
Что такое конъюгация? Примеры конъюгации
Многие, вероятно, не знали, что бактерии могут заниматься половым размножением. Это не совсем то, что можно себе представить. Сегодня рассмотрим процесс бактериальной конъюгации, что такое конъюгация в микробиологии и ее влияние на генетическую изменчивость бактерий.
Половое размножение бактерий
Это не станет большим шоком, но у бактерий нет полового пути размножения, по крайней мере, в обычном смысле. Бактериальные клетки размножаются путем создания клонов самих себя. Материнская клетка копирует свою ДНК-хромосому, затем разделяет ее клетку пополам, удерживая одну хромосому и отдавая ее новой дочерней клетке. По соглашению, эти клетки называются материнскими и дочерними, но на деле они являются клонами.
У них есть тот же самый генетический материал. В бактериальной популяции этот процесс продолжается, одна клетка делится на две снова и снова и снова, в результате чего появляются огромные популяции, которые являются всеми клонами друг друга. Это называется вертикальным переносом генов, когда ДНК передается от матери к потомству. И это то, что происходит в природе подавляющее большинство времени.

Бактерии — хитрые маленькие существа
Что такое конъюгация? У бактерий есть некоторые гениальные способы генерации генетического разнообразия. Например, бактерии способны обмениваться генами со своими соседями.
Это называется переносом горизонтального гена и относится к способности некоторых бактериальных клеток приобретать новые гены из соседних в их среде. Теперь вместо того, чтобы быть дочерней ячейкой клона, клетка имеет новое генетическое разнообразие, смесь между переданной по вертикали ДНК материнских клеток и переносимой по горизонтали соседней клеточной ДНК.
Помните, что у бактерий нет полового размножения? В эволюционном смысле это имеет решающее значение для разрешения смешивания и сопоставления генов, что приводит к генетическому разнообразию в пределах одного вида. Теперь мы можем видеть, что у бактерий нет обычного пола, у них есть горизонтальные механизмы переноса генов для генерации генетического разнообразия. В биологии это конъюгация.

Горизонтальный перенос гена
Существует три способа, которыми бактерии способны выполнять горизонтальный перенос генов:
- трансдукция использует бактериальные вирусы, называемые бактериофагами, для переноса ДНК из одной инфицированной клетки в другую;
- трансформация — это способность некоторых клеток принимать свободно плавающие ДНК, обнаруженные в окружающей среде;
- конъюгация позволяет переносить ДНК через структуру, называемую пилюсом, то есть из одной клетки в другую.
Что такое конъюгация?
Для конъюгации две живые бактериальные клетки должны вступать в прямой контакт друг с другом. Контакт между клетками осуществляется с помощью сопряжения особого придатка (пилус), напоминающего волос на поверхности бактерии. Он имеет белковую основу, подобную структуре волоса, которая простирается от бактериальной клетки.
Некоторые придатки используются для прикрепления к поверхностям, но специальное сопряжение применяется исключительно для прикрепления к другим клеткам и облегчения переноса ДНК.
Клетка, которая будет передавать ДНК, называется донорской и строит конъюгированный пилус. Его конъюгация представляет собой полую трубчатую структуру, которая соединяет цитоплазму донорной клетки с цитоплазмой реципиента.
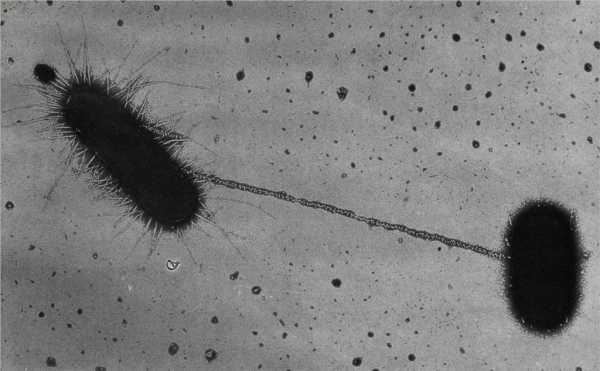
Перенос плазмиды
Когда цитоплазма клетки-донора и реципиента физически связана, настало время для переноса ДНК. Если донорская клетка содержит плазмиду, круглую часть внехромосомной ДНК, то ее можно перенести в клетку-реципиент. Это делается путем копирования плазмиды и отправки цепи копируемой ДНК получателю через сопряжение пилуса. Конечным результатом является копия плазмиды как у донора, так и у реципиента. Возможно, наиболее интересным является тот факт, что плазмида несет гены, которые позволяют клетке-реципиенту стать самим донором конъюгации! Теперь клетка-получатель также может распространять плазмиду на новые клетки, с которыми она сталкивается в своей среде.

Значение конъюгации
Бактериальное конъюгация — это передача генетического материала между бактериальными клетками путем прямого контакта или мостоподобной связью. Это механизм горизонтального переноса генов, как и трансформация, и трансдукция. Бактериальное сопряжение часто рассматривается как бактериальный эквивалент полового размножения или спаривания, поскольку оно связано с обменом генетическим материалом. Однако это не половое размножение, так как обмен гаметой не происходит.
Во время конъюгации (микробиология) донорская клетка обеспечивает конъюгативный или мобилизуемый генетический элемент, который чаще всего является плазмидой или транспозоном. Большинство конъюгативных плазмид имеют системы, гарантирующие, что клетка-получатель уже не содержит подобный элемент. Передаваемая генетическая информация часто выгодна получателю. Преимущества могут включать устойчивость к антибиотикам, ксенобиотическую толерантность или способность использовать новые метаболиты.
Такие полезные плазмиды можно рассматривать как бактериальные эндосимбионты. А другие элементы как бактериальные паразиты, конъюгацию как механизм, разработанный ими для обеспечения их распространения. Этот процесс был открыт в 1946 году Джошуа Ледербергом и Эдвардом Татумом.

Механизм конъюгации
Сначала донорская клетка производит пилус. Он присоединяется к ячейке-получателю и объединяет две клетки. Мобильная плазмида зазубривается, и одна нить ДНК затем переносится в клетку-реципиент. Обе синтезируют комплементарную цепь для получения циркулярной плазмиды, а также размножают пилусы. Обе клетки в настоящее время являются жизнеспособным донором для F-фактора.
F-плазмида представляет собой эписому (плазмиду, которая может интегрироваться в бактериальную хромосому посредством гомологичной рекомбинации). Она несет в себе происхождение репликации и источник передачи. В данной бактерии, свободной или интегрированной, может быть только одна копия F-плазмиды, а бактерии, обладающие копией, называются F-позитивными и обозначаются F+. Клетки, у которых отсутствуют F-плазмиды, называются F-отрицательными (F—), они могут функционировать как ячейки-получатели.
Хотя есть некоторые споры о точном механизме конъюгации, может показаться, что пилусы не являются структурами, через которые происходит обмен ДНК, однако все же трансформация ДНК продолжается. Несколько белков, закодированных в локус, открывают канал между бактериями. Считается, что фермент, расположенный у основания пилуса, инициирует слияние мембран.
Примеры
Яркий пример конъюгации демонстрируют бактерии, связанные с азотфиксирующими тризобиями, которые представляют собой интересный случай внутреннего сопряжения. Например, индуцирующая опухоль (Ti) плазмида Agrobacterium и индуцирующая корневую опухоль (Ri) плазмида A. rhizogenes содержат гены, которые способны переносить растительные клетки. Экспрессия этих генов эффективно преобразовывает растительные клетки в опин-продуцирующие растения. Опины используются бактериями в качестве источников азота и энергии. Зараженные клетки образуют коронарные желчные или корневые опухоли.
Таким образом, плазмиды Ti и Ri являются эндосимбионтами бактерий, которые в свою очередь являются паразитами зараженного растения. Плазмиды Ti и Ri также могут переноситься между бактериями. Такие передачи создают вирулентные штаммы от ранее авирулентных штаммов.
