
Ткани — урок. Биология, Человек (8 класс).
Ткани состоят из клеток и межклеточного вещества. Каждая ткань выполняет строго определённую функцию. Выполняемые функции взаимосвязаны со строением. Поэтому ткани отличаются высокой специфичностью.
Ткань — это группа клеток и межклеточного вещества, которые имеют общее происхождение и развитие, сходное строение и выполняют определённую функцию.
В организме человека выделяют следующие виды тканей:
- эпителиальную;
- соединительную;
- мышечную;
- нервную.
Рис. \(1\). Виды тканей
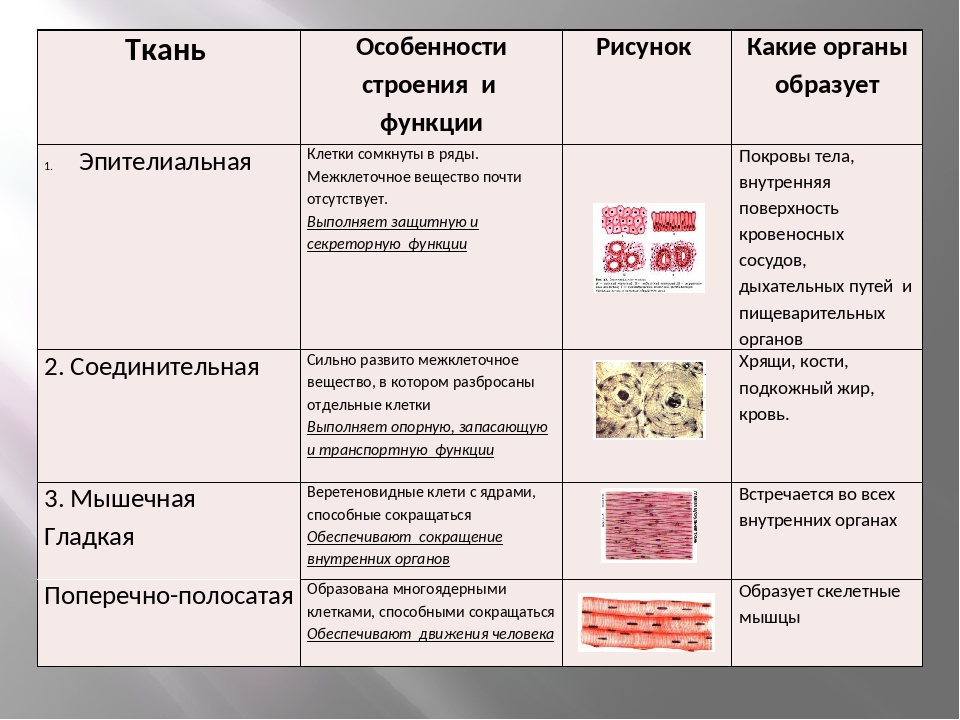
Эпителиальная ткань состоит из плотно прижатых клеток (межклеточного вещества мало), которые выполняют барьерную, защитную и секреторную функции. Она образует покровы тела, слизистые оболочки, железы.
В соединительной ткани клетки хорошо развито межклеточные вещество. Оно представлено волокнами, жидкостями, костными пластинками и т.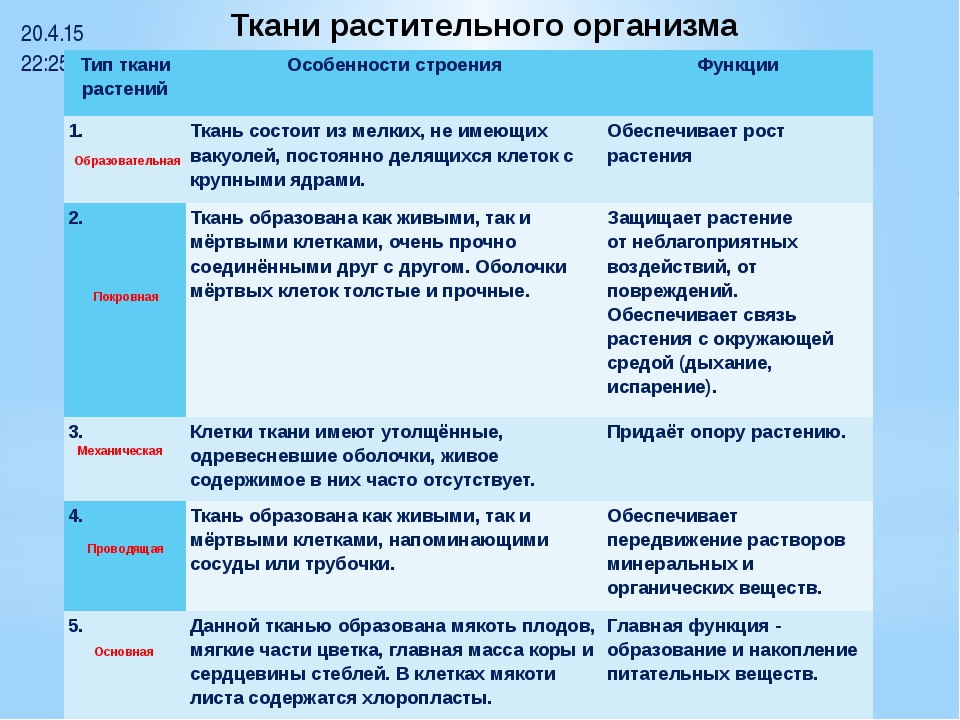 д.
д.
Эти особенности строения позволяют соединительной ткани выполнять опорную (кости, хрящи, сухожилия), защитную (подкожный жир), питательную (кровь, лимфа) функции.
Мышечные ткани выполняют сокращение сердечной и скелетных мышц, внутренних органов, изменение просвета кровеносных сосудов.
В зависимости от особенностей местонахождения и выполняемых функций мышечная ткань бывает:
- поперечно-полосатая скелетная;
- поперечно-полосатая сердечная;
- гладкая (мышечная ткань кровеносных сосудов и внутренних органов: желудка, мочевого пузыря и др.).
Более интенсивная работа сердечной и скелетных мышц обусловила особенности строения поперечно-полосатой ткани в отличие от гладкой.
Поперечно-полосатая мышечная ткань состоит из развитых многоядерных мышечных волокон. Гладкая ткань образована короткими одноядерными мышечными волокнами.
Нервная ткань состоит из нейронов и нейроглии.
Она обеспечивает передачу возбуждения от нервных окончаний (рецепторов) к центральной нервной системе, а от неё к органу.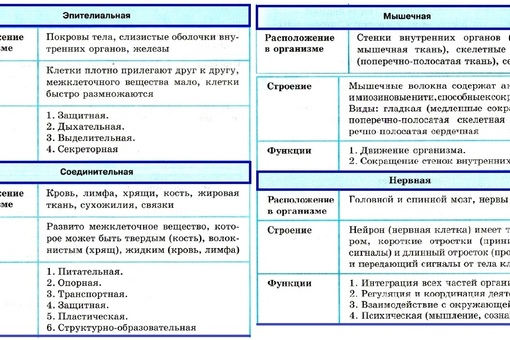 Это возможно благодаря особому строению нейронов. Нейрон имеет тело, длинный отросток (аксон) и короткие отростки (дендриты).
Это возможно благодаря особому строению нейронов. Нейрон имеет тело, длинный отросток (аксон) и короткие отростки (дендриты).
Нервная ткань расположена в головном и спинном мозге, нервных узлах, нервных волокнах.
Источники:
Рис. 1. Виды тканей: © ЯКласс
Типы тканей и их свойства
Тема: Ткани. Типы тканей и их свойства
Цель: познакомить учащихся со строением и функциями тканей организма человека, развивать навыки самостоятельной работы с учебником, составления таблиц, распознавания микроструктур по описанию.
Ход урока
Организационный момент
Проверка домашнего задания.
Отчеты учащихся о заполнении таблицы «Функции органоидов и частей клетки».
Изучение нового материала
Познавательные вопросы по теме урока:
Что вам известно о тканях?
Какие ткани составляют организм человека?
Каковы их особенности строения, свойства и функции?
/Ответы обучающихся/
Слайд 1 — Во многом вы правы.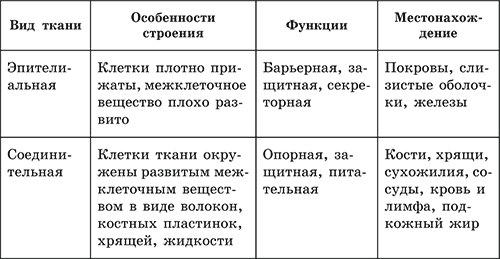 Давайте подробно остановимся на особенностях тканей человека. Запишите тему нашего урока.
Давайте подробно остановимся на особенностях тканей человека. Запишите тему нашего урока.
Слайд 2 — В процессе изучения материала вам необходимо заполнить следующую таблицу:
Вспомним основные понятия:Ткань – группа клеток и межклеточного вещества, объединенные общим строением, происхождением и функцией.
Тканевая жидкость пополняется из вышедшей из кровеносного сосуда жидкой части крови, состав которой при этом изменяется, межклеточное вещество выделяется клетками
Слайд 3 – В организме человека выделяются четыре группы тканей.
Эпителиальная. Клетки плотно прилегают друг к другу, межклеточного вещества мало
Соединительная. Клетки расположены рыхло, сильно развито межклеточное вещество
Мышечная. Образована мышечными волокнами, способна возбуждаться и сокращаться
Нервная. Состоит из клеток с отростками. Способна возбуждаться и передавать возбуждение
Слайды 4-9 – Эпителиальная (покровная) ткань.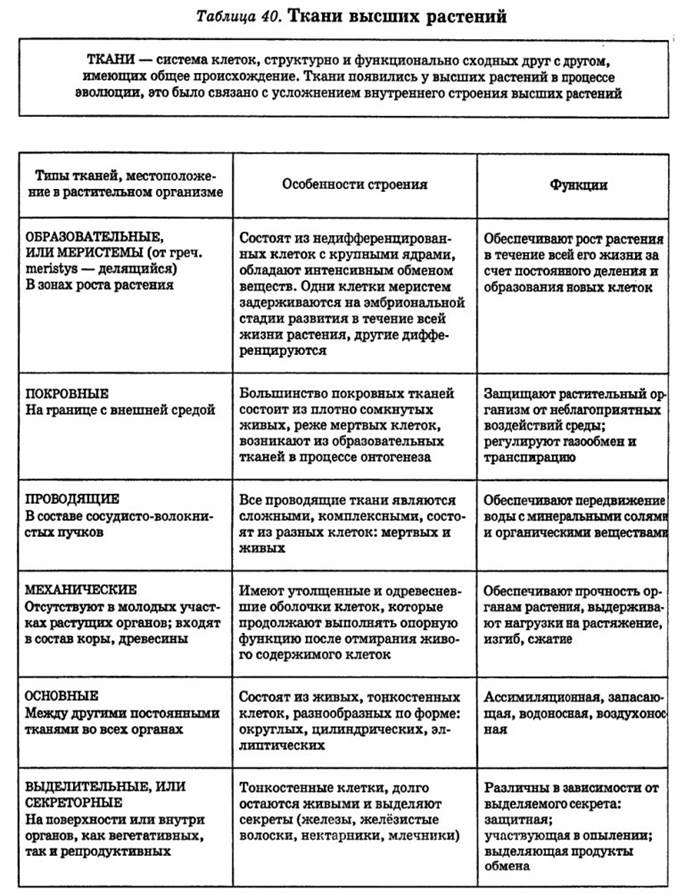
Однослойная. Расположение: смежные оболочки внутренних органов. Функции: защитная, всасывающая
Многослойная. Расположение: покровы тела. Функции: защитная.
Железистая. Расположение: железы внешней и внутренней секреции. Функции: секреторная
Эпителиальная ткань – ткань, покрывающая тело и выстилающая его полости в виде пласта.
1) Образуется в онтогенезе раньше всех других тканей из зародышевых листков.
2) Способна к регенерации.
3) Лишена кровеносных сосудов.
4) Клетки плотно прилегают друг к другу.
5) Имеет мало межклеточного вещества.
6) Может состоять из нескольких слоев клеток.
7) Основные функции — защита (кожа), всасывание (кишечник), избирательный транспорт (почки, сосуды).
Слайды 10-16 – Соединительная ткань.
Костная. Расположение: скелет. Функции: Опорная, защитная, кроветворная
Хрящевая. Расположение: скелет, органы дыхания, ушная раковина. Функции: опорная, защитная
Волокнистая. Расположение: связки, сухожилия, дерма, прослойки между органами. Функции: опорно-защитная
Расположение: связки, сухожилия, дерма, прослойки между органами. Функции: опорно-защитная
Жировая. Расположение: подкожная клетчатка, между внутренними органами.
Функции: запасающая, защитная
К соединительной ткани относится и жидкая ткань – кровь.
Соединительная ткань — ткань, развивающаяся из мезодермы и выполняющая следующие функции: опорную (костная и хрящевая), трофическую (жировая и лимфа), защитную (лимфоидная и кровь).
1) Клетки не прилегают друг к другу.
2) Много межклеточного вещества.
3) Отличается большим разнообразием клеток.
Слайды 17-22 – Мышечная ткань.
Поперечно-полосатая скелетная . Расположение: опорно-двигательный аппарат тела и некоторых внутренних органов (язык, глотка, начальная часть пищевода). Функции: сократительная
Состоят из многоядерных мышечных волокон, покрытых возбудимой мембраной.
а) Волокна объединяются в мышечные пучки, из которых состоит мышца.
б) Основа скелетной мускулатуры.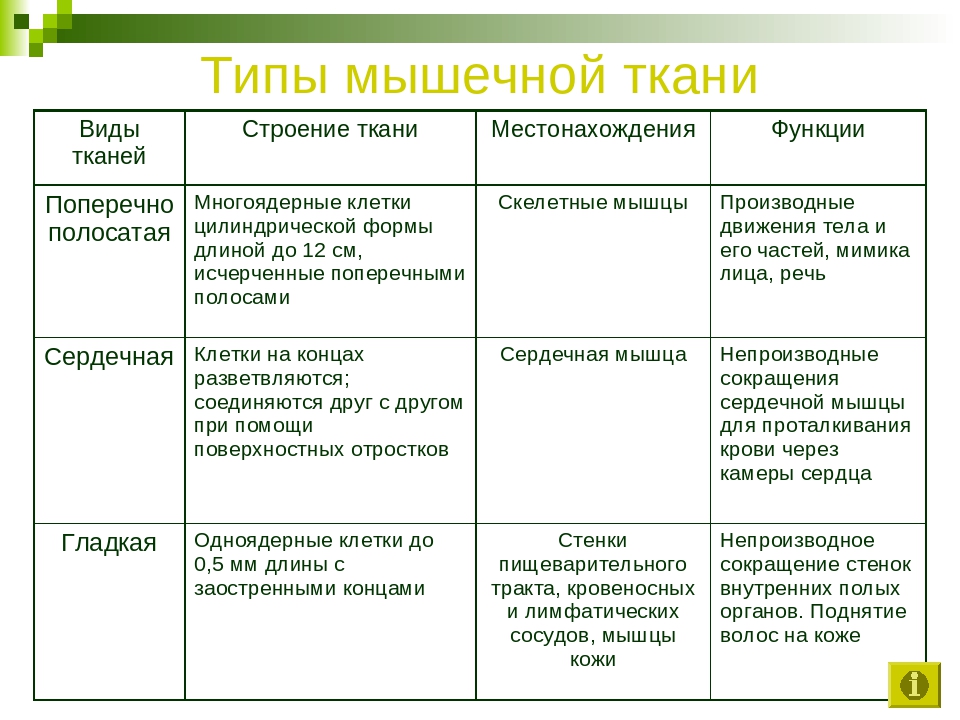
в) Белые поперечнополосатые мышцы содержат много миофибрилл, сильно сокращаются, но быстро утомляются.
г) Красные — мало миофибрилл, имеют меньшую силу, но могут долго работать.
д) Клетки имеют поперечную исчерченность за счет миофибрилл.
МИОФИБРИЛЛЫ — мышечные нити,состоящие из саркомеров, способные к сокращению за счет актино-миозинового взаимодействия.
Сердечная мышца. Состоит из прямоугольных сократительных поперечнополосатых клеток.
а) Сокращения более медленные, чем у скелетных мышц.
б) Волокна переплетены в пучки.
в) Клетки не сливаются как в поперечнополосатой мышце.
г) Способны к автоматическим сокращениям.
д) Имеет большой рефрактерный период (не может сокращаться).
е) Может управляться вегетативной нервной системой.
ж) Сокращается в объеме, уменьшая просвет полостей сердца.
Мышечная ткань — ткань, состоящая из клеток мезодермального происхождения, способных к возбуждению и сокращению.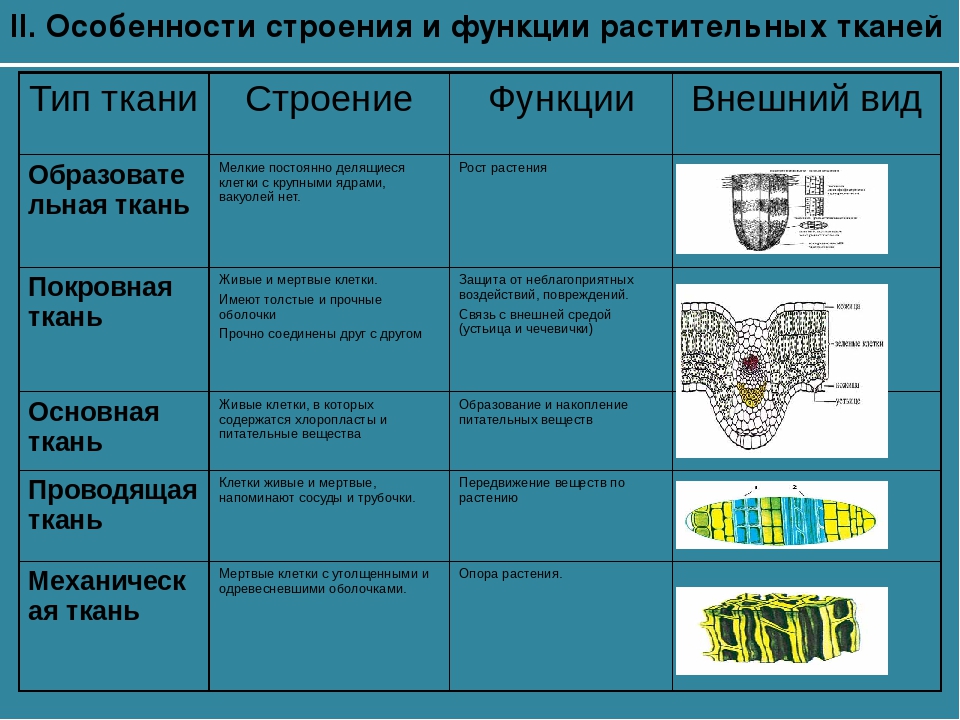
Гладкая мышечная ткань — сократимая ткань, состоящая из отдельных клеток и не имеющая поперечной исчерченности.
Клетки сильно вытянуты. Способны к медленным длительным сокращениям.
Управляется вегетативной нервной системой. Входят в состав внутренних органов и сосудов.
Слайды 23-27 – Нервная ткань. Расположение: Головной и спинной мозг, нервные узлы и волокна Функции: Обеспечение согласованной деятельности разных систем органов, обеспечение связи организма с внешней средой, приспособление обмена веществ к изменяющимся условиям
Строение нервной ткани:
Нейроглия – вспомогательная роль (опора, питание)
Нейрон = тело + отростки (дендриты + аксон)
Дендрит – отросток, передающий возбуждение к телу нейрона.
Аксон – длинный единственный отросток, передающий информацию от тела нейрона к другому нейрону или рабочему органу.
Нервная ткань образована клетками (нейронами) эктодермального происхождения.
Нейроны не делятся.
Они способны к возбуждению и проведению нервного импульса.
Образуют стабильные контакты с другими клетками.
Образуют группы — ганглии, серое и белое вещество, нервные волокна.
Рассмотреть строение нейрона по схеме.
Закрепление изученного материала.
Провести фронтальное уточнение по заполнению таблицы
Покровы тела
Защита
Железистая
Железы внешней и внутренней секреции
Секреторная
Соединительная – клетки расположены рыхло, сильно развито межклеточное вещество.
Костная
Скелет
Опорная, защитная, кроветворная
Хрящевая
Скелет, органы дыхания, ушная раковина
Опорная, защитная
Волокнистая
Связки, сухожилия, дерма, прослойки между органами
Опорно-защитная
Жировая
Подкожная клетчатка, между внутренними органами
Запасающая, защитная
Кровь
Полости сердца и кровеносные сосуды
Дыхательная, транспортная, защитная
Мышечная – образована мышечными волокнами, способна возбуждаться и сокращаться.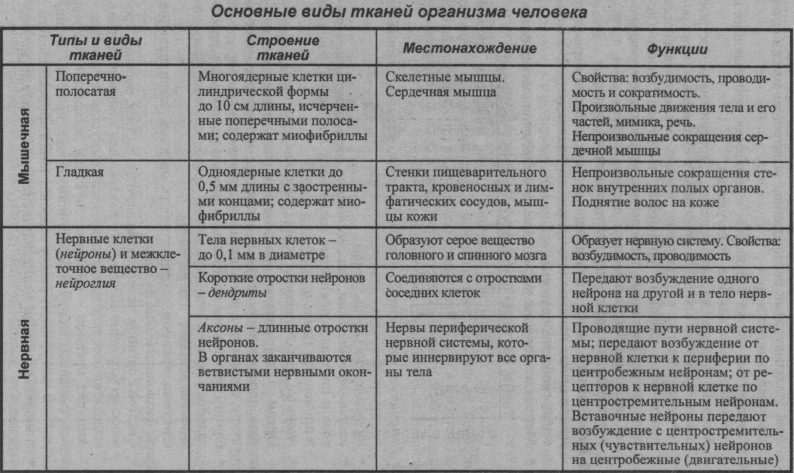
Поперечно-полосатая скелетная
Опорно-двигательный аппарат тела и некоторых внутренних органов (язык, глотка, пищевод)
Сократительная
Поперечно-полосатая сердечная
Сердце
Сократительная
Гладкая
Мускулатура пищеварительного тракта, мочевого пузыря, кровеносных и лимфатических сосудов и др. внутренних органов
Сократительная
Нервная – состоит из клеток с отростками. Способна возбуждаться и передавать возбуждение.
Нейроны + нейроглия
Головной и спинной мозг, нервные узлы, волокна
Согласованная работа всех систем органов, связь с окружающей средой
Слайды 28-30 — Проверочная работа:
Запишите номера суждений, напротив верных поставьте «+», напротив ошибочных — «—»:
1. Эпителий желудка и кишечника относится к эпителиальным тканям.
2. для эпителиальной ткани характерно слабое развитие межклеточного вещества.
3. для эпителиальной ткани характерны свойства возбудимости и проводимости.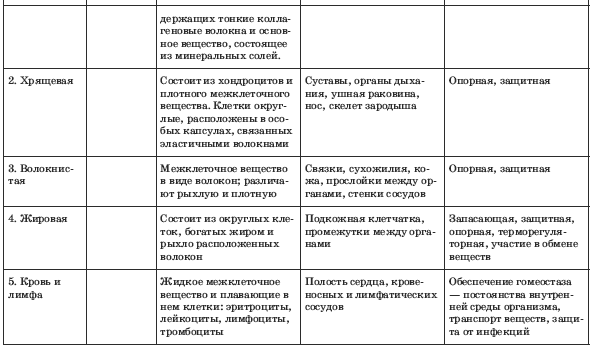
5. Эндотелий кровеносных сосудов относится к эпителиальной ткани.
6. Подкожная жировая клетчатка относится к эпителиальной ткани.
7. Для соединительных тканей характерно наличие хорошо развитого межклеточного вещества.
8. У соединительных тканей межклеточное вещество может быть твердым, жидким, эластичным.
9. К клеткам соединительной ткани относятся клетки крови, жировые клетки, клетки хряща.
10. Для мышечной ткани характерны свойства: возбудимость и сократимость.
11. Гладкая мышечная ткань входит в состав внутренних органов.
12. Поперечно-полосатая мышечная ткань образована мышечными клетками.
13. Сердечная мышца образована гладкой мышечной тканью.
14. Скелетные мышцы образованы мышечными волокнами, имеющими длину около 4 сантиметров, в каждом волокне сотни ядер находятся на периферии.
15. Возбуждение по аксону может идти только от тела нейрона.
16. По двигательному нейрону возбуждение от тела нейрона передается по аксону.
17. Нейрон всегда имеет только один аксон.
18. С одним нейроном могут контактировать более тысячи нервных клеток.
/По окончании работы проводится взаимопроверка/
Слайд 31 – Ответы:
Критерии оценивания:16-18 правильных ответов – «5»,
12-15 правильных ответов – «4»,
8-11 правильных ответов – «3»,
До 7 правильных ответов – «2».
Подведение итога урока. Выставление оценок
Слайд 32 — Домашнее задание: § 8 прочитать. Ответить устно на вопросы к параграфу. Закончить оформление таблицы схемами.
Вид ткани | Особенности строения | Функции | Местонахождение |
Эпителиальная ткань | |||
Покровный эпителий | Однослойный и многослойный Клетки плотно прилегают
друг к другу, практически отсутствует межклеточное вещество, нет кровеносных
сосудов. Однослойный — плоский, кубический, цилиндрический, мерцательный (есть реснички — колебательные движения, выстилает органы). | Защита от внешних воздействий, всасывание компонентов пищи, выведение продуктов обмена, подвижность органов. | Покровы тела, внутренние поверхности полых органов, полостей. |
Железистый эпителий | Клетки располагаются поодиночке или группами образуя органы – железы. Межклеточное вещество содержит кровеносные, лимфатические сосуды, нервные окончания. Железы внутренней секреции – лишены выводных протоков – вещества в кровь и лимфу. Железы внешней секреции – через протоки секрет на поверхность тела (потовые) или в полость (желудочные) | Выработка гормонов, пищеварительных ферментов, молока, слюны. | Ж внутренней секреции (гипофиз, щитовидная железа) Ж внешней секреции (слюнные, потовые, сальные) Ж смешанной секреции (поджелудочная, половая) |
Мышечные ткани | |||
Гладкая | Состоит из веретеновидных
одноядерных клеток, межклеточного вещества мало. | Сокращение без участия сознания, перистальтика ЖКТ, тонус сосудов, сокращение мочевого пузыря. | В стенках кровеносных сосудов, полых внутренних органов. |
Поперечнополосатая:
1) Скелетная
2) Сердечная | Имеет поперечную исчерченность за счет чередования участков с разными оптическими свойствами.
Вытянутые многоядерные мышечные волокна
Клетки с 1-2 ядрами |
Подконтрольна сознанию – физическая активность, натяжение голосовых связок, дыхание
Обладает автоматизмом – ритмичное сокращение |
Скелетная мускулатура,
мышцы языка и гортани.
Сердце |
Нервная ткань | |||
| Состоит из нейронов и клеток глии. Нейрон – тело с ядром и отростки. Слабоветвящийся отросток – аксон. Сильноветвящиеся многочисленные – дендриты. | Дендрит – прием и передача информации телу, аксон – от тела к другим клеткам. Клетки глии – снабжение питательными веществами, опора, защита. | Головной и спинной мозг, ганглии (нервные узлы), нервы. |
Соединительные ткани Сильно развито межклеточное вещество. Ткани не контактируют с внешней средой | |||
Кровь и лимфа | Жидкие ткани, межклеточное вещество жидкое, в нем располагаются форменные элементы (эритроциты, тромбоциты, лейкоциты) | Транспортировка веществ (кровь — глюкоза, кислород; лимфа – белки, вода, соли из тканей в кровь) | Внутреннее содержимое
полостей и сосудов, лимфатическая система. |
Рыхлая волокнистая соединительная ткань | Из клеток, разбросанных в межклеточном веществе и переплетенных неупорядоченных волокон. | Механическая (оболочка сосудов, нервов, органов) | Стенки кровеносных сосудов |
Плотная соединительная ткань | Состоит из волокон | Опорная, защитная | Связки и сухожилия |
Жировая | Почти всю цитоплазму занимает вакуоль | Защита органов от повреждений, накопление и хранение жиров, энергетическое обеспечение. | Под кожей и между внутренними органами |
Пигментная ткань | В клетках содержится меланин | Защита от УФ | Радужная оболочка глаза, кожа |
Хрящевая скелетная ткань | Из хрящевых клеток и большого количества плотного межклеточного вещества | Опора, защита | Хрящи ушных раковин, дыхательные пути |
Костная скелетная ткань | Костные пластинки, между которыми клетки | Опора, защита, обмен минеральных веществ (содержится почти 98% Ca от общего количества) | Кости скелета |



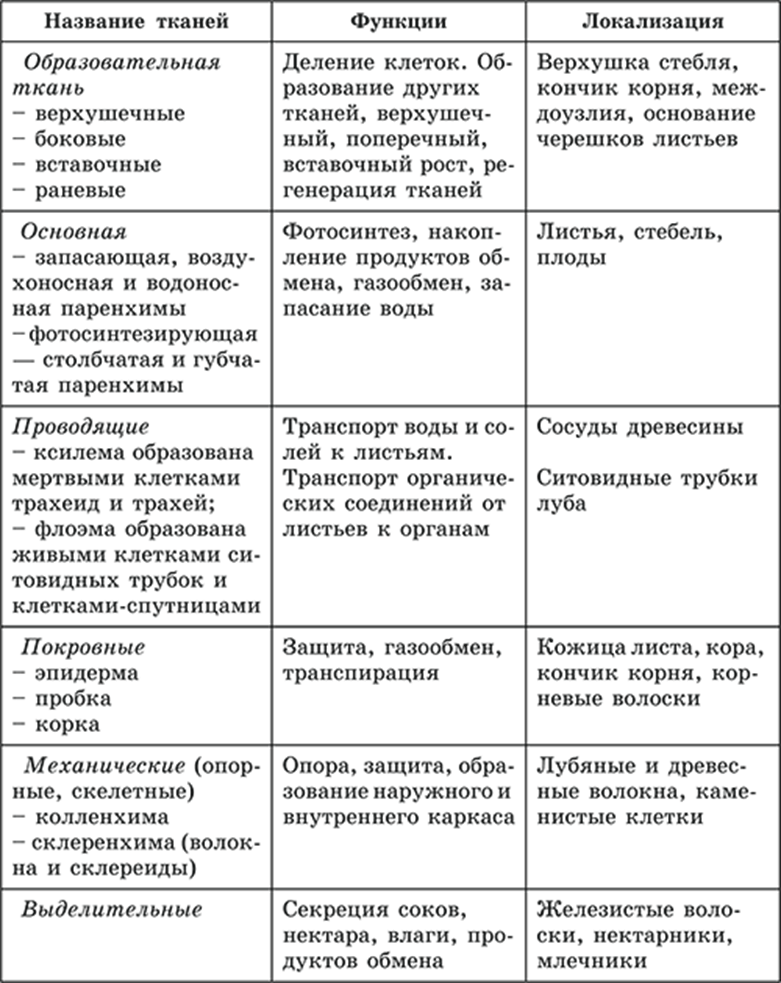
Тип тканей и месторасположение | Строение и жизнедеятельность клеток | Функции |
Образовательная Верхушка корня и конус нарастания стебля, в почке, камбий | — клетки круглые, мелкие; оболочки тонкие — крупные ядра, нет хлоропластов, вакуолей или они мелкие; — постоянное деление | рост органов и образование других тканей |
Основная: — запасающая — всасывающая — фотосинтезирующая. Мякоть листа, плодов. Середина корня, стебля Мягкие части цветка Главная масса коры | — Клетки округлые, крупные и средние с тонкими оболочками — клетки вытянутые тонкостенные, — содержат хлоропласты; — ядро у стенки, крупные вакуоли, много хлоропластов; — хорошо развито межклеточное вещество; — образуются органические вещества из неорганических — превращение и накопление веществ. | — запас питательных веществ (крахмал, белки, сахар) — поглощение воды с минеральными веществами — фотосинтез |
Механическая: Жилки листа Волокна стебля Косточка абрикоса Скорлупа грецкого ореха | — мёртвые и живые клетки вытянутые, их оболочка утолщена, одревесневела; — цитоплазма часто отсутствует | опора, упругость, эластичность |
Покровная: Кожица (с устьицами) листа, пробка (с чечевичками) дерева кора | — мелкие и средние живые клетки с тонкими оболочками, плотно прилегают друг к другу; — мёртвые клетки, оболочка одревесневает, оболочки толстые | защита связь с внешней средой (газообмен) |
Проводящая Жилки листа, в корне, стебле | — сосуды – мёртвые вытянутые клетки с разрушенными поперечными оболочками — ситовидные трубки – живые клетки без ядра, вакуолей и пластид, между ними есть отверстия | передвижение воды и минеральных веществ по древесине снизу вверх передвижение органических веществ по коре от листьев вниз |
Урок 7.
 Ткани | Поурочные планы по биологии 6-7 класс
Ткани | Поурочные планы по биологии 6-7 классУрок 7. Ткани
07.01.2015 5908 0Цели урока: Сформировать у учащихся понятие о тканях, их многообразии и значении; дать представление о взаимосвязи строения ткани и выполняемых ею функций.
Оборудование: Таблица «Виды тканей животных».
I. Актуализация знаний
Биологический диктант.
1) Наука, изучающая строение и функции клеток — ….
2) Основная структурно — функциональная единица всех живых организмов, элементарная живая система —
3) Тело клетки и ее содержимое отделено от внешней среды — ….
4)
Основное
содержимое клетки, заполняющее весь ее объем —. …
…
5) Органоид клетки, в котором происходит синтез белков —….
6) Главнейшая структура ядра, обеспечивающая передачу наследственной информации —….
7) Впервые клетки были обнаружены в 1665 году ….
8) Основное содержимое клетки, заполняющее весь ее объем —….
9) В клетках животных, в отличие от растительных, отсутствуют….
10) Органоиды клетки, ферменты которых расщепляют белки, жиры, углеводы —….
II. Изучение нового материала
1. Рассказ учителя.
У многоклеточных животных клетки специализируются на выполнении определенных функций. Группа клеток, сходных по строению, происхождению и выполняемым функциям, называется тканью.
— Какие типы тканей встречаются у растений, какие функции они выполняют?
Наука, изучающая ткани, называется гистологией.
Дифференцировка клеток на различные типы тканей это своего рода разделение труда в организме. Такое разделение труда выгодно для организма, поскольку ткани выполняют специфические функции эффективнее отдельных клеток. Такая специализация повышает эффективность работы всего организма в целом. Работа клеток в тканях скоординирована и они представляют собой самостоятельную функциональную единицу. Кроме того, совместная деятельность различных тканей тоже строго скоординирована, так как только таким образом организм может сохранить свою жизнеспособность.
В теле животных выделяют четыре вида тканей: эпителиальная (покровная), соединительная, мышечная, нервная. Каждый из видов ткани имеет свое характерное строение и выполняет в организме свою строго определенную функцию. Внешний вид и строение ткани четко соответствуют ее назначению.
2. Самостоятельная работа учащихся с учебником.
О
Заполнить таблицу, пользуясь материалом учебника §7, стр. 28-30.
28-30.
Название ткани | Строение | Функции | Примеры |
Эпителиальная |
|
|
|
Соединительная |
|
|
|
Мышечная |
|
|
|
Нервная |
|
|
|
III.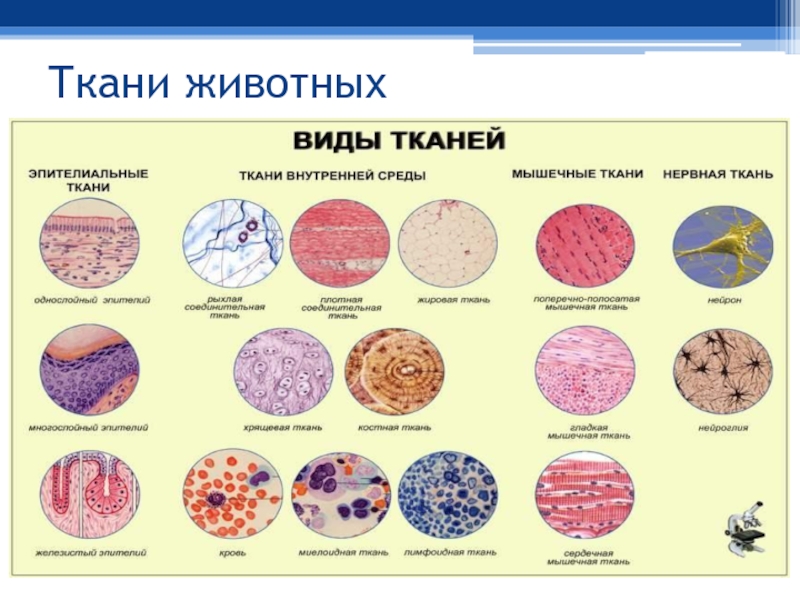 Закрепление знаний
Закрепление знаний
1. Фронтальный опрос.
— Что такое ткань? Какие виды тканей вы знаете?
— Какова функция эпителиальной ткани? Где может быть расположена эпителиальная ткань?
— Какова функция сальных желез?
— Какие еще железы могут встречаться у животных?
— Какие элементы организма состоят из соединительной ткани?
— Какова функция соединительной ткани?
— Какие вы знаете виды мышечной ткани?
— Какова особенность клеток нервной ткани?
2. Игра «Слабое звено».
Учитель называет орган или структурный элемент
организма и указывает на одного из учеников. Ученик должен назвать ткань, из
которой состоит данная структура и назвать следующую, указав на другого
ученика (или назвав его имя).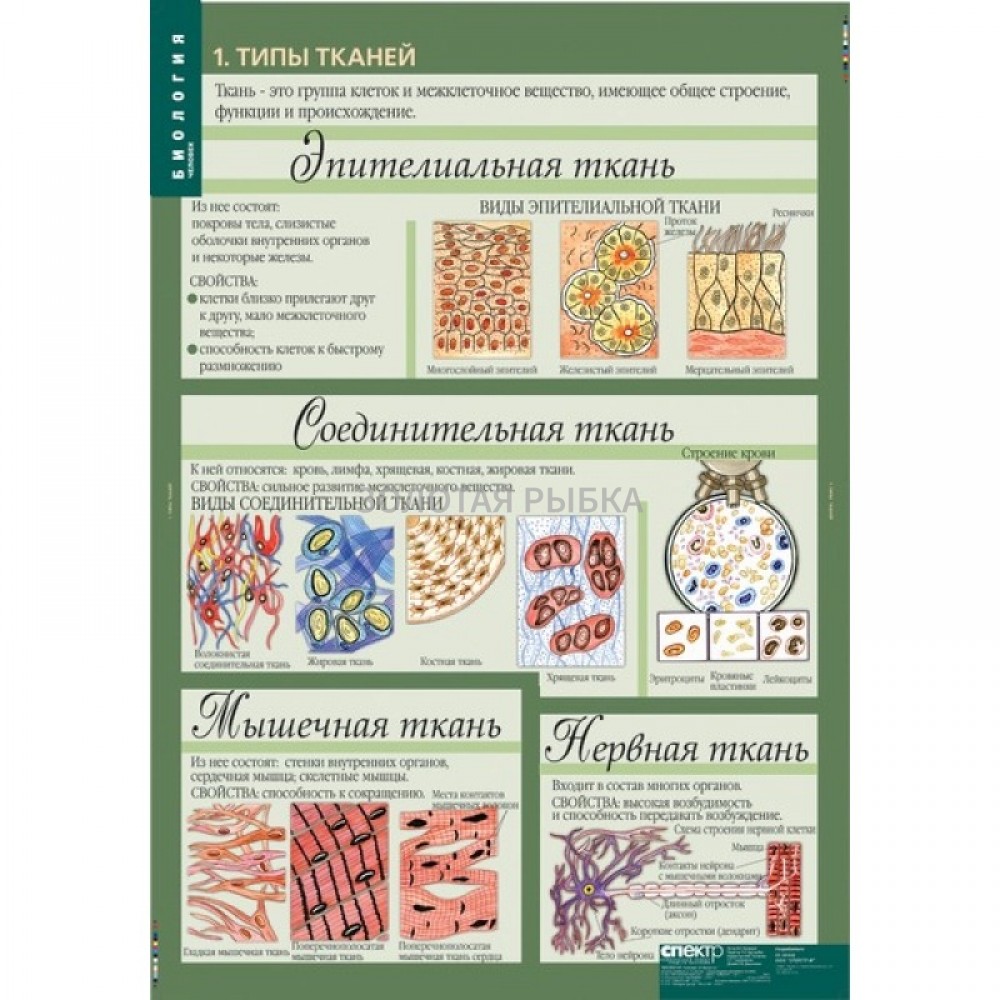 Если какой-либо ученик затрудняется ответить,
учитель сам отвечает на вопрос и дает задание другому ученику. Задача игры —
составить как можно более длинную цепочку.
Если какой-либо ученик затрудняется ответить,
учитель сам отвечает на вопрос и дает задание другому ученику. Задача игры —
составить как можно более длинную цепочку.
Домашнее задание
§7 учебника, вопросы в конце параграфаТкани, органы и системы органов. Типы тканей рестений
Стр. 27
Вспомните
1.Какие типы тканей характерны для растений?
Образовательные, покровные, основные, проводящие, механические.
2.Какие органы имеются у растений, и какие функции они выполняют?
Цветок — половое размножение, образование плода и семян.
Лист — фотосинтез, газообмен, испарение воды.
Стебель — проводник, опора, запасает питательные вещества, фотосинтез.
Корни — опора, минеральное питание, вегетативное размножение.
Стр. 32
32
Вопросы после параграфа
1.Выделите существенные признаки ткани.
Ткань — группа клеток сходных по строению, происхождению и выполняющих определенную функцию.
2.Используя материал параграфа, составьте таблицу «Характеристика типов тканей». Кратко укажите особенности строения и функции каждого из типов тканей.
3.Докажите существование взаимосвязи строения и функций организмов на примере какой — либо системы органов.
Опорно-двигательная система выполняет опорную и двигательную функции, а так же защитную функцию. Особенно выраженной функцией обладают череп у позвоночных животных и панцирь у рака, скорпиона, и некоторых насекомых.
4.Обоснуйте взаимосвязь типа симметрии животного образа жизни.
Двусторонняя симметрия — активно передвигающиеся животные.
Радиальная симметрия — неподвижный образ жизни.
5.Используя рисунок 18, составьте с помощью компьютера таблицу «Основные системы органов многоклеточных животных» с пустыми графами. Вы также можете приготовить её бумажный вариант. Для этого выделите несколько разворотов в тетради или возьмите отдельные листы бумаги. В таблицу вы будете заносить сведения о строении многоклеточных организмов, которые вам предстоит изучать в течение года. Эти записи помогут вам готовиться к контрольным работам и к экзаменам в старших классах.
Оборот стабильных изотопов и период полураспада в тканях животных: синтез литературы
Abstract
Стабильные изотопы углерода, азота и серы используются в качестве экологических индикаторов для различных приложений, таких как исследования миграций животных, источников энергии и путей пищевых цепей. Тем не менее, неопределенность, относящаяся к периоду времени, интегрированному с помощью изотопных измерений тканей животных, может затруднить интерпретацию изотопных данных. Было проведено большое количество экспериментальных исследований изменения изотопной диеты, направленных на количественную оценку скорости изотопного оборота в тканях животных λ (% · день –1 , часто выражаемое как период полураспада изотопов, ln (2) / λ, дни).Тем не менее, ни в одном исследовании не проводилась оценка или обобщение многих индивидуальных оценок периода полураспада с целью как поиска широкомасштабных закономерностей, так и характеристики степени изменчивости. Здесь мы собираем ранее опубликованные оценки периода полураспада, изучаем, как период полураспада связан с размером тела, и тестируем аллометрические отношения, варьирующиеся от тканей и таксонов. Период полураспада обычно увеличивается с увеличением массы тела животного и дольше в мышцах и крови по сравнению с плазмой и внутренними органами. Период полураспада был самым продолжительным у экотерм, за ними следовали млекопитающие и, наконец, птицы.Что касается эктотермов, разные комбинации таксонов и тканей имели сходные аллометрические наклоны, которые в целом совпадали с предсказаниями метаболической теории.
Было проведено большое количество экспериментальных исследований изменения изотопной диеты, направленных на количественную оценку скорости изотопного оборота в тканях животных λ (% · день –1 , часто выражаемое как период полураспада изотопов, ln (2) / λ, дни).Тем не менее, ни в одном исследовании не проводилась оценка или обобщение многих индивидуальных оценок периода полураспада с целью как поиска широкомасштабных закономерностей, так и характеристики степени изменчивости. Здесь мы собираем ранее опубликованные оценки периода полураспада, изучаем, как период полураспада связан с размером тела, и тестируем аллометрические отношения, варьирующиеся от тканей и таксонов. Период полураспада обычно увеличивается с увеличением массы тела животного и дольше в мышцах и крови по сравнению с плазмой и внутренними органами. Период полураспада был самым продолжительным у экотерм, за ними следовали млекопитающие и, наконец, птицы.Что касается эктотермов, разные комбинации таксонов и тканей имели сходные аллометрические наклоны, которые в целом совпадали с предсказаниями метаболической теории. Период полураспада для эктотермов можно приблизительно рассчитать следующим образом: ln (период полураспада) = 0,22 * ln (масса тела) + перехват, зависящий от группы; n = 261, p <0,0001, r 2 = 0,63. Для эндотермических групп связь с массой тела была слабой, а уклоны и пересечения моделей были неоднородными. Хотя период полураспада изотопов может быть приблизительно определен с использованием простых аллометрических соотношений для некоторых таксонов и типов тканей, в наших моделях также наблюдается высокая степень необъяснимых вариаций.Наше исследование подчеркивает несколько сильных и общих закономерностей, хотя точное предсказание периода полураспада изотопов на основе легко доступных переменных, таких как масса тела животного, остается неуловимым.
Период полураспада для эктотермов можно приблизительно рассчитать следующим образом: ln (период полураспада) = 0,22 * ln (масса тела) + перехват, зависящий от группы; n = 261, p <0,0001, r 2 = 0,63. Для эндотермических групп связь с массой тела была слабой, а уклоны и пересечения моделей были неоднородными. Хотя период полураспада изотопов может быть приблизительно определен с использованием простых аллометрических соотношений для некоторых таксонов и типов тканей, в наших моделях также наблюдается высокая степень необъяснимых вариаций.Наше исследование подчеркивает несколько сильных и общих закономерностей, хотя точное предсказание периода полураспада изотопов на основе легко доступных переменных, таких как масса тела животного, остается неуловимым.
Образец цитирования: Vander Zanden MJ, Clayton MK, Moody EK, Solomon CT, Weidel BC (2015) Стабильный оборот изотопов и период полураспада в тканях животных: синтез литературы. PLoS ONE 10 (1):
e0116182. https://doi.org/10.1371/journal.pone.0116182
https://doi.org/10.1371/journal.pone.0116182
Академический редактор: Дэвид Уильям Понд, Шотландская ассоциация морских наук, СОЕДИНЕННОЕ КОРОЛЕВСТВО
Поступило: 16 сентября 2014 г .; Одобрена: 4 декабря 2014 г .; Опубликован: 30 января 2015 г.
Это статья в открытом доступе, свободная от всех авторских прав, и ее можно свободно воспроизводить, распространять, передавать, изменять, строить или иным образом использовать в любых законных целях.Работа сделана доступной по лицензии Creative Commons CC0, выделенная в общественное достояние
Доступность данных: Все соответствующие данные находятся в документе и файлах с вспомогательной информацией.
