятся под угрозой исчезновения, тогда как 3654 растения находятся под угрозой исчезновения. 5
В 2016 году МСОП сообщил, что 2493 растения находятся под угрозой исчезновения, тогда как 3654 растения находятся под угрозой исчезновения. 5
Растения взаимодействуют с другими организмами и образуют симбиоз. Примеры следующие:
8. На Земле насчитывается 7 миллионов 90 574 видов эукариот, и из этого числа около 298 000, по прогнозам, составляют общее количество видов растений. 215, 644 уже были описаны и каталогизированы. 6
На Земле насчитывается 7 миллионов 90 574 видов эукариот, и из этого числа около 298 000, по прогнозам, составляют общее количество видов растений. 215, 644 уже были описаны и каталогизированы. 6
Эволюция растений
Согласно эндосимбиотической теории , такие органеллы, как пластиды и митохондрии, представляют собой ранее свободно жившие прокариоты. Хлоропласты, по-видимому, связаны с прокариотическими цианобактериями. В основе лежит структурное сходство между цианобактериями и хлоропластами.Более того, оба они имеют одинаковые фотосинтетические пигменты и одну кольцевую молекулу ДНК, что и геном. По-видимому, эндосимбиотические события привели к появлению первых фотосинтезирующих эукариот миллиард лет назад. Считается, что Charophyta (подгруппа зеленых водорослей) возникла из эмбриофитов. Харофиты и эмбриофиты имеют много общих черт, т.е. образование фрагмопластов во время митоза.
Краткая хронология эволюции эмбриофитов представлена ниже:
Рекомендуемый источник: Darwin Reviews – Журнал экспериментальной ботаники самых престижных обзоров и тем, тщательно отобранных в самых прогрессивных областях исследований.
Таксономия растений
Первоначальное определение растений включает зеленые водоросли, грибы и эмбриофиты, поскольку все они имеют хлоропласты и клеточную стенку. Однако со временем водоросли и грибы были перемещены в свои царства.
В самом узком смысле растения (т. е. Plantae sensu strictissimo ) — это растения, которые в основном являются многоклеточными, с клеточными стенками, содержащими целлюлозу, и имеют хлоропласты для фотосинтеза. В этом случае царство Plantae состоит из эмбриофитов, таких как сосудистые растения, печеночники, мхи и другие ископаемые растения, которые имеют те же признаки.
Plantae sensu stricto («растения в узком смысле») включает эмбриофиты и зеленые водоросли (Chlorophyta и Charophyta). Это до сих пор широко признанное определение растений. Они составляют кладу Viridiplantae (или Chlorobionta), которую обычно называют зелеными растениями . Различные подразделения королевства Plantae в буквальном смысле слова заключаются в следующем:
Это до сих пор широко признанное определение растений. Они составляют кладу Viridiplantae (или Chlorobionta), которую обычно называют зелеными растениями . Различные подразделения королевства Plantae в буквальном смысле слова заключаются в следующем:
Значение
Растения необходимы для жизни различных организмов, так как они являются производителями в пищевой цепи.Они запасают крахмал. Они также служат важным источником минералов и соединений.
Растения служат средой обитания для некоторых организмов ( например, насекомых и древесных организмов). Они также являются основным источником кислорода, необходимого аэробным животным.
Они также являются основным источником кислорода, необходимого аэробным животным.
Некоторые растения обладают лечебными свойствами. Одуванчик ( Taraxacum officinale ) в качестве мягкого слабительного, листья подорожника ( Plantago major ) для уменьшения воспаления и боли, а также корни и листья лопуха ( Arctium minus ) для облегчения экземы или потрескавшейся кожи — вот лишь некоторые из разнообразных средств. лекарственные растения.
Люди используют растения для производства различных продуктов, таких как эфирные масла, пигменты, смолы, дубильные вещества, алкалоиды, янтарь, воски, косметика, пластмассы, каучук, лаки, смазки, чернила и т.д.
Древесина растений используется при строительстве зданий, музыкальных инструментов, лодок и мебели. Он также используется в производстве бумаги.
Исследования
Раздел науки, изучающий растения, называется ботаника (или биология растений). Специалист в этой области называется ботаник . Некоторыми из областей исследования являются морфоанатомия, цитология, гистология, физиология, экология, эволюция, таксономия и патология. Различные группы растений привели к возникновению таких субдисциплин, как:
Некоторыми из областей исследования являются морфоанатомия, цитология, гистология, физиология, экология, эволюция, таксономия и патология. Различные группы растений привели к возникновению таких субдисциплин, как:
Прикладная ботаника занимается коммерческим и экономическим использованием растений.Он охватывает сельское хозяйство (, например, , агрономия, садоводство, растениеводство), лесное хозяйство (, например, , дендрология, технология обработки древесины), фармацевтическую ботанику и ландшафтную архитектуру.
Научные классификации
см.
 Также
ТакжеСсылка
Дополнительная литература
Примечания
Таксономическая классификация организмов может измениться на основе новых данных.
© Biology Online. Контент предоставлен и модерируется Онлайн-редакторами биологии
Биология растений — Неукротимая наука
Этот раздел предназначен для того, чтобы вы быстро ознакомились с основами биологии растений.Здесь, в Untamed Science, мы думаем, что растения довольно удивительны. У нас есть все виды забавных видеороликов о растениях. Вот забавный короткометражный фильм, который мы создали о лекарственных растениях.
И почему бы не добавить к этому видео второе видео, посвященное основным формам растений. Эти видеоролики были созданы в сотрудничестве с Pearson Publishing и в настоящее время сопровождают учебник по биологии для средней школы Миллера и Левина.
Биология растений — это раздел биологии, изучающий жизнь и развитие растений.Ботаника — другое название этой области биологии. Она охватывает широкий спектр научных дисциплин, изучающих растения. Некоторые из них включают структуру, рост, размножение, обмен веществ, развитие, болезни, химические свойства и эволюционные отношения между различными группами. Ботаника началась с усилий племен по выявлению съедобных, лекарственных и ядовитых растений, что сделало ботанику одной из древнейших наук. Из-за этого древнего интереса к растениям объем ботаники увеличился и теперь включает изучение более 550 000 видов или видов живых организмов.На этих страницах Untamed Science мы хотим охватить основы биологии растений, включая все, что можно было бы охватить на вводном уроке биологии. Для начала мы создали краткое введение в несколько основных форм растений.
Ботаника началась с усилий племен по выявлению съедобных, лекарственных и ядовитых растений, что сделало ботанику одной из древнейших наук. Из-за этого древнего интереса к растениям объем ботаники увеличился и теперь включает изучение более 550 000 видов или видов живых организмов.На этих страницах Untamed Science мы хотим охватить основы биологии растений, включая все, что можно было бы охватить на вводном уроке биологии. Для начала мы создали краткое введение в несколько основных форм растений.
Темы растений
Поскольку в этом году ботаники из UntamedScience добавят больше информации, мы также добавим на эти страницы образовательные видео. Будьте терпеливы с нами, все эти страницы находятся в разработке …
Биологическое разнообразие растений
Не забывайте, что у нас есть целый сайт, посвященный разнообразию растений, к которым мы постепенно добавляем новые виды.Вот несколько примеров. Кроме того, вот хорошее введение в ботанику для дальнейшего чтения.
растений | Короткие увлекательные видеоролики о биологии растений
Видео — отличный способ рассказать о науке о растениях. Они могут возбудить любопытство, объяснить сложную идею или сделать невидимое видимым. Вот тщательно подобранный список некоторых из наших любимых коротких видеороликов о растениях.
Ускоренный курс биологии: клетки растений (10 мин), фотосинтез (13 мин), половая жизнь несосудистых растений (10 мин), сосудистые растения = победа (12 мин), растения и пчелы: размножение растений (10 мин)
Быть умным — нормально: что появилось раньше — цветы или пчелы? (5 мин), Думают ли растения? (6 мин), «Твой салат пытается тебя убить» (6 мин), Сколько здесь деревьев? (6 мин)
Veritasium: Самое удивительное в деревьях (7 мин), Откуда деревья берут свою массу? (4 мин), Внутри хранилища семян на Шпицбергене (9 мин)
Minute Earth: почему (питательные вещества) бедные места более разнообразны (3 мин), Тайная социальная жизнь растений (2 мин), Как деревья переживают зиму? (3 мин), Почему листья зеленые? (Часть 1 и Часть 2) (по 2 мин каждая), Настоящая причина, по которой листья меняют цвет осенью (2 мин), Что было раньше — дождь или тропический лес? (3 мин), Кого пытаются соблазнить цветы? (3 мин), Орхидеи: Мастера лжи, обмана и воровства (4 мин)
BBSRC: Как наука о растениях переписывает наши истории о будущем (3 мин), Как растения выползли в море (2 мин), Энергия растений: растения против взрывчатых веществ (2 мин), Сила растений: растения против бензина (2 мин), Растения сила: Растения против рыб (2 мин), Сила растений: Растения против наводнений (2 мин), Сила растений: Растения против ожирения (2 мин), Еда будущего (2 мин)
PBS NewsHour: чтобы бороться с изменением климата, эти ученые превращают CO 2 в камень (2016, 9 мин). Конец бананам, какими мы их знаем? (2016, 5 мин), Восстановление водно-болотных угодий залива Сан-Франциско по одному местному растению за раз (2016, 6 мин), Тайная жизнь растений и автор «Лаборантки» Хоуп Джарен (2016, 6 мин), Медицинские исследования марихуаны выходит из тени (2016 г., 9 мин.), Как вырастить вакцину против Эболы с помощью растения табака (2015 г., 6 мин.), Калифорнийские секвойи, страдающие от нехватки воды, проявляют признаки стресса (2015 г., 6 мин.), Почему выращивание салата в Нью-Йорке Город Йорк – растущий бизнес (2015 г., 8 мин.) Разумно ли выращивать миндаль во время засухи? (2015, 8 мин), Почему почти половина еды в Америке выбрасывается? (2015, 9 мин), Итальянские оливковые деревья увядают от этой смертельной бактерии (2015, 7 мин), Фитопланктон, который оживил Землю (2014, 5 мин), Найден рай: История ананасов, сахара и семян на Гавайях (2013 г., 3 мин.), Баланс между сельским хозяйством Коста-Рики и сохранением природы (2013 г., 9 мин.), Калифорнийские работы по спасению францисканского растения Мансанита от вымирания (2011 г.
