
Книгопечатание Иван Федоров Презентация Ученицы 6 класса Б
Книгопечатание. Иван Федоров. Презентация Ученицы 6 класса «Б» Шевчик Варвары
Термины Выводы До книгопеча тания Иван Федоров СОДЕРЖАНИЕ Начало книгопеча тания на Руси Наше время Шаг Вперед
До книгопечатания. К половине XV века русские земли объединились вокруг Москвы. Русское государство не только окрепло политически, но и быстро развивалось в культурном отношении. В связи с общим подъемом русской национальной культуры увеличился спрос на книгу. В то время книги на Руси были только в рукописном виде. Переписывание книг считалось занятием почетным. К концу XV – началу XVI веков искусство рукописания настолько развилось, что кроме писцов, трудившихся при монастырях, образовался целый класс переписчиков. Книги все больше и больше возрастали в цене.
Иван Федоров родился 1510 -1530 годах, точного сведений о дате и месте рождения нет.
Начало книгопечатание на Руси В 1553 году по приказу Ивана IV в Москве был построен Печатный двор. Как мы знаем первая книга Ивана Федорова «Апостол» 1563 -1564. На следующий год в типографии Фёдорова вышла его вторая книга, «Часовник» . Через некоторое время начались нападки на печатников со стороны переписчиков, обвиняющей Федорова в волшебстве. После поджога, уничтожившего их мастерскую , Фёдоров со Мстиславцем уехали в Великое княжество Литовское. Там их радушно принял гетман Ходкевич, который основал типографию в своём имении Заблудове. Однако Иван Федоров продолжал писать для Руси.
Шаг вперед Таким образам мы видим что Иван Федоров сделал огромный прорыв для развития культуры Руси. Был изобретен печатный станок. Была построена первая типография.
Наше время В наше время было разработана удобные способы печатание. Теперь мы используем компьютеры если хотим что-то написать. Книгопечатание началось использоваться не только для священного и исторического писания, но и для написание книг для всеобщего чтения. Ноутбук и его удобная клавиатура.
Теперь книги существует разных жанров. Книги читают все. Учебники, газеты, журналы и просто книжки все это печатное издание. Мы можем задуматься на сколько Иван Федоров улучшил нашу жизнь и облегчил ее.
ВЫВОДЫ Развитие книгопечатание на Руси. Изобретена первая печатная машинка. Создана первая на Руси типография. Иван Федоров сделал огромный вклад в развитие культуры Руси. Иван Федоров улучшил и облегчил нашу жизнь.
Термины Энциклопедия-справочник, содержащий сведения по широкому кругу знаний. Книга-сшитые в один переплет листы бумаги, на котором написан текст. Книгопечатание-печатное издание книг. Типография-предприятие, в котором производится набор и печатание книг, газет, журналов и других изданий.
Иван Федоров. Книгопечатание — История и исторические личности презентация – Telegraph
Главная
История и исторические личности
Иван Федоров. Книгопечатание
Начало книгопечатания в России и на Украине. Обучение Ивана Федорова в университете в г. Кракове. Создание первой московской печатни. Выход в свет первой московской книги. Первое напечатанное кириллическим шрифтом издание Библии. Начало печати «Азбуки».
посмотреть текст работы
скачать работу можно здесь
полная информация о работе
весь список подобных работ
Нужна помощь с учёбой? Наши эксперты готовы помочь!
Нажимая на кнопку, вы соглашаетесь с
политикой обработки персональных данных
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
HTML-версии работы пока нет. Cкачать архив работы можно перейдя по ссылке, которая находятся ниже.
Причины образования единого русского государства и возвышения Москвы. Начало московской династии. Иван Данилович Калита и Дмитрий Иванович Донской. Значение Куликовской битвы. Феодальная война 1433-1453 гг. Начало правления Ивана IV Васильевича Грозного. презентация [1,7 M], добавлен 10.04.2014
Письмо и письменность в древности. Книжное и печатное дело в Средние века. Начало книгопечатания и печатного дела в Европе (XV-XVI века). Начало славянского книгопечатания и печатного дела в русском государстве. Современное полиграфическое производство. реферат [5,7 M], добавлен 31.05.2014
Психологический портрет Ивана Грозного. Внутренняя и внешняя политика. Реформы, направленные на дальнейшее развитие и укрепление Русского централизованного государства. Приговор о местничестве. Борьба за выход к Балтийскому морю. Начало освоения Сибири. реферат [45,9 K], добавлен 07.04.2016
Начало правления Ивана Грозного. Торжественное венчание великого князя Ивана IV. Реформы по централизации государства, преобразования в армии. Сыновья и жены Ивана Грозного. Присоединение Казани и Астрахани. Ливонская война. Наследие Ивана Грозного. презентация [1,1 M], добавлен 21.12.2011
Торжественное венчание великого князя Ивана IV. Реформы по централизации государства, преобразования в армии. Сыновья и жены Ивана Грозного. Присоединение Казани и Астрахани. Ливонская война. Наследие Ивана Грозного. презентация [1,1 M], добавлен 21.12.2011
Процесс объединения раздробленных русских земель. Начало правление Ивана Грозного. Венчание на царство. Правление при «Избранной раде» и ее падение. Война со Швецией. Начало Ливонской войны. Период опричнины. Последние годы правления Ивана Грозного. контрольная работа [54,3 K], добавлен 09.10.2014
Иван IV (Грозный) – первый Российский царь. Реформы середины XVI в. Формирование сословно-представительской монархии. Опричнина, ее причины и последствия. Внешняя политика Ивана IV. Внешняя политика Ивана Грозного на восточном и западном направлениях. контрольная работа [25,9 K], добавлен 23.04.2007
Начало правления Ивана. Венчание на царство. Пожар и восстание в Москве. Реформы Избранной рады. Россия в середине XVI в. Государственно-политический строй России. Падение Избранной рады. Опричнина. Безумный самодержец. Смерть Ивана Грозного. реферат [38,9 K], добавлен 15.01.2003
Падение Избранной рады. Опричнина. Безумный самодержец. Смерть Ивана Грозного. реферат [38,9 K], добавлен 15.01.2003
Работы в архивах красиво оформлены согласно требованиям ВУЗов и содержат рисунки, диаграммы, формулы и т.д. PPT, PPTX и PDF-файлы представлены только в архивах. Рекомендуем скачать работу .
© 2000 — 2021
Иван Федоров. Книгопечатание презентация. История и исторические личности.
Реферат: Главные требования к этике управления
Реферат по теме Логические законы
Қуаныш Сәтбаев Роман Эссе
Сахарный Диабет Курсовая Работа Введение
Дипломная работа: Финансово-правовые основы деятельности государственных внебюджетных фондов в РФ
Дневник По Практике Механика
Реферат На Тему Древние Славяне
Сочинение Рассуждение На Тему Бездушие 9 Класс
Сочинение 11
Дипломная работа по теме Особливості інноваційної стратегії телекомунікаційної компанії на прикладі міжнародної корпорації ‘Вимпелком’
Курсовая работа по теме База даних студії веб-дизайну
Управление Организационными Управлениями Курсовая Работа
Правильное Оформление Курсовой Работы По Госту Пример
Дипломная работа по теме Лікування цукрового діабету з використанням хризантеми
Реферат: Иверский монастырь, как историко-архитектурная ценность России
Дипломные Темы Для Программистов
Реферат по теме Социологическое исследование различных аспектов трудовой занятости молодежи
Оптимизация Денежных Расходов Курсовая
Реферат На Тему Право На Отдых
Реферат: Основы теории цепей
Учет расчетов с использованием векселей — Бухгалтерский учет и аудит курсовая работа
Реализация федерального закона N44-ФЗ в части закупки лекарственных средств — Государство и право курсовая работа
Убийство — Государство и право курсовая работа
Презентація на тему «Діячі культури козацької доби.
 Федоров Іван»
Федоров Іван»Про матеріал
Обличчя історії України формують особистості. Це ті видатні історичні постаті, які залишили свій важливий внесок в історії і стали гордістю України. Дана презентація може бути використана на уроках з історії України в 8 класі під час вивчення історії України, на заняттях з факультативу «Видатні постаті України», на заняттях історичного гуртка та в позакласній роботі з історії. Перегляд файлуЗміст слайдів
Номер слайду 1
Український «Гуттенберг» – Іван Федоров
Номер слайду 2
Відомо, що раніше люди писали книжки від руки. Написати цілу книжку було складно, тому в давнину книги мали дуже велику цінність. М. М. Антокольский. Нестор – літописець .
Номер слайду 3
Йоган Гуттенберг. З гравюри 16 ст.. Реконструкція друкарського верстата Йогана Гуттенберга. В 15 ст. Йоган Гуттенберг винайшов друкарський верстат. З тих пір книжки в Європі почали друкувати.
Номер слайду 4
Історія розуму представляє дві головні епохи — винахід букв та типографії. .. Микола Карамзін. Видатний східно-слов’янський друкар, продовжувач справи Швайполя Філя та Франциска Скорини, художник-гравер, інженер, ливарник. Організатор чотирьох перших слов’янських кириличних друкарень у Москві, Заблудові, Львові, Дермані, Острозі. Першим почав видавати книги кирилицею. Його можна сміливо зарахувати до справжніх титанів сили думки, пристрасті до своєї справи і цілеспрямованості, глибокого розуміння своєї місії — «духовна сємєна по всєлєнии разсєвати и всєму почину раздавати духовную сию пищу…» (Апостол, післямова, 1574 р.)
.. Микола Карамзін. Видатний східно-слов’янський друкар, продовжувач справи Швайполя Філя та Франциска Скорини, художник-гравер, інженер, ливарник. Організатор чотирьох перших слов’янських кириличних друкарень у Москві, Заблудові, Львові, Дермані, Острозі. Першим почав видавати книги кирилицею. Його можна сміливо зарахувати до справжніх титанів сили думки, пристрасті до своєї справи і цілеспрямованості, глибокого розуміння своєї місії — «духовна сємєна по всєлєнии разсєвати и всєму почину раздавати духовную сию пищу…» (Апостол, післямова, 1574 р.)
Номер слайду 5
Першодрукар Іван Федоров народився приблизно в 1510 році. Навчався в Краківському університеті. Володів латинською, давньогрецькою мовами. В 1532 р. отримав ступінь бакалавра. Був він на всі руки майстер, зробив не тільки друкарський верстат, але і багатоствольну гармату — мортиру – давню попередницю «катюши».
Номер слайду 6
Народився близько 1510, або 1525 року. Існує декілька версій його родового походження, так як в своєму друкарському знакові (власне клеймо) Іван Федоров використовує білоруський шляхецький герб Рогоза.
Номер слайду 7
З 1550 року Іван Федоров — диякон церкви Миколи Гостунського при Московському Кремлі.1551 року московський цар Іван Грозний та російський Стоглавий Собор, спираючись на думку просвітителя Максима Грека мати книги писані «кирилицею», вирішили з метою уникнення помилок при переписуванні в церковних книгах, запровадити друкування книг, заснувавши Московський Печатний Двір. Облаштування типографії було доручене диякону Гостунської Кремлівської церкви Івану Федорову, досвіченому граверу-художнику, палітурнику, переписувачу рукописних книг.
Номер слайду 8
У 1552 році диякон Іван Федоров, під керівництвом датського місіонера-протестанта Місінгейма, який привіз друкарський верстат і літери, та за допомогою підмайстрів Петра Мстиславця й Маруші Нефедьєва започаткували друкарську справу.
Номер слайду 9
Місця діяльності Івана Федорова
Номер слайду 10
Найбільш значимі поліграфічні досягнення Івана Федорова відбулися на Рівненщині (тогочасній Волині) — в культурно-просвітницьких осередках Дермані і Острога. Тут, під егідою князя Костянтина Острозького Федоровим була видана одна з найвеличніших книг всіх наступних епох — Острозька Біблія.
Номер слайду 11
Близько десяти років було витрачено на будівництво та відлиття потрібних шрифтів. Всі ці роки Федоров самотужки створював власну техніку книгодрукування. Відливаючи форми своїх власних літер, він спирався на традиції слов’янської писемності і слов’янської писемної графіки букв.
Номер слайду 12
Шрифти Федорова
Номер слайду 13
Шрифти Федорова І.
Номер слайду 14
17 квітня 1563 року вони почали набір першої своєї книги і 1 березня 1564 року, майже рівно через рік, випустили першу книгу московського друку «Апостол». Це перша друкована російська книга, про яку відомо, хто, де, коли і навіщо її надрукував. Ці відомості містяться в літописі на титульному листі книги та в післямові Івана Федорова.
Це перша друкована російська книга, про яку відомо, хто, де, коли і навіщо її надрукував. Ці відомості містяться в літописі на титульному листі книги та в післямові Івана Федорова.
Номер слайду 15
Книга «Апостол» — 1564 рік
Номер слайду 16
Через рік, в серпні — жовтні 1565 року в Московському Печатному Дворі була видана ще одна знакова книга — «Часословець». До 1565 року Іван Федоров надрукував ще близько 20 літургійних церковно-слов’янських книг. Після смерті московського митрополита Макарія, протектора друкарської справи, почалися конфлікти між конкурентами — переписувачами книг і новозаснованою друкарнею, що переросли в ворожнечу і підпали, які закінчились вигнанням друкарів з Москви.
Номер слайду 17
Друкарський знак Івана Федорова
Номер слайду 18
З 1670 року друкування книг у Заблудові припинилось у зв’язку з тяжкою хворобою Хоткевича та погіршенням його фінансових справ. В нагороду за друкарську працю гетьман дарує Федорову село, де першодрукар мав змогу жити достатньо заможно. Проте він відмовляється від осідлого життя. Бувши людиною віруючою, та вважаючи свою друкарську діяльність апостольським служінням, переїздить у 1572 році до Львова. Там, у 1573 — 1574 роках «друковане занедбане обновил», як сповіщає напис на його могилі у Львові.
Проте він відмовляється від осідлого життя. Бувши людиною віруючою, та вважаючи свою друкарську діяльність апостольським служінням, переїздить у 1572 році до Львова. Там, у 1573 — 1574 роках «друковане занедбане обновил», як сповіщає напис на його могилі у Львові.
Номер слайду 19
В Україні пройшли два періоди життя і діяльності Івана Федорова: львівський — 1572-1574 років та 1582-1583 років, і дермансько-острозький — 1575-1581 років. Будинок друкарні Ставропігійського братства, де були надруковані книги Івана Федорова, Львів
Номер слайду 20
Першодрукар Іван Федоров і князь Василь Острозький
Номер слайду 21
Перша слов’янська азбука. Львів, 1574 рік
Номер слайду 22
Сторінка з «Азбуки» Івана Федорова м. Львів 1574 рік.
Номер слайду 23
«Острозька Біблія»Лебединою піснею Івана Федорова стала «Острозька Біблія» — книга, надрукована в замку князя Костянтина Острозького в 1581 р. , — великий том об’ємом 1256 сторінок. Колосальний на той час тираж, тільки до наших днів збереглося не більше 300 екземплярів.
, — великий том об’ємом 1256 сторінок. Колосальний на той час тираж, тільки до наших днів збереглося не більше 300 екземплярів.
Номер слайду 24
Номер слайду 25
«Апостол» поч. 17 ст. Переписаний з «Апостола» Івана Федоровича 1574 року. Музей Острозької академії. 2007 рік.
Номер слайду 26
Номер слайду 27
Номер слайду 28
Наприкінці весни 1583 року Федоров від’їжджає до Відня, щоб запропонувати австрійському імператору Рудольфу ІІ швидкострільну багатоствольну гармату. Гармату можна було розбирати на п’ятдесят, сто, або й двісті частин і змонтувати для бою за три дні і три ночі. На той час такої гармати світ ще знав. Проте, багато грошей за свій винахід Федоров не здобув. Подальша доля винаходу невідома.
Номер слайду 29
1583-й рік був трагічним і останнім в житті Івана Федорова. Не здобувши достатньо коштів ні у Римі, ні у Відні, він потрапляє у скруту, його майно, друкарське обладнання, книги описують за борги. Не витримавши цього, Федоров захворів і в ніч на 6 грудня 1583 помер на руках свого сина Івана в домі у Підзамче (Львів). Похований у монастирському дворі Святого Онуфрія у Львові.
Не витримавши цього, Федоров захворів і в ніч на 6 грудня 1583 помер на руках свого сина Івана в домі у Підзамче (Львів). Похований у монастирському дворі Святого Онуфрія у Львові.
Номер слайду 30
Могила Івана Федорова На його могилу в Онуфрієвському монастирі поклали плиту з зображенням типографської марки першодрукаря. Навколо вився надпис: «Иоанн Фёдоровия, друкарь Московитин, который своим тщанием печатание небывалое обновил…»
Номер слайду 31
Надгробок Івана Федорова
Номер слайду 32
У Львові, на розі вулиць Краківської та Вірменської, де тепер розташований дитячий майданчик, як свідчать історики, була колись друкарня Івана Федорова.
Номер слайду 33
Львівський музей мистецтва давньої української книги був відкритий в 1976 році в приміщенні колишнього Онуфріївського монастиря і називався «Музей імені Івана Федорова». Перед фасадом музею мистецтва давньої української книги встановлено пам’ятник першодрукареві. Це трифігурна композиція: друкар і два майстри.
Це трифігурна композиція: друкар і два майстри.
Номер слайду 34
5 (15) 1583 р. Іван Федоров помер і був похований у Львові в Святоонуфрієвському монастирі
Номер слайду 35
Пам’ятник Івану Федорову в м. Львові
Номер слайду 36
Номер слайду 37
Номер слайду 38
Пам’ятник Івану Федорову в м. Москва
Номер слайду 39
Номер слайду 40
Номер слайду 41
Музей книги. Києво-печерська лавра
Номер слайду 42
Номер слайду 43
Номер слайду 44
Номер слайду 45
Номер слайду 46
Номер слайду 47
Номер слайду 48
На могилі була покладена плита з написом: “Друкарь книг, перед тем невиданных”. В колишньому монастирі зараз музей І. Федорова. Започаткувавши книгодрукування в Росії і на Україні, І. Федоров сіяв “розумне, добре, вічне”.
В городе Запорожской области на строительство ледовой арены хотят потратить 100 миллионов – Индустриалка
Ледовая арена в Мелитополе появится в центре города рядом с больничными учреждениями и на месте зеленой зоны
В планах на 2021 год у мелитопольских чиновников — добиться создания в городе ледовой арены на 250 зрительских мест. В мэрии надеются, что этот объект войдет в перечень строительств, приуроченных в стране к 30-й годовщине независимости Украины.
В мэрии надеются, что этот объект войдет в перечень строительств, приуроченных в стране к 30-й годовщине независимости Украины.
Свой выбор представители местной власти объясняют повышенным спросом услуг временного катка, открытого этой зимой в центральном парке, а также наличием четырех хоккейных команд, которые не имеют своей тренировочной базы, и вынуждены ездить для оттачивания мастерства аж в Херсон.