Что такое конъюгация? Это удобное средство для переноса генетического материала со множеством целей. Сообщалось об успешных передачах от бактерий к дрожжам, растениям, клеткам млекопитающих, диатомовым и изолированным митохондриям млекопитающих. Конъюгация имеет преимущества перед другими формами генетического переноса, включая минимальное нарушение клеточной оболочки цели и способность передавать относительно большие количества генетического материала.
fb.ru
Половое размножение
Половое размножение
Половой процесс. Половое размножение отличается наличием полового процесса, который обеспечивает обмен наследственной информацией и создает условия для возникновения наследственной изменчивости. В нем, как правило, участвуют две особи — женская и мужская, которые образуют гаплоидные женские и мужские половые клетки — гаметы. В результате оплодотворения, т. е. слияния женской и мужской гамет, образуется диплоидная зигота с новой комбинацией наследственных признаков, которая и становится родоначальницей нового организма.
Половое размножение по сравнению с бесполым обеспечивает появление наследственно более разнообразного потомства. Формами полового процесса являются конъюгация и копуляция.
Конъюгация — своеобразная форма полового процесса, при которой оплодотворение происходит путем взаимного обмена мигрирующими ядрами, перемещающимися из одной клетки в другую по цитоплазматическому мостику, образуемому двумя особями. При конъюгации обычно не происходит увеличения количества особей, но происходит обмен генетическим материалом между клетками, что обеспечивает перекомбинацию наследственных свойств. Конъюгация типична для ресничных простейших (например, инфузорий), некоторых водорослей (спирогиры).
Копуляция (гаметогамия) — форма полового процесса, при которой две различающиеся по полу клетки — гаметы — сливаются и образуют зиготу. При этом ядра гамет образуют одно ядро зиготы.
Различают следующие основные формы гаметогамии: изогамия, анизогамия и оогамия.
При изогамии образуются подвижные, морфологически одинаковые гаметы, однако физиологически они различаются на «мужскую» и «женскую». Изогамия встречается у многих водорослей.
При анизогамии (гетерогамии) формируются подвижные, различающиеся морфологически и физиологически гаметы. Такой тип полового процесса характерен для многих водорослей.
В случае оогамии гаметы сильно отличаются друг от друга. Женская гамета — крупная неподвижная яйцеклетка, содержащая большой запас питательных веществ. Мужские гаметы —
Гаметогенез. Процесс образования и развития гамет называется гаметогенезом. У многоклеточных водорослей, многих грибов и высших споровых растений формирование гамет происходит в специальных органах полового размножения — гаметангиях. У высших споровых растений женские гаметангии называются архегониями, мужские — антеридиями. У животных гаметогенез протекает в специальных половых железах —гонадах. Однако, например, у губок и кишечнополостных половые железы отсутствуют и гаметы возникают из различных соматических клеток.
Сперматозоиды и яйцеклетки обычно формируются соответственно особями мужского и женского пола. Биологические виды, у которых все организмы делятся в зависимости от производимых ими клеток на самцов и самок, называются раздельно-полыми. Встречаются виды, у которых один и тот же организм может образовывать как мужские, так и женские половые клетки. Такие организмы называются гермафродитами (в греческой мифологии гермафродит — дитя Гермеса и Афродиты — обоеполое существо, несущее в себе и женское, и мужское начало). Гермафродитизм наблюдаются у многих беспозвоночных животных (моллюсков, плоских и кольчатых червей), а также у круглоротых (миксины) и рыб (морской окунь). В этом случае организмы, как правило, имеют ряд приспособлений, препятствующих самооплодотворению. У некоторых моллюсков половая железа продуцирует попеременно мужские и женские половые клетки. Это зависит от условий существования особи и ее возраста.
У большинства низших животных гаметы вырабатываются в течение всей жизни, у высших — только в период половой активности, с момента полового созревания до затухания деятельности желез в старости.
Половые клетки в своем развитии претерпевают ряд сложных преобразований. Процесс формирования мужских половых клеток называется сперматогенез, женских — оогенез.
Сперматогенез и строение мужских гамет у высших животных. Сперматогенез происходит в мужских половых железах — семенниках. Семенник высших животных состоит из семенных канальцев. В каждом канальце можно обнаружить отдельные зоны, в которых клетки расположены концентрическими кругами. В каждой зоне клетки находятся на соответствующих стадиях развития. Сперматогенез складывается из четырех периодов: размножения, роста, созревания и формирования (рис. 2.1).По периферии семенного канальца располагается зона размножения. Клетки этой зоны называются сперматогониями. Они усиленно делятся митозом, благодаря чему увеличивается их количество и сам семенник. Период интенсивного деления сперма-то гониев называется
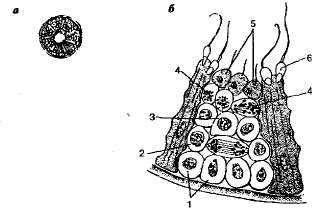
Рис 2.1. Сперматогенез в семенном канальце: а—срез через каналец; 6 —участок канальца при большем увеличении; I — спериатогонии; 2 — делящийся сперматацит первого порядка; 3 — сперматоцит второго порядка; 4 — делящийся сперматоцит второго порядка; 5 — сперматиды; 6 — сперматозоид.
После наступления половой зрелости некоторые сперматого-нии перемещаются в следующую зону — зону роста, расположенную ближе к просвету канальца. Здесь клетки увеличиваются в размерах за счет возрастания количества цитоплазмы и превращаются в сперматоциты первого порядка (период роста).
Третий период развития мужских гамет называется периодом созревания. В это время сперматоциты первого порядка делятся мейозом. После первого деления образуется два сперматоцита второго порядка, а после второго — четыре сперматиды, имеющие овальную форму и значительно меньшие размеры. Сперматиды перемещаются в зону, ближайшую к просвету канальца (зона формирования). Здесь сперматиды изменяют свою форму и превращаются в зрелые сперматозоиды, которые затем выносятся из семенников по семявыносящим путям.
В семенниках формируется огромное количество сперматозоидов. Так, при каждом половом акте у человека наружу выносится около 200 млн. сперматозоидов.
Форма мужских гамет у разных видов животных различна. Наиболее типичны для высших животных сперматозоиды, имеющие головку, шейку и длинный хвост, служащий для активного передвижения. Именно такое строение имеют сперматозоиды человека. Ширина их овальной головки 1,5—2 мкм, длина хвоста — около 60 мкм. Головка содержит ядро и незначительное количество цитоплазмы с органеллами. На переднем конце головки расположена акросома, представляющая собой видоизмененный аппарат Гольджи. В ней содержатся ферменты, растворяющие оболочку яйцеклетки при оплодотворении. В шейке находятся центриоли и митохондрия.
Сперматозоиды не имеют запасов питательных веществ и обычно быстро погибают. Однако у некоторых животных, например у пчел, они обладают большой жизнеспособностью и сохраняются живыми в течение нескольких лет, находясь в специальном органе самки — семяприемнике.
Оогенез н строение яйцеклеток у высших животных. Оогенез происходит в особых железах — яичниках —и включаеттри периода: размножение, рост и созревание. Период формирования здесь отсутствует.
В период размножения интенсивно делятся предшественники половых клеток — оогонии. У млекопитающих этот период заканчивается еще до рождения. К этому времени формируется около 30 тыс. оогониев, которые сохраняются долгие годы без изменения. С наступлением половой зрелости отдельные оогонии периодически вступают в период роста. Клетки увеличиваются, в них накапливается желток — образуются
Зрелая яйцеклетка, как и сперматозоид, содержит в себе половинное число хромосом, так как в период созревания ооциты первого порядка претерпевают мейоз. Яйцеклетки чаще всего имеют сферическую форму (рис. 2.3). Они обычно значительно крупнее соматических клеток. Яйцеклетка человека, например, имеет в диаметре 150—200 мкм. Особенно больших размеров достигают яйцеклетки животных, эмбриональное развитие которых происходит вне тела матери (яйца птиц, рептилий, амфибий и рыб).