Конец бананам, какими мы их знаем? (2016, 5 мин), Восстановление водно-болотных угодий залива Сан-Франциско по одному местному растению за раз (2016, 6 мин), Тайная жизнь растений и автор «Лаборантки» Хоуп Джарен (2016, 6 мин), Медицинские исследования марихуаны выходит из тени (2016 г., 9 мин.), Как вырастить вакцину против Эболы с помощью растения табака (2015 г., 6 мин.), Калифорнийские секвойи, страдающие от нехватки воды, проявляют признаки стресса (2015 г., 6 мин.), Почему выращивание салата в Нью-Йорке Город Йорк – растущий бизнес (2015 г., 8 мин.) Разумно ли выращивать миндаль во время засухи? (2015, 8 мин), Почему почти половина еды в Америке выбрасывается? (2015, 9 мин), Итальянские оливковые деревья увядают от этой смертельной бактерии (2015, 7 мин), Фитопланктон, который оживил Землю (2014, 5 мин), Найден рай: История ананасов, сахара и семян на Гавайях (2013 г., 3 мин.), Баланс между сельским хозяйством Коста-Рики и сохранением природы (2013 г., 9 мин.), Калифорнийские работы по спасению францисканского растения Мансанита от вымирания (2011 г. , 7 мин.)
, 7 мин.)
Пифле: что такое ГМО? (5 мин)
Вашингтонский университет, ул.Луи: Почему растения не растут вверх ногами? (3 мин)
HHMI Interactive: раскрытый секрет: загадочное происхождение кукурузы (18 мин)
SAPS: анимация — транспорт воды и сахара в растениях (4 мин)
Видеотека BBC – Растения (несколько роликов разной длины)
Герцог Фан Тран, Университет Галле: Xanthomonas (компьютерная анимация процесса заражения) (6 мин)
IntoBiology: от клетки до планеты за 90 секунд (1,5 мин)
Глобальная инициатива Borlaug по борьбе с ржавчиной: жизненный цикл стеблевой ржавчины пшеницы (7 мин)
CIMMYT: зачем инвестировать в исследования пшеницы (5 мин)
Королевский ботанический сад Эдинбурга: движение вперед после отмирания ясеня (3 мин)
Сады Кью: забытый дом кофе (6 мин)
BBC Unplugged: взрывающиеся семена, замедленная съемка (5 мин)
Университет Монаша: Накормить будущее с участием Рос Глидоу (4 мин)
Университет Глазго: Исследование растений (PSI) (обман CSI) (7 мин)
Научный журнал: Здоровье лесов в меняющемся мире (2. 5 мин)
5 мин)
TED-Ed: Как одноклеточный организм почти уничтожил жизнь на Земле (Анусуя Уиллис, 4 мин), Могут ли растения разговаривать друг с другом? (Ричард Карбан, 5 мин.), Простая история о фотосинтезе и еде (Аманда Утен, 4 мин.)
Энциклопедия жизни: инвазивные виды, рассказы о биоразнообразии в движении (7 мин)
Американский музей естественной истории: Зеленый рост саламандр (6,5 мин)
Рамеш Бунратана: Мангровые леса и живородок (2,5 мин)
Международный институт питания растений: Роль управления питательными веществами 4R в снижении выбросов парниковых газов (5 мин)
Biolapse: Carnivora Gardinum (замедленная съемка плотоядных растений) (4 мин)
Коллекции и каналы
Серия Криса Мартина «Растения тоже крутые» – несколько видеороликов (7–15 мин.)
Серия видеороликов Клэр Хопкинс Brilliant Botany — несколько видеороликов (большинство < 5 минут)
На сайте Роджера Хангартера «Растения в движении» можно найти много видеороликов с интервальной съемкой (большинство < 3 мин)
На закрытом конкурсе видеороликов ChloroFilms представлены отличные видеоролики
Продовольственная и сельскохозяйственная организация (ФАО): десятки видеороликов на многих языках, в том числе видеоролики Года зернобобовых
МФСР: Международный фонд сельскохозяйственного развития. Видео о проблемах фермерства по всему миру
Видео о проблемах фермерства по всему миру
Смитсоновский канал: Растения
Видео CABI: много видео об инвазивных видах и агрономических методах
AgriumInc: Удобрение 101 (8 видео об удобрениях)
Видео с песнями
Кембриджская команда iGEM: Песня сборки Gibson (4 мин)
Калифорнийский университет в Ирвине: химики знают (пародия на Let it Go из Frozen) (4 мин)
Musical Cells: Power Pack: The mitochondria rock song (4 мин)
Biology Fortified Inc: Play it hard — Норман Борлоуг, посвященный 100-летию (3 мин)
Kamoun Lab: Оомицет Фифи (2 мин) и Вы можете звать меня Ал (для Альбуго) (4 мин)
Лаборатория Йоникаса ♫ Ученые спасают мир с помощью марионеток и водорослей ♫ (3.5 мин)
HarvestPlus: лучшее питание благодаря фасоли с высоким содержанием железа (5 мин)
Крис Холмс: La Bloomba (2,5 мин)
Science Rap Academy: Они растут (2 мин)
ZhengLabProductions: Bad Project (Пародия на Леди Гагу) (5 мин)
Лаборатория массажа: не прекращайте пипетирование (5 мин)
CohenfordMU: The Lab Song (пародия на Бруно Марса) (3,5 мин)
Weigelworld: Shake it off (пародия) (4 мин)
Weigel Style (пародия на Gangnam Style) (5 мин)
И многие другие пародии на научные песни можно найти на YouTube….
Биология растений | Отдел биологии
Давление развития, особенно в тропических странах, вызывает тревожное увеличение скорости вымирания видов, что делает нынешнее возрождение систематики особенно своевременным. Учитывая разумную оценку того, что систематики открыли и назвали только около 10% видов на Земле, и тот факт, что лишь крошечная часть этих видов была изучена сколь-либо подробно, за короткое время предстоит проделать большую работу.Многие виды вымрут еще до того, как мы их узнаем; неудивительно, что систематикам кажется, будто они наблюдают, как огромная разнообразная библиотека сгорает еще до того, как будет подготовлен картотека (или до того, как кто-либо прочитает хотя бы 1% книг!). Недавно разработанные методы сбора данных и анализа филогенетических отношений ставят нас на порог глубокого понимания истории биологического мира. Таким образом, утрата биологического разнообразия является катастрофой как с экономической точки зрения (сколько организмов, пригодных для еды, медицины или технологий, вымрет?), так и с более широкой интеллектуальной точки зрения (как разнообразие видов стало таким, каким оно стало сейчас). является?).
является?).