— Предварительная стоимость проекта ледовой арены площадью более 1600 квадратных метров, — 100 миллионов гривен, — цитирует городского голову Мелитополя Ивана Федорова сайт мэрии. — Мы уже подали документы в Министерство молодежи и спорта, провели предварительные консультации и рассчитываем получить целевую субвенцию на строительство ледовой арены. Второй источник финансирования — Государственный фонд регионального развития, третий — Фонд социально-экономического развития, также будет участвовать местный бюджет.
Строить ледовую арену в мэрии было решено на проспекте Богдана Хмельницкого — прямо в зеленой зоне перед зданием городской больницы №1 — подразделения многопрофильной больницы интенсивных методов лечения и скорой медицинской помощи, здесь же рядом расположен городской морг, станция переливания крови и онкологический диспансер.
Иван Федоров обращает внимание на тот факт, что благодаря данному расположению ледовой арены в шаговой доступности будут все главные спортивные объекты города — стадион «Спартак» имени Олега Олексенко, водно-спортивный комплекс и ДЮСШ №1.
Мелитопольцы активно обсуждают новость в соцсетях и многие недоумевают, почему именно такое расположение выбрано для ледовой арены. В зеленой зоне, которую присмотрели в мэрии для строительства на сегодня около двух сотен деревьев, а заменить большинство из них, как видно на картинке-презентации, планируется на газон и хвойные кустики. К слову, на том месте, где сейчас в парке расположен временный каток когда-то также было вырублено десятки деревьев. Большинство времени это место просто пустовало, так как попытки оборудовать здесь место для катания на коньках и игру в хоккей на постоянной основе успехом не увенчались.
Читайте также: Очарование зимней Хортицы показал запорожский фотограф — фото
Фото автора и с сайта Мелитопольского горсовета.
Project MUSE — Новые взгляды на раннюю кириллицу
Slavonic and East European Review, 95, 4, 2017 ОБЗОРНОЕ ОПИСАНИЕ Новые взгляды на раннее кириллическое книгопечатание ЧАРЛЬЗ Дж. ГАЛЬПЕРИН Богатырев, Сергей (ред.). Путешествия Ивана Федорова: новые взгляды на раннее кириллическое книгопечатание. Специальный выпуск: Canadian-American Slavic Studies, 51, 2017, 2–3, стр. 173–408 (235 стр.). Иллюстрации. Таблицы. Примечания. В 2014 г. исполнилось 450 лет со дня издания Иваном Федоровым и Петром Мстиславским первой датированной книги, появившейся в Московии (России), Апостола (Деяния и Послания) в 1564 году1 и 440 лет со дня издания Федоровым первой книги, появившейся на Украине. , Букварь 1574 года.В честь этих событий Сергей Богатырев из Университетского колледжа Лондона (UCL) организовал в рамках совместного проекта UCL и Британской библиотеки международную конференцию «Возвращение к наследию Ивана Федорова в Европе раннего Нового времени», которая состоялась 9 мая 2014 г. Британская библиотека, в которой хранятся копии каждой исторической книги, спонсируется Школой славянских и восточноевропейских исследований UCL, Центром региональных языковых исследований Восточной Европы, Европейским институтом и Центром раннего современного обмена. В настоящее время Богатырев отредактировал избранные статьи с этой конференции2 с одним дополнением в виде двойного выпуска журнала Canadian-American Slavic Studies, в котором он был приглашенным редактором.3 Двойной выпуск содержит введение Богатырева, девять основных статей, сгруппированных в три. Чарльз Дж. Гальперин — независимый ученый, проживающий в Блумингтоне, штат Индиана. Хочу выразить искреннюю признательность Сергею Богатыреву за консультации при подготовке данной обзорной статьи. Я один остаюсь ответственным за его ошибки и выводы. 1 В первых книгах, напечатанных в Московии, вероятно, в 1550-х годах, отсутствуют даты или имена печатников. 2 И отредактировано очень хорошо. Я нашел только две опечатки, в «Введении» Богатырева Федоров стал «скитальцем», должен быть «скитальцем» (с.
Британская библиотека, в которой хранятся копии каждой исторической книги, спонсируется Школой славянских и восточноевропейских исследований UCL, Центром региональных языковых исследований Восточной Европы, Европейским институтом и Центром раннего современного обмена. В настоящее время Богатырев отредактировал избранные статьи с этой конференции2 с одним дополнением в виде двойного выпуска журнала Canadian-American Slavic Studies, в котором он был приглашенным редактором.3 Двойной выпуск содержит введение Богатырева, девять основных статей, сгруппированных в три. Чарльз Дж. Гальперин — независимый ученый, проживающий в Блумингтоне, штат Индиана. Хочу выразить искреннюю признательность Сергею Богатыреву за консультации при подготовке данной обзорной статьи. Я один остаюсь ответственным за его ошибки и выводы. 1 В первых книгах, напечатанных в Московии, вероятно, в 1550-х годах, отсутствуют даты или имена печатников. 2 И отредактировано очень хорошо. Я нашел только две опечатки, в «Введении» Богатырева Федоров стал «скитальцем», должен быть «скитальцем» (с. 181), а у Рогачевской «советское экспертное агентство» должно быть «советским экспортным агентством» (с. 386). 3 Название специального выпуска, приведенное выше, появляется в печатной версии, а не в онлайн-версии двойного выпуска. РАННЯЯ КИРИЛЛИЧНАЯ ПЕЧАТЬ 721 раздел, по три статьи в каждом,4 в том числе одна Богатырева, примечание также Богатырева (всего Богатырев написал четверть всего двойного номера)5 и очень полезный «Список соавторов» (стр. 405–08) с биографическими и библиографическими данными о соавторах.Шесть очерков снабжены иллюстрациями, многие полностраничные в цвете (Клеминсон, статья и заметка Богатырева, Ерусалимский, Франклин и Рогачевская). Эта впечатляющая публикация дает представление о большом разнообразии современных исследований появления книгопечатания у восточных славян, особенно у русских и украинцев. из России. Двойной выпуск не содержит двух докладов конференций ученых из России и Великобритании соответственно.7 Кроме того, один россиянин представил для публикации не ту статью, которую он представил в Лондоне.
181), а у Рогачевской «советское экспертное агентство» должно быть «советским экспортным агентством» (с. 386). 3 Название специального выпуска, приведенное выше, появляется в печатной версии, а не в онлайн-версии двойного выпуска. РАННЯЯ КИРИЛЛИЧНАЯ ПЕЧАТЬ 721 раздел, по три статьи в каждом,4 в том числе одна Богатырева, примечание также Богатырева (всего Богатырев написал четверть всего двойного номера)5 и очень полезный «Список соавторов» (стр. 405–08) с биографическими и библиографическими данными о соавторах.Шесть очерков снабжены иллюстрациями, многие полностраничные в цвете (Клеминсон, статья и заметка Богатырева, Ерусалимский, Франклин и Рогачевская). Эта впечатляющая публикация дает представление о большом разнообразии современных исследований появления книгопечатания у восточных славян, особенно у русских и украинцев. из России. Двойной выпуск не содержит двух докладов конференций ученых из России и Великобритании соответственно.7 Кроме того, один россиянин представил для публикации не ту статью, которую он представил в Лондоне. 8 Таким образом, в двойной выпуск вошли статьи четырех ученых из Великобритании (Богатырев, Клеминсон, Франклин, Рогачевская), двух из Украины (Бондарь, Ульяновский) и трое из России (Филюшкин, Усачев, Ерусалимский). Четыре статьи опубликованы на русском языке с аннотациями на английском языке9. В книге Богатриева «Введение: принтер в пути» (стр. 173–183) содержится справочная информация о конференции, представленной выше, а также об Иване Федорове.Федоров, диакон Московского храма святителя Николая Гостунского, начал печатать в Москве вместе с Петром Мстиславцем под покровительством царя Ивана IV и митрополита (глава 4-го отдела). но не в «Оглавлении» онлайн. В обеих версиях они появляются над заголовком в первой статье каждого раздела. 5 Одиннадцатистраничное введение, сорок одна страница статьи и пятистраничное примечание, пятьдесят семь страниц. из 231 страницы (не считая «Списка авторов», который он составил).6 Печать на кириллице началась раньше за пределами Московии или Украины, в Кракове, Праге, Венеции, Черногории и Румынии.
8 Таким образом, в двойной выпуск вошли статьи четырех ученых из Великобритании (Богатырев, Клеминсон, Франклин, Рогачевская), двух из Украины (Бондарь, Ульяновский) и трое из России (Филюшкин, Усачев, Ерусалимский). Четыре статьи опубликованы на русском языке с аннотациями на английском языке9. В книге Богатриева «Введение: принтер в пути» (стр. 173–183) содержится справочная информация о конференции, представленной выше, а также об Иване Федорове.Федоров, диакон Московского храма святителя Николая Гостунского, начал печатать в Москве вместе с Петром Мстиславцем под покровительством царя Ивана IV и митрополита (глава 4-го отдела). но не в «Оглавлении» онлайн. В обеих версиях они появляются над заголовком в первой статье каждого раздела. 5 Одиннадцатистраничное введение, сорок одна страница статьи и пятистраничное примечание, пятьдесят семь страниц. из 231 страницы (не считая «Списка авторов», который он составил).6 Печать на кириллице началась раньше за пределами Московии или Украины, в Кракове, Праге, Венеции, Черногории и Румынии. Ни одна статья не посвящена источнику печатной техники в Москве или не посвящена итальянскому словарю раннемосковского книгопечатания: М.Н.Тихомиров и др. (ред.), У истоков русского книгопечатания. К трехсот-семидесятипятилетию со дня смерти Ивана Федорова…
Ни одна статья не посвящена источнику печатной техники в Москве или не посвящена итальянскому словарю раннемосковского книгопечатания: М.Н.Тихомиров и др. (ред.), У истоков русского книгопечатания. К трехсот-семидесятипятилетию со дня смерти Ивана Федорова…
Россия взаимодействует с миром — NYPL
Эта хронология не является исчерпывающей, а скорее дает ориентиры для история представлена в Russia Engages the World, 14531825 .
| 1453 | Османы захватывают Константинополь, положив конец Византии. Империя. | |
| 145355 | Иоганн Гутенберг издает первую книгу, напечатанную на Западе с использованием подвижный тип. | |
| 1472 | Московский великий князь Иван III (р. 142562) женится на Зои (Софии)
Палеолог, племянница последнего византийского императора. 142562) женится на Зои (Софии)
Палеолог, племянница последнего византийского императора. | |
| 1488 | Бартоломеу Диаш достигает мыса Доброй Надежды и открывает морской путь в Индию. | |
| 1492 | Христофор Колумб достигает Карибского моря от имени Королевства Кастилии, подготовив почву для колониальной экспансии и эксплуатации Европы. в Африке и Америке. | |
| 1502 | Шах Исмаил основывает династию Сефевидов, первую коренную Персидское государство с 11 века. | |
| 1505 | г. Василий III (годы правления 150533) становится великим князем Московским. | |
| 1517 | 90 013 Вызов Мартина Лютера практике католической церкви дал о себе знать протестантская Реформация, процесс, расколовший века религиозного единство Западной, Центральной и Южной Европы. | |
| 1520 | Слейман I Великолепный (годы правления 152066) становится османским султаном. | |
| 1529 | Первая османская осада Вены; будет предпринята вторая осада в 1683 году. | |
| 1533 | Иван IV Грозный (годы правления 154784) становится великим князем. Москвы.В 1547 году он провозглашает себя «царем Московии». | |
| 1552 | Северный морской путь в Россию открыт англичанином Ричардом Канцлер. | |
| 1556 | Начинается правление Акбара I Великого (годы правления 1556–1605) в Индии;
он будет представлять собой апогей Империи Великих Моголов с центром в Дели. | |
| 155859 | Английский купец Энтони Дженкинсон выполняет миссии в Персию и Среднюю Азию в качестве личного посланника царя Ивана IV. | |
| 1564 | Первая датированная книга в Москве напечатана Иваном Федоровым. | |
| 1577 | Россия устанавливает торговые связи с Голландией. | |
| 1584 | Федор I (р. 158498) становится последним из династии Рюриковичей, которая правил Московией с 862 г. |

ученый ОИЯИ стал лауреатом Федоровского конкурса
В декабре на заседании Президиума Российской инженерной академии были подведены итоги Всероссийского конкурса имени первооткрывателя книгопечатания Ивана Федорова на лучшую научно-методическую работу. Среди лауреатов конкурса 2021 года – известный российский физик, заведующий научно-экспериментальным отделом физики тяжелых ионов Лаборатории физики высоких энергий ОИЯИ, профессор Александр Малахов.
Среди лауреатов конкурса 2021 года – известный российский физик, заведующий научно-экспериментальным отделом физики тяжелых ионов Лаборатории физики высоких энергий ОИЯИ, профессор Александр Малахов.
Организаторами конкурса выступили Российская инженерная академия, Ассоциация технических университетов и Российский государственный университет имени Косыгина. В 26 секциях конкурса приняли участие профессора российских и зарубежных вузов, специалисты и инженеры предприятий, организаций, научно-исследовательских институтов, доктора и кандидаты наук. Конкурс учрежден для поощрения ученых, внесших значительный вклад в науку и образование.
Прочитать книгу по ссылке
«Издание такого пособия помогает ориентировать студентов и молодых сотрудников в большом информационном потоке, освещая основные направления исследований в ОИЯИ. Большую роль в этом играют теоретические доклады, дополненные краткими описаниями основной массы экспериментов по релятивистской ядерной физике на синхрофазотроне и нуклотроне ОИЯИ. В пособии представлены ускорительные центры, выполняющие исследования релятивистских ядерных взаимодействий.Особое внимание автор уделяет мегасайенс-проекту NICA, который имеет большое значение в настоящее время», — комментирует необходимость издания книги Александр Иванович.
В пособии представлены ускорительные центры, выполняющие исследования релятивистских ядерных взаимодействий.Особое внимание автор уделяет мегасайенс-проекту NICA, который имеет большое значение в настоящее время», — комментирует необходимость издания книги Александр Иванович.
Кроме того, в пособии подробно представлен разработанный академиком А. М. Балдиным подход к изучению релятивистских ядерных взаимодействий в пространстве четырехмерных скоростей. Такой подход позволил нашим ученым получить множество значительных результатов в этой области. В книге широко представлены результаты исследований по релятивистской ядерной физике в других крупных научных центрах мира: в БНЛ (США) на RHIC, в ЦЕРНе на SPS и LHC.
«Особого внимания заслуживает прекрасное издание книги. Это большая заслуга Издательского отдела ОИЯИ. В связи с этим хочу выразить огромную благодарность коллективу Издательского отдела ОИЯИ за работу по изданию книги и особую благодарность заведующей отделом Анастасии Шабашовой и редактору Екатерине Григорьевой», – подчеркнул лауреат, говоря о особенности руководства.
Как издателям следует подготовиться к отказу от сторонних файлов cookie?
30 марта команда Admixer представила решения, которые помогут онлайн-издателям сохранить свои доходы от рекламы после устранения сторонних файлов cookie в начале 2022 года.
Ярослав Холод , директор Programmatic Operations, Елена Подшувейт , директор по продуктам, и Иван Федоров , директор по новым направлениям бизнеса, представили решения Admixer, которые помогут отрасли преодолеть критический вызов, с которым она столкнется в ближайшем будущем. год — изменения в идентификации пользователя в сети.
Технологии, представленные на вебинаре, будут полезны владельцам сайтов и СМИ, медиаменеджерам и всем, кто работает с рекламой и данными на стороне издателя.
Посмотреть веб-семинар полностьюКак издателям подготовиться к поэтапному отказу от сторонних файлов cookie?
Ярослав Холод, директор по программным операциям Admixer
Полная ликвидация сторонних файлов cookie — одно из важнейших событий для рекламного рынка за последние 15 лет.
Что сегодня могут делать интернет-издатели?
- Укрепляйте отношения с прямыми рекламодателями
- Научитесь делать контекстную разметку страниц или начните работать с платформами, которые это делают и передают данные рекламодателям (контекст страницы без привязки к персональным данным пользователя может стать хорошей альтернативой таргетингу на аудиторию )
- Обратите внимание на работу со сторонними данными и идентификаторами, рассмотрите возможность развертывания альтернативных решений для идентификации (например, Admixer ID)
- Переход на технологию header bidding
Admixer ID — решение для идентификации пользователей в открытом Интернете
Иван Федоров, новый бизнес-директор Admixer
Будущее идентификации браузера имеет три наиболее вероятных сценария: Песочница конфиденциальности Google (идентифицирует когорты, а не конкретных пользователей), 1st-party ID (идентифицирует браузер пользователя), Unified ID (идентифицирует конкретного пользователя).
Песочница конфиденциальности
Вместе с Google другие рекламные компании разрабатывают новые подходы к таргетингу рекламы без использования идентификатора пользователя. Предлагаемый API будет основан на когортном анализе истории посещенных страниц, а не на данных отдельных пользователей.
Помимо инициативы Google FLoC, другие компании представили свои предлагаемые API будущего, например SPARROW от Criteo. Эти решения будут действовать только в Chrome — другие браузеры пока не проявили интереса к этим инициативам.
Собственный идентификатор
Другой подход заключается в использовании собственных идентификаторов издателей. Для веб-сайтов это собственные файлы cookie , которые устанавливаются в браузере пользователя. Эта система может работать только для определенного домена и для определенного браузера.
Но если на сайте есть система регистрации, например решение единого входа (SSO), которое собирает универсальные идентификаторы, то его можно использовать для идентификации пользователей на разных сайтах и платформах.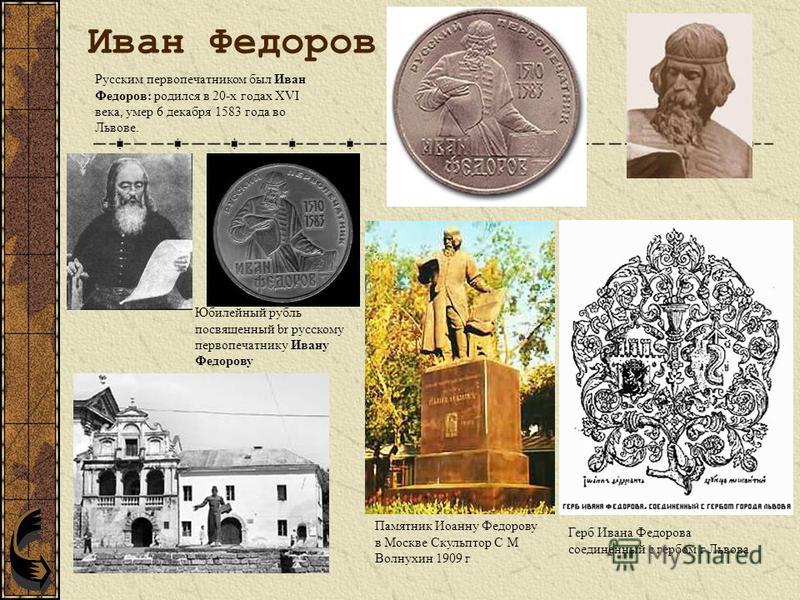
Компания Google уже объявила, что планирует поддерживать собственные идентификаторы для своих издателей.Для этого в Google Ad Manager уже есть механизм pubID, но для массового внедрения решение еще нуждается во многих доработках.