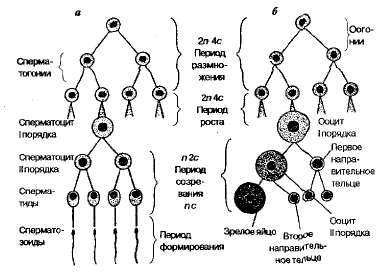
Рис 2.2. Схема сперматогенеза (а) и оогенеза (б).
Рис 2.3. Строение яйцеклетки млекопитающего: I — цитоплазма; 2 — ядро; 3 — оболочка; 4 — фолликулярные клетки.
В яйцеклетках содержится ряд веществ, необходимых для формирования зародыша. В первую очередь это питательный материал — желток. В зависимости от количества желтка и характера его распределения различают несколько типов яйцеклеток.
Яйцеклетки покрыты оболочками. По происхождению оболочки делят на первичные, вторичные и третичные. Первичная оболочка яйцеклетки является производной цитоплазмы и называется желточной оболочкой. Ее наличие характерно для яйцеклеток всех животных. Вторичные оболочки образуются за счет деятельности клеток, питающих яйцеклетку. Вторичная оболочка характерна, например, для членистоногих (хитиновая оболочка). Третичные оболочки возникают в результате деятельности желез половых путей. К третичным относятся скорлуповая, подскорлу-повая и белковая оболочки яиц птиц и пресмыкающихся, студенистая оболочка яйцеклеток земноводных.
Оболочки выполняют защитные функции, обеспечивают обмен веществ с окружающей средой, а у плацентарных служат для внедрения зародыша в стенку матки.
Осеменение и оплодотворение. Процесс, обусловливающий встречу мужских и женских половых клеток у животных, называется осеменением. Различают наружное и внутреннее осеменение.
При наружном осеменении, характерном для большинства водных животных, сперматозоиды и яйцеклетки выделяются в воду, где и происходит их слияние. Для такого осеменения не обязательна непосредственная встреча мужских и женских особей, но необходимо большое количество гамет, так как большая часть их гибнет.
Внутреннее осеменение характерно для обитателей суши, где отсутствуют условия для сохранения и встречи гамет во внешней среде. При таком типе осеменения сперматозоиды вводятся в половые пути самки. У самцов для этого обычно имеются специальные совокупительные органы. Внутреннее осеменение характерно для всех наземных позвоночных (рептилий, птиц, млекопитающих), а также червей, пауков и насекомых.
При достижении сперматозоидами яйцеклеток происходит процесс оплодотворения. Осуществляется он следующим образом. При контакте с яйцеклеткой акросома сперматозоида разрывается и ее содержимое высвобождается. Под воздействием ферментов акросомы оболочка яйцеклетки в месте контакта растворяется. Внутренняя поверхность акросомы вытягивается, и формируется акро-сомальный отросток, который проникает через растворенную зону яйцевых оболочек и сливается с мембраной яйцеклетки. В этом месте из цитоплазмы образуется воспринимающий бугорок. Он захватывает ядро, центриоли и митохондрии сперматозоида и увлекает их внутрь яйцеклетки. Цитоплазматическая мембрана сперматозоида встраивается в мембрану яйцеклетки.
Проникновение сперматозоида в яйцеклетку вызывает отслаивание от яйцеклетки оболочки оплодотворения. Между ней и поверхностью яйцеклетки возникает пространство, заполненное жидкостью. Образование оболочки оплодотворения препятствует проникновению других сперматозоидов в яйцеклетку.
Проникшее в цитоплазму яйцеклетки ядро сперматозоида набухает, достигает величины ядра яйцеклетки. Ядра сближаются и сливаются. Этот момент и есть собственно оплодотворение. В результате из двух гамет образуется одна диплоидная зигота, т. е. восстанавливается диплоидный набор хромосом.
При оплодотворении в яйцеклетку обычно проникает один сперматозоид. Однако у насекомых, рыб, птиц и других животных в яйцеклетку может проникать несколько сперматозоидов. Это явление получило название полиспермии. При этом с ядром яйцеклетки сливается ядро только одного сперматозоида. Ядра других сперматозоидов разрушаются. Тем не менее для оплодотворения требуется участие многих сперматозоидов, так как они выделяют ферменты, обеспечивающие их проникновение в яйцеклетку. Если ферментов недостаточно, оплодотворение не наступает.
Источник : Н.А. Лемеза Л.В.Камлюк Н.Д. Лисов «Пособие по биологии для поступающих в ВУЗы»
sbio.info
Копуляция (изогамная, анизогамная, оогамная), Конъюгация
Половое размножение
Половое размножение связано с образованием растениями особого типа клеток — гамет. Растение, на котором происходит образование гамет, называют гаметофитом. Процесс формирования гамет называют гаметогенезом. Он происходит в особых органах — гаметангиях. У равноспоровых растений гаметофит обычно обоеполый: несет и женские, и мужские гаметангии. У разноспоровых растений из микроспор развивается гаметофит с мужскими гаметангиями, а из мегаспор — гаметофит с женскими гаметангиями.
Гаметы всегда гаплоидны. При слиянии мужской и женской гамет происходит образование зиготы, из которой развивается новый организм. Процесс слияния гамет называют оплодотворением.
Сущность полового процесса едина для всех живых организмов, а его формы разнообразны. Различают следующие типы полового процесса: хологамия, изогамия, гетерогамия и оогамия (рис. 39).
| Рис. 39. Типы полового размножения: 1 — изогамия; 2 — гетерогамия; 3 — оогамия; 4 — конъюгация. |
Хологамия — слияние гаплоидных одноклеточных, внешне неотличимых организмов друг с другом. Этот тип полового процесса характерен для некоторых примитивных водорослей. В данном случае сливаются не гаметы, а целые организмы, выступающие в роли гамет. Образовавшаяся диплоидная зигота обычно сразу же делится мейотически и образуется 4 дочерних гаплоидных одноклеточных организма.
Особой формой полового процесса является конъюгация, характерная для некоторых нитчатых водорослей. Отдельные гаплоидные клетки нитевидных талломов, расположенных близко друг от друга, начинают образовывать выросты. Они растут навстречу друг другу, соединяются, перегородки в месте
стыка растворяются, и содержимое одной клетки (мужской) переходит в другую (женскую). В результате конъюгации образуется диплоидная зигота.
При изогамии (равный брак) гаметы морфологически сходны между собой, то есть одинаковы по форме и размерам, но физиологически они разнокачественны. Данный половой процесс характерен для многих водорослей и некоторых грибов. Изогамия происходит только в воде, для передвижения в которой гаметы снабжены жгутиками. Они очень похожи на зооспоры, но имеют меньшие размеры.
При гетерогамии (разный брак) происходит слияние подвижных половых клеток, сходных по форме, но различающихся размерами. Женская гамета в несколько раз больше мужской и менее подвижна. Гетерогамия характерна для тех же групп организмов, что и изогамия, и также происходит в воде.
Характерна для некоторых водорослей и всех высших растений. Женская гамета — яйцеклетка — крупная и неподвижная. У низших растений образуется в одноклеточных гаметангиях — оогониях, у высших растений (исключая покрытосеменные) — в многоклеточных архегониях. Мужская гамета (сперматозоид) мала и подвижна, образуется у грибов и водорослей в одноклеточных, а у высших растений (исклю-
чая покрытосеменные) — в многоклеточных гаметангиях — антеридиях. Сперматозоиды способны передвигаться только в воде. Поэтому наличие воды — обязательное условие для оплодотворения у всех растений, за исключением семенных. У большинства семенных растений мужские гаметы утратили жгутики и называются спермиями.