Систематики должны иметь технические навыки для извлечения информации на всех уровнях исследования (например, морфология, цитология, генетика, последовательности ДНК, органическая химия, анатомия, экология) и теоретические основы для правильной ее интерпретации. Современная биологическая систематика объединяет широкий спектр дисциплин, от молекулярной, клеточной биологии и биологии развития до экологии и эволюционной биологии. Методы сбора данных включают секвенирование ДНК, электрофорез белков, электронную и световую микроскопию, эксперименты с контролируемым ростом и полевые исследования экологии и распределения.Аналитические методы требуют больших вычислительных ресурсов: для автоматизированного описания морфологии (морфометрия) используются такие аппаратные средства, как планшеты для оцифровки и видеокамеры, для описания и сравнения видов и других таксонов используются многомерные статистические данные, для реконструкции филогении используются числовые кладистические программы.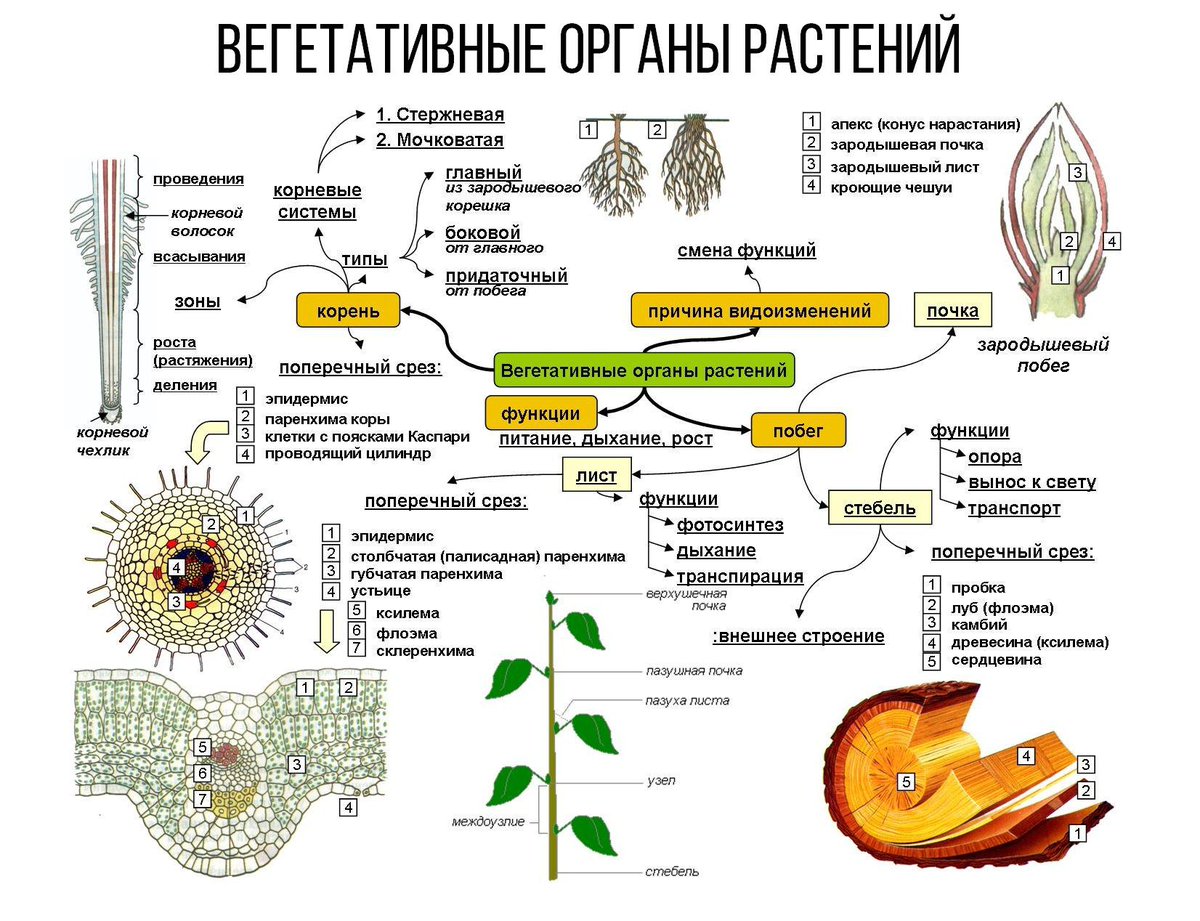
Требуются специалисты по всем группам растений, цветковым, а также менее изученным водорослям, мхам, папоротникам и грибам (включая лишайники). Эта концентрация подходит для студентов, планирующих обучение в аспирантуре в этих областях.Более непосредственные возможности трудоустройства находятся в Службе национальных парков, программах государственного и природного наследия и исчезающих растений, частных консалтинговых фирмах, природоохранных организациях, ботанических садах и гербариях. Систематическая биология — это хороший способ потворствовать желанию путешествовать, заниматься наукой и вносить свой вклад в общество одновременно.
Студенты, выполнившие требования Концентрации по биологии растений, получат отметку в своем официальном транскрипте.
Районный советник
| Районный консультант | Районный факультет |
|---|---|
Др. Пол Манос, кафедра биологии Пол Манос, кафедра биологии телефон: 660-7358 электронная почта: [email protected] | Бенфей, Бернхардт, Чен, Донг, Донохью, Луцони, Манос, Митчелл-Олдс, Пей, Прайер, Раушер, Шоу, Сан, Вилгалис, Уиллис, Райт
|
Предварительные условия
Химия
| Для B.S. Степень | Для Б.А. Степень |
|---|---|
ПРИМЕЧАНИЕ: CHEM 210DL и CHEM 202L рекомендуются для студентов, изучающих медицинские, ветеринарные, биохимические и фармацевтические специальности.
Математика
| Для B.S. Степень | Для Б.А. Степень |
|---|---|
|
Физика
Для Б. С. Степень С. Степень | Для Б.А. Степень |
|---|---|
ПРИМЕЧАНИЕ. PHYSICS 142L рекомендуется для студентов, изучающих медицинские, ветеринарные, биохимические и фармацевтические специальности.
Основные требования по биологии (минимум 10 курсов)
Следующие курсы обязательны для студентов, изучающих как степень бакалавра, так и степень бакалавра наук. и Б.А. документы по биологии.
Курсы Gateway (2 или 1 курс)
Шлюзы не упорядочены и могут быть выбраны в любом порядке.
ИЛИ
Требования к области биологии растений (3 курса)
Выберите 1 курс из КАЖДОЙ из следующих трех областей. Курсы, перечисленные более чем в одной области, могут использоваться только для удовлетворения требований одной области.
Курсы, перечисленные более чем в одной области, могут использоваться только для удовлетворения требований одной области.
Факультативы (5 или 6 курсов)
Максимум 2 независимых исследования или урока могут быть засчитаны для получения диплома.
Требования к опыту работы в лаборатории
В рамках вашего региона и факультативных курсов вы должны пройти как минимум 2 полных лабораторных курса в дополнение к базовым курсам. Курсы должны быть на уровне 200 или выше. Лабораторным курсом может считаться не более 1 независимого исследования.
Курсы должны быть на уровне 200 или выше. Лабораторным курсом может считаться не более 1 независимого исследования.
Требование к курсу Capstone
Вы должны пройти как минимум 1 полный курс биологии «Capstone» уровня 400 или выше или другой утвержденный курс Capstone. Самостоятельное обучение может засчитываться в соответствии с этим требованием, если оно является продолжением второго семестра.
Развитие растений. Молекулярная биология клетки
Растения и животные разделены примерно 1,5 миллиардами лет эволюционной истории.Они развили свою многоклеточную организацию независимо, но с использованием одного и того же исходного набора инструментов — набора генов, унаследованных от их общего одноклеточного эукариотического предка. Большинство контрастов в их стратегиях развития проистекают из двух основных особенностей растений. Во-первых, они получают энергию от солнечного света, а не от других организмов. Это диктует строение тела, отличное от животного. Во-вторых, их клетки заключены в полужесткие клеточные стенки и сцементированы вместе, что не позволяет им двигаться, как клетки животных.Это диктует иной набор механизмов формирования тела и иные процессы развития, чтобы справиться с изменчивой средой.
Во-вторых, их клетки заключены в полужесткие клеточные стенки и сцементированы вместе, что не позволяет им двигаться, как клетки животных.Это диктует иной набор механизмов формирования тела и иные процессы развития, чтобы справиться с изменчивой средой.
Развитие животных в значительной степени защищено от изменений окружающей среды, и эмбрион создает ту же самую генетически детерминированную структуру тела, на которую не влияют внешние условия. Развитие большинства растений, напротив, сильно зависит от окружающей среды. Поскольку они не могут соответствовать окружающей среде, перемещаясь с места на место, растения приспосабливаются вместо этого, изменяя ход своего развития.Их стратегия оппортунистична. Определенный тип органа — скажем, лист, цветок или корень — может образоваться из оплодотворенной яйцеклетки многими различными путями в соответствии с сигналами окружающей среды. Лист бегонии, привязанный к земле, может пустить корень; корень может выбросить побег; на побеге при солнечном свете могут вырасти листья и цветы.
Зрелое растение обычно состоит из множества копий небольшого набора стандартизированных модулей, как описано в разделе . Позиции и время, в которые генерируются эти модули, сильно зависят от окружающей среды, что приводит к изменению общей структуры завода.Выбор между альтернативными модулями и их организация в целое растение зависят от внешних сигналов и долгосрочных гормональных сигналов, которые играют гораздо меньшую роль в контроле развития животных.
Рисунок 21-106
Простой пример модульной конструкции установок. Каждый модуль (показан разными оттенками зеленого цвета ) состоит из стебля, листа и почки, содержащей потенциальный центр роста, или меристему . Почка образуется в месте разветвления или узла, там, где лист (подробнее…)
Но хотя глобальная структура растения — структура его корней или ветвей, количество листьев или цветков — может сильно различаться, его детальная организация в малом масштабе — нет. Лист, цветок или даже ранний зародыш растения так же точно определены, как и любой орган животного, обладающий детерминированной структурой, в отличие от неопределенной схемы ветвления и прорастания растения в целом. Внутренняя организация растительного модуля ставит по существу те же проблемы генетического контроля за формированием паттерна, что и развитие животных, и решаются они аналогичным образом.В этом разделе мы сосредоточимся на клеточных механизмах развития цветковых растений. Мы исследуем как контрасты, так и сходства с животными.
Внутренняя организация растительного модуля ставит по существу те же проблемы генетического контроля за формированием паттерна, что и развитие животных, и решаются они аналогичным образом.В этом разделе мы сосредоточимся на клеточных механизмах развития цветковых растений. Мы исследуем как контрасты, так и сходства с животными.
Arabidopsis служит модельным организмом для молекулярной генетики растений
Цветковые растения, несмотря на их удивительное разнообразие, возникли сравнительно недавно. Возраст самых ранних известных окаменелостей составляет 125 миллионов лет, по сравнению с 350 миллионами лет и более для позвоночных животных. Таким образом, в основе разнообразия форм лежит высокая степень сходства молекулярных механизмов.Как мы увидим, небольшое генетическое изменение может трансформировать крупномасштабную структуру растения; и точно так же, как физиология растений позволяет выживать во многих различных средах, она также позволяет выживать множеству форм с различной структурой. Мутация, дающая животному две головы, обычно смертельна; тот, который удваивает количество цветов или ветвей на растении, как правило, не является.
Мутация, дающая животному две головы, обычно смертельна; тот, который удваивает количество цветов или ветвей на растении, как правило, не является.