Универсальный идентификатор
Более 100 компаний сотрудничают в рамках этой инициативы в рамках проекта Rearc в рамках IAB Tech Lab.
Платформы могут использовать универсальный идентификатор поверх других идентификаторов, что может значительно расширить охват рекламных кампаний и идентифицировать пользователей так же, как сторонние файлы cookie, но с использованием зашифрованных идентификаторов.
Универсальный идентификатор также является кросс-канальным и кросс-платформенным, что делает его более совершенным, чем файлы cookie, которые могут идентифицировать пользователей только в браузере.
В универсальном идентификаторе два идентификатора работают вместе. зарегистрироваться в обмен на доступ к контенту. Основным ограничением этого решения является масштабирование: для его эффективности требуется соглашение с несколькими издателями. Так что для одних рекламодателей это может хорошо сработать, для других охват в некоторых регионах и нишах может быть весьма ограниченным.
Основным ограничением этого решения является масштабирование: для его эффективности требуется соглашение с несколькими издателями. Так что для одних рекламодателей это может хорошо сработать, для других охват в некоторых регионах и нишах может быть весьма ограниченным.
Что такое Admixer ID?
Admixer ID — это универсальный идентификатор, который позволяет отслеживать пользователей в разных браузерах, платформах и устройствах без сторонних файлов cookie.
Admixer ID предоставляет рекламодателям и издателям единую систему для сегментации аудитории, таргетинга и измерения.
Поскольку представители Admixer входят в состав рабочей группы Rearc в IAB Tech Lab, решение было основано на принципах идентификации пользователей, установленных организацией.
Как работает Admixer ID?
Admixer ID использует хешированные адреса электронной почты и номера телефонов, полученные с явного согласия пользователя. Персональные данные из идентификатора невозможно расшифровать, поэтому пользователи могут быть уверены, что их PII не будет использована злоумышленниками.
- Каждому пользователю веб-сайта или приложения, который оставляет свой адрес электронной почты или номер телефона, присваивается уникальный идентификатор.
- User ID связан с Admixer ID.
- Идентификатор передается в Admixer User Graph, который соответствует идентификаторам пользователей из разных источников и сред:
- Веб-идентификаторы (собственные данные входа, используемые для доступа к онлайн-ресурсам)
- Мобильные идентификаторы (IDFA для iOS) и Android ID)
- Другой сторонний идентификатор , разработанный различными поставщиками технологий (например, Unified ID 2.0, LiveRamp, ID5, Net ID и т. д.)
- Проверка и измерение кроссплатформенный Admixer ID отправляется рекламодателям, которые затем могут активировать его в рекламных кампаниях.
- Рекламодатели связывают идентификаторы Admixer пользователей со своими собственными идентификаторами 1st-party из CRM / CDP и других платформ, чтобы повысить эффективность кампании.

- Возможность использования с существующей системой авторизации и идентификации пользователей
- Готовые коннекторы для издателей (Tag Manager, SSO, HB Wrapper, SDK) и для рекламодателей (Tag Manager, Onboarding Tools)
- Интеграция с другими системами идентификации для глобального использования (Liveramp IDL, Unified ID 2.0, HB User Module) и измерения аудитории (IAB Rearc Compliant)
- Готовые инструменты для активации кампаний Header Bidding, Direct Deals, Open Auction и монетизации аудитории
- Конфиденциальность пользователей и механизмы отказа
- 52% издателей считают, что решения HB не предоставляют достаточно данных для оптимизации
- 37% убеждены, что управление HB отнимает много времени в установке и настройке
- Обеспечивает полную прозрачность данных об источниках доходов
- Предоставляет подробную статистику в режиме реального времени
- Подключает новых покупателей в один клик
- Поддерживает отображение и видеообъявления
- Быстро и легко настраивается путем одноразовой установки кода рекламного блока
- Поэтапный отказ от сторонних файлов cookie повлияет на всю цифровую рекламу: планирование кампании, таргетинг на аудиторию , атрибуция и измерение эффективности рекламы. Большинство пользователей открытой сети станут неидентифицируемыми.
- Если издатели не изменят свой подход к рекламе и управлению данными, они могут потерять около 70% дохода от рекламы после 2021 года.
- Данные и идентификаторы 1-й стороны , так как собственные файлы cookie веб-сайта будут продолжать работать.
- Издателям уже нужно выбирать решение, которое поможет сохранить таргетинг на аудиторию после отключения файлов cookie в Chrome, например, Admixer ID — универсальный идентификатор, который работает без сторонних файлов cookie и позволяет идентифицировать пользователей в разных браузерах , на разных платформах и устройствах.

- Возрастающая сложность рекламной экосистемы не позволяет вам эффективно взаимодействовать со многими партнерами одновременно и получать более высокие ставки, доступные на рынке. Технология Header Bidding может решить эти проблемы.
- Заголовочные ставки позволяют вам подключать и управлять многими рекламодателями и получать несколько ставок одновременно . Это увеличивает конкуренцию за рекламное место, среднюю цену за тысячу показов и доход сайта.
Важность поля. Адаптация CRISPR представляет собой сложный многоэтапный процесс, который остается наименее изученным этапом адаптивного прокариотического иммунитета. В то время как большинство практических применений CRISPR построено на стадии интерференции, понимание адаптации, в дополнение к фундаментальному пониманию процесса, позволит эффективно создавать клетки с определенными профилями устойчивости и записывать «воспоминания» о конкретных событиях с временным разрешением.
 Эти разработки были бы невозможны без высокочувствительных методов обнаружения редких случаев приобретения спейсеров.
Эти разработки были бы невозможны без высокочувствительных методов обнаружения редких случаев приобретения спейсеров.Краткое изложение текущих мыслей. Несмотря на значительные усилия, в отсутствие селекции наилучшие доступные методы могут обнаружить адаптацию, когда клетки с расширенными массивами составляют не менее 0,01% популяции. Эти уровни, вероятно, на несколько порядков выше наблюдаемых в естественных условиях, где высокие уровни адаптации CRISPR, по крайней мере, при отсутствии вирусной инфекции, должны быть весьма неблагоприятными из-за последующего вредного аутоиммунитета.Действительно, большинство используемых в настоящее время методов используются для изучения искусственных систем, в которых адаптация активируется за счет сверхэкспрессии cas ответственных генов.
Будущие направления. Поскольку некоторые доступные методы, такие как SENECA, использование вырожденных праймеров и супПЦР, ортогональны друг другу, их комбинирование может несколько повысить чувствительность.
 При отсутствии дальнейших разработок очень глубокое секвенирование, вероятно, станет методом выбора для изучения адаптации, особенно у организмов, для которых не доступны методы генетических манипуляций.Разработка и применение таких методов, как FragSeq, для изучения коротких нуклеиновых кислот внутри клеток должно позволить проводить дополнительные исследования преспейсеров. Наибольший интерес представляет сравнительный анализ преспейсеров, генерируемых различными типами систем CRISPR-Cas, и установление взаимосвязи между генерацией преспейсеров и процессами поддержания ДНК в клетке. Выбор преспейсеров практически не изучен. Необходимо разработать новые чувствительные и высокопроизводительные методы, чтобы раскрыть детали этой самой ранней стадии формирования адаптивного иммунитета.
При отсутствии дальнейших разработок очень глубокое секвенирование, вероятно, станет методом выбора для изучения адаптации, особенно у организмов, для которых не доступны методы генетических манипуляций.Разработка и применение таких методов, как FragSeq, для изучения коротких нуклеиновых кислот внутри клеток должно позволить проводить дополнительные исследования преспейсеров. Наибольший интерес представляет сравнительный анализ преспейсеров, генерируемых различными типами систем CRISPR-Cas, и установление взаимосвязи между генерацией преспейсеров и процессами поддержания ДНК в клетке. Выбор преспейсеров практически не изучен. Необходимо разработать новые чувствительные и высокопроизводительные методы, чтобы раскрыть детали этой самой ранней стадии формирования адаптивного иммунитета.
Внедрение Admixer ID поможет владельцам веб-сайтов и СМИ повысить эффективную цену за тысячу показов и спрос на их ресурсы среди рекламодателей, а также даст им возможность конкурировать с богатыми данными Walled Gardens. Решение позволит рекламодателям проводить многоканальные кампании, измерять и проверять охват без ущерба для личности пользователя.
Преимущества Admixer ID
Елена Подшувейт, директор по продуктам Admixer
Недостатки водопадной модели
Большинство цифровой рекламы в Восточной Европе сейчас продается по водопадной модели, которая предлагает трафик сначала к прямым рекламодателям, затем к агентствам, а затем в рекламные сети.
Рекламный сервер, установленный на сайте издателя, постоянно предлагает размещение рекламы различным партнерам по спросу. Как только рекламодатель готов платить выше установленной минимальной ставки, он выигрывает показ, и сервер объявлений не ищет более высокие ставки.
Header Bidding (HB) — это технология автоматического аукциона , которая позволяет сайтам получать предложения от нескольких рекламодателей одновременно . HB помогает увеличить конкуренцию за рекламное место, а значит, среднюю эффективную цену за тысячу показов и доход сайта.
Многие интернет-издатели считают, что это решение технически сложное, непрозрачное и не может гарантировать эффективную монетизацию. Согласно исследованию Open X:
Несмотря на множество преимуществ, Head Bidding не так-то просто развернуть самостоятельно. Согласно исследованию Admixer, 38% сайтов в Украине не используют header bidding, а 43% внедрили это решение, но не добились желаемых результатов.
Согласно исследованию Admixer, 38% сайтов в Украине не используют header bidding, а 43% внедрили это решение, но не добились желаемых результатов.
Мы спросили онлайн-издателей, с какими трудностями они столкнулись и что могло бы помочь им лучше использовать эту технологию для создания лучшего решения, которое устранило бы камни преткновения.
Учитывая самые распространенные опасения паблишеров, мы создали Admixer HB, который:
Кроме того, платформа Admixer Header Bidding легко интегрируется с внешними рекламными серверами.
Как только вы получаете доступ к своей учетной записи, все необходимые настройки выполняются в интерфейсе платформы. Admixer Header Bidding интегрирован с Google Ad Manager, поэтому вы можете автоматически создавать позиции в GAM через интерфейс.
Admixer Header Bidding интегрирован с Google Ad Manager, поэтому вы можете автоматически создавать позиции в GAM через интерфейс.
Вы можете настроить HB-аукцион с GAM и внешними источниками спроса, доступными через платформу:
Подводя итог
Кривцун Николай
Программист
Летчик-герой Иван Федоров
Летчик-герой Иван Федоров — Главный Мюнхгаузен советской авиации (часть 1) Эрик Почему люди врут? Часто преследует корыстные или какие-то другие цели. Зажгли, чтобы скрыть иногородние действия или «скелеты в шкафу». Типа «скажу жене, что иду к любовнице, любовнице, что иду к жене, а сам на чердак и работаю». Я даже могу понять, когда человек, живущий скучной повседневной жизнью, в рассказах незнакомых людей всячески ее приукрашивает, притворяясь важным и значимым. Так мой друг детства Игорь Михайлович, будучи обычным слесарем по ремонту лифтов, постоянно выдавал себя за секретного агента спецслужб. Забавная, забойная шапка из нутрии и полное безденежье объясняются тем, что многомиллионные гонорары идут на счет сверхпроводника, которым в силу конспирации он не имеет права пользоваться.
Так мой друг детства Игорь Михайлович, будучи обычным слесарем по ремонту лифтов, постоянно выдавал себя за секретного агента спецслужб. Забавная, забойная шапка из нутрии и полное безденежье объясняются тем, что многомиллионные гонорары идут на счет сверхпроводника, которым в силу конспирации он не имеет права пользоваться.
А вот зачем нафантазировал и нафантазировал Иван Евграфович Федоров, мне совершенно непонятно. Летчик-динамик, испытания новых образцов самолетов, Герой Советского Союза, кавалер многих орденов и наград, участник войны в Испании и Великой Отечественной, человек несомненно смелый, по воспоминаниям современников, один из лучших летчиков ВВС.Почему он должен лгать? Отличное материальное положение, интересная и значимая жизнь. Как-то не надо было себя приукрашивать.
Но Федоров, мягко говоря, пофантазировал, и, грубо говоря, заблокировался. Всячески принимая ваши и без того симпатичные и достойные подвиги. И всю жизнь. И жил он долго. Умер в 2011 году в возрасте 94 лет. Нехорошо так говорить, но на старости лет заслуженный ветеран видимо впал в маразм старческий.
Нехорошо так говорить, но на старости лет заслуженный ветеран видимо впал в маразм старческий.
Сам верил в свои фантазии.Ко времени участников Великой войны осталось немного, а героев Советского Союза вообще читаются единицы. А к Федорову — многочисленные писатели и журналисты, люди, прямо скажем, ничего не знающие о войне. Федорова, которого все друзья ветеранов считали классным летчиком, но в то же время и Великий Трепл, найдя свободные и доверчивые уши, стал щедро подвешивать им и клюкву, и лапшу. Чего только от него не слышали девушки-журналисты.И около 134 сбитых самолетов (больше, чем у обшивки и ташкина вместе взятых). Иногда «под градусом» количество сбитых самолетов дорастало до 148, и про шесть, а то и про восемь воздушных таранов, которые он израсходовал и остался жив, и про золотую саблю, которую снял с немца Асса. Но апофеозом стал рассказ о том, как перед войной Гитлер наградил советского летчика Рыцарским крестом!!! В отличие от ветеранов, журналисты поверили. А потом с апломбом и ссылкой на ветерана их печатали в журналах и газетах, о них рассказывали в телепередачах.И приходится сомневаться в словах ветерана, а тем более героя не принято. Так что пока боимся фантазии Ивана Федорова и еще дольше будем разваливаться.
А потом с апломбом и ссылкой на ветерана их печатали в журналах и газетах, о них рассказывали в телепередачах.И приходится сомневаться в словах ветерана, а тем более героя не принято. Так что пока боимся фантазии Ивана Федорова и еще дольше будем разваливаться.
«На перроне, где долго надо было долго ждать, долго, долго, долго, Ивану Евграфовичу было тяжело — он победил начальника в серпах и рассказывал сказки, думая о них сразу, на морозе «Наверное, Женька подозревал, что мы не верим ни одному его слову. И мы действительно не верили.Но рассказ был показан с такой самоотверженностью, что ни прерывать, ни сомневаться в нем просто не хотелось. (Из воспоминаний летчика-испытателя Салеста).
Так что к его биографии, особенно рассказанной с его слов, следует относиться скептически. Слишком много романтической флоры и смешного вранья. Иван Евграфович Федоров родился 23 февраля, 1914 г. в Харькове в родной семье.Есть и другие версии его рождения.Иногда Федоров говорил, что он родился в кургане, который возил соль между хутором Харьковкой и станицей Каменской. Настоящая фамилия — Денисов. Его отец, будевен первой конной армии, вернувшийся с Гражданской войны в Луганск, переписал Сына на фамилию деда. Это, как говорится, от греха подальше, так как 8-летний Иван, будучи баркотом у местных богачей, в отместку хозяину хозяина растратил свое имение, нанеся немалый ущерб рабочему спокойствию.
Настоящая фамилия — Денисов. Его отец, будевен первой конной армии, вернувшийся с Гражданской войны в Луганск, переписал Сына на фамилию деда. Это, как говорится, от греха подальше, так как 8-летний Иван, будучи баркотом у местных богачей, в отместку хозяину хозяина растратил свое имение, нанеся немалый ущерб рабочему спокойствию.
Приходит время рассказа о легендарном дедушке. Как Ильф и Петров: «Венец всего — рассказ о могучем дедушке, который, будучи крепостными в одних плавках, в 60-градусный мороз купил в проруби.Дед Федоров, по его рассказам, жил 123 года, до самой смерти не болел, ходил в одной рубахе и водки ведрами в одной рубахе и водки. Прожил бы дед, еще неизвестно сколько, да попал на ржавый гвоздь и умер от заражения крови.
Ваня вместе с братьями безнаказан до 1927 года.В 14 лет он «взял на себя ум» и за два года успел окончить 5 классов.Работал слесарем-инструментальщиком Луганского завода парового труда и учился на Вечернем Рабатаке при Донецком институте народного просвещения, который окончил в 1930 году. Без отрыва от производства он занимался в Луганском летном училище и летал на самодельных планерах, вечно «больных» небесами. Полеты на планерах ушли в будущее, и уже в августе 1932 года Федоров окончил Луганскую 11-ю авиационную школу военных летчиков и начал службу в строевых частях ВВС младшим летчиком. Летчик он был очень хороший, как говорится, от бога. В 19 лет командир звена, по его рассказам — командир эскадрильи.
Без отрыва от производства он занимался в Луганском летном училище и летал на самодельных планерах, вечно «больных» небесами. Полеты на планерах ушли в будущее, и уже в августе 1932 года Федоров окончил Луганскую 11-ю авиационную школу военных летчиков и начал службу в строевых частях ВВС младшим летчиком. Летчик он был очень хороший, как говорится, от бога. В 19 лет командир звена, по его рассказам — командир эскадрильи.
В 1937 году Федоров вместе с группой летчиков отправляется в Испанию.Русские летчики называли себя Педро, Хосе, Хуанс. Иван Евграфович тоже стал Хуаном, или Жуаном. Вернувшись на родину, преобразился в Женю. А имя Женя осталось за ним и, видимо, ему очень понравилось.
По рассказам самого Федорова, за год он совершил 286 боевых вылетов в Испанию, провел 36 авиасвязей, показавших исключительные образцы воздушного боя. Он уничтожил лично 11 самолетов противника и 13 в группе, в том числе 2 Ме-109 — новую немецкую машину, которая была неуязвима для фашистов. Дважды помечал вражеские машины — 18 июля и 21 августа 1937 года. Один раз, перехватив одиночный «Фиат», вымотал его в 20-минутном бою и заставил сесть на свой аэродром.
Дважды помечал вражеские машины — 18 июля и 21 августа 1937 года. Один раз, перехватив одиночный «Фиат», вымотал его в 20-минутном бою и заставил сесть на свой аэродром.
По официальным данным и воспоминаниям других летчиков, Федоров на своем И-16 совершил более 150 боевых вылетов (боевой налет составил 180-190 часов). В районе города Картахена лично были сбиты два оставшихся без прикрытия итальянских бомбардировщика «Savoy Marketti SM.79». Не было таранцев, сбитых «мессерами» и посаженных итальянцев.Генерал Борис Смирнов и старший советник по военной авиации Яков Смеушевич (легендарный генерал Дуглас), командовавшие в Испании нашими летчиками, о Федорове даже не упоминают.