Дата добавления: 2016-05-30; просмотров: 802;
Похожие статьи:
Формы полового размножения: Копуляция (изогамная, анизогамная, оогамная), Конъюгация
Копуляция – Копуляция представляет собой процесс слияния двух гаплоидных гамет и их ядер с образованием диплоидной зиготы.
Различают три разновидности копуляции: гаметогамию, автогамию и гамонтогамию.
При гаметогамии происходит копуляция (слияние) свободноподвижных гамет от двух различных гамонтов (клеток, образующих гаметы).
Если гаметы морфологически не отличаются друг от друга, говорят об изогамии; при анизогамии размеры, форма или структура гамет различны.
Наконец, оогамия—это вариант анизогамии, при котором мужские и женские гаметы напоминают по своему облику и различиям в размерах сперматозоиды и яйцеклетки многоклеточных.
Автогамными называют тех одноклеточных, у которых копулируют гаметы (или гаметные ядра), происходящие от одного гамонта (самооплодотворение).
Этот вид полового размножения известен у солнечников, фораминифер и инфузорий.
При гамонтогамии половой процесс начинается со слияния гамонтов (специальные половые клетки), как например у многих фораминифер и у некоторых Apicomplexa.
На последующих этапах процесса появляются гаметы (или, по крайней мере, гаметные ядра), которые сливаются между собой.
Конъюгация — совершенно своеобразный тип полового процесса, свойственный только инфузориям (Ciliophora) и одной группе зеленых водорослей.
И в том и в другом случае между конъюгирующими клетками образуется цитоплазматический мостик, через который осуществляется слияние ядер партнеров. При этом у конъюгат, как, впрочем, и у некоторых инфузорий, сливается частично или полностью цитоплазма.
Поэтому, строго говоря, четкой границы между копуляцией и конъюгацией провести нельзя. Для инфузорий особенно характерно отсутствие прямой связи полового процесса с размножением.
В конъюгацию вступают 2 особи, и в результате этого процесса образуются также 2 особи, но обладающие уже другим (гибридным) геномом. Число особей при этом не увеличивается, но физиологическое состояние их меняется, а также изменяется генотип. Таким образом, завершившая конъюгацию особь представляет собой в генетическом смысле новое поколение.
Жизненным циклом называется совокупность закономерно сменяющих друг друга стадий развития организма между двумя одинаковыми стадиями.
В принципе в качестве точки отсчета может быть выбрана любая из таких стадий, но, как правило, для этого выбирают момент образования оплодотворенного яйца (зиготы), из-за чего часто говорят, что жизненным циклом называется период в жизни организма «от яйца до яйца».
Если смена способа размножения не сопряжена с изменением состояния хромосомного набора, то чередование поколений является гомофазным; в иных случаях его называют гетерофазным.
В итоге различают 3 основные формы соотношения гаплоидной и диплоидной фазы в жизненном цикле простейших:
При зиготической редукции диплоидна лишь зигота — продукт оплодотворения, слияния гамет.
Уже первое деление зиготы является мейозом, приводящим к редукции числа хромосом. Все остальные этапы жизненного цикла протекают в гаплоидной фазе.
Описанная форма соотношения гапло- и дипло-фазы свойственна многим водорослям и гетеротрофным жгутиконосцам, а также всем споровикам.
Гаметическая редукция является прямой противоположностью зиготической. Здесь все фазы жизненного цикла диплоидны, мейоз наблюдается при образовании гамет, которые являются единственной гаплоидной фазой в жизненном цикле.
Гаметическая редукция имеет место в жизненном цикле солнечников, инфузорий, некоторых жгутиконосцев.
Гетерофазное чередование поколений (спорическая редукция) характеризуется закономерной сменой диплоидной фазы гаплоидной. Типичный пример этого явления демонстрируют фораминиферы. Из зиготы развивается диплоидное поколение, размножающееся агамно путем образования бесполых одноядерных агамет.
Их формирование происходит посредством мейоза. Из гаплоидных агамет развивается следующее поколение — гамонты.
Биология для школьного учителя
В конце жизни этого гаплоидного поколения происходит формирование гамет (без мейоза). Последние копулируют, и из зиготы развивается диплоидное поколение.
Если смена способа размножения не сопряжена с изменением состояния хромосомного набора, то чередование поколений является гомофазным; в иных случаях его называют гетерофазным.
В итоге получается три типа чередования поколений:
1. Гапло-гомофазное: диплоидна только зигота, все остальные стадии гаплоидные (например, у Apicomplexa).
Дипло-гомофазное: гаплоидны только гаметы (или гаметные ядра), все остальные стадии диплоидны (например, у инфузорий).
3. Гетерофазное: за (гаплоидным или диплоидным) поколением с бесполым размножением следует поколение, которое размножается половым путем (например, у фораминифер).
В ходе жизненных циклов протистов при наступлении определенных условий формируются покоящиеся стадии — цисты, что имеет место как у свободноживущих, так и у паразитических форм. У последних инцистирование обычно бывает связано с выходом из хозяина в окружающую среду и с заражением новых особей хозяина.
У протистов, обладающих сложной организацией (солнечники, инфузории, некоторые жгутиконосцы) при образовании цист имеет место дедифференцировка: ресничные образования, скелет, органоиды захвата и приема пищи и другие, связанные с активной жизнью, резорбируются, а иногда отбрасываются.
Обычно происходит обезвоживание и уплотнение цитоплазмы, исчезают вакуоли, в том числе пищеварительные. Вокруг тела простейшего выделяются 2 или более защитных оболочек (цисты). Обладая слабой проницаемостью, оболочки служат не только механической, но и химической защитой особи. В инцистированном состоянии простейшие могут сохранять жизнеспособность месяцами и даже годами, цисты могут иметь различное физиологическое назначение.
Цисты покоя служат для перенесения неблагоприятных условий среды — чаще всего они позволяют простейшему переносить высыхание в водоеме или почве. Широко распространены у многих свободно-живущих простейших цисты размножения. Например, некоторые инфузории и гетеротрофные жгутиконосцы перед делением инцистируются, а после завершения деления снова эксцистируются и переходят к свободному существованию.
Предыдущая12345678910111213141516Следующая
Дата добавления: 2016-02-24; просмотров: 1098;
ПОСМОТРЕТЬ ЕЩЕ:
Размножение водорослей
Каким образом размножаются водоросли?
Водоросли размножаются половым и бесполым путем.
Как правило, бесполым путем водоросли размножаются в благоприятный период.
Формы полового размножения: Копуляция (изогамная, анизогамная, оогамная), Конъюгация
Бесполое размножение осуществляется вегетативно (у одноклеточных — делением надвое, у многоклеточных — частями слоевища), спорами (неподвижными клетками) и зооспорами (подвижными клетками). Рассмотрим для примера бесполое (зооспорами) и половое размножение одноклеточной водоросли. Если водоросль подвижна, то перед размножением она теряет жгутики. Ядро и цитоплазма делятся пополам; затем происходит еще одно или два деления, в результате которых в одной и той же оболочке образуются 4–8 клеток.
Эти мелкие подвижные клетки — зооспоры — выходят из оболочки материнской клетки и вырастают во взрослую особь.
В неблагоприятный период (высокая или низкая температура, накопление продуктов обмена в среде обитания при высокой плотности заселения, загрязнение водоемов) происходит половое размножение. Каждая клетка делится на много мелких половых клеток — гамет, которые потом попарно сливаются с гаметами другой особи, образуя зиготу, несущую в результате признаки обеих родительских особей.
Последняя покрывается плотной оболочкой и зимует. Весной из зиготы выходит 4 зооспоры. Каждая из них дает начало новому растению.
Половое размножение водорослей
Половое размножение у водорослей связано с половым процессом, который заключается в слиянии двух клеток, в результате чего образуется зигота, вырастающая в новую особь или дающая зооспоры.