Чтобы идентифицировать гены, управляющие развитием растений, и выяснить, как они функционируют, биологи растений выбрали небольшой сорняк, кресс-салат обыкновенный Arabidopsis thaliana (), в качестве основного модельного организма.Как и Drosophila или Caenorhabditis elegans , он маленький, быстро размножается и удобен для генетики. Его можно выращивать в помещении в чашках Петри или крошечных горшках для растений в больших количествах, и через 8-10 недель оно дает сотни семян на растение. Он имеет, как и C. elegans , значительное преимущество перед Drosophila или позвоночными животными с точки зрения генетики: как и многие цветковые растения, он может размножаться как гермафродит, потому что один цветок производит как яйца, так и мужские гаметы, которые могут оплодотворяться. их.Следовательно, когда цветок, гетерозиготный по рецессивной летальной мутации, самоопыляется, четверть его семян будет иметь гомозиготный эмбриональный фенотип. Это позволяет легко проводить генетический скрининг () и, таким образом, получать каталог генов, необходимых для определенных процессов развития.
Это позволяет легко проводить генетический скрининг () и, таким образом, получать каталог генов, необходимых для определенных процессов развития.
Рисунок 21-107
Arabidopsis thaliana. Это небольшое растение относится к семейству горчичных (или крестоцветных) (см. также рис. 1-47). Это сорняк, не имеющий хозяйственного значения, но представляющий большую ценность для генетических исследований развития растений.(Из MA Estelle and CR Somerville, Trends (подробнее…)
Рисунок 21-108
Получение мутантов Arabidopsis. Семя, содержащее многоклеточный зародыш, обрабатывают химическим мутагеном и оставляют для вырасти в растение.В общем, это растение будет представлять собой мозаику клонов клеток, несущих различные индуцированные мутации.Особь (подробнее…)
Геном
Arabidopsis богат генами контроля развития Arabidopsis имеет один из самых маленьких геномов растений — 125 миллионов пар нуклеотидов, наравне с C.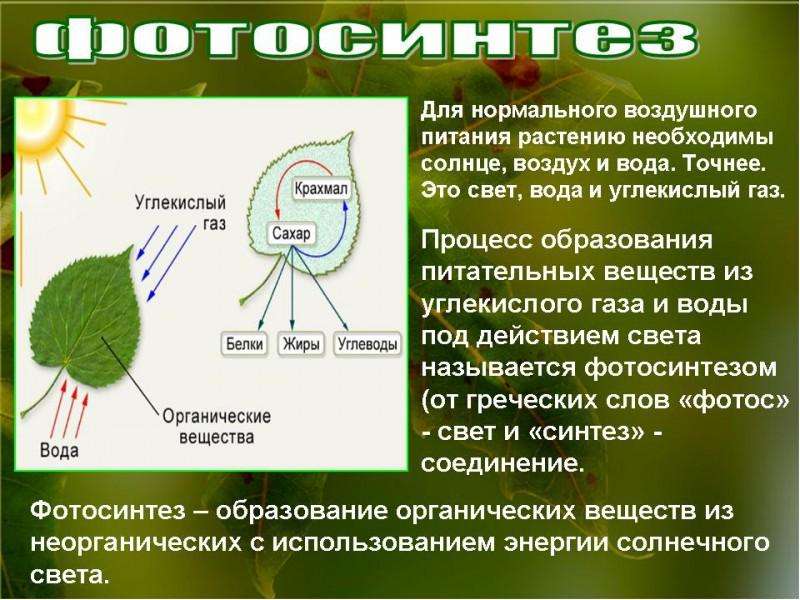 elegans и Drosophila — теперь известна полная последовательность ДНК. Он содержит около 26 000 генов. Однако это общее количество включает множество недавно созданных дубликатов, так что количество представленных функционально различных типов белков может быть значительно меньше. Были созданы методы культивирования клеток и генетической трансформации, а также обширные библиотеки семян, несущих мутации, полученные путем случайных вставок мобильных генетических элементов, так что растения с мутациями в любом выбранном гене могут быть получены на заказ.Таким образом, доступны мощные инструменты для анализа функций генов. Хотя пока экспериментально охарактеризована лишь небольшая часть общего набора генов, функции могут быть предварительно отнесены ко многим генам (около 18 000) на основе сходства их последовательностей с хорошо охарактеризованными генами Arabidopsis и других организмов.
elegans и Drosophila — теперь известна полная последовательность ДНК. Он содержит около 26 000 генов. Однако это общее количество включает множество недавно созданных дубликатов, так что количество представленных функционально различных типов белков может быть значительно меньше. Были созданы методы культивирования клеток и генетической трансформации, а также обширные библиотеки семян, несущих мутации, полученные путем случайных вставок мобильных генетических элементов, так что растения с мутациями в любом выбранном гене могут быть получены на заказ.Таким образом, доступны мощные инструменты для анализа функций генов. Хотя пока экспериментально охарактеризована лишь небольшая часть общего набора генов, функции могут быть предварительно отнесены ко многим генам (около 18 000) на основе сходства их последовательностей с хорошо охарактеризованными генами Arabidopsis и других организмов.
Даже больше, чем геномы многоклеточных животных, геном Arabidopsis богат генами, кодирующими белки, регулирующие гены (). Некоторые основные семейства белков, регулирующих гены животных (такие как семейство ДНК-связывающих белков Myb), в значительной степени расширены, в то время как другие (например, ядерные рецепторы гормонов), по-видимому, полностью отсутствуют, и существуют большие семейства белков, регулирующих гены. растения, не имеющие животных гомологов.
Некоторые основные семейства белков, регулирующих гены животных (такие как семейство ДНК-связывающих белков Myb), в значительной степени расширены, в то время как другие (например, ядерные рецепторы гормонов), по-видимому, полностью отсутствуют, и существуют большие семейства белков, регулирующих гены. растения, не имеющие животных гомологов.
Таблица 21-2
Некоторые основные семейства генов-регуляторных белков у арабидопсиса, дрозофилы, C. elegans и дрожжей Saccharomyces cerevisiae .
Хотя гомологичные белки, регулирующие гены (такие как гомеодоменовые белки), могут быть обнаружены как у растений, так и у животных, они имеют мало общего в отношении генов, которые они регулируют, или типов решений, связанных с развитием, которые они контролируют, и очень мало консервативных белковой последовательности вне ДНК-связывающих доменов.
Arabidopsis , подобно многоклеточным животным, обладает многими генами для клеточной коммуникации и передачи сигналов (1900 генов из 18000 классифицированных), но конкретные детали этих наборов генов сильно различаются, как обсуждалось в главе 15. Wnt, Hedgehog , Notch и механизмы передачи сигналов TGFβ отсутствуют у Arabidopsis . В качестве компенсации сильно развиты другие сигнальные пути, свойственные растениям. Рецепторы клеточной поверхности класса тирозинкиназ, по-видимому, полностью отсутствуют, хотя многие из сигнальных компонентов ниже этих рецепторов у животных присутствуют.Наоборот, рецепторов класса серин/треонинкиназ очень много, но они не действуют через ту же систему внутриклеточных мессенджеров, что и рецепторы серин/треонинкиназы у животных. Значительные наборы генов предназначены для процессов развития, имеющих особое значение у растений: например, более 700 для синтеза и ремоделирования клеточной стенки растений и более 100 для обнаружения и реагирования на свет.
Wnt, Hedgehog , Notch и механизмы передачи сигналов TGFβ отсутствуют у Arabidopsis . В качестве компенсации сильно развиты другие сигнальные пути, свойственные растениям. Рецепторы клеточной поверхности класса тирозинкиназ, по-видимому, полностью отсутствуют, хотя многие из сигнальных компонентов ниже этих рецепторов у животных присутствуют.Наоборот, рецепторов класса серин/треонинкиназ очень много, но они не действуют через ту же систему внутриклеточных мессенджеров, что и рецепторы серин/треонинкиназы у животных. Значительные наборы генов предназначены для процессов развития, имеющих особое значение у растений: например, более 700 для синтеза и ремоделирования клеточной стенки растений и более 100 для обнаружения и реагирования на свет.
Теперь мы должны изучить, как гены растения используются для управления его развитием.
Эмбриональное развитие начинается с образования оси отростков и затем останавливается внутри семени
Основная стратегия полового размножения у цветковых растений кратко изложена на Панели 21-1. Оплодотворенная яйцеклетка, или зигота, высшего растения начинает с асимметричного деления, чтобы установить полярность будущего зародыша. Одним из продуктов этого деления является маленькая клетка с плотной цитоплазмой, которая станет собственно зародышем. Другая представляет собой крупную вакуолизированную клетку, которая далее делится и образует структуру, называемую суспензором , которая в некоторых отношениях сравнима с пуповиной у млекопитающих.Подвеска прикрепляет эмбрион к соседней питательной ткани и обеспечивает путь для транспортировки питательных веществ.
Оплодотворенная яйцеклетка, или зигота, высшего растения начинает с асимметричного деления, чтобы установить полярность будущего зародыша. Одним из продуктов этого деления является маленькая клетка с плотной цитоплазмой, которая станет собственно зародышем. Другая представляет собой крупную вакуолизированную клетку, которая далее делится и образует структуру, называемую суспензором , которая в некоторых отношениях сравнима с пуповиной у млекопитающих.Подвеска прикрепляет эмбрион к соседней питательной ткани и обеспечивает путь для транспортировки питательных веществ.
Панель 21-1
Особенности раннего развития цветковых растений.