И.Е. Федоров: воевал в секретной группе летчиков Асова вместе с П. Леччаговым, П. Пумпером и М. Якушевым.
Архивные данные: Федоры с П.И. Пумпуром и П.В. Рычаг в Испании даже не встречал. Все они в командировке в разное время.
Летчик-герой Иван Федоров — Главный Мюнхгаузен советской авиации (часть 1) Эрик И. Федоров: За подвиги награжден Высшим республиканским орденом Испании «Лавра Мадрид». Восхищенные героизмом испанцы прозвали его «Дибле Рохо» («Красный дьявол»). Но самой ценной наградой для него стал страстный поцелуй Долорес Ибаррури, вдобавок к которому она подарила пилоту трехэтажную виллу, «Пэтетон» и пушку «Астра».
Федоров: За подвиги награжден Высшим республиканским орденом Испании «Лавра Мадрид». Восхищенные героизмом испанцы прозвали его «Дибле Рохо» («Красный дьявол»). Но самой ценной наградой для него стал страстный поцелуй Долорес Ибаррури, вдобавок к которому она подарила пилоту трехэтажную виллу, «Пэтетон» и пушку «Астра».
Официальные источники: Никаких документальных подтверждений о награждении И.Федорова орденом «Мадридская Лавра» нет. Вроде нет подтверждения про трехэтажную виллу.В воспоминаниях летчиков-участников войны в Испании среди знатных имен Фёдоров не встречается. А прозвищем «Красный дьявол» фашисты в то время окрестили бесстрашного испанского летчика Франсиско Ошана, впоследствии погибшего в воздушном бою.
Летчик-герой Иван Федоров — Главный Мюнхгаузен советской авиации (часть 1) Эрик И. Федоров: За подвиги в Испании представлен к высшей награде Советского Союза — Золотой Звезде Героя. С большой группой офицеров из Испании Федоров приехал в Москву на торжественное представление. Среди награжденных, кроме летчиков, были моряки и танкисты. На одном из «банкетов» представители дружественного труда войск стали выяснять, какие вооруженные силы лучше. Спор дошел до драки, в которой участвовало сто человек, а затем и до перестрелки. В результате 11 санитарных машин доставили пострадавших в московские больницы и морги. Иван Федоров особого участия в драке не принял, но, не очнувшись, ударил приставленного к нему офицера НКВД. Пилот был первоклассным боксером — на вторые сутки человек, не приходя в сознание, скончался.В итоге Федорова объявили одним из зачинщиков скандала. На это происшествие надавило руководство защиты наркомана, но наград никому не дали. Нас всех ругали по воинским частям с совершенно не подходящими для дальнейшей карьеры характеристиками. От ареста и смерти Федорова сам Сталин приказал Берии не трогать летчика, дабы не осложнять отношения с испанцами, для которых Иван был национальным героем. Однако Федорова из ВВС уволили и перевели летчиком-испытателем в КБ С.
Среди награжденных, кроме летчиков, были моряки и танкисты. На одном из «банкетов» представители дружественного труда войск стали выяснять, какие вооруженные силы лучше. Спор дошел до драки, в которой участвовало сто человек, а затем и до перестрелки. В результате 11 санитарных машин доставили пострадавших в московские больницы и морги. Иван Федоров особого участия в драке не принял, но, не очнувшись, ударил приставленного к нему офицера НКВД. Пилот был первоклассным боксером — на вторые сутки человек, не приходя в сознание, скончался.В итоге Федорова объявили одним из зачинщиков скандала. На это происшествие надавило руководство защиты наркомана, но наград никому не дали. Нас всех ругали по воинским частям с совершенно не подходящими для дальнейшей карьеры характеристиками. От ареста и смерти Федорова сам Сталин приказал Берии не трогать летчика, дабы не осложнять отношения с испанцами, для которых Иван был национальным героем. Однако Федорова из ВВС уволили и перевели летчиком-испытателем в КБ С. А. Лавочкин.
А. Лавочкин.
Из официальных данных, милицейских протоколов и воспоминаний очевидцев: После Испании Иван Федоров был представлен к награждению орденом Красного Знамени, который получил. Бой после награждения состоялся. Но участвовало в нем не сто человек, а всего восемь. Федоров бросился с танкистом, который не поверил его рассказам. Мертвых вредителей не было, всех участников разделяли только синяки, стрельбы не было. Сам Федоров после награждения был назначен командиром эскадрильи 17-го истребительного Авиамара.За короткое время вывел эскадрилью на 1-е место в части, после чего в июне того же года был назначен помощником командира полка, а с 10 октября 1938 г. вступил в командование этим полком.
Как видите, все правда, но сильно приукрашено и преувеличено. О дальнейших подвигах отважного летчика Федорова-Мюнгаузена, как реальных, так и мифических, расскажет мне во второй части статьи.
Если вам понравился пост, то ставьте лайки, пишите комментарии, подписывайтесь на канал.
Острожская Библия
Выставка приурочена ко Дню православной книги и 450-летию со дня рождения апостола Ивана Федорова.
14 марта Белорусская Православная Церковь отмечает День православной книги. Ежегодное празднование учреждено в 2009 году Синодом Русской Православной Церкви. Он приурочен к выходу в свет первой русской датированной печатной книги «Апостол » Ивана Федорова (1564 г.). В 2014 году исполняется 450 лет этому знаменательному изданию.
Острожская Библия – это первый наиболее полный и лучший по тем временам перевод Библии на церковнославянский язык. Книга была напечатана Иваном Федоровым в Остроге в 1581 году по инициативе и при финансовой поддержке князя Константина Острожского и по благословению игумена Дубенского Иова.
Острожская Библия состоит из 76 книг Ветхого и Нового Завета. Писатели, переводчики и редакторы того времени собрались в Остроге, чтобы подготовить это издание Библии, и проделали огромную работу по сравнительному анализу, переводу и изданию различных текстов. Руководил этим процессом, скорее всего, Герасим Смотрицкий, ректор Острожской академии. Человек с высшим для того времени образованием, он стал первым официальным русским поэтом, а его стихи в Острожской Библии с описанием герба князя Константина Острожского — первыми русскими литературными стихами.
Руководил этим процессом, скорее всего, Герасим Смотрицкий, ректор Острожской академии. Человек с высшим для того времени образованием, он стал первым официальным русским поэтом, а его стихи в Острожской Библии с описанием герба князя Константина Острожского — первыми русскими литературными стихами.
Издание Острожской Библии сыграло важнейшую роль в православном воспитании. Для Русской Православной Церкви эта Библия стала образцом языкового стандарта и основой для Грамматики (1619) Мелетия Смотрицкого, которая с XVII века определяла формы церковнославянского языка.
Иван Федоров руководил печатью Библии. Как и все издания этого выдающегося первопечатника, Острожская Библия отлично набрана и выполнена. Для основного текста был специально отлит так называемый «Острожский» шрифт. Многие приемы декорирования, использованные Иваном Федоровым при создании Острожской Библии, стали впоследствии характерными для украинского и белорусского книгоиздания.
Издана Острожская Библия в фолиантах. Она состоит из 628 листов и украшена ксилографической рамкой обложки, гербом князя Константина Острожского и печатным знаком Ивана Федорова.Центральная часть заставки, открывающая основной текст Острожской Библии, оттиснута с пластины из Апостола (1564 г.), а также форма картуша для герба князя Константина Острожского на оборотной стороне титульного листа . В рисунках преобладают растительные элементы. Весь дизайн Острожской Библии строг и прост, без фигурных гравюр.
Она состоит из 628 листов и украшена ксилографической рамкой обложки, гербом князя Константина Острожского и печатным знаком Ивана Федорова.Центральная часть заставки, открывающая основной текст Острожской Библии, оттиснута с пластины из Апостола (1564 г.), а также форма картуша для герба князя Константина Острожского на оборотной стороне титульного листа . В рисунках преобладают растительные элементы. Весь дизайн Острожской Библии строг и прост, без фигурных гравюр.
Объем 1256 страниц был издан тиражом 1500 экземпляров, что по тем временам было немалым тиражом.Инициатор издания князь Константин Острожский подарил Библию папе Григорию XIII, а Иван Грозный – посланнику королевы Джерому Горсею. Также известна книга, принадлежавшая шведскому королю Густаву II Адольфу. В настоящее время сохранилось (полностью или частично) всего 260 экземпляров Острожской Библии. Они хранятся в музеях, библиотеках и архивах Украины, России, Белоруссии, Болгарии, Германии, Франции и многих других стран. Два подлинных экземпляра этого древнего памятника православных верований хранятся в фондах Национальной библиотеки Беларуси.
Два подлинных экземпляра этого древнего памятника православных верований хранятся в фондах Национальной библиотеки Беларуси.
Дополнительную информацию можно получить по телефону: (375 17) 293 25 85.
|
Обнаружение адаптации CRISPR
Biochem Soc Trans. 2020 28 февраля; 48(1): 257–269.
Ширяева Анна
1 Центр наук о жизни Сколковского института науки и технологий, Москва 121205, Россия
2 Институт Ваксмана, Рутгерс, Государственный университет Нью-Джерси, Пискатауэй, NJ 08854, U.СА
Иван Федоров
1 Центр наук о жизни Сколковского института науки и технологий, Москва 121205, Россия
3 Институт биологии гена Российской академии наук, Москва 119334, Россия Даныловыховский
7
1 Центр наук о жизни Сколковского института науки и технологий, Москва 121205, Россия
Северинов Константин
1 Центр наук о жизни Сколковского института науки и технологий, Москва 121205 1
2, Россия 0 Институт Ваксмана, Рутгерс, Государственный университет Нью-Джерси, Пискатауэй, Нью-Джерси 08854, Ю. ЮА
ЮА4 Центр точного редактирования генома и генетических технологий в биомедицине Института биологии гена РАН, Москва 119334, Россия
1 Центр наук о жизни Сколковского института науки и технологий, Москва 121205 , Россия
2 Институт им. за прецизионное редактирование генома и генетические технологии в биомедицине, Институт биологии гена РАН, Москва 119334, Россия
Поступила в редакцию 15.11.2019 г.; Пересмотрено 26 декабря 2019 г.; Принято 3 января 2020 г.
Эта статья была процитирована другими статьями в PMC.Abstract
Прокариотический адаптивный иммунитет строится, когда короткие фрагменты ДНК, называемые спейсерами, встраиваются в массивы CRISPR (группированные регулярно расположенные короткие палиндромные повторы). Адаптация CRISPR — это многоэтапный процесс, который включает в себя выбор, создание и включение преспейсеров в массивы. После адаптации спейсеры обеспечивают иммунитет за счет распознавания комплементарных последовательностей нуклеиновых кислот, направляя их на разрушение. Таким образом, чтобы предотвратить вредный аутоиммунитет, адаптация CRISPR должна быть строго регулируемым и нечастым процессом, по крайней мере, в отсутствие генетических захватчиков. За прошедшие годы были разработаны оригинальные методы изучения адаптации CRISPR. В этой статье мы обсуждаем и сравниваем методы, которые обнаруживают адаптацию CRISPR и ее промежуточные продукты in vivo , и предлагаем подавление ПЦР в качестве простой модификации популярного анализа для мониторинга приобретения спейсера с повышенной чувствительностью.
Таким образом, чтобы предотвратить вредный аутоиммунитет, адаптация CRISPR должна быть строго регулируемым и нечастым процессом, по крайней мере, в отсутствие генетических захватчиков. За прошедшие годы были разработаны оригинальные методы изучения адаптации CRISPR. В этой статье мы обсуждаем и сравниваем методы, которые обнаруживают адаптацию CRISPR и ее промежуточные продукты in vivo , и предлагаем подавление ПЦР в качестве простой модификации популярного анализа для мониторинга приобретения спейсера с повышенной чувствительностью.
Ключевые слова: Адаптация CRISPR, массивы CRISPR, системы CRISPR-Cas, спейсеры другие мобильные генетические элементы (МГЭ) [4,5].Они состоят из массивов CRISPR и CRISPR-ассоциированных генов cas [6,7]. Массивы CRISPR состоят из идентичных или очень похожих повторов, разделенных уникальными последовательностями ДНК, называемыми спейсерами [6,7]. Общее количество спейсеров в массиве варьируется от одной до нескольких сотен [6,8]. Источник подавляющего большинства (~93%) спейсеров остается неизвестным, они составляют «темную материю» CRISPR [9]. Большинство оставшихся спейсеров сопоставляются с MGE и могут рассматриваться как воспоминания о предыдущих встречах, которые клетки хранят в массивах CRISPR [9].Выше локусов CRISPR находится AT-богатая последовательность, называемая «лидером» [7]. Массивы CRISPR транскрибируются с промотора, расположенного в лидере, и первичный транскрипт процессируется в CRISPR РНК (crРНК), содержащих один спейсер и фланкирующие последовательности, полученные из повторов [10–17]. Белки Cas вместе с crРНК образуют эффекторные комплексы (комплекс Cascade в системе типа I-E Escherichia coli ), которые узнают «протоспейсеры» — последовательности ДНК или, иногда, РНК, комплементарные спейсеру crРНК [13,18–20].Распознавание протоспейсеров в МГЭ приводит к их разрушению [18–20].
Источник подавляющего большинства (~93%) спейсеров остается неизвестным, они составляют «темную материю» CRISPR [9]. Большинство оставшихся спейсеров сопоставляются с MGE и могут рассматриваться как воспоминания о предыдущих встречах, которые клетки хранят в массивах CRISPR [9].Выше локусов CRISPR находится AT-богатая последовательность, называемая «лидером» [7]. Массивы CRISPR транскрибируются с промотора, расположенного в лидере, и первичный транскрипт процессируется в CRISPR РНК (crРНК), содержащих один спейсер и фланкирующие последовательности, полученные из повторов [10–17]. Белки Cas вместе с crРНК образуют эффекторные комплексы (комплекс Cascade в системе типа I-E Escherichia coli ), которые узнают «протоспейсеры» — последовательности ДНК или, иногда, РНК, комплементарные спейсеру crРНК [13,18–20].Распознавание протоспейсеров в МГЭ приводит к их разрушению [18–20].
Иммунитет к CRISPR создается во время адаптации CRISPR, процесса, который влечет за собой включение новых спейсеров в массив [4]. Новые спейсеры обычно включаются на границе между лидером и первым повтором, поэтому хронологический порядок приобретения спейсеров соответствует обратному порядку спейсеров в массиве [4, 21, 22]. Для каждого полученного спейсера создается новая копия повтора [4,21,22]. Два наиболее консервативных белка Cas, Cas1 и Cas2, общие почти для всех систем CRISPR-Cas, катализируют интеграцию предшественников спейсеров (преспейсеров) в массивы [23–25].Как правило, приобретение спейсеров не направлено конкретно на МГЭ, и, таким образом, спейсеры также могут быть получены из собственного генома клетки [23,26]. Это может привести к аутоиммунному ответу, подавляющему рост клеток [27–29]. Неудивительно, что адаптация CRISPR — это жестко контролируемый процесс, обычно протекающий с очень низкой эффективностью, и его бывает трудно обнаружить как в естественных условиях, так и в лабораторных экспериментах. Было разработано несколько методов обнаружения адаптации CRISPR, которые помогли пролить свет на молекулярные механизмы, управляющие выбором спейсера.
Новые спейсеры обычно включаются на границе между лидером и первым повтором, поэтому хронологический порядок приобретения спейсеров соответствует обратному порядку спейсеров в массиве [4, 21, 22]. Для каждого полученного спейсера создается новая копия повтора [4,21,22]. Два наиболее консервативных белка Cas, Cas1 и Cas2, общие почти для всех систем CRISPR-Cas, катализируют интеграцию предшественников спейсеров (преспейсеров) в массивы [23–25].Как правило, приобретение спейсеров не направлено конкретно на МГЭ, и, таким образом, спейсеры также могут быть получены из собственного генома клетки [23,26]. Это может привести к аутоиммунному ответу, подавляющему рост клеток [27–29]. Неудивительно, что адаптация CRISPR — это жестко контролируемый процесс, обычно протекающий с очень низкой эффективностью, и его бывает трудно обнаружить как в естественных условиях, так и в лабораторных экспериментах. Было разработано несколько методов обнаружения адаптации CRISPR, которые помогли пролить свет на молекулярные механизмы, управляющие выбором спейсера. Эти методы и их ограничения обсуждаются ниже.
Эти методы и их ограничения обсуждаются ниже.
Методы обнаружения адаптации CRISPR в отдельных клетках или клонах, основанные на селекции
Очевидным способом обнаружения приобретения нового спейсера является амплификация лидер-проксимального конца массива CRISPR с парой праймеров: один соответствует лидеру , а другой соответствует внутренней, уже существующей прокладке [23,30,31]. Поскольку новые спейсеры обычно встраиваются перед первым, лидерно-проксимальным повтором, и приводят к дублированию повтора [4, 21–23], обнаружение продуктов ПЦР, расширенных на целое число единиц спейсер-повтор, выявляет события адаптации CRISPR.Однако, поскольку приобретение спейсеров может быть очень редким, требуется специфический отбор адаптированных клеток (14). Примеры такой селекции включают получение колоний BIM (мутанты, нечувствительные к бактериофагам) () [4,21,22,31,32] или PIM (мутанты, интерферирующие с плазмидами) () [30,31,33]. Эти методы дешевы и не требуют генетических манипуляций с изучаемыми клетками, но они ориентированы на способные к интерференции спейсеры, полученные из МГЭ, и поэтому не могут использоваться для обнаружения спейсеров, которые не приводят к интерференции против МГЭ или приводят к самоинтерференции из-за приобретение спейсера из собственного генома клетки (в зависимости от подтипа CRISPR-Cas, при инактивации интерференции такие спейсеры могут составлять от 2 до 99% приобретенных спейсеров [23,26,34,35]).