Финансирование: Авторы не получали специального финансирования на эту работу.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.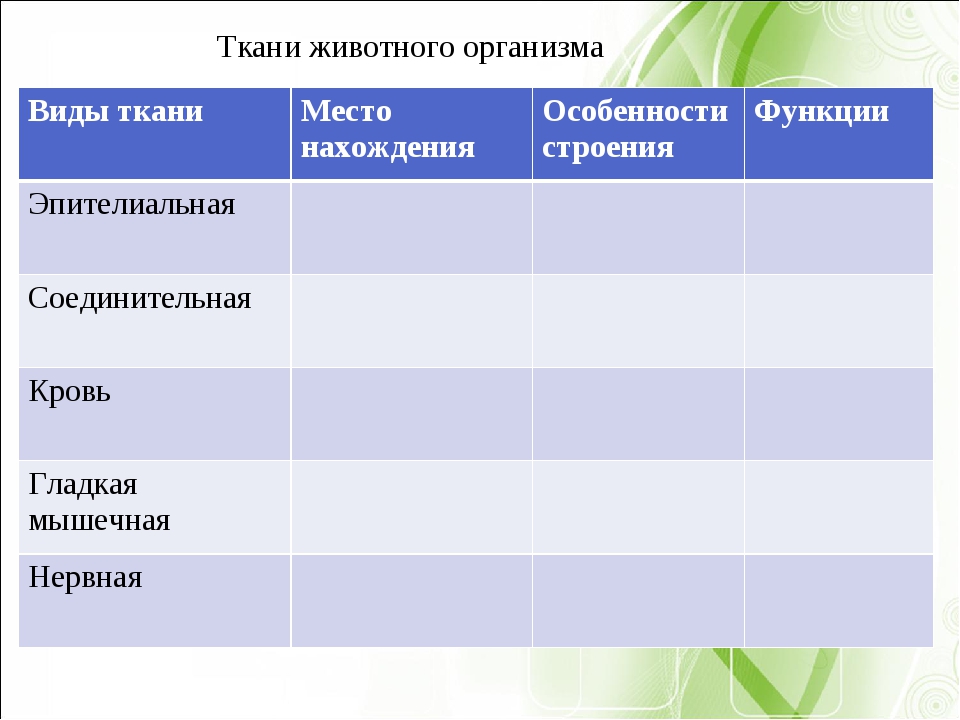
Введение
Естественная изменчивость стабильных изотопных соотношений углерода, азота и серы (δ 13 C, δ 15 N, δ 34 S) широко используются в экологии животных, включая исследования миграции животных, трофические сети, оценка трофического положения и зависимость от источников пищи [1–3].Использование стабильных изотопов в исследованиях пищевых сетей основывается на понимании взаимосвязи изотопного состава рациона животных и тканей. В этом есть два основных аспекта. Во-первых, это факторы различения диеты и ткани (иногда называемые трофическим фракционированием). Обобщены закономерности и вариабельность факторов различения диеты и ткани [4–6]. Вторая связана с временной изотопной динамикой — в частности, с идеей о том, что ткань животного не сразу отражает изотопный состав ее рациона, а, скорее, интегрируется в течение некоторого периода времени.Примечательно, что многие полевые исследования стабильных изотопов, такие как те, которые используют модели смешения изотопов [7,8], неявно предполагают, что изотопный состав тканей животных находится в равновесии с диетой (т. е. предполагает устойчивое состояние диета-ткань). Очевидно, что во многих ситуациях это не так, и это может привести к весьма вводящим в заблуждение интерпретациям пищевой сети [9].
е. предполагает устойчивое состояние диета-ткань). Очевидно, что во многих ситуациях это не так, и это может привести к весьма вводящим в заблуждение интерпретациям пищевой сети [9].
Фрай и Арнольд [10] были первым исследованием, в котором количественно определена скорость включения изотопов в ткани животных. Путем изменения изотопного состава углерода рациона выращенных в лаборатории креветок они количественно определили временную шкалу включения изотопов и расчетную скорость изотопного оборота (λ,% · день -1 ) и период полураспада изотопов (ln (2) / λ, дни), определяемое как время, необходимое для достижения 50% равновесия с диетой.Фрай и Арнольд также отметили, что изотопный оборот происходит в результате двух различных процессов: роста ткани и катаболического обмена. Последующие лабораторные исследования позволили оценить изотопный оборот для самых разных животных и типов тканей [1,11]. Эти исследования выявили тканеспецифические различия, например, внутренние органы и плазма крови, как правило, имеют высокие скорости включения изотопов по сравнению с мышечной тканью и клетками крови [11]. Поскольку ткани объединяют рационы потребителей в разных временных масштабах, исследование нескольких тканей потенциально может предоставить информацию о временной динамике использования ресурсов [12].
Поскольку ткани объединяют рационы потребителей в разных временных масштабах, исследование нескольких тканей потенциально может предоставить информацию о временной динамике использования ресурсов [12].
Интерпретация изотопной ценности ткани из полевых исследований должна учитывать, по крайней мере в общем смысле, скорость включения изотопов. Что мы в настоящее время знаем о временном масштабе включения изотопов для различных тканей и животных? Различаются ли скорости включения изотопов систематически среди таксонов, типов тканей или элементов (δ 13 C, δ 15 N, δ 34 S)? Насколько он изменчив и в какой степени мы можем сделать общие выводы при отсутствии информации об инкорпорации изотопов, специфичной для таксофона или системы? За последние несколько десятилетий в большом количестве экспериментов по изменению изотопной диеты был оценен период полураспада изотопов для различных таксонов животных и типов тканей.Количественный синтез этих результатов изотопного периода полураспада был недавно определен как «плодотворная и, возможно, срочная задача» в недавнем обзоре [1]. Несколько недавних исследований суммировали период полураспада изотопов для конкретных таксонов и типов тканей [13–16]. В наиболее исчерпывающем на сегодняшний день синтезе [11] изучались различия между типами тканей для трех основных таксонов животных (птицы, млекопитающие, рыбы), но не учитывалась роль размера тела. Можно было бы ожидать, что изотопный оборот сильно зависит от размера тела, при этом ткани мелких животных интегрируются в течение короткого периода времени по сравнению с тканями крупных животных.Основание для роли размера тела было формализовано Карлтоном и Мартинесом дель Рио [15], которые отметили, что скорость обмена белков обычно пропорциональна массе тела примерно в 3/4 степени [17]. Поскольку масса отдельных тканей примерно пропорциональна массе тела, можно ожидать, что изотопный оборот примерно пропорционален массе тела -1/4 (т.е. массе 3/4 / масса). Аллометрические исследования периода полураспада изотопов для конкретных таксонов и типов тканей в целом соответствовали этому предсказанному наклону ~ 0.
Несколько недавних исследований суммировали период полураспада изотопов для конкретных таксонов и типов тканей [13–16]. В наиболее исчерпывающем на сегодняшний день синтезе [11] изучались различия между типами тканей для трех основных таксонов животных (птицы, млекопитающие, рыбы), но не учитывалась роль размера тела. Можно было бы ожидать, что изотопный оборот сильно зависит от размера тела, при этом ткани мелких животных интегрируются в течение короткого периода времени по сравнению с тканями крупных животных.Основание для роли размера тела было формализовано Карлтоном и Мартинесом дель Рио [15], которые отметили, что скорость обмена белков обычно пропорциональна массе тела примерно в 3/4 степени [17]. Поскольку масса отдельных тканей примерно пропорциональна массе тела, можно ожидать, что изотопный оборот примерно пропорционален массе тела -1/4 (т.е. массе 3/4 / масса). Аллометрические исследования периода полураспада изотопов для конкретных таксонов и типов тканей в целом соответствовали этому предсказанному наклону ~ 0.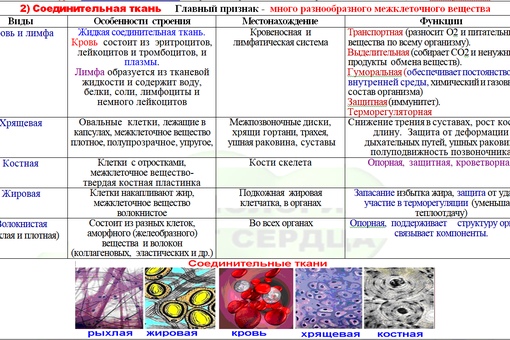 25 [14,15]. В этом исследовании мы собрали обширную коллекцию опубликованных оценок периода полураспада изотопов и проверили, изменяется ли период полураспада в зависимости от массы тела, как это предсказано в теории [1,18]. Мы также проверяем, существуют ли различия в аллометических отношениях между таксонами, типами тканей и тремя широко используемыми изотопами (δ 13 C, δ 15 N и δ 34 S). Срочно необходимо общее понимание того, как период полураспада изотопов изменяется в зависимости от массы тела для различных типов тканей и таксонов, что поможет при интерпретации изотопных данных различных лабораторных и полевых исследований.
25 [14,15]. В этом исследовании мы собрали обширную коллекцию опубликованных оценок периода полураспада изотопов и проверили, изменяется ли период полураспада в зависимости от массы тела, как это предсказано в теории [1,18]. Мы также проверяем, существуют ли различия в аллометических отношениях между таксонами, типами тканей и тремя широко используемыми изотопами (δ 13 C, δ 15 N и δ 34 S). Срочно необходимо общее понимание того, как период полураспада изотопов изменяется в зависимости от массы тела для различных типов тканей и таксонов, что поможет при интерпретации изотопных данных различных лабораторных и полевых исследований.
Материалы и методы
Мы провели поиск литературы с помощью ISI Web of Science (поисковые запросы: углерод, азот, сера, стабильный изотоп, оборот, период полураспада) для исследований, которые оценивали или содержали данные, которые могли быть использованы для оценки углерода. , период полураспада изотопов азота и серы (λ) для любых таксонов животных и типов тканей.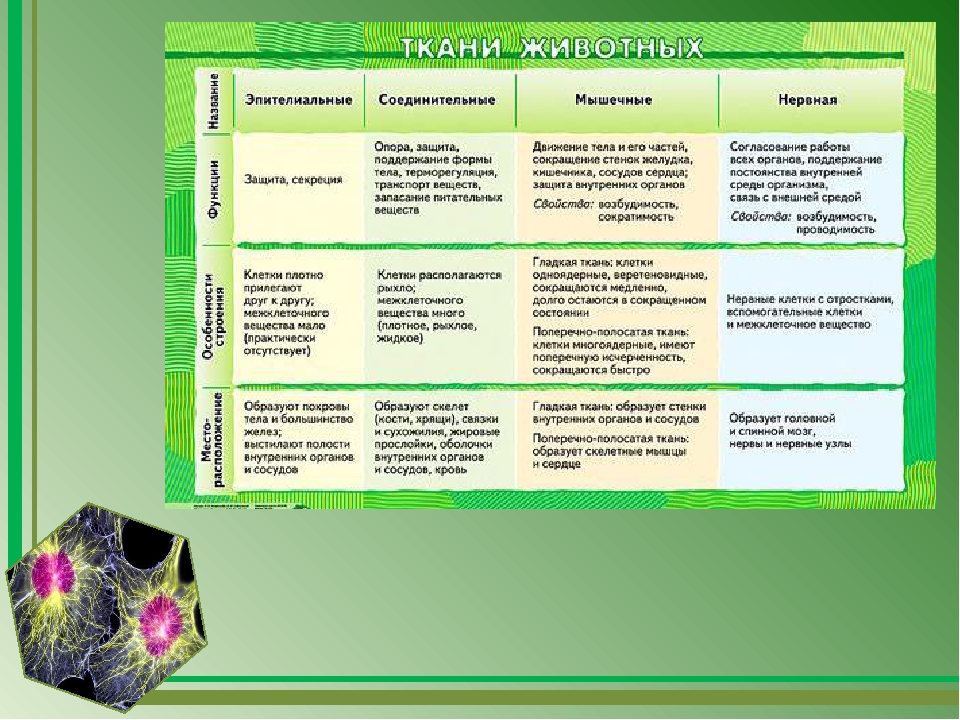 Разделы «Ссылки» соответствующих исследований были использованы для определения дополнительных исследований для включения.
Разделы «Ссылки» соответствующих исследований были использованы для определения дополнительных исследований для включения.
Скорость обмена изотопов может быть оценена путем моделирования соотношений стабильных изотопов в тканях как функции времени или массы тела с использованием одной из нескольких базовых схем моделирования (текст S1): Hobson and Clark [19], Hesslein et al.[20], а также Фрай и Арнольд [10], все из которых обычно дают аналогичные результаты, но различаются по их применимости в зависимости от ситуации (т.е. растущие животные по сравнению с нерастущими). Несколько недавних исследований оценили, лучше ли мультикомпартментные модели описывают изотопный оборот, чем однокамерные модели [21,22]. Результаты были неоднозначными. Для этих исследований мы использовали оценку оборачиваемости из подхода, который лучше всего подходил для описания изотопных данных из теоретико-информационного подхода (AIC).
Животные, включенные в это исследование, имеют массу тела примерно 10 порядков (белоногие креветки, 0,009 мг коровам, 493 кг).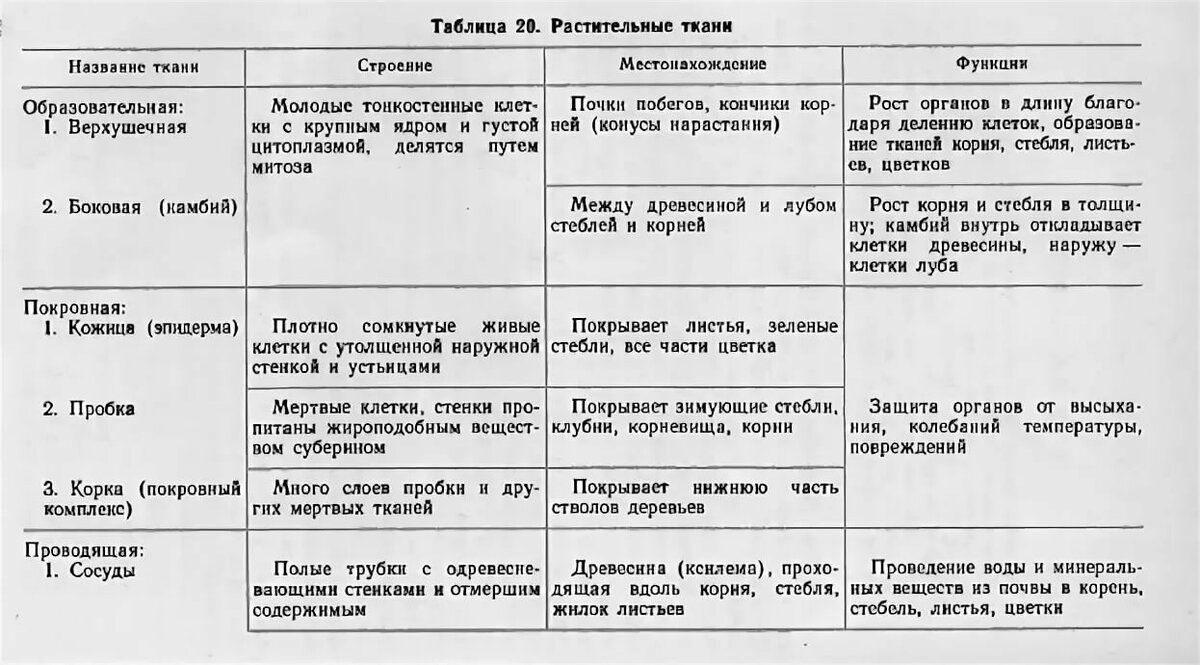 Мы собрали оценки оборота для широкого диапазона типов тканей. Оценки оборота были разделены на пять групп тканей: мышцы, все тело, кровь, плазма крови и внутренние органы. Следующие типы тканей либо имели небольшой размер выборки, либо не соответствовали нашей структуре и поэтому были исключены из дальнейшего анализа: жабры, костный коллаген, гонады, плавники, глаза, волосы, чешуя, кожа и ткани щитков.Группа тканей «внутренние органы» была довольно широкой и включала следующие типы тканей: печень, мозг, почки, поджелудочная железа, селезенка, легкие, желудочно-кишечный тракт и сердце. Дальнейшая оценка гетерогенности изотопного обмена среди типов органов не выявила общего эффекта размера тела и высокой степени перекрытия между различными внутренними органами. Для целей этого анализа мы рассматривали эти различные органы как единую группу, хотя признавали, что отдельные типы внутренних органов могут различаться в отношении изотопного обмена.Наш окончательный набор данных включал 486 оценок изотопного круговорота 86 видов, взятых из 85 отдельных рецензируемых исследований, опубликованных за 1982–2014 гг.
Мы собрали оценки оборота для широкого диапазона типов тканей. Оценки оборота были разделены на пять групп тканей: мышцы, все тело, кровь, плазма крови и внутренние органы. Следующие типы тканей либо имели небольшой размер выборки, либо не соответствовали нашей структуре и поэтому были исключены из дальнейшего анализа: жабры, костный коллаген, гонады, плавники, глаза, волосы, чешуя, кожа и ткани щитков.Группа тканей «внутренние органы» была довольно широкой и включала следующие типы тканей: печень, мозг, почки, поджелудочная железа, селезенка, легкие, желудочно-кишечный тракт и сердце. Дальнейшая оценка гетерогенности изотопного обмена среди типов органов не выявила общего эффекта размера тела и высокой степени перекрытия между различными внутренними органами. Для целей этого анализа мы рассматривали эти различные органы как единую группу, хотя признавали, что отдельные типы внутренних органов могут различаться в отношении изотопного обмена.Наш окончательный набор данных включал 486 оценок изотопного круговорота 86 видов, взятых из 85 отдельных рецензируемых исследований, опубликованных за 1982–2014 гг. (Рис. 1; таблица S1).
(Рис. 1; таблица S1).
Там, где это возможно, мы пересчитали полученные оценки периода полураспада, чтобы проверить указанные значения, и рассчитали период полураспада, если значения оборачиваемости или полураспада не были указаны. Большинство оценок было получено на основе лабораторных экспериментов по смене диеты или экспериментов по поглощению / очистке изотопных меток. Мы также включили небольшое количество полевых ситуаций, которые очень четко имитировали экспериментальную смену диеты [14,23,24].Изотопный оборот λ был выражен как период полужизни изотопов, ln (2) / λ, который представляет собой время (в днях), необходимое для 50% -ного уравновешивания с экспериментальной диетой. Мы решили представить результаты как период полураспада изотопов, потому что они более легко интерпретируемы и интуитивно понятны, чем изотопный оборот.
Значения температуры представляют собой зарегистрированную среднюю температуру окружающей среды, в которой животное содержалось во время исследования смены рациона. В начале исследования смены диеты мы использовали массу животных в качестве индикатора массы тела.Возможно, это несовершенный показатель, но он был единственным постоянно сообщаемым показателем массы тела животных в первоначальных исследованиях. Для нескольких исследований, в которых не сообщалось о начальной массе тела экспериментальных животных, это значение было оценено из литературы для одного и того же или близкородственных видов на одной и той же стадии жизненного цикла. Мы исключили оценки периода полураспада, по которым не хватало информации для оценки массы тела.
В начале исследования смены диеты мы использовали массу животных в качестве индикатора массы тела.Возможно, это несовершенный показатель, но он был единственным постоянно сообщаемым показателем массы тела животных в первоначальных исследованиях. Для нескольких исследований, в которых не сообщалось о начальной массе тела экспериментальных животных, это значение было оценено из литературы для одного и того же или близкородственных видов на одной и той же стадии жизненного цикла. Мы исключили оценки периода полураспада, по которым не хватало информации для оценки массы тела.
Статистический подход
Мы предположили, что период полураспада изотопов будет увеличиваться в зависимости от массы тела животного.Кроме того, мы ожидали, что период полураспада и взаимосвязь с массой тела потенциально будут зависеть от таких факторов, как таксон, тип ткани, изотоп и температура. Небольшие размеры выборки для определенных таксонов и типов тканей, а также ограниченный диапазон размеров тела для некоторых таксонов ограничивали наши возможности по разработке надежных эмпирических моделей, которые широко применимы ко всем таксонам и тканям. Мы разработали модели периода полураспада, которые использовали комбинации типа ткани и таксона в качестве категориальной переменной и массу тела в качестве ковариаты в рамках ANCOVA.Мы создали таблицу (матрицу) сопряженности 5×4 наших ключевых категориальных переменных: тип ткани (мышцы, все тело, внутренние органы, кровь, плазма крови) и таксон (беспозвоночные, эктотермия позвоночных, птицы, млекопитающие). Комбинации тип ткани-таксон (далее называемые «группами») широко различались по размеру выборки, при этом несколько комбинаций ткань-таксон представлены небольшими оценками периода полураспада или отсутствием их. Группы с четырьмя или менее оценками периода полураспада были исключены из сравнений по группам, поскольку считалось, что это будет абсолютный минимальный размер выборки, необходимый для разумной оценки наклона и пересечения в рамках ANCOVA.Перед анализом значения массы тела и периода полураспада преобразовали в натуральный логарифм. Статистический анализ проводился в SAS v 9.
Мы разработали модели периода полураспада, которые использовали комбинации типа ткани и таксона в качестве категориальной переменной и массу тела в качестве ковариаты в рамках ANCOVA.Мы создали таблицу (матрицу) сопряженности 5×4 наших ключевых категориальных переменных: тип ткани (мышцы, все тело, внутренние органы, кровь, плазма крови) и таксон (беспозвоночные, эктотермия позвоночных, птицы, млекопитающие). Комбинации тип ткани-таксон (далее называемые «группами») широко различались по размеру выборки, при этом несколько комбинаций ткань-таксон представлены небольшими оценками периода полураспада или отсутствием их. Группы с четырьмя или менее оценками периода полураспада были исключены из сравнений по группам, поскольку считалось, что это будет абсолютный минимальный размер выборки, необходимый для разумной оценки наклона и пересечения в рамках ANCOVA.Перед анализом значения массы тела и периода полураспада преобразовали в натуральный логарифм. Статистический анализ проводился в SAS v 9. 3 (Кэри, Северная Каролина, США).
3 (Кэри, Северная Каролина, США).
Результаты
Широкие узоры
В наиболее всеобъемлющем анализе, который включает оценки периода полураспада для всех изотопов (δ 13 C, δ 15 N, δ 34 S), таксонов и типов тканей, период полураспада увеличивается в зависимости от животных масса тела (рис. 2А):
л (период полураспада) = 0,11 * л (масса тела) + 2.66, n = 486, p <0,0001, F = 117,02, RMSE = 1,03, r 2 = 0,19 (уравнение 1).
Рис. 2. Связь между ln (масса тела животного, граммы) и ln (период полураспада изотопов, дни).
A) График для всех типов тканей и таксономических групп вместе взятых. B – E) Отмечены отдельно для каждого таксона. F – J) Графики нанесены отдельно для каждого типа ткани. Информация о регрессии для каждого типа ткани и таксона представлена в таблице 1.
https://doi.org/10.1371/journal.pone.0116182.g002
Добавление «изотопа» (δ 13 C, δ 15 N, δ 34 S) в качестве категориальной переменной к вышеприведенной модели выявило предельное значение (p = 0,06) и отсутствие значимого изотопа массы тела. взаимодействие (p = 0,29). Включение «изотопа» не привело к заметному снижению ошибки модели (RMSE = 1,02). В результате оценки периода полураспада для этих трех стабильных изотопов в дальнейшем были объединены.
взаимодействие (p = 0,29). Включение «изотопа» не привело к заметному снижению ошибки модели (RMSE = 1,02). В результате оценки периода полураспада для этих трех стабильных изотопов в дальнейшем были объединены.
Мы исследовали период полураспада как функцию массы тела отдельно для каждого широко определенного таксона (рис.2B – E) и тип ткани (рис. 2F – J) и представляют индивидуальные регрессионные модели для широких таксонов и типов тканей (таблица 1). Чтобы изучить более тонкие закономерности среди широких таксонов и типов тканей, мы разделили оценки периода полураспада на группы на основе комбинаций ткань-таксон (далее именуемые «группой»; Таблица 2).
Взаимосвязь между массой тела и периодом полураспада для эктотермов
Для эктотермов модель, прогнозирующая период полураспада с использованием массы тела и комбинации ткани-таксон (группа), выявила значительные эффекты массы тела (p <0.0001) и «группа» (p <0,0001), но без значимого члена взаимодействия (p = 0,28). Таким образом, модель, в которой группы имеют общий аллометрический наклон и групповые точки пересечения (таблица 3), описывает данные: ln (период полураспада) = 0,22 * ln (масса тела) + [групповая точка пересечения из таблицы 3]; n = 261, p <0,0001, F = 634,00, RMSE = 0,76, r 2 = 0,63 (уравнение 2).
Таким образом, модель, в которой группы имеют общий аллометрический наклон и групповые точки пересечения (таблица 3), описывает данные: ln (период полураспада) = 0,22 * ln (масса тела) + [групповая точка пересечения из таблицы 3]; n = 261, p <0,0001, F = 634,00, RMSE = 0,76, r 2 = 0,63 (уравнение 2).
Попарное сравнение перехватов и связанных стандартных ошибок показало, что многие групповые перехваты статистически не отличались друг от друга, давая три существенно разных и различных кластера (обозначенных номерами в верхнем индексе 1-3 в Таблице 3).Плазма и органы эктотерма позвоночных имели самые низкие значения перехватов (т.е. самый низкий период полураспада) и существенно не отличались друг от друга («1» в таблице 3). Вторую группу составили кровь и мышцы эктотерм позвоночных, а также мышцы и все тело беспозвоночных («2» в таблице 3). Эктотерм всего тела позвоночных было последней группой («3» в Таблице 3).
В качестве альтернативы использованию отдельных групп-специфичных перехватов (уравнение 2) мы объединили группы, которые существенно не отличались друг от друга, для упрощенной модели периода полураспада для эктотерм: ln (период полураспада) = 0. 21 * ln (масса тела) + [2,47 (эктотермия плазмы и органов позвоночных), 3,23 (эктотермия крови и мышц позвоночных, мышцы беспозвоночных и все тело), 3,60 (эктотерм позвоночных всего тела)]; n = 261, p <0,0001, F = 1276,3, RMSE = 0,76, r 2 = 0,63 (уравнение 3). Сравнение наблюдаемых и прогнозируемых значений периода полураспада для эктотерм (из уравнения 3) показывает, что прогнозируемые значения попадают в линию 1: 1 (рис. 3).
21 * ln (масса тела) + [2,47 (эктотермия плазмы и органов позвоночных), 3,23 (эктотермия крови и мышц позвоночных, мышцы беспозвоночных и все тело), 3,60 (эктотерм позвоночных всего тела)]; n = 261, p <0,0001, F = 1276,3, RMSE = 0,76, r 2 = 0,63 (уравнение 3). Сравнение наблюдаемых и прогнозируемых значений периода полураспада для эктотерм (из уравнения 3) показывает, что прогнозируемые значения попадают в линию 1: 1 (рис. 3).
Наконец, учитывая, что метаболизм эктотерма и широкий диапазон других скоростей сильно зависят от температуры [18], мы проверили, объясняет ли температура какие-либо дополнительные различия в соотношении масса тела-период полураспада для эктотерм.Влияние температуры было слабым, но статистически значимым (p = 0,02) и имело небольшое влияние RMSE модели (уменьшилось с 0,76 до 0,73). В целом наши результаты указывают на относительно незначительное влияние температуры на период полураспада изотопов для эктотерм.
Соотношение масса тела и период полураспада для эндотермов
Для эндотерм существует слабая положительная связь между периодом полураспада и массой тела: ln (период полураспада) = 0,13 * ln (масса тела) + 2,01; n = 220, p <0,0001, F = 24,15, RMSE = 0. 87, г 2 = 0,10 (4)
87, г 2 = 0,10 (4)
Сравнение между группами выявило неоднородные наклоны и пересечения — общая модель, включающая массу тела и комбинацию ткани и таксона (группа), выявила значимые эффекты «группы» (p <0,0001), а также значительный член взаимодействия «масса тела * группа» (p <0,0001). ). Таким образом, значительный член взаимодействия исключает использование общей модели с общим наклоном.
Для трех групп: плазма млекопитающих, кровь млекопитающих и кровь птиц не было значимой связи с массой тела (обозначены 1,2 и 3 в таблице 4 и на рис.4). Таким образом, период полураспада для этих групп можно просто приблизительно представить как среднее значение (ln (период полураспада): плазма млекопитающих = 1,69, кровь млекопитающих = 3,19, кровь птиц = 2,57). Для органов млекопитающих, мышц млекопитающих и плазмы птиц (4, 5 и 6) период полураспада значительно увеличивался с увеличением массы тела, и его можно оценить с помощью групповых уравнений (таблица 4 и рис. 4). Наклоны и пересечения моделей для этих трех групп заметно различались. Наконец, для органов и мышц птиц наблюдалась отрицательная связь между периодом полураспада и массой тела (7 и 8 в таблице 4 и на рис.4). Обе эти группы имели чрезвычайно узкий диапазон массы тела (рис. 4). В результате мы меньше доверяем наклонам и пересечениям, указанным в Таблице 4.
Наклоны и пересечения моделей для этих трех групп заметно различались. Наконец, для органов и мышц птиц наблюдалась отрицательная связь между периодом полураспада и массой тела (7 и 8 в таблице 4 и на рис.4). Обе эти группы имели чрезвычайно узкий диапазон массы тела (рис. 4). В результате мы меньше доверяем наклонам и пересечениям, указанным в Таблице 4.
Рис. 4. Связь между ln (массой тела) и ln (периодом полураспада) для типов тканей у A) птиц и B) млекопитающих.
Числа соответствуют таблице 4. Символы: закрашенные треугольники = кровь; белые кружки = органы, закрашенные кружки = мышцы, закрашенные квадраты = плазма.
https://doi.org/10.1371/journal.pone.0116182.g004
Чтобы облегчить более общее сравнение эндотермических групп, мы оценили период полувыведения гипотетического животного весом 50 г для каждой группы (таблица 4, последний столбец). У млекопитающих период полураспада был самым высоким для мышц, за которыми следовали кровь, органы и плазма. У птиц самый высокий период полураспада имел кровь, за ней следовали мышцы, органы и плазма крови. В целом период полураспада тканей млекопитающих был в 1,8–3,7 раза выше, чем у птиц.
У птиц самый высокий период полураспада имел кровь, за ней следовали мышцы, органы и плазма крови. В целом период полураспада тканей млекопитающих был в 1,8–3,7 раза выше, чем у птиц.
Обсуждение
Широко признано, что стабильные изотопы могут обеспечить интегрированное во времени описание рациона потребителя и трофических отношений, и что интегрирующий во времени характер подхода является ценным атрибутом подхода стабильных изотопов [2,25].Возможно, это так, хотя в значительной степени нерешенным вопрос заключается в том, «в течение какого периода времени интегрируется использование ресурсов»? Многие пищевые сети сильно структурированы по размеру. Например, в водных пищевых сетях масса тела животных увеличивается на много порядков от основания пищевой сети до высших хищников. Поскольку временной период интеграции изотопов аллометрически масштабируется с массой тела (как мы демонстрируем здесь), результатом является одновременная интеграция изотопов в нескольких временных масштабах в пределах одной пищевой сети. На примере пелагической пищевой цепи озера Танганьика O’Reilly et al.[9] обсуждали, как игнорирование того факта, что ткани животных с разных трофических уровней интегрируются в разных временных масштабах, может затруднить экологическую интерпретацию данных о стабильных изотопах. Учитывая, что многие полевые исследования стабильных изотопов, такие как те, которые используют модели смешения стабильных изотопов [8], неявно предполагают изотопное установившееся состояние между ресурсами и потребителями, мы считаем, что проблема усреднения изотопного времени неадекватно рассматривается во многих полевых исследованиях, в которых используются стабильные изотопы. Это общее беспокойство мотивировало наши усилия по синтезу опубликованных оценок периода полураспада изотопов в попытке сделать широкомасштабные статистические обобщения о том, как период полураспада зависит от таких факторов, как масса тела, таксоны и тип ткани.
На примере пелагической пищевой цепи озера Танганьика O’Reilly et al.[9] обсуждали, как игнорирование того факта, что ткани животных с разных трофических уровней интегрируются в разных временных масштабах, может затруднить экологическую интерпретацию данных о стабильных изотопах. Учитывая, что многие полевые исследования стабильных изотопов, такие как те, которые используют модели смешения стабильных изотопов [8], неявно предполагают изотопное установившееся состояние между ресурсами и потребителями, мы считаем, что проблема усреднения изотопного времени неадекватно рассматривается во многих полевых исследованиях, в которых используются стабильные изотопы. Это общее беспокойство мотивировало наши усилия по синтезу опубликованных оценок периода полураспада изотопов в попытке сделать широкомасштабные статистические обобщения о том, как период полураспада зависит от таких факторов, как масса тела, таксоны и тип ткани.
Определители периода полураспада
Скорость включения изотопов для данной ткани является следствием двух различных процессов: накопления новой биомассы (роста) и катаболического замещения тканей (катаболического обмена) [10,20]. Для роста ткани изотопная ценность новой ткани постепенно разбавляет существующий пул биомассы. Катаболический оборот включает в себя элементарный оборот в ткани, независимый от роста ткани, и было предложено, чтобы он в некоторой степени соответствовал скорости обмена белка [15,17].Ожидается, что рост будет доминировать над оборотом у маленьких, быстрорастущих животных, тогда как катаболический оборот, вероятно, будет доминировать у медленно растущих животных, и ожидается, что относительная важность катаболического обмена будет увеличиваться с увеличением размера тела животного [26]. Конечно, масса тела и скорость роста также могут быть разделены. Например, рассмотрим двух животных разных видов одинакового размера. Первое животное достигло максимального размера, а второе — быстрорастущее молодое животное. Можно было бы ожидать, что у второго животного будет заметно более высокая скорость изотопного обмена, чем у первого из-за вкладов как процессов роста, так и катаболических процессов в изотопный оборот второго животного.
Для роста ткани изотопная ценность новой ткани постепенно разбавляет существующий пул биомассы. Катаболический оборот включает в себя элементарный оборот в ткани, независимый от роста ткани, и было предложено, чтобы он в некоторой степени соответствовал скорости обмена белка [15,17].Ожидается, что рост будет доминировать над оборотом у маленьких, быстрорастущих животных, тогда как катаболический оборот, вероятно, будет доминировать у медленно растущих животных, и ожидается, что относительная важность катаболического обмена будет увеличиваться с увеличением размера тела животного [26]. Конечно, масса тела и скорость роста также могут быть разделены. Например, рассмотрим двух животных разных видов одинакового размера. Первое животное достигло максимального размера, а второе — быстрорастущее молодое животное. Можно было бы ожидать, что у второго животного будет заметно более высокая скорость изотопного обмена, чем у первого из-за вкладов как процессов роста, так и катаболических процессов в изотопный оборот второго животного. Этот гипотетический пример подчеркивает важность рассмотрения основных процессов, ответственных за оборот изотопов, и может помочь объяснить относительно высокую степень вариации периода полураспада при данной массе тела. В то время как некоторые лабораторные эксперименты действительно разделяют оборот на компоненты роста и катаболические компоненты, в большинстве случаев этого не происходит. Хотя, возможно, было бы полезно изучить, мы не стали дополнительно оценивать относительную важность этих способствующих процессов, а вместо этого сосредоточились на изучении общих закономерностей полураспада изотопов.
Этот гипотетический пример подчеркивает важность рассмотрения основных процессов, ответственных за оборот изотопов, и может помочь объяснить относительно высокую степень вариации периода полураспада при данной массе тела. В то время как некоторые лабораторные эксперименты действительно разделяют оборот на компоненты роста и катаболические компоненты, в большинстве случаев этого не происходит. Хотя, возможно, было бы полезно изучить, мы не стали дополнительно оценивать относительную важность этих способствующих процессов, а вместо этого сосредоточились на изучении общих закономерностей полураспада изотопов.
В нашем подходе к моделированию масса тела использовалась в качестве ковариаты, а комбинации типов тканей и таксонов — в качестве категориальных переменных. Хорошо известно, что разные ткани одного и того же организма могут иметь разные скорости включения изотопов [27,28], что предположительно указывает на разные скорости обмена белков [1,15]. Кроме того, мы ожидаем различий в обороте среди таксономических групп (беспозвоночные, рыбы, млекопитающие, птицы), в значительной степени отражающих более высокую скорость обновления эндотермических животных из-за их более высокой скорости кормления и метаболизма (метаболизм в широком смысле). Мы исследовали роль типа ткани и таксона, рассматривая комбинации категорий тканей (мышцы, все тело, органы, кровь, плазма крови) и таксонов (беспозвоночные, эктотермия позвоночных, птицы, млекопитающие; таблица 2). Несколько комбинаций ткани и таксона имели либо несколько наблюдений, либо их отсутствие, либо небольшие вариации в массе тела. Доступно достаточно данных, чтобы охарактеризовать 13 из 20 возможных комбинаций ткани и таксона.
Мы исследовали роль типа ткани и таксона, рассматривая комбинации категорий тканей (мышцы, все тело, органы, кровь, плазма крови) и таксонов (беспозвоночные, эктотермия позвоночных, птицы, млекопитающие; таблица 2). Несколько комбинаций ткани и таксона имели либо несколько наблюдений, либо их отсутствие, либо небольшие вариации в массе тела. Доступно достаточно данных, чтобы охарактеризовать 13 из 20 возможных комбинаций ткани и таксона.
Мы использовали аллометрический подход для изучения изменчивости периода полураспада тканей животных.Интуитивно мы могли бы ожидать, что период полураспада будет увеличиваться с увеличением массы тела: небольшое животное, вероятно, будет иметь высокие удельные темпы роста и, возможно, высокие удельные скорости оборота белка, так что измерение изотопов будет отражать несколько дней или недель кормления. Напротив, ожидается, что у крупного животного будет низкая удельная скорость роста и скорость оборота белка, так что можно ожидать, что измерение изотопов будет интегрироваться в течение периода от месяцев до лет. Более формально Карлтон и Мартинес дель Рио (2005) предположили, что оборот λ можно интерпретировать как v / P, где v — чистая скорость притока элемента в ткань, а P — размер пула элементов.Ожидается, что v будет пропорционален массе тела 3/4 , тогда как P должен масштабироваться изометрически с массой тела, так что оборот должен быть пропорционален массе тела -1/4 (т. е. масса 3/4 / масса ). Кроме того, конкретные темпы роста также имеют тенденцию масштабироваться с массой тела -1/4 [29]. Таким образом, ожидается, что оба процесса, которые вносят вклад в обмен изотопов (рост ткани и катаболический оборот), будут масштабироваться с массой тела с показателем -1 / 4.
Более формально Карлтон и Мартинес дель Рио (2005) предположили, что оборот λ можно интерпретировать как v / P, где v — чистая скорость притока элемента в ткань, а P — размер пула элементов.Ожидается, что v будет пропорционален массе тела 3/4 , тогда как P должен масштабироваться изометрически с массой тела, так что оборот должен быть пропорционален массе тела -1/4 (т. е. масса 3/4 / масса ). Кроме того, конкретные темпы роста также имеют тенденцию масштабироваться с массой тела -1/4 [29]. Таким образом, ожидается, что оба процесса, которые вносят вклад в обмен изотопов (рост ткани и катаболический оборот), будут масштабироваться с массой тела с показателем -1 / 4.
Ectotherms
Период полураспада для эктотермов описывался моделью с общим наклоном и групповыми пересечениями.Отношение массы тела к периоду полураспада имело наклон ~ 0,22 (эквивалент показателя оборачиваемости -0,22), что достаточно хорошо соответствует прогнозируемому наклону ~ 0,25 [1,15], а также наблюдаемому аллометрическому наклону. для изотопов углерода в мышцах рыб [14].
для изотопов углерода в мышцах рыб [14].
Мы оценили групповые перехваты в рамках структуры ANCOVA (уравнение 2; таблица 3), а затем упростили модель, объединив группы, которые существенно не отличались друг от друга (уравнение 3; таблица 3, цифры в верхнем индексе).Органы эктотерма позвоночных и плазма крови имели самый низкий период полураспада. Этот результат согласуется с многочисленными другими исследованиями, в которых сообщается, что внутренние органы и плазма крови рыб обновляются быстрее по сравнению с кровью или мышечной тканью рыб [28,30,31]. Остальные группы с эктотермией (за исключением эктотермии всего тела позвоночных) статистически неразличимы. перехватывает, тем самым указывая на общие аллометрические отношения. Эктотерм позвоночных для всего тела имел незначительно более высокий перехват, чем в других группах (Таблица 3).В исследованиях обычно используются измерения всего тела для животных, которые слишком малы для разделения отдельных тканей. Таким образом, существуют только оценки «всего тела» для беспозвоночных и очень мелких рыб, и есть небольшое совпадение по массе тела между оценками всего тела и другими оценками (сравните диапазон массы тела на рис. 2G с диапазоном массы тела других типов тканей). Мы решили хранить оценки эктотермы всего тела позвоночных отдельно от других групп в уравнении. 3, но отметим, что эта разница может быть просто результатом ограниченного совпадения масс тела.
2G с диапазоном массы тела других типов тканей). Мы решили хранить оценки эктотермы всего тела позвоночных отдельно от других групп в уравнении. 3, но отметим, что эта разница может быть просто результатом ограниченного совпадения масс тела.
Эндотерм
В отличие от эктотерм, комбинации эндотермических тканей и таксонов демонстрируют групповые наклоны и пересечения (Таблица 4; Рис. 4). Таким образом, хотя мы все еще можем предоставить статистическую модель для аппроксимации периода полураспада (таблица 4), интерпретация менее проста, чем для эктотерм.
Для трех групп (плазма млекопитающих, кровь млекопитающих и кровь птиц) не было значительного изменения периода полувыведения в зависимости от массы тела животного (таблица 4; рис. 4). Этот результат противоречит нашим ожиданиям о положительной взаимосвязи между оборотом и массой тела и предполагает, что период полураспада для этих групп можно оценить без ссылки на массу тела.В отличие от наших результатов, два более ранних исследования показали положительную взаимосвязь между периодом полураспада и массой тела для крови птиц [15,16]. Причины этих различий неясны, хотя мы отмечаем, что все выводы основаны на относительно небольших размерах выборки. Никакие предыдущие исследования не изучали, связан ли период полураспада изотопов с массой тела для тканей млекопитающих любого типа.
Причины этих различий неясны, хотя мы отмечаем, что все выводы основаны на относительно небольших размерах выборки. Никакие предыдущие исследования не изучали, связан ли период полураспада изотопов с массой тела для тканей млекопитающих любого типа.
Для трех групп эндотерма (органы млекопитающих, мышцы млекопитающих и плазма птиц) ожидаемая положительная взаимосвязь между массой тела и периодом полураспада с отклонениями от 0.18 до 0,3. Аллометрические наклоны для мышц млекопитающих и органов млекопитающих были аналогичными, хотя мышца млекопитающих имела более высокий перехват (таблица 4, рис. 4B). Этот вывод согласуется с предыдущей литературой, показывающей, что внутренние органы имеют тенденцию к более быстрому обновлению, чем мышечная ткань (Tieszen et al. 1983). В то время как аллометрический наклон для плазмы птиц был высоким (0,3), период полувыведения из плазмы птиц был низким по сравнению с другими тканями (таблица 4, рис. 4A). Вызывает недоумение то, что плазма млекопитающих и плазма птиц имели такие разные значения массы тела (-0. 01 против 0,30). Тем не менее, наши результаты показывают, что у плазмы крови короткий период полураспада по сравнению с кровью в целом, что и ожидалось, поскольку клетки плазмы крови недолговечны по сравнению с эритроцитами и лейкоцитами.
01 против 0,30). Тем не менее, наши результаты показывают, что у плазмы крови короткий период полураспада по сравнению с кровью в целом, что и ожидалось, поскольку клетки плазмы крови недолговечны по сравнению с эритроцитами и лейкоцитами.
Наконец, для органов и мышц птиц действительно существует отрицательная связь между массой тела и периодом полураспада (Таблица 4; Рис. 4A). Этот результат был неожиданным. Для этих двух групп наблюдалась небольшая разница в массе тела (рис. 4), и возможно, что наши результаты вместо этого основаны на небольшом размере выборки и одной или небольшом количестве аномальных точек данных.Аллометрические взаимосвязи мышц и органов птиц явно требуют дальнейшего изучения.
Изотопные различия
Мы обнаружили незначительные различия в соотношении масса тела-период полураспада между тремя оцениваемыми нами изотопами (δ 13 C, δ 15 N и δ 34 S). Этот результат несколько удивителен, учитывая уникальные метаболические пути, биохимические роли и функции этих трех элементов. Отдельные исследования обнаружили различия в периоде полураспада между изотопами углерода и азота, хотя направление этих тенденций не было согласованным.Карлтон и Мартинес дель Рио [15] обнаружили, что период полураспада азота у птиц на 50% выше, чем у углерода. Напротив, исследование молодых стальных головок показало, что период полураспада азота ниже, чем у углерода [32]. Другие исследования не сообщают об отсутствии разницы между периодом полураспада для δ 13 C и δ 15 N [33]. По сравнению с углеродом и азотом сера присутствует в тканях животных в низких концентрациях, и большая часть серы связана с белками (аминокислотами цистеином и метионином). Мы отмечаем, что было относительно мало оценок периода полураспада для δ 34 S (18 оценок периода полураспада из шести отдельных статей по сравнению с 468 комбинированными оценками для углерода и азота), и большинство оценок серы было получено из одного исследования, сравнивающего δ . 13 C, δ 15 N и δ 34 S оборот для набора тканей мышей (Arneson et al.
Отдельные исследования обнаружили различия в периоде полураспада между изотопами углерода и азота, хотя направление этих тенденций не было согласованным.Карлтон и Мартинес дель Рио [15] обнаружили, что период полураспада азота у птиц на 50% выше, чем у углерода. Напротив, исследование молодых стальных головок показало, что период полураспада азота ниже, чем у углерода [32]. Другие исследования не сообщают об отсутствии разницы между периодом полураспада для δ 13 C и δ 15 N [33]. По сравнению с углеродом и азотом сера присутствует в тканях животных в низких концентрациях, и большая часть серы связана с белками (аминокислотами цистеином и метионином). Мы отмечаем, что было относительно мало оценок периода полураспада для δ 34 S (18 оценок периода полураспада из шести отдельных статей по сравнению с 468 комбинированными оценками для углерода и азота), и большинство оценок серы было получено из одного исследования, сравнивающего δ . 13 C, δ 15 N и δ 34 S оборот для набора тканей мышей (Arneson et al.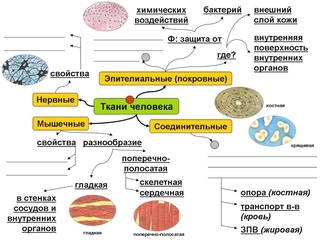 2006 г.). Отдельные исследования с участием изотопов серы показали, что оценки периода полураспада серы, как правило, не сильно отличаются от других элементов (Tarboush 2006, Arneson et al. 2006, Bahar 2009, Hesslein et al. 1993, MacAvoy 2001). Несмотря на глубокие биохимические различия между этими тремя элементами, не было резких различий в аллометрических отношениях между ними. В целом, различия между изотопами кажутся незначительным источником шума в более широком контексте довольно общей аллометрической взаимосвязи.
2006 г.). Отдельные исследования с участием изотопов серы показали, что оценки периода полураспада серы, как правило, не сильно отличаются от других элементов (Tarboush 2006, Arneson et al. 2006, Bahar 2009, Hesslein et al. 1993, MacAvoy 2001). Несмотря на глубокие биохимические различия между этими тремя элементами, не было резких различий в аллометрических отношениях между ними. В целом, различия между изотопами кажутся незначительным источником шума в более широком контексте довольно общей аллометрической взаимосвязи.
Температурный эффект
Хотя на биологические показатели, как правило, сильно влияет температура [18], после учета массы тела наблюдалось слабое (p = 0,02) влияние температуры на период полураспада для эктотерм. Размер нашей выборки, включая данные о температуре, был довольно большим (247 оценок), а диапазон температур был большим (приблизительно 35 ° C). Аналогичным образом Weidel et al. [14] практически не обнаружили влияния температуры на период полураспада изотопа углерода в мышечной ткани рыб. Напротив, отдельные эксперименты, которые включали температуру в качестве обработки, иногда обнаруживали влияние температуры на период полураспада. Например, Bosley et al. [34] и Witting et al. [35] обнаружили, что выращивание личинок рыб при более высоких температурах воды приводит к более низкому периоду полураспада углерода. Это могло быть связано с положительным влиянием температуры на скорость роста, которая, как ожидается, будет доминировать в обмене изотопов у быстрорастущих животных. В целом, наши результаты показывают, что влияние температуры на период полураспада невелико в контексте общего изменения периода полураспада.