Половое размножение у водорослей бывает нескольких типов:
гологамия (коньюгация) – без образования специализированных клеток;
гаметогамия – с помощью специализированных клеток – гамет.
Гологамия.
В простейшем случае процесс происходит путем слияния двух неподвижных, лишенных клеточных оболочек вегетативных клеток. У одноклеточных жгутиковых форм водорослей половой процесс осуществляется путем слияния двух особей.
При слиянии содержимого двух безжгутиковых вегетативных клеток половой процесс называют коньюгацией.
Во время коньюгации происходит слияние двух клеток, которые выполняют функцию половых клеток – гамет. Слияние содержимого клеток происходит через специально образующийся коньюгационный канал, получается зигота, которая впоследствии покрывается толстой оболочкой и превращается в зигоспору. Если скорость перетекания содержимого клеток одинакова, зигота образуется в коньюгационном канале. В этом случае разделение клеток на мужскую и женскую условно.
Гаметогамия.
Половое размножение у водорослей, в том числе и одноклеточных, чаще происходит путем деления содержимого клеток и образования в них специализированных половых клеток – гамет. У всех зеленых и бурых водорослей мужские гаметы имеют жгутики, а женские гаметы имеют их не всегда. У примитивных водорослей гаметы образуются в вегетативных клетках. У более высокоорганизованных форм гаметы находятся в особых клетках, называемых гаметангиями.
В вегетативной клетке или гаметангии может быть от одной до нескольких сотен гамет. В зависимости от размеров сливающихся гамет различают несколько типов гаметогамии: изогамия, гетерогамия, оогамия.
Если сливающиеся гаметы имеют одинаковую форму и размеры, такой половой процесс называют изогамией.
Если сливающиеся гаметы имеют одинаковую форму, но разные размеры (женская гамета крупнее мужской), то говорят о гетерогамии.
Половой процесс, при котором сливаются неподвижная крупная клетка – яйцеклетка и подвижная мелкая мужская клетка – сперматозоид, называется оогамией.
Гаметангии с яйцеклетками называют архегониями или оогониями, а со сперматозоидами – антеридиями. Мужские и женские гаметы могут развиваться на одной особи (однодомные) или на разных особях (двудомные).
Образующаяся в результате слияния гамет зигота после некоторых изменений превращается в зигоспору. Последняя обычно одета плотной оболочкой. Зигоспора может находиться длительное время в покое (до нескольких месяцев) или прорастать без периода покоя.
Автогамия. Особый тип полового процесса. Он заключается в том, что ядро клетки делится мейотически, из четырех образовавшихся ядер, два разрушаются, а оставшиеся два ядра сливаются, образуя зиготу, которая без периода покоя увеличивается в размерах и превращается в ауксоспору.
Так происходит омоложение особей.
ЖИЗНЕННЫЙ ЦИКЛ ВОДОРОСЛЕЙ
У одного и того же вида водорослей в зависимости от времени года и внешних условий наблюдаются разные формы размножения (бесполое и половое), при этом происходит смена ядерных фаз (гаплоидной и диплоидной). Исключение составляют виды, лишенные полового процесса. Изменения, претерпеваемые особями вида между одноименными стадиями (моментами жизни), составляют его цикл развития.
У одних видов органы бесполого и полового размножения развиваются на разных особях; тогда растения, образующие споры, называют спорофитами, а растения, производящие гаметы,— гаметофитами.
У других водорослей споры и гаметы образуются на одних и тех же растениях; одновременно у таких видов могут существовать и особи, дающие только споры, т.
е. спорофиты (порфира). Сейчас растения, способные производить и споры (зооспоры) и гаметы, как правило, называют гаметофитами. Однако во избежание путаницы с настоящими гаметофитами, производящими только гаметы, их лучше называть гаметоспорофитами.
Развитие органов размножения того или иного типа у гаметоспорофитов определяется температурой. Например, пластинчатые слоевища одного из видов порфиры (Porphyra tenera) при температуре ниже +15, + 17 °С производят органы полового размножения, а при более высокой температуре— органы бесполого размножения.
И у других водорослей гаметы обычно появляются при более низкой температуре, чем споры. При промежуточной температуре развитие тех или иных органов размножения на гаметоспорофитах определяется другими факторами — интенсивностью света, длиной дня, сезонными изменениями химического состава воды или солености (для морских водорослей). Гаметоспорофиты существуют у улотриксовых, ульвовых и кладофоровых из зеленых водорослей, у эктокарповых, хордариевых, сфацеляриевых и пунктариевых из бурых водорослей, бангиевых и некоторых немалиевых из красных.
Спорофиты и гаметофиты (гаметоспорофиты) бывают одинакового строения или разного, и соответственно существуют понятия изоморфной (сходной) и гетероморфной (разной) смены форм развития (чередования поколений).
В отношении большинства водорослей неправильно говорить о чередовании поколений спорофитов и гаметофитов (гаметоспорофитов), так как они нередко существуют одновременно. Иногда они могут расти в несколько различных экологических условиях.
Например, спорофит порфиры имеет вид ветвящихся нитей из одного ряда клеток, которые внедряются в известковый субстрат (раковины моллюсков, известковые скалы) и предпочитают слабое освещение, проникая на большую глубину.
Гаметоспорофит порфиры пластинчатый и растет вблизи уреза воды, в том числе в приливно-отливной зоне.
Разница в строении спорофитов и гаметофитов (гаметоспорофитов) при гетероморфной смене форм развития может быть очень значительной.
Гаметоспорофит или гаметофит может быть многоклеточным, высотой в несколько сантиметров, а спорофит — микроскопическим, одноклеточным (акросифония из зеленых). Возможна и обратная картина, когда гаметофит микроскопический и даже одноклеточный, а спорофит достигает в длину 12 м (ламинария японская из бурых).
Гаметофиты и спорофиты подавляющего большинства водорослей — самостоятельные растения. У ряда водорослей спорофиты растут на гаметофитах (филлофора Броди из красных) или гаметофиты развиваются внутри слоевищ спорофитов (циклоспоровые из бурых).
Поскольку при половом процессе в результате слияния гамет и их ядер происходит удвоение набора хромосом в ядре, то в последующем в какой-то момент цикла развития наступает редукционное деление ядра (мейоз), в результате которого дочерние ядра получают одинарный набор хромосом.
Спорофиты многих водорослей диплоидные, и мейоз в цикле их развития совпадает с моментом образования спор, из которых развиваются гаплоидные гаметоспорофиты или гаметофиты. Такой мейоз называют спорической редукцией (рис. 25,1).
В спорофитах примитивных водорослей (кладофора, эктокарпус и многие другие) наряду с гаплоидными спорами могут образовываться диплоидные споры, которые снова развиваются в спорофиты.
Споры, появляющиеся на гаметоспорофитах, служат для воспроизведения материнских растений. Спорофиты и гаметофиты водорослей, стоящих на верхних ступенях эволюции, строго чередуются без самовозобновления (ламинариевые из бурых, многие флоридеи из красных).
Ряд водорослей имеют мейоз в зиготе, т. е. зиготическую редукцию (рис.
25, 2). Она характерна для конъюгат из зеленых водорослей.
Зиготы части пресноводных зеленых водорослей, таких, как вольвоксовые, улотриксовые и др., представляют собой одноклеточные спорофиты. Они производят до 32 зооспор, что по массе во много раз больше, чем пара родительских гамет.
Таким образом, у этих водорослей, по существу, наблюдается спорическая редукция.
Некоторые группы водорослей имеют гаметическую редукцию, которая характерна для царства животных. Мейоз происходит при образовании гамет, остальные клетки всегда диплоидные (рис. 25, 3).
Такая смена ядерных фаз присуща диатомовым и циклоспоровым водорослям, а также одному из видов кладофоры (Cladophora glomerata). Интересно отметить, что диатомовые преобладают по количеству видов над другими водорослями и встречаются во всех местообитаниях, где только способны расти водоросли.