На следующем этапе развития диплоидная клетка зародыша пролиферирует, образуя клубок клеток, который быстро приобретает поляризованную структуру. Он включает две ключевые группы пролиферирующих клеток: одну на суспензорном конце эмбриона, которая будет сотрудничать с самой верхней суспензорной клеткой для образования корня, и другую на противоположном конце, которая будет генерировать побег (4). Установленная таким образом основная ось «корень-побег» аналогична оси «голова-хвост» животного. При этом становится возможным различать будущие клетки эпидермиса , образующие самый наружный слой зародыша, будущие клетки основной ткани , занимающие большую часть внутренней части, и будущие клетки сосудистой ткани , образующие центральное ядро (панель 21-2). Эти три набора клеток можно сравнить с тремя зародышевыми листками эмбриона животного. Несколько позже в развитии зачаток побега начинает давать зачаточные семенные листочки, или семядолей — одну у однодольных и две у двудольных.Вскоре после этой стадии развитие обычно прекращается, и зародыш упаковывается в семя (корпус, образованный тканями материнского растения), специализированный для расселения и выживания в суровых условиях. Зародыш в семени стабилизируется обезвоживанием и может оставаться в состоянии покоя очень долго — даже сотни лет. При регидратации семена прорастают, и эмбриональное развитие возобновляется.
Установленная таким образом основная ось «корень-побег» аналогична оси «голова-хвост» животного. При этом становится возможным различать будущие клетки эпидермиса , образующие самый наружный слой зародыша, будущие клетки основной ткани , занимающие большую часть внутренней части, и будущие клетки сосудистой ткани , образующие центральное ядро (панель 21-2). Эти три набора клеток можно сравнить с тремя зародышевыми листками эмбриона животного. Несколько позже в развитии зачаток побега начинает давать зачаточные семенные листочки, или семядолей — одну у однодольных и две у двудольных.Вскоре после этой стадии развитие обычно прекращается, и зародыш упаковывается в семя (корпус, образованный тканями материнского растения), специализированный для расселения и выживания в суровых условиях. Зародыш в семени стабилизируется обезвоживанием и может оставаться в состоянии покоя очень долго — даже сотни лет. При регидратации семена прорастают, и эмбриональное развитие возобновляется.
Рисунок 21-109
Две стадии эмбриогенеза у Arabidopsis thaliana. (Из G. Jürgens et al., Development [Suppl.] 1:27–38, 1991. © The Company of Biologist.)
Панель 21-2
Типы клеток и ткани высших растений Построен.
Генетический скрининг можно использовать для Arabidopsis , так же как и для Drosophila или C. elegans , для выявления генов, управляющих организацией эмбриона, и их группировки по категориям в соответствии с их гомозиготными мутантными фенотипами.Часть необходима для образования корня сеянца, часть – для стебля сеянца, часть – для верхушки сеянца с его семядолями. Другой класс необходим для образования трех основных типов тканей — эпидермиса, основной ткани и сосудистой ткани — и еще один класс для организованных изменений формы клеток, которые придают зародышу и проростку их удлиненную форму (1).
Рисунок 21-110
Мутант Саженцы арабидопсиса . Нормальный проросток (A) по сравнению с четырьмя типами мутантов (BE), дефектных в разных частях их апикально-базального рисунка: (B) у него отсутствуют структуры на его верхушке, (C) есть верхушка и корень, но отсутствует стебель между ними. их, (Д) (подробнее…)
их, (Д) (подробнее…)
Части растения образуются последовательно меристемами
Грубо говоря, зародыш насекомого или позвоночного животного представляет собой рудиментарную миниатюрную модель более позднего организма, а детали строения тела постепенно заполняются по мере увеличения. Зародыш растения вырастает во взрослое растение совершенно по-другому: части взрослого растения создаются последовательно группами клеток, которые пролиферируют, образуя дополнительные структуры на периферии растения.Эти важнейшие группы клеток называются апикальными меристемами (см. ). Каждая меристема состоит из самообновляющейся популяции стволовых клеток. Когда они делятся, они оставляют след потомства, которое смещается из области меристемы, увеличивается и, наконец, дифференцируется. Хотя апикальные меристемы побега и корня генерируют все основные разновидности клеток, которые необходимы для построения листьев, корней и стеблей, многие клетки вне апикальных меристем также сохраняют способность к дальнейшей пролиферации и сохраняют меристемный потенциал. Таким образом, деревья и другие многолетние растения, например, могут с годами увеличивать обхват своих стеблей и корней и могут дать новые побеги из спящих участков, если растение повреждено.
Таким образом, деревья и другие многолетние растения, например, могут с годами увеличивать обхват своих стеблей и корней и могут дать новые побеги из спящих участков, если растение повреждено.
Зачатки апикальных меристем корня и побега определяются уже у зародыша. Как только семенная кожура разрывается во время прорастания, происходит резкое увеличение немеристематических клеток, приводящее к прорастанию сначала корня, который сразу же закрепляется в почве, а затем ростка.Это сопровождается быстрыми и непрерывными клеточными делениями в апикальных меристемах: например, в апикальной меристеме корня кукурузы клетки делятся каждые 12 часов, производя 5·10 5 клеток в день. Быстро растущие корень и побег исследуют окружающую среду: корень увеличивает способность растения поглощать воду и минеральные вещества из почвы, побег увеличивает его способность к фотосинтезу (см. рис. 21-1).
Рисунок 21-111
Саженец арабидопсиса . коричневый объект справа молодого сеянца — это две половинки сброшенной семенной кожуры. (С любезного разрешения Кэтрин Дакетт.)
(С любезного разрешения Кэтрин Дакетт.)
Развитие проростка зависит от сигналов окружающей среды
Начиная с момента прорастания, ход развития растения находится под сильным влиянием сигналов из окружающей среды. Побег должен быстро пробиться сквозь почву, открыть свои семядоли и начать фотосинтез только после того, как достигнет света.Время этого перехода от быстрого подземного прорастания к световому росту не может быть запрограммировано генетически, потому что невозможно предсказать глубину, на которую закапывают семя. Вместо этого переключатель развития управляется светом, который, помимо прочего, воздействует на проросток, подавляя выработку класса растительных гормонов, называемых брассиностероидами , обсуждаемыми в главе 15. Мутации в генах, необходимых для производства или приема брассиностероидного сигнала. вызывают позеленение стебля сеянца, замедляют его удлинение и преждевременно раскрывают семядоли, пока он еще находится в темноте.
Формирование каждой новой структуры зависит от направленного клеточного деления и расширения
Растительные клетки, заключенные в свои клеточные стенки, не могут ползать и не могут перетасовываться по мере роста растения; но они могут делиться, набухать, растягиваться и изгибаться. Таким образом, морфогенез развивающегося растения зависит от упорядоченных клеточных делений, за которыми следует строго ориентированное расширение клеток. Например, большинство клеток, образующихся в меристеме кончика корня, проходят три различные фазы развития: деление, рост (удлинение) и дифференцировку.Эти три шага, которые перекрываются как в пространстве, так и во времени, приводят к характерной архитектуре кончика корня. Хотя процесс клеточной дифференцировки часто начинается, когда клетка еще увеличивается в размерах, в кончике корня сравнительно легко различить зону клеточного деления, зону ориентированного удлинения клеток (за счет чего происходит увеличение длины корня), и зона дифференцировки клеток ().
Таким образом, морфогенез развивающегося растения зависит от упорядоченных клеточных делений, за которыми следует строго ориентированное расширение клеток. Например, большинство клеток, образующихся в меристеме кончика корня, проходят три различные фазы развития: деление, рост (удлинение) и дифференцировку.Эти три шага, которые перекрываются как в пространстве, так и во времени, приводят к характерной архитектуре кончика корня. Хотя процесс клеточной дифференцировки часто начинается, когда клетка еще увеличивается в размерах, в кончике корня сравнительно легко различить зону клеточного деления, зону ориентированного удлинения клеток (за счет чего происходит увеличение длины корня), и зона дифференцировки клеток ().
Рисунок 21-112
Растущий кончик корня. (A) Организация последних 2 мм растущего кончика корня.Указаны примерные зоны, в которых можно обнаружить делящиеся, удлиняющиеся и дифференцирующиеся клетки. (B) Апикальная меристема и корневой чехлик кончика корня кукурузы, показывающий (подробнее. ..)
..)
В фазе контролируемого расширения, которая обычно следует за делением клеток, дочерние клетки часто могут увеличиваться в объеме в 50 или более раз. более. Это расширение обусловлено осмотическим тургорным давлением, которое давит наружу на клеточную стенку растения, а его направление определяется ориентацией фибрилл целлюлозы в клеточной стенке, которые сдерживают расширение вдоль одной оси (см. ).Ориентация целлюлозы, в свою очередь, по-видимому, контролируется ориентацией массивов микротрубочек непосредственно внутри плазматической мембраны, которые, как считается, направляют отложение целлюлозы (обсуждается в главе 19). Эти ориентации могут быть быстро изменены регуляторами роста растений, такими как этилен и гибберелловая кислота, но молекулярные механизмы, лежащие в основе этих драматических перестроек цитоскелета, до сих пор неизвестны.
Рисунок 21-113
Различные эффекты регуляторов роста растений этилена и гибберелловой кислоты.Эти регуляторы оказывают быстрое и противоположное влияние на ориентацию массива кортикальных микротрубочек в клетках молодых побегов гороха. Типичная клетка в обработанном этиленом (подробнее…)
Типичная клетка в обработанном этиленом (подробнее…)
Каждый растительный модуль вырастает из микроскопического набора зачатков в меристеме
Апикальные меристемы самовоспроизводятся: в многолетнем растении они продолжают выполнять свои функции на неопределенный срок, пока выживает растение, и они несут ответственность за его непрерывный рост и развитие. Но апикальные меристемы также дают начало второму типу выростов, развитие которых строго ограничено и завершается образованием структуры типа листа или цветка с определенными размерами и формой и коротким сроком жизни.Так, по мере удлинения вегетативного (нецветущего) побега его верхушечная меристема откладывает за собой упорядоченную последовательность из узлов, в которых выросли листья, и междоузлий, (члеников стебля). Таким образом, непрерывная деятельность меристемы производит все большее число подобных модулей, каждый из которых состоит из стебля, листа и почки (см.). Модули соединены друг с другом поддерживающей и транспортной тканью, а последовательные модули точно расположены относительно друг друга, создавая структуру с повторяющимся рисунком. Этот итеративный способ развития характерен для растений и наблюдается во многих других структурах, помимо системы стебель-лист.