Обнаружение приобретения способных к интерференции спейсеров (S +1 ) у нечувствительных к бактериофагам мутантов ( A ) или в клетках, утративших плазмиды ( B ). Вверху показаны структуры нерасширенных (UA) массивов CRISPR в клетках из исходной культуры или расширенных (EA) массивов в клетках, которые либо выжили после заражения фагом, либо потеряли плазмиду. Показаны лидер (светло-персиковый прямоугольник), повторы CRISPR (белые прямоугольники), ранее существовавший спейсер S 0 (темно-персиковый прямоугольник) и вновь приобретенный спейсер S +1 (бирюзовый прямоугольник).Приобретение спейсеров проверяется с помощью ПЦР с праймерами, соответствующими лидеру (светло-персиковая стрелка) и спейсеру S 0 (темно-персиковая стрелка). Ампликоны из расширенных и нерасширенных массивов показаны в виде скобок под праймерами. В A колония, образовавшаяся после инфицирования фагом, непосредственно тестируется с помощью ПЦР, и результаты электрофореза ампликонов в агарозном геле, полученных с исходными клетками (1), и устойчивой к фагам колонии (2) схематически показаны справа. В B жидкая культура инокулируется клетками из устойчивой к антибиотикам (Ab-устойчивой) колонии, несущей плазмиду (1) с нерасширенным массивом CRISPR.После выращивания и посева на неселективную (-Ab) среду присутствие плазмиды проверяют путем посева штрихом колоний (2, 3, 4) на чашки с Ab и без него. Клетки из колонии, чувствительной к антибиотикам (2), дополнительно тестируют с помощью ПЦР, чтобы выявить событие приобретения спейсера. M, маркер размера молекулярной массы. Обнаружение адаптации CRISPR с использованием репортерных систем cat ( C ) или yfp ( D ). Вставка массива CRISPR с перевернутым лидером/одиночным спейсером нарушает рамку считывания репортерных генов cat ( C ) или yfp ( D ).Элементы массивов CRISPR показаны как A и B . P репортерный — промотор, направляющий синтез репортерной мРНК; P CRISPR — промотор, направляющий синтез пре-крРНК.
В B жидкая культура инокулируется клетками из устойчивой к антибиотикам (Ab-устойчивой) колонии, несущей плазмиду (1) с нерасширенным массивом CRISPR.После выращивания и посева на неселективную (-Ab) среду присутствие плазмиды проверяют путем посева штрихом колоний (2, 3, 4) на чашки с Ab и без него. Клетки из колонии, чувствительной к антибиотикам (2), дополнительно тестируют с помощью ПЦР, чтобы выявить событие приобретения спейсера. M, маркер размера молекулярной массы. Обнаружение адаптации CRISPR с использованием репортерных систем cat ( C ) или yfp ( D ). Вставка массива CRISPR с перевернутым лидером/одиночным спейсером нарушает рамку считывания репортерных генов cat ( C ) или yfp ( D ).Элементы массивов CRISPR показаны как A и B . P репортерный — промотор, направляющий синтез репортерной мРНК; P CRISPR — промотор, направляющий синтез пре-крРНК. Состав репортерных мРНК показан ниже. Каждый прямоугольник соответствует одному кодону; rbs — сайт связывания рибосомы; AUG — стартовый кодон, UAA — стоп-кодон, расположенный в рамке с AUG. Трансляция мРНК, транскрибированных с нерасширенных репортеров CRISPR, останавливается на кодоне UAA в лидерной последовательности, что приводит к ( C ) чувствительности к хлорамфениколу (Cm-чувствительность) или ( D ) к отсутствию флуоресценции.Вставка 61-пн (спейсер 33 пн/повтор 28 пн) в массив CRISPR сдвигает стоп-кодон UAA в лидере за пределы рамки и восстанавливает рамку считывания репортерного гена, что приводит к устойчивости к хлорамфениколу ( C ) или флуоресценция ( D ) клеток с расширенными массивами CRISPR.
Состав репортерных мРНК показан ниже. Каждый прямоугольник соответствует одному кодону; rbs — сайт связывания рибосомы; AUG — стартовый кодон, UAA — стоп-кодон, расположенный в рамке с AUG. Трансляция мРНК, транскрибированных с нерасширенных репортеров CRISPR, останавливается на кодоне UAA в лидерной последовательности, что приводит к ( C ) чувствительности к хлорамфениколу (Cm-чувствительность) или ( D ) к отсутствию флуоресценции.Вставка 61-пн (спейсер 33 пн/повтор 28 пн) в массив CRISPR сдвигает стоп-кодон UAA в лидере за пределы рамки и восстанавливает рамку считывания репортерного гена, что приводит к устойчивости к хлорамфениколу ( C ) или флуоресценция ( D ) клеток с расширенными массивами CRISPR.
Две мощные экспериментальные системы были разработаны для преодоления этих ограничений и повышения чувствительности обнаружения событий приобретения спейсеров [36,37] (). Обе системы основаны на репортерном гене, введенном выше лидерной последовательности специально разработанного миниатюрного штамма E. coli массив CRISPR. Репортер транскрибируется с промотора, расположенного ниже матрицы, в направлении, противоположном направлению транскрипции матрицы CRISPR, инициируемой лидером. Полученная мРНК включает стартовый кодон, за которым следует сегмент массива лидер-CRISPR (клонированный в обратной ориентации) и последовательность репортера, которая не имеет собственного начала трансляции. В клетках с нерасширенными массивами CRISPR трансляция репортерной ORF предотвращается из-за внутрирамочного стоп-кодона внутри лидера.Вставка дополнительной единицы длиной 61 п.н. (спейсер 33 п.н./повтор 28 п.н.) изменяет рамку считывания и позволяет синтезировать репортер, приводящий либо к устойчивости к хлорамфениколу [36] (), либо к флуоресценции [37] () клеток что приобрел распорку. Редкие колонии, устойчивые к хлорамфениколу, могут быть подвергнуты прямому скринингу на расширение массива CRISPR с помощью ПЦР. В системе на основе флуоресцентного белка используется живая флуоресцентная микроскопия для наблюдения и количественного определения клеток, которые приобрели спейсеры [37].
coli массив CRISPR. Репортер транскрибируется с промотора, расположенного ниже матрицы, в направлении, противоположном направлению транскрипции матрицы CRISPR, инициируемой лидером. Полученная мРНК включает стартовый кодон, за которым следует сегмент массива лидер-CRISPR (клонированный в обратной ориентации) и последовательность репортера, которая не имеет собственного начала трансляции. В клетках с нерасширенными массивами CRISPR трансляция репортерной ORF предотвращается из-за внутрирамочного стоп-кодона внутри лидера.Вставка дополнительной единицы длиной 61 п.н. (спейсер 33 п.н./повтор 28 п.н.) изменяет рамку считывания и позволяет синтезировать репортер, приводящий либо к устойчивости к хлорамфениколу [36] (), либо к флуоресценции [37] () клеток что приобрел распорку. Редкие колонии, устойчивые к хлорамфениколу, могут быть подвергнуты прямому скринингу на расширение массива CRISPR с помощью ПЦР. В системе на основе флуоресцентного белка используется живая флуоресцентная микроскопия для наблюдения и количественного определения клеток, которые приобрели спейсеры [37]. Хотя это еще не реализовано, использование FACS (сортировка клеток, активируемая флуоресценцией) должно позволить обогатить популяцию клеток расширенными массивами для последующего анализа.В обеих системах приобретение спейсеров, несущих стоп-кодоны, расположенные в рамке считывания репортера, остается незамеченным. Аналогично, включение более чем одной единицы повтора спейсера или включение одного нестандартного спейсера, который не может восстановить рамку считывания репортера, не будет обнаружено. Наконец, системы CRISPR-Cas, в которых включение стандартной единицы спейсер-повтор не сдвигает рамку считывания (т. е. вводит вставку длиной n × 3 п.н., где n — целое число нуклеотидов), не подлежат изучению. .
Хотя это еще не реализовано, использование FACS (сортировка клеток, активируемая флуоресценцией) должно позволить обогатить популяцию клеток расширенными массивами для последующего анализа.В обеих системах приобретение спейсеров, несущих стоп-кодоны, расположенные в рамке считывания репортера, остается незамеченным. Аналогично, включение более чем одной единицы повтора спейсера или включение одного нестандартного спейсера, который не может восстановить рамку считывания репортера, не будет обнаружено. Наконец, системы CRISPR-Cas, в которых включение стандартной единицы спейсер-повтор не сдвигает рамку считывания (т. е. вводит вставку длиной n × 3 п.н., где n — целое число нуклеотидов), не подлежат изучению. .
Обнаружение адаптации CRISPR в клеточных популяциях
В ранних исследованиях адаптации последовательности вновь приобретенных спейсеров определяли для отдельных колоний с помощью секвенирования по Сэнгеру [4,21–23,30–33]. Для анализа миллионов CRISPR-массивов в одном эксперименте обычно используют высокопроизводительное секвенирование (HTS) [38–40]. Это позволяет изучать отклонения в длине спейсеров, распределение соответствующих протоспейсеров по разным источникам ДНК и их нуклеотидный состав [34,35,38-52].В принципе, при достаточной глубине секвенирования HTS тотальной геномной ДНК, очищенной от культуры, должны выявлять чтения, соответствующие расширенным массивам [53]. В модельной системе из культур E. coli , сверхпродуцирующих комплекс адаптационных белков Cas1-Cas2 и трансформированных олигонуклеотидами спейсерного размера, покрытие генома ~350× позволило уверенно обнаружить расширение массива CRISPR, которое произошло примерно в 10% клеток [54]. . Более того, также были обнаружены более редкие нецелевые интеграционные события в других частях генома [54].
Это позволяет изучать отклонения в длине спейсеров, распределение соответствующих протоспейсеров по разным источникам ДНК и их нуклеотидный состав [34,35,38-52].В принципе, при достаточной глубине секвенирования HTS тотальной геномной ДНК, очищенной от культуры, должны выявлять чтения, соответствующие расширенным массивам [53]. В модельной системе из культур E. coli , сверхпродуцирующих комплекс адаптационных белков Cas1-Cas2 и трансформированных олигонуклеотидами спейсерного размера, покрытие генома ~350× позволило уверенно обнаружить расширение массива CRISPR, которое произошло примерно в 10% клеток [54]. . Более того, также были обнаружены более редкие нецелевые интеграционные события в других частях генома [54].
Несмотря на очевидную эффективность и беспристрастность, метод дробового секвенирования требует высокого охвата секвенирования и обеспечивает очень небольшое (десятки) количество прочтений, соответствующих расширенным массивам, что делает его непригодным для исследований, направленных на качественное понимание предпочтений выбора спейсеров [54]. Поэтому общепринятой стратегией является подготовка ПЦР-ампликонов массивов из культур, подвергающихся адаптации CRISPR, а затем их подвергание HTS [38–40]. Гель-электрофорез используется для отделения ампликонов исходных нерасширенных массивов CRISPR (+0) от тех, которые приобрели одну (+1), две (+2) или более единицы спейсерных повторов ().
Поэтому общепринятой стратегией является подготовка ПЦР-ампликонов массивов из культур, подвергающихся адаптации CRISPR, а затем их подвергание HTS [38–40]. Гель-электрофорез используется для отделения ампликонов исходных нерасширенных массивов CRISPR (+0) от тех, которые приобрели одну (+1), две (+2) или более единицы спейсерных повторов ().
На всех панелях нерасширенные массивы CRISPR помечены как UA; расширенные массивы — советник, лидер показан светло-персиковым прямоугольником; повторы — в виде белых прямоугольников; ранее существовавшая проставка S 0 — в виде темно-персикового прямоугольника; недавно приобретенная проставка S +1 — в виде бирюзового прямоугольника. Праймеры показаны стрелками под массивами CRISPR. Перевернутый конец стрелки праймера указывает на отсутствие комплементарности между концом праймера и матрицей.Скобки под парами праймеров указывают продукты ПЦР (штриховые линии соответствуют низкоэффективной ПЦР с вырожденными праймерами, 3′-концы которых не комплементарны матрице). ( A ) Стандартный анализ получения спейсера на основе ПЦР. Лидерно-проксимальная часть массива CRISPR амплифицируется с помощью праймера, специфичного для лидера (светлая персиковая стрелка), и праймера, отжигающего со спейсером S 0 (темно-персиковая стрелка). Продукты ПЦР разделяют на агарозном геле. M, маркер размера молекулярной массы.Полоса, соответствующая расширенным массивам CRISPR (EA), очищается от геля и подвергается HTS (высокопроизводительному секвенированию). ( B ) Селективная амплификация расширенных массивов с помощью ПЦР с праймером, специфичным для лидера, и праймером, специфичным для вырожденного повтора (белые стрелки с бирюзовыми наконечниками стрелок). Все нуклеотиды вырожденного праймера, кроме последнего, отжигают до повтора, помещая 3′-концевой нуклеотид напротив последнего нуклеотида лидера или последнего нуклеотида приобретенного спейсера.Последнее положение вырожденного праймера включает один из трех нуклеотидов, не комплементарных последнему нуклеотиду лидера, что позволяет проводить эффективную амплификацию только тогда, когда последний нуклеотид приобретенного спейсера отличается от последнего нуклеотида лидера.
( A ) Стандартный анализ получения спейсера на основе ПЦР. Лидерно-проксимальная часть массива CRISPR амплифицируется с помощью праймера, специфичного для лидера (светлая персиковая стрелка), и праймера, отжигающего со спейсером S 0 (темно-персиковая стрелка). Продукты ПЦР разделяют на агарозном геле. M, маркер размера молекулярной массы.Полоса, соответствующая расширенным массивам CRISPR (EA), очищается от геля и подвергается HTS (высокопроизводительному секвенированию). ( B ) Селективная амплификация расширенных массивов с помощью ПЦР с праймером, специфичным для лидера, и праймером, специфичным для вырожденного повтора (белые стрелки с бирюзовыми наконечниками стрелок). Все нуклеотиды вырожденного праймера, кроме последнего, отжигают до повтора, помещая 3′-концевой нуклеотид напротив последнего нуклеотида лидера или последнего нуклеотида приобретенного спейсера.Последнее положение вырожденного праймера включает один из трех нуклеотидов, не комплементарных последнему нуклеотиду лидера, что позволяет проводить эффективную амплификацию только тогда, когда последний нуклеотид приобретенного спейсера отличается от последнего нуклеотида лидера. ( C ) Выборочное усиление расширенных массивов с помощью ЗАХВАТА. Вверху массивы CRISPR амплифицируются, а полоса, соответствующая расширенным массивам CRISPR, очищается после электрофореза, как в A . Внизу, чтобы еще больше увеличить процент расширенных массивов в библиотеках секвенирования, продукты первой ПЦР повторно амплифицируют с праймерами (белые стрелки), отжигая до повторов.Расширенные массивы селективно амплифицируются, так как продукты первой стадии ПЦР нерасширенных и расширенных массивов содержат один и два повтора соответственно. ( D ) Селективное усиление расширенных массивов с помощью SENECA. Сайт узнавания эндонуклеазы FaqI (бордовый прямоугольник) вводится сразу после первого повтора CRISPR. FaqI расщепляет ДНК выше своего сайта узнавания, создавая липкие концы в повторяющейся последовательности. К этому липкому концу лигируют соответствующий адаптер и проводят ПЦР-амплификацию с праймерами, специфичными для адаптера (светло-синий) и повторно совпадающими (белый) праймерами.
( C ) Выборочное усиление расширенных массивов с помощью ЗАХВАТА. Вверху массивы CRISPR амплифицируются, а полоса, соответствующая расширенным массивам CRISPR, очищается после электрофореза, как в A . Внизу, чтобы еще больше увеличить процент расширенных массивов в библиотеках секвенирования, продукты первой ПЦР повторно амплифицируют с праймерами (белые стрелки), отжигая до повторов.Расширенные массивы селективно амплифицируются, так как продукты первой стадии ПЦР нерасширенных и расширенных массивов содержат один и два повтора соответственно. ( D ) Селективное усиление расширенных массивов с помощью SENECA. Сайт узнавания эндонуклеазы FaqI (бордовый прямоугольник) вводится сразу после первого повтора CRISPR. FaqI расщепляет ДНК выше своего сайта узнавания, создавая липкие концы в повторяющейся последовательности. К этому липкому концу лигируют соответствующий адаптер и проводят ПЦР-амплификацию с праймерами, специфичными для адаптера (светло-синий) и повторно совпадающими (белый) праймерами.
Основной проблемой внутрикультурных методов выявления адаптации CRISPR на основе ПЦР является их низкая чувствительность из-за более эффективной амплификации более коротких (а в большинстве интересных случаев гораздо более распространенных) нерасширенных массивов CRISPR [55]. В случае системы E. coli типа I-E стандартный метод позволяет достоверно выявлять расширенные массивы ампликонов только в культурах, содержащих, по нашему опыту, не менее 5% адаптированных клеток. Разработано несколько модификаций, направленных на повышение чувствительности.Самый простой основан на амплификации с помощью лидер-специфического праймера и праймера, соответствующего вновь полученному спейсеру, последовательность которого известна [40]. После калибровки по количеству продукта ПЦР, амплифицированного из области за пределами массива CRISPR, и отражая общее количество молекул ДНК в образце, этот метод можно использовать для определения эффективности адаптации с помощью количественной ПЦР [56]. Очевидным недостатком этого метода является то, что он требует предварительных знаний о приобретенных спейсерах и, следовательно, не может применяться для изучения приобретения спейсеров в системах с неизвестными адаптационными предпочтениями.Однако он очень эффективен при изучении приобретения олигонуклеотидов размером со спейсер, трансформированных в клетки [51, 54, 57].
Очевидным недостатком этого метода является то, что он требует предварительных знаний о приобретенных спейсерах и, следовательно, не может применяться для изучения приобретения спейсеров в системах с неизвестными адаптационными предпочтениями.Однако он очень эффективен при изучении приобретения олигонуклеотидов размером со спейсер, трансформированных в клетки [51, 54, 57].
Другая модификация использует лидер-специфический праймер и «вырожденные» праймеры, соответствующие последовательности повтора и содержащие один дополнительный 3′-концевой нуклеотид () [42]. Дополнительная позиция содержит в равных пропорциях три нуклеотида, кроме одного, комплементарного последнему нуклеотиду лидера. В то время как продукты амплификации ожидаются только в том случае, если (i) был получен спейсер и (ii) его последний нуклеотид отличается от последнего нуклеотида лидера, на практике все еще наблюдаются ампликоны из нерасширенных массивов [42].Сообщалось, что этот метод обнаруживает всего 0,01% клеток с расширенными массивами [42]. Однако по замыслу до ~25% приобретенных спейсеров остаются необнаруженными. Более того, метод эффективен только при применении к сконструированным миниатюрным массивам CRISPR, сокращенным до одного повтора, поскольку из нерасширенных массивов с множеством повторов получаются множественные продукты амплификации, которые невозможно отличить от ампликонов из расширенных массивов.
Однако по замыслу до ~25% приобретенных спейсеров остаются необнаруженными. Более того, метод эффективен только при применении к сконструированным миниатюрным массивам CRISPR, сокращенным до одного повтора, поскольку из нерасширенных массивов с множеством повторов получаются множественные продукты амплификации, которые невозможно отличить от ампликонов из расширенных массивов.