В свою очередь, циклоспоровые относятся к наиболее массовым морским водорослям. Видимо, цикл развития с гаметической редукцией дает этим водорослям какие-то преимущества.
Ботаника. 1. 12. Основные органеллы цитоплазмы
У зеленой водоросли празиолы (Prasiola stipitata) обнаружена соматическая редукция — мейоз совершается в вегетативных клетках верхней части диплоидного гаметофита, при этом появляются участки гаплоидных клеток, в которых вслед за тем образуются гаплоидные гаметы (рис. 25, 4).
В цикле развития водорослей, лишенных полового размножения (сине-зеленые, криптофитовые и эвгленовые) или имеющих его в редких случаях (золотистые, желто-зеленые и динофитовые), наблюдаются только изменения в строении тела.
Поэтому применительно к таким водорослям принято говорить о цикломорфозе. Он может охватывать несколько поколений или ограничиваться периодом роста и развития одной особи. В наиболее резкой форме цикломорфоз выражен у гиеллы дернистой (Hyella caespitosa) из сине-зеленых водорослей и у гленодиниума Борге (Glenodinium borgei) из динофитовых.
Как циклы развития, так и цикломорфозы у водорослей отличаются большой пластичностью.
Их прохождение во многом определяется экологическими условиями. Поэтому они далеко пе всегда сопровождаются строго последовательным проявлением всех стадий. В зависимости от условий произрастания отдельные стадии и формы развития могут выпадать полностью (например, спорофит или гаметоспорофит и гаметофит) или, наоборот, существовать на протяжении нескольких поколений, с тем чтобы на период жизни одного поколения уступить место другой форме развития.
Строго упорядоченные циклы развития существуют у водорослей, стоящих на верхних ступенях эволюции (рис. 26).
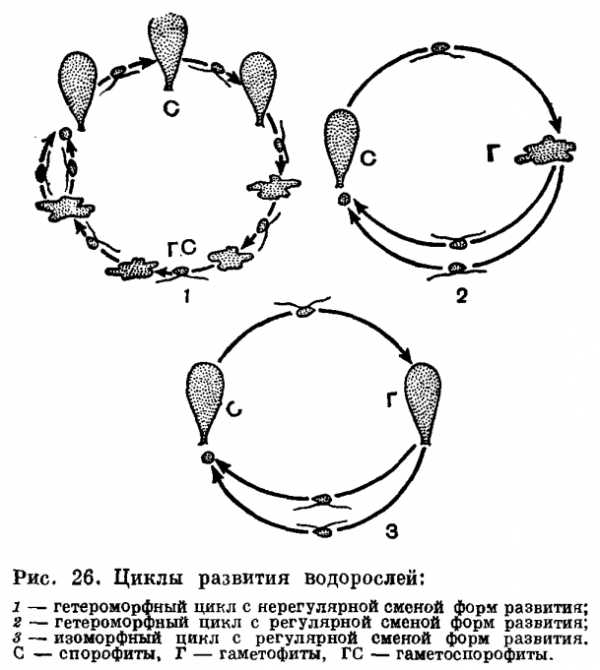
Соотношение диплоидной и гаплоидной фаз в жизненном цикле разных водорослей неодинаково.
В одних случаях прорастание зиготы сопровождается редукционным делением конуляционного ядра (зиготическая редукция) и развивающиеся при этом растения оказываются гаплоидными. Так, у многих зеленых водорослей (вольвокальные, большинство хлорококкальных, ко-нъюгатофициевые, харофициевые) зигота —единственная диплоидная стадия в цикле развития, вся вегетативная жизнь проходит у них в гаплоидном состоянии, они являются гаплонтами.
У других водорослей, наоборот, вся вегетативная жизнь осуществляется в диплоидном состоянии, а гаплоидная фаза представлена лишь гаметами, перед образованием которых и происходит редукционное деление ядра (гаметическая редукция). Зигота без редукционного деления ядра прорастает в диплоидный таллом. Эти водоросли— диплонты. Таковы многие зеленые водоросли, имеющие сифоновое строение, все диатомовые, из бурых — представители порядка фукальные.
У третьих редукционное деление ядра предшествует образованию зооспор или анланоспор, развивающихся, как правило, на диплоидных талломах (спорическая редукция). Эти клетки бесполого размножения вырастают в гаплоидные растения, размножающиеся только половым путем. После слияния гамет зигота развивается в диплоидное растение, несущее только органы бесполого размножения. Таким образом, у этих водорослей имеет место чередование форм развития (генераций): диплоидного бесполого спорофита и гаплоидного полового гаметофита.
Оба поколения могут быть одинаковы морфологически (изоморфная смена генераций) или же резко различны по внешнему виду (гетероморфная смена генераций).
Изоморфная смена генераций характерна для морских видов ульвы, энтероморфы, кладофоры, хетоморфы из зеленых водорослей, для ряда порядков бурых и большинства красных водорослей (несколько усложненная), Гетероморфная смена генераций особенно распространена среди бурых водорослей, но встречается у зеленых и красных.
Читайте также:
ekoshka.ru
Размножение у простейших. Конъюгация и копуляция.
К типу Простейшие относят организмы, тело которых состоит из одной клетки, функционирующей как целый организм.
Простейшие могут размножаться бесполым и половым путем. Бесполое размножение осуществляется путем простого и множественного деления. Для многих групп простейших свойственен половой процесс — копуляция и конъюгация. При конъюгации происходит слияние ядер разных клеток, а при копуляции сливаются целые клетки (в этом плане клетка простейших аналогична гамете многоклеточных).
Амеба размножается только путем деления надвое. Бесполое размножение плазмодия осуществляется в теле человека, а половой процесс — в малярийном комаре рода анофелес. Для инфузории характерен половой процесс по типу конъюгации. Две инфузории соединяются «ротовыми» сторонами, между ними возникают анастомозы. В каждой клетке макронуклеус (тип ядра) растворяется, а микронуклеус мейотически делится. Образуется 4 ядра, 3 из которых растворяются, а оставшееся ядро делится митотически. Одно из образовавшихся ядер мигрирует к партнеру, второе сливается с пришедшим ядром, при этом восстанавливается диплоидность микронуклеуса. Клетки расходятся, а образовавшееся ядро делится, образуется микро — и макронуклеус. Позже макронуклеус путем эндомитоза восстанавливает своюполиплоидность. В результате конъюгации происходит рекомбинация генетического материала инфузорий.
38- 40 по книге!!!
41. Балантидий. Назвать по-русски и по латыни. Систематическое положение, морфология, цикл развития, пути заражения. Название болезни и ее лабораторная диагностика. Профилактика
Балантидий — Balantidium Coli
Тип Простейшие, класс Ресничные инфузории
Морфологические особенности. Балантидий существует в двух формах: трофозоита (вегетативная стадия) и цисты. Несимметричное овальное тело балантидия покрыто короткими ресничками, которые расположены продольными рядами. На переднем конце брюшной поверхности располагается перистом, который открывается наружу широким отверстием и заканчивается в клетке глоткой. Ротовое отверстие окружено длинными ресничками, которые продолжаются в глубину глотки. На заднем конце тела помещается перфорированное анальное отверстие. В цитоплазме находятся две сократительные вакуоли. Имеет два ядра: макронуклеус и микронуклеус. Размножается балантидий поперечным делением. Описан также половой процесс — конъюгация. Цисты (Balantidium coli) диаметром около 50–60 мкм имеют округлую форму и окружены двойной оболочкой. В цитоплазме цисты сохраняется макронуклеус и сократительные вакуоли.