Этот итеративный способ развития характерен для растений и наблюдается во многих других структурах, помимо системы стебель-лист.
Рисунок 21-114
Повторяющиеся узоры у растений. Точное размещение последовательных модулей из одной апикальной меристемы создает эти сложные, но регулярные узоры в листьях (А), цветках (В) и плодах (С). (A, from John Sibthorp, Flora Graeca. London: R. Taylor, 1806–1840; (подробнее…)
Хотя последний модуль большой, его организация, как и у зародыша животного, сначала намечена. в микроскопическом масштабе.На вершине побега, на расстоянии миллиметра или меньше, можно обнаружить небольшой низкий центральный купол, окруженный набором отчетливых вздутий на разных стадиях увеличения (). Центральный купол — это сама апикальная меристема; каждое из окружающих вздутий является зачатком листа. Таким образом, эта небольшая область содержит уже отчетливые зачатки нескольких целых модулей. Благодаря четко определенной программе клеточной пролиферации и увеличения клеток каждый зачаток листа и прилегающие к нему клетки будут расти, образуя лист, узел и междоузлие. Тем временем сама апикальная меристема будет давать начало новым листовым зачаткам, чтобы генерировать все больше и больше модулей в потенциально бесконечной последовательности. Таким образом, последовательная организация модулей растения контролируется событиями на верхушке побега. Именно система локальных сигналов внутри этой крошечной области определяет рисунок зачатков — положение одного зачатка листа относительно другого, расстояние между ними и их положение относительно самой апикальной меристемы.
Тем временем сама апикальная меристема будет давать начало новым листовым зачаткам, чтобы генерировать все больше и больше модулей в потенциально бесконечной последовательности. Таким образом, последовательная организация модулей растения контролируется событиями на верхушке побега. Именно система локальных сигналов внутри этой крошечной области определяет рисунок зачатков — положение одного зачатка листа относительно другого, расстояние между ними и их положение относительно самой апикальной меристемы.
Рисунок 21-115
Верхушка побега молодого растения табака.(A) Сканирующая электронная микрофотография показывает верхушку побега с двумя последовательно появляющимися листовыми зачатками, которые здесь видны как боковые вздутия по обе стороны от куполообразной апикальной меристемы. (B) Тонкий срез аналогичного (подробнее…)
Вариации на эту основную повторяющуюся тему могут привести к более сложной архитектуре, включая такие структуры, как усики, шипы, ветки и цветы.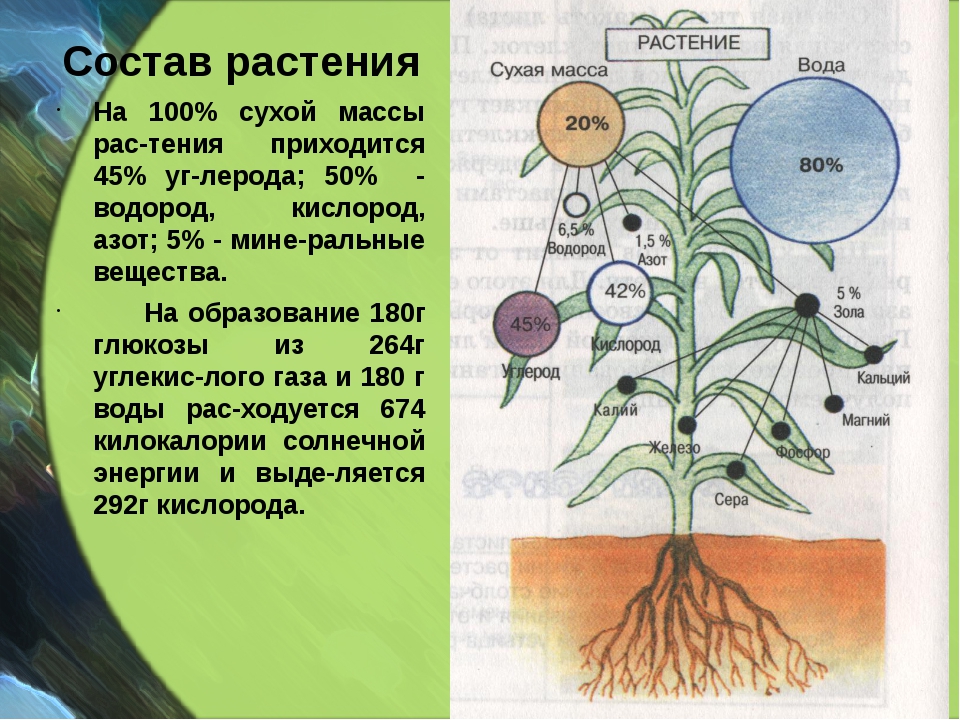 Таким образом, включив разные наборы генов на верхушке побега, растение может производить разные типы зачатков в разных пространственных структурах.
Таким образом, включив разные наборы генов на верхушке побега, растение может производить разные типы зачатков в разных пространственных структурах.
Передача сигналов клетками поддерживает меристему
Центральным во всех этих явлениях является вопрос о том, как апикальная меристема поддерживает себя. Клетки меристемы должны продолжать размножаться в течение недель, лет или даже столетий по мере роста растения, заменяя себя и постоянно производя потомство, которое дифференцируется. При всем этом размер скопления клеток, составляющих меристему, остается практически постоянным (около 100 клеток у Arabidopsis , например).Новые меристемы могут возникать в виде ветвей растений, но и они сохраняют прежний размер.
Генетические скрининги выявили гены, необходимые для поддержания меристемы. Например, мутации, нарушающие работу гена WUSCHEL , кодирующего гомеодоменовый белок, превращают апикальную меристему в немеристематическую ткань, так что проросток не дает прорастания. Наоборот, мутации в группе генов CLAVATA , кодирующих компоненты межклеточного сигнального пути (см. ), делают меристему аномально большой.Эти гены экспрессируются в разных слоях клеток в области меристемы (). Два самых поверхностных слоя клеток, называемые слоями L1 и L2, вместе с самой верхней частью слоя L3 содержат клетки собственно меристемы, способные неограниченно делиться, чтобы дать начало будущим частям растения. Меристематические клетки слоев L1 и L2 экспрессируют Clavata3, небольшой секретируемый сигнальный белок. Чуть ниже, в слое L3, находится группа клеток, экспрессирующих Clavata1 (рецептор для Clavata3).В центре этого пластыря Clavata1 находятся клетки, которые экспрессируют регуляторный белок гена Wuschel.
Наоборот, мутации в группе генов CLAVATA , кодирующих компоненты межклеточного сигнального пути (см. ), делают меристему аномально большой.Эти гены экспрессируются в разных слоях клеток в области меристемы (). Два самых поверхностных слоя клеток, называемые слоями L1 и L2, вместе с самой верхней частью слоя L3 содержат клетки собственно меристемы, способные неограниченно делиться, чтобы дать начало будущим частям растения. Меристематические клетки слоев L1 и L2 экспрессируют Clavata3, небольшой секретируемый сигнальный белок. Чуть ниже, в слое L3, находится группа клеток, экспрессирующих Clavata1 (рецептор для Clavata3).В центре этого пластыря Clavata1 находятся клетки, которые экспрессируют регуляторный белок гена Wuschel.
Рисунок 21-116
Петли обратной связи, которые, как считается, поддерживают апикальную меристему побега. (A) Расположение клеточных слоев, составляющих апикальную меристему побега. (B) Схема межклеточной коммуникации, которая поддерживает меристему. Искусственная сверхэкспрессия Wuschel (подробнее…)
Искусственная сверхэкспрессия Wuschel (подробнее…)
Характер клеточных делений подразумевает, что клетки, экспрессирующие Wuschel, сами по себе не являются частью меристемы; новые клетки, экспрессирующие Wuschel, по-видимому, постоянно рекрутируются из меристематической части популяции L3, прямо над доменом Wuschel.Тем не менее, Wuschel-экспрессирующие клетки находятся в центре механизма, поддерживающего меристему. Сигнал, который они продуцируют, поддерживает меристематическое поведение в указанных выше клетках, стимулирует экспрессию генов CLAVATA и, предположительно, заставляет новые клетки, рекрутированные в домен Wuschel, переключаться на Wuschel. Отрицательная обратная связь от верхних меристематических клеток, доставляемая сигнальным путем Clavata, воздействует обратно на нижние области, ограничивая размер домена Вушеля, тем самым предотвращая чрезмерное увеличение меристемы (4).
Этот отчет о меристеме растений все еще неясен во многих деталях, и в него вовлечены и другие гены, кроме тех, которые мы упомянули. Тем не менее, математическое моделирование показывает, что системы подобного рода, основанные на петле обратной связи, включающей короткодействующий активирующий сигнал и дальнодействующий тормозной сигнал, могут стабильно поддерживать сигнальный центр четко определенного размера даже в условиях непрерывной пролиферации. и оборот клеток, образующих этот центр. Считают, что аналогичные системы сигналов функционируют в процессе развития животных для поддержания локализованных сигнальных центров, таких как Организатор гаструлы земноводных или зона поляризующей активности в зачатке конечности.