Напротив, отдельные эксперименты, которые включали температуру в качестве обработки, иногда обнаруживали влияние температуры на период полураспада. Например, Bosley et al. [34] и Witting et al. [35] обнаружили, что выращивание личинок рыб при более высоких температурах воды приводит к более низкому периоду полураспада углерода. Это могло быть связано с положительным влиянием температуры на скорость роста, которая, как ожидается, будет доминировать в обмене изотопов у быстрорастущих животных. В целом, наши результаты показывают, что влияние температуры на период полураспада невелико в контексте общего изменения периода полураспада.
Прогнозы модели и ошибки
Наши эмпирические модели, предсказывающие период полураспада изотопов для тканей животных, основаны на большом объеме экспериментальных данных, полученных от животных, охватывающих диапазон размеров тела. Наша модель может использоваться в качестве инструмента для непосредственной аппроксимации временной шкалы интеграции диеты в зависимости от размера тела, типа ткани и таксономической группы. В качестве примера, используя уравнение. 3, период полураспада мышечной ткани от 10-граммовой эктотермы позвоночных (группа 2 в таблице 3) оценивается в 44 дня с интервалом прогноза 95% в диапазоне от 10 до 196 дней.В качестве другого примера, мышечная ткань млекопитающего весом 10 грамм оценивается в 21 день (95% интервал прогноза составляет 12–38 дней). Хотя наша модель позволяет прогнозировать период полураспада на основе массы тела и типа ткани, широкие интервалы прогнозирования подчеркивают тот факт, что этот подход обеспечивает грубую аппроксимацию периода полураспада изотопов.
В качестве примера, используя уравнение. 3, период полураспада мышечной ткани от 10-граммовой эктотермы позвоночных (группа 2 в таблице 3) оценивается в 44 дня с интервалом прогноза 95% в диапазоне от 10 до 196 дней.В качестве другого примера, мышечная ткань млекопитающего весом 10 грамм оценивается в 21 день (95% интервал прогноза составляет 12–38 дней). Хотя наша модель позволяет прогнозировать период полураспада на основе массы тела и типа ткани, широкие интервалы прогнозирования подчеркивают тот факт, что этот подход обеспечивает грубую аппроксимацию периода полураспада изотопов.
Высокая степень неопределенности модели напрямую связана с высокой степенью изменчивости исходных оценок периода полураспада — другими словами, животные одного размера, типа ткани и широкой таксономической группы могут иметь сильно различающиеся периоды полураспада изотопов.Эксперименты по смене диеты, включенные в наш синтез, проводились широким кругом исследователей и в большом диапазоне экспериментальных условий (диеты, скорость роста, температуры и т.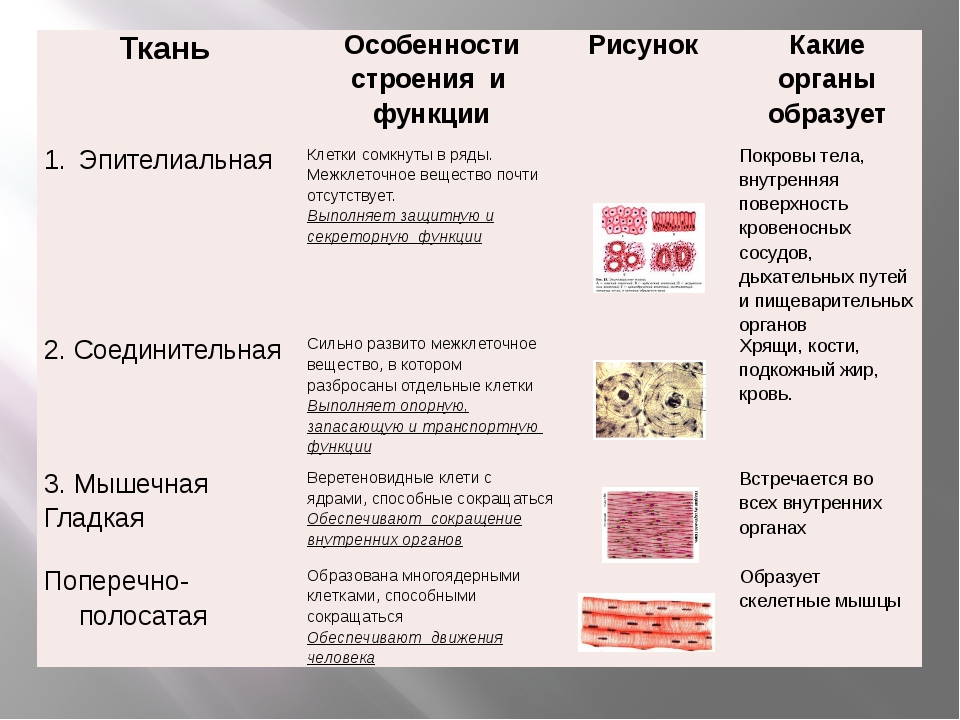 Д.). Это само по себе может помочь объяснить высокую степень шума в соотношении масса тела — период полураспада. Общий вывод состоит в том, что оценку периода полураспада из одного экспериментального исследования нельзя использовать в качестве основы для точного определения периода полураспада в полевых условиях. Наш подход к синтезу многих опубликованных оценок периода полураспада (обзор литературы актуален до мая 2014 г.) позволяет нам сделать более строгие статистические обобщения и явно признать высокую степень изменчивости, которая в данном случае приводит к неопределенности модели.
Д.). Это само по себе может помочь объяснить высокую степень шума в соотношении масса тела — период полураспада. Общий вывод состоит в том, что оценку периода полураспада из одного экспериментального исследования нельзя использовать в качестве основы для точного определения периода полураспада в полевых условиях. Наш подход к синтезу многих опубликованных оценок периода полураспада (обзор литературы актуален до мая 2014 г.) позволяет нам сделать более строгие статистические обобщения и явно признать высокую степень изменчивости, которая в данном случае приводит к неопределенности модели.
Ряд факторов, явно не учтенных в нашем исследовании, могут повлиять на динамику включения изотопов. К ним относятся различия в скорости роста животных, диете и качестве пищи, стадии жизненного цикла, физиологическом состоянии и маршрутизации изотопов [1,36,37]. Кроме того, исходные оценки периода полураспада сами по себе подвержены нескольким источникам ошибок. К ним могут относиться недостаточная разница изотопов между экспериментальными диетами, ошибка в предполагаемом факторе обогащения диетических тканей, невозможность провести эксперимент до изотопного устойчивого состояния и использование принципиально несоответствующей модели.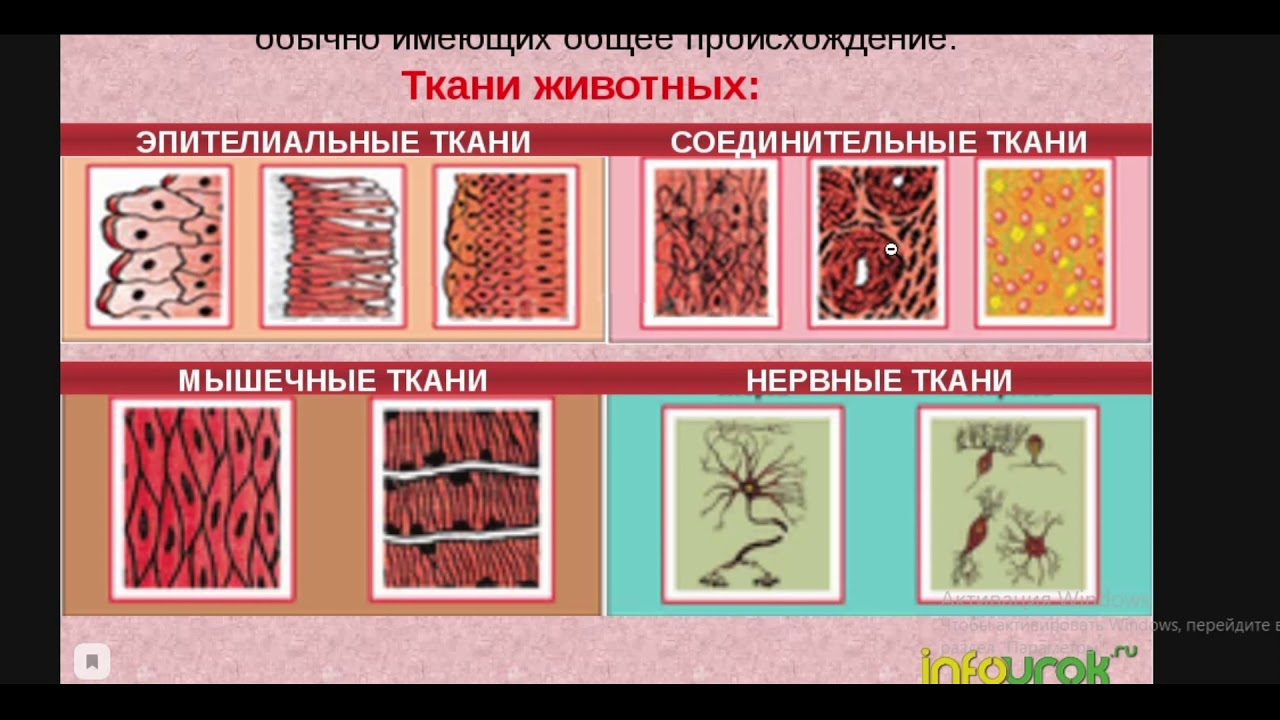 В качестве примера последнего, несколько моделей, используемых для оценки периода полураспада, предполагают экспоненциальный соматический рост животного в эксперименте по смене диеты — отклонение от этого предположения приведет к ошибочным оценкам периода полураспада. Фактически, существует неопределенность, связанная с величиной периода полураспада, о которой сообщалось в первоначальных исследованиях, хотя эта неопределенность редко оценивалась или сообщалась, и здесь явно не рассматривается.
В качестве примера последнего, несколько моделей, используемых для оценки периода полураспада, предполагают экспоненциальный соматический рост животного в эксперименте по смене диеты — отклонение от этого предположения приведет к ошибочным оценкам периода полураспада. Фактически, существует неопределенность, связанная с величиной периода полураспада, о которой сообщалось в первоначальных исследованиях, хотя эта неопределенность редко оценивалась или сообщалась, и здесь явно не рассматривается.
Другие переменные, использованные в нашем анализе, также были подвержены ошибкам. Например, мы использовали массу тела животного в начале эксперимента в качестве меры массы тела в нашем анализе.Хотя это несовершенный способ измерения массы тела животного, это был лучший и самый надежный вариант, учитывая доступные данные.
Сводка
Наша цель состояла в том, чтобы оценить, можно ли сделать широкие эмпирические обобщения на основе большого количества оценок периода полураспада, которые были опубликованы за последние несколько десятилетий. Чтобы обеспечить основу для сравнения, было проведено несколько широких эмпирических синтезов факторов различения диет и тканей [4–6]. Эти синтезы впоследствии обеспечили основу для количественного применения подходов к изучению стабильных изотопов, которые обычно требуют четких предположений о различении тканей и диеты.Дискриминация ткани и питания в настоящее время достаточно хорошо изучена, и исследования обычно включают наблюдаемую изменчивость в различении тканей и питания в результаты моделей смешивания. Напротив, тема изотопного оборота и периода полураспада не была предметом широкого эмпирического синтеза. Рассмотрение временной шкалы включения изотопов в ткани животных может быть жизненно важным при интерпретации результатов по стабильным изотопам [9,36]. Для эктотермов период полураспада предсказуемо увеличивался с увеличением размера тела, что давало относительно простую эмпирическую модель для приблизительного определения периода полураспада.Для птиц и млекопитающих связь с массой тела включает разные аллометрические наклоны и пересечения для разных тканей.
Чтобы обеспечить основу для сравнения, было проведено несколько широких эмпирических синтезов факторов различения диет и тканей [4–6]. Эти синтезы впоследствии обеспечили основу для количественного применения подходов к изучению стабильных изотопов, которые обычно требуют четких предположений о различении тканей и диеты.Дискриминация ткани и питания в настоящее время достаточно хорошо изучена, и исследования обычно включают наблюдаемую изменчивость в различении тканей и питания в результаты моделей смешивания. Напротив, тема изотопного оборота и периода полураспада не была предметом широкого эмпирического синтеза. Рассмотрение временной шкалы включения изотопов в ткани животных может быть жизненно важным при интерпретации результатов по стабильным изотопам [9,36]. Для эктотермов период полураспада предсказуемо увеличивался с увеличением размера тела, что давало относительно простую эмпирическую модель для приблизительного определения периода полураспада.Для птиц и млекопитающих связь с массой тела включает разные аллометрические наклоны и пересечения для разных тканей. Несмотря на наличие некоторых широких эмпирических закономерностей, в наших моделях периода полураспада изотопов также наблюдались существенные необъяснимые вариации, указывающие на то, что ошибка измерения и неизмеренные факторы влияют на период полураспада изотопов. Мы надеялись, что широкомасштабный синтез существующих данных сможет выявить общие закономерности, которые нельзя было обнаружить в отдельных исследованиях.Хотя наш сравнительный подход не может напрямую прояснить лежащие в основе механизмы, он действительно выявляет пробелы в наших знаниях и понимании, которые, мы надеемся, послужат источником информации для будущих экспериментальных работ на этом важном стыке экологии и физиологии.
Несмотря на наличие некоторых широких эмпирических закономерностей, в наших моделях периода полураспада изотопов также наблюдались существенные необъяснимые вариации, указывающие на то, что ошибка измерения и неизмеренные факторы влияют на период полураспада изотопов. Мы надеялись, что широкомасштабный синтез существующих данных сможет выявить общие закономерности, которые нельзя было обнаружить в отдельных исследованиях.Хотя наш сравнительный подход не может напрямую прояснить лежащие в основе механизмы, он действительно выявляет пробелы в наших знаниях и понимании, которые, мы надеемся, послужат источником информации для будущих экспериментальных работ на этом важном стыке экологии и физиологии.
Благодарности
Мы благодарим Скотта Хиггинса, членов лабораторной группы Вандера Зандена, и анонимных рецензентов за ценные комментарии к рукописи.
Вклад авторов
Задумал и спроектировал эксперименты: MJVZ EKM CTS BCW.Проведены эксперименты: MJVZ EKM CTS BCW. Проанализированы данные: MJVZ MKC EKM CTS. Написал документ: MJVZ MKC EKM CTS BCW.
Проанализированы данные: MJVZ MKC EKM CTS. Написал документ: MJVZ MKC EKM CTS BCW.
Ссылки
- 1.
Мартинес дель Рио К., Вольф Н., Карлтон С.А., Ганнес Л. (2009) Изотопная экология через десять лет после призыва к новым лабораторным экспериментам. Биологические обзоры 84: 91–111. pmid: 1
98
- 2. Петерсон Б.Дж., Фрай Б. (1987) Стабильные изотопы в исследованиях экосистем. Ежегодный обзор экологии и систематики 18: 293–320.
- 3. Хобсон К.А., Велч Х.Э. (1992) Определение трофических взаимоотношений в пределах высокоразвитой сети морских ферм в Арктике с использованием анализа d13C и d15N. Серия «Прогресс морской экологии» 84: 9–18.
- 4. Vander Zanden MJ, Rasmussen JB (2001) Изменения в 15 N и 13 C трофическое фракционирование: значение для исследований водных пищевых сетей. Лимнология и океанография 46: 2061–2066.
- 5.
Вандерклифт М.А., Понсард С. (2003) Источники различий в обогащении дельты N-15 в рационе потребителей: метаанализ.
 Oecologia 136: 169–182. pmid: 12802678
Oecologia 136: 169–182. pmid: 12802678 - 6. McCutchan JH, Lewis WM, Kendall C, McGrath CC (2003) Изменение трофического сдвига для соотношений стабильных изотопов углерода, азота и серы. Ойкос 102: 378–390.
- 7. Парнелл А.К., Ингер Р., Bearhop С., Джексон А.Л. (2010) Разделение источника с использованием стабильных изотопов: борьба со слишком большим разнообразием. Plos One 5. pmid: 21283510
- 8. Филлипс Д.Л., Грегг Дж. В. (2003) Разделение источников с использованием стабильных изотопов: работа со слишком большим количеством источников.Oecologia 136: 261–269. pmid: 12759813
- 9. O’Reilly CM, Hecky RE (2002) Интерпретация стабильных изотопов в пищевых сетях: признание роли временного усреднения на разных трофических уровнях. Лимнология и океанография 47: 306–309. pmid: 25564461
- 10.
Fry B, Arnold C (1982) Быстрый оборот 13C / 12C во время роста коричневой креветки ( Penaeus aztecus ).
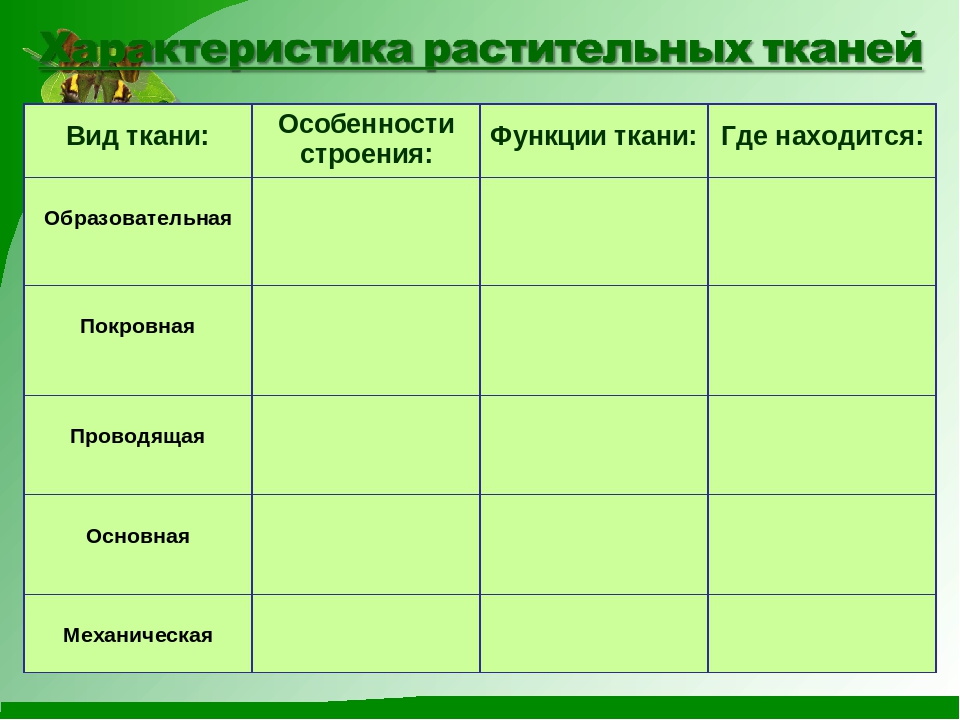 Oecologia 54: 200–204.
Oecologia 54: 200–204. - 11. Беклен В.Дж., Ярнес СТ, Кук Б.А., Джеймс А.С. (2011) Об использовании стабильных изотопов в трофической экологии.Ежегодный обзор экологии, эволюции и систематики 42: 411–440.
- 12. Филлипс Д.Л., Элдридж П.М. (2006) Оценка сроков смены рациона с использованием стабильных изотопов. Oecologia 147: 195–203. pmid: 16341714
- 13. McIntyre PB, Flecker AS (2006) Быстрый оборот тканевого азота основных потребителей в тропических пресных водах. Oecologia 148: 12–21. pmid: 16456686
- 14. Weidel BC, Carpenter SR, Kitchell JF, Vander Zanden MJ (2011) Темпы и компоненты кругооборота углерода в мышцах рыб: выводы из биоэнергетических моделей и добавление углерода для всего озера (13).Канадский журнал рыболовства и водных наук 68: 387–399.
- 15.
Карлтон С.А., Мартинес дель Рио С. (2005) Влияние повышенного уровня метаболизма, вызванного холодом, на скорость включения (13) C и (15) N у домашних воробьев (Passer domesticus).
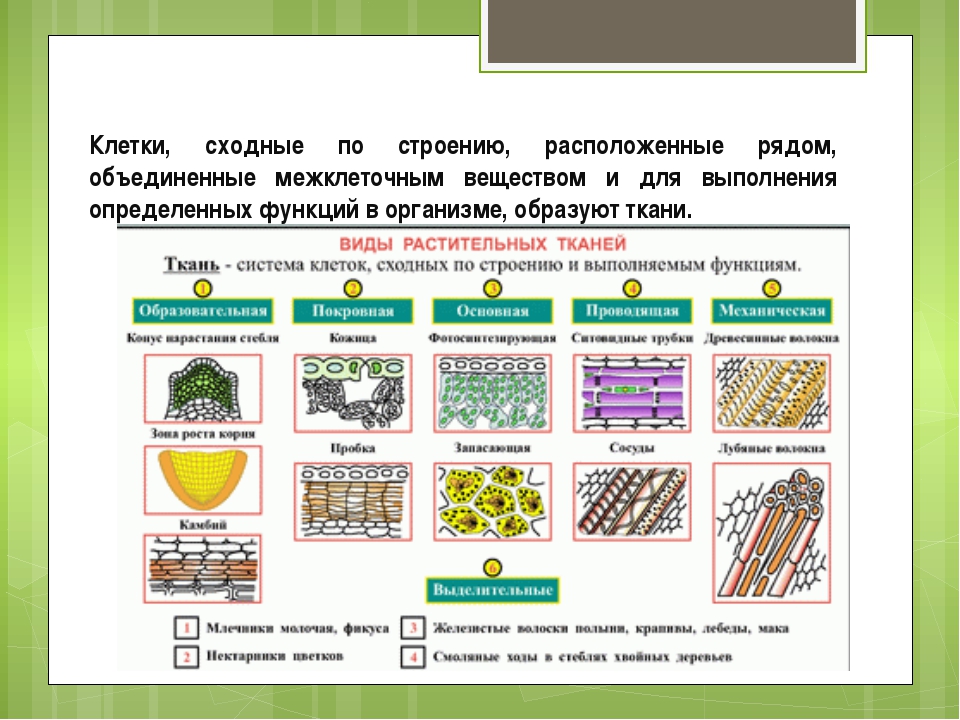 Oecologia 144: 226–232. pmid: 158
Oecologia 144: 226–232. pmid: 158 - 16. Bauchinger U, McWilliams SR (2009) Круговорот углерода в тканях воробьиных птиц: аллометрия, изотопные часы и фенотипическая гибкость в размере органа. Интегративная и сравнительная биология 49: E11 – E11.
- 17. Houlihan DF, Carter CG, McCarthy I (1995) Белковый обмен у животных. В: Уолш П.Дж., Райт П., редакторы. Метаболизм и выведение азота. Бока-Ратон: CRC Press. С. 1–32.
- 18. Браун Дж. Х., Гиллули Дж. Ф., Аллен А. П., Сэвидж В. М., Вест ГБ (2004) К метаболической теории экологии. Экология 85: 1771–1789.
- 19. Хобсон К.А., Кларк Р.Г. (1992) Оценка рациона птиц с использованием стабильных изотопов I: оборот 13C в тканях. Кондор 94: 181–188.
- 20.
Hesslein RH, Hallard KA, Ramlal P (1993) Замена серы, углерода и азота в тканях растущего сига ( Coregonus nasus ) в ответ на изменение рациона, отслеживаемое с помощью 34S, 13C и 15N. Канадский журнал рыболовства и водных наук 50: 2071–2076.

- 21. Cerling TE, Ayliffe LK, Dearing MD, Ehleringer JR, Passey BH и др. (2007) Определение оборота биологической ткани с использованием стабильных изотопов: переменная хода реакции.Oecologia 151: 175–189. pmid: 17186277
- 22. Карлтон С.А., Келли Л., Андерсон-Спречер Р., дель Рио С.М. (2008) Должны ли мы использовать одно- или многокамерные модели для описания включения C-13 в ткани животных? Быстрые коммуникации в масс-спектрометрии 22: 3008–3014. pmid: 18773411
- 23. Vander Zanden MJ, Hulshof M, Ridgway MS, Rasmussen JB (1998) Применение методов стабильных изотопов к трофическим исследованиям малорозного окуня возраста 0. Сделки Американского рыболовного общества 127: 729–739.
- 24. Маруяма А., Ямада Ю., Русува Б., Юма М. (2001) Изменение соотношения стабильных изотопов азота в мышечной ткани мигрирующего бычка, Rhinogobius sp., В естественных условиях. Может ли J Fish Aquat Sci 58: 2125–2128.
- 25.
 Клинг Г.В., Фрай Б., О’Брайен В.Дж. (1992) Стабильные изотопы и планктонная трофическая структура в арктических озерах. Экология 73: 561–566.
Клинг Г.В., Фрай Б., О’Брайен В.Дж. (1992) Стабильные изотопы и планктонная трофическая структура в арктических озерах. Экология 73: 561–566. - 26. Вольф Н., Карлтон С.А., дель Рио С.М. (2009) Десять лет экспериментальной изотопной экологии животных.Функциональная экология 23: 17–26.
- 27. Логан Дж., Хаас Х., Диган Л., Гейнс Э. (2006) Скорость оборота стабильных изотопов азота в мумихоге солончака, Fundulus heteroclitus, после смены диеты в лаборатории. Oecologia 147: 391–395. pmid: 16249895
- 28. Suzuki KW, Kasai A, Nakayama K, Tanaka M (2005) Дифференциальное изотопное обогащение и период полураспада среди тканей молоди японского умеренного окуня (Lateolabrax japonicas): значение для анализа миграции.Канадский журнал рыболовства и водных наук 62: 671–678.
- 29. Петерс Р.Х. (1983) Экологические последствия размера тела. Кембридж: Издательство Кембриджского университета.
- 30.
MacNeil MA, Drouillard KG, Fisk AT (2006) Переменное поглощение и удаление стабильных изотопов азота между тканями у рыб.
 Канадский журнал рыболовства и водных наук 63: 345–353.
Канадский журнал рыболовства и водных наук 63: 345–353. - 31. Guelinckx J, Maes J, Van Den Driessche P, Geysen B, Dehairs F и др.(2007) Изменения дельта C-13 и дельта N-15 в различных тканях молоди песчаного бычка Pomatoschistus minutus: лабораторный эксперимент по смене диеты. Серия «Прогресс морской экологии» 341: 205–215.
- 32. Church MR, Ebersole JL, Rensmeyer KM, Couture RB, Barrows FT и др. (2009) Слизь: новое фракционирование тканей для быстрого определения смены рациона рыб с использованием анализа стабильных изотопов. Канадский журнал рыболовства и водных наук 66: 1–5.
- 33. Ogden LJE, Hobson KA, Lank DB (2004) Изотопный обмен крови (дельта C-13 и дельта C-15) факторы фракционирования диеты и ткани у чернозобиков в неволе (Calidris alpina pacifica).Аук 121: 170–177
- 34.
Босли К.Л., Виттинг Д.А., Чемберс Р.К., Уэйнрайт С.К. (2002) Оценка скорости оборота углерода и азота стабильными изотопами у недавно подвергшейся метаморфизации зимней камбалы Pseudopleuronectes americanus.
 Серия «Морская экология-Прогресс» 236: 233–240.
Серия «Морская экология-Прогресс» 236: 233–240. - 35. Witting DA, Chambers RC, Bosley KL, Wainright SC (2004) Экспериментальная оценка онтогенетических изменений диеты у летней камбалы (Paralichthys dentatus) с использованием стабильных изотопов в качестве индикаторов рациона.Канадский журнал рыболовства и водных наук 61: 2069–2084.
- 36. Мартинес дель Рио К., Карлтон С.А. (2012) Насколько быстро и насколько достоверно: динамика включения изотопов в ткани животных. Journal of Mammalogy 93: 353–359.
- 37. Ньюсом С.Д., Клеменц М.Т., Кох П.Л. (2010) Использование биогеохимии стабильных изотопов для изучения экологии морских млекопитающих. Наука о морских млекопитающих 26: 509–572.
Ткань — определение и примеры
Ткань
n., множественное число: ткани
[ˈtɪʃu]
Определение: совокупность клеток, имеющих одинаковую структуру и функцию
Определение ткани
Что такое ткань в организме? Ткань тела представляет собой совокупность клеток, которые функционируют вместе и выполняют схожие функции. Клетки многоклеточного организма обычно не функционируют независимо, вместо этого они обычно связаны с другими клетками, образующими ткань. В основном ткани состоят из нескольких типов клеток, которые удовлетворяют различные потребности тканевых функций.Исследование тканей называется гистология . Эта наука изучает анатомию различных тканей с помощью микроскопов. Гистология микроскопической биологической науки помогла в классификации тканей растений и животных, а также в определении их детальной структуры.
Клетки многоклеточного организма обычно не функционируют независимо, вместо этого они обычно связаны с другими клетками, образующими ткань. В основном ткани состоят из нескольких типов клеток, которые удовлетворяют различные потребности тканевых функций.Исследование тканей называется гистология . Эта наука изучает анатомию различных тканей с помощью микроскопов. Гистология микроскопической биологической науки помогла в классификации тканей растений и животных, а также в определении их детальной структуры.
«Когда клетки организма со схожими функциями и структурами объединяются в группу, эта единица называется тканью». Получите информацию прямо от эксперта: гладкие мышцы против плотной нормальной соединительной ткани.Присоединяйтесь к нам сейчас!
Биологическое определение: A ткань представляет собой совокупность клеток в организме, которые имеют аналогичную структуру и функции.
 Ткани, которые работают в унисон для выполнения определенного набора функций, образуют орган. Примерами тканей растений являются ткани меристемы и ткани сосудов. Примерами тканей животных являются эпителиальные ткани, соединительные ткани, мышечные ткани и нервные ткани. Этимология: Среднеанглийский тиссу, богатый вид ткани, от старофранцузского, от причастия прошедшего времени tistre, до ткать, от латинского texere.
Ткани, которые работают в унисон для выполнения определенного набора функций, образуют орган. Примерами тканей растений являются ткани меристемы и ткани сосудов. Примерами тканей животных являются эпителиальные ткани, соединительные ткани, мышечные ткани и нервные ткани. Этимология: Среднеанглийский тиссу, богатый вид ткани, от старофранцузского, от причастия прошедшего времени tistre, до ткать, от латинского texere.Типы тканей животных
Что такое ткани? Определение ткани у животных — это группа клеток, объединенных вместе для выполнения определенной функции. У животных существует четыре типа тканей, каждый из которых имеет свою особую структуру и функцию. Это (1) эпителиальных тканей , (2) соединительных тканей , (3) мышечных тканей и (4) нервных тканей . Каждая из этих тканей имеет свою уникальную структуру и биологическую функцию.Эти различные типы тканей обсуждаются ниже:
Эпителиальная ткань
Эпителиальные ткани (или эпителий ) являются наиболее распространенными тканями человека.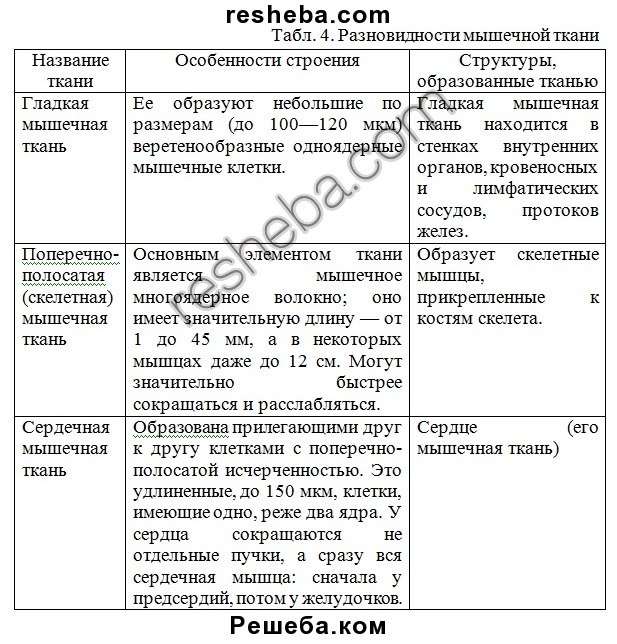 Они покрывают всю поверхность тела, образуя слой эпидермиса кожи. Кроме того, он покрывает слизистую оболочку органов тела, например слизистую оболочку пищеварительной системы. Таким образом, они защищают орган от внешних факторов, таких как травмы, патогенные микроорганизмы и потеря жидкости.Эпителий участвует в выделительном процессе, в котором клетки железистого эпителия участвуют в синтезе и выведении шлаков. Эпителиальные ткани также отвечают за секрецию и абсорбцию тела. Они отвечают за секрецию слюны, слизи, пота, и т. Д. , поэтому они составляют значительную часть многоклеточных желез, таких как печень и поджелудочная железа. Другая эпителиальная ткань покрывает различные полости тела, вырабатывая секреторные смазки.
Они покрывают всю поверхность тела, образуя слой эпидермиса кожи. Кроме того, он покрывает слизистую оболочку органов тела, например слизистую оболочку пищеварительной системы. Таким образом, они защищают орган от внешних факторов, таких как травмы, патогенные микроорганизмы и потеря жидкости.Эпителий участвует в выделительном процессе, в котором клетки железистого эпителия участвуют в синтезе и выведении шлаков. Эпителиальные ткани также отвечают за секрецию и абсорбцию тела. Они отвечают за секрецию слюны, слизи, пота, и т. Д. , поэтому они составляют значительную часть многоклеточных желез, таких как печень и поджелудочная железа. Другая эпителиальная ткань покрывает различные полости тела, вырабатывая секреторные смазки.
Эпителиальная ткань состоит либо из однослойных эпителиальных клеток (называемых простым эпителием ), либо из множественных слоев клеток (называемых многослойным эпителием ).Эпителиальные клетки, образующие единый слой эпителиальной ткани, имеют три поверхности: (1) апикальную поверхность, (2) базальную поверхность и (3) боковую поверхность. Каждая из этих поверхностей выполняет свою функцию.
Каждая из этих поверхностей выполняет свою функцию.
Апикальная поверхность эпителиальных клеток отвечает за обмен материала, входящего и выходящего из клетки. Апикальная поверхность эпителиальных клеток внутренней оболочки тонкой кишки содержит микроворсинки, которые выступают в тонкую кишку, тогда как подвижные эпителиальные клетки имеют реснички, выступающие с апикальной поверхности.Напротив, поверхность эпидермиса не имеет или имеет минимальную специализацию на апикальной поверхности, поскольку эти клетки подвергаются отслаиванию и истиранию.
Базальная поверхность служит для соединения нижнего слоя эпителиальной ткани с базальной мембраной. Базальная мембрана представляет собой тонкий неклеточный слой, похожий на оболочку. Он покрывает периферических нервов и мышц, а также подлежащую эпителиальной ткани. Базальная мембрана секретируется разными клетками.Он состоит из белков, таких как ламинин и коллаген. Интегрины — это белки плазматических мембран эпителиальных клеток, которые соединяют их с базальными мембранами.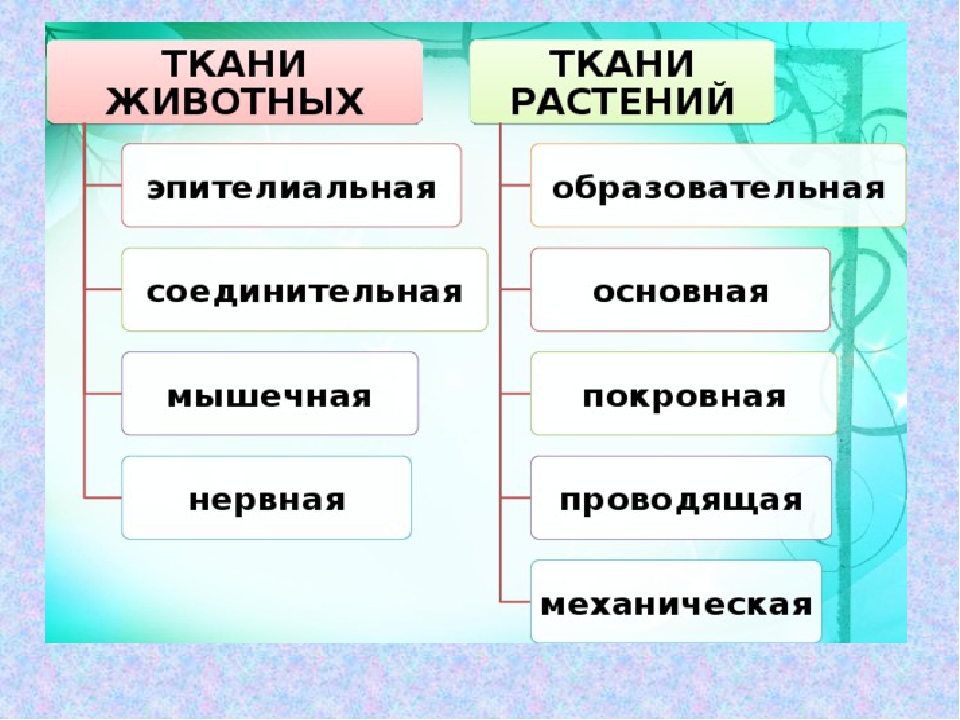 В секретирующих и поглощающих клетках функция апикальной поверхности увеличивает площадь поверхности.
В секретирующих и поглощающих клетках функция апикальной поверхности увеличивает площадь поверхности.
Боковая поверхность обеспечивает внутриклеточные связи между эпителиальными клетками. Эпителиальные клетки соединяются вместе, образуя эпителиальные ткани через плотные соединения около их апикальных концов; эти соединения не позволяют ионам, воде и растворенным веществам проникать в межклеточные пространства.
Рисунок 1: Различные типы эпителияСоединительная ткань
Чем соединительная ткань отличается от трех других основных типов тканей? Соединительные ткани содержат неживой внеклеточный материал больше, чем живые клетки. Среди соединительных тканей относительно мало клеток. Они встроены во внеклеточный матрикс. Клетки часто находятся далеко друг от друга, за исключением клеток жировой ткани, в которых адипоциты расположены близко друг к другу.
Основная функция соединительной ткани заключается в соединении различных клеток и тканей с помощью некоторых молекул, выделяемых из соединительной ткани во внеклеточный матрикс, образующий волокнистые ткани. Другие функции: обеспечивают защиту и поддержку мягких тканей, передают механические силы, и производят углеводы и белки.
Другие функции: обеспечивают защиту и поддержку мягких тканей, передают механические силы, и производят углеводы и белки.
Классификация соединительных тканей
Соединительные ткани человека и других позвоночных можно классифицировать следующим образом:
I. Собственно соединительная ткань
A. Плотная соединительная ткань — имеет больше волокон, чем основное вещество.
- Плотная правильная соединительная ткань — характеризуется волокнами коллагена, которые расположены явно в одном направлении, в частности, параллельно. Например, сухожилия и связки
- Плотная нерегулярная соединительная ткань — характеризуется коллагеновыми волокнами, которые, кажется, расположены во многих направлениях. Например, соединительные ткани, обнаруженные в нижних слоях кожи (дермы) и в защитном белом слое глазного яблока
Присоединяйтесь к нашему обсуждению тканей: гладкие мышцы против плотной нормальной соединительной ткани.
Будем рады услышать ваши идеи!
B. Рыхлая соединительная ткань — имеет больше основного вещества, чем волокна.
II. Особая соединительная ткань — соединительные ткани, не являющиеся волокнистыми. Примеры: ретикулярная соединительная ткань, жировая ткань, хрящ, кость и кровь.
В других источниках соединительная ткань классифицируется иначе; например, жировая ткань, особый тип соединительной ткани, классифицируется как рыхлая соединительная ткань (собственно соединительная ткань).
Клетки соединительной ткани
Клетки соединительной ткани различаются в зависимости от их структуры и функций. Например, фибробласты представляют собой тип клеток соединительной ткани, характеризующийся наличием большого количества эндоплазматического ретикулума и аппарата Гольджи. Они производят и секретируют волокна, такие как коллаген, ретикулярные волокна и эластичные волокна. Таким образом, они несут ответственность за поддержание внеклеточного матрикса (особенно во время заживления ран или восстановления тканей). Они обеспечивают структурную основу для многих тканей.
Они обеспечивают структурную основу для многих тканей.
На эмбриональной стадии мезенхимные клетки дифференцируются в различные типы соединительных тканей. Некоторые из них дифференцируются в остеобласты, образующие кости. Другие дифференцируются в хондробласты, образующие хрящи глубоких костей черепа.
Какой тип ткани — кровь? Кровь — одна из важнейших соединительных тканей в организме. Он в основном состоит из клеток крови и плазмы. Кровяные тельца — это белые кровяные тельца (лейкоциты) и красные кровяные тельца (эритроциты).Тромбоциты (тромбоциты) представляют собой клеточные фрагменты, поэтому в некоторых источниках они не рассматриваются как клетки крови. Все три типа происходят из мультипотентных стволовых клеток красного костного мозга. Функция белых кровяных телец — защита от инфекций и удаление клеточного мусора. Красные кровяные тельца переносят кислород по всему телу. Тромбоциты участвуют в образовании тромбов.
Волокна соединительной ткани
Волокнистые соединительные ткани богаты неклеточными компонентами, такими как волокна.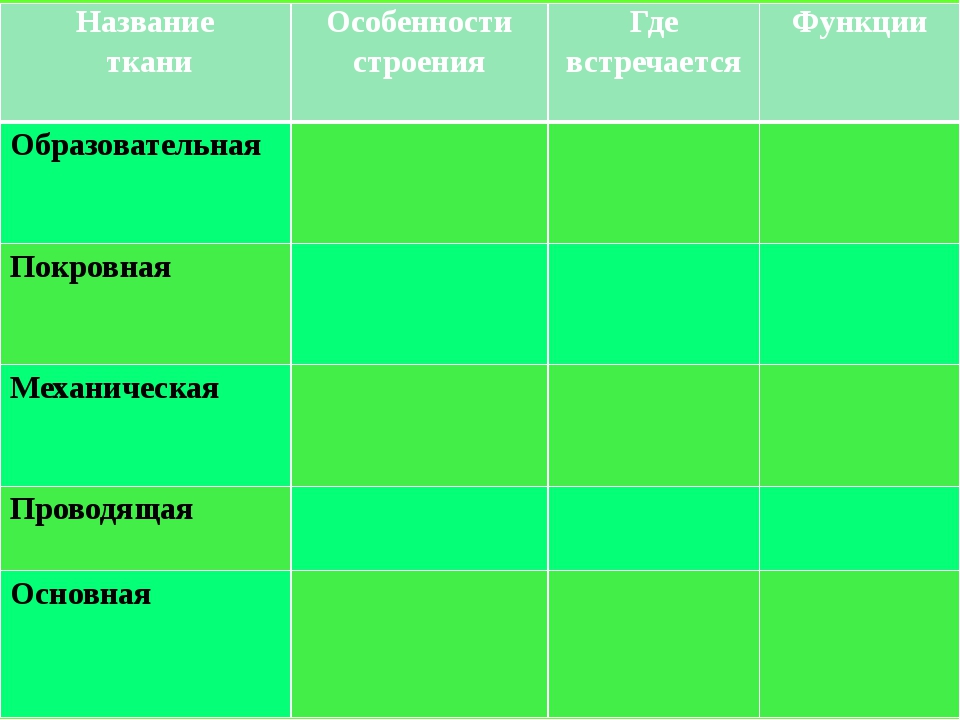 Эти волокна бывают разных типов: коллагеновые волокна, эластичные волокна и ретикулярные волокна. Коллагеновые волокна состоят из коллагена. Они находятся в сухожилиях и связках. Они поддерживают стенки внутренних органов. Они также составляют около одной трети веса кости.
Эти волокна бывают разных типов: коллагеновые волокна, эластичные волокна и ретикулярные волокна. Коллагеновые волокна состоят из коллагена. Они находятся в сухожилиях и связках. Они поддерживают стенки внутренних органов. Они также составляют около одной трети веса кости.
Другой тип соединительнотканных волокон — это эластичных волокон . Они в основном состоят из эластина. Их можно сравнить с резинками из-за их эластичных свойств. Эластичные волокна могут сохранять свою первоначальную форму после деформации, например, дерма кожи, крупные артерии и легкие, где эти ткани, содержащие эластичные волокна, могут растягиваться и сокращаться в ответ на различные раздражители.
Ретикулярные волокна — это волокна, состоящие в основном из коллагена III типа, секретируемого ретикулярными клетками. Эти волокна находятся в разных частях тела, таких как соединительные ткани, поддерживающие жиры, или лимфоидная ткань.
Основное вещество
Основное вещество состоит в основном из протеогликанов, гликозаминогликанов, гликопротеинов, воды и ионов.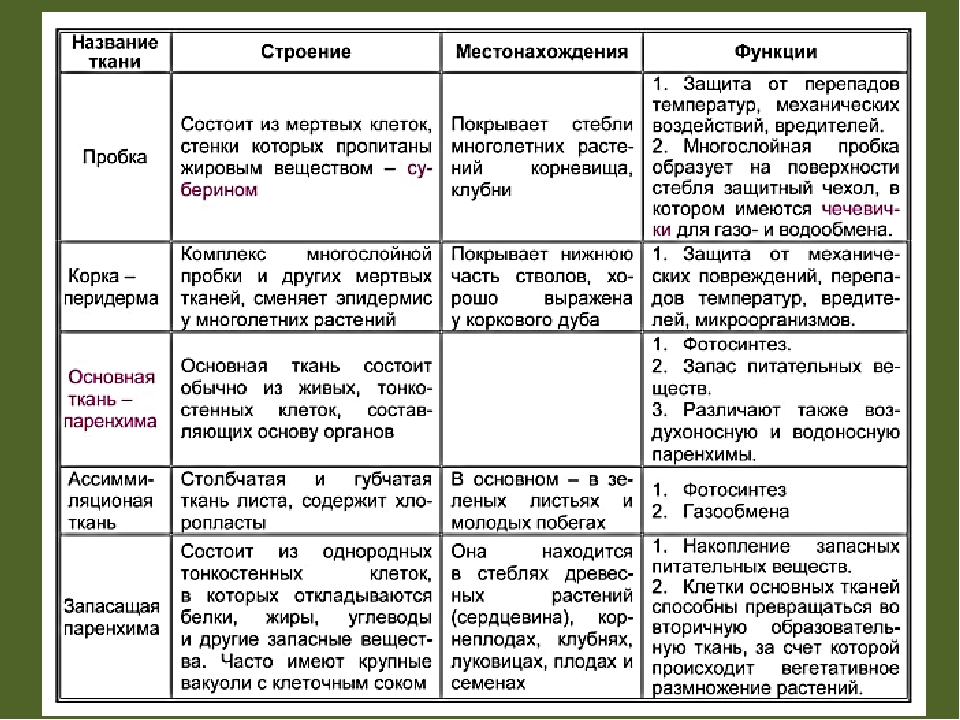 Они служат заполнителями пространства, поглощают удары, смазывают и ограничивают движение бактерий в соединительных тканях.
Они служат заполнителями пространства, поглощают удары, смазывают и ограничивают движение бактерий в соединительных тканях.
Мышечная ткань
Мышцы — это сократительные ткани в организме. Мышечная ткань подразделяется на три типа: (1) скелетных мышц , которые прикреплены к костям и помогают в движении тела, (2) гладких мышц , которые образуют внутреннюю оболочку кровеносных сосудов, и (3) сердечные мышцы, образующие сердце.
Мышечная ткань подразделяется на три типа: (1) скелетных мышц , которые прикреплены к костям и помогают в движении тела, (2) гладких мышц , которые образуют внутреннюю оболочку кровеносных сосудов, и (3) сердечные мышцы, образующие сердце.
Скелетные мышцы отвечают за произвольные движения тела. Гладкие мышцы предназначены для непроизвольного движения, подобно сердечным мышцам. Однако сердечные мышцы похожи на скелетные мышцы тем, что имеют заметные бороздки при просмотре под микроскопом. Сердечные мышцы также отличаются от двух других наличием вставных дисков.