Жизненный цикл. Балантидии в кишечнике свиней образуют цисты, которые выходят наружу с фекалиями. Заражение человека происходит перорально при проглатывании цист с загрязненными овощами, водой, с немытых рук (при забое свиней, использовании свиного навоза в качестве удобрения и при загрязнении им источников воды). Возможна передача инфекции от человека к человеку. Цисты могут распространяться мухами. Из проглоченных цист выходят трофозоиты, которые живут и размножаются в просвете толстого кишечника.
Действие на организм человека. Во многих случаях заболевание — балантидиаз протекает бессимптомно и развивается цистоносительство. У больных с клиническими проявлениями в стенке толстого кишечника находят такие же изменения, как при амебиазе, — поражение слизистой кишечника, язвы на стенках кишечника, развивается диарея. В отличие от амебиаза, при балантидиазе паразиты не распространяются в организме гематогенно.
Диагностика. Обнаружение трофозоитов в фекалиях или биоптате толстого кишечника.
Профилактические мероприятия. Выявление и лечение больных людей и цистоносителей. Личная гигиена: пить только кипяченую воду, соблюдать чистоту рук (особенно перед едой). Проводить ветеринарный контроль свиней на фермах, уничтожать мух. Санитарно-просветительская работа среди населения.
Жизненный цикл паразитов. Чередование поколений и феномен смены хозяев; промежуточные и основные хозяева. Понятие о био- и гео-гельминтах. Примеры.
Жизненный цикл – это совокупность всех стадий развития «от яйца до яйца», а при отсутствии яйца от любой стадии до ближайшей такой же.
Чередование поколений и феномен смены хозяев.Гельминтам со сменой хозяев в жизненном цикле для его реализации необходимо пройти через строго обязательный круг промежуточных и окончательного хозяев. Это минимально необходимая, и в некоторых случаях достаточная основа паразитарной системы. Однако у многих эндопаразитических гельминтов при широкой специфичности личиночных и ювинильных стадий количество особей хозяев на данной стадии личиночного развития может быть достаточно велико (от 2–3 и в условиях океанской пелагиали до 5–6 и более). Особенно характерно это явление для океанических гельминтов, реализующих свои жизненные циклы по трофическим каналам биотической структуры сообществ. Эти хозяева могут быть как разно-размерными представителями одного вида, так и представителями разных типов, классов и т.д. Это явление связано с последовательным рядом актов поедания хищниками зараженных жертв (принцип «эстафетной передачи»).
В одних случаях за счет тех или иных причин (высокий иммунный барьер и т.п.) гельминты, попав в хозяина живыми и активными, погибают. В других случаях в первом подходящем хищнике (хозяине) у личинок происходят морфо-физиологические изменения, специфичные для данной стадии онтогенеза (промежуточный хозяин), а в последующих хозяевах они могут жить, не претерпевая дальнейшего развития (дополнительные паратенические – транспортные хозяева). Дальнейшее онтогенетическое развитие возможно лишь при попадании личинки гельминта в промежуточных хозяев следующего уровня, которые необходимы для прохождения следующей стадии онтогенеза. При этом, благодаря трофическим, пространственным или временным особенностям экологической ниши данного хозяина личинки гельминтов могут или оставаться в нем неопределенно долго (в соответствии с продолжительностью онтогенеза хозяина), или попадать к хищникам (хозяевам) не имеющих трофических контактов с необходимыми для дальнейшего развития хозяевами.
Промежуточные и основные хозяева.Далее, если новый хозяин имеет «подходящие» биоценотические связи, он «используется» паразитом для оптимизации путей достижения окончательного хозяина в соответствии с особенностями данной экосистемы и положения в ее пространственной и трофической структуре промежуточных и окончательных хозяев с одной стороны и «нового» хозяина – с другой. Наиболее далеко зашедшее взаимоотношение с такими хозяевами – использование их в качестве паратенических (транспортных) хозяев. Это явление приобретает особенно важное значение в случае значительного накопления личинок гельминтов в теле хозяина. Последних в отечественной литературе обычно называют резервуарными хозяевами, и они являются источником массового заражения следующих хозяев. В этих резервуарных хозяевах иногда сосредоточена основная численность личинок данной стадии жизненного цикла популяции гельминта.
В конечном варианте развития этой тенденции «освоения» новых хозяев, некоторые из них могут стать с экологической точки зрения необходимыми (облигатны) для замыкания жизненного цикла данной популяции гельминта, и далее попасть в категорию промежуточных и или реже окончательных хозяев.
Обязательные хозяева – генетически и соответственно физиологически жестко детерминированный круг обязательных хозяев, в которых происходят обязательные подготовительные морфо-физиологические стадийные процессы онтогенетического развития и достижение взрослого половозрелого состояния гельминтов. Эти обязательные хозяева необходимы для завершения жизненного цикла, через них «проходит» в процессе жизненного цикла основная часть популяции гельминта.
Факультативные хозяева: в которых также происходит полный или частичный морфогенез и достижение половой зрелости, но они не являются обязательными хозяевами и их заражение данным гельминтом носит случайный характер. Это обычно близкородственные виды по отношению к хозяевам первичной структуры с отличающимися от них экологическими, и в первую очередь, трофическими характеристиками. Они не являются полноценными и необходимыми членами паразито-хозяинных систем, но могут быть использованы частью гельминтов данной популяции для замыкания онтогенеза.
Понятие о био- и геогельминтах.В зависимости от условий развития паразитических червей делят на две большие группы: биогельминты и геогельминты. К биогельминтам относятся те паразиты, которые развиваются с участием двух или более организмов. В одном организме живут взрослые формы червя, в другом – личиночные стадии. Организм, в котором паразитируют взрослые формы и происходит половое размножение, называют окончательным (или дефинитивным) хозяином. Организм, в котором развиваются личиночные формы, – промежуточным хозяином. Например, бычий солитер во взрослом состоянии паразитирует в кишечнике человека, а развитие его личинки происходит в организме крупного рогатого скота. Таким образом, для этого солитера человек является окончательным хозяином, а корова – промежуточным хозяином. К биогельминтам относится большинство представителей типа плоских червей. Геогельминтами называют тех паразитов, которые в процессе своего развития не требуют смены хозяев. Их яйца выводятся из организма вместе с калом во внешнюю среду и при определенной температуре и влажности в них развиваются личинки. Такое яйцо, содержащее личинку, становится заразным. Попадая в организм человека (в его кишечник) личинки освобождаются от оболочек яйца, проникают в те или иные органы и вырастают в половозрелую форму. У некоторых гельминтов личинка освобождается из яйца во внешней среде. Такая личинка живет в воде или в почве, претерпевает определенные стадии развития и в дальнейшем активно проникает в организм через кожные покровы. К геогельминтам относятся многие круглые черви (за исключением трихинеллы).
infopedia.su
15. Стадии конъюгации инфузорий. Биологический смысл конъюгации.
Размножение – бесполое (поперечное деление клетки надвое), которое чередуется с половым процессом. В последнее время ряд авторов называют этот половой процесс, встречающийся только у инфузорий, примитивной формой полового размножения типа конъюгации. При бесполом размножении микронуклеус делится митозом, макронуклеус – амитозом. Конъюгация инфузорий-туфелек – сложный процесс, продолжающийся несколько часов. В конъюгации можно выделить следующие этапы.
Соединение двух инфузорий (конъюгантов) друг с другом перистомальными областями с образованием в месте контакта цитоплазматического мостика, соединяющего обе инфузории.
Демонтаж макронуклеусов, деление микронуклеусов мейозом с образованием четырех гаплоидных ядер.
Разрушение трех гаплоидных ядер, деление четвертого митозом. Из двух образовавшихся гаплоидных ядер, одно останется на месте, второе переместится по цитоплазматическому мостику в другую инфузорию. Первое ядро условно называют женским, второе – мужским.
Обмен мужскими ядрами, следствием которого является перекомбинация генетической информации.
Слияние мужского и женского ядер собразованием диплоидного синкариона, расхождение инфузорий. После расхождения инфузории называют эксконъюгантами.