Тем не менее, математическое моделирование показывает, что системы подобного рода, основанные на петле обратной связи, включающей короткодействующий активирующий сигнал и дальнодействующий тормозной сигнал, могут стабильно поддерживать сигнальный центр четко определенного размера даже в условиях непрерывной пролиферации. и оборот клеток, образующих этот центр. Считают, что аналогичные системы сигналов функционируют в процессе развития животных для поддержания локализованных сигнальных центров, таких как Организатор гаструлы земноводных или зона поляризующей активности в зачатке конечности.
До сих пор неизвестно, как клетки, экспрессирующие Wuschel, передают сигнал своим соседям. Одна возможность состоит в том, что сам белок Wuschel диффундирует непосредственно из клетки в клетку через плазмодесмы — сигнальный путь, свойственный растениям. Было показано, что некоторые другие белки, регулирующие гены, путешествуют таким же образом в меристемах, распространяясь из клеток, содержащих соответствующую мРНК, в соседние клетки, которые ее не содержат.
Регуляторные мутации могут трансформировать топологию растения путем изменения поведения клеток в меристеме
Если стебель должен разветвляться, должны быть созданы новые апикальные меристемы побега, и это также зависит от событий, происходящих вблизи верхушки побега.В каждом развивающемся узле в остром углу (пазухе) между зачатком листа и стеблем образуется почка (). Он содержит гнездо клеток, полученных из апикальной меристемы, которые сохраняют меристематический характер. Они способны стать апикальной меристемой новой ветви или зачатком такой структуры, как цветок; но у них также есть альтернативный вариант оставаться в состоянии покоя как пазушные почки . Схема ветвления растения регулируется этим выбором судьбы, и влияющие на нее мутации могут трансформировать структуру растения.Кукуруза является прекрасным примером.
Рисунок 21-117
Пазушные почки вблизи верхушки побега. На фотографии показан продольный срез Coleus blumei, обыкновенного комнатного растения.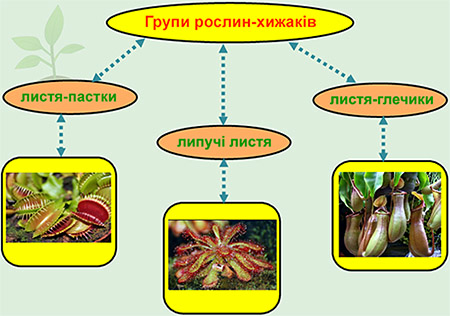 (Из PH Raven, RF Evert и SE Eichhorn, Biology of Plants, 6th edn. New York: Freeman/Worth, 1999, используется с (подробнее…)
(Из PH Raven, RF Evert и SE Eichhorn, Biology of Plants, 6th edn. New York: Freeman/Worth, 1999, используется с (подробнее…)
Кукуруза представляет собой одно из самых замечательных достижений человечества в области генной инженерии. Коренные американцы создали его путем селекционного размножения в течение нескольких столетий или, возможно, тысячелетий между 5000 и 10000 лет назад.Они начались с дикой травы, известной как теосинте, с сильно разветвленными лиственными стеблями и крошечными колосьями с несъедобными твердыми зернами. Детальный генетический анализ выявил несколько генетических локусов — около пяти — как участки мутаций, которые объясняют большую часть различий между этим бесперспективным предком и современной кукурузой. Один из этих локусов с особенно впечатляющим эффектом соответствует гену, называемому teosinte Branched-1 ( tb1 ). У кукурузы с мутациями с потерей функции в tb1 обычный простой неразветвленный стебель с несколькими крупными листьями с промежутками вдоль него превращается в плотную, ветвящуюся листовую массу, напоминающую теосинте (). Паттерн ветвления у мутанта подразумевает, что пазушные почки, возникающие в нормальном положении, избежали ингибирования, которое мешает им у нормальной кукурузы вырасти в ветви.
Паттерн ветвления у мутанта подразумевает, что пазушные почки, возникающие в нормальном положении, избежали ингибирования, которое мешает им у нормальной кукурузы вырасти в ветви.
Рисунок 21-118
Преобразование архитектуры растений в результате мутации: сравнение теосинте, нормальной кукурузы и tb1 — дефектной кукурузы. (A) Фотографии трех видов растений. (B) Архитектура теосинта, нормальной кукурузы и кукурузы с дефектом tb1 — схематически сравнивается.(подробнее…)
У обычной кукурузы единственный стебель увенчан кисточкой — мужским цветком, а несколько пазушных почек вдоль стебля развиваются в женские цветки и после оплодотворения образуют початки, которые мы едим. У мутантной кукурузы с дефектным геном tb1 эти плодоносные пазушные почки трансформируются в ветви с кистями. Дикое растение teosinte похоже на tb1 -дефектную кукурузу своим облиственным, сильно разветвленным видом, но в отличие от этого мутанта оно делает уши на многих боковых ветвях, как если бы tb1 был активен. Анализ ДНК дает объяснение. И теосинте, и нормальная кукуруза обладают функциональным геном tb1 с почти идентичной кодирующей последовательностью, но у кукурузы регуляторная область претерпела мутацию, которая повышает уровень экспрессии гена. Таким образом, у нормальной кукурузы ген экспрессируется на высоком уровне в каждой пазушной почке, подавляя образование ветвей, в то время как у теосинте экспрессия во многих пазушных почках низка, так что образование ветвей разрешено.
Анализ ДНК дает объяснение. И теосинте, и нормальная кукуруза обладают функциональным геном tb1 с почти идентичной кодирующей последовательностью, но у кукурузы регуляторная область претерпела мутацию, которая повышает уровень экспрессии гена. Таким образом, у нормальной кукурузы ген экспрессируется на высоком уровне в каждой пазушной почке, подавляя образование ветвей, в то время как у теосинте экспрессия во многих пазушных почках низка, так что образование ветвей разрешено.
Этот пример показывает, как простые мутации, меняющие поведение мери-стволовых клеток, могут трансформировать структуру растений — принцип огромной важности в селекции пищевых растений.В более общем случае случай tb1 иллюстрирует, как новые формы тела, будь то растения или животные, могут развиваться посредством изменений в регуляторной ДНК без изменения свойств образующихся белков.
Гормональные сигналы дальнего действия координируют события развития в отдельных частях растения
Судьба пазушной почки определяется не только ее генами, но и условиями окружающей среды. Отдельные части растения находятся в разных условиях и индивидуально реагируют на них изменением способа своего развития.Однако завод должен продолжать функционировать как единое целое. Это требует, чтобы выбор развития и события в одной части растения влияли на выбор развития в другом месте. Для такой координации должны быть сигналы дальнего действия.
Отдельные части растения находятся в разных условиях и индивидуально реагируют на них изменением способа своего развития.Однако завод должен продолжать функционировать как единое целое. Это требует, чтобы выбор развития и события в одной части растения влияли на выбор развития в другом месте. Для такой координации должны быть сигналы дальнего действия.
Как известно садовникам, например, отщипыванием кончика ветки можно стимулировать боковой рост: удаление апикальной меристемы избавляет от торможения покоящиеся пазушные меристемы и позволяет им образовывать новые побеги. В этом случае был идентифицирован дальнодействующий сигнал от апикальной меристемы или, по крайней мере, ключевой компонент.Это ауксин, член одного из шести известных классов регуляторов роста растений (иногда называемых растительными гормонами ), каждый из которых оказывает сильное влияние на развитие растений. Пятью другими известными классами являются гиббереллины , цитокинины , абсцизовая кислота , газ этилен и брассиностероиды . Как показано на рисунке, все они представляют собой небольшие молекулы, которые легко проникают через клеточные стенки. Все они синтезируются большинством растительных клеток и могут либо действовать локально, либо транспортироваться для воздействия на клетки-мишени на расстоянии.Ауксин, например, транспортируется от клетки к клетке со скоростью около 1 см в час от кончика побега к его основанию. Каждый регулятор роста имеет несколько эффектов, и они модулируются другими регуляторами роста, а также сигналами окружающей среды и состоянием питания. Таким образом, ауксин сам по себе может способствовать корнеобразованию, но в сочетании с гиббереллином он может способствовать удлинению стебля, с цитокинином ауксин может подавлять рост боковых побегов, а с этиленом он может стимулировать рост боковых корней.Рецепторы, распознающие некоторые из этих регуляторов роста, обсуждаются в главе 15.
Как показано на рисунке, все они представляют собой небольшие молекулы, которые легко проникают через клеточные стенки. Все они синтезируются большинством растительных клеток и могут либо действовать локально, либо транспортироваться для воздействия на клетки-мишени на расстоянии.Ауксин, например, транспортируется от клетки к клетке со скоростью около 1 см в час от кончика побега к его основанию. Каждый регулятор роста имеет несколько эффектов, и они модулируются другими регуляторами роста, а также сигналами окружающей среды и состоянием питания. Таким образом, ауксин сам по себе может способствовать корнеобразованию, но в сочетании с гиббереллином он может способствовать удлинению стебля, с цитокинином ауксин может подавлять рост боковых побегов, а с этиленом он может стимулировать рост боковых корней.Рецепторы, распознающие некоторые из этих регуляторов роста, обсуждаются в главе 15.
Рисунок 21-119
Регуляторы роста растений. Показана формула одной встречающейся в природе репрезентативной молекулы из каждой из шести групп молекул, регулирующих рост растений.
Гомеотические селекторные гены определяют части цветка
Меристемы сталкиваются с другими вариантами развития, помимо выбора между состоянием покоя и ростом, как мы уже видели в нашем обсуждении кукурузы, и они также часто регулируются окружающей средой.Наиболее важным является решение сформировать цветок ().