Реамплификация гель-очищенных ампликонов расширенных массивов позволяет повысить чувствительность обнаружения адаптации CRISPR в культурах клеток [58].Продукты стандартной амплификации разделяют гель-электрофорезом и очищают. Стадию реамплификации повторяют до тех пор, пока на геле не станет отчетливо виден фрагмент ожидаемой длины (). Использование автоматизированной системы BluePippin (электрофорез в агарозном геле с автоматическим элюированием для выбора размера) позволяет улучшить качество разделения, снизить контаминацию нерасширенными массивами ампликонов и повысить воспроизводимость анализа. Даже когда ампликоны расширенных массивов невидимы после первого электрофоретического разделения, ДНК, извлеченная из соответствующего положения геля, может быть использована для повторной амплификации. Сообщается, что в зависимости от набора праймеров, предназначенных для реамплификации («внутренние», «дегенеративные» или «повторноспецифичные»), этот метод выявляет, соответственно, 1, 0,01 и 0,1% клеток с расширенными массивами в пределах E. .coli культуры. «Внутренние» праймеры могут быть либо такими же, как те, которые использовались во время начальной амплификации, либо для повышения специфичности и предотвращения амплификации ДНК, отличной от CRISPR, можно использовать лидер-специфический «вложенный» праймер для повторной амплификации, отжигающий ближе к матрице.В любом случае усиление нерасширенных массивов, совместно очищенных с расширенными, не подавляется. «Вырожденные» праймеры избирательно подавляют повторную амплификацию нерасширенного массива, как описано выше. Повторная амплификация с «специфическими для повтора» праймерами основывается на том факте, что ампликоны, соответствующие расширенным массивам, имеют два повтора после первого шага ПЦР (2). Таким образом, амплификация с праймерами, соответствующими половинам повторяющейся последовательности, дает продукт ПЦР только для расширенных массивов.
Сообщается, что в зависимости от набора праймеров, предназначенных для реамплификации («внутренние», «дегенеративные» или «повторноспецифичные»), этот метод выявляет, соответственно, 1, 0,01 и 0,1% клеток с расширенными массивами в пределах E. .coli культуры. «Внутренние» праймеры могут быть либо такими же, как те, которые использовались во время начальной амплификации, либо для повышения специфичности и предотвращения амплификации ДНК, отличной от CRISPR, можно использовать лидер-специфический «вложенный» праймер для повторной амплификации, отжигающий ближе к матрице.В любом случае усиление нерасширенных массивов, совместно очищенных с расширенными, не подавляется. «Вырожденные» праймеры избирательно подавляют повторную амплификацию нерасширенного массива, как описано выше. Повторная амплификация с «специфическими для повтора» праймерами основывается на том факте, что ампликоны, соответствующие расширенным массивам, имеют два повтора после первого шага ПЦР (2). Таким образом, амплификация с праймерами, соответствующими половинам повторяющейся последовательности, дает продукт ПЦР только для расширенных массивов.
Конвейер SENECA [59] выборочно усиливает расширенные массивы CRISPR.В основе метода лежит конструирование плазмидного массива CRISPR с сайтом узнавания эндонуклеазы FaqI непосредственно за миниатюризированным «массивом» CRISPR, состоящим из одного повтора, которому предшествует лидерная последовательность (). В отличие от большинства эндонуклеаз рестрикции типа II, FaqI, фермент типа II-S, расщепляет ДНК за пределами сайта узнавания, образуя липкий конец. Матрица CRISPR, используемая в SENECA, разработана таким образом, что распознавание сайта FaqI приводит к расщеплению восходящего повтора.Адаптер Illumina с липким концом, комплементарным генерируемому FaqI, лигируется, и ПЦР с парой праймеров, один комплементарный повтору, а другой — адаптеру, избирательно амплифицирует расширенные массивы, так как исходная последовательность повторов теряется после обработки FaqI и таким образом, ампликоны из нерасширенных массивов не амплифицируются. Хотя опубликованный протокол SENECA основан на использовании FaqI, вместо FaqI можно было бы использовать другие эндонуклеазы рестрикции типа II-S.
Первоначальный протокол SENECA был применен для анализа спейсеров, полученных системой CRISPR-Cas типа III Fusicatenibacter saccharivorans , гетерологично экспрессированных в E.coli [59]. Хотя сообщалось о примерно 700-кратном обогащении расширенных массивов в библиотеке секвенирования, чувствительность SENECA с точки зрения определения процента клеток с расширенными массивами не определялась. Чтобы сравнить SENECA с другими методами обнаружения адаптации CRISPR, мы сконструировали штамм E. coli со спейсером, содержащим сайт распознавания FaqI, S FaqI , включенным в массив геномного типа I-E CRISPR (и дополнительный рисунок S1A). За исключением S FaqI , штамм изогенен для KD263, штамма с генами cas под контролем индуцируемых промоторов и миниатюрным массивом CRISPR с одним спейсером (дополнительная рис.S1A и дополнительную таблицу S1). KD263 широко использовался для изучения различных аспектов адаптации CRISPR [41, 49, 56, 60–64], и сообщалось об эффективности адаптации до 50–80% в условиях прайминга [41, 62]. Для сравнения чувствительности стандартной ПЦР-детекции и СЕНЕКА ДНК, полученную из клеток с нерасширенным массивом CRISPR, содержащих спейсер S FaqI , смешивали в разных пропорциях с ДНК, полученной из клеток, содержащих дополнительную единицу спейсер-повтор (). При стандартном протоколе расширенные массивы были четко обнаружены, когда 10% ДНК имели дополнительный спейсер.С SENECA ожидаемый продукт амплификации все еще наблюдался, когда ДНК с расширенным массивом составляла 0,4% от общего количества, что примерно в 25 раз увеличивает чувствительность (). Чтобы проверить, вносит ли SENECA систематические ошибки в амплификацию отдельных спейсеров, мы проанализировали спейсеры, полученные во время адаптации с праймированием типа I-E по стандартному протоколу или SENECA (дополнительный рис. S1). Между двумя методами не было обнаружено никаких изменений в схемах выбора прокладок (дополнительная рис. S1C).
Для сравнения чувствительности стандартной ПЦР-детекции и СЕНЕКА ДНК, полученную из клеток с нерасширенным массивом CRISPR, содержащих спейсер S FaqI , смешивали в разных пропорциях с ДНК, полученной из клеток, содержащих дополнительную единицу спейсер-повтор (). При стандартном протоколе расширенные массивы были четко обнаружены, когда 10% ДНК имели дополнительный спейсер.С SENECA ожидаемый продукт амплификации все еще наблюдался, когда ДНК с расширенным массивом составляла 0,4% от общего количества, что примерно в 25 раз увеличивает чувствительность (). Чтобы проверить, вносит ли SENECA систематические ошибки в амплификацию отдельных спейсеров, мы проанализировали спейсеры, полученные во время адаптации с праймированием типа I-E по стандартному протоколу или SENECA (дополнительный рис. S1). Между двумя методами не было обнаружено никаких изменений в схемах выбора прокладок (дополнительная рис. S1C).
 палочка .
палочка . ( A ) Для создания штамма E. coli , пригодного для обнаружения праймированной адаптации с помощью SENECA, родительские клетки E. coli KD263, содержащие индуцируемые гены cas , и массив CRISPR с одним спейсером S g8 трансформируют плазмидой, несущей затравочный протоспейсер PPS g8 , соответствующий S g8 , и сайт FaqI, введенный в ранее охарактеризованный широко используемый «горячий» протоспейсер (PS FaqI ), показанный бирюзовым прямоугольником.После распознавания PPS g8 эффекторным комплексом Cascade-crRNA происходит первичная адаптация, и новые спейсеры, происходящие из плазмиды, интегрируются в массив CRISPR [31,41,65]. Среди колоний, которые потеряли плазмиду и расширили свой массив CRISPR (см. ), выбирают клон, который приобрел спейсер S FaqI . ( B ) Геномную ДНК, очищенную от родительских клеток, несущих спейсер S FaqI или производное с расширенным массивом (+1), смешивали в указанных соотношениях, и состояние массивов CRISPR оценивали с помощью стандартной ПЦР (см. ) или SENECA. (видеть ).Процент +1 геномной ДНК в образце указан над гелем. С левой стороны ампликоны, соответствующие нерасширенному и расширенному массивам, выявленным с помощью стандартной ПЦР, помечены соответственно +0 и +1. Ампликон расширенного массива, сгенерированный SENECA, также помечен как +1. Нижняя минорная полоса (серый треугольник) соответствует результатам линейного усиления родительского массива [59].
) или SENECA. (видеть ).Процент +1 геномной ДНК в образце указан над гелем. С левой стороны ампликоны, соответствующие нерасширенному и расширенному массивам, выявленным с помощью стандартной ПЦР, помечены соответственно +0 и +1. Ампликон расширенного массива, сгенерированный SENECA, также помечен как +1. Нижняя минорная полоса (серый треугольник) соответствует результатам линейного усиления родительского массива [59].
Подавляющая ПЦР
В качестве еще одной альтернативы мы стремились повысить эффективность обнаружения редких событий адаптации путем селективной амплификации более длинных фрагментов с помощью подавляющей ПЦР (супПЦР), основанной на амплификации фрагментов, фланкированных инвертированными концевыми повторами длиной около 30 нуклеотидов ( ИТР) [66,67].Фрагменты ДНК, содержащие ITR, образуют паноподобные структуры за счет внутримолекулярной гибридизации (). Поскольку более короткие фрагменты с большей вероятностью образуют pan-подобные структуры, отжиг праймера, идентичного ITR и используемого на втором этапе реамплификации, ингибируется [68]. Чтобы определить, может ли supPCR повысить чувствительность обнаружения приобретения спейсера, мы использовали стандартные праймеры, специфичные для лидера и первого спейсера, дополненные ITR. Как видно из дополнительного рисунка S2, предпочтительная амплификация расширенных массивов CRISPR на втором этапе ПЦР с праймером ITR повысила чувствительность обнаружения событий адаптации примерно в 10 раз.
Чтобы определить, может ли supPCR повысить чувствительность обнаружения приобретения спейсера, мы использовали стандартные праймеры, специфичные для лидера и первого спейсера, дополненные ITR. Как видно из дополнительного рисунка S2, предпочтительная амплификация расширенных массивов CRISPR на втором этапе ПЦР с праймером ITR повысила чувствительность обнаружения событий адаптации примерно в 10 раз.
Схематическое представление анализа supPCR для обнаружения адаптации CRISPR представлено в A . Показаны обе нити расширенного (EA) и нерасширенного (UA) массивов CRISPR. В первой стадии ПЦР (I) используется один праймер, соответствующий лидеру массива CRISPR (бежевая стрелка), и другой, соответствующий ранее существовавшему проксимальному спейсеру S 0 (бордовая стрелка). Оба праймера несут на своих 5′-концах идентичные 30-нуклеотидные последовательности (показаны сиреневым цветом), которые после амплификации становятся ITR. Второй этап ПЦР (II) проводят с использованием праймера, последовательность которого совпадает с последовательностью ITR. Ампликоны, соответствующие расширенным массивам, с меньшей вероятностью подвергаются внутримолекулярной гибридизации и, следовательно, более эффективно амплифицируются с праймером ITR. Праймеры показаны стрелками под или над массивами CRISPR. В скобках под массивами CRISPR указаны области, амплифицированные с помощью ПЦР (штриховая линия соответствует низкоэффективной ПЦР на матрице UA). ( B ) Геномная ДНК, очищенная от штамма E.coli с одним дополнительным спейсером (+1) смешивали в указанных пропорциях с ДНК штамма с нерасширенным (+0) массивом. Смеси использовали в качестве матриц для супПЦР. Показаны продукты «стандартной» амплификации (), а также продукты первого (I) и второго (II) этапов супПЦР, разделенные электрофорезом в агарозном геле.
Второй этап ПЦР (II) проводят с использованием праймера, последовательность которого совпадает с последовательностью ITR. Ампликоны, соответствующие расширенным массивам, с меньшей вероятностью подвергаются внутримолекулярной гибридизации и, следовательно, более эффективно амплифицируются с праймером ITR. Праймеры показаны стрелками под или над массивами CRISPR. В скобках под массивами CRISPR указаны области, амплифицированные с помощью ПЦР (штриховая линия соответствует низкоэффективной ПЦР на матрице UA). ( B ) Геномная ДНК, очищенная от штамма E.coli с одним дополнительным спейсером (+1) смешивали в указанных пропорциях с ДНК штамма с нерасширенным (+0) массивом. Смеси использовали в качестве матриц для супПЦР. Показаны продукты «стандартной» амплификации (), а также продукты первого (I) и второго (II) этапов супПЦР, разделенные электрофорезом в агарозном геле.
Отношение матрицы ДНК к праймеру ITR, используемому на втором этапе, является критическим параметром для успешной супПЦР, поскольку подавление зависит от конкуренции между внутримолекулярной гибридизацией ITR и отжигом праймера ITR для амплификации целевого продукта (дополнительная рис. S2A) [68]. Это зависит от длины спейсерного повтора, включенного в расширенный массив, и должно быть определено экспериментально. Поскольку один и тот же ITR может использоваться для амплификации массивов CRISPR из разных систем, другие параметры, такие как температура отжига и длина супрессорного праймера, остаются одинаковыми для разных реакций амплификации. Мы применили supPCR для анализа спейсеров, полученных во время праймированной адаптации системой E. coli типа IE, и обнаружили аналогичные паттерны приобретенных спейсеров для стандартного протокола и supPCR (дополнительная рис.С3). В дополнение к системе E. coli типа IE мы успешно использовали supPCR для выявления приобретения спейсера в массивах Thermus thermophilus CRISPR типа III (наши неопубликованные наблюдения).
S2A) [68]. Это зависит от длины спейсерного повтора, включенного в расширенный массив, и должно быть определено экспериментально. Поскольку один и тот же ITR может использоваться для амплификации массивов CRISPR из разных систем, другие параметры, такие как температура отжига и длина супрессорного праймера, остаются одинаковыми для разных реакций амплификации. Мы применили supPCR для анализа спейсеров, полученных во время праймированной адаптации системой E. coli типа IE, и обнаружили аналогичные паттерны приобретенных спейсеров для стандартного протокола и supPCR (дополнительная рис.С3). В дополнение к системе E. coli типа IE мы успешно использовали supPCR для выявления приобретения спейсера в массивах Thermus thermophilus CRISPR типа III (наши неопубликованные наблюдения).
Обнаружение предшественников спейсеров
in vivo Методы, описанные выше, обнаруживают конечный результат адаптации CRISPR, спейсеры, интегрированные в массив CRISPR. Новые спейсеры образуются из «преспейсеров», коротких фрагментов ДНК, которые генерируются внутри клетки с помощью различных процессов, а затем направляются в массивы CRISPR с помощью белков Cas1 и Cas2 [24, 25, 34, 50–52, 69–75].Знание структуры преспейсера необходимо для понимания предпочтений адаптации CRISPR и возможности их контролировать. In vitro , комплекс Cas1-Cas2 предпочтительно использует фрагменты двухцепочечной ДНК с тупыми концами или выступающими концами 3′ в качестве субстратов для интеграции [24,70,76-78]. Шипман и др. были первыми, кто проверил требования к преспейсерам для интеграции in vivo [57]. Они продемонстрировали, что двухцепочечные синтетические олигонуклеотиды электропорируются в E.coli , экспрессирующие тип I-E cas1 и cas2 , можно обрезать до размера канонического спейсера E. coli (33 п.н.) и интегрировать в массив CRISPR () [57]. Присутствие консенсусной последовательности PAM (AAG/CTT) увеличивало эффективность приобретения преспейсеров примерно в 5 раз и обеспечивало специфическую ориентацию интеграции с G/C, полученными из PAM, сразу после первого повтора [57].
Новые спейсеры образуются из «преспейсеров», коротких фрагментов ДНК, которые генерируются внутри клетки с помощью различных процессов, а затем направляются в массивы CRISPR с помощью белков Cas1 и Cas2 [24, 25, 34, 50–52, 69–75].Знание структуры преспейсера необходимо для понимания предпочтений адаптации CRISPR и возможности их контролировать. In vitro , комплекс Cas1-Cas2 предпочтительно использует фрагменты двухцепочечной ДНК с тупыми концами или выступающими концами 3′ в качестве субстратов для интеграции [24,70,76-78]. Шипман и др. были первыми, кто проверил требования к преспейсерам для интеграции in vivo [57]. Они продемонстрировали, что двухцепочечные синтетические олигонуклеотиды электропорируются в E.coli , экспрессирующие тип I-E cas1 и cas2 , можно обрезать до размера канонического спейсера E. coli (33 п.н.) и интегрировать в массив CRISPR () [57]. Присутствие консенсусной последовательности PAM (AAG/CTT) увеличивало эффективность приобретения преспейсеров примерно в 5 раз и обеспечивало специфическую ориентацию интеграции с G/C, полученными из PAM, сразу после первого повтора [57].
( А ) Электропорация двухцепочечных олигонуклеотидов (бирюзового цвета) в клетки с CRISPR-массивом (лидер CRISPR — светло-персиковый прямоугольник; повторы — белые прямоугольники; ранее существовавший спейсер — темно-персиковый прямоугольник). Cas1 и Cas2 (светло-зеленые овалы) образуют комплекс, который связывает олигонуклеотиды, которые могут функционировать как преспейсеры, и интегрирует их в массив CRISPR в качестве новых спейсеров (бирюзовый прямоугольник). ( B ) Конвейер FragSeq. Высокомолекулярную ДНК выделяют из клеток вместе с короткими фрагментами (цепи двухцепочечной ДНК показаны синим и красным цветом).Короткие фрагменты ДНК извлекают, денатурируют и лигируют с одноцепочечными адаптерами (светло-голубые и светло-зеленые прямоугольники). Полученные молекулы амплифицируют праймерами со штрих-кодом (голубая и светло-зеленая стрелки). ( C ) Генерация преспейсера во время праймированной адаптации типа I-E в E. coli , как показано с помощью FragSeq [79] и удлинения праймера [63]. Преспейсер (синие и красные прямоугольники) в ДНК (синие и красные линии) связан комплексом Cas1-Cas2 (светло-зеленый). На вставке показан состав нуклеотидов на конце преспейсера, полученном из PAM, согласно FragSeq.Красная цепь перед PAM деградирует (пунктирная линия), что предотвращает отжиг праймера, используемого для удлинения праймера (синяя стрелка). Синяя нить надрезана между AA и G AAG PAM. Реакция удлинения праймера с отжигом праймера до синей нити (красная стрелка) ниже преспейсера приводит к образованию двух продуктов удлинения (показаны тонкими черными стрелками), что означает, что 5′-конец, полученный из PAM, образуется первым, и что PAM- дистальный 3′-конец образуется в результате эндонуклеолитического расщепления.Механизм образования 5′-концов преспейсера неизвестен (двойные сплошные/пунктирные линии со знаком вопроса).
coli , как показано с помощью FragSeq [79] и удлинения праймера [63]. Преспейсер (синие и красные прямоугольники) в ДНК (синие и красные линии) связан комплексом Cas1-Cas2 (светло-зеленый). На вставке показан состав нуклеотидов на конце преспейсера, полученном из PAM, согласно FragSeq.Красная цепь перед PAM деградирует (пунктирная линия), что предотвращает отжиг праймера, используемого для удлинения праймера (синяя стрелка). Синяя нить надрезана между AA и G AAG PAM. Реакция удлинения праймера с отжигом праймера до синей нити (красная стрелка) ниже преспейсера приводит к образованию двух продуктов удлинения (показаны тонкими черными стрелками), что означает, что 5′-конец, полученный из PAM, образуется первым, и что PAM- дистальный 3′-конец образуется в результате эндонуклеолитического расщепления.Механизм образования 5′-концов преспейсера неизвестен (двойные сплошные/пунктирные линии со знаком вопроса).