ЧИТАТЬ: Физиология человека — Мышцы
Гладкая мышечная ткань
Гладкие мышцы не имеют бороздок при просмотре под микроскопом.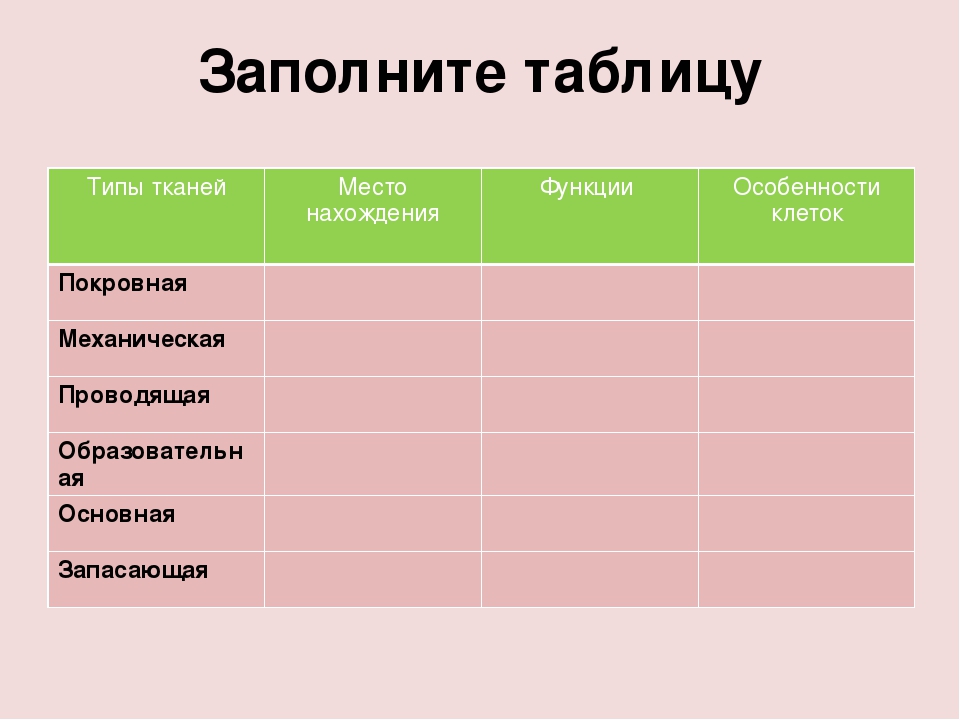 Они состоят из узких веретенообразных клеток с одним центрально расположенным ядром. У них отсутствуют характерные бороздки, потому что их актиновые и миозиновые филаменты не организованы аналогично скелетным и сердечным мышечным тканям. Расположение актина и миозина в клетках гладких мышц позволяет гладким мышцам сокращаться дольше, чем скелетным мышцам, с меньшим количеством энергии. Следовательно, гладкие мышцы обладают способностью поддерживать тонус сосудов или сокращение матки во время родов.Гладкие мышцы выстилают стенки трубок тела, таких как пищеварительная система, потому что они отвечают за движение материала по этим трубам. Наблюдайте за сокращением гладких мышц ниже.
Они состоят из узких веретенообразных клеток с одним центрально расположенным ядром. У них отсутствуют характерные бороздки, потому что их актиновые и миозиновые филаменты не организованы аналогично скелетным и сердечным мышечным тканям. Расположение актина и миозина в клетках гладких мышц позволяет гладким мышцам сокращаться дольше, чем скелетным мышцам, с меньшим количеством энергии. Следовательно, гладкие мышцы обладают способностью поддерживать тонус сосудов или сокращение матки во время родов.Гладкие мышцы выстилают стенки трубок тела, таких как пищеварительная система, потому что они отвечают за движение материала по этим трубам. Наблюдайте за сокращением гладких мышц ниже.
Гладкие мышцы, а также сердечные мышцы сокращаются непроизвольно, означает, что они не контролируются сознательными требованиями человека. Напротив, скелетные мышцы — это произвольные мышцы, которые следуют за сознательным контролем.
Чем соединительная ткань отличается от мышечной? Получите ответ здесь: гладкие мышцы против плотной нормальной соединительной ткани.
Присоединяйтесь к нашему форуму прямо сейчас!
Скелетная мышечная ткань
Скелетные мышцы представляют собой многоядерные ткани из слитых клеток. Мышечные клетки сливаются вместе, образуя мышечных трубок . Ядра мышечных трубок крупные, удлиненные, с большим ядрышком. Ткани скелетных мышц производят большое количество белков и РНК. РНК и белки стимулируют сокращение мышц после того, как белки организуются в массивы, называемые саркомерами. Посмотрите видео ниже, чтобы увидеть, как сокращаются скелетные мышцы.
Скелетные мышцы выделяют базальную пластинку, окружающую мышцу. Базальная пластинка помогает передавать сокращение мышц окружающим тканям. Клетки-сателлиты расположены между базальной пластинкой и мышечным волокном. Клетки-сателлиты помогают в развитии и регенерации мышечных волокон.
Существует два основных типа тканей скелетных мышц: быстрые и медленные мышцы. Они классифицируются по потреблению кислорода при заключении контракта.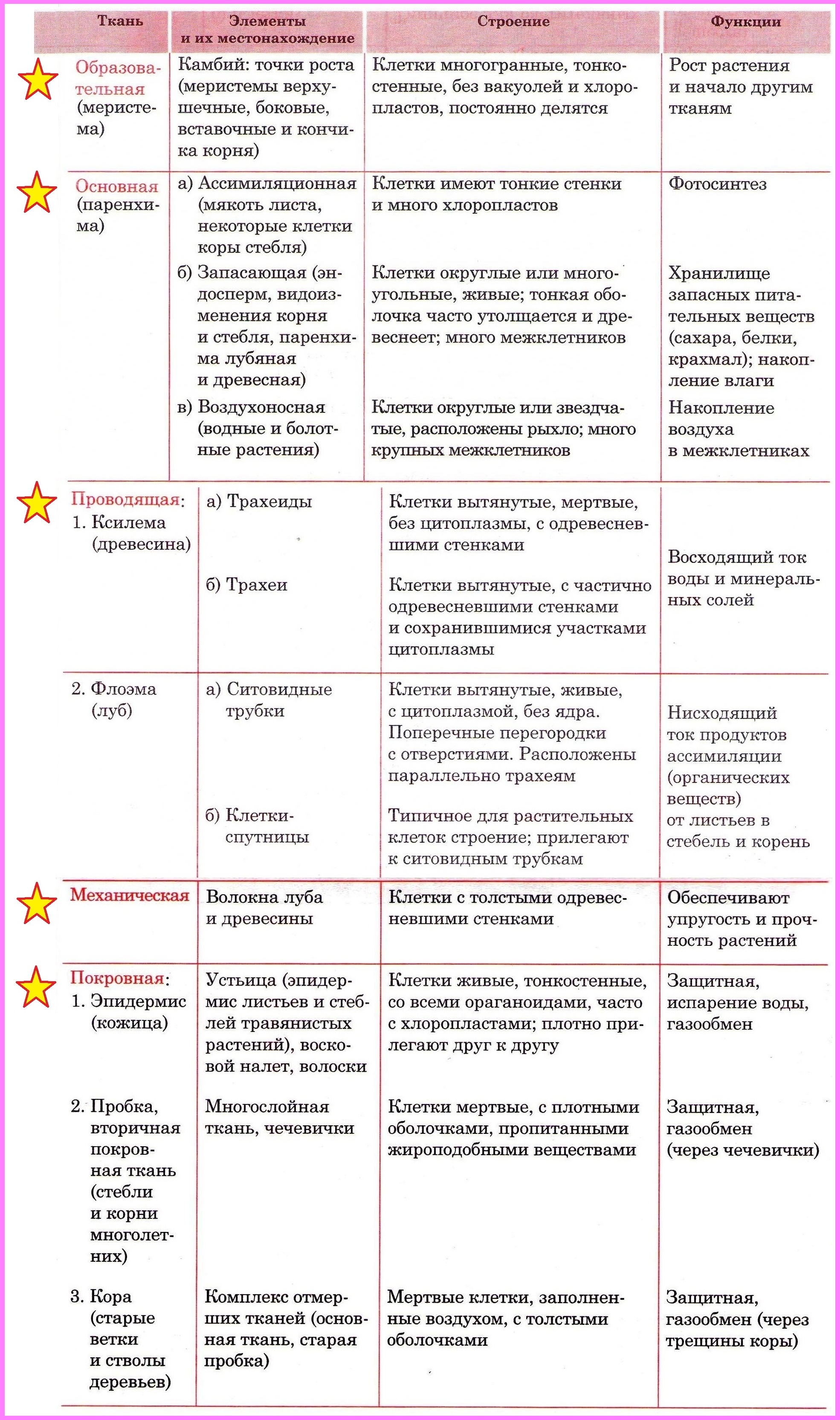 Быстрые мышцы толще, сильнее и сокращаются на меньшее время, чем медленные.
Быстрые мышцы толще, сильнее и сокращаются на меньшее время, чем медленные.
Ткань сердечной мышцы
Сердечная мышца — поперечно-полосатая мышца. Они контактируют как скелетные мышцы; однако это непроизвольные мышцы. Еще одно различие между скелетными и сердечными мышцами состоит в том, что сердечные мышцы содержат только одно ядро, в отличие от многоядерных скелетных мышц.
Сердечные мышцы богаты кровеносными сосудами и митохондриями, поскольку они нуждаются в активном метаболизме для поддержания их непрерывного сокращения.В отличие от скелетных мышц сердечные мышцы должны одновременно сокращаться, чтобы перекачивать кровь из сердца. Этот эффект достигается наличием соединений между сердечными мышцами; Эти соединения связывают каждую мышцу со следующей, что помогает в передаче сигнала через все сердце.
Кардиомиоциты (сердечные мышцы) не содержат сателлитных клеток, как в скелетных мышцах; следовательно, сердечные мышцы не делятся и не дифференцируются. Вместо этого они растут только за счет увеличения своего размера.
Вместо этого они растут только за счет увеличения своего размера.
ПРОЧИТАЙТЕ: Анатомия сердца — онлайн-уроки биологии
Нервная ткань
Нервные ткани — это ткани, которые образуют центральную нервную систему и периферическую нервную систему. Нервная клеточная ткань образует головной и спинной мозг в центральной нервной системе, тогда как в периферической нервной системе они образуют сенсорные, двигательные и вегетативные нервы.
Нервная ткань состоит из крупных клеток, называемых нейронами; у них длинное тело, достигающее метра.Нейроны имеют клеточные отростки дендритов или аксонов . Эти клеточные процессы переносят сигналы через разные нейроны от тела клетки к нервному окончанию. В частности, дендриты несут сигналы от периферии к телу клетки. Тело клетки содержит многочисленные органеллы, которые выполняют его функцию, включая рибосомы, цитоплазму и аппарат Гольджи. С другой стороны, аксоны содержат большое количество цитоплазмы по сравнению с телом клетки. Они также содержат микрофиламенты и микротрубочки, которые переносят сигналы и поддерживают клетку.
Они также содержат микрофиламенты и микротрубочки, которые переносят сигналы и поддерживают клетку.
Каждый аксон разветвляется на области, известные как пресинаптические окончания. На пресинаптических окончаниях электрический импульс преобразуется в химические нейротрансмиттеры, которые посылают сигналы следующему нейрону или мышечной клетке в цепи. Нейротрансмиттеры хранятся в карманах и высвобождаются в результате изменения проницаемости клеточной мембраны, когда электрический сигнал достигает синапса.Нейротрансмиттеры высвобождаются для связывания со своим рецептором в следующей клетке, продолжая передачу нервного импульса, пока он не достигнет ткани-мишени. Функция нервной ткани заключается в передаче нервных импульсов по разным нервным клеткам для управления всеми произвольными и непроизвольными действиями в организме.
Основные типы тканей у животных следующие:
Примеры тканей растений
Что такое ткань в биологии растений? Мы уже знаем, что биологическая ткань состоит из ряда клеток, выполняющих определенную функцию.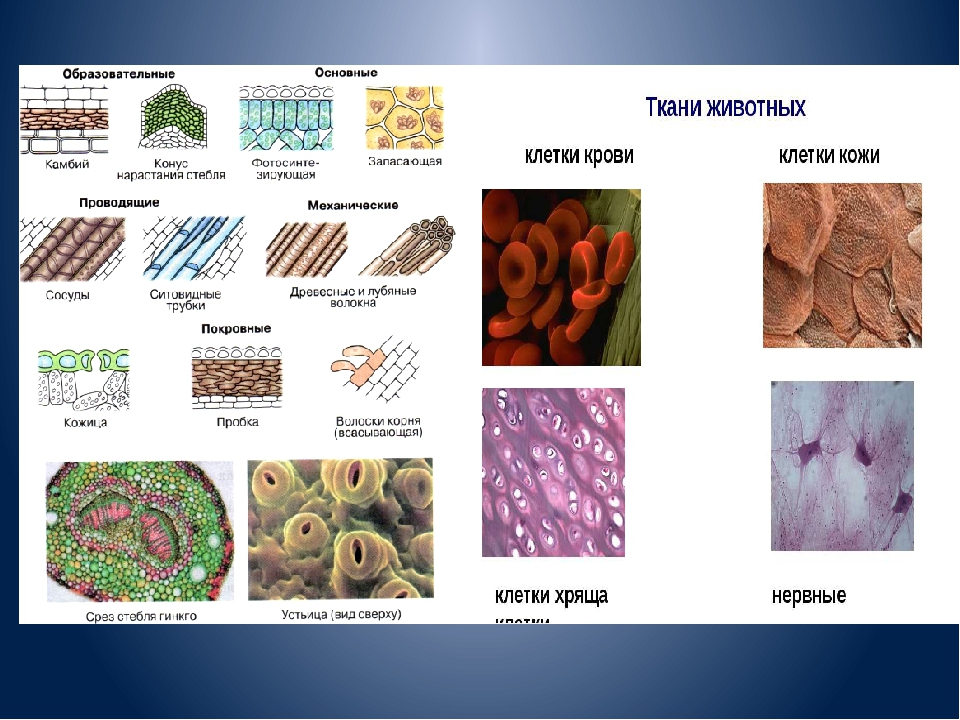 У растений ткани также состоят из клеток, выполняющих определенную функцию. Ниже приведены некоторые примеры тканей растений, которые выполняют определенную функцию. Например, эпидермис растений — это ткань, обеспечивающая защиту. Наземные ткани — это ткани, состоящие из основных растительных клеток, которые выполняют определенную функцию. Сосудистые ткани — это ткани растений, участвующие в транспортировке питательных веществ по телу растения. Давайте рассмотрим их подробнее ниже.
У растений ткани также состоят из клеток, выполняющих определенную функцию. Ниже приведены некоторые примеры тканей растений, которые выполняют определенную функцию. Например, эпидермис растений — это ткань, обеспечивающая защиту. Наземные ткани — это ткани, состоящие из основных растительных клеток, которые выполняют определенную функцию. Сосудистые ткани — это ткани растений, участвующие в транспортировке питательных веществ по телу растения. Давайте рассмотрим их подробнее ниже.
Эпидермальная ткань растения
Эпидермальная ткань растения (эпидермис) является самой внешней защитной тканью растения.Он покрывает поверхность растения, защищая внутренние слои от внешней среды. Он покрыт слоем кутикулы, который делает эпидермис непроницаемым для воды и, следовательно, помогает предотвратить потерю воды.
Рисунок 5: Верхний и нижний эпидермис листа растения. Кредит: Zephyris CC BY-SA 3.0 Рис. 6. Кутикула на поверхности листа делает его непроницаемым для воды. Эпидермис растения защищает его от ультрафиолетового излучения солнца. Однако он обеспечивает обмен газов между окружающей средой и растением через устьица.Устьица эпидермальной ткани состоят из двух замыкающих клеток . Вместе они образуют устьичную пору. Внутренние стенки замыкающих ячеек толще их наружных. Замыкающие клетки имеют почковидную форму. Они прикреплены друг к другу своими концами с помощью окружающих их микрофибрилл целлюлозы. Защитные клетки расширяются, когда они поглощают воду, но из-за наличия микрофибрилл целлюлозы они расширяются в продольном направлении, а не широко, создавая поры, которые позволяют газообмену.
Однако он обеспечивает обмен газов между окружающей средой и растением через устьица.Устьица эпидермальной ткани состоят из двух замыкающих клеток . Вместе они образуют устьичную пору. Внутренние стенки замыкающих ячеек толще их наружных. Замыкающие клетки имеют почковидную форму. Они прикреплены друг к другу своими концами с помощью окружающих их микрофибрилл целлюлозы. Защитные клетки расширяются, когда они поглощают воду, но из-за наличия микрофибрилл целлюлозы они расширяются в продольном направлении, а не широко, создавая поры, которые позволяют газообмену.
Эпидермальные клетки могут иметь отростки, называемые трихомами . Трихомы защищают растение от травоядных и насекомых, выделяя токсины или блокируя доступ к поверхности растения.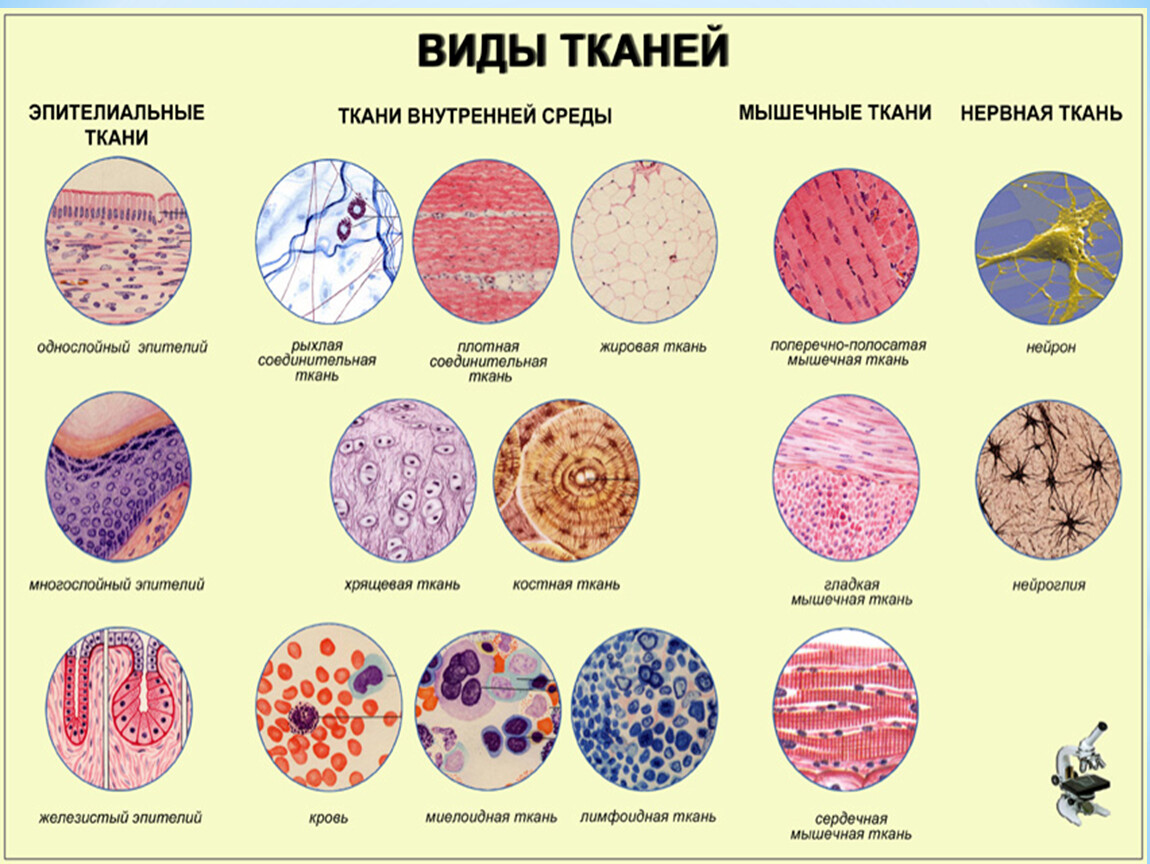
В отличие от эпидермиса побегов, эпидермис корня пропускает воду из почвы к растениям.Он покрыт более тонкой кутикулой, содержащей более короткий воск. Корни разрастаются, образуя корневые волоски; эти волоски увеличивают впитывающую площадь корня (повышенное потребление воды и питательных веществ). Эпидермис корня обычно производит гидрофильный углевод, который задерживает воду, чтобы облегчить ее абсорбцию из почвы и смазывать область, через которую проходит корень.
Наземная ткань
Наземная ткань представляет собой все ткани, образующие растение, кроме эпидермальных или сосудистых тканей.Наземные ткани делятся на три основных типа растительных клеток: паренхима, колленхима и склеренхима в соответствии со структурой их клеточной стенки.
Клетки паренхимы имеют тонкую стенку. Они занимают большую часть коры, которая является областью между другими слоями, эпидермальным, сосудистым и сердцевинным. Паренхима накапливает в растении белки, крахмал и масла. Они также способствуют росту, развитию и поддержке растения.
Они также способствуют росту, развитию и поддержке растения.
Клетки колленхимы подобны паренхиматозным клеткам тем, что имеют первичную клеточную стенку.Материалы клеточной стенки откладываются по краям клетки, образуя колленхиму. Клетки колленхимы способствуют удлинению стеблей растений. Однако они обеспечивают поддержку только тогда, когда растение набухло, потому что в их клеточных стенках отсутствуют гидрофильные вещества.
Клетки склеренхимы имеют толстые стенки, содержащие лигнин. Такая структура делает склеренхиму водонепроницаемой и прочной. Склеренхима поддерживает растение больше, чем паренхима и колленхима. Однако сделать это труднее, поэтому его не так много, как других видов измельченной ткани.Благодаря своей жесткости склеренхима может включаться в структуру тканей сосудов, образовывать трахеиды и сосуды ксилемы.
Рис. 8: Различные типы тканей растений. Кредит: Кельвинсонг — диаграмма, CC BY 3.0.Сосудистая ткань
Сосудистая ткань растения представляет собой главную систему, отвечающую за транспортировку в растении, она состоит из ксилемы и флоэмы. Сосудистая ткань присутствует во всех частях растения в виде сосудистых пучков, стеблей, листьев и пересекающих корней.Ксилема транспортирует различные материалы, такие как вода и ионы, от корней к листьям и стеблям растений. Флоэма транспортирует метаболиты, такие как аминокислоты, сахара и ионы, вырабатываемые растением, с места производства в листьях в различные области, такие как корни, плоды и листья.
Сосудистая ткань присутствует во всех частях растения в виде сосудистых пучков, стеблей, листьев и пересекающих корней.Ксилема транспортирует различные материалы, такие как вода и ионы, от корней к листьям и стеблям растений. Флоэма транспортирует метаболиты, такие как аминокислоты, сахара и ионы, вырабатываемые растением, с места производства в листьях в различные области, такие как корни, плоды и листья.
Флоэма в основном состоит из клеток-компаньонов , клеток паренхимы флоэмы , ситчатых трубок, и волокон флоэмы . Ситовые пластины разделяют ситовые трубки, эти трубки имеют поры, обозначенные как , ситовые поры .Несмотря на то, что в элементах сита отсутствуют ядра, они представляют собой живые клетки, содержащие плазмалемму и белки. Элементы-компаньоны окружают элементы сита. Они содержат крупные ядра, вакуоли и плотную цитоплазму. Клетки-компаньоны транспортируют метаболиты в ситовые элементы и из них.
Ксилема состоит из ксилемных сосудов, трахеид, паренхимных клеток ксилемы, и ксилемных волокон . Большие пластинки соединяют сосуды вместе, оставляя промежутки между сосудами, в то время как концы трахеид перекрывают следующие клетки без промежутков между ними.Лигнин распределен в сосудах в различных формах, таких как спиралевидные, сетчатые, кольцевые, чешуйчатые и с ямками. Его можно транспортировать как в продольном, так и в поперечном направлении через поры или ямки пластин.
У растений различные типы тканей:(1) Эмбриональные или меристематические ткани (например, апикальная меристема и камбий)
(2) Постоянные ткани (например, эпидермис, пробка, трихома)
- Фундаментальные ( например, паренхима, колленхима, склеренхима)
- Комплекс (e.г. ткани флоэмы и ксилемы)
(3) Репродуктивные ткани (т. е. спорогенные ткани).
Тканевая инженерия
Тканевая инженерия — это метод получения тканей вне тела, он сочетает в себе биологические и инженерные науки для восстановления или создания новых тканей. Эта наука может сильно повлиять на сферу медицины, улучшая восстановление поврежденных тканей из-за ожогов, травм или заболеваний путем создания искусственного трансплантата тканей. Несмотря на то, что этот подход кажется многообещающим, он сталкивается с некоторыми трудностями, такими как создание искусственных кровеносных сосудов для питания и снабжение тканей циркулирующей кровью.Другие проблемы включают производство и использование биореакторов для создания тканей с использованием точного количества питательных веществ, ферментов и клеточных компонентов. Дифференциация клеток в культуре может не контролироваться, поэтому произведенная ткань будет гипертрофирована и может содержать более одного ядра на клетку, что приведет к образованию деформированной ткани. Таким образом, тканевая инженерия — это новая многообещающая наука, которая подразумевает важность и влияние культивирования тканей в области медицины.
Эта наука может сильно повлиять на сферу медицины, улучшая восстановление поврежденных тканей из-за ожогов, травм или заболеваний путем создания искусственного трансплантата тканей. Несмотря на то, что этот подход кажется многообещающим, он сталкивается с некоторыми трудностями, такими как создание искусственных кровеносных сосудов для питания и снабжение тканей циркулирующей кровью.Другие проблемы включают производство и использование биореакторов для создания тканей с использованием точного количества питательных веществ, ферментов и клеточных компонентов. Дифференциация клеток в культуре может не контролироваться, поэтому произведенная ткань будет гипертрофирована и может содержать более одного ядра на клетку, что приведет к образованию деформированной ткани. Таким образом, тканевая инженерия — это новая многообещающая наука, которая подразумевает важность и влияние культивирования тканей в области медицины.
Попробуйте ответить на приведенный ниже тест, чтобы проверить, что вы узнали о тканях.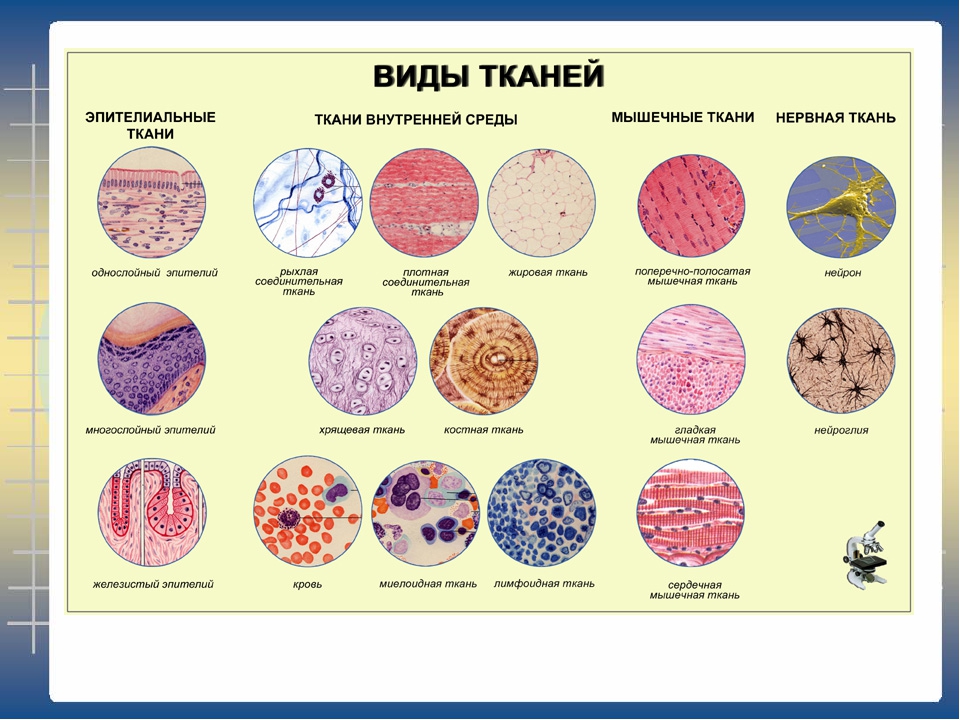
Обзор заживления ран в различных типах тканей — постоянные нейронные имплантаты
1.1. ВВЕДЕНИЕ И ОБЗОР
Неизбежный ответ на любой имплант — заживление ран, состоящее из гемостаза, воспаления, восстановления и ремоделирования. Для неразлагаемых имплантатов с гладкой поверхностью ремонт и ремоделирование приводит к изоляции имплантата посредством инкапсуляции ткани. Природа инкапсулируемой ткани и клеточных участников иммунной реакции, приводящей к этому результату, варьируется в зависимости от места имплантации и типа ткани, в которой находится имплант (не говоря уже о навыках хирурга).
В настоящее время точно установлено, что процесс заживления ран оказывает существенное пагубное влияние на точность и надежность имплантированных датчиков и электродов. Ряд обзоров [1–6] и множество первичных статей посвящены этой проблеме для имплантированных сенсоров, в первую очередь для глюкозных электродов; однако существует только один обзор [7] и ограниченное количество первичных статей, касающихся электродов, имплантированных в ткань центральной нервной системы (ЦНС) [8–11].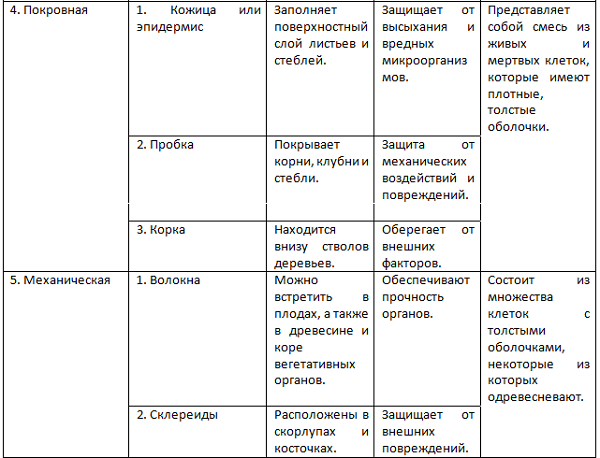 В этой главе приводится краткое изложение процесса заживления ран в ткани ЦНС и сравнивается его с процессом заживления ран в периферической нервной системе (ПНС), костях и коже.
В этой главе приводится краткое изложение процесса заживления ран в ткани ЦНС и сравнивается его с процессом заживления ран в периферической нервной системе (ПНС), костях и коже.
Недавние сообщения показали, что имплантированные электроды могут контролировать нервные сигналы в ЦНС и, следовательно, могут использоваться для контроля протезирования у пациентов, страдающих полным или частичным параличом [12,13]. Хотя первоначальные исследования являются многообещающими, при хронической имплантации датчиков и электродов часто и непредсказуемо возникают неисправности компонентов или осложнения, связанные с реакцией на заживление ран [7]. Процесс заживления ран начинается, как только электроды вводятся в мозг, что неизбежно нарушает гематоэнцефалический барьер [14].Нарушение гематоэнцефалического барьера приводит к гемостазу и начальным стадиям воспаления в ткани ЦНС, которые типичны для всех васкуляризированных тканей. Однако отчетливые различия возникают в ткани ЦНС на более поздних стадиях воспаления, восстановления и ремоделирования ().
Рисунок 1.1
Резюме реакции заживления ран в головном мозге по сравнению с неневральной тканью.
Первое отличие — гематоэнцефалический барьер. ЦНС сильно васкуляризована; однако поддержание барьера между капиллярами и окружающей тканью важно для регулирования концентрации ионов и других молекул как в сосудистом, так и в внесосудистом пространстве ЦНС [15].Эндотелиальные клетки, образующие капилляры в ЦНС, отличаются от клеток остального тела в двух отношениях. Во-первых, они способны образовывать плотные контакты, ограничивая параклеточный поток, и, во-вторых, у них очень мало эндоцитотических пузырьков, ограничивающих трансцеллюлярное движение растворенных веществ из крови в межклеточные промежутки мозга (обзор Rubin and Staddon [16]). В результате гематоэнцефалический барьер препятствует проникновению практически всех молекул крови, за исключением небольших и липофильных, таких как стероиды.Однако некоторые более крупные молекулы, такие как глюкоза и определенные аминокислоты, все еще могут перемещаться в ткань мозга через определенные белки-переносчики [16].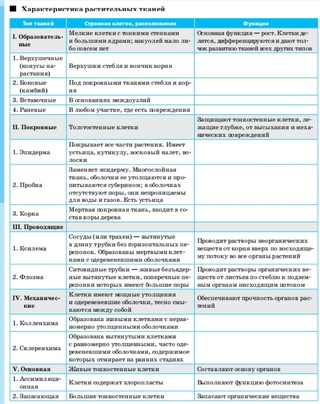 Плотная базальная мембрана и отростки астроцитов, называемые концевыми ножками, окружают эндотелиальные клетки капилляров, внося дополнительный вклад в гематоэнцефалический барьер. В нормальных непатологических условиях эндотелий сосудов ЦНС с его плотными контактами полностью отделяет ткань мозга от циркулирующей крови, изолируя уникальные типы клеток мозга, одновременно защищая нервную ткань от потенциально вредных иммунных реакций, происходящих вне нервной ткани в организме [17].Это важно, потому что, в отличие от большинства периферических тканей, ЦНС функционирует через сеть нейронов, которая обычно неспособна к пролиферации и не может быть заменена при повреждении.
Плотная базальная мембрана и отростки астроцитов, называемые концевыми ножками, окружают эндотелиальные клетки капилляров, внося дополнительный вклад в гематоэнцефалический барьер. В нормальных непатологических условиях эндотелий сосудов ЦНС с его плотными контактами полностью отделяет ткань мозга от циркулирующей крови, изолируя уникальные типы клеток мозга, одновременно защищая нервную ткань от потенциально вредных иммунных реакций, происходящих вне нервной ткани в организме [17].Это важно, потому что, в отличие от большинства периферических тканей, ЦНС функционирует через сеть нейронов, которая обычно неспособна к пролиферации и не может быть заменена при повреждении.
Ткань ЦНС отличается от тканей, не относящихся к ЦНС, наличием типов клеток, которые уникальны для ЦНС. Нейроны, которые состоят из тела клетки и ее аксона, являются функциональной единицей ткани мозга, хотя на них приходится менее 25% от общей популяции клеток [7]. Примерно половину общего объема ЦНС занимают три типа поддерживающих глиальных клеток соединительной ткани: олигодендроциты, астроциты и микроглия.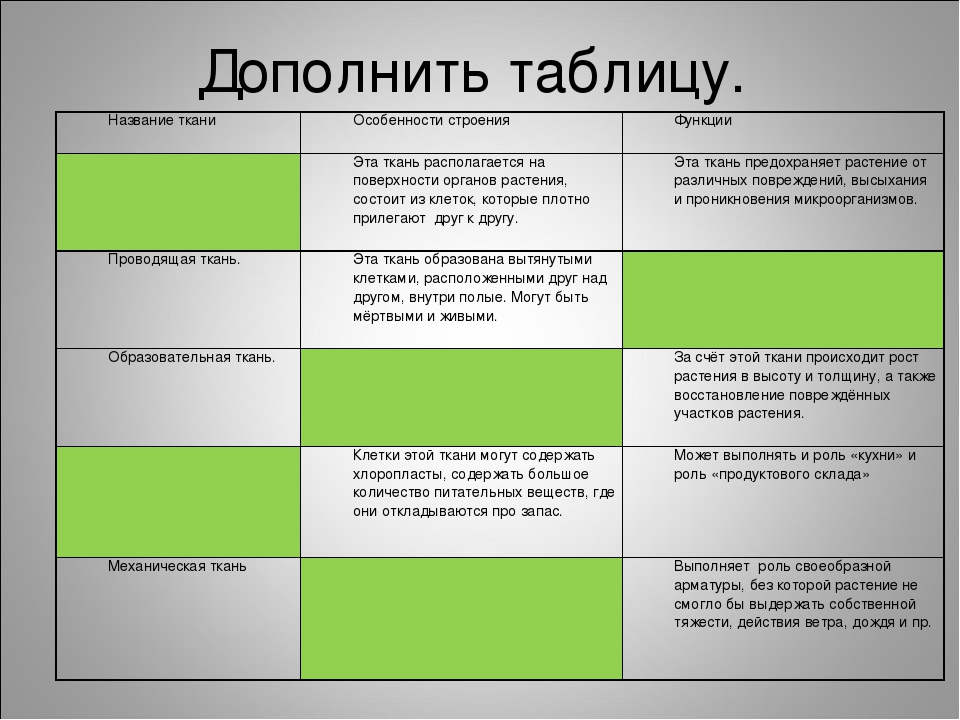 Каждый тип глиальных клеток выполняет функции, обеспечивающие здоровье нейронов ЦНС. Олигодендроциты обвиваются вокруг аксонов, чтобы обеспечить поддержку и произвести богатую липидами миелиновую оболочку для защиты нервных процессов; астроциты выполняют множество функций нервной поддержки, но примечательны тем, что формируют глиальную границу между ЦНС и структурами, не относящимися к ЦНС, для создания гематоэнцефалического барьера; микроглия имеет много общих фенотипических и функциональных характеристик с макрофагами, полученными из крови, существующими в состоянии покоя в тканях мозга до тех пор, пока они не активируются, чтобы управлять врожденными иммунными ответами мозга [7,18].В отличие от ЦНС, аксоны в ПНС имеют непрерывную миелиновую оболочку, продуцируемую шванновскими клетками. Сплошная миелиновая оболочка увеличивает скорость распространения нервного импульса, что важно для аксонов, которые проходят через ПНС на большие расстояния.
Каждый тип глиальных клеток выполняет функции, обеспечивающие здоровье нейронов ЦНС. Олигодендроциты обвиваются вокруг аксонов, чтобы обеспечить поддержку и произвести богатую липидами миелиновую оболочку для защиты нервных процессов; астроциты выполняют множество функций нервной поддержки, но примечательны тем, что формируют глиальную границу между ЦНС и структурами, не относящимися к ЦНС, для создания гематоэнцефалического барьера; микроглия имеет много общих фенотипических и функциональных характеристик с макрофагами, полученными из крови, существующими в состоянии покоя в тканях мозга до тех пор, пока они не активируются, чтобы управлять врожденными иммунными ответами мозга [7,18].В отличие от ЦНС, аксоны в ПНС имеют непрерывную миелиновую оболочку, продуцируемую шванновскими клетками. Сплошная миелиновая оболочка увеличивает скорость распространения нервного импульса, что важно для аксонов, которые проходят через ПНС на большие расстояния.
В последующих разделах этой главы освещаются характеристики каждой стадии реакции заживления ран (т.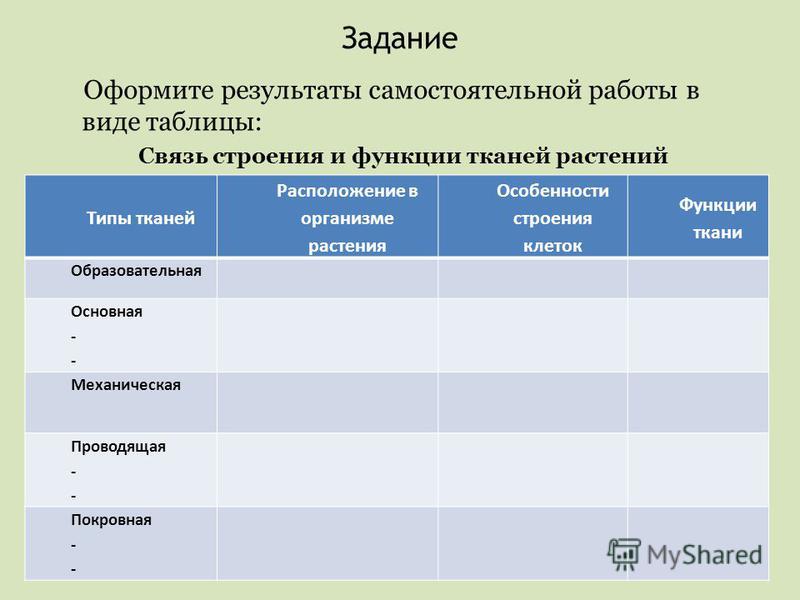 Е. Гемостаза, воспаления, заживления, ремоделирования), при этом выявляются сходства и различия в основных типах тканей.Однако важно отметить, что не существует определенных границ между стадиями, поскольку реакция заживления раны «переходит» в следующую стадию заживления.
Е. Гемостаза, воспаления, заживления, ремоделирования), при этом выявляются сходства и различия в основных типах тканей.Однако важно отметить, что не существует определенных границ между стадиями, поскольку реакция заживления раны «переходит» в следующую стадию заживления.
1.2. ГЕМОСТАЗ (ОТ СЕКУНД ДО ЧАСОВ)
Общий для всех типов тканей процесс заживления ран начинается сразу после травмы, поскольку ткань разрушается и кровеносные сосуды разрываются, высвобождая плазму крови и клетки периферической крови в место раны. Самыми ранними сигналами повреждения тканей являются выброс таких молекул, как АТФ, и воздействие коллагена на стенку кровеносных сосудов [19].В ЦНС черепно-мозговая травма разрушает гематоэнцефалический барьер, поскольку сыворотка и клетки крови попадают в место раны [14]. Во всех тканях образуется сгусток, который действует как временный барьер, предотвращающий чрезмерное кровотечение и ограничивающий распространение болезнетворных микроорганизмов в кровоток. Первичный гемостаз происходит, когда тромбоциты прикрепляются к коллагеновым волокнам, обнаженным в поврежденном эндотелии, с использованием специфических гликопротеинов рецептора коллагена (GPIb / IX / V) для формирования первичной гемостатической пробки [19]. Когда тромбоциты прикрепляются к участку поражения, они быстро активируют высокоаффинный интегрин тромбоцитов αIIbβ3, который опосредует агрегацию тромбоцитов [19].Как только тромбоциты связываются, они активируются и дегранулируют, высвобождая свое содержимое в плазму, чтобы стимулировать местную активацию факторов свертывания плазмы (). Эти факторы запускают образование фибринового сгустка () [20]. Воздействие на плазму крови тканевого фактора, продуцируемого субэндотелиальными клетками, обычно не контактирующими с кровью, такими как гладкомышечные клетки и фибробласты [21], или с чужеродными поверхностями, такими как имплантаты, инициирует ускоренный каскад активированных белков, который приводит к образованию фибрина. .Расщепление фибриногена на мономеры фибрина, его полимеризация и поперечное сшивание формируют переплетенную желатиноподобную тромбоцитарную пробку, производящую стабильный сгусток [22].
Первичный гемостаз происходит, когда тромбоциты прикрепляются к коллагеновым волокнам, обнаженным в поврежденном эндотелии, с использованием специфических гликопротеинов рецептора коллагена (GPIb / IX / V) для формирования первичной гемостатической пробки [19]. Когда тромбоциты прикрепляются к участку поражения, они быстро активируют высокоаффинный интегрин тромбоцитов αIIbβ3, который опосредует агрегацию тромбоцитов [19].Как только тромбоциты связываются, они активируются и дегранулируют, высвобождая свое содержимое в плазму, чтобы стимулировать местную активацию факторов свертывания плазмы (). Эти факторы запускают образование фибринового сгустка () [20]. Воздействие на плазму крови тканевого фактора, продуцируемого субэндотелиальными клетками, обычно не контактирующими с кровью, такими как гладкомышечные клетки и фибробласты [21], или с чужеродными поверхностями, такими как имплантаты, инициирует ускоренный каскад активированных белков, который приводит к образованию фибрина. .Расщепление фибриногена на мономеры фибрина, его полимеризация и поперечное сшивание формируют переплетенную желатиноподобную тромбоцитарную пробку, производящую стабильный сгусток [22].
Рисунок 1.2
Гемостаз в поврежденном кровеносном сосуде. (а) Повреждение стенки сосуда вызывает выброс переносимых кровью клеток, белков и тромбоцитов на периферию. Тромбоциты активируются при контакте с окружающим коллагеном вне крови (подробнее …)
Активация тромбоцитов также вызывает высвобождение ряда сигнальных молекул, таких как фактор роста тромбоцитов (PDGF) [23], трансформируя фактор роста-β (TGF-β) [24] и фактор роста эндотелия сосудов (VEGF) [25] из их цитоплазматических гранул.Воспалительные и репаративные клетки хемотаксически притягиваются к «резервуару» молекул, хранящемуся внутри сгустка, который вызывает воспаление, следующий шаг в последовательности заживления.
1.3. ВОСПАЛЕНИЕ (ЧАСЫ И ДНЯ)
1.3.1. Начальные события
Процесс воспаления содержит, нейтрализует или разбавляет вызывающий травму агент или поражение, независимо от типа ткани [26]. Как в ЦНС, так и вне ЦНС тканевая среда, в которой начинается воспаление, представляет собой смесь поврежденной ткани, компонентов сгустка (тромбоцитов, эритроцитов и фибрина), экстравазированных белков сыворотки и инородного материала, введенного во время травмы. [27].Воспаление инициируется высвобождением сигнальных молекул из раны во время гемостаза [28]. Молекулы хемоаттрактантов, высвобождаемые тромбоцитами, также увеличивают вазодилатацию и проницаемость сосудов, впоследствии увеличивая рекрутирование лейкоцитов. Поступающие лейкоциты распознают белки плазмы, такие как фибронектин, витронектин и тромбоспондин, которые пассивно абсорбируются сгустком. Первоначальный процесс воспаления является общим для всех типов тканей, поскольку любая физическая травма обычно приводит к образованию тромбоцитарной пробки и привлечению воспалительных клеток, переносимых кровью, к месту повреждения.В ткани ЦНС повреждение также приводит к активации микроглии.
[27].Воспаление инициируется высвобождением сигнальных молекул из раны во время гемостаза [28]. Молекулы хемоаттрактантов, высвобождаемые тромбоцитами, также увеличивают вазодилатацию и проницаемость сосудов, впоследствии увеличивая рекрутирование лейкоцитов. Поступающие лейкоциты распознают белки плазмы, такие как фибронектин, витронектин и тромбоспондин, которые пассивно абсорбируются сгустком. Первоначальный процесс воспаления является общим для всех типов тканей, поскольку любая физическая травма обычно приводит к образованию тромбоцитарной пробки и привлечению воспалительных клеток, переносимых кровью, к месту повреждения.В ткани ЦНС повреждение также приводит к активации микроглии.
Нейтрофилы — это первые воспалительные клетки, попавшие в рану, и они прибывают в течение 24 часов после травмы. Они мигрируют в рану, отвечая на хемоаттрактанты, выделяемые тромбоцитами, а также на хемокины, представленные на поверхности эндотелиальных клеток [29]. Рецептор лейкоцитов PSGL-1 связывается с Р-селектином, экспрессируемым как на тромбоцитах, так и на эндотелиальных клетках [30]. Низкоаффинное связывание селектина с PSGL-1 на мембране нейтрофилов заставляет текущие нейтрофилы катиться и ненадолго связываться с эндотелиальными клетками.Во время катания нейтрофилы дополнительно активируются интерлейкином-8 (IL-8) и воспалительным белком макрофагов (MIP-1β), высвобождаемым из эндотелиальных клеток. Связывание рецептора нейтрофилов с хемоаттракантами активирует их интегрины β2, которые затем прочно прикрепляются к молекулам внутриклеточной адгезии эндотелиальных клеток (ICAM) -1 [31]. Нейтрофилы впоследствии экстравазируются через стенку сосуда в место раны, где они высвобождают протеолитические ферменты для переваривания инородных тел и уничтожения бактерий посредством фагоцитоза и образования супероксида и перекиси водорода [32].Нейтрофилы подвергаются апоптозу через 24–48 часов, если дезинфекция раны завершена. Они заменяются экстравазирующими моноцитами и макрофагами, доставленными к месту повреждения путем дальнейшего высвобождения MIP-1α и MIP-1β, моноцитарного хемоаттрактантного белка-1 (MCP-1) и хемотаксического цитокина, называемого RANTES, все из которых продуцируются посредством активированные эндотелиальные клетки [33].
Низкоаффинное связывание селектина с PSGL-1 на мембране нейтрофилов заставляет текущие нейтрофилы катиться и ненадолго связываться с эндотелиальными клетками.Во время катания нейтрофилы дополнительно активируются интерлейкином-8 (IL-8) и воспалительным белком макрофагов (MIP-1β), высвобождаемым из эндотелиальных клеток. Связывание рецептора нейтрофилов с хемоаттракантами активирует их интегрины β2, которые затем прочно прикрепляются к молекулам внутриклеточной адгезии эндотелиальных клеток (ICAM) -1 [31]. Нейтрофилы впоследствии экстравазируются через стенку сосуда в место раны, где они высвобождают протеолитические ферменты для переваривания инородных тел и уничтожения бактерий посредством фагоцитоза и образования супероксида и перекиси водорода [32].Нейтрофилы подвергаются апоптозу через 24–48 часов, если дезинфекция раны завершена. Они заменяются экстравазирующими моноцитами и макрофагами, доставленными к месту повреждения путем дальнейшего высвобождения MIP-1α и MIP-1β, моноцитарного хемоаттрактантного белка-1 (MCP-1) и хемотаксического цитокина, называемого RANTES, все из которых продуцируются посредством активированные эндотелиальные клетки [33].
1.3.2. Более поздние события (центральная нервная система [ЦНС] по сравнению с не-ЦНС)
В течение 2–3 дней после травмы воспаление продолжается, поскольку моноциты рекрутируются из крови и дифференцируются в макрофаги.Попадая в ткань, макрофаги выделяют дополнительные провоспалительные цитокины, такие как IL-1, TGF-β и фактор некроза опухоли-α (TNF-α) [26]. Макрофаги удаляют инородный мусор и могут оставаться в нем до нескольких месяцев, в зависимости от степени повреждения и количества инородного и некротического мусора, подлежащего удалению [26]. Считается, что макрофаги более важны, чем нейтрофилы, для успешного разрешения воспаления. В исследованиях, в которых нейтрофилы были истощены, заживление ран не нарушалось [34], но когда вместо них удалялись макрофаги, наблюдалось ограниченное удаление некротических остатков в месте раны, что приводило к измененному процессу заживления ран [32,25].Тучные клетки в ткани вносят свой вклад в воспалительную реакцию, высвобождая гистамин и серотонин, чтобы повысить проницаемость кровеносных сосудов и способствовать миграции макрофагов.
Как только гематоэнцефалический барьер восстанавливается путем наложения заплат на поврежденные кровеносные сосуды на более поздних стадиях гемостаза, воспалительные реакции ЦНС и тканей, не связанных с ЦНС, начинают расходиться (). В частности, селектины, экспрессируемые на эндотелиальных клетках сосудов, которые привлекают лейкоциты, менее распространены в ткани ЦНС, тем самым ограничивая рекрутирование [36–38].Следовательно, аксон и миелиновый мусор из поврежденных нейронов может занять месяцы, чтобы очистить ЦНС, в то время как в ПНС требуется всего несколько дней [39]. Снижение активности лейкоцитов и инфильтрация может рассматриваться как метод защиты мозга от чрезмерного воспалительного повреждения, при котором пониженные уровни рекрутирования лейкоцитов заменяются резидентными макрофагоподобными клетками микроглии.
Микроглия, единственные резидентные фагоцитозные клетки в ЦНС, обычно находятся в сильно разветвленном состоянии покоя или «разветвленном состоянии» и активируются только в том случае, если необходимо очистить от мусора или если нарушен гематоэнцефалический барьер [7]. После активации микроглия прекращает свои отростки и изменяет свое разветвленное состояние покоя на более компактную стержнеобразную форму [40,41]. Микроглия действует согласованно с макрофагами, которые проникли через гематоэнцефалический барьер, чтобы фагоцитировать дегенерирующие аксоны и миелин в месте повреждения [42]. На этом этапе микроглию можно отличить от макрофагов, полученных из крови, с помощью анализа проточной цитометрии in vitro по различиям в экспрессии CD45 (микроглия: низкий уровень CD45; макрофаги: высокий уровень CD45) [43,44].
После активации микроглия прекращает свои отростки и изменяет свое разветвленное состояние покоя на более компактную стержнеобразную форму [40,41]. Микроглия действует согласованно с макрофагами, которые проникли через гематоэнцефалический барьер, чтобы фагоцитировать дегенерирующие аксоны и миелин в месте повреждения [42]. На этом этапе микроглию можно отличить от макрофагов, полученных из крови, с помощью анализа проточной цитометрии in vitro по различиям в экспрессии CD45 (микроглия: низкий уровень CD45; макрофаги: высокий уровень CD45) [43,44].
Микроглия, как и макрофаги, экспрессирует поверхностные рецепторы, участвующие в врожденном иммунном ответе после воздействия различных патогенов или продуктов патогенов, таких как липополисахарид (ЛПС) [45]. Активированная микроглия также вносит вклад в аспекты адаптивного иммунитета, активируя главный комплекс гистосовместимости класса II и костимулирующие молекулы для презентации антигенов поступающим Т-клеткам [45,46]. Было показано, что микроглия стимулируется в течение 1 часа после первоначального повреждения ткани [10,47]. Количество активированной микроглии достигает пика через 3 дня после воспалительного поражения, хотя микроглия может оставаться активированной до 1 или 2 недель после травмы [48].
Количество активированной микроглии достигает пика через 3 дня после воспалительного поражения, хотя микроглия может оставаться активированной до 1 или 2 недель после травмы [48].
Микроглия в месте повреждения продуцирует множество провоспалительных и нейротоксических факторов, таких как MCP-1 [49], TNF-α [50], IL-1β [51], IL-6 [52], оксид азота [53] ] и супероксид [54]. Парадоксально, но также сообщалось, что микроглия увеличивает выживаемость нейронов за счет высвобождения противовоспалительных факторов, таких как IL-10 [55,56], и нейротрофических факторов, таких как фактор роста нервов (NGF), нейротрофический фактор головного мозга (BDNF). , и нейротрофин-3 (NT-3) [57,58] (обзор в [58]).
Астроциты, которые активируются в течение нескольких часов после травмы, являются вторым типом эффекторных клеток иммунного ответа, которые находятся в ЦНС. Астроциты также выполняют широкий спектр функций помимо воспаления, таких как обеспечение сигналов роста и механическая поддержка нейронов во время развития ЦНС, создание и поддержание гематоэнцефалического барьера и помощь в контроле окружающей химической среды посредством буферизации нейротрансмиттеров и ионов, высвобождаемых во время нейрональная передача сигналов [7]. Активированные астроциты идентифицируются во время воспаления по активации белка глиальной фибриллярной кислоты (GFAP), повышенной пролиферации и гипертрофии цитоплазмы и ядер. GFAP активируется уже через 1 час после травмы и, таким образом, является чувствительным ранним маркером реактивных астроцитов [59]. Однако активация GFAP не является единственным показателем активации астроцитов, поскольку микроглия может поглощать некротические фрагменты GFAP астроцитов и тем самым экспрессировать астроглиальный маркер [60]. Следовательно, клеточная гипертрофия является важным дополнительным индикатором астроглиоза.Было показано, что астроциты помогают опосредовать воспаление за счет высвобождения как провоспалительных, так и противовоспалительных факторов, таких как: (а) оксид азота, который способствует врожденному иммунному ответу, а также вызывает нейродегенерацию; (b) экспрессия молекул MHC как класса I, так и класса II, которые способствуют вторичному иммунному ответу; и (c) регулирование диаметра кровеносных сосудов путем высвобождения вазоконстрикторов [60].
Активированные астроциты идентифицируются во время воспаления по активации белка глиальной фибриллярной кислоты (GFAP), повышенной пролиферации и гипертрофии цитоплазмы и ядер. GFAP активируется уже через 1 час после травмы и, таким образом, является чувствительным ранним маркером реактивных астроцитов [59]. Однако активация GFAP не является единственным показателем активации астроцитов, поскольку микроглия может поглощать некротические фрагменты GFAP астроцитов и тем самым экспрессировать астроглиальный маркер [60]. Следовательно, клеточная гипертрофия является важным дополнительным индикатором астроглиоза.Было показано, что астроциты помогают опосредовать воспаление за счет высвобождения как провоспалительных, так и противовоспалительных факторов, таких как: (а) оксид азота, который способствует врожденному иммунному ответу, а также вызывает нейродегенерацию; (b) экспрессия молекул MHC как класса I, так и класса II, которые способствуют вторичному иммунному ответу; и (c) регулирование диаметра кровеносных сосудов путем высвобождения вазоконстрикторов [60].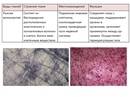
Через три дня после воспалительной реакции в ЦНС активированные астроциты наблюдаются на периферии повреждения, в то время как макрофаги и микроглия локализуются в центре раны [48] ().Микроглия, вероятно, играет главную роль в ответной реакции, поскольку гематоэнцефалический барьер в значительной степени исключает возможность повреждения макрофагов [61]. Тем не менее, макрофаги и лимфоциты из неневральной ткани также могут проникать в мозг на поздней стадии воспалительного процесса, чтобы еще больше усилить реакцию заживления ран организма за счет секреции различных цитокинов. Однако только лимфоциты, находящиеся в активированном состоянии, способны проходить через гематоэнцефалический барьер и попадать в ЦНС (см. Обзор [62]).
Рисунок 1.3
Заживление ран различных типов тканей. (A) В ЦНС микроглия и макрофаги мигрируют в очаг поражения, секретируя различные цитокины и факторы роста, а также удаляя некротическую ткань. Активированные астроциты образуют физический барьер вокруг области (подробнее .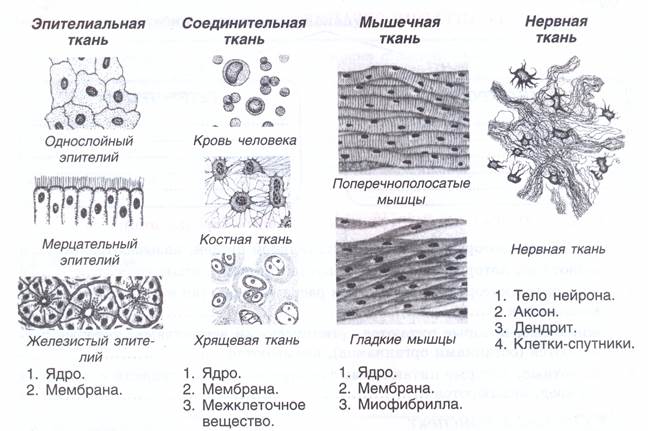 ..)
..)
Механизм экстравазации лимфоцитов во время воспаления изменен в ЦНС. Во-первых, лимфоциты не катятся по эпителию в ЦНС, а, во-вторых, активированные лимфоциты могут экстравазировать непосредственно через эндотелиальную клетку, оставляя соседние плотные контакты нетронутыми для сохранения гематоэнцефалического барьера [63].Этот процесс может занять несколько часов, тогда как диапедез по кровеносным сосудам в лимфатические узлы вне ЦНС может происходить в течение нескольких минут [64]. Уровень входа лимфоцитов варьируется в тканях ЦНС, с самым высоким входом в спинной мозг и самым низким входом в головной мозг [36]. Однако Т-клетки обнаруживают враждебную среду ЦНС и быстро умирают из-за апоптоза или покидают ЦНС через спинномозговую жидкость [36,65]. Снижение уровня лимфоцитов приводит к значительно более низким уровням иммунного надзора в ЦНС, чем в других тканях, и является причиной, по которой ЦНС часто называют участком «иммунологической привилегии» [36].Следствием воспалительной реакции в головном мозге является повреждение нейронов, о чем свидетельствует отсутствие нейрофиламентов вокруг поражения всего через несколько дней после инъекции провоспалительного агента [48].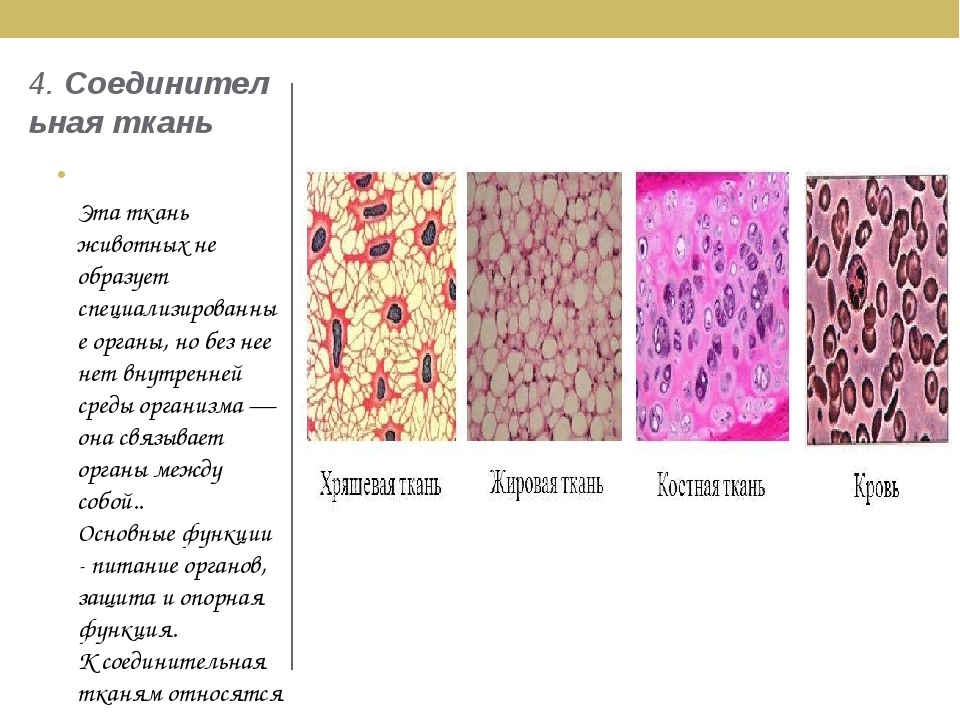
Воспалительная фаза разрешается смесью противовоспалительных цитокинов, хотя конкретные отдельные цитокины, которые задействованы, и механизмы их действия в значительной степени неизвестны [66]. TGF-β, секретируемый макрофагами, считается одним из основных аттенюаторов воспаления как в ЦНС, так и в тканях вне ЦНС [66].TGF-β участвует в стимулировании набора и пролиферации фибробластов, чтобы подтолкнуть реакцию заживления ран к фазе заживления. Также известно, что макрофаги продуцируют другие факторы заживления раны, такие как VEGF, чтобы инициировать ангиогенез в гипоксической тканевой среде [67].
1.4. РЕМОНТ (ОТ ДНЕЙ ЗА НЕДЕЛЮ)
Во время воспаления общая прочность ткани раны минимальна, поскольку ткани не восстанавливают свою нормальную функциональную прочность до тех пор, пока воспаление не перейдет в восстановление.Этот переход опосредуется макрофагами и их противовоспалительными цитокинами и обычно происходит через неделю после первоначального повреждения. Конечный результат процесса восстановления жизненно важен для заживления ран, потому что он устанавливает каркас, необходимый для поддержки и восстановления поврежденной ткани.
Конечный результат процесса восстановления жизненно важен для заживления ран, потому что он устанавливает каркас, необходимый для поддержки и восстановления поврежденной ткани.
Восстановление ткани характеризуется повышенной пролиферацией клеток, зачатком капилляров и синтезом внеклеточного матрикса (ЕСМ) для заполнения поврежденной ткани, которая была очищена во время воспаления.Материал матрикса изначально состоит из фибриногена и фибронектина [68]. После этого протеогликаны, большие макромолекулы с коровым белком и одной или несколькими ковалентно присоединенными молекулами гликозаминогликана, синтезируются клетками, чтобы составить основное вещество ЕСМ. Клетки, продуцирующие ЕСМ, которые продуцируют поддерживающий матрикс, необходимый для восстановления структурной целостности, включают фибробласты в соединительной ткани, хондроциты в хряще, остеобласты в кости и клетки Шванна в ПНС.
Репаративная реакция организма на повреждение различается в зависимости от мягкой коллагеновой кожной ткани, костей, ПНС и ЦНС (). Хотя полное восстановление поврежденной ткани является идеальным, этот ответ может постоянно происходить только в минерализованной ткани или кости, поскольку восстановление полной структурной целостности имеет решающее значение для функционального восстановления. В неминерализованной соединительной ткани, особенно в кожных ранах, глубина утраты ткани определяет реакцию восстановления. В то время как эпидермис может регенерироваться, глубокие тканевые раны, в которых теряется дерма, подвергаются вторичному заживлению, что требует избыточного производства ECM, что приводит к образованию фиброзной рубцовой ткани.В правильных условиях регенерация может происходить в ПНС, поскольку поврежденные аксоны могут вырасти через каркас шванновских клеток. В ЦНС утраченные аксоны не восстанавливаются, а глиальный рубец образуется реактивными астроцитами, действующими как физический и молекулярный барьер, препятствующий регенерации аксонов.
Хотя полное восстановление поврежденной ткани является идеальным, этот ответ может постоянно происходить только в минерализованной ткани или кости, поскольку восстановление полной структурной целостности имеет решающее значение для функционального восстановления. В неминерализованной соединительной ткани, особенно в кожных ранах, глубина утраты ткани определяет реакцию восстановления. В то время как эпидермис может регенерироваться, глубокие тканевые раны, в которых теряется дерма, подвергаются вторичному заживлению, что требует избыточного производства ECM, что приводит к образованию фиброзной рубцовой ткани.В правильных условиях регенерация может происходить в ПНС, поскольку поврежденные аксоны могут вырасти через каркас шванновских клеток. В ЦНС утраченные аксоны не восстанавливаются, а глиальный рубец образуется реактивными астроцитами, действующими как физический и молекулярный барьер, препятствующий регенерации аксонов.
1.4.1. Ткань, не связанная с ЦНС
Первым этапом восстановления ткани является стабилизация разрывов, образовавшихся в результате травмы.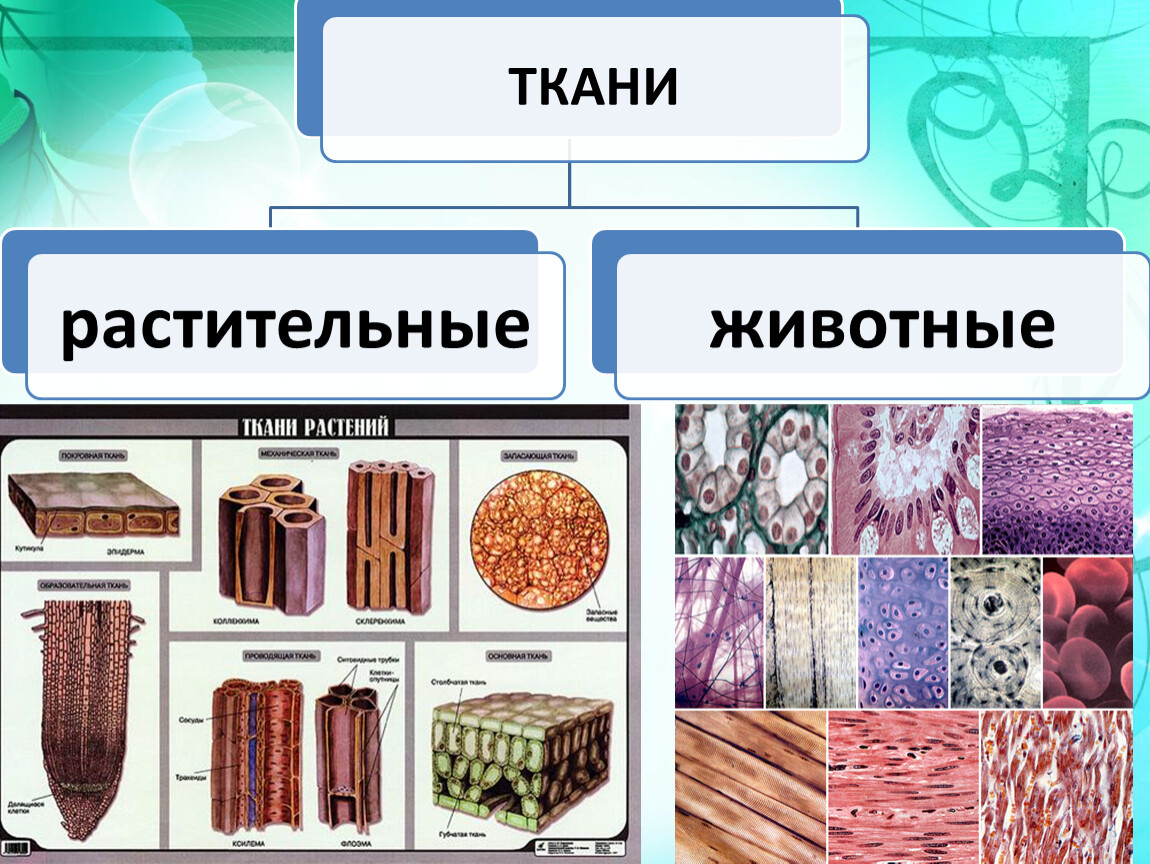 Традиционно существует две широких классификации исцеления. Ткань, которая почти не имеет зазора, разделяющего границы раны, подвергнется «первичному заживлению» от соприкасающихся краев ткани.Ткань, которая нестабильна с большим разрывом или прерывистым повреждением, подвергнется «вторичному заживлению», когда вырабатывается избыток внеклеточного матрикса, чтобы закрепить и заполнить поражение. ECM вторичного заживления, который впоследствии становится васкуляризованным, называется грануляционной тканью — термин, возникающий из-за его внешнего вида. Как правило, количество образовавшейся грануляционной ткани пропорционально возможному уровню рубцевания.
Традиционно существует две широких классификации исцеления. Ткань, которая почти не имеет зазора, разделяющего границы раны, подвергнется «первичному заживлению» от соприкасающихся краев ткани.Ткань, которая нестабильна с большим разрывом или прерывистым повреждением, подвергнется «вторичному заживлению», когда вырабатывается избыток внеклеточного матрикса, чтобы закрепить и заполнить поражение. ECM вторичного заживления, который впоследствии становится васкуляризованным, называется грануляционной тканью — термин, возникающий из-за его внешнего вида. Как правило, количество образовавшейся грануляционной ткани пропорционально возможному уровню рубцевания.
1.4.1.1. Первичная репарация (внеклеточный матрикс [ЕСМ] не продуцируется)
1.4.1.1.1. Восстановление частичной толщины кожи
Кожа состоит из двух отдельных слоев: эпидермиса и дермы. В ранах с частичной толщиной эпидермиса повреждается только эпидермис, оставляя неповрежденной базальную мембрану вместе с волосяными фолликулами, потовыми и сальными железами.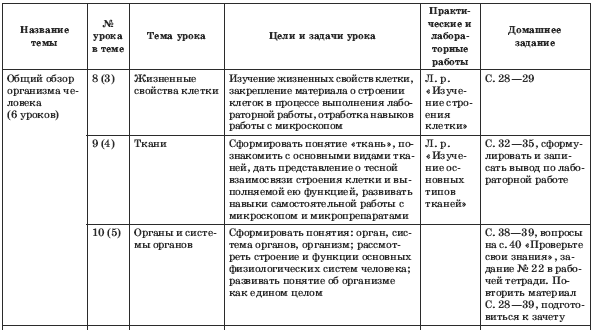 Поскольку необходимо заменить только поверхность эпидермиса, а клетки-предшественники эпителия остаются нетронутыми под раной, синтез и отложение коллагена не требуется. Чтобы исправить поверхностное повреждение, такое как «разрез бумаги», необходимо реэпителизацию этого участка только за счет миграции кератиноцитов из-под раны и по краю раны ().Степень реэпителизации зависит от количества утраченной ткани, глубины и ширины раны.
Поскольку необходимо заменить только поверхность эпидермиса, а клетки-предшественники эпителия остаются нетронутыми под раной, синтез и отложение коллагена не требуется. Чтобы исправить поверхностное повреждение, такое как «разрез бумаги», необходимо реэпителизацию этого участка только за счет миграции кератиноцитов из-под раны и по краю раны ().Степень реэпителизации зависит от количества утраченной ткани, глубины и ширины раны.
Реэпителизация начинается в течение 24-48 часов, когда неповрежденные кератиноциты отделяются от базальной пластинки и проходят через или под фибриновым сгустком, ползая внутрь и поперек раны — процесс, называемый ламеллопоидным ползанием (см. Обзор [69]). Миграция начинается, когда кератиноциты на краю раны активируют свою продукцию матриксных металлопротеиназ (ММП), высвобождая клетки из их привязок в базальную пластинку [70].Кератиноциты, покоящиеся на базальной пластинке, мигрируют через участок раны со скоростью 1-2 мм / день [71], прикрепляясь к фибронектину и витронектину, содержащимся в сгустке, путем усиления экспрессии их α 5 β 1 и α V β 6 интегринов [69,72]. Двигаясь через плотный фибриновый сгусток, кератиноциты растворяют плотный фибриновый матрикс за счет активации нескольких протеаз, таких как тканевый активатор плазминогена (tPA) и активатор плазминогена урокиназного типа (uPA), активируя фибринолитический фермент плазмин [73].
Двигаясь через плотный фибриновый сгусток, кератиноциты растворяют плотный фибриновый матрикс за счет активации нескольких протеаз, таких как тканевый активатор плазминогена (tPA) и активатор плазминогена урокиназного типа (uPA), активируя фибринолитический фермент плазмин [73].
Передвижение кератиноцитов происходит за счет сокращения актиномиозиновых филаментов цитоскелета [69]. Из-за изменений в экспрессии интегрина и сборки актиномиозиновых филаментов, когда кератиноциты прикрепляются к временной матрице сгустка, наблюдается задержка на несколько часов, прежде чем наблюдается миграция базальных кератиноцитов [74]. Как только начинается миграция, кератиноциты перемещаются один за другим по месту раны, пока рана не будет покрыта полным слоем кератиноцитов [72]. После завершения миграции клетки за краем раны подвергаются пролиферативному взрыву, чтобы заменить кератиноциты, потерянные в результате повреждения, а также формируют дополнительные слои кератиноцитов над базальным слоем [75]. Считается, что эпидермальный фактор роста (EGF), фактор роста кератиноцитов (KGF) и трансформирующий фактор роста (TGF-α) могут управлять пролиферацией клеток и закрытием ран [69]. Миграция и пролиферация продолжаются до тех пор, пока кератиноциты не получат стоп-сигнал, вероятно, из-за контактного ингибирования [28]. В это время экспрессия MMP прерывается, и образуется новая базальная мембрана, посредством чего устанавливаются новые адгезии клеточного матрикса [28].
Считается, что эпидермальный фактор роста (EGF), фактор роста кератиноцитов (KGF) и трансформирующий фактор роста (TGF-α) могут управлять пролиферацией клеток и закрытием ран [69]. Миграция и пролиферация продолжаются до тех пор, пока кератиноциты не получат стоп-сигнал, вероятно, из-за контактного ингибирования [28]. В это время экспрессия MMP прерывается, и образуется новая базальная мембрана, посредством чего устанавливаются новые адгезии клеточного матрикса [28].
1.4.1.1.2. Стабилизированное восстановление костей
Когда движение в месте перелома предотвращается и концы сломанных костей жестко удерживаются на месте сразу после травмы, происходит восстановление первичной минерализованной ткани.При первичном восстановлении кости нет необходимости в структурной поддержке, богатой ECM, потому что ткань уже стабилизирована. Могут происходить два типа первичного заживления минерализованной ткани: заживление зазора, при котором имеется разрыв менее 0,5 мм, разделяющий стабилизированные костные фрагменты, и заживление контакта, при котором сломанные концы удерживаются в прямом соприкосновении [76].
При заживлении промежутков заживление начинается с заполнения раны кровеносными сосудами и рыхлой соединительной тканью. Через 2 недели плюрипотентные мезенхимальные клетки, полученные из костного мозга, достигают места повреждения и дифференцируются в клетки, продуцирующие кость, которые называются остеобластами.Остеобласты заполняют любые пробелы в ткани, секретируя слои неминерализованного костного матрикса, называемого остеоидом, со скоростью от 1 до 2 мкм в день, который становится минерализованным примерно через 10 дней [77]. Новая кость, сформированная на этом этапе, расположена перпендикулярно длинной оси кости (). По мере того как остеоид становится минерализованным, некоторые остеобласты остаются в матрице и захватываются по мере минерализации ткани. Эти остеобласты становятся остеоцитами и отвечают за механотрансдукцию в ткани.Остеобласты, вероятно, активируются белками, принадлежащими к семейству TGF-β, такими как костные морфогенные белки (BMP) [78]. Новая кость заполняет разрыв, но не полностью соединяет концы перелома. На этом этапе граница между новой и исходной костью является самым слабым звеном в соединении [79]. Эта новообразованная кость действует как каркас для будущего ремоделирования остеокластами и остеобластами.
На этом этапе граница между новой и исходной костью является самым слабым звеном в соединении [79]. Эта новообразованная кость действует как каркас для будущего ремоделирования остеокластами и остеобластами.
Рисунок 1.4
Сравнение первичного и вторичного восстановления минерализованной ткани. (A) Когда возникает небольшая щель перелома и кость остается стабилизированной, остеобласты откладывают кость перпендикулярно длинной оси кости, заполняя щель и служа каркасом для будущего ремоделирования.(подробнее …)
Контактное восстановление происходит при прямом контакте между концами кости. Это позволяет сформированной пластинчатой кости формироваться прямо поперек линии перелома. Однако, прежде чем новую кость можно будет отложить для заживления раны, необходимо удалить некротизированную кость. Остеокласты, большие многоядерные клетки, которые происходят из гемопоэтических клеток-предшественников в костном мозге и тесно связаны с макрофагами, проходят через линию перелома и удаляют старую кость и некротическую ткань за счет высвобождения протеаз и лизосомальных ферментов (). Они прикрепляются к матрице ткани через интегрины β3, создавая волнистую клеточную границу между клеткой и поверхностью кости [80,81]. Ионы водорода перекачиваются к мембране остеокласта, чтобы создать кислую микросреду для переваривания минерального компонента кости. Органический матрикс кости разрушается лизосомальными протеазами и затем подвергается эндоцитозу остеокластами [82]. Скорость разрушения кости остеокластами зависит от ориентации ткани. Остеокласты способны разрезать кость со скоростью от 20 до 40 мкм в день параллельно и от 5 до 10 мкм в день перпендикулярно длинной оси кости [83].Внутри этих «режущих конусов» образуются капилляры, которые снабжают восстанавливаемую ткань питательными веществами [84]. За режущим конусом ряды остеобластов выстилают резорбтивный канал, откладывая слои остеоида. Эта область называется «закрывающимся конусом», где остеобласты пытаются заменить примерно столько кости, сколько было удалено [83] (). Слои остеоида изначально состоят из коллагена I и IV типов, который постепенно минерализуется за счет отложения кристаллов гидроксиапатита с образованием пластинчатой кости.
Они прикрепляются к матрице ткани через интегрины β3, создавая волнистую клеточную границу между клеткой и поверхностью кости [80,81]. Ионы водорода перекачиваются к мембране остеокласта, чтобы создать кислую микросреду для переваривания минерального компонента кости. Органический матрикс кости разрушается лизосомальными протеазами и затем подвергается эндоцитозу остеокластами [82]. Скорость разрушения кости остеокластами зависит от ориентации ткани. Остеокласты способны разрезать кость со скоростью от 20 до 40 мкм в день параллельно и от 5 до 10 мкм в день перпендикулярно длинной оси кости [83].Внутри этих «режущих конусов» образуются капилляры, которые снабжают восстанавливаемую ткань питательными веществами [84]. За режущим конусом ряды остеобластов выстилают резорбтивный канал, откладывая слои остеоида. Эта область называется «закрывающимся конусом», где остеобласты пытаются заменить примерно столько кости, сколько было удалено [83] (). Слои остеоида изначально состоят из коллагена I и IV типов, который постепенно минерализуется за счет отложения кристаллов гидроксиапатита с образованием пластинчатой кости. Кость, сформированная этим методом, при котором на промежуточном этапе не образуется хрящ, называется внутримембранозным окостенением.
Кость, сформированная этим методом, при котором на промежуточном этапе не образуется хрящ, называется внутримембранозным окостенением.
1.4.1.1.3. Восстановление периферической нервной системы (ПНС)
Нерв состоит из набора нескольких нейронов и поддерживающих их клеток, называемых шванновскими клетками (). Шванновские клетки действуют как поддерживающие клетки, окружающие процессы передачи сигналов нейронов, называемые аксонами, и изолируют их от ECM, производя миелин, что увеличивает скорость распространения сигнала.Каждый аксон и его шванновские клетки окружены матрицей соединительной ткани, называемой эндоневрием, впоследствии сгруппированы в пучки и ограничены другой тонкой матрицей соединительной ткани, называемой периневрием. Некоторые из этих пучков вместе с сосудистой сетью составляют нерв с внешней оболочкой, называемой эпиневрием [85,86].
Рисунок 1.5
Анатомия поперечного сечения периферического нерва. На вставке слева показано немиелинизированное волокно.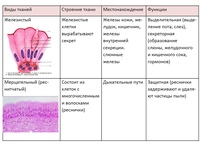 На вставке внизу показано миелинизированное волокно. (© 2000 Американская академия хирургов-ортопедов.Перепечатано из J. Am. Акад. Ортоп. Surg. , 8 (4), 243, 2000. (подробнее …)
На вставке внизу показано миелинизированное волокно. (© 2000 Американская академия хирургов-ортопедов.Перепечатано из J. Am. Акад. Ортоп. Surg. , 8 (4), 243, 2000. (подробнее …)
Возможность ремонта в ПНС зависит от степени травмы. Наиболее серьезная травма, которую можно исправить, классифицируется как травма третьей степени, когда аксон и эндоневрий повреждаются или разрываются, в то время как периневрий и эпиневрий остаются нетронутыми [87]. При более травматических повреждениях, когда нарушаются не только аксоны, но и весь эпиневрий или периневрий, гемато-нервный барьер разрушается, что требует обширного фиброзного ВКМ для физического восстановления поврежденной ткани.Этот плотный ECM предотвращает регенерацию, так как разрезанный аксон неспособен найти свою иннервируемую мишень [87].
Независимо от степени повреждения, если восстановление в ПНС должно быть успешным, тело клетки нейрона должно сначала пережить повреждение его расширяющегося аксона [39]. Тело клетки нейрона необходимо для метаболизма в аксональных процессах нейрона; следовательно, любой расширяющийся аксон, оторванный от его тела, быстро дегенерирует вниз по течению от того места, где произошло повреждение.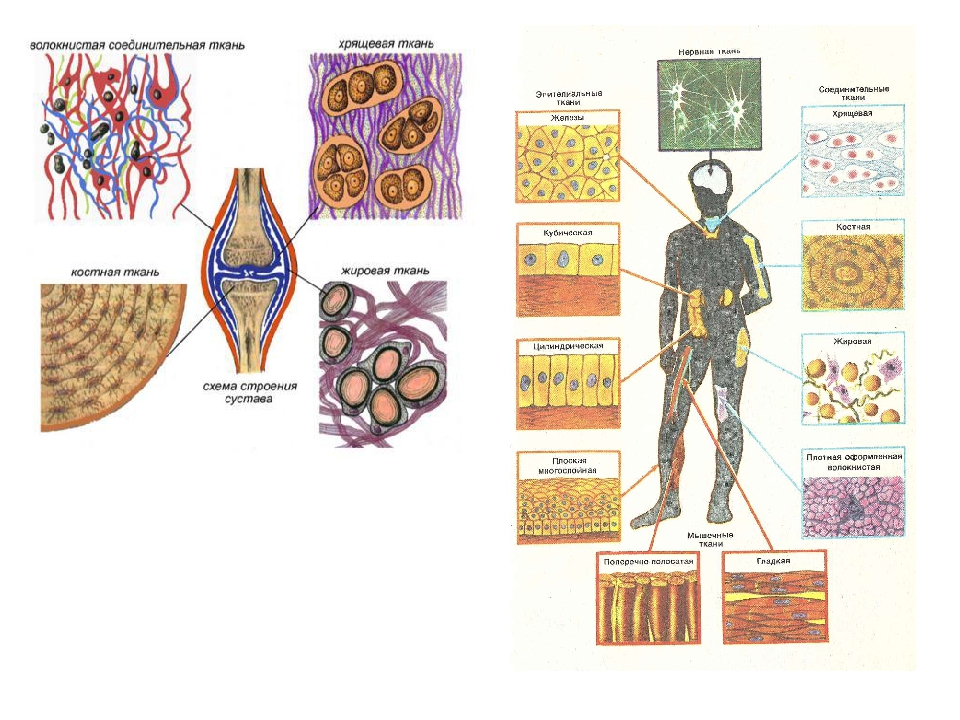 В течение нескольких минут или часов после повреждения нейрона и окружающей его среды дегенерация начинается, когда шванновские клетки, окружающие аксон, перестают вырабатывать миелиновые белки.Внутри аксона ферментные протеазы активируются в ответ на приток кальция, который вызывает фрагментацию аксона и заставляет окружающий миелин формировать частицы в форме капли, процесс, называемый дегенерацией Валлера [39]. К 3–4 дням проведение импульса становится невозможным, так как расширяющийся аксон разрушается. Обломки аксонов и миелина очищаются на более поздних стадиях воспаления и в начале фазы восстановления за счет притока макрофагов в течение нескольких недель, в отличие от ЦНС, которое продолжается в течение нескольких месяцев [61].
В течение нескольких минут или часов после повреждения нейрона и окружающей его среды дегенерация начинается, когда шванновские клетки, окружающие аксон, перестают вырабатывать миелиновые белки.Внутри аксона ферментные протеазы активируются в ответ на приток кальция, который вызывает фрагментацию аксона и заставляет окружающий миелин формировать частицы в форме капли, процесс, называемый дегенерацией Валлера [39]. К 3–4 дням проведение импульса становится невозможным, так как расширяющийся аксон разрушается. Обломки аксонов и миелина очищаются на более поздних стадиях воспаления и в начале фазы восстановления за счет притока макрофагов в течение нескольких недель, в отличие от ЦНС, которое продолжается в течение нескольких месяцев [61].
При травмах, при которых нейрон выживает и наибольшая степень повреждения расширяющегося нерва — раздавливание или разрыв аксона и эндоневрия, восстановление начинается по мере пролиферации шванновских клеток проксимальнее повреждения (). Шванновские клетки помогают удалить дегенерированный аксональный и миелиновый остатки и заполнить область, ранее занимаемую расширяющимся аксоном и миелиновой оболочкой () [88]. Пролиферирующие шванновские клетки образуют взаимосвязанные клеточные трубки, которые действуют как каналы для регенерации аксонов [42].Внутри канала шванновские клетки увеличивают производство факторов роста, таких как NGF и мозговой фактор роста (BDGF), одновременно синтезируя белки ECM, такие как ламинин и фибронектин [39]. Аксоны начинают регенерировать в течение 3–24 часов, прорастая множественные новые аксоны малого диаметра в месте повреждения, называемом «конусом роста», и пытаются прорасти через канал шванновских клеток () [39]. Регенерирующие отростки аксонов перемещаются через очаг поражения на 0,5–5,0 мм в день, стимулируемые контактом и хемотаксическим контролем, обеспечиваемым шванновскими клетками [89, 90].Каждый аксональный отросток содержит филоподии, которые связываются через интегрины β1 с молекулами ЕСМ, такими как ламинин и фибронектин, которые продуцируются шванновскими клетками [91]. Вновь регенерированные аксоны изначально немиелинизированы, но являются митогенными, заставляя соседние шванновские клетки пролиферировать дальше и производить миелин вокруг аксона по мере его регенерации [39].
Пролиферирующие шванновские клетки образуют взаимосвязанные клеточные трубки, которые действуют как каналы для регенерации аксонов [42].Внутри канала шванновские клетки увеличивают производство факторов роста, таких как NGF и мозговой фактор роста (BDGF), одновременно синтезируя белки ECM, такие как ламинин и фибронектин [39]. Аксоны начинают регенерировать в течение 3–24 часов, прорастая множественные новые аксоны малого диаметра в месте повреждения, называемом «конусом роста», и пытаются прорасти через канал шванновских клеток () [39]. Регенерирующие отростки аксонов перемещаются через очаг поражения на 0,5–5,0 мм в день, стимулируемые контактом и хемотаксическим контролем, обеспечиваемым шванновскими клетками [89, 90].Каждый аксональный отросток содержит филоподии, которые связываются через интегрины β1 с молекулами ЕСМ, такими как ламинин и фибронектин, которые продуцируются шванновскими клетками [91]. Вновь регенерированные аксоны изначально немиелинизированы, но являются митогенными, заставляя соседние шванновские клетки пролиферировать дальше и производить миелин вокруг аксона по мере его регенерации [39]. Отростки аксонов продолжают расти в попытке реиннервировать ткань, которая была деиннервирована дегенерацией аксонов, вызванной повреждением.
Отростки аксонов продолжают расти в попытке реиннервировать ткань, которая была деиннервирована дегенерацией аксонов, вызванной повреждением.
1,4.1.2. Вторичный ремонт (производство ЭБУ)
1.4.1.2.2. Восстановление полной толщины кожной ткани
Если базальная мембрана повреждена в ране на всю толщину кожи и значительная часть дермы потеряна, рана не может зажить только за счет реэпителизации. Дермальные фибробласты, продуцирующие ВКМ, прилегающие к месту раны, активируются и пролиферируют, мигрируя в гематому раны в течение 3-4 дней [69].
При вторичной репарации, примерно в тот же период времени, когда кератиноциты пытаются реэпителизировать рану (как ранее было показано в первичной репарации кожи), фибробласты мигрируют через временный матрикс за счет сокращения своего актиномиозинового цитоскелета.Мигрирующие фибробласты первоначально синтезируют фибронектин [92], но в ответ на TGF-β1, высвобождаемый макрофагами, фибробласты переключаются на более фибротический фенотип, синтезируя коллагеновый матрикс, который начинает обеспечивать структурную поддержку раны [93]. Кератиноциты используют этот вновь образованный ECM для миграции и эпителизации места повреждения (). ЕСМ действует также как канал, по которому формируются новые капилляры, когда эндотелиальные клетки мигрируют в участок раны, отвечая на факторы роста, такие как фактор роста фибробластов-2 (FGF-2) и VEGF [94].Неоваскуляризация доставляет питательные вещества к мигрирующим фибробластам в месте повреждения, придавая замещающей ткани розовый гранулярный вид — отсюда и название «грануляционная ткань». Новые кровеносные сосуды образуются при изменении баланса между относительными количествами молекул, которые индуцируют, по сравнению с молекулами, которые ингибируют васкуляризацию. Проангиогенные факторы роста, такие как VEGF, секретируются преимущественно кератиноцитами на краю раны, тогда как FGF-2 и TNF-α высвобождаются из поврежденных эндотелиальных клеток и макрофагов [69].Гиалуронан, олигосахаридный компонент ЕСМ, также способствует ангиогенезу и помогает в репарации [95]. Реваскуляризация раневого участка имеет решающее значение, поскольку ангиогенная недостаточность может привести к хроническим ранам, таким как венозные язвы, которые не могут заживать.
Кератиноциты используют этот вновь образованный ECM для миграции и эпителизации места повреждения (). ЕСМ действует также как канал, по которому формируются новые капилляры, когда эндотелиальные клетки мигрируют в участок раны, отвечая на факторы роста, такие как фактор роста фибробластов-2 (FGF-2) и VEGF [94].Неоваскуляризация доставляет питательные вещества к мигрирующим фибробластам в месте повреждения, придавая замещающей ткани розовый гранулярный вид — отсюда и название «грануляционная ткань». Новые кровеносные сосуды образуются при изменении баланса между относительными количествами молекул, которые индуцируют, по сравнению с молекулами, которые ингибируют васкуляризацию. Проангиогенные факторы роста, такие как VEGF, секретируются преимущественно кератиноцитами на краю раны, тогда как FGF-2 и TNF-α высвобождаются из поврежденных эндотелиальных клеток и макрофагов [69].Гиалуронан, олигосахаридный компонент ЕСМ, также способствует ангиогенезу и помогает в репарации [95]. Реваскуляризация раневого участка имеет решающее значение, поскольку ангиогенная недостаточность может привести к хроническим ранам, таким как венозные язвы, которые не могут заживать.
1.4.1.2.2. Восстановление кости
При восстановлении кости вторичное заживление более часто наблюдается, чем заживление первичного перелома кости, потому что большинство костей не имеют жесткой поддержки после травмы [96]. Когда травма приводит к разрушению внешнего сосудистого покрытия кости (надкостницы), а также окружающих мягких тканей, поврежденная ткань обычно остается без поддержки.Нестабилизированная минерализованная ткань подвергнется вторичной репарации, где первым шагом будет создание мостовидного протеза, богатого внеклеточным матриксом, для поддержки перелома ().
Наиболее компактные поверхности костей, составляющие внешний слой минерализованной ткани, покрыты выстилкой из остеобластов, которые становятся активными и производят небольшое внутримембранозное окостенение уже через 24 часа после травмы по обе стороны от перелома () [96 ]. Однако это ограниченное раннее формирование кости обеспечивает небольшую стабильность [96].Место раны не начинает восстанавливать механическую прочность в течение 3-4 дней после травмы, когда фибробласты и недифференцированные мезенхимальные клетки попадают в надкостницу через кровообращение [97].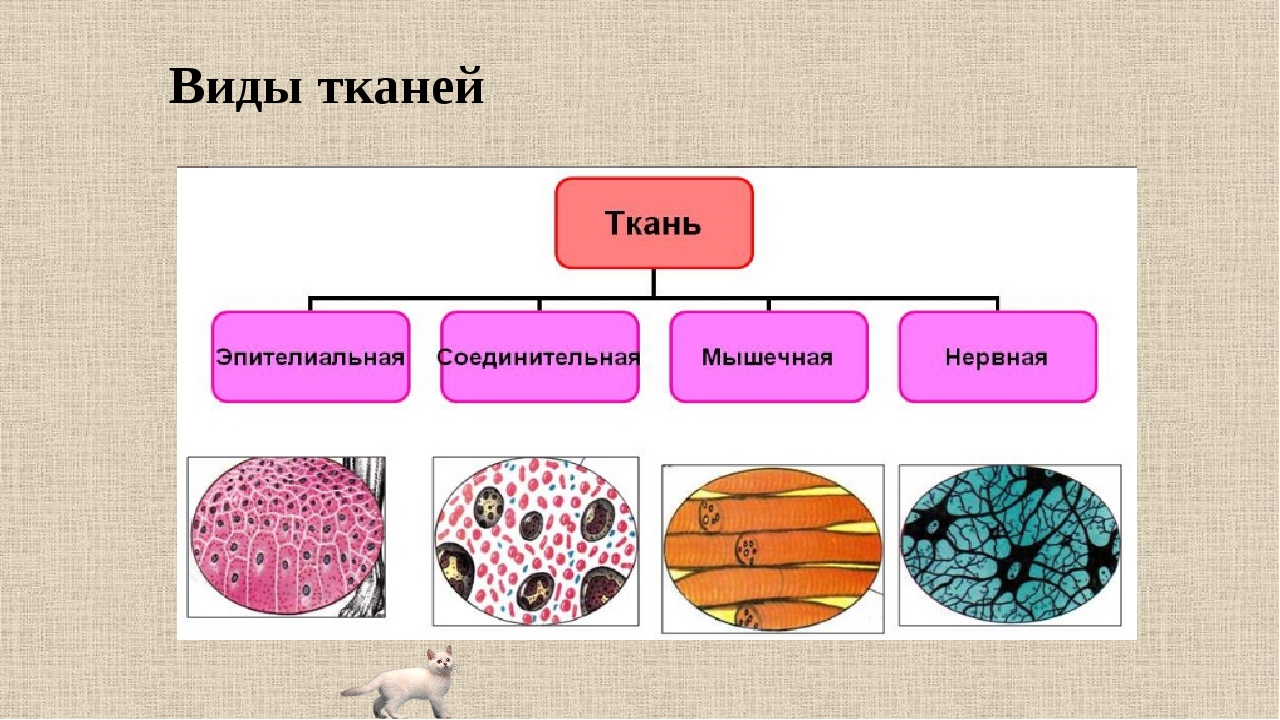 При вторичном заживлении остеопрогениторы, поступающие из сосудистой сети, дифференцируются в хондроциты, тогда как при первичном заживлении они дифференцируются в остеобласты. Хондроциты и фибробласты объединяются для образования коллагеновой и фиброзной ткани, которая формируется вокруг трещины снаружи надкостницы и внутри костного мозга, образуя внутреннюю шину () [98].Эта богатая коллагеном ткань называется мягкой костной мозолью и может быть классифицирована как тип грануляционной ткани, которая имеет решающее значение для обеспечения васкуляризации и структурной поддержки перелома. Максимум мозоли обычно наблюдается через неделю после травмы (7-е сутки) [99]. Как правило, количество мягкой костной мозоли зависит от относительной стабильности фрагментов перелома. Чем сильнее движение в месте перелома, тем больше костной мозоли требуется для предотвращения этого движения [100]. Факторы роста, такие как PDGF, TGF-β и FGF, высвобождаемые из тромбоцитов сразу после повреждения, являются возможными инициаторами образования каллуса [100].
При вторичном заживлении остеопрогениторы, поступающие из сосудистой сети, дифференцируются в хондроциты, тогда как при первичном заживлении они дифференцируются в остеобласты. Хондроциты и фибробласты объединяются для образования коллагеновой и фиброзной ткани, которая формируется вокруг трещины снаружи надкостницы и внутри костного мозга, образуя внутреннюю шину () [98].Эта богатая коллагеном ткань называется мягкой костной мозолью и может быть классифицирована как тип грануляционной ткани, которая имеет решающее значение для обеспечения васкуляризации и структурной поддержки перелома. Максимум мозоли обычно наблюдается через неделю после травмы (7-е сутки) [99]. Как правило, количество мягкой костной мозоли зависит от относительной стабильности фрагментов перелома. Чем сильнее движение в месте перелома, тем больше костной мозоли требуется для предотвращения этого движения [100]. Факторы роста, такие как PDGF, TGF-β и FGF, высвобождаемые из тромбоцитов сразу после повреждения, являются возможными инициаторами образования каллуса [100].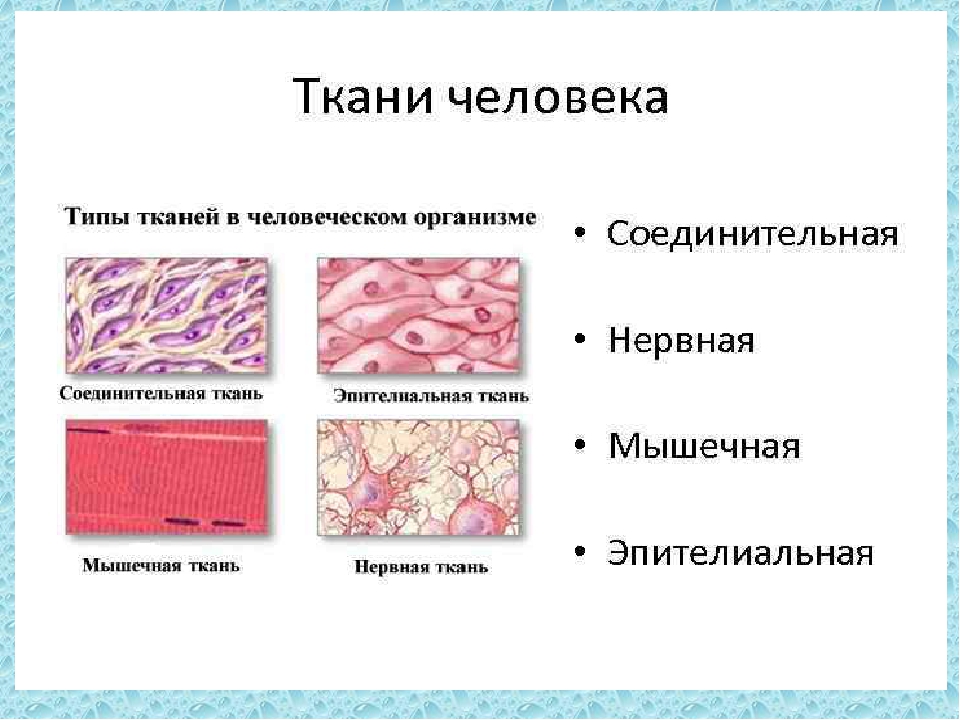
Рис. 1.6
Гистологический анализ вторичного заживления перелома кости, показывающий прогрессирование восстановления на 1, 3, 14, 21 и 28 дни. Перелом костей кажется более плотным, чем окружающие ткани. На 7-й день вокруг поврежденной кости образуется обширная мягкая мозоль. (подробнее …)
Через 2 недели коллагеновая мягкая мозоль постепенно минерализуется, образуя твердую мозоль, повышая стабильность места перелома (, 14 день) [99]. Минерализация коллагена с образованием тканой кости отличается от минерализации остеоида с образованием пластинчатой кости.При минерализации коллагена хондроциты прекращают производство коллагена, удлиняются и высвобождают протеазы [101]. Гликозаминогликаны в коллагеновой матрице ингибируют минерализацию и должны удаляться протеогликаназами хондроцитов, чтобы произошла минерализация [101]. По мере разрушения хрящевого матрикса хондроциты дифференцируются и секретируют ангиогенные факторы, такие как VEGF, чтобы вызвать прорастание капилляров из соседних тканей [102]. Остеобласты, выстилающие поверхность кости, затем секретируют не содержащие коллаген органические матрицы, такие как остеонектин и остеопонтин, обеспечивая места зародышеобразования для инициации минерализации нанокристаллического фосфата кальция.Кость, образованная этим методом, когда коллаген минерализуется с образованием кости, называется эндохондральным окостенением.
Остеобласты, выстилающие поверхность кости, затем секретируют не содержащие коллаген органические матрицы, такие как остеонектин и остеопонтин, обеспечивая места зародышеобразования для инициации минерализации нанокристаллического фосфата кальция.Кость, образованная этим методом, когда коллаген минерализуется с образованием кости, называется эндохондральным окостенением.
К третьей неделе большая часть хряща становится костью, и достигается сращение (21 день) [99]. В этот момент заживающая кость обычно способна выдерживать нагрузки. Однако даже после стабилизации вновь сформированная кость все еще слабее, чем нормальная неповрежденная кость. Только после фазы ремоделирования, в которой тканая кость становится более компактной ламинарной костью, ткань достигает полной прочности [98].
1.4.2. Восстановление ЦНС (глиоз)
Восстановление раны в ЦНС не сопровождается регенерацией нейронов. В отличие от реакции, наблюдаемой в ПНС, где дегенерированные аксоны могут регенерироваться, поврежденные аксоны ЦНС сначала прорастают, но регенерация затрудняется, поскольку конусы роста разрушаются в течение дня. Восстановление ткани ЦНС начинается через несколько часов после травмы, когда активируются астроциты за пределами ядра поражения и окружающей области. Этот выраженный глиальный ответ, обычно называемый глиозом, состоит из многослойного слоя активированных астроцитов, которые образуют границу вокруг участков повреждения ткани ().Подобно ответу, наблюдаемому при восстановлении неминерализованной ткани, астроциты изменяют свою экспрессию интегрина, мигрируя к поражению, а также секретируют ММП для деградации ECM. Экспрессия ММР астроцитами и нейронами через 1-2 недели после повреждения также может способствовать восстановлению за счет стимуляции продукции VEGF для инициации ангиогенеза [103]. Как только астроциты достигают места повреждения, их отростки охватывают поражение и плотно переплетаются, придавая глиальному барьеру крайне беспорядочный вид, образуя так называемый глиальный рубец.Общая величина глиальной активации примерно коррелирует с степенью нарушения гематоэнцефалического барьера и повреждения тканей [104].
Восстановление ткани ЦНС начинается через несколько часов после травмы, когда активируются астроциты за пределами ядра поражения и окружающей области. Этот выраженный глиальный ответ, обычно называемый глиозом, состоит из многослойного слоя активированных астроцитов, которые образуют границу вокруг участков повреждения ткани ().Подобно ответу, наблюдаемому при восстановлении неминерализованной ткани, астроциты изменяют свою экспрессию интегрина, мигрируя к поражению, а также секретируют ММП для деградации ECM. Экспрессия ММР астроцитами и нейронами через 1-2 недели после повреждения также может способствовать восстановлению за счет стимуляции продукции VEGF для инициации ангиогенеза [103]. Как только астроциты достигают места повреждения, их отростки охватывают поражение и плотно переплетаются, придавая глиальному барьеру крайне беспорядочный вид, образуя так называемый глиальный рубец.Общая величина глиальной активации примерно коррелирует с степенью нарушения гематоэнцефалического барьера и повреждения тканей [104].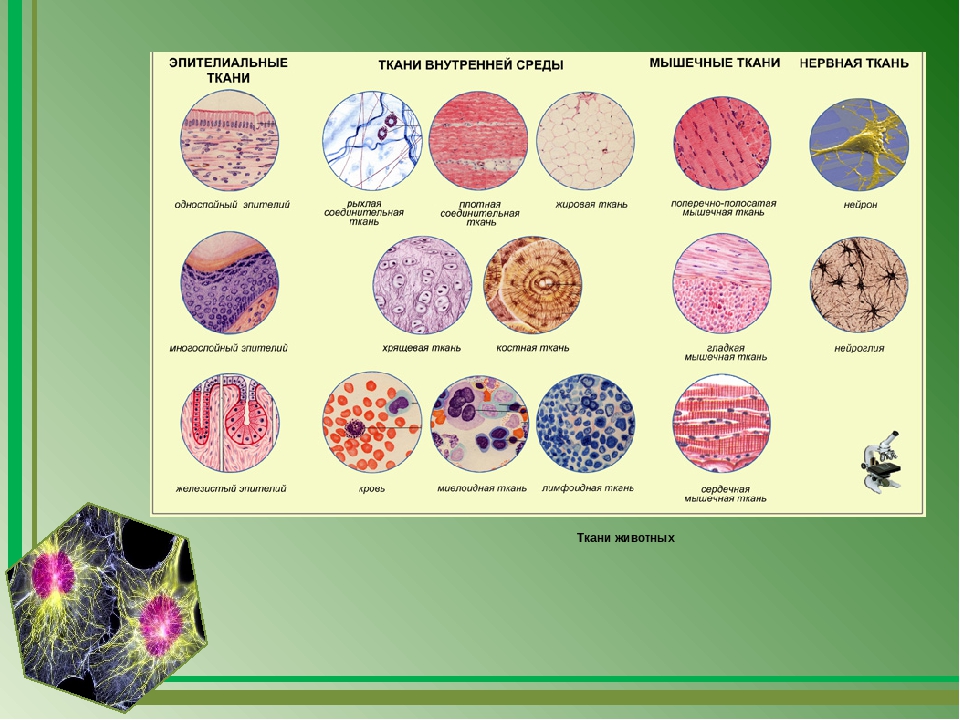
Внутри поражения и в окружении реактивных астроцитов микроглия и макрофаги продолжают попытки удалить потенциальные патогены и переваривать фибриновый сгусток [105] (). Из-за сильной воспалительной реакции, вызываемой микроглией и макрофагами, через 3 дня после травмы в месте раны обычно нет нейрофиламентов [48,105]. Без жизнеспособности нейронов ЦНС теряет свою функциональность в месте повреждения.Со структурной точки зрения, в то время как более мелкие поражения в ЦНС могут быть заполнены реактивными астроцитами [48], гипертрофия и пролиферация глии не могут компенсировать большую потерю ткани. Эти большие раны остаются полостями с реактивными астроцитами, образующими плотный барьер вокруг поражения. Поскольку поражение не заполнено клетками или внеклеточным матриксом, хирурги могут визуализировать прошлые травмы головного мозга во время визуализации из-за отсутствия структуры ткани () [106]. В случаях, когда проникает внешняя менингеальная поверхность головного мозга, поражение заполняется фиброзной тканью, поскольку менингеальные фибробласты, выстилающие внешнюю часть мозга, могут мигрировать в очаг [9,105,107].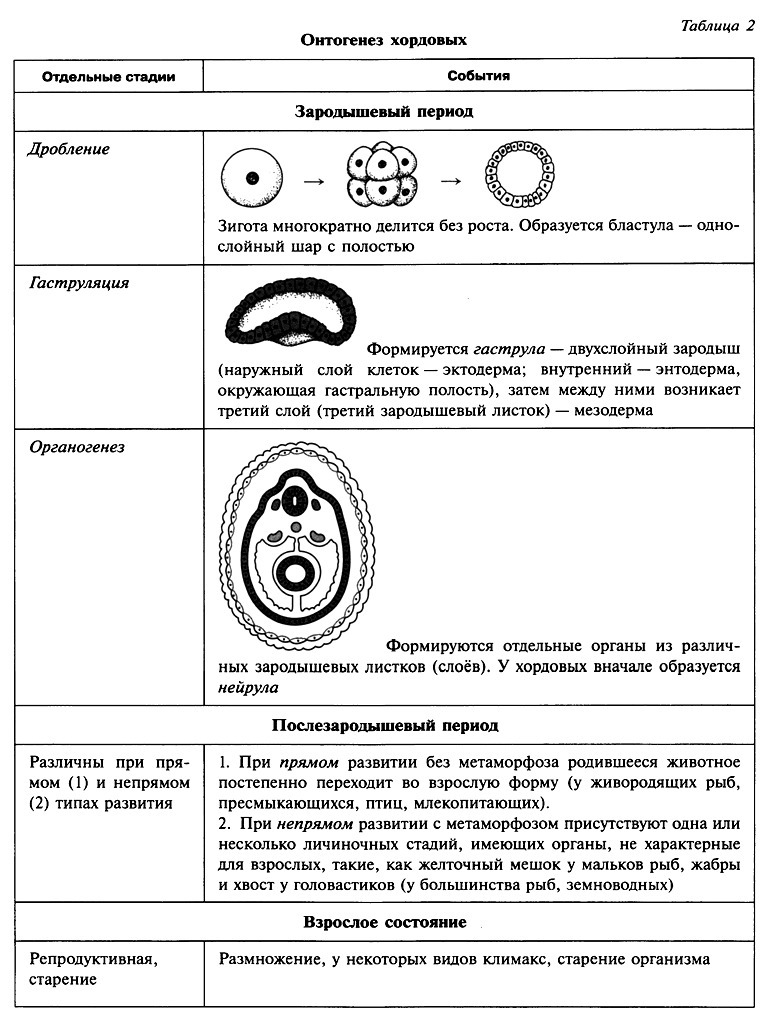 Менингеальные фибробласты, как и те, которые восстанавливают соединительную ткань, способны продуцировать коллаген (типы I, III, IV) и белки ECM (ламинин, фибронектин) для заполнения места раны [108,109]. Другие неглиальные клетки, такие как эндотелиальные клетки сосудов и мезенхимальные клетки, присутствуют в глиальном рубце. Эндотелиальные клетки пытаются сформировать новые кровеносные сосуды, тогда как мезенхимальные клетки откладывают базальную пластинку, которая, как известно, ингибирует повторный рост аксонов [60, 110].
Менингеальные фибробласты, как и те, которые восстанавливают соединительную ткань, способны продуцировать коллаген (типы I, III, IV) и белки ECM (ламинин, фибронектин) для заполнения места раны [108,109]. Другие неглиальные клетки, такие как эндотелиальные клетки сосудов и мезенхимальные клетки, присутствуют в глиальном рубце. Эндотелиальные клетки пытаются сформировать новые кровеносные сосуды, тогда как мезенхимальные клетки откладывают базальную пластинку, которая, как известно, ингибирует повторный рост аксонов [60, 110].
Рисунок 1.7
Ремоделирование ран различных типов тканей.(A) В месте повреждения фибриновый сгусток и некротическая ткань удаляются микроглией и макрофагами. В отличие от неневральной ткани утраченная ткань не восстанавливается, оставляя поражение с спинномозговой жидкостью — (подробнее …)
1.4.2.1. Назначение глиального рубца
Исследования повреждений спинного мозга, когда астроциты были инактивированы, привели к лучшему пониманию назначения глиального рубца.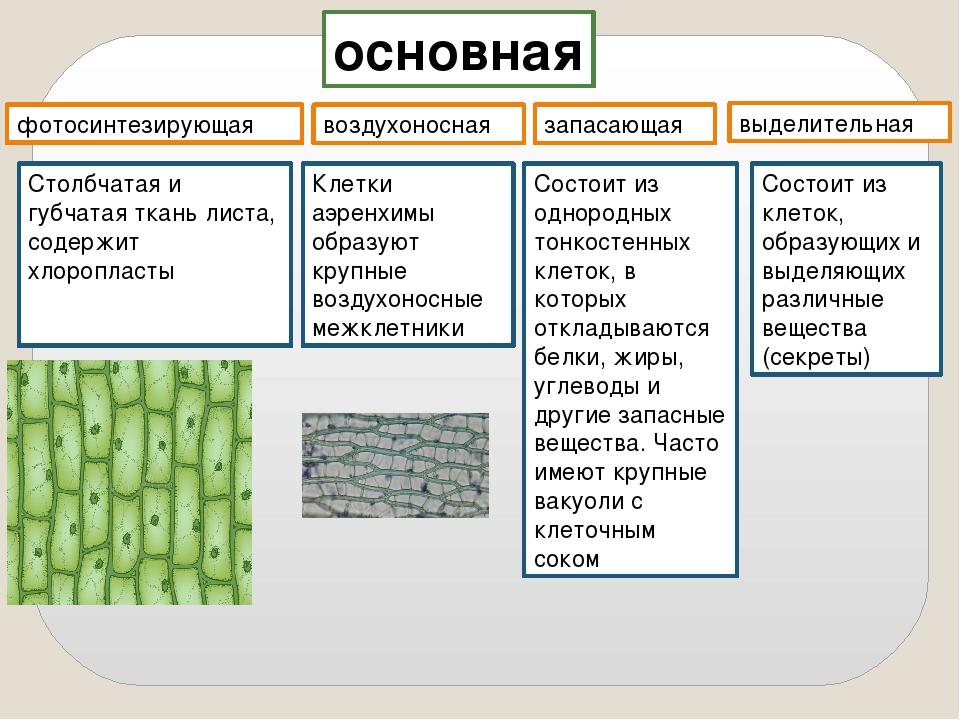 Селективное удаление реактивных астроцитов показывает отсутствие образования глиальных рубцов через 2 недели после колотых ран [109].Однако удаление астроцитов привело к потере сдерживания повреждения, поскольку воспалительные клетки разлились в ткани, окружающие исходную рану, увеличивая дегенерацию нейронов рядом с повреждением [109]. Считается, что глиальный рубец защищает функцию нейронов после повреждения, восстанавливая гематоэнцефалический барьер, и впоследствии ограничивает воспалительный ответ на клетки ЦНС [60,104].
Селективное удаление реактивных астроцитов показывает отсутствие образования глиальных рубцов через 2 недели после колотых ран [109].Однако удаление астроцитов привело к потере сдерживания повреждения, поскольку воспалительные клетки разлились в ткани, окружающие исходную рану, увеличивая дегенерацию нейронов рядом с повреждением [109]. Считается, что глиальный рубец защищает функцию нейронов после повреждения, восстанавливая гематоэнцефалический барьер, и впоследствии ограничивает воспалительный ответ на клетки ЦНС [60,104].
В отличие от ПНС, аксоны в ЦНС неспособны к регенерации. В среде глиального рубца две основные группы ингибирующих молекул препятствуют регенерации аксонов: те, которые связаны с глиальным рубцом, и те, которые связаны с миелином [111].Внутри глиального рубца хондроитинсульфат протеогликаны (CSPG) активируются астроцитами, предшественниками олигодендроцитов и менингеальными фибробластами. CPSG состоят из корового белка, к которому присоединяется различное количество повторяющихся дисахаридных хондроитинсульфатных цепей [111]. CSPG секретируются реактивными астроцитами в течение 24 часов после травмы и могут продолжаться в течение нескольких месяцев после этого [104, 112, 113]. Экспрессия протеогликана наиболее высока в центре поражения и снижается кнаружи [114].
CSPG секретируются реактивными астроцитами в течение 24 часов после травмы и могут продолжаться в течение нескольких месяцев после этого [104, 112, 113]. Экспрессия протеогликана наиболее высока в центре поражения и снижается кнаружи [114].
Миелин в ЦНС также содержит лиганды, ингибирующие рост, которые высвобождаются локально после травмы аксонов.Эти ингибирующие молекулы представляют собой Nogo, миелин-ассоциированный гликопротеин (MAG), олигодендроцит-миелиновый гликопротеин (OMgp), семафорин 4D и миелин-ассоциированные CSPG (см. Обзор [111]). И глиальный рубец, и миелин-ассоциированные молекулы предотвращают повторный рост, отталкивая или разрушая ростовые конусы через путь передачи сигналов Rho GTPase, блокируя сборку микротрубочек [115].
Основное различие между ПНС и ЦНС состоит в том, что в то время как поддерживающие шванновские клетки и астроциты как пролиферируют, так и активируются после травмы, шванновские клетки ПНС претерпевают изменения, создавая поддерживающую среду для регенерации, в то время как астроциты ЦНС претерпевают изменения, чтобы произвести ингибирующие молекулы, препятствующие регенерации нейронов. Это было продемонстрировано, поскольку аксоны ЦНС способны к регенерации посредством трансплантатов периферических нервов, имплантированных в раны за пределами ЦНС [116]. Было высказано предположение, что глиальный рубец не только защищает ткань ЦНС, прилегающую к повреждению, от дальнейшего повреждения, но также предотвращает реформирование нейронами несоответствующих нейронных связей после повреждения [60,113,117]. Рубцевание глии также связано с клиренсом глутамата и выработкой противовоспалительных цитокинов [110].
Это было продемонстрировано, поскольку аксоны ЦНС способны к регенерации посредством трансплантатов периферических нервов, имплантированных в раны за пределами ЦНС [116]. Было высказано предположение, что глиальный рубец не только защищает ткань ЦНС, прилегающую к повреждению, от дальнейшего повреждения, но также предотвращает реформирование нейронами несоответствующих нейронных связей после повреждения [60,113,117]. Рубцевание глии также связано с клиренсом глутамата и выработкой противовоспалительных цитокинов [110].
1.4.2.2. Индукция глиального рубца
Было показано, что цитокины, такие как IL-1, TGF-α, TGF-β, TNF-α, INF-α и INF-γ, активируются в рубцовой ткани после повреждения, способствуют пролиферации астроцитов в vitro и увеличивают глиальные рубцы in vivo (недавний обзор см. [104]) [110]. Было показано, что экспрессия TGF-β1 и TGF-β2 увеличивается сразу после повреждения головного и спинного мозга. TGF-β2 увеличивал как продукцию протеогликанов [118,119], так и рубцевание глии астроцитами [120].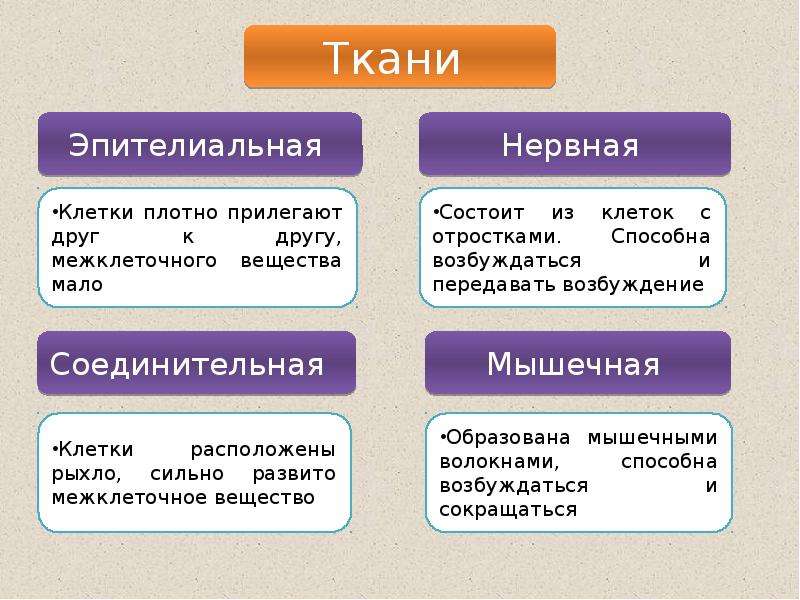 INF-γ и FGF2 также увеличивают степень глиального рубцевания и увеличивают пролиферацию астроцитов в культуре [104].
INF-γ и FGF2 также увеличивают степень глиального рубцевания и увеличивают пролиферацию астроцитов в культуре [104].
1.5. ПЕРЕМОДЕЛИРОВАНИЕ (НЕДЕЛИ И МЕСЯЦЫ)
Конечная конечная точка после ремоделирования зависит от типа ткани. В тканях, не относящихся к ЦНС, которые подвергаются первичному заживлению, происходит очень незначительное ремоделирование из-за отсутствия ECM, образующихся во время восстановления. Вторичное заживление, напротив, включает выравнивание и сокращение волокон для уменьшения размера раны и восстановления прочности тканей.Полное восстановление первоначальной прочности ткани редко достигается при вторичном заживлении, потому что восстановленная ткань остается менее организованной, чем неповрежденная ткань, что приводит к образованию рубца. Шрамы, богатые коллагеном, морфологически характеризуются отсутствием специфической организации клеточных и матричных элементов, составляющих окружающую неповрежденную ткань. В ткани ЦНС, где нет восстановления или регенерации поврежденных нейронов, восстановление структурной целостности в этой области также относительно невелико. Вместо этого во время ремоделирования ЦНС глиальный рубец вокруг поражения становится более плотным, поскольку астроцитарные процессы становятся более переплетенными и более или менее изолированными, но не восстанавливают поврежденную область.
Вместо этого во время ремоделирования ЦНС глиальный рубец вокруг поражения становится более плотным, поскольку астроцитарные процессы становятся более переплетенными и более или менее изолированными, но не восстанавливают поврежденную область.
1.5.1. Ремоделирование без ЦНС
1.5.1.1. Первичное ремоделирование
1.5.1.1.1. Ремоделирование кожной ткани частичной толщины
При поверхностных повреждениях раны могут зажить только за счет эпителизации, при этом для заполнения ткани требуется небольшой дополнительный ECM или не требуется его вовсе. Поскольку эти первичные тканевые повреждения могут зажить за счет регенерации кератиноцитов и минимального образования внеклеточного матрикса, требуется очень небольшое дополнительное ремоделирование ().В результате не остается рубцов, а восстановленная ткань практически неотличима от неповрежденной ткани.
В клинических условиях, когда есть небольшое загрязнение или некротический мусор, врачи используют наложение швов, чтобы привести дермальные края в прямое соприкосновение. После тщательного выравнивания и устранения напряжения эпидермальный и дермальный слои могут заживать в первую очередь за счет эпителизации внутри эпидермиса и с ограниченным образованием ECM в дерме, что приводит к ограниченному рубцеванию [121]. В течение периода от недель до месяцев, рана постепенно увеличивает свою прочность на растяжение по мере ремоделирования ВЦМ.Во время этого процесса ECM поврежденной ткани переориентируется относительно силы растяжения. Ремоделирование более важно для вторичного заживления и, следовательно, будет более подробно рассмотрено в разделе, посвященном полнослойным неминерализованным тканевым ранам.
После тщательного выравнивания и устранения напряжения эпидермальный и дермальный слои могут заживать в первую очередь за счет эпителизации внутри эпидермиса и с ограниченным образованием ECM в дерме, что приводит к ограниченному рубцеванию [121]. В течение периода от недель до месяцев, рана постепенно увеличивает свою прочность на растяжение по мере ремоделирования ВЦМ.Во время этого процесса ECM поврежденной ткани переориентируется относительно силы растяжения. Ремоделирование более важно для вторичного заживления и, следовательно, будет более подробно рассмотрено в разделе, посвященном полнослойным неминерализованным тканевым ранам.
1.5.1.1.2. Ремоделирование стабилизированной кости
Ремоделирование минерализованной ткани — это активный и динамичный процесс. Кость уникальна тем, что ремоделирование происходит на протяжении всей жизни ткани, поскольку механическое напряжение заставляет кость переориентировать себя и производить новую кость, чтобы лучше справляться с возложенными на нее требованиями. Согласно закону Вольфа, ткань адаптируется к окружающей среде, ориентируясь по линиям максимального стресса [112].
Согласно закону Вольфа, ткань адаптируется к окружающей среде, ориентируясь по линиям максимального стресса [112].
Ремоделирование после восстановления первичного перелома, при котором кость стабилизируется, аналогично реакции ремоделирования, которая происходит в течение жизни ткани, длится до нескольких лет до восстановления полной прочности до травмы [76]. При первичном заживлении промежутков ремоделирование важно для восстановления прочности тканей. Однако при первичном заживлении контактов ремоделирование связано с процессом восстановления.Во время ремоделирования контакта режущие конусы созревают, центростремительно откладывая пластинчатую кость, образуя кольцеобразные структуры с центральным каналом, содержащим кровеносные сосуды ().
Рисунок 1.8
Сравнение первичного и вторичного ремоделирования кости. (A) Пластинчатая кость, отложенная перпендикулярно длинной оси кости во время первичного ремонта, используется в качестве основы для разрезания конусов. Остеокласты создают туннели, по которым следуют новые кровеносные сосуды, стимулируя (подробнее …)
Остеокласты создают туннели, по которым следуют новые кровеносные сосуды, стимулируя (подробнее …)
При ремоделировании первичного зазора пластинчатая кость, расположенная перпендикулярно длинной оси внутри повреждения во время ремонта, используется в качестве каркаса ().Этот процесс обычно называют гаверсовским ремоделированием, когда кость ремоделируется в небольшие пакеты клеток, называемые базовыми многоклеточными единицами (BMU) [123]. Это тот же процесс, который происходит во время восстановления первичного контакта, когда остеокласты образуют режущие конусы, обеспечивая приток эндотелиальных клеток, которые образуют капилляры (). Почкующиеся капилляры приносят плюрипотентные мезенхимные клетки, которые после прикрепления к поверхности внутреннего режущего конуса дифференцируются в остеобласты. Остеобласты активируются и концентрически синтезируют новую пластинчатую кость, постепенно уменьшая диаметр туннеля ().Примерно через четыре недели образование кости прекращается, так как туннель закрывается, оставляя после себя васкуляризованную полость, называемую остеоном, которая проходит параллельно длинной оси [96].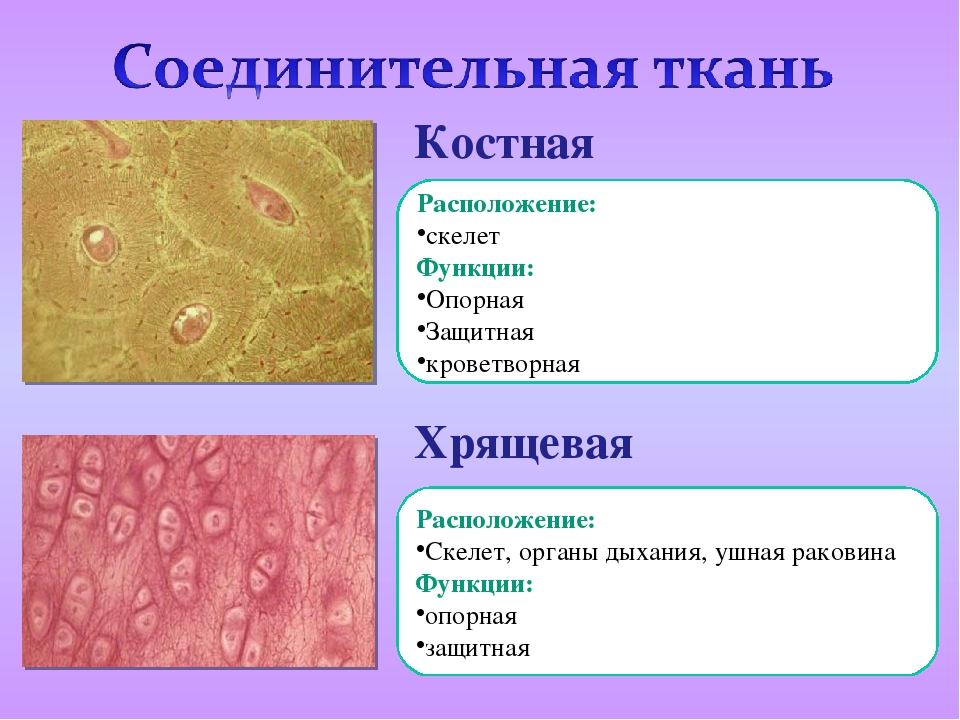 Считается, что чем большее количество остеонов пересекает место повреждения, тем выше предел прочности [79].
Считается, что чем большее количество остеонов пересекает место повреждения, тем выше предел прочности [79].
1.5.1.1.3. Ремоделирование ПНС
В ПНС, когда регенерирующие аксоны находят свою цель, ткань созревает, поскольку аксоны постепенно увеличиваются в толщине за счет синтеза нейрофиламентов (). Диаметр аксонов и толщина миелиновой оболочки регенерированных нейронов обычно меньше и никогда не достигают нормального уровня до травмы [39].Дочерние аксоны, не контактирующие с целью, отщепляются. Во время регенерации родительский аксон прорастает в среднем 3 дочерних аксона, хотя было обнаружено до 25 дочерних аксонов [39]. Большее количество нейронной гибели обычно приводит к большему прорастанию, поскольку конкуренция за доступ к цели меньше [39]. ПНС способна поддерживать регенеративный ответ как минимум через 12 месяцев после травмы [124]. Каркасы шванновских клеток, которые остаются неиннервируемыми, медленно уменьшаются в диаметре, и если они не получают регенерирующий аксон, они теряют опорную способность, поскольку они постепенно заполняются фиброзной тканью [124].
Восстановление и ремоделирование аксонов зависит от тяжести травмы. Большинство травм в ПНС связано с растяжением, когда растягивающая сила, приложенная к ткани, превышает способность нерва растягиваться. Обычно при этих повреждениях внешний соединительнотканный слой нерва, эндоневрий, не поврежден, и аксон способен регенерироваться, как показано ранее () [124]. Однако при более травматических повреждениях, таких как разрывы, эндоневрий и аксон разрываются, и ткань обычно не восстанавливается.После травм фибробласты, содержащиеся в эндоневрии, разрастаются и образуют коллагеновый рубец вокруг нервного ствола во время фаз восстановления и ремоделирования. Этот коллагеновый рубец неверно направляет или блокирует регенерацию аксонов [124]. Прорастающие аксоны, которые не могут найти свою цель или контактировать с каналом шванновских клеток, либо расщепляются, либо вырастают в неорганизованную массу, что приводит к образованию невромы. Между тем, целевая мышца остается неактивной, а тела нервных клеток атрофируются и в конечном итоге умирают. Даже когда эндоневрий не разорван, регенерация целевого участка чаще всего терпит неудачу, и даже в лучшем случае регенерация периферического нерва не восстанавливает полностью ткань до ее исходного состояния, поскольку во время попытки неизбежно возникает неточность. возвращение аксона к его исходной цели [88].
Даже когда эндоневрий не разорван, регенерация целевого участка чаще всего терпит неудачу, и даже в лучшем случае регенерация периферического нерва не восстанавливает полностью ткань до ее исходного состояния, поскольку во время попытки неизбежно возникает неточность. возвращение аксона к его исходной цели [88].
Хирургическое введение аутологичного нервного трансплантата может использоваться для восстановления поражений ПНС, которые слишком велики для того, чтобы их перекрыли шванновские клетки. Назначение трансплантата — восстановить соединение поврежденных нервов от конца к концу, не вызывая напряжения.Для соединения соединительнотканной оболочки поврежденного нерва с аутотрансплантатом используется наложение швов. Свежеповрежденные тканевые оболочки плохо держат швы, поэтому хирургическое вмешательство, как правило, не проводят до трех недель после травмы, когда оболочки успевают утолщаться. Главный недостаток этого метода восстановления заключается в том, что забор аутологичного нерва влечет за собой жертву одного или нескольких нервов. Ряд групп работают над созданием синтетических нервных трансплантатов для восстановления ПНС [86,125].
Ряд групп работают над созданием синтетических нервных трансплантатов для восстановления ПНС [86,125].
1.5.1.2. Вторичный ремонт
1.5.1.2.1. Ремоделирование кожной ткани полной толщины
Основная цель ремоделирования вторичной раны — уменьшить количество избыточного ВКМ и выровнять ВКМ посредством сокращения. Если размер раны относительно невелик и реорганизация внеклеточного матрикса эффективна, возникает относительно небольшая контрактура и наблюдается незначительное рубцевание или его отсутствие. Однако при более крупных травмах, когда происходит более обширное восстановление тканей за счет образования внеклеточного матрикса, требуется значительное ремоделирование, которое приводит к рубцеванию.Ремоделирование происходит в течение длительного периода времени. Эта фаза может совпадать с фазой восстановления, так как она может начаться уже через 1 неделю после травмы и может длиться до 2 лет, в зависимости от раны.
Во время репарации фибробласты мигрируют в место повреждения и производят ECM для замещения утраченной ткани. Силы тракции, которые создают фибробласты при движении через раневую ткань, создают механическое напряжение, которое способствует закрытию раны [126]. Фаза ремоделирования официально начинается, когда TGF-β и другие цитокины, высвобождаемые из тромбоцитов и активированных макрофагов, заставляют фибробласты дифференцироваться в более сократительный фенотип, называемый миофибробластами.Миофибробласты характеризуются экспрессией альфа-актина гладких мышц и выработкой коллагена I [28,127]. После активации они увеличивают свои цитоскелетные стрессовые волокна и фокальные спайки, обеспечивая постоянное напряжение для сокращения раневого ложа (см. Обзор [127]). Сокращение миофибробластов похоже на сокращение гладкомышечных клеток, хотя способ активации сильно отличается. Гладкомышечные клетки сокращаются из-за повышения Ca 2+ . Поскольку уровни Ca 2+ вокруг клеток могут быстро изменяться, сократимость гладкомышечных клеток может быстро меняться [128].Считается, что миофибробласты регулируются Rho-Rho киназным путем, который является менее временным и вызывает более длительную силу сокращения.
Силы тракции, которые создают фибробласты при движении через раневую ткань, создают механическое напряжение, которое способствует закрытию раны [126]. Фаза ремоделирования официально начинается, когда TGF-β и другие цитокины, высвобождаемые из тромбоцитов и активированных макрофагов, заставляют фибробласты дифференцироваться в более сократительный фенотип, называемый миофибробластами.Миофибробласты характеризуются экспрессией альфа-актина гладких мышц и выработкой коллагена I [28,127]. После активации они увеличивают свои цитоскелетные стрессовые волокна и фокальные спайки, обеспечивая постоянное напряжение для сокращения раневого ложа (см. Обзор [127]). Сокращение миофибробластов похоже на сокращение гладкомышечных клеток, хотя способ активации сильно отличается. Гладкомышечные клетки сокращаются из-за повышения Ca 2+ . Поскольку уровни Ca 2+ вокруг клеток могут быстро изменяться, сократимость гладкомышечных клеток может быстро меняться [128].Считается, что миофибробласты регулируются Rho-Rho киназным путем, который является менее временным и вызывает более длительную силу сокращения. Возможны контрактуры от 10 до 20 мкм в день без необходимости создания большого количества силы из-за низкого уровня базального натяжения кожи [127]. Миофибробласты продолжают поддерживать нагрузки в сокращенной ткани до тех пор, пока ECM не будет продуцироваться и сшиться, что приводит к экранированию стресса [127]. Коллагеновые волокна постепенно утолщаются и вместе с миофибробластами становятся ориентированными параллельно ложу раны по линиям напряжения, в результате чего появляется полосатая рубцовая ткань ().Это прямо контрастирует с рисунком плетения корзины, наблюдаемым на неповрежденной коже [129].
Возможны контрактуры от 10 до 20 мкм в день без необходимости создания большого количества силы из-за низкого уровня базального натяжения кожи [127]. Миофибробласты продолжают поддерживать нагрузки в сокращенной ткани до тех пор, пока ECM не будет продуцироваться и сшиться, что приводит к экранированию стресса [127]. Коллагеновые волокна постепенно утолщаются и вместе с миофибробластами становятся ориентированными параллельно ложу раны по линиям напряжения, в результате чего появляется полосатая рубцовая ткань ().Это прямо контрастирует с рисунком плетения корзины, наблюдаемым на неповрежденной коже [129].
Когда происходит сокращение раны, расслабление стресса заставляет миофибробласты возвращаться в состояние покоя. Затем клетки получают сигнал к апоптозу, трансформируя рану из богатой клетками грануляционной ткани в бедную клетками рубцовую ткань с избытком ECM [130]. В то же время плотность капилляров постепенно уменьшается, и рана теряет розовый цвет, становясь все бледнее [27]. Конечной конечной точкой процесса ремоделирования является образование бесклеточной рубцовой ткани, которая плохо реорганизуется в плотные параллельные пучки, в отличие от плотно сплетенной сети нормальной кожной ткани () [72].Кожные структуры, такие как волосяные фолликулы, потовые и сальные железы, утраченные во время травмы, не регенерируются [69]. Через три месяца неминерализованная ткань может иметь максимум 80% прочности неизвлекаемой ткани [68]. Обычно это наивысший уровень прочности, которого может достичь зажившая ткань.
Конечной конечной точкой процесса ремоделирования является образование бесклеточной рубцовой ткани, которая плохо реорганизуется в плотные параллельные пучки, в отличие от плотно сплетенной сети нормальной кожной ткани () [72].Кожные структуры, такие как волосяные фолликулы, потовые и сальные железы, утраченные во время травмы, не регенерируются [69]. Через три месяца неминерализованная ткань может иметь максимум 80% прочности неизвлекаемой ткани [68]. Обычно это наивысший уровень прочности, которого может достичь зажившая ткань.
1.5.1.2.2. Нестабилизированное ремоделирование кости
Конечная цель вторичного ремоделирования минерализованной ткани — полное структурное восстановление с минимальным рубцеванием или без него ().Хотя ремоделирование может длиться до 6 месяцев, конечной точкой является зажившая ткань, которая очень похожа на неповрежденную ткань с точки зрения прочности, но может выглядеть немного менее организованной [78]. Единственное существенное различие между первичным и вторичным ремоделированием минерализованной ткани — это обширное удаление костной ткани, необходимое для удаления избыточной костной мозоли, образовавшейся во время восстановления кости.
При вторичном заживлении остеокласты попадают в участок, нуждающийся в ремоделировании, через кровообращение. Они распознают и прикрепляются к белкам клеточной адгезии, таким как остеопонтин, остеокальцин и остеонектин [83].Остеокласты присутствуют не только для удаления лишней кости, не необходимой для структурной поддержки, но и для переваривания тканой кости, синтезированной из мягкой мозоли во время восстановления, чтобы ее можно было заменить пластинчатой костью, выровненной в ответ на стресс. Подобно первичному ремоделированию, вторичное ремоделирование происходит в основном за счет режущих конусов. Внутри коры остеокласты удаляют тканые и некротические пластинки кости. Остеобласты следуют друг за другом и образуют новую компактную пластинчатую кость, придавая ткани большую прочность ().Каждый концентрический ламеллярный слой толщиной 100 мкм направлен особым образом, чтобы укрепить фрагменты трещин [96]. Вторичное ремоделирование отличается от первичного ремоделирования из-за большой внешней костной мозоли, образовавшейся во время ремонта. Наружная мозоль образуется за пределами коры и обычно ремоделируется остеокластами без образования режущих конусов и остеонов. Это возможно, потому что остеокласты имеют непосредственный доступ к тканой кости внешней костной мозоли через надкостницу ().
Наружная мозоль образуется за пределами коры и обычно ремоделируется остеокластами без образования режущих конусов и остеонов. Это возможно, потому что остеокласты имеют непосредственный доступ к тканой кости внешней костной мозоли через надкостницу ().
Большая часть кости, удаленной во время ремоделирования коры, заменяется пластинчатой костью.Во время ремоделирования механические нагрузки, прикладываемые к минерализованной ткани, способны генерировать сигналы на клеточном уровне для увеличения производства кости (см. Обзор [80]). Остеоциты, являющиеся наиболее многочисленными клетками в кости, заключены в костном матриксе и способны связываться с соседними клетками через сеть отростков, соединенных щелевыми соединениями. Механический стресс в минерализованной ткани вызывает сдвиг жидкости на остеоциты и вызывает приток внеклеточных ионов кальция, а также высвобождение АТФ, что приводит к активации ионных каналов [80].Это, в свою очередь, может стимулировать синтез пластинчатой кости за счет активации определенных простагландинов и увеличения продукции оксида азота [80].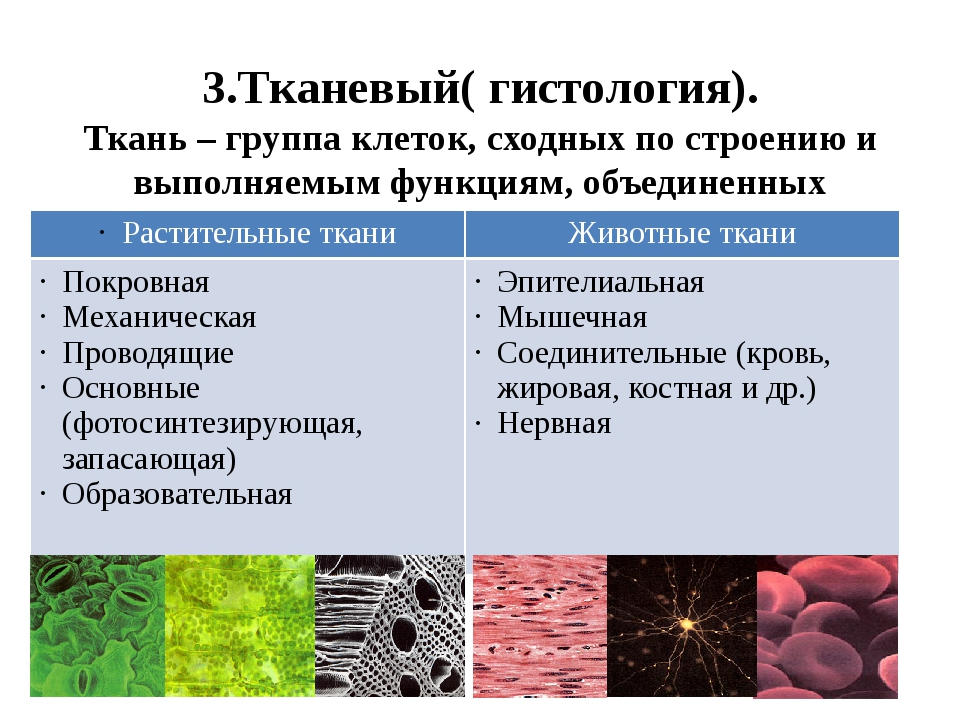 Было показано, что активность остеобластов увеличивается в присутствии некоторых простагландинов [131], в то время как оксид азота, как известно, ингибирует резорбцию кости остеокластами [132].
Было показано, что активность остеобластов увеличивается в присутствии некоторых простагландинов [131], в то время как оксид азота, как известно, ингибирует резорбцию кости остеокластами [132].
1.5.2. Ремоделирование ЦНС
Ремоделирование ЦНС ограничено. Из-за необходимости защиты ЦНС от сильных воспалительных реакций организма реактивные астроцитарные процессы еще больше переплетаются, образуя плотную оболочку вокруг места раны ().Во время стадий восстановления реактивность GFAP наблюдается примерно в 400 мкм за пределами поражения. Во время стадий ремоделирования реактивность GFAP наблюдается только на расстоянии менее 100 мкм от места повреждения ткани [133]. Плотность глиальных рубцов увеличивается по мере того, как гипертрофические астроциты становятся более конденсированными вокруг места повреждения () [11].
Рис. 1.9
Динамика формирования глиального рубца в четырех временных точках, как показано с помощью окрашивания GFAP. Через 2 и 4 недели астроцитарные процессы возвращаются в пустоту, оставшуюся после извлечения зонда перед обработкой ткани.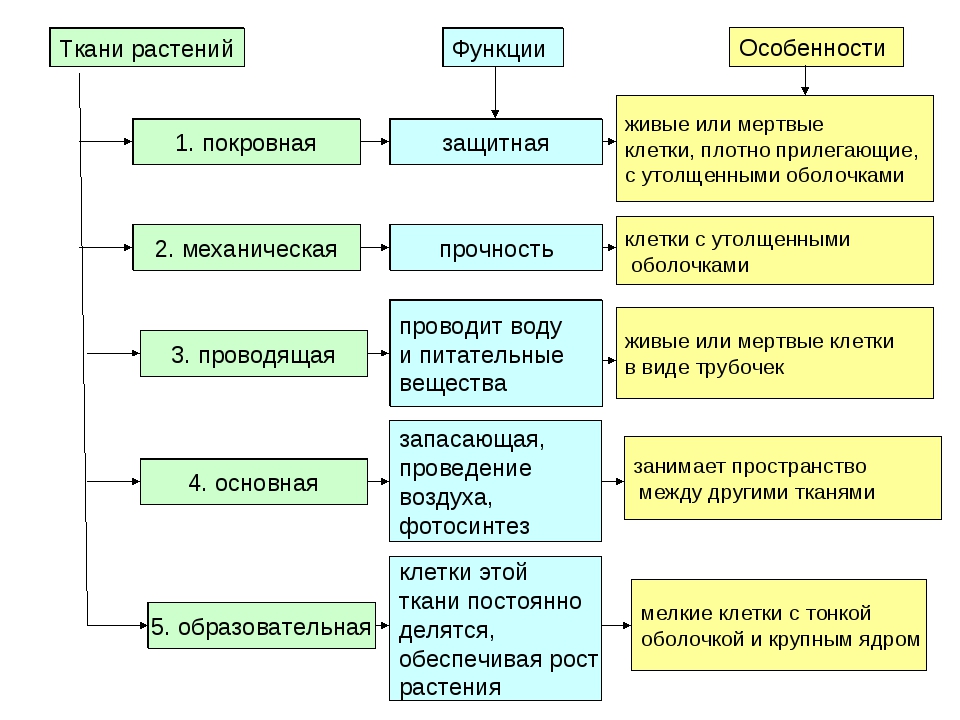 К 6 неделям отростки переплелись (подробнее …)
К 6 неделям отростки переплелись (подробнее …)
Внутри поражения удаление фибринового сгустка, некротических нейронов и поддерживающих глиальных клеток завершается микроглией и макрофагами () [105]. После завершения микроглия и макрофаги подвергаются апоптозу, оставляя кисту, заполненную спинномозговой жидкостью, в центре поражения, где возникла первоначальная рана [105]. Киста граничит с тонким плотным слоем реактивных астроцитов, которые служат барьером между здоровой и утраченной тканью и могут помочь защитить нейроны за пределами травмы.Аксоны нейронов, защищенные за пределами глиального рубца, не могут снова превратиться в потерянную ткань из-за тормозных молекул внутри рубца, и, таким образом, функция ткани никогда не восстанавливается ().
1.6. ЗАКЛЮЧЕНИЕ
Клеточная реакция после травмы зависит от типа ткани, а также от размера раны. При повреждении ткани ЦНС, которое повреждает нейроны и поддерживающие глиальные клетки, реакция организма неумолима, поскольку регенерация потерянных нейронов невозможна. Активированные астроциты защищают очаг поражения, создавая глиальный рубец.Эти активированные астроциты могут предотвращать дальнейшее повреждение тканей, хотя повторный рост аксонов нейронов подавляется. Напротив, в тканях, не относящихся к ЦНС, один тип ткани может давать несколько ответов в зависимости от степени повреждения. Например, поверхностная кожная рана часто имеет более низкий уровень воспалительного инфильтрата и может подвергаться первичному заживлению, в то время как более глубокая рана с более обширным повреждением тканей и потерей клеток подвергнется вторичному заживлению. В этой главе основное внимание уделялось первичной реакции заживления, наблюдаемой после травм кожи, костей и ПНС, когда общая структура ткани сохраняется после травмы, позволяя тканям подвергаться восстановлению преимущественно за счет регенерации потерянных клеток для восстановления нормальной структуры ткани.При вторичном заживлении раны часто бывают более глубокими, и общая структура ткани нарушается.
Активированные астроциты защищают очаг поражения, создавая глиальный рубец.Эти активированные астроциты могут предотвращать дальнейшее повреждение тканей, хотя повторный рост аксонов нейронов подавляется. Напротив, в тканях, не относящихся к ЦНС, один тип ткани может давать несколько ответов в зависимости от степени повреждения. Например, поверхностная кожная рана часто имеет более низкий уровень воспалительного инфильтрата и может подвергаться первичному заживлению, в то время как более глубокая рана с более обширным повреждением тканей и потерей клеток подвергнется вторичному заживлению. В этой главе основное внимание уделялось первичной реакции заживления, наблюдаемой после травм кожи, костей и ПНС, когда общая структура ткани сохраняется после травмы, позволяя тканям подвергаться восстановлению преимущественно за счет регенерации потерянных клеток для восстановления нормальной структуры ткани.При вторичном заживлении раны часто бывают более глубокими, и общая структура ткани нарушается.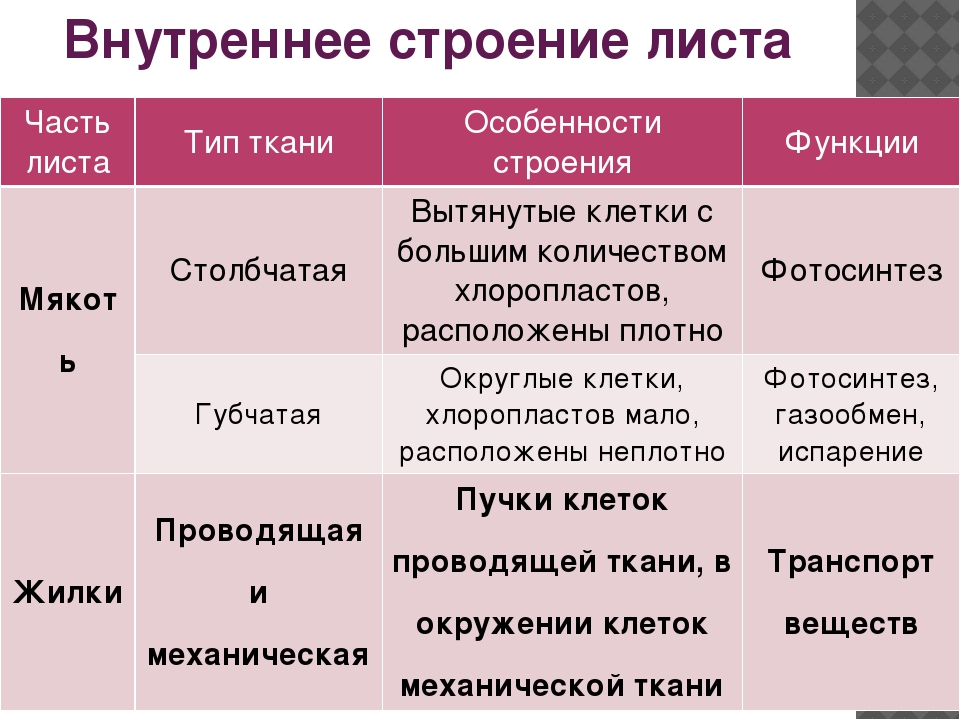 Воспалительная реакция может быть более интенсивной и продолжительной, так как в нее вовлекаются фибробласты и эндотелиальные клетки, которые продуцируют грануляционную ткань. В грануляционной ткани отложение коллагена фибробластами изменяет структуру и функцию ткани и в конечном итоге приводит к образованию рубцов по завершении стадии ремоделирования. Вторичное заживление кости является исключением, поскольку костная ткань способна удалить исходную фиброзную мозоль и заменить ее пластинчатой костью.Другие примеры рубцевания после вторичного заживления возникают в тканях, не способных к регенерации, таких как сердце.
Воспалительная реакция может быть более интенсивной и продолжительной, так как в нее вовлекаются фибробласты и эндотелиальные клетки, которые продуцируют грануляционную ткань. В грануляционной ткани отложение коллагена фибробластами изменяет структуру и функцию ткани и в конечном итоге приводит к образованию рубцов по завершении стадии ремоделирования. Вторичное заживление кости является исключением, поскольку костная ткань способна удалить исходную фиброзную мозоль и заменить ее пластинчатой костью.Другие примеры рубцевания после вторичного заживления возникают в тканях, не способных к регенерации, таких как сердце.
Таким образом, хотя общий результат заживления ран ЦНС и тканей, не связанных с ЦНС, был описан, для терапевтических целей в ЦНС важно учитывать различные стыки в процессе заживления ран, при которых реакция может быть изменена на подтолкнуть реакцию заживления ран тела к желаемому результату. Эти точки вмешательства в основном находятся на стадиях гемостаза, воспаления и восстановления.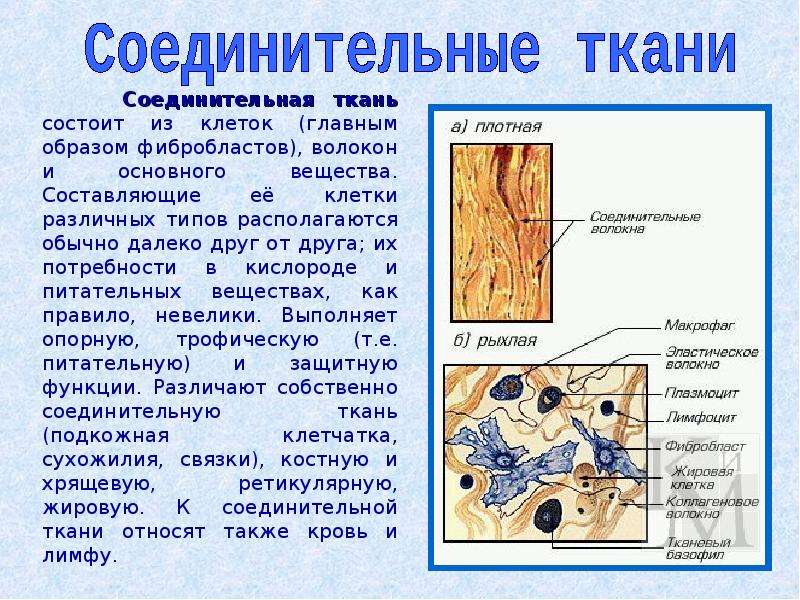 Внутри каждого этапа есть точки, которые могут быть полезны для регулирования реакции заживления ран. Например, чтобы ограничить обширное образование сгустка, было предложено, что существует три основных этапа в образовании сгустка, для которых могут быть разработаны терапевтические средства: инициирование (активация тромбоцитов), распространение (каскад коагуляции) и образование фибрина (тромбин) [134] . Во время воспаления стратегии модуляции сосредоточены вокруг ограничения активации иммунных клеток и уменьшения миграции воспалительных клеток в рану [67].Поскольку нейроны в ЦНС особенно чувствительны к воспалительному повреждению [48], и поскольку хроническое воспаление участвует во многих других заболеваниях за пределами ЦНС, было приложено много усилий для устранения механизмов, с помощью которых воспаление ослабляется, с надеждой на то, что достижение баланса между профилактикой инфекций и разрешением воспаления [67]. Наконец, чтобы изменить процесс восстановления ЦНС для улучшения функции ЦНС после травмы, исследователи попытались снизить активность астроцитов [135] и стимулировать регенерацию нервов [111].
Внутри каждого этапа есть точки, которые могут быть полезны для регулирования реакции заживления ран. Например, чтобы ограничить обширное образование сгустка, было предложено, что существует три основных этапа в образовании сгустка, для которых могут быть разработаны терапевтические средства: инициирование (активация тромбоцитов), распространение (каскад коагуляции) и образование фибрина (тромбин) [134] . Во время воспаления стратегии модуляции сосредоточены вокруг ограничения активации иммунных клеток и уменьшения миграции воспалительных клеток в рану [67].Поскольку нейроны в ЦНС особенно чувствительны к воспалительному повреждению [48], и поскольку хроническое воспаление участвует во многих других заболеваниях за пределами ЦНС, было приложено много усилий для устранения механизмов, с помощью которых воспаление ослабляется, с надеждой на то, что достижение баланса между профилактикой инфекций и разрешением воспаления [67]. Наконец, чтобы изменить процесс восстановления ЦНС для улучшения функции ЦНС после травмы, исследователи попытались снизить активность астроцитов [135] и стимулировать регенерацию нервов [111].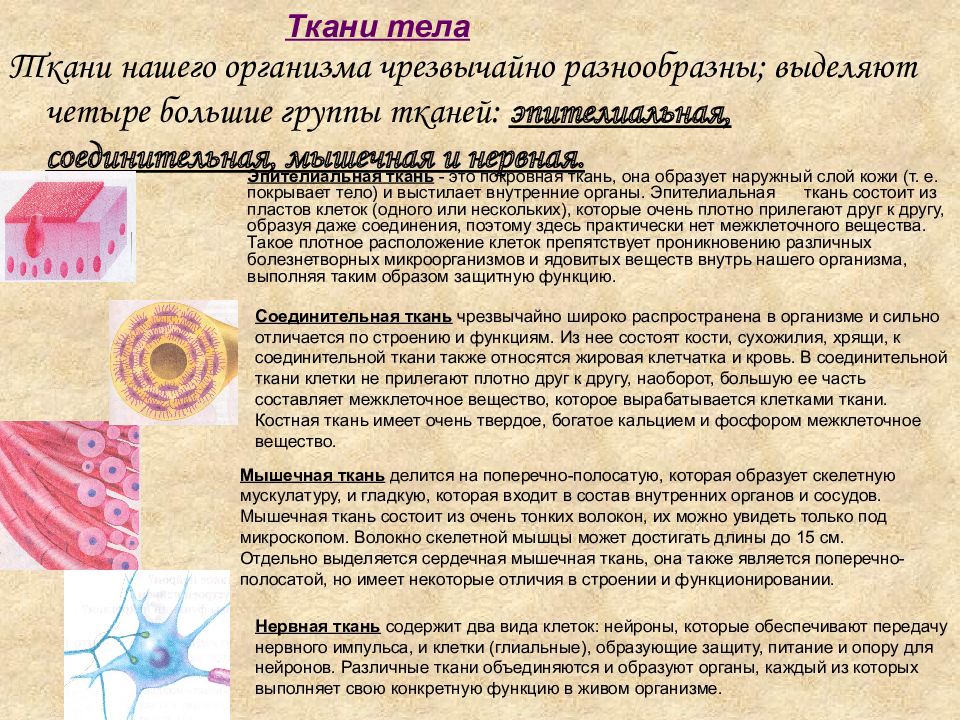
К вмешательствам по заживлению ран ЦНС, направленным на изменение глиального рубца и воспалительных процессов, следует подходить с осторожностью. Уменьшение глиальных рубцов может привести к ненужному повреждению тканей из-за неспособности восстановить нормальный гомеостаз и восстановление гематоэнцефалического барьера [109,136]. Одной из основных проблем, связанных с противовоспалительными подходами, является потеря защиты от инфекции, хотя методы до сих пор не продемонстрировали существенного увеличения восприимчивости к инфекции, что позволяет предположить, что полностью остановить воспалительный процесс сложно из-за дублирования пути [67]. ].
СПИСОК ЛИТЕРАТУРЫ
- 1.
- Вишневски Н., Райхерт М. Методы уменьшения биообрастания мембран биосенсоров. Коллоиды и поверхности B: Биоинтерфейсы. 2000. 18 (3-4): 197. [PubMed: 10
- 4]
- 2.
- Герритсен М., Янсен Дж. Характеристики подкожно имплантированных сенсоров глюкозы: обзор. Журнал следственной хирургии.
 1998; 11: 163. [PubMed: 9743484]
1998; 11: 163. [PubMed: 9743484] - 3.
- Уилсон Г.С., Гиффорд Р. Биосенсоры для измерений in vivo в реальном времени. Биосенсоры и биоэлектроника.2005; 20 (12): 2388. [PubMed: 15854814]
- 4.
Апдайк С., Шульц М., Роудс Р. Принципы долгосрочных полностью имплантированных датчиков с упором на радиотелеметрический мониторинг уровня глюкозы в крови внутри подкожной капсулы с инородным телом (FBC) In: Fraser Д, редактор. Биосенсоры в организме. Нью-Йорк: Уайли; 1997. стр. 117.
- 5.
Райхерт В., Шаркави А. Биосенсоры. В: фон Рекум А, редактор. Справочник по оценке биоматериалов. Анн-Арбор, Мичиган: Тейлор и Фрэнсис; 1999 г.
- 6.
- Wisniewski N, Moussy F, Reichert WM. Характеристика биообрастания имплантируемой мембраны биосенсора. Журнал аналитической химии Фрезениуса. 2000. 366 (6–7): 611. [PubMed: 11225773]
- 7.
- Поликов В.С., Треско П.А., Райхерт В.М. Ответ мозговой ткани на хронически имплантированные нервные электроды.
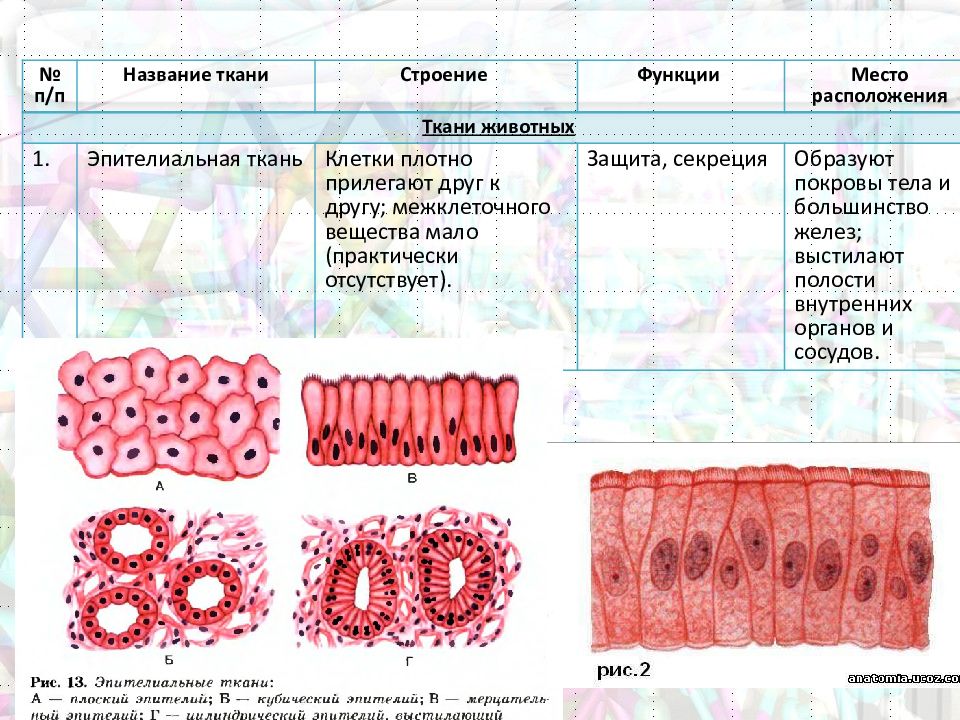 Журнал методов неврологии. 2005; 148 (1): 1. [PubMed: 16198003]
Журнал методов неврологии. 2005; 148 (1): 1. [PubMed: 16198003] - 8.
- Биран Р., Мартин, округ Колумбия, Треско, Пенсильвания. Потеря нейрональных клеток сопровождает реакцию ткани мозга на хронически имплантированные кремниевые микроэлектродные матрицы.Экспериментальная неврология. 2005; 195 (1): 115. [PubMed: 16045910]
- 9.
- Ким И-Т, Хичкок Р.В., Бридж М.Дж., Треско, Пенсильвания. Хронический ответ мозговой ткани взрослой крысы на имплантаты, прикрепленные к черепу. Биоматериалы. 2004; 25 (12): 2229. [PubMed: 14741588]
- 10.
- Саровски Д.Х., Андерсен М.Д., Реттерер С., Спенс А.Дж., Исааксон М., Крейгхед Х.Г., Тернер Дж.Н., Шейн У. Исследование мозга. 2003. 983 (1-2): 23. [PubMed: 12
- 3]
- 11.
- Тернер Дж. Н., Шайн В., Саровски Д. Х., Андерсен М., Мартинс С., Исааксон М., Крейгхед Х. Ответ церебральных астроцитов на микромашинные силиконовые имплантаты. Экспериментальная неврология. 1999; 156 (1): 33. [PubMed: 10192775]
- 12.
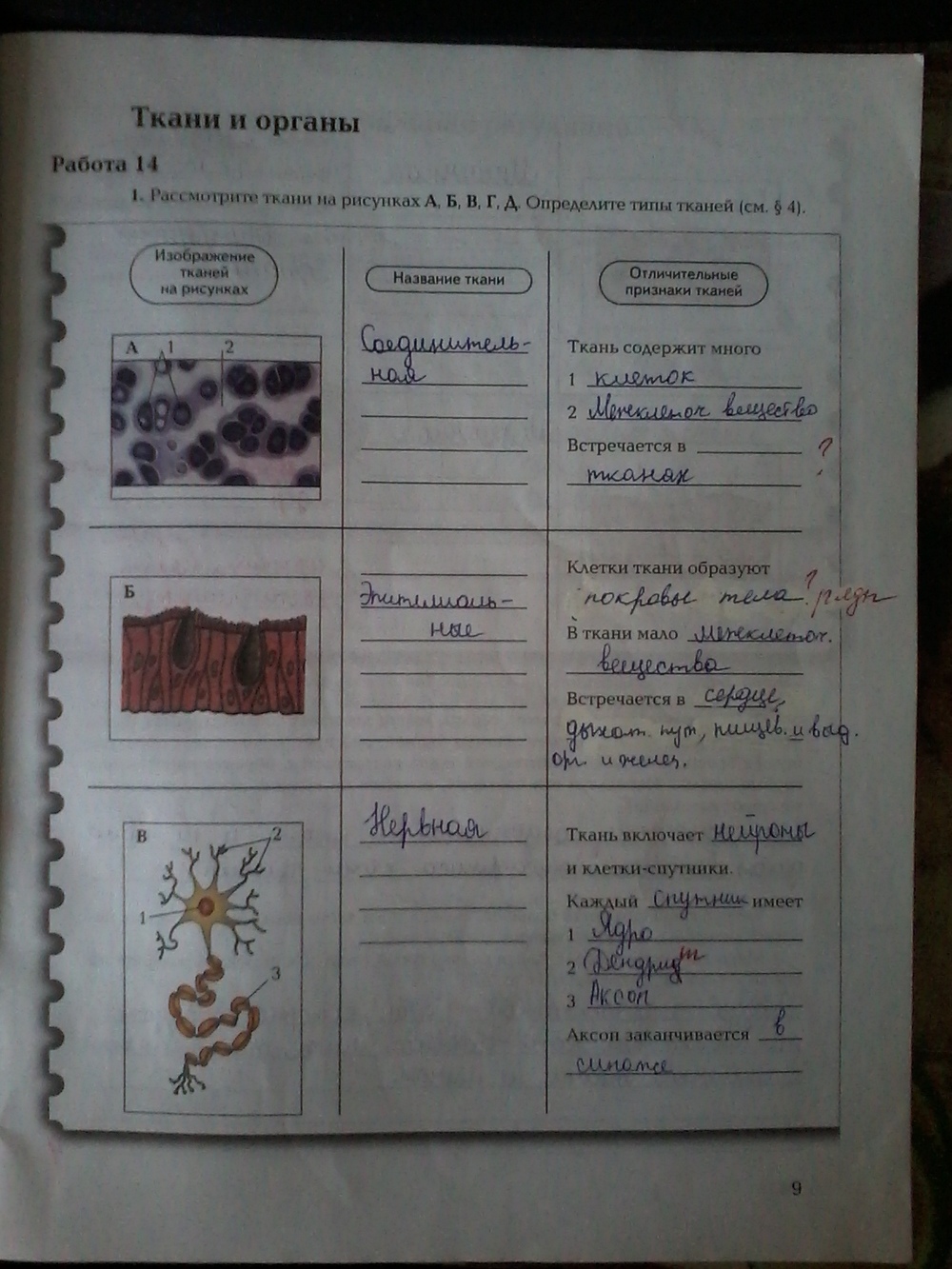
- Кармена Дж. М., Лебедев М. А., Крист Р. Э., О’Догерти Дж. Э., Сантуччи Д. Ф., Димитров Д. Ф., Патил П. Г., Энрикес К. С., Николелис МАЛ. Научиться управлять интерфейсом мозг-машина для достижения и хватания приматов. PLoS Биология. 2003; 1 (2): 193. [Бесплатная статья PMC: PMC261882] [PubMed: 14624244]
- 13.
- Hochberg LR, Serruya MD, Friehs GM, Mukand JA, Saleh M, Caplan AH, Branner A, Chen D., Penn RD, Donoghue JP. Контроль нейронного ансамбля протезных устройств у человека с тетраплегией. Природа. 2006; 442 (7099): 164. [PubMed: 16838014]
- 14.
- Кларк Р., Шидинг Дж., Качоровски С., Марион Д., Кочанек П. Накопление нейртрофилов после черепно-мозговой травмы у крыс: сравнение моделей падения веса и контролируемого кортикального воздействия. Журнал нейротравмы. 1994; 11: 499. [PubMed: 7861443]
- 15.
Первес Д., Августин Дж., Фицпатрик Д., Кац Л., Ламантия А., Макнамара Дж., Уильямс В. Сандерленд, Массачусетс: Sinauer Associates; 2001.
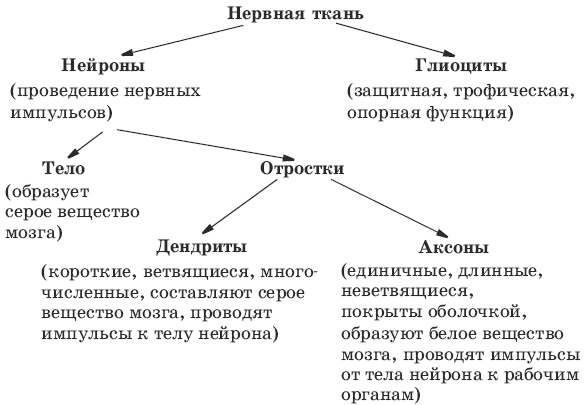 Организация нервной системы.
Организация нервной системы.- 16.
- Рубин Л.Л., Стаддон Дж. М.. Клеточная биология гематоэнцефалического барьера. Ежегодный обзор неврологии. 1999; 22 (1): 11. [PubMed: 10202530]
- 17.
- Шварц М. Макрофаги и микроглия при повреждении центральной нервной системы: полезны они или вредны? Журнал мозгового кровотока и метаболизма.2003; 23: 385. [PubMed: 12679714]
- 18.
- Seung UK, de Vellis J. Microglia в здоровье и болезнях. Журнал неврологических исследований. 2005; 81 (3): 302. [PubMed: 15954124]
- 19.
- Ruggeri ZM. Тромбоциты при атеротромбозе. Природная медицина. 2002; 8 (11): 1227. [PubMed: 12411949]
- 20.
- Стандевен К.Ф., Ариенс РАН, Грант П.Дж. Молекулярная физиология и патология структуры / функции фибрина. Обзоры крови. 2005; 19 (5): 275. [PubMed: 15963835]
- 21.
- del Zoppo F, Poeck K, Pessin M, Wolpert S, Furlan A, Febert A, Alberts M, Zivin J, Wechsler L., Busse O.
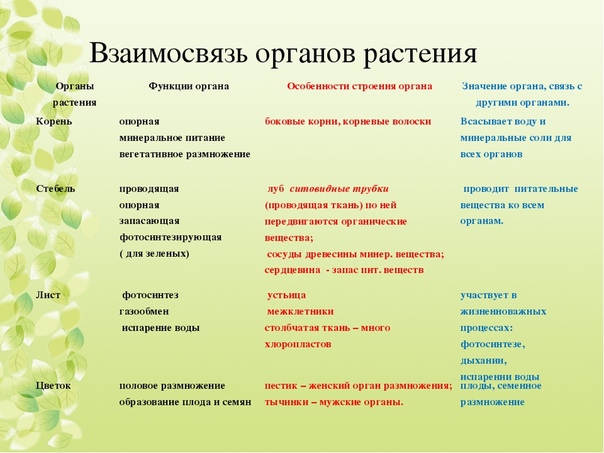 Рекомбинантный тканевый активатор плазминогена при остром тромботическом и эмболическом инсульте. Анналы неврологии. 1992; 68: 642. [PubMed: 1642475]
Рекомбинантный тканевый активатор плазминогена при остром тромботическом и эмболическом инсульте. Анналы неврологии. 1992; 68: 642. [PubMed: 1642475] - 22.
- Rosenberg RD, Aird WC. Гемостаз, специфичный для сосудистого русла, и состояния гиперкоагуляции. Медицинский журнал Новой Англии. 1999; 340 (20): 1555. [PubMed: 10332019]
- 23.
- Heldin CH, Westermark B, Wasteson A.Фактор роста тромбоцитов. Выделение с помощью крупномасштабной процедуры и анализ субъединичного состава. Биохимический журнал. 1981; 193 (3): 907. [Бесплатная статья PMC: PMC1162684] [PubMed: 7305965]
- 24.
- Assoian RK, Komoriya A, Meyers CA, Miller DM, Sporn MB. Трансформирующий фактор роста бета в тромбоцитах человека. Идентификация основного места хранения, очистка и характеристика. Журнал биологической химии. 1983; 258 (11): 7155. [PubMed: 6602130]
- 25.
- Mohle R, Green D, Moore Malcolm AS, Nachman RL, Rafii S.Конститутивная продукция и тромбин-индуцированное высвобождение фактора роста эндотелия сосудов мегакариоцитами и тромбоцитами человека.
 Труды Национальной академии наук США. 1997; 94 (2): 663. [Бесплатная статья PMC: PMC19570] [PubMed:
Труды Национальной академии наук США. 1997; 94 (2): 663. [Бесплатная статья PMC: PMC19570] [PubMed:41]
- 26.
Андерсон Дж. Биологические реакции на материалы. Ежегодный обзор материаловедения. 2001; 31: 81.
- 27.
Fine N, Mustoe T. Заживление ран. Филадельфия: Липпинкотт Уильямс и Уилкинс; 2006 г.
- 28.
- Мартин П. Заживление ран — стремление к идеальной регенерации кожи. Наука. 1997; 276 (5309): 75. [PubMed:
89]
- 29.
- Ягер Д., Нвомех Б. Протеолитическая среда хронических ран. Регенерация заживления ран. 1999; 7: 433. [PubMed: 10633002]
- 30.
- Diacovo TG, Roth SJ, Buccola JM, Bainton DF, Springer TA. Катание, задержка и трансмиграция нейтрофилов через активированные, приклеенные к поверхности тромбоциты за счет последовательного действия Р-селектина и бета 2-интегрина CD11b / CD18.Кровь. 1996; 88 (1): 146. [PubMed: 8704169]
- 31.
- Springer TA. Сигналы светофора для рециркуляции лимфоцитов и эмиграции лейкоцитов: многоэтапная парадигма.
 Клетка. 1994; 76 (2): 301. [PubMed: 7507411]
Клетка. 1994; 76 (2): 301. [PubMed: 7507411] - 32.
- Мартин П., Лейбович С.Дж. Воспалительные клетки при заживлении ран: хорошее, плохое и уродливое. Тенденции в клеточной биологии. 2005; 15 (11): 599. [PubMed: 16202600]
- 33.
- Шукаляк Дж., Доровини-Зис К. Экспрессия β-хемокинов RANTES и MIP-1β эндотелиальными клетками микрососудов головного мозга человека в первичной культуре.Журнал невропатологии экспериментальной неврологии. 2000; 59 (5): 339. [PubMed: 10888363]
- 34.
- 35.
- 36.
- Hickey W. Основные принципы иммунологического надзора за нормальной центральной нервной системой. Глия. 2001; 36 (2): 118. [PubMed: 11596120]
- 37.
- Перри В.Х., Браун М.С., Гордон С. Реакция макрофагов на повреждение центрального и периферического нерва. Возможная роль макрофагов в регенерации. Журнал экспериментальной медицины.1987; 165 (4): 1218. [Бесплатная статья PMC: PMC2188570] [PubMed: 3559478]
- 38.
- Авеллино А., Харт Д., Дейли А., Маккиннон М., Эллегала Д., Клиот М. Дифференциальный ответ макрофагов в периферической и центральной нервной системе во время Валлеровской дегенерации Аксоны. Экспериментальная неврология. 1995; 136: 183. [PubMed: 7498408]
- 39.
Бисби М. Регенерация аксонов периферической нервной системы. В: Waxman S, Kocsis J, Stys P, редакторы. Аксон: структура, функция и патофизиология.Нью-Йорк: издательство Оксфордского университета; 1995. стр. 553.
- 40.
- Fawcett JW. Глиальный ответ на повреждение и его роль в ингибировании восстановления ЦНС. В: Бахр М., редактор. Ремонт мозга. Vol. 557. Нью-Йорк: Спрингер; [PubMed: 16955702] Наука. 2006: 11.
- 41.
Берри М., Батт А, Логан А. Клеточные реакции на проникающее повреждение ЦНС. В: Берри М., Логан А., редакторы. Повреждения ЦНС: клеточные реакции и фармакологические стратегии. Бока-Ратон, Флорида: CRC Press; 1999 г.п. 1.
- 42.
Griffin J, George E, Hsieh S, Glass J. Дегенерация аксонов и нарушения аксонального цитоскелета. В: Waxman S, Kocsis J, Stys P, редакторы. Аксон: структура, функция и патофизиология. Нью-Йорк: издательство Оксфордского университета; 1995. стр. 375.
- 43.
- Ford AL, Goodsall AL, Hickey WF, Sedgwick JD. Нормальная взрослая разветвленная микроглия, отделенная от других макрофагов центральной нервной системы с помощью проточной цитометрии. Определены фенотипические различия и проведено сравнение прямой презентации антигена ex vivo на Т-лимфоцитах CD4 +, реагирующих с основным белком миелина.Журнал иммунологии. 1995; 154 (9): 4309. [PubMed: 7722289]
- 44.
Streit W. Клетки микроглии. В: Кеттенманн Х., Рэнсом Б., редакторы. Нейроглия. Оксфорд, Великобритания: Издательство Оксфордского университета; 2005.
- 45.
- Олсон Дж. К., Миллер С. Д.. Микроглия инициирует врожденные и адаптивные иммунные ответы центральной нервной системы через несколько TLR. Журнал иммунологии. 2004; 173 (6): 3916. [PubMed: 15356140]
- 46.
- Hayes G, Woodroofe M, Cuzner M. Микроглия является основным типом клеток, экспрессирующих MHC класса II в белом веществе человека.Журнал неврологических наук. 1987; 80: 25. [PubMed: 3302117]
- 47.
- Фудзита Т., Йошимин Т., Маруно М., Хаякава Т. Клеточная динамика макрофагов и микроглиальных клеток в ответ на колотые ранения в коре головного мозга крысы. Acta Neurochirurgica. 1998; 140 (3): 275. [PubMed: 9638265]
- 48.
- Fitch MT, Doller C, Combs CK, Landreth GE, Silver J. Клеточные и молекулярные механизмы глиального рубцевания и прогрессирующей кавитации: анализ in vivo и in vitro вторичного повреждения, вызванного воспалением, после Травма ЦНС.Журнал неврологии. 1999; 19 (19): 8182. [Бесплатная статья PMC: PMC6783021] [PubMed: 10493720]
- 49.
- Бэбкок А.А., Кузил В.А., Ривест С., Оуэнс Т. Экспрессия хемокинов глиальными клетками направляет лейкоциты к участкам повреждения аксонов в ЦНС. Журнал неврологии. 2003; 23 (21): 7922. [Бесплатная статья PMC: PMC6740601] [PubMed: 12944523]
- 50.
- Джулиан Д., Ли Дж., Ли Х, Джордж Дж., Рутеки П. Влияние цитокинов, полученных из микроглии, на глиоз в ЦНС. Развитие нейробиологии.1994. 16 (3-4): 128. [PubMed: 7535679]
- 51.
- Джулиан Д., Ли Дж., Леара Б., Кинен С. Фагоцитарная микроглия выделяет цитокины и цитотоксины, которые регулируют выживание астроцитов и нейронов в культуре. Neurochemistry International. 1994; 25 (3): 227. [PubMed: 7833791]
- 52.
- Woodroofe M, Sarna G, Wadhwa M, Hayes G, Loughlin A, Tinker A, Cuzner M. Обнаружение интерлейкина-1 и интерлейкина-6 в головном мозге взрослых крыс после механического повреждения с помощью микродиализа in vivo — свидетельство роли микроглии в продукции цитокинов.Журнал Нейроиммунология. 1991; 33 (3): 227. [PubMed: 1874973]
- 53.
- Zielasek J, Muller B, Hartung H-P. Ингибирование цитокин-индуцируемой синтазы оксида азота в микроглии крыс и макрофагах мышей метил-2,5-дигидроксициннаматом. Neurochemistry International. 1996; 29 (1): 83. [PubMed: 8808792]
- 54.
- 55.
- Morgan SC, Taylor DL, Pocock JM. Микроглия высвобождает активаторы нейрональной пролиферации, опосредованные активацией митоген-активируемой протеинкиназы, фосфатидилинозитол-3-киназы / Akt и сигнальных каскадов дельта-Notch.Журнал нейрохимии. 2004; 90 (1): 89. [PubMed: 15198670]
- 56.
- Polazzi E, Gianni T., Contestabile A. Клетки микроглии защищают нейроны гранул мозжечка от апоптоза: доказательства реципрокной передачи сигналов. Глия. 2001; 36 (3): 271. [PubMed: 11746765]
- 57.
- Elkabes S, DiCicco-Bloom EM, Black IB. Микроглия / макрофаги мозга экспрессируют нейротрофины, которые выборочно регулируют пролиферацию и функцию микроглии. Журнал неврологии. 1996; 16 (8): 2508. [Бесплатная статья PMC: PMC6578768] [PubMed: 8786427]
- 58.
- Накадзима К., Хонда С., Тохьяма Ю., Имаи Ю., Косака С., Курихара Т. Секреция нейротрофинов из культивированной микроглии. Журнал неврологических исследований. 2001; 65 (4): 322. [PubMed: 11494368]
- 59.
- Mucke L, Oldstone M, Morris J, Nerenberg M. Быстрая активация астроцит-специфической экспрессии трансгена GFAP-lacZ путем очагового повреждения. Новая биология. 1991; 3 (5): 465. [PubMed: 1883812]
- 60.
Норенберг М. Реактивный астроцит. В: Ашнер М., Коста Л., редакторы.Роль глии в нейротоксичности. Бока-Ратон, Флорида: CRC Press; 2005. с. 73.
- 61.
- Джордж Р., Гриффин Дж. У. Задержанные реакции макрофагов и клиренс миелина во время валлеровской дегенерации в центральной нервной системе: модель дорсальной радикулотомии. Экспериментальная неврология. 1994; 129 (2): 225. [PubMed: 7957737]
- 62.
- Niklason LE, Abbott W, Gao J, Klagges B, Hirschi KK, Ulubayram K, Conroy N, Jones R, Vasanawala A, Sanzgiri S, Langer R. Морфологические и механические характеристики спроектированных бычьи артерии.Журнал сосудистой хирургии. 2001; 33 (3): 628. [PubMed: 11241137]
- 63.
- Вольбург Х., Вольбург-Бухгольц К., Энгельгардт Б. Диапедез мононуклеарных клеток через венулы головного мозга во время экспериментального аутоиммунного энцефаломиелита оставляет плотные соединения неповрежденными. Acta Neuropathologica. 2005; 109 (2): 181. [PubMed: 15549331]
- 64.
- Мясник Э., Уильямс М., Янгман К., Ротт Л., Брискин М. Торговля лимфоцитами и региональный иммунитет. Успехи иммунологии. 1999; 72: 209.[PubMed: 10361577]
- 65.
- Bauer J, Bradl M, Hickey WF, Forss-Petter S, Breitschopf H, Linington C, Wekerle H, Lassmann H. Апоптоз Т-клеток при воспалительных поражениях головного мозга: разрушение Т-клеток не зависит от распознавания антигена. Американский журнал патологии. 1998; 153 (3): 715. [Бесплатная статья PMC: PMC1853020] [PubMed: 9736022]
- 66.
Лоддик С., Ротвелл Н. Цитокины и нейродегенерация. В: Ротвелл Н., Лоддик С., редакторы. Иммунные и воспалительные реакции в нервной системе.Оксфорд, Великобритания: Издательство Оксфордского университета; 2002. с. 90.
- 67.
- Henson PM. Смягчение воспаления. Иммунология природы. 2005; 6 (12): 1179. [PubMed: 16369556]
- 68.
- Витте М., Абарбул А. Общие принципы заживления ран. Хирургические клиники Северной Америки. 1997; 3: 509. [PubMed: 78]
- 69.
Mehendale F, Martin P. Клеточные и молекулярные процессы заживления ран. В: Фаланга V, редактор. Заживление кожных ран. Лондон: Мартин Дуниц; 2001 г.
- 70.
- Saarialho-Kere UK, Chang ES, Welgus HG, Parks WC. Четкая локализация коллагеназы и тканевого ингибитора экспрессии металлопротеиназ при заживлении ран, связанных с язвенной пиогенной гранулемой. Журнал клинических исследований. 1992; 90 (5): 1952–1957. [Бесплатная статья PMC: PMC443257] [PubMed: 1430217]
- 71.
Могфорд Дж., Мустоу Т. Экспериментальные модели заживления ран. В: Фаланга V, редактор. Заживление кожных ран. Лондон: Мартин Дуниц; 2001 г.п. 109.
- 72.
Сюй Дж., Кларк Р. Регулирование интегрина при заживлении ран. В: Гарг Х., Лонгакер М., редакторы. Исцеление без рубцов. Нью-Йорк: Марсель Деккер; 2000.
- 73.
- Romer J, Bugge T, Pyke C, Lund L, Flick M, Degen J, Dano K. Нарушение заживления ран у мышей с нарушенным геном плазминогена. Природная медицина. 1996; 2 (3): 287. [PubMed: 8612226]
- 74.
- Гриннелл Ф. Заживление ран, активация кератиноцитов и модуляция интегрина.Журнал клеточной науки. 1992; 101 (1): 1. [PubMed: 1569118]
- 75.
- Matoltsy A, Viziam C. Дальнейшие наблюдения эпителизации небольших ран: авторадиографическое исследование включения и распределения 3H-тимидина в эпителии, покрывающем кожные раны. Журнал следственной дерматологии. 1970; 55 (1): 20. [PubMed: 5425061]
- 76.
Sfeir C, Ho L, Doll B, Azari K, Hollinger J. Ремонт перелома. В: Либерман Дж., Фридлендер Дж., Редакторы. Восстановление и восстановление костей: биология и клиническое применение.Тотова, Нью-Джерси: Humana Press; 2005. с. 21.
- 77.
Дельмас П., Малавал Л. Белки костей. В: Манди Дж., Редактор. Физиология и фармакология костей. Нью-Йорк: Спрингер-Верлаг; 1993. с. 673.
- 78.
- Мачта Б. Заживление ран в других тканях. Хирургические клиники Северной Америки. 1997; 77 (3): 529. [PubMed: 79]
- 79.
- Chao E, Aro H, Lewallen D, Kelly P. Влияние жесткости на заживление переломов при внешней фиксации.Клиническая ортопедия и смежные исследования. 1989; 241: 24. [PubMed: 2647334]
- 80.
- Роблинг А., Кастильо А., Тернер С. Биомеханическое и молекулярное регулирование ремоделирования кости. Ежегодный обзор биомедицинской инженерии. 2006; 8: 455. [PubMed: 16834564]
- 81.
- МакХью К.П., Ходивала-Дилке К., Чжэн М.Х., Намба Н., Лам Дж., Новак Д., Фенг Х, Росс Ф.П., Хайнс Р.О., Тейтельбаум С.Л. Мыши, лишенные интегринов β3, остеосклеротичны из-за дисфункциональных остеокластов. Журнал клинических исследований.2000; 105 (4): 433. [Бесплатная статья PMC: PMC289172] [PubMed: 10683372]
- 82.
- Teitelbaum SL. Резорбция кости остеокластами. Наука. 2000; 289 (5484): 1504. [PubMed: 10968780]
- 83.
Ли К., Эйнхорн Т. Система костных органов: форма и функция. В: Маркус Р., редактор. Остеопороз. Vol. 1. Сан-Диего, Калифорния: Academic Press; 2001.
- 84.
- МакКиббин Б. Биология заживления переломов длинных костей. Журнал костной и совместной хирургии.1978; 60В: 150. [PubMed: 350882]
- 85.
- Ли С., Вулф С. Повреждение и восстановление периферических нервов. Журнал Американской академии ортопедической хирургии. 2000; 8 (4): 243. [PubMed: 10951113]
- 86.
- Schmidt CE, Leach JB. Инженерия нервных тканей: стратегии восстановления и регенерации. Ежегодный обзор биомедицинской инженерии. 2003; 5 (1): 293. [PubMed: 14527315]
- 87.
Сандерленд С. Нервные травмы и их восстановление: критическая оценка.Нью-Йорк: Черчилль Ливингстон; 1991.
- 88.
- Frostick S, Yin Q, Kemp G. Шванновские клетки, нейротрофические факторы и регенерация периферических нервов. Микрохирургия. 1998; 18: 397. [PubMed: 9880154]
- 89.
- Додд Дж., Джессел Т. Руководство аксона и формирование паттерна нейрональных проекций у позвоночных. Наука. 1988; 242: 692. [PubMed: 3055291]
- 90.
- 91.
- Grabham PW, Foley M, Umeojiako A, Goldberg DJ. Фактор роста нервов стимулирует связывание интегрина бета1 с различными транспортными механизмами в филоподиях конусов роста.Журнал клеточной науки. 2000; 113 (17): 3003. [PubMed: 10934039]
- 92.
- Гриннелл Ф., Биллингем Р., Берджесс Л. Распределение фибронектина во время заживления ран in vivo. Журнал следственной дерматологии. 1981; 76 (3): 181. [PubMed: 7240787]
- 93.
Eckes B, Aumailley M, Kreig T. Collagens и восстановление целостности дермы. В: Кларк Р., редактор. Молекулярная и клеточная биология заживления ран. Нью-Йорк: Пленум Пресс; 1996. с. 493.
- 94.
- Певец AJ, Кларк РАФ. Заживление кожных ран. Медицинский журнал Новой Англии. 1999; 341 (10): 738. [PubMed: 10471461]
- 95.
- Chen WYJ, Abatangelo G. Функции гиалуронана при заживлении ран. Ремонт и регенерация ран. 1999; 7 (2): 79. [PubMed: 10231509]
- 96.
Doll B, Sfeir C, Azari K, Holland S, Hollinger J. Черепно-лицевой ремонт. В: Либерман Дж., Фридлендер Дж., Редакторы. Восстановление и восстановление костей: биология и клиническое применение.Тотова, Нью-Джерси: Humana Press; 2005. с. 337.
- 97.
- Nakahara H, Bruder SP, Haynesworth SE, Holecek JJ, Baber MA, Goldberg VM, Caplan AI. Формирование костей и хрящей в диффузионных камерах субкультивированными клетками, происходящими из надкостницы. Кость. 1990; 11 (3): 181. [PubMed: 23]
- 98.
Wornom I., Buchman S. Кости и хрящевые ткани. В: Коэн К., Дигельманн Р., Линдглад В., редакторы. Заживление ран: биохимические и клинические аспекты. Филадельфия: У. Б. Сондерс; 1992 г.п. 356.
- 99.