Рисунок 21-120
Строение цветка Arabidopsis . (Фотограф. (Б) Рисунки. (C) Схематический вид в поперечном сечении. Основной план, как показано на (С), является общим для большинства цветковых двудольных растений. (А, любезно предоставлено Лесли Зибуртом.)
Переход от меристематического роста к формированию цветка обычно вызывается светом. Благодаря плохо изученным механизмам, основанным на поглощении света фитохромными и криптохромными белками (обсуждаемыми в главе 15), растение может очень точно ощущать изменение продолжительности дня.Он отвечает включением экспрессии набора из идентичности цветочной меристемы генов в апикальной меристеме.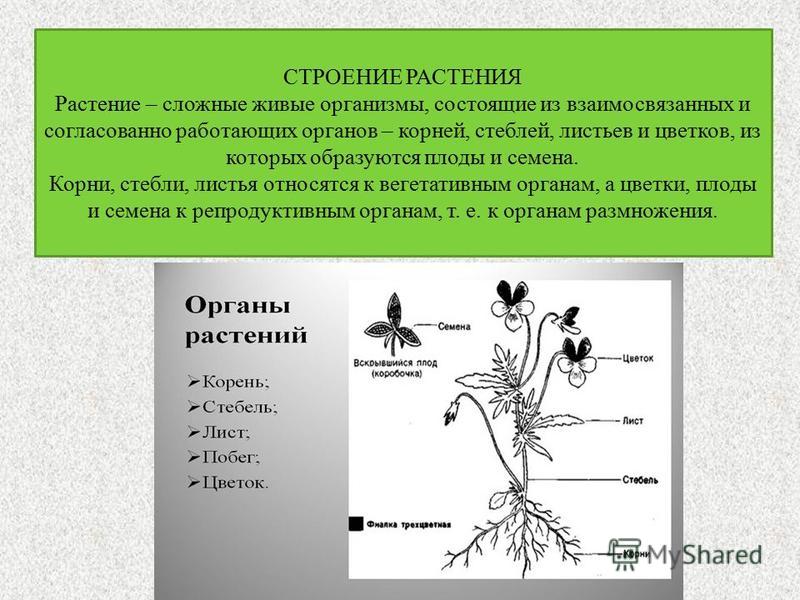 Включая эти гены, апикальная меристема отказывается от своих шансов на продолжение вегетативного роста и делает ставку на производство гамет. Его клетки выполняют строго конечную программу роста и дифференциации: путем модификации обычных механизмов образования листьев в точном порядке образуются ряды мутовок специализированных придатков — обычно сначала чашелистики, затем лепестки, затем тычинки, несущие пыльники, содержащие пыльцу и, наконец, плодолистики, содержащие яйца (см. рис. 21-1).К концу этого процесса меристема исчезла, но в своем потомстве создала половые клетки.
Включая эти гены, апикальная меристема отказывается от своих шансов на продолжение вегетативного роста и делает ставку на производство гамет. Его клетки выполняют строго конечную программу роста и дифференциации: путем модификации обычных механизмов образования листьев в точном порядке образуются ряды мутовок специализированных придатков — обычно сначала чашелистики, затем лепестки, затем тычинки, несущие пыльники, содержащие пыльцу и, наконец, плодолистики, содержащие яйца (см. рис. 21-1).К концу этого процесса меристема исчезла, но в своем потомстве создала половые клетки.
Ряд видоизмененных листьев, образующих цветок, можно сравнить с рядом сегментов тела, образующих муху. У растений, как и у мух, можно обнаружить гомеотические мутации, превращающие одну часть паттерна в характер другой. Мутантные фенотипы могут быть сгруппированы по крайней мере в четыре класса, в которых изменены разные, но перекрывающиеся наборы органов (4). У первого или класса «А», представленного мутантом apetala2 Arabidopsis , два крайних оборота трансформированы: чашелистики преобразованы в плодолистики, а лепестки — в тычинки. У второго или «В» класса, представленного apetala3, , два средних оборота трансформированы: лепестки преобразованы в чашелистики, а тычинки в плодолистики. Третий или «С» класс, представленный агамным, имеет две самые внутренние мутовки, преобразованные с более радикальными последствиями: тычинки превращаются в лепестки, плодолистики отсутствуют, а на их месте ведут себя центральные клетки цветка. как цветочная меристема, которая начинает процесс развития сначала, порождая другой ненормальный набор чашелистиков и лепестков, вложенных в первый и, возможно, еще один, вложенный в него, и так далее до бесконечности.У четвертого класса, мутантов sepallata , три внутренних оборота превратились в чашелистики.
У второго или «В» класса, представленного apetala3, , два средних оборота трансформированы: лепестки преобразованы в чашелистики, а тычинки в плодолистики. Третий или «С» класс, представленный агамным, имеет две самые внутренние мутовки, преобразованные с более радикальными последствиями: тычинки превращаются в лепестки, плодолистики отсутствуют, а на их месте ведут себя центральные клетки цветка. как цветочная меристема, которая начинает процесс развития сначала, порождая другой ненормальный набор чашелистиков и лепестков, вложенных в первый и, возможно, еще один, вложенный в него, и так далее до бесконечности.У четвертого класса, мутантов sepallata , три внутренних оборота превратились в чашелистики.
Рисунок 21-121
Цветки арабидопсиса , показывающие набор гомеозисных мутаций. (A) У apetala2 чашелистика превращаются в плодолистики, а лепестки — в тычинки; (B) У apetala3 лепестка преобразованы в чашелистики, а тычинки в плодолистики; (C) У агамного тычинки преобразованы (подробнее. ..)
..)
Эти фенотипы идентифицируют четыре класса гомеотических селекторных генов, которые, как и гомеозисные селекторные гены дрозофилы , все кодируют регуляторные белки генов.Они экспрессируются в разных доменах и определяют различия в состоянии клеток, которые придают разным частям нормального цветка разные характеристики, как показано на рис. Генные продукты взаимодействуют, образуя белковые комплексы, которые управляют экспрессией соответствующих нижестоящих генов. У тройного мутанта, у которого отсутствуют все генетические функции А, В и С, вместо цветка получается неопределенная последовательность тесно расположенных листьев (см. ). И наоборот, в трансгенном растении, где все гены классов A, B и sepallata экспрессируются вместе за пределами своих нормальных доменов, листья трансформируются в лепестки.Таким образом, листья представляют собой «основное состояние», в котором ни один из этих гомеозисных селекторных генов не экспрессируется, в то время как другие типы органов возникают в результате экспрессии генов в различных комбинациях.
Рисунок 21-122
Экспрессия гена гомеозисного селектора в цветке Arabidopsis . (А) Схема нормальных паттернов экспрессии трех генов, мутантные фенотипы которых проиллюстрированы на фиг. 21-121А-С. Все три гена кодируют регуляторные белки генов. Цветное затенение (подробнее…)
Аналогичные исследования проводились и на других видах растений, и был выявлен аналогичный набор фенотипов и генов: растения не в меньшей степени, чем животные, сохранили свои гомеозисные системы генов-селекторов. Дупликация генов сыграла большую роль в эволюции этих генов: некоторые из них, необходимые в разных органах цветка, имеют явно гомологичные последовательности. Они не относятся к классу гомеобоксов, но являются членами другого семейства белков, регулирующих гены (так называемого семейства MADS), также обнаруженного у дрожжей и позвоночных.
Ясно, что растения и животные независимо друг от друга нашли очень похожие решения многих фундаментальных проблем многоклеточного развития.
Резюме
Развитие цветкового растения, как и у животного, начинается с деления оплодотворенной яйцеклетки с образованием зародыша с поляризованной организацией: апикальная часть зародыша образует побег, базальная часть, корень, а среднюю часть — стебель. Сначала деление клеток происходит по всему телу зародыша.Однако по мере роста эмбриона добавление новых клеток ограничивается небольшими областями, известными как меристемы. Апикальные меристемы на кончиках побегов и корнях сохраняются на протяжении всей жизни растения, позволяя ему расти за счет последовательного добавления новых частей тела на его периферии. Обычно побег образует повторяющуюся серию модулей, каждый из которых состоит из сегмента стебля, листа и пазушной почки. Подмышечная почка — это потенциальная новая меристема, способная дать начало боковой ветви; окружающая среда и гормональные сигналы дальнего действия внутри растения могут контролировать развитие растения, регулируя активацию почек.Мутации, изменяющие правила активизации пазушных почек, могут сильно повлиять на форму и структуру растения; одна такая мутация — одна из примерно пяти ключевых генетических изменений — объясняет большую часть резкого различия между современной кукурузой и ее диким предком, теосинте.
Небольшой сорняк Arabidopsis thaliana широко используется в качестве модельного организма для генетических исследований и является первым растением, геном которого полностью секвенирован. Как и у животных, гены, управляющие развитием растений, могут быть идентифицированы с помощью генетического скрининга, а их функции проверены с помощью генетических манипуляций.Такие исследования начали раскрывать молекулярные механизмы, с помощью которых внутренняя организация каждого растительного модуля очерчивается в микроскопическом масштабе посредством межклеточных взаимодействий вблизи апикальной меристемы. Сама меристема, по-видимому, поддерживается локальной петлей обратной связи, в которой клетки, экспрессирующие регуляторный белок гена Wuschel, обеспечивают положительный стимул, а отрицательная обратная связь, зависящая от межклеточного сигнального пути Clavata, не дает меристеме стать слишком большой.
Сигналы окружающей среды, особенно правильно подобранный по времени свет, могут вызывать экспрессию генов, которые переключают апикальную меристему из листообразующей в цветкообразующую.