Мы разработали метод специфичного для нити HTS коротких фрагментов ДНК, сгенерированных in vivo , который мы назвали FragSeq и который позволил нам обнаружить преспейсеры в клетках E. coli , подвергшихся примированной адаптации CRISPR () [79]. Метод использует протоколы, которые позволяют избежать потери коротких фрагментов во время очистки ДНК из клеточных лизатов (очистка ДНК фенол-хлороформом) и выбора размера фрагментов размера спейсера перед подготовкой библиотеки.Наш опыт показывает, что даже если всего лишь 0,1% геномной ДНК фрагментируется случайным образом во время очистки ДНК и выбора размера, это создает фоновый шум, который усложняет обнаружение специфического сигнала от коротких фрагментов ДНК, присутствующих в клетке. Чтобы избежать этого осложнения, эксперимент FragSeq должен быть спланирован таким образом, чтобы специфический сигнал от определенной области генома/плазмиды, из которой ожидаются преспейсеры, сравнивался с контрольной областью, неспецифически фрагментированной во время последующей обработки.В нашем случае мы достигаем этого с помощью самонацеливания клеток E. coli , в которых crRNA направляет механизм интерференции на распознавание протоспейсера, расположенного в бактериальном геноме.
coli , подвергшихся примированной адаптации CRISPR () [79]. Метод использует протоколы, которые позволяют избежать потери коротких фрагментов во время очистки ДНК из клеточных лизатов (очистка ДНК фенол-хлороформом) и выбора размера фрагментов размера спейсера перед подготовкой библиотеки.Наш опыт показывает, что даже если всего лишь 0,1% геномной ДНК фрагментируется случайным образом во время очистки ДНК и выбора размера, это создает фоновый шум, который усложняет обнаружение специфического сигнала от коротких фрагментов ДНК, присутствующих в клетке. Чтобы избежать этого осложнения, эксперимент FragSeq должен быть спланирован таким образом, чтобы специфический сигнал от определенной области генома/плазмиды, из которой ожидаются преспейсеры, сравнивался с контрольной областью, неспецифически фрагментированной во время последующей обработки.В нашем случае мы достигаем этого с помощью самонацеливания клеток E. coli , в которых crRNA направляет механизм интерференции на распознавание протоспейсера, расположенного в бактериальном геноме. После индукции самонацеливания клетки перестают делиться, но остаются живыми в течение как минимум нескольких часов. Чтобы гарантировать обнаружение фрагментов с различными концевыми структурами, требуется одноцепочечное специфическое секвенирование для правильного определения длины каждой цепи преспейсера, а это означает, что адаптеры необходимо лигировать как с одноцепочечными, так и с двухцепочечными концами фрагментов преспейсера в библиотеке. стадия строительства.Мы использовали комбинацию термостабильной 5′-App ДНК/РНК-лигазы (NEB), которая представляет собой вариант K97A РНК-лигазы из Methanobacterium thermoautotrophicum [80], и T4 РНК-лигазы 1 (NEB). Тем не менее, мы наблюдали сильную предвзятость 5′-App ДНК / РНК-лигазы для лигирования адаптеров с фрагментами ДНК, несущими мотив CGN на их 3′-концах, что искажает библиотеки. Таким образом, возможно, что менее предвзятая Т4 РНК-лигаза 1 сама по себе может лучше подходить для создания библиотеки. В качестве альтернативы можно использовать коммерческие наборы для одноцепочечного специфического секвенирования, такие как Accel-NGS 1S Plus DNA Library Kit (Swift Biosciences).
После индукции самонацеливания клетки перестают делиться, но остаются живыми в течение как минимум нескольких часов. Чтобы гарантировать обнаружение фрагментов с различными концевыми структурами, требуется одноцепочечное специфическое секвенирование для правильного определения длины каждой цепи преспейсера, а это означает, что адаптеры необходимо лигировать как с одноцепочечными, так и с двухцепочечными концами фрагментов преспейсера в библиотеке. стадия строительства.Мы использовали комбинацию термостабильной 5′-App ДНК/РНК-лигазы (NEB), которая представляет собой вариант K97A РНК-лигазы из Methanobacterium thermoautotrophicum [80], и T4 РНК-лигазы 1 (NEB). Тем не менее, мы наблюдали сильную предвзятость 5′-App ДНК / РНК-лигазы для лигирования адаптеров с фрагментами ДНК, несущими мотив CGN на их 3′-концах, что искажает библиотеки. Таким образом, возможно, что менее предвзятая Т4 РНК-лигаза 1 сама по себе может лучше подходить для создания библиотеки. В качестве альтернативы можно использовать коммерческие наборы для одноцепочечного специфического секвенирования, такие как Accel-NGS 1S Plus DNA Library Kit (Swift Biosciences). Но следует учитывать, что эти наборы включают стадию добавления несложного хвоста на 3′-концы, что может усложнить картирование концов преспейсеров.
Но следует учитывать, что эти наборы включают стадию добавления несложного хвоста на 3′-концы, что может усложнить картирование концов преспейсеров.
В нашей работе мы использовали FragSeq для анализа преспейсеров, образующихся в ходе праймированной адаптации [79]. Отличительной чертой праймированной адаптации системой CRISPR-Cas типа I-E является приобретение спейсеров с последовательностью нетранскрибируемой цепи, происходящей из нецелевой цепи ДНК, расщепленной с помощью механизма интерференции CRISPR [30,31].В соответствии с этим мы обнаружили фрагменты длиной 32–34 нт (начиная с G или AG на их 5′-концах), которые произошли от нецелевой цепи выше праймирующего протоспейсера (PPS) за счет надрезов внутри мотива AAG [79]. Более длинные 37–38-н. фрагменты происходили из цепи-мишени и имели мотив CTTNN на 3′-концах [79]. Используя подход Shipman et al. мы проверили комплементарные пары наиболее распространенных фрагментов и пришли к выводу, что преспейсеры с тупым дистальным концом PAM, двухцепочечной областью 33–34 п. интегрированы в массив CRISPR с наибольшей эффективностью [79].Таким образом, оказывается, что асимметрия в структуре преспейсера типа IE способствует специфической ориентации интеграции преспейсера, поскольку дистальный 3′-конец PAM обрабатывается (становится тупым) раньше, чем конец, производный от PAM, и, следовательно, имеет более высокие шансы вовлечься в процесс. интеграция на границе лидер-повтор.
интегрированы в массив CRISPR с наибольшей эффективностью [79].Таким образом, оказывается, что асимметрия в структуре преспейсера типа IE способствует специфической ориентации интеграции преспейсера, поскольку дистальный 3′-конец PAM обрабатывается (становится тупым) раньше, чем конец, производный от PAM, и, следовательно, имеет более высокие шансы вовлечься в процесс. интеграция на границе лидер-повтор.
Ранее Мушарова и соавт. [63] выполнили анализ удлинения праймера на тотальной геномной ДНК, очищенной от клеток E. coli , подвергшихся праймированной адаптации, чтобы проверить, есть ли какие-либо специфические сайты расщепления рядом с двумя протоспейсерами, соответствующими часто приобретаемым спейсерам.Реакция с отжигом праймера на нецелевую цепь ниже последовательности протоспейсера выявила два продукта, один из которых соответствует ДНК, расщепляемой внутри ААГ-ПАМ, а другой — на границе ПАМ-дистальная () [63]. Это наблюдение полностью согласуется с обнаружением 32–34-н фрагментов с помощью FragSeq. Хотя с помощью FragSeq были обнаружены 37–38-н. СТТ-содержащие фрагменты, продукты удлинения праймера для цепи-мишени обнаружены не были. Возможное объяснение этого несоответствия может заключаться в том, что в то время как, по крайней мере, PAM-дистальный 3′-конец преспейсера образуется посредством эндонуклеолитического расщепления нецелевой цепи, 3′-конец, полученный из PAM, образуется благодаря экзонуклеолитическому расщеплению целевой цепи выше по течению. протоспейсера, что приводит к потере последовательности, комплементарной праймеру, использованному в Musharova et al.().
Хотя с помощью FragSeq были обнаружены 37–38-н. СТТ-содержащие фрагменты, продукты удлинения праймера для цепи-мишени обнаружены не были. Возможное объяснение этого несоответствия может заключаться в том, что в то время как, по крайней мере, PAM-дистальный 3′-конец преспейсера образуется посредством эндонуклеолитического расщепления нецелевой цепи, 3′-конец, полученный из PAM, образуется благодаря экзонуклеолитическому расщеплению целевой цепи выше по течению. протоспейсера, что приводит к потере последовательности, комплементарной праймеру, использованному в Musharova et al.().
Перспективы
Благодарности
Мы посвящаем эту статью памяти доктора Екатерины Савицкой, сыгравшей центральную роль в исследованиях CRISPR в нашей лаборатории.
Сокращения
| Ab | антибиотик |
| Bims | бактериофага нечувствительные мутанты |
| CAPTURE | адаптации CRISPR ПЦР технологии с использованием повторной амплификации и электрофореза |
| Cm | хлорамфеникол |
| CRISPR | кластерные регулярно interspaced короткие палиндромные повторы |
| EA | расширенные CRISPR массивы |
| FACS | флуоресценции активированных клеток сортировки |
| HTS | с высокой пропускной секвенирования |
| ITR | перевернутом терминал повторяет |
| мобильных генетических элементов | |
| PAM | |
| PAM | Protospacer соседний Motif |
| PIMS | плазмида мешают мутанты |
| PPS | PR IMing protospacer |
| PS | protospacer |
| руб | рибосомы сайт связывания |
| S | спейсер |
| Сенека | селективное усиление расширенных CRISPR массивов |
| UA | нераскрытый CRISPR массивы |
Вклад авторов
K. С. задумал статью. Рукопись написали А.С., К.С., И.Ф. и Д.В. А.С., И.Ф. и Д.В. подготовили рисунки. Д.В. провел эксперименты SENECA и анализ данных. ЕСЛИ. провели эксперименты с супПЦР и анализ данных.
С. задумал статью. Рукопись написали А.С., К.С., И.Ф. и Д.В. А.С., И.Ф. и Д.В. подготовили рисунки. Д.В. провел эксперименты SENECA и анализ данных. ЕСЛИ. провели эксперименты с супПЦР и анализ данных.
Финансирование
Работа выполнена при поддержке гранта 075-15-2019-1661 Министерства науки и высшего образования Российской Федерации и гранта NIH RO1 GM104071 для КС.
Конкурирующие интересы
Авторы заявляют об отсутствии конкурирующих интересов, связанных с рукописью.
Дополнительный материал
Дополнительные рисунки S1-S3:Ссылки
1. Макарова К.С., Вольф Ю.И., Алхнбаши О.С., Коста Ф., Шах С.А., Сондерс С.Дж. и другие. (2015) Обновленная эволюционная классификация систем CRISPR-Cas. Нац. Преподобный Микробиолог. 13, 722–736 10.1038/nrmicro3569 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]2. Шмаков С., Абудайе О.О., Макарова К.С., Вольф Ю.И., Гутенберг Дж.С., Семенова Е. и др. (2015)Обнаружение и функциональная характеристика различных систем CRISPR-Cas класса 2. Мол. Клетка
60, 385–397
10.1016/j.molcel.2015.10.008 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]3. Макарова К.С., Вольф Ю.И. и Кунин Е.В. (2018) Классификация и номенклатура систем CRISPR-Cas: откуда?
КРИСПР Дж.
1, 325–336
10.1089/crispr.2018.0033 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]4. Баррангу Р., Фремо К., Дево Х., Ричардс М., Бояваль П., Муано С. и др. (2007) CRISPR обеспечивает приобретенную устойчивость к вирусам у прокариот. Наука
315, 1709–1712 гг.
10.1126/science.1138140 [PubMed] [CrossRef] [Google Scholar]5. Марраффини Л.А. и Зонтхаймер Э.Дж. (2008) Интерференция CRISPR ограничивает горизонтальный перенос генов у стафилококков путем нацеливания на ДНК. Наука
322, 1843–1845 гг.
10.1126/science.1165771 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]6. Mojica F., Díez-Villaseñor C., Soria E. and Juez G. (2000) Биологическое значение семейства регулярно расположенных повторов в геномах архей, бактерий и митохондрий. Мол. микробиол.
Мол. Клетка
60, 385–397
10.1016/j.molcel.2015.10.008 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]3. Макарова К.С., Вольф Ю.И. и Кунин Е.В. (2018) Классификация и номенклатура систем CRISPR-Cas: откуда?
КРИСПР Дж.
1, 325–336
10.1089/crispr.2018.0033 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]4. Баррангу Р., Фремо К., Дево Х., Ричардс М., Бояваль П., Муано С. и др. (2007) CRISPR обеспечивает приобретенную устойчивость к вирусам у прокариот. Наука
315, 1709–1712 гг.
10.1126/science.1138140 [PubMed] [CrossRef] [Google Scholar]5. Марраффини Л.А. и Зонтхаймер Э.Дж. (2008) Интерференция CRISPR ограничивает горизонтальный перенос генов у стафилококков путем нацеливания на ДНК. Наука
322, 1843–1845 гг.
10.1126/science.1165771 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]6. Mojica F., Díez-Villaseñor C., Soria E. and Juez G. (2000) Биологическое значение семейства регулярно расположенных повторов в геномах архей, бактерий и митохондрий. Мол. микробиол. 36, 244–246
10.1046/j.1365-2958.2000.01838.x [PubMed] [CrossRef] [Google Scholar]7. Янсен Р., ван Эмбден Дж. Д. А., Гаастра В. и Шоулс Л. М. (2002) Идентификация генов, связанных с повторами ДНК у прокариот. Мол. микробиол.
43, 1565–1575 гг.
10.1046/j.1365-2958.2002.02839.x [PubMed] [CrossRef] [Google Scholar]8. Pourcel C., Touchon M., Villeriot N., Vernadet J.-P., Couvin D., Toffano-Nioche C. et al. (2019) CRISPRCasdb — преемник CRISPRdb, содержащий массивы CRISPR и гены cas из полных последовательностей генома, а также инструменты для загрузки и запроса списков повторов и спейсеров.Нуклеиновые Кислоты Res.
48, Д535–Д544
10.1093/nar/gkz915 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]9. Шмаков С.А., Ситник В., Макарова К.С., Вольф Ю.И., Северинов К.В. и Кунин Е.В. (2017) В спейсерном пространстве CRISPR преобладают последовательности видоспецифичных мобиломов. МБио
8, e01397-17
10.1128/mBio.01397-17 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]10. Танг Т.
36, 244–246
10.1046/j.1365-2958.2000.01838.x [PubMed] [CrossRef] [Google Scholar]7. Янсен Р., ван Эмбден Дж. Д. А., Гаастра В. и Шоулс Л. М. (2002) Идентификация генов, связанных с повторами ДНК у прокариот. Мол. микробиол.
43, 1565–1575 гг.
10.1046/j.1365-2958.2002.02839.x [PubMed] [CrossRef] [Google Scholar]8. Pourcel C., Touchon M., Villeriot N., Vernadet J.-P., Couvin D., Toffano-Nioche C. et al. (2019) CRISPRCasdb — преемник CRISPRdb, содержащий массивы CRISPR и гены cas из полных последовательностей генома, а также инструменты для загрузки и запроса списков повторов и спейсеров.Нуклеиновые Кислоты Res.
48, Д535–Д544
10.1093/nar/gkz915 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]9. Шмаков С.А., Ситник В., Макарова К.С., Вольф Ю.И., Северинов К.В. и Кунин Е.В. (2017) В спейсерном пространстве CRISPR преобладают последовательности видоспецифичных мобиломов. МБио
8, e01397-17
10.1128/mBio.01397-17 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]10. Танг Т. -Х., Башеллери Ж.-П., Рождественский Т., Бортолин М.-Л., Хубер Х., Друнговски М. и соавт. (2002) Идентификация 86 кандидатов на малые РНК, не являющиеся информационными, от археона Archaeoglobus fulgidus .проц. Натл акад. науч. США.
99, 7536–7541
10.1073/pnas.112047299 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]11. Танг Т.-Х., Полачек Н., Живицкий М., Хубер Х., Бруггер К., Гарретт Р. и соавт. (2005) Идентификация новых некодирующих РНК в качестве потенциальных антисмысловых регуляторов у архея Sulfolobus solfataricus . Мол. микробиол.
55, 469–481
10.1111/j.1365-2958.2004.04428.x [PubMed] [CrossRef] [Google Scholar]13. Броунс С.Дж.Дж., Джор М.М., Лундгрен М., Вестра Э.Р., Слейкхус Р.Дж.Х., Снейдерс А.П.Л. и другие. (2008) Малые РНК CRISPR направляют противовирусную защиту у прокариот. Наука
321, 960–964
10.1126/science.1159689 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]15. Карт Дж., Ван Р., Ли Х., Тернс Р.М. и Тернс М.П. (2008) Cas6 представляет собой эндорибонуклеазу, которая генерирует направляющие РНК для защиты от захватчиков у прокариот.
-Х., Башеллери Ж.-П., Рождественский Т., Бортолин М.-Л., Хубер Х., Друнговски М. и соавт. (2002) Идентификация 86 кандидатов на малые РНК, не являющиеся информационными, от археона Archaeoglobus fulgidus .проц. Натл акад. науч. США.
99, 7536–7541
10.1073/pnas.112047299 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]11. Танг Т.-Х., Полачек Н., Живицкий М., Хубер Х., Бруггер К., Гарретт Р. и соавт. (2005) Идентификация новых некодирующих РНК в качестве потенциальных антисмысловых регуляторов у архея Sulfolobus solfataricus . Мол. микробиол.
55, 469–481
10.1111/j.1365-2958.2004.04428.x [PubMed] [CrossRef] [Google Scholar]13. Броунс С.Дж.Дж., Джор М.М., Лундгрен М., Вестра Э.Р., Слейкхус Р.Дж.Х., Снейдерс А.П.Л. и другие. (2008) Малые РНК CRISPR направляют противовирусную защиту у прокариот. Наука
321, 960–964
10.1126/science.1159689 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]15. Карт Дж., Ван Р., Ли Х., Тернс Р.М. и Тернс М.П. (2008) Cas6 представляет собой эндорибонуклеазу, которая генерирует направляющие РНК для защиты от захватчиков у прокариот. Гены. Дев.
22, 3489–3496
10.1101/gad.1742908 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]16. Лиллестёль Р.К., Шах С.А., Брюггер К., Реддер П., Phan H., Christiansen J. et al. (2009) CRISPR-семейства кренархей рода Sulfolobus: двунаправленная транскрипция и динамические свойства. Мол. микробиол.
72, 259–272
10.1111/j.1365-2958.2009.06641.x [PubMed] [CrossRef] [Google Scholar]17. Пул У., Вурм Р., Арслан З., Гейссен Р., Хофманн Н. и Вагнер Р. (2010) Идентификация и характеристика промоторов E. coli CRISPR-Cas и их подавление с помощью H-NS. Мол. микробиол.
75, 1495–1512 гг.
10.1111/j.1365-2958.2010.07073.x [PubMed] [CrossRef] [Google Scholar]18.Hale C.R., Zhao P., Olson S., Duff M.O., Graveley B.R., Wells L. et al. (2009)Расщепление РНК под управлением РНК белковым комплексом CRISPR РНК-Cas. Клетка
139, 945–956
10.1016/j.cell.2009.07.040 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]19. Джинек М., Чилински К., Фонфара И., Хауэр М., Дудна Ю.А. и Charpentier E.
Гены. Дев.
22, 3489–3496
10.1101/gad.1742908 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]16. Лиллестёль Р.К., Шах С.А., Брюггер К., Реддер П., Phan H., Christiansen J. et al. (2009) CRISPR-семейства кренархей рода Sulfolobus: двунаправленная транскрипция и динамические свойства. Мол. микробиол.
72, 259–272
10.1111/j.1365-2958.2009.06641.x [PubMed] [CrossRef] [Google Scholar]17. Пул У., Вурм Р., Арслан З., Гейссен Р., Хофманн Н. и Вагнер Р. (2010) Идентификация и характеристика промоторов E. coli CRISPR-Cas и их подавление с помощью H-NS. Мол. микробиол.
75, 1495–1512 гг.
10.1111/j.1365-2958.2010.07073.x [PubMed] [CrossRef] [Google Scholar]18.Hale C.R., Zhao P., Olson S., Duff M.O., Graveley B.R., Wells L. et al. (2009)Расщепление РНК под управлением РНК белковым комплексом CRISPR РНК-Cas. Клетка
139, 945–956
10.1016/j.cell.2009.07.040 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]19. Джинек М., Чилински К., Фонфара И., Хауэр М., Дудна Ю.А. и Charpentier E. (2012) Программируемая ДНК-эндонуклеаза с двойной РНК-управляемой функцией в адаптивном бактериальном иммунитете. Наука
337, 816–821
10.1126/science.1225829 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]20.Гасюнас Г., Баррангоу Р., Хорват П. и Сикснис В. (2012) Рибонуклеопротеиновый комплекс Cas9-crРНК опосредует специфическое расщепление ДНК для адаптивного иммунитета у бактерий. проц. Натл. акад. науч. США.
109, Е2579–Е2586
10.1073/pnas.1208507109 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]21. Дево Х., Баррангу Р., Гарно Дж. Э., Лабонте Дж., Фремо К., Бояваль П. и др. (2008) Фаговый ответ на закодированную CRISPR устойчивость у Streptococcus thermophilus . Дж. Бактериол.
190, 1390–1400
10.1128/JB.01412-07 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]22. Хорват П., Ромеро Д.А., Куте-Монвуазен А.-К., Ричардс М., Дево Х., Муано С. и др. (2008) Разнообразие, активность и эволюция локусов CRISPR у Streptococcus thermophilus .
(2012) Программируемая ДНК-эндонуклеаза с двойной РНК-управляемой функцией в адаптивном бактериальном иммунитете. Наука
337, 816–821
10.1126/science.1225829 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]20.Гасюнас Г., Баррангоу Р., Хорват П. и Сикснис В. (2012) Рибонуклеопротеиновый комплекс Cas9-crРНК опосредует специфическое расщепление ДНК для адаптивного иммунитета у бактерий. проц. Натл. акад. науч. США.
109, Е2579–Е2586
10.1073/pnas.1208507109 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]21. Дево Х., Баррангу Р., Гарно Дж. Э., Лабонте Дж., Фремо К., Бояваль П. и др. (2008) Фаговый ответ на закодированную CRISPR устойчивость у Streptococcus thermophilus . Дж. Бактериол.
190, 1390–1400
10.1128/JB.01412-07 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]22. Хорват П., Ромеро Д.А., Куте-Монвуазен А.-К., Ричардс М., Дево Х., Муано С. и др. (2008) Разнообразие, активность и эволюция локусов CRISPR у Streptococcus thermophilus . Дж. Бактериол.
190, 1401–1412 гг.
10.1128/JB.01415-07 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]23. Йосеф И., Горен М.Г. и Qimron U. (2012) Белки и элементы ДНК, необходимые для процесса адаптации CRISPR в Escherichia coli .Нуклеиновые Кислоты Res.
40, 5569–5576
10.1093/nar/gks216 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]24. Нуньес Дж.К., Ли А.С.Ю., Энгельман А. и Дудна Дж.А. (2015)Опосредованное интегразой приобретение спейсера во время адаптивного иммунитета CRISPR-Cas. Природа
519, 193–198
10.1038/nature14237 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]25. Сяо Ю., Нг С., Нам К.Х. и Ке А. (2017) Как CRISPR-Cas типа II создают иммунитет посредством интеграции спейсеров, опосредованной Cas1-Cas2. Природа
550, 137–141
10.1038/nature24020 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]26.Вэй Ю., Тернс Р.М. и Тернс М.П. (2015)Функция Cas9 и выборка генома хозяина в адаптации CRISPR-Cas типа II-A. Гены Дев.
29, 356–361
10.
Дж. Бактериол.
190, 1401–1412 гг.
10.1128/JB.01415-07 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]23. Йосеф И., Горен М.Г. и Qimron U. (2012) Белки и элементы ДНК, необходимые для процесса адаптации CRISPR в Escherichia coli .Нуклеиновые Кислоты Res.
40, 5569–5576
10.1093/nar/gks216 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]24. Нуньес Дж.К., Ли А.С.Ю., Энгельман А. и Дудна Дж.А. (2015)Опосредованное интегразой приобретение спейсера во время адаптивного иммунитета CRISPR-Cas. Природа
519, 193–198
10.1038/nature14237 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]25. Сяо Ю., Нг С., Нам К.Х. и Ке А. (2017) Как CRISPR-Cas типа II создают иммунитет посредством интеграции спейсеров, опосредованной Cas1-Cas2. Природа
550, 137–141
10.1038/nature24020 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]26.Вэй Ю., Тернс Р.М. и Тернс М.П. (2015)Функция Cas9 и выборка генома хозяина в адаптации CRISPR-Cas типа II-A. Гены Дев.
29, 356–361
10. 1101/gad.257550.114 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]27. Стерн А., Керен Л., Вуртцель О., Амитай Г. и Сорек Р. (2010) Самонацеливание с помощью CRISPR: генная регуляция или аутоиммунитет?
Тенденции Жене.
26, 335–340
10.1016/j.tig.2010.05.008 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]28. Эдгар Р. и Кимрон У. (2010) Система Escherichia coli CRISPR защищает от λ-лизогенизации, лизогенов и индукции профагов.Дж. Бактериол.
192, 6291–6294
10.1128/JB.00644-10 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]29. Верко Р.Б., Чанг Дж.Т., Дай Р.Л., Тейлор С., Гриствуд Т., Клулоу Дж.С. и другие. (2013) Цитотоксическое нацеливание на хромосомы с помощью систем CRISPR/Cas может изменять форму бактериальных геномов и удалять или реконструировать островки патогенности. Генетика PLoS.
9, е1003454
10.1371/journal.pgen.1003454 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]30. Свортс Д.К., Мостерд К., ван Пассел М.В.Дж. и Браунс С.
1101/gad.257550.114 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]27. Стерн А., Керен Л., Вуртцель О., Амитай Г. и Сорек Р. (2010) Самонацеливание с помощью CRISPR: генная регуляция или аутоиммунитет?
Тенденции Жене.
26, 335–340
10.1016/j.tig.2010.05.008 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]28. Эдгар Р. и Кимрон У. (2010) Система Escherichia coli CRISPR защищает от λ-лизогенизации, лизогенов и индукции профагов.Дж. Бактериол.
192, 6291–6294
10.1128/JB.00644-10 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]29. Верко Р.Б., Чанг Дж.Т., Дай Р.Л., Тейлор С., Гриствуд Т., Клулоу Дж.С. и другие. (2013) Цитотоксическое нацеливание на хромосомы с помощью систем CRISPR/Cas может изменять форму бактериальных геномов и удалять или реконструировать островки патогенности. Генетика PLoS.
9, е1003454
10.1371/journal.pgen.1003454 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]30. Свортс Д.К., Мостерд К., ван Пассел М.В.Дж. и Браунс С. Дж.Дж. (2012)Интерференция CRISPR направляет приобретение спейсера, специфичного для нити.ПЛОС ОДИН
7, е35888
10.1371/journal.pone.0035888 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]31. Даценко К.А., Пугач К., Тихонов А., Ваннер Б.Л., Северинов К., Семенова Е. (2012) Молекулярная память о предшествующих инфекциях активирует систему адаптивного бактериального иммунитета CRISPR/Cas. Нац. коммун.
3 945
10.1038/ncomms1937 [PubMed] [CrossRef] [Google Scholar]32. van der Ploeg J.R. (2009) Анализ CRISPR в Streptococcus mutans предполагает частое возникновение приобретенного иммунитета против инфекции M102-подобными бактериофагами.микробиология
155, 1966–1976 гг.
10.1099/mic.0.027508-0 [PubMed] [CrossRef] [Google Scholar]33. Garneau J.E., Dupuis M.-E., Villion M., Romero D.A., Barrangou R., Boyaval P. et al. (2010) Бактериальная иммунная система CRISPR/Cas расщепляет ДНК бактериофага и плазмиды. Природа
468, 67–71
10.1038/nature09523 [PubMed] [CrossRef] [Google Scholar]34.
Дж.Дж. (2012)Интерференция CRISPR направляет приобретение спейсера, специфичного для нити.ПЛОС ОДИН
7, е35888
10.1371/journal.pone.0035888 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]31. Даценко К.А., Пугач К., Тихонов А., Ваннер Б.Л., Северинов К., Семенова Е. (2012) Молекулярная память о предшествующих инфекциях активирует систему адаптивного бактериального иммунитета CRISPR/Cas. Нац. коммун.
3 945
10.1038/ncomms1937 [PubMed] [CrossRef] [Google Scholar]32. van der Ploeg J.R. (2009) Анализ CRISPR в Streptococcus mutans предполагает частое возникновение приобретенного иммунитета против инфекции M102-подобными бактериофагами.микробиология
155, 1966–1976 гг.
10.1099/mic.0.027508-0 [PubMed] [CrossRef] [Google Scholar]33. Garneau J.E., Dupuis M.-E., Villion M., Romero D.A., Barrangou R., Boyaval P. et al. (2010) Бактериальная иммунная система CRISPR/Cas расщепляет ДНК бактериофага и плазмиды. Природа
468, 67–71
10.1038/nature09523 [PubMed] [CrossRef] [Google Scholar]34. Леви А., Горен М.Г., Йосеф И., Остер О., Манор М., Амитай Г. и др. (2015) Предвзятость адаптации CRISPR объясняет предпочтение приобретения чужой ДНК. Природа
520, 505–510
10.1038/nature14302 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]35. Шиимори М., Гаррет С.С., Чемберс Д.П., Гловер К.В.К. III, Гравли Б.Р. и Тернс М.П. (2017) Роль свободных концов ДНК и мотивов, прилегающих к протоспейсеру, в поглощении ДНК CRISPR у Pyrococcus furiosus . Нуклеиновые Кислоты Res.
45, 11281–11294
10.1093/nar/gkx839 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]36. Диес-Вильясеньор К., Гусман Н.М., Альмендрос К., Гарсия-Мартинес Х. и Мохика Ф.Х.М. (2013) Репортерные плазмиды интеграции CRISPR-спейсера обнаруживают отчетливую подлинную специфичность приобретения среди вариантов CRISPR-Cas IE Escherichia coli .РНК биол.
10, 792–802
10.4161/rna.24023 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]37. Амлингер Л., Хёкзема М., Вагнер Э.Г.Х., Коскиниеми С.
Леви А., Горен М.Г., Йосеф И., Остер О., Манор М., Амитай Г. и др. (2015) Предвзятость адаптации CRISPR объясняет предпочтение приобретения чужой ДНК. Природа
520, 505–510
10.1038/nature14302 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]35. Шиимори М., Гаррет С.С., Чемберс Д.П., Гловер К.В.К. III, Гравли Б.Р. и Тернс М.П. (2017) Роль свободных концов ДНК и мотивов, прилегающих к протоспейсеру, в поглощении ДНК CRISPR у Pyrococcus furiosus . Нуклеиновые Кислоты Res.
45, 11281–11294
10.1093/nar/gkx839 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]36. Диес-Вильясеньор К., Гусман Н.М., Альмендрос К., Гарсия-Мартинес Х. и Мохика Ф.Х.М. (2013) Репортерные плазмиды интеграции CRISPR-спейсера обнаруживают отчетливую подлинную специфичность приобретения среди вариантов CRISPR-Cas IE Escherichia coli .РНК биол.
10, 792–802
10.4161/rna.24023 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]37. Амлингер Л., Хёкзема М., Вагнер Э.Г.Х., Коскиниеми С. и Лундгрен М. (2017) Флуоресцентный репортер адаптации CRISPR для быстрой количественной оценки приобретения спейсера. науч. Респ.
7, 10392
10.1038/s41598-017-10876-z [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]38. Паес-Эспино Д., Морович В., Сан К.Л., Томас Б.К., Уеда К.-И., Шталь Б. и соавт. (2013) Сильная предвзятость в бактериальных элементах CRISPR, которые обеспечивают иммунитет к фагам.Нац. коммун.
4, 1430
10.1038/ncomms2440 [PubMed] [CrossRef] [Google Scholar]39. Савицкая Е., Семенова Е., Дедков В., Метлицкая А. и Северинов К. (2013) Высокопроизводительный анализ приобретения спейсера CRISPR/Cas типа I-E в E. coli . РНК биол.
10, 716–725
10.4161/rna.24325 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]40. Йосеф И., Шитрит Д., Горен М.Г., Бурштейн Д., Пупко Т. и Кимрон У. (2013) Мотивы ДНК, определяющие эффективность адаптации в массиве Escherichia coli CRISPR.проц. Натл акад. науч. США.
110, 14396–41401
10.1073/pnas.1300108110 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]41.
и Лундгрен М. (2017) Флуоресцентный репортер адаптации CRISPR для быстрой количественной оценки приобретения спейсера. науч. Респ.
7, 10392
10.1038/s41598-017-10876-z [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]38. Паес-Эспино Д., Морович В., Сан К.Л., Томас Б.К., Уеда К.-И., Шталь Б. и соавт. (2013) Сильная предвзятость в бактериальных элементах CRISPR, которые обеспечивают иммунитет к фагам.Нац. коммун.
4, 1430
10.1038/ncomms2440 [PubMed] [CrossRef] [Google Scholar]39. Савицкая Е., Семенова Е., Дедков В., Метлицкая А. и Северинов К. (2013) Высокопроизводительный анализ приобретения спейсера CRISPR/Cas типа I-E в E. coli . РНК биол.
10, 716–725
10.4161/rna.24325 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]40. Йосеф И., Шитрит Д., Горен М.Г., Бурштейн Д., Пупко Т. и Кимрон У. (2013) Мотивы ДНК, определяющие эффективность адаптации в массиве Escherichia coli CRISPR.проц. Натл акад. науч. США.
110, 14396–41401
10.1073/pnas.1300108110 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]41. Шмаков С., Савицкая Е., Семенова Е., Логачева М.Д., Даценко К.А. и Северинов К. (2014) Повсеместное образование противоположно ориентированных спейсеров во время адаптации CRISPR. Нуклеиновые Кислоты Res.
42, 5907–5916
10.1093/nar/gku226 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]42. Хелер Р., Самаи П., Моделл Дж.В., Вайнер К., Голдберг Г.В., Бикард Д. и др. (2015) Cas9 определяет функциональные вирусные мишени во время адаптации CRISPR-Cas.Природа
519, 199–202
10.1038/nature14245 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]43. Воронцова Д., Даценко К.А., Медведева С., Бонди-Деномы Дж., Савицкая Е.Е., Пугач К. и соавт. (2015) Приобретение чужеродной ДНК с помощью системы IF CRISPR-Cas требует наличия всех компонентов механизма интерференции. Нуклеиновые Кислоты Res.
43, 10848–10860
10.1093/nar/gkv1261 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]44. Сайлас С., Мор Г., Сидот Д.Дж., Маркхэм Л.М., Санчес-Амат А., Бхая Д. и др.(2016)Прямое приобретение спейсера CRISPR из РНК с помощью естественного слитого белка обратной транскриптазы-Cas1.
Шмаков С., Савицкая Е., Семенова Е., Логачева М.Д., Даценко К.А. и Северинов К. (2014) Повсеместное образование противоположно ориентированных спейсеров во время адаптации CRISPR. Нуклеиновые Кислоты Res.
42, 5907–5916
10.1093/nar/gku226 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]42. Хелер Р., Самаи П., Моделл Дж.В., Вайнер К., Голдберг Г.В., Бикард Д. и др. (2015) Cas9 определяет функциональные вирусные мишени во время адаптации CRISPR-Cas.Природа
519, 199–202
10.1038/nature14245 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]43. Воронцова Д., Даценко К.А., Медведева С., Бонди-Деномы Дж., Савицкая Е.Е., Пугач К. и соавт. (2015) Приобретение чужеродной ДНК с помощью системы IF CRISPR-Cas требует наличия всех компонентов механизма интерференции. Нуклеиновые Кислоты Res.
43, 10848–10860
10.1093/nar/gkv1261 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]44. Сайлас С., Мор Г., Сидот Д.Дж., Маркхэм Л.М., Санчес-Амат А., Бхая Д. и др.(2016)Прямое приобретение спейсера CRISPR из РНК с помощью естественного слитого белка обратной транскриптазы-Cas1. Наука
351, ад4234
10.1126/science.aad4234 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]45. Стаалс Р.Х.Дж., Джексон С.А., Бисвас А., Браунс С.Дж.Дж., Браун С.М. и Финеран П.К. (2016) Приобретение спейсеров, управляемое интерференцией, доминирует над наивной и подготовленной адаптацией в нативной системе CRISPR-Cas. Нац. коммун.
7, 12853
10.1038/ncomms12853 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]46.Хелер Р., Райт А.В., Вучеля М., Бикард Д., Дудна Дж.А. и Marraffini L.A. (2017) Мутации в