- Kon T, Cho T, Aizawa T., Yamazaki M, Nooh N, Graves D., Gerstenfeld L, Einhorn T. Экспрессия остеопротегерина, активатора рецептора лиганда NF-kappaB (лиганда остеопротегерина) и родственного ему провоспалительные цитокины при заживлении переломов. Журнал исследований костных минералов. 2001; 16 (6): 1004. [PubMed: 11393777]
- 100.
Баквалтер Дж., Эйнхорн Т., Марш Дж. Заживление костей и суставов. В: Бухольц Р., Хекман Дж., Редакторы. Переломы Роквуда и Грина у взрослых.Vol. 1. Филадельфия: Липпинкотт Уильямс и Уилкинс; 2001.
- 101.
Пойнтон А., Лейн Дж. Заживление костей и отказ. В: Petite H, Quarto R, редакторы. Искусственная кость. Джорджтаун, Техас: Landes Bioscience; 2005.
- 102.
Miclau T, Schneider R, Eames B, Helms J. Общие молекулярные механизмы, регулирующие формирование костной ткани плода и восстановление переломов у взрослых. В: Либерман Дж., Фридлендер Дж., Редакторы. Восстановление и восстановление костей: биология и клиническое применение.Тотова, Нью-Джерси: Humana Press; 2005. с. 45.
- 103.
- Чжао Би-Кью, Ван С., Ким Х-И, Сторри Х, Розен Б.Р., Муни Ди-джей, Ван Х, Ло Э. Роль матриксных металлопротеиназ в задержке кортикальных реакций после инсульта. Природная медицина. 2006; 12 (4): 441. [PubMed: 16565723]
- 104.
- Сильвер Дж., Миллер Дж. Х. Регенерация за пределами глиального шрама. Обзоры природы Неврология. 2004; 5 (2): 146. [PubMed: 14735117]
- 105.
- Clark R, Lee E, Fish C, White R, Price W, Jonak Z, Feuerstein G, Barone F.Развитие повреждения тканей, воспаление и разрешение после инсульта: иммуногистохимическое и количественное планиметрическое исследование. Бюллетень исследований мозга. 1993; 31: 565. [PubMed: 8495380]
- 106.
Фрош М., Энтони Д., ДеГиролами В. Центральная нервная система. В: Кумар В., Аббас А., Фаусто Н., редакторы. Патологические основы болезни Роббинса и Котрана. Филадельфия: Эльзевьер; 2005.
- 107.
- Fawcett JW, Asher RA. Восстановление глиального рубца и центральной нервной системы.Бюллетень исследований мозга. 1999; 49 (6): 377. [PubMed: 10483914]
- 108.
Берри М., Логан А. Преобразование бета-фактора роста и рубцевание ЦНС. В: Берри М., Логан А., редакторы. Повреждения ЦНС: клеточные реакции и фармакологические стратегии. Бока-Ратон, Флорида: CRC Press; 1999.
- 109.
- Faulkner JR, Herrmann JE, Woo MJ, Tansey KE, Doan NB, Sofroniew MV. Реактивные астроциты защищают ткань и сохраняют функцию после травмы спинного мозга. Журнал неврологии.2004; 24 (9): 2143. [Бесплатная статья PMC: PMC6730429] [PubMed: 14999065]
- 110.
Zurn A, Bandtlow C. Нарушение регенерации в ЦНС: клеточные и молекулярные механизмы. В: Бахр М., редактор. Ремонт мозга. Vol. 557. Нью-Йорк: Спрингер; Science: 2006.
- 111.
- Fawcett JW. Преодоление торможения в поврежденном спинном мозге. Журнал нейротравмы. 2006. 23 (3-4): 371. [PubMed: 16629623]
- 112.
- МакКеон Р.Дж., Юринек М.Дж., Бак С.Р. Хондроитинсульфат протеогликаны нейрокан и фосфакан экспрессируются реактивными астроцитами в хроническом глиальном рубце ЦНС.Журнал неврологии. 1999; 19 (24): 10778. [Бесплатная статья PMC: PMC6784959] [PubMed: 10594061]
- 113.
- Davies SJA, Goucher DR, Doller C, Silver J. Надежная регенерация сенсорных аксонов взрослых в дегенерирующем белом веществе спинного мозга взрослых крыс. Журнал неврологии. 1999; 19 (14): 5810. [Бесплатная статья PMC: PMC6783087] [PubMed: 10407022]
- 114.
- Дэвис Ю., Льюис Д., Фуллвуд Н.Дж., Недушинский И.А., Марцинюк Б., Альбон Дж., Талло А. Протеогликаны на нормальном и мигрирующем эндотелии роговицы человека.Экспериментальные исследования глаза. 1999; 68 (3): 303. [PubMed: 10079138]
- 115.
- Sandvig A, Berry M, Barrett L, Butt A, Logan A. Миелиновые, реактивные глии и происходящие из рубца ингибиторы роста аксонов ЦНС: экспрессия, передача сигналов рецептора и корреляция с регенерация аксонов. Глия. 2004; 46 (3): 225. [PubMed: 15048847]
- 116.
- Дэвид С., Агуайо А. Удлинение аксонов в «мосты» PN после повреждения центральной нервной системы у взрослых крыс. Наука. 1981; 214: 931. [PubMed: 6171034]
- 117.
- Davies SJA, Field PM, Raisman G. Регенерация разрезанных взрослых аксонов не удается даже при наличии непрерывных выровненных глиальных путей. Экспериментальная неврология. 1996; 142 (2): 203. [PubMed: 8934554]
- 118.
- Лагорд С., Берри М., Логан А. Экспрессия TGFβ2, но не TGFβ1 коррелирует с отложением рубцовой ткани в пораженном спинном мозге. Молекулярная и клеточная неврология. 2002; 20 (1): 69. [PubMed: 12056841]
- 119.
- Ашер Р.А., Моргенштерн Д.А., Фидлер П.С., Адкок К.Х., Охира А., Брейстед Дж. Э., Левин Дж. М., Марголис РУ, Роджерс Дж. Х., Фосетт Дж. У.Нейрокан активируется в поврежденном мозге и в астроцитах, обработанных цитокинами. Журнал неврологии. 2000; 20 (7): 2427. [Бесплатная статья PMC: PMC6772249] [PubMed: 10729323]
- 120.
- Логан А., Грин Дж., Хантер А., Джексон Р., Берри М. Ингибирование глиальных рубцов в поврежденном мозге крысы с помощью рекомбинантных человеческих моноклональных антител к трансформирующим фактор роста-бета2. Европейский журнал нейробиологии. 1999; 11 (7): 2367. [PubMed: 10383626]
- 121.
Фергюсон М., Ли И.Лечение раны. В: Чемпион Р., редактор. Rook / Wilkinson / Ebling Учебник дерматологии. Vol. 1. Оксфорд, Великобритания: Blackwell Science; 1998. с. 337.
- 122.
Wolff J. Das Gestz der Transformation der Knochen. Берлин: Хиршвальд; 1892.
- 123.
- Парфитт А. Остеональное и гемиостеональное ремоделирование: пространственная и временная основа для передачи сигналов в кости взрослого человека. Журнал клеточной биохимии. 1994; 55: 273. [PubMed: 7962158]
- 124.
- Бернетт М., Загер Э. Патофизиология повреждения периферических нервов: краткий обзор. Нейрохирургия. 2004; 16 (5): 1. [PubMed: 15174821]
- 125.
- Bellamkonda RV. Регенерация периферических нервов: мнение о каналах, каркасах и анизотропии. Биоматериалы. 2006; 27 (19): 3515. [PubMed: 16533522]
- 126.
- Эрлих Х., Раджаратнам Дж. Силы локомоции клеток в сравнении с силами сокращения клеток для сокращения коллагеновой решетки: модель in vitro сокращения раны.Ткань и клетка. 1990; 22 (4): 407. [PubMed: 2260082]
- 127.
- Томасек Дж. Дж., Габбиани Дж., Хинц Б., Шапонье С., Браун Р. А.. Миофибробласты и механорегуляция ремоделирования соединительной ткани. Обзоры природы Молекулярная клеточная биология. 2002; 3 (5): 349. [PubMed: 11988769]
- 128.
- Gong MC, Iizuka K, Nixon G, Browne JP, Hall A, Eccleston JF, Sugai M, Kobayashi S, Somlyo AV, Somlyo AP. Роль гуаниновых нуклеотид-связывающих белков — белков семейства ras или тримерных белков или обоих — в сенсибилизации гладких мышц Ca2 +.Труды Национальной академии наук США. 1996; 93 (3): 1340. [Бесплатная статья PMC: PMC40082] [PubMed: 8577766]
- 129.
- Hinz B, Mastrangelo D, Iselin CE, Chaponnier C, Gabbiani G. Механическое напряжение контролирует сократительную активность грануляционной ткани и дифференциацию миофибробластов. Американский журнал патологии. 2001; 159 (3): 1009. [Бесплатная статья PMC: PMC1850455] [PubMed: 11549593]
- 130.
- Desmouliere A, Redard M, Darby I., Gabbiani G. Апоптоз опосредует уменьшение клеточности во время перехода между грануляционной тканью и рубцом.Американский журнал патологии. 1995; 1: 56. [Бесплатная статья PMC: PMC1870783] [PubMed: 7856739]
- 131.
- Jee WSS, Mori S, Li XJ, Chan S. Простагландин E2 увеличивает массу кортикальной кости и активирует внутрикортикальное ремоделирование кости у интактных и подвергшихся овариэктомии самок крыс. Кость. 1990; 11 (4): 253. [PubMed: 2242291]
- 132.
- Fan X, Roy E, Zhu L, Murphy TC, Ackert-Bicknell C, Hart CM, Rosen C, Nanes MS, Rubin J. Оксид азота регулирует рецепторный активатор ядерного фактора — { экспрессия лиганда каппа} B и остеопротегерина в стромальных клетках костного мозга.Эндокринология. 2004; 145 (2): 751. [PubMed: 14563699]
- 133.
- Spataro L, Dilgen J, Retterer S, Spence AJ, Isaacson M, Turner JN, Shain W. Лечение дексаметазоном снижает реакцию астроглии на вставленные нейропротезные устройства в неокортекс крыс. Экспериментальная неврология. 2005; 194 (2): 289. [PubMed: 16022859]
- 134.
- Linkins L, Weitz J. Новая антикоагулянтная терапия. Ежегодный обзор медицины. 2005; 56: 63. [PubMed: 15660502]
- 135.
- МакГроу Дж., Хиберт Дж., Стивс Дж.Модулирующий астроглиоз после нейротравмы. Журнал неврологических исследований. 2001; 63 (2): 109. [PubMed: 11169620]
- 136.
- Софронев М.В. Реактивные астроциты в восстановлении и защите нейронов. Невролог. 2005; 11 (5): 400. [PubMed: 16151042]
Ткани человека — обзор
5.31.5.2 Европейские директивы и законодательство
В Европе Европейская комиссия разработала дополнительные нормативные документы посредством набора директив и законодательства. В Англии и Уэльсе Закон о тканях человека 2004 г. сделал согласие ключевым элементом легального изъятия, хранения и использования ткани или клеток от живых доноров (см. Ссылку 12 и ссылки в ней).Европейские стандарты использования клеток пуповинной крови и тканей для применения человеком подробно описаны в Директивах Европейского Союза по тканям и клеткам (2004/23 / EC, 2006/17 / EC и 2006/86 / EC), которые были включены в законодательство Великобритании через Положение о человеческих тканях (качество и безопасность для человека) 2007 г. С 5 июля 2008 г. эти Правила по качеству и безопасности теперь регулируют закупку, тестирование, обработку, хранение, распространение и импорт или экспорт клеток и тканей UCB для применения на людях в Соединенное Королевство (http: // www.hta.gov.uk). Они направлены на обеспечение качества и безопасности человеческих тканей и клеток (включая ОКБ), используемых для лечения, с момента закупки до трансплантации. Эти правила применяются к установкам UCB с минимальным манипулированием. Европейские директивы и законодательство контролируются в отдельных европейских странах компетентным органом, которым в Соединенном Королевстве является Управление по тканям человека (HTA). Банк пуповинной крови NHS в Англии, например, имеет лицензию на сбор, хранение и распространение UCB в соответствии с настоящими постановлениями и директивами Европейской комиссии через этот компетентный орган [12].В Соединенном Королевстве незаконно собирать и хранить UCB для использования людьми вне рамок лицензирования HTA (http://www.hta.gov.uk). Проверки ОТЗ на соответствие Правилам качества и безопасности в настоящее время проводятся с интервалом в 2 года, при этом требуются отчеты о соблюдении требований дважды в год и годовые отчеты о деятельности. Для тестирования, если оно не проводится самим банком пуповинной крови, должны быть аккредитованы сторонние микробиологические и испытательные лаборатории (например, в Соединенном Королевстве это аккредитованные лаборатории по клинической патологии (CPA) для микробиологии и мазков крови и CPA / EFI. -аккредитованные лаборатории для HLA-типирования).Кроме того, нормативные требования могут периодически меняться или обновляться (http://www.cpa-uk.co.uk; http://www.efiweb.org). Например, поскольку некоторые практики, связанные с частными закупками UCB в Соединенном Королевстве, были сочтены неуместными, HTA недавно выпустило новые руководящие принципы, гарантирующие, что лицо, собирающее UCB, является соответствующим образом подготовленным профессионалом и должно действовать на основании лицензии HTA или при необходимости, посредством стороннего соглашения с учреждением, имеющим лицензию HTA.Ключевыми вопросами, связанными с этими новыми руководящими принципами ОМТ, являются: (1) безопасность, сводящая к минимуму риск для матери и ребенка; (2) наилучшее качество собранных образцов UCB, также обеспечивающее надежное отслеживание; (3) информированное согласие матери на сбор UCB с объяснением рисков, преимуществ и вариантов; и (4) деятельность должна быть законной, то есть сбор, тестирование, обработка, хранение, распространение или импорт / экспорт UCB должны осуществляться на основании лицензии HTA или, где это уместно, соглашения с третьей стороной. с учреждением, имеющим лицензию HTA.В Соединенном Королевстве обязательно проверять материнскую кровь на вирусы, такие как ВИЧ, HBV, HCV и HTLV, если она находится в зоне повышенного риска, и на сифилис в течение 7 дней после родов, а также проверять блок UCB. путем тестирования нуклеиновых кислот на ВИЧ, ВГВ и ВГС во время банковского обслуживания.
Описанные правила HTA регулируют использование устройств UCB с минимальным управлением. Если блок UCB подвергается существенным манипуляциям (например, с помощью генной терапии, тканевой инженерии или терапии соматических клеток), эти продукты будут классифицироваться как лекарственные препараты для современной терапии (ATMP) для использования человеком и регулируются Регламентом (EC) No.1394/2007, введенный в декабре 2008 г. для ATMP (http://www.ema.europa.eu; [электронная почта защищена]; http://www.ema.europa.eu/SME/SMEapplication.htm; http: // www.ema.europa.eu/htms/human/advanced_therapies/certification.htm). Европейским органом, контролирующим это регулирование, является Европейское агентство по оценке лекарственных средств (EMEA), а компетентным органом Великобритании является Агентство по регулированию лекарственных средств и товаров медицинского назначения (MHRA). Конкретные цели европейского регулирования в этой области для всех ATMP, которые включают более индивидуализированную терапию, разработку наномедицины и регенеративную медицину и имеют отношение к UCB, где этим существенно манипулируют, являются:
- •
, чтобы гарантировать наивысший уровень защиты пациентов и безопасности в этой области;
- •
для облегчения доступа на рынок Европейского союза (ЕС) этих продуктов и новых методов лечения;
- •
для обеспечения беспрепятственного перемещения лекарств в пределах ЕС; и
- •
для повышения конкурентоспособности европейских фармацевтических компаний.
Ключевые регулирующие элементы включают:
- •
создание нового экспертного комитета Европейского медицинского агентства по передовой терапии или CAT в регионе EMEA для оценки ATMP и отслеживания соответствующих научных разработок;
- •
технические требования к банкоматам;
- •
централизованная европейская процедура разрешения на продажу; и
- •
специальные льготы для малых и средних предприятий.
Научные разработки для персонализированных лекарств также будут включать новые генетические технологии, такие как фармакогеномика и другие омики, и они, вероятно, также повлияют на банкинг и производство UCB для использования человеком в будущем.
Поскольку в Великобритании есть ряд регулирующих агентств или компетентных органов, охватывающих использование биологических материалов для применения человеком, британский инструментарий стволовых клеток был разработан при поддержке регулирующих органов, таких как HTA, MHRA и Консультативный комитет по генной терапии. , ученых и клиницистов, чтобы предоставить единый ресурс для разработки программы производства и исследования терапевтических клеток (http: // www.sc-toolkit.ac.uk/home.cfm).
Основные типы клеток — Неукротимая наука
Четыре основных типа клеток
Несмотря на то, что в организме существует несколько сотен типов клеток, все они могут быть сгруппированы всего в четыре основные категории, или ткани. Это облегчает их понимание.
Эти четыре основные ткани образованы из:
- Эпителиальные клетки. Эти клетки плотно связаны друг с другом. Они покрывают внутреннюю часть полых органов, таких как кровеносные сосуды или органы пищеварения, или же образуют поверхность вещей, например кожу.Есть десятки типов эпителиальных клеток. Без эпителиальных клеток у вас не было бы кожи, защищающей тело от травм, и не было бы желудка, чтобы переваривать пищу!
- нервных клеток. Эти клетки предназначены для общения. Они посылают сигналы от мозга к мышцам и железам, которые контролируют их функции. Они также получают сенсорную информацию от кожи, глаз и ушей и отправляют эту информацию в мозг. В организме существуют десятки разновидностей нервных клеток, каждая из которых имеет свою форму и функции.Без нервных клеток у вас не было бы сознания или контроля над своим телом.
- мышечных клеток. Эти клетки специализируются на сокращении. Без мышечных клеток вы не смогли бы двигаться! Есть три вида мышечных клеток. Они тянут и тянут кости и сухожилия, чтобы произвести движение. Они также образуют толстые внешние стенки полых органов, таких как кровеносные сосуды и органы пищеварения, и могут сокращаться, регулируя диаметр этих полых органов.
- Клетки соединительной ткани.Эти клетки обеспечивают структурную прочность тела, а также защищают от чужеродных захватчиков, таких как бактерии. Два типа клеток — фибробласты и жировые клетки — присущи соединительной ткани. Другие клетки мигрируют в соединительную ткань из кровотока для борьбы с болезнями. Особые типы соединительной ткани — хрящ и кость — более прочны и жестки, чем большинство соединительных тканей.
Классификация тканей — соединительная, мышечная, нервная и эпителиальная ткани
Классификация тканей
Ткани образуют промежуточную организационную структуру между клетками и системами органов.Различные группы тканей с различными функциями группируются вместе, образуя орган. Таким образом, у животных группа мышечных клеток объединяется, образуя мышечную ткань.
Классификация тканей животных подразделяется на четыре основные категории. Это:
Соединительные ткани.
Мышечные ткани.
Нервные ткани.
Эпителиальные ткани.
Прежде чем вы узнаете больше о функциях тканей, вот вам викторина:
Pop Quiz 1:
Изучение тканей известно как ________.
Анатомия растений.
Морфология.
Гистология.
Естествознание.
Соединительные ткани
В соединительной ткани группа клеток, разделенных неживым внеклеточным матриксом, объединяется в ткань. Соединительные ткани помогают придать органам механическую прочность, форму и жесткость, а также удерживать их на месте. Примеры соединительной ткани включают сухожилия, связки, кости, кровь, жировую ткань и ареолярную ткань.Далее, существует три типа соединительной ткани. Это:
Соединительные ткани выполняют несколько различных функций в организме человека, а именно:
Они обеспечивают изоляцию и защиту тела от холода.
Они помогают связывать органы вместе и обеспечивают им механическую поддержку.
Соединительные ткани помогают транспортировать газы, воду, гормоны, питательные вещества и отходы в организме.
Мышечная ткань
Мышечные ткани помогают нам выполнять различные действия и участвовать в таких действиях, как ходьба, бег и передвижение.Есть три типа мышечной ткани:
Во-первых, мышечные ткани помогают нам поддерживать осанку. Во-вторых, они также помогают в наших непроизвольных и произвольных движениях. Например, сердечные мышцы помогают перекачивать кровь и регулировать ее кровоток в артериях и капиллярах.
Нервная ткань
Нервные ткани присутствуют в центральной нервной системе, которая включает головной и спинной мозг. В периферической нервной системе нервные ткани составляют черепные и спинномозговые нервы.
Нервные ткани выполняют несколько жизненно важных функций, таких как:
Они контролируют и координируют метаболические процессы в организме.
Они помогают передавать информацию в теле.
Они помогают поддерживать стабильность и помогают лучше понять окружающую нас среду.
Они также помогают нам реагировать на внешние раздражители.
Эпителиальная ткань
Эпителиальные ткани состоят из клеток, которые образуют внешнее покрытие тела.Они покрывают поверхности органов, такие как кожа, дыхательные пути, внутренние стенки пищеварительного тракта, а также репродуктивный тракт.
Эпителиальные ткани обеспечивают поддержку и механическую прочность клеток и тканей. Они также помогают в транспортировке материалов путем диффузии, секреции и фильтрации.
Более того, эпителиальные ткани, такие как те, что находятся под нашей кожей, помогают нам в сенсорном восприятии и защищают нас от патогенов и физических травм. Они также выделяют гормоны, слизь и ферменты из различных эндокринных органов.
Теперь, когда вы прошли классификацию тканей, пришло время пересмотреть несколько концепций с помощью следующего упражнения.
Pop Quiz 2
Какие бывают типы тканей?
Соединительные ткани.
Нервная ткань.
Эпителиальная ткань.
Мышечная ткань.
Все вышеперечисленное.
Вот таблица классификации тканей, которая поможет вам лучше понять.
(изображение будет загружено в ближайшее время)
Узнайте больше о темах, связанных с тканями и типами тканей, на наших живых занятиях в приложении Vedantu. Загрузите учебные материалы из нашего диапазона заметок о пересмотрах и образцов работ и сделайте обучение увлекательным.
7.4 Ткани — Биология человека
Создано Christine Miller
Рисунок 7.4.1 Строительство. Для работы важно иметь подходящие материалы.
Строительство дома — большая работа, и для ее выполнения требуется много разных материалов.Как вы можете видеть на рис. 7.4.1, для строительства дома используется множество различных типов материалов, но каждый тип материала выполняет определенные функции. Вы не будете использовать изоляцию, чтобы покрыть крышу, и вы не будете использовать пиломатериалы для прокладки проводов в доме. Подобно тому, как строитель выбирает подходящие материалы для строительства каждого аспекта дома (провода для электричества, пиломатериалы для каркаса, черепица для кровли), ваше тело использует правильные ячейки для каждого типа роли. Когда многие клетки работают вместе для выполнения определенной функции, это называется тканью.
Группы связанных клеток образуют ткани. Все клетки в ткани могут быть одного или нескольких типов. В любом случае клетки ткани работают вместе, чтобы выполнять определенную функцию, и они всегда специализированы, чтобы выполнять эту функцию лучше, чем любой другой тип ткани. Существует четыре основных типа тканей человека: соединительная, эпителиальная, мышечная и нервная. Мы используем ткани для построения органов и систем органов. 200 типов клеток, которые организм может производить на основе нашего единого набора ДНК, могут создавать все типы тканей в организме.
Эпителиальная ткань состоит из клеток, выстилающих внутреннюю и внешнюю поверхности тела, такие как кожа и внутренняя поверхность пищеварительного тракта. Эпителиальная ткань, выстилающая внутренние поверхности тела и отверстия тела, называется слизистой оболочкой . Эпителиальная ткань этого типа продуцирует слизь , слизистое вещество, которое покрывает слизистые оболочки и улавливает патогены, частицы и мусор. Эпителиальная ткань защищает тело и его внутренние органы, помимо слизи выделяет вещества (например, гормоны) и поглощает вещества (например, питательные вещества).
Ключевым отличительным признаком эпителиальной ткани является то, что она содержит свободную поверхность и базальную мембрану. Свободная поверхность не прикреплена к каким-либо другим клеткам и либо открыта снаружи тела, либо открыта внутрь полого органа или трубки тела. Базальная мембрана прикрепляет эпителиальную ткань к нижележащим клеткам.
Эпителиальная ткань идентифицируется и получает название по форме и слою. Эпителиальные клетки существуют в трех основных формах: плоском, кубовидном и столбчатом.Эти клетки особой формы, в зависимости от функции, могут быть разделены на несколько слоев: простой, стратифицированный, псевдостратифицированный и переходный.
Эпителиальная ткань образует покрытия и подкладки и отвечает за ряд функций, включая диффузию, абсорбцию, секрецию и защиту. Форма эпителиальной клетки может максимизировать ее способность выполнять определенную функцию. Чем тоньше эпителиальная клетка, тем легче веществам проходить через нее для диффузии и / или абсорбции.Чем больше эпителиальная клетка, тем больше места в ее цитоплазме для выработки продуктов для секреции, и тем большую защиту она может обеспечить нижележащим тканям. У них есть три основных формы эпителиальных клеток: плоскоклеточный (имеющий форму плоского и овального блинчика), кубовидный (кубовидный) и столбчатый (высокий и прямоугольный).
Рис. 7.4.2 Форма эпителиальных тканей важна.
Эпителиальная ткань также может быть разделена на разные слои в зависимости от их функции.Например, несколько слоев ячеек обеспечивают отличную защиту, но больше не будут эффективны для диффузии, тогда как один слой будет очень хорошо работать для диффузии, но уже не будет такой защитной; Для растягивающихся органов, таких как мочевой пузырь, требуется специальный тип наслоения, называемый переходным. Ваши ткани обладают многослойной структурой, которая делает их наиболее эффективными для выполнения той функции, которую они должны выполнять. В эпителиальной ткани обнаруживаются четыре основных слоя: простой (один слой клеток), стратифицированный (много слоев клеток), псевдостратифицированный (кажется многослойным, но при более близком рассмотрении на самом деле простой) и переходный (может растягиваться, переходя из многих слоев. к меньшему количеству слоев).
Рис. 7.4.3. Слои, обнаруженные в эпителиальных тканях, имеют большое значение.
См. Таблицу 7.4.1 для обзора различных типов слоев и форм, которые могут образовываться эпителиальными клетками, а также их связанных функций и местоположений.
Таблица 7.4.1
Сводка по клеткам эпителиальной ткани
На данный момент мы идентифицировали эпителиальную ткань по форме и слою. Представительные диаграммы, которые мы видели до сих пор, полезны для визуализации тканевых структур, но важно смотреть на реальные примеры этих клеток.Поскольку клетки слишком малы, чтобы их можно было увидеть невооруженным глазом, мы полагаемся на микроскопы, которые помогут нам изучить их. Гистология — это исследование микроскопической анатомии, клеток и тканей. См. Таблицу 7.4.2, чтобы увидеть некоторые примеры слайдов эпителиальных тканей, подготовленных для гистологии.
Таблица 7.4.2
Эпителиальные ткани и гистологические образцы
Кость и кровь являются примерами соединительной ткани. Соединительная ткань очень разнообразна.В целом, он образует каркас и опорную структуру для тканей и органов тела. Он состоит из живых клеток, разделенных неживым материалом, называемым внеклеточным матриксом, который может быть твердым или жидким. Например, внеклеточный матрикс кости — это жесткий минеральный каркас. Внеклеточный матрикс крови — жидкая плазма.
Ключевой отличительной чертой соединительной ткани является то, что она состоит из множества клеток в неклеточном матриксе. В зависимости от природы матрикса существует три основных категории соединительной ткани.Они сильно отличаются друг от друга, что отражает их разные функции:
- Волокнистая соединительная ткань: характеризуется гибкой матрицей, состоящей из белковых волокон, включая коллаген, эластин и, возможно, ретикулярные волокна. Эти ткани составляют сухожилия, связки и оболочки тела.
- Поддерживающая соединительная ткань: характеризуется твердым матриксом и используется для образования костей и хрящей. Эти ткани используются для поддержки и защиты.
- Жидкая соединительная ткань: характеризуется жидким матриксом и включает кровь и лимфу.
Волокнистая соединительная ткань
Волокнистая соединительная ткань содержит клетки, называемые фибробластами. Эти клетки производят волокна коллагена, эластина или ретикулярные волокна, которые составляют матрицу соединительной ткани этого типа. В зависимости от того, насколько плотно упакованы эти волокна и как они ориентированы, меняются свойства и, следовательно, функция волокнистой соединительной ткани.
- Рыхлая волокнистая соединительная ткань: состоит из рыхлого и беспорядочного переплетения коллагеновых и эластиновых волокон, образуя ткань, которая является тонкой и гибкой, но все же жесткой. Эта ткань, которую также иногда называют «ареолярной тканью», находится в мембранах и окружающих кровеносных сосудах, а также в большинстве органов тела. Как вы можете видеть из диаграммы на рисунке 7.4.4, рыхлая волокнистая соединительная ткань соответствует определению соединительной ткани, поскольку это рассеяние клеток (фибробластов) в неклеточном матриксе (сетка из коллагеновых и эластиновых волокон).Существует два типа специализированной рыхлой волокнистой соединительной ткани: ретикулярная и жировая. Жировая ткань хранит жир, а ретикулярная ткань образует селезенку и лимфатические узлы.
Рис. 7.4.4. Схема рыхлой волокнистой соединительной ткани состоит из рассеяния фибробластов в неклеточном матриксе из рыхлых волокон коллагена и эластина. Рис. 7.4.5. Микроскопический вид рыхлой волокнистой соединительной ткани. - Плотная волокнистая соединительная ткань Ткань: состоит из плотного мата из параллельных коллагеновых волокон и рассеянных фибробластов, образующих очень прочную ткань.Плотная волокнистая соединительная ткань образует сухожилия и связки, которые соединяют кости с мышцами и / или кости с соседними костями.
Рис. 7.4.6 Плотная волокнистая соединительная ткань состоит из фибробластов и плотной параллельной упаковки коллагеновых волокон. Рис. 7.4.7. Микроскопический вид плотной волокнистой соединительной ткани.
Поддерживающая соединительная ткань
Поддерживающая соединительная ткань демонстрирует определяющую особенность соединительной ткани в том, что она представляет собой разброс клеток в неклеточном матриксе; что отличает его от других соединительных тканей, так это твердый матрикс.В этой группе тканей матрикс твердый — кость или хрящ. В то время как волокнистая соединительная ткань содержала клетки, называемые фибробластами, которые производили волокна, поддерживающая соединительная ткань содержит клетки, которые либо образуют кость (остеоциты), либо клетки, которые образуют хрящ (хондроциты).
Хрящ
Хондроциты производят хрящевой матрикс, в котором они находятся. Хрящ состоит из белковых волокон и хондроцитов в лакунах. Это прочная, но гибкая ткань, которая используется во многих частях тела для защиты и поддержки.Хрящ — одна из немногих тканей, которые не являются сосудистыми (не имеют прямого кровоснабжения), а это означает, что для получения питательных веществ и газов требуется диффузия; это причина медленного заживления повреждений хряща. Выделяют три основных типа хрящей:
- Гиалиновый хрящ : гладкая, прочная и гибкая ткань. Находится на концах ребер и длинных костей, в носу и составляет весь скелет плода.
- Фиброхрящ : очень прочная ткань, содержащая толстые пучки коллагена.Встречается в суставах, которые нуждаются в амортизации при сильных ударах (колени, челюсть).
- Эластичный хрящ : помимо коллагена содержит эластичные волокна, обеспечивающие поддержку и эластичность. Встречается в мочках ушей и надгортаннике.
Рис. 7.4.8 Три типа хряща, каждый из которых имеет свои характеристики в зависимости от природы матрикса.
Кость
Остеоциты производят костный матрикс, в котором они находятся. Поскольку кость очень твердая, эти клетки располагаются в небольших пространствах, называемых лакунами.Эта костная ткань состоит из волокон коллагена, встроенных в фосфат кальция, что придает ей прочность без хрупкости. Кость бывает двух типов: плотная и губчатая.
- Компактная кость: имеет плотный матрикс, организованный в цилиндрические единицы, называемые остеонами. Каждый остеон содержит центральный канал (иногда называемый каналом Харверса), который позволяет разместить кровеносные сосуды и нервы, а также концентрические кольца костного матрикса и остеоцитов в лакунах, как показано на схеме.Компактная кость находится в длинных костях и образует оболочку вокруг губчатой кости.
- Губчатая кость: очень пористая кость, которая чаще всего содержит костный мозг. Он находится на конце длинных костей и составляет большую часть ребер, лопаток и плоских костей черепа.
Жидкая соединительная ткань
Жидкая соединительная ткань имеет жидкий матрикс; в отличие от двух других категорий соединительной ткани, клетки, которые находятся в матрице, на самом деле не производят матрицу. Фибробласты образуют волокнистый матрикс, хондроциты образуют хрящевой матрикс, остеоциты создают костный матрикс, но клетки крови не образуют жидкий матрикс лимфы или плазмы.Эта ткань по-прежнему соответствует определению соединительной ткани в том смысле, что это все еще рассеяние клеток в неклеточном матриксе.
Есть два типа жидкой соединительной ткани:
- Кровь: Кровь содержит три типа клеток, взвешенных в плазме, и содержится в сердечно-сосудистой системе.
- Эритроциты, чаще называемые эритроцитами, присутствуют в большом количестве (примерно 5 миллионов клеток на мл) и отвечают за доставку кислорода из легких во все другие части тела.Эти клетки относительно небольшие по размеру, около 7 микрометров в диаметре, и живут не более 120 дней.
- Лейкоциты, часто называемые лейкоцитами, присутствуют в меньшем количестве (примерно 5 тысяч клеток на мл) и отвечают за различные иммунные функции. Обычно они больше, чем эритроциты, но могут жить намного дольше, особенно белые кровяные тельца, отвечающие за долговременный иммунитет. Количество лейкоцитов в крови может увеличиваться или уменьшаться в зависимости от того, боретесь ли вы с инфекцией.
- Тромбоциты, также известные как тромбоциты, представляют собой очень маленькие клетки, отвечающие за свертывание крови. На самом деле тромбоциты — это не настоящие клетки, это фрагменты гораздо более крупной клетки, называемой мегакариоцитом.
- Лимфа: содержит жидкий матрикс и белые кровяные тельца и содержится в лимфатической системе, которая в конечном итоге стекает в сердечно-сосудистую систему.
Рис. 7.4.11. Окрашенный лимфоцит, окруженный эритроцитами, просматриваемый под световым микроскопом.
Мышечная ткань состоит из клеток, которые обладают уникальной способностью сокращаться, что является определяющей особенностью мышечной ткани. Как показано на рис. 7.4.12, существует три основных типа мышечной ткани: скелетная, гладкая и сердечная мышечные ткани.
Скелетная мышца
Скелетные мышцы — это произвольные мышцы, что означает, что вы осуществляете над ними сознательный контроль. Скелетные мышцы прикрепляются к костям с помощью сухожилий — разновидности соединительной ткани.Когда эти мышцы укорачиваются, чтобы тянуть за кости, к которым они прикреплены, они позволяют телу двигаться. Когда вы занимаетесь спортом, читаете книгу или готовите ужин, вы используете скелетные мышцы, чтобы двигать своим телом для выполнения этих задач.
Под микроскопом скелетные мышцы имеют поперечнополосатый (или полосатый) вид из-за их внутренней структуры, которая содержит чередующиеся белковые волокна актина и миозина. Скелетные мышцы описываются как многоядерные, что означает, что одна «клетка» имеет много ядер.Это происходит потому, что в утробе матери отдельные клетки, которым суждено стать скелетными мышцами, сливаются, образуя мышечные волокна в процессе, известном как миогенез. Вы узнаете больше о скелетных мышцах и о том, как они сокращаются в мышечной системе.
Рис. 7.4.12 Скелетная мышца поперечно-полосатая и многоядерная.Гладкие мышцы
Гладкие мышцы — это гладкие мышцы — они все еще содержат актин и миозин мышечных волокон, но не в таком же чередующемся расположении, как в скелетных мышцах.Гладкая мускулатура находится в трубках тела — в стенках кровеносных сосудов, в репродуктивных, желудочно-кишечных и дыхательных путях. Гладкие мышцы не находятся под произвольным контролем, что означает, что они действуют бессознательно через вегетативную нервную систему. Гладкие мышцы перемещают вещества посредством волны сокращения, которая спускается вниз по длине трубки, этот процесс называется перистальтикой .
Посмотрите видео на YouTube «Что такое перистальтика» от Mister Science, чтобы увидеть перистальтику в действии.
Что такое перистальтика, Mister Science, 2018.
Рис. 7.4.13. Гладкая мышца не имеет поперечно-полосатой части, и каждая клетка овальной формы содержит одно ядро. (Микрофотография предоставлена Медицинской школой Риджентс Мичиганского университета © 2012) Рис. 7.4.14 Перистальтика — это волнообразное сокращение гладкой мускулатуры, которое выталкивает содержимое трубки перед волной сокращения.Сердечная мышца
Сердечные мышцы работают непроизвольно, то есть регулируются вегетативной нервной системой.Вероятно, это хорошо, так как вам не нужно сознательно концентрироваться на том, чтобы ваше сердце постоянно билось! Сердечная мышца, которая находится только в сердце, является одноядерной и поперечно-полосатой (из-за чередования полос миозина и актина). Их сокращения заставляют сердце перекачивать кровь. Чтобы обеспечить синхронное сокращение всех участков сердца, ткань сердечной мышцы содержит специальные клеточные соединения, называемые вставными дисками, которые проводят электрические сигналы, используемые, чтобы «сообщать» камерам сердца, когда нужно сокращаться.
Нервная ткань
Нервная ткань состоит из нейронов и группы клеток, называемых нейроглией (также известной как глиальные клетки). Нервная ткань составляет центральную нервную систему (в основном головной и спинной мозг) и периферическую нервную систему (сеть нервов, которая проходит по всему остальному телу).Отличительной чертой нервной ткани является то, что она специализируется на генерации и проведении нервных импульсов. Эту функцию выполняют нейроны, а цель нейроглии — поддерживать нейроны.
Структура нейрона состоит из нескольких частей:
- Дендриты, улавливающие входящие нервные импульсы
- Тело клетки или сома, которая содержит большинство органелл нейрона, включая ядро
- Аксон, который переносит нервные импульсы от сомы к следующему нейрону в цепи
- Миелиновая оболочка, которая окружает аксон и увеличивает скорость проведения нервных импульсов
- Терминалы аксонов, которые поддерживают физический контакт с дендритами соседних нейронов
Neuroglia можно рассматривать как вспомогательный персонал для нейрона. Нейроны выполняют такую важную работу, что им нужны клетки, чтобы доставлять им питательные вещества, удалять клеточные отходы и строить свою милеиновую оболочку. Существует много типов нейроглии, которые классифицируются в зависимости от их функции и / или расположения в нервной системе. Нейроглии в 50 раз больше, чем нейроны, и они намного меньше. См. Схему в 7.4.17, чтобы сравнить размер и количество нейронов и нейроглии.
- Ткани состоят из взаимодействующих клеток.
- Существует четыре основных типа тканей: эпителиальная, соединительная, мышечная и нервная.
- Эпителиальная ткань составляет выстилку и покровы тела и характеризуется наличием свободной поверхности и базальной мембраны.Типы эпителиальной ткани различаются по форме клеток (плоскоклеточная, кубовидная или столбчатая) и по слоистости (простая, стратифицированная, псевдостратифицированная и переходная). Различные эпителиальные ткани могут осуществлять диффузию, секрецию, абсорбцию и / или защиту в зависимости от их конкретной формы клеток и их слоев.
- Соединительная ткань обеспечивает структуру и опору для тела и характеризуется разбросом клеток в неклеточном матриксе. Существует три основных категории соединительной ткани, каждая из которых характеризуется определенным типом матрикса:
- Волокнистая соединительная ткань содержит белковые волокна.К этой категории относятся как рыхлая, так и плотная волокнистая соединительная ткань.
- Поддерживающая соединительная ткань содержит очень прочный матрикс и включает в себя как кость, так и хрящ.
- Жидкая соединительная ткань содержит клетки в жидком матриксе с двумя типами крови и лимфы.
- Отличительной чертой мышечной ткани является ее сократимость. Существует три типа мышечной ткани: скелетная мышца, которая прикреплена к скелету для произвольного движения, гладкая мышца, которая перемещает вещества по трубкам тела, и сердечная мышца, которая перемещает кровь через сердце.
- Нервная ткань содержит специализированные клетки, называемые нейронами, которые могут проводить электрические импульсы. В нервной ткани также содержится нейроглия, которая поддерживает нейроны, обеспечивая питательными веществами, удаляя отходы и создавая миелиновую оболочку.
- Определите термин ткань.
- Если бы части тела требовалась подкладка, которая была бы одновременно защитной, но все же способной поглощать питательные вещества, какой тип эпителиальной ткани лучше всего использовать?
- Где вы найдете скелетные мышцы? Гладкая мышца? Сердечная мышца?
- Каковы некоторые функции нейроглии?
Типы тканей человеческого тела, MoomooMath and Science, 2017.
Как напечатать человеческую ткань на 3D-принтере — Танека Джонс, TED-Ed, 2019.
Как кости производят кровь — Мелоди Смит, TED-Ed, 2020.
Атрибуции
Рисунок 7.4.1
Рисунок 7.4.2 и Рисунок 7.4.3
Рисунок 7.4.4
Рыхлая волокнистая соединительная ткань по CNX OpenStax. Биология. на Викискладе используется под лицензией CC BY 4.0. (https: // creativecommons.org / licenses / by / 4.0) лицензия.
Рисунок 7.4.5
Connective Tissue: Loose Aerolar от Berkshire Community College Bioscience Image Library на Flickr используется в соответствии с лицензией CC0 1.0, предназначенной для универсального общественного достояния (https://creativecommons.org/publicdomain/zero/1.0/).
Рисунок 7.4.6
Плотная волокнистая соединительная ткань от CNX OpenStax. Биология. на Викискладе используется под лицензией CC BY 4.0. (https://creativecommons.org/licenses/by/4.0) лицензия.
Рисунок 7.4.7
Dense_connective_tissue-400x от J Jana на Wikimedia Commons используется под лицензией CC BY-SA 4.0 (https://creativecommons.org/licenses/by-sa/4.0/deed.en).
Рисунок 7.4.8
Types_of_Cartilage-new от OpenStax College в Wikipedia Commons используется под лицензией CC BY 3.0 (https://creativecommons.org/licenses/by/3.0).
Рисунок 7.4.9
Compact_bone_histology_2014 пользователя Athikhun.suw на Викискладе используется по лицензии CC BY-SA 4.0 (https://creativecommons.org/licenses/by-sa/4.0/deed.en).
Рисунок 7.4.10
Bone_normal_and_degraded_micro_structure от Gtirouflet на Wikimedia Commons используется под лицензией CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0/deed.en).
Рисунок 7.4.11
Lymphocyte2, созданный Николасом Гранджином на Wikimedia Commons, используется в соответствии с CC BY-SA 3.0 (https: // creativecommons.org / licenses / by-sa / 3.0 / deed.en) лицензия. [Машиночитаемый автор не предоставлен. Предполагается, что Николас Гранджан на основании заявлений об авторском праве.]
Рисунок 7.4.12
Skeletal_muscle_ 横纹肌 1 от 乌拉 跨 氪 на Wikimedia Commons используется под лицензией CC BY-SA 4.0 (https://creativecommons.org/licenses/by-sa/4.0/deed.en).
Рисунок 7.4.13
Smooth_Muscle_new от OpenStax на Wikimedia Commons используется в соответствии с CC BY 4.0 (https://creativecommons.org/licenses/by/4.0 / deed.en) лицензия.
Рисунок 7.4.14
Peristalsis от OpenStax на Викискладе используется по лицензии CC BY 3.0 (https://creativecommons.org/licenses/by/3.0/deed.en).
Рисунок 7.4.15
400x Cardiac Muscle от Jessy731 на Flickr используется и адаптирована Кристин Миллер под лицензией CC BY-NC 2.0 (https://creativecommons.org/licenses/by-nc/2.0/).
Рисунок 7.4.16
Neuron.svg пользователя: Dhp1080 на Wikimedia Commons используется в соответствии с CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0/deed.en) лицензии.
Рисунок 7.4.17
400x Nervous Tissue от Jessy731 на Flickr используется под лицензией CC BY-NC 2.0 (https://creativecommons.org/licenses/by-nc/2.0/).
Таблица 7.4.1
Summary of Epithelial Tissue Cells, разработанное OpenStax College в Wikipedia Commons, используется под лицензией CC BY 3.0 (https://creativecommons.org/licenses/by/3.0).
Таблица 7.4.2
Список литературы
Беттс, Дж.Дж., Янг, К.А., Уайз, Дж. А., Джонсон, Э., По, Б., Круз, Д. Х., Корол, О., Джонсон, Дж. Э., Уомбл, М., ДеСе, П. (25 апреля 2013 г.). Рисунок 4.8. Сводка клеток эпителиальной ткани [цифровое изображение]. В Анатомия и физиология (Раздел 4.2). OpenStax. https://openstax.org/books/anatomy-and-physiology/pages/4-2-epithelial-tissue
Беттс, Дж. Г., Янг, К. А., Уайз, Дж. А., Джонсон, Э., По, Б., Круз, Д. Х., Корол, О., Джонсон, Д. Э., Уомбл, М., Де Сикс, П. (2013, 25 апреля ). Рисунок 4.16 Типы хрящей [цифровое изображение]. В Анатомия и физиология (Раздел 4.3). OpenStax. https://openstax.org/books/anatomy-and-physiology/pages/4-3-connective-tissue-supports-and-protects
Беттс, Дж. Г., Янг, К. А., Уайз, Дж. А., Джонсон, Э., По, Б., Круз, Д. Х., Корол, О., Джонсон, Д. Э., Уомбл, М., Де Сикс, П. (2013, 25 апреля ). Рис. 10.23. Гладкая мышца [цифровая микрофотография]. В Анатомия и физиология (Раздел 10.8). OpenStax. https://openstax.org/books/anatomy-and-physiology/pages/10-8-smooth-muscle (Микрофотография предоставлена Риджентсами Медицинской школы Мичиганского университета © 2012)
Беттс, Дж.Дж., Янг, К.А., Уайз, Дж. А., Джонсон, Э., По, Б., Круз, Д. Х., Корол, О., Джонсон, Дж. Э., Уомбл, М., ДеСе, П. (25 апреля 2013 г.). Рис. 22.5. Псевдостратифицированный мерцательный столбчатый эпителий [цифровая микрофотография]. В Анатомия и физиология (Раздел 22.1). OpenStax. https://openstax.org/books/anatomy-and-physiology/pages/22-1-organs-and-structures-of-the-respiratory-system
Беттс, Дж. Г., Янг, К. А., Уайз, Дж. А., Джонсон, Э., По, Б., Круз, Д. Х., Король, О., Джонсон, Дж. Э., Уомбл, М., ДеСе, П. (25 апреля 2013 г.). Рисунок 23.5 Перистальтика [диаграмма]. В Анатомия и физиология (Раздел 23.2). OpenStax. https://openstax.org/books/anatomy-and-physiology/pages/23-2-digestive-system-processes-and-regulation
Мистер Наука. (2018). Что такое перистальтика? YouTube. https://www.youtube.com/watch?v=kVjeNZA5pi4
MoomooMath and Science. (2017, 18 мая). Типы тканей человеческого тела. YouTube. https://www.youtube.com/watch?v=O0ZvbPak4ck&feature=youtu.быть
Open Stax. (2016, 27 мая). Рис. 6. Рыхлая соединительная ткань [цифровое изображение]. В OpenStax Biology (Раздел 33.2). OpenStax CNX. https://cnx.org/contents/[email protected]:-LfhWRES@4/Animal-Primary-Tissues
Open Stax. (2016, 27 мая). Рис. 7. Волокнистая соединительная ткань сухожилия [цифровое изображение]. В OpenStax Biology (Раздел 33.2). OpenStax CNX. https://cnx.org/contents/[email protected]:-LfhWRES@4/Animal-Primary-Tissues
TED-Ed. (2019, 17 октября).Как напечатать человеческую ткань на 3D-принтере — Танека Джонс.