Деление синкариона три раза митозом, образование восьми диплоидных ядер, из которых четыре являются микронуклеусами, четыре – будущими макронуклеусами.
Демонтаж трех микронуклеусов, образование инфузории с пятью ядрами: одно – микронуклеус (диплоидный), четыре – макронуклеусы, тоже диплоидные.
Деление каждого из эксконъюгантов надвое, причем генеративные ядра делятся митозом, а макронуклеусы расходятся попарно в дочерние клетки. Образование четырех дочерних инфузорий, каждая из которых имеет один микронуклеус и два макронуклеуса.
Деление каждой из четырех дочерних инфузорий надвое, вновь микронуклеусы делятся митозом, а макронуклеусы расходятся в дочерние инфузории. В итоге из одной материнской инфузории образуются четыре дочерних инфузории, в каждой из которых один макронуклеус и один микронуклеус. Микронуклеус остается диплоидным. Макронуклеус становится полиплоидным, поскольку перед каждым из расхождений в нем реплицируется ДНК.
Половой процесс инфузорий — конъюгация. В ходе конъюгации макронуклеос распадается, а микронуклеос проходит сложный цикл созревания. Половой процесс состоит в том, что две особи разного полового типа сближаются и прикладываются друг к другу теми поверхностями тела, на которых находится рот. Макронуклеосы распадаются, а микронуклеосы претерпевают мейотическое деление, так что в каждой инфузории образуется по четыре гаплоидных ядра. Три из них дегенерируют, остается по одному, каждое из которых делится затем митотически. Образуется по два гаплоидных ядра, одно из этих идентичных ядер остается в клетке, а другое через область рта переходит в другую особь и сливается с ее гаплоидным ядром. Таким образом, при конъюгации происходит как бы взаимное оплодотворение, а оба новых диплоидных ядра идентичны. Затем инфузории расходятся, макронуклеосы восстанавливаются. Следует заметить, что в данном случае размножения как такового не происходит (увеличения числа особей не наблюдается), а происходит обновление генетического материала.
У инфузорий имеется еще одна особенность — передача наследственной информации не через хромосомы ядра, а через цитоплазму (цитоплазматическая наследственность).
Смысл конъюгации в правильном делении набора хромосом между двумя клетками, когда гомологичные хромосомы образуют перед делением пары. При половом размножении каждая соматическая клетка содержит двойной набор хромосом: из «отцовского» и «материнского» организмов. Если бы конъюгации не было, то половые клетки получали бы не равноценные половинные наборы исходного генома, а случайную комбинацию, как это происходит при образовании половых клеток у стерильных гибридов
studfiles.net
Коньюгация | Дистанционные уроки
07-Ноя-2013 | Нет комментариев | Лолита Окольнова
Всем известно преимущество полового размножения над бесполым. Половой процесс позволяет разнообразить генетическую информацию. Это позволяет проявиться новым признакам, которые, возможно, будут полезными.
Природа знает много таких механизмов, в результате которых, генетическая информация приобретает большее разнообразие: начиная от случайного распределения хромосом при мейозе, заканчивая кроссинговером.

Эта статья посвящена одному из таких процессов. Он свойственен инфузориям. Называется очень умным словом конъюгация.
В генетике этим словом словом обозначают тесное сближение хромосом для осуществления ранее упомянутого кроссинговера.
Конъюгация у инфузорий — это процесс, при котором инфузории сближаются и обмениваются генетической информацией. Похожие, к общем, процессы.
Давайте поподробнее рассмотрим процесс более подробно.
Плавает, значит, себе инфузория, вдруг понимает, что ее генетическая информация «устарела», ее нужно разнообразить. Для этого она тесно прижимается к другой такой же инфузории, между ними образуется так называемый цитоплазматический мостик, он соединяет клетки. Как вам, наверное, известно, у инфузории два ядра: полиплоидный макронуклеус (∞n) (вегетативное ядро) и диплоидный (2n) микронуклеус (генеративное ядро).
- Макронуклеус управляет всеми процессами в клетке;
- микронуклеус участвует в конъюгации.

После сближения и коньюгации, у инфузории микронуклеус проходит два деления мейозом, в результате которого образуются четыре гаплоидных (1n) ядра, три из которых рассасываются. Одно оставшееся гаплоидное (1n) ядро делится митозом и образует два пронуклеуса, которые тоже гаплоидны (1n). В это время макронуклеус рассасывается. То же самое происходит и у второй инфузории.
Далее инфузории обмениваются пронуклеусами. То есть инфузория отдает один свой пронуклеус и принимает пронуклеус другой инфузории. Затем «родной» пронуклеус сливается с пронуклеусом, полученным из второй инфузории. Образуется диплоидное (2n) ядро. Потом цитоплазматический мостик исчезает, инфузории больше не связаны, коньюгация завершена и каждая «идет своей дорогой».
Затем образовавшееся в результате слияния ядро делится митозом. Получаются два диплоидных (2n) ядра. Одно ядро — теперь новый микронуклеус инфузории. А второе ядро претерпевает множественное деления, образуя полиплоидное ядро — это макронуклеус инфузории.

Таким образом, в результате коньюгации генетическая информация инфузории обновилась, произошел половой процесс.
автор статьи Саид Лутфуллин
Еще на эту тему:
Обсуждение: «Коньюгация»
(Правила комментирования)distant-lessons.ru
Размножение инфузорий — урок. Биология, Животные (7 класс).
Инфузории могут размножаться как половым, так и бесполым путями.
Бесполое размножение
Инфузории-туфельки обычно размножаются бесполым путём — делением надвое. Однако, в отличие от жгутиковых, инфузории делятся поперек тела: посреди тела инфузории образуется перетяжка и, таким образом, из одной клетки образуется две.
Инфузории-туфельки делятся один-два раза в сутки.

Половой процесс
Ядра делятся на две части, и в каждой новой инфузории оказывается по одному большому и по одному малому ядру. Каждая из двух дочерних инфузорий получает часть органоидов, а другие образуются заново.
Конъюгация — половой процесс одноклеточных животных, при котором те обмениваются своим наследственным материалом.
Этапы конъюгации инфузорий

1. Сближение двух инфузорий (МИ — микронуклеус; МА — макронуклеус) и образование между ними цитоплазматического мостика.
2. Первое деление микронуклеуса (как результат, образование двух ядер).
3. Второе деление микронуклеуса (как результат, образование четырех ядер).
4. Трое из четырёх образовавшихся микронуклеусов погибают. Также погибает макронуклеус.
5. Третье деление микронуклеуса (как результат, образование двух ядер).
6. Обмен ядрами (одно из двух образовавшихся ядер переходит к клетку другой инфузории).
7. Слияние двух новых микронуклеусов в одно ядро.
8. Деление ядра надвое.
9. Из двух имеющихся ядер, одно становится макронуклеусом, другое — микронуклеусом.
Биологическое значение конъюгации состоит в обновлении генетического материала и возможном появлении новых признаков.
Источники:
Биология. Животные. 7 кл.:учеб. для общеобразоват. Учреждений/ В.В. Латюшин, В.А. Шапкин. – 10-е изд., стереотип. М.: Дрофа, 2009. – 302, [2] с. :ил.
Вахрушев А.А., Бурский О.В., Раутиан А.С. Биология (От амёбы до человека). 7 класс. Учебник общеобразовательной школы. – М:Баласс, 2013. – 320с., ил. (Образовательная система «Школа 2100»)
Константинов В.М., Бабенко В.Г., Кучменко B.C./Под ред. Константинова В.М. Биология. 7 класс Издательский центр ВЕНТАНА-ГРАФ
Иллюстрации:
http://cmd4win.ucoz.hu/blog/prezentacija_na_temu_bespoloe_razmnozhenie/2013-05-27-44
www.yaklass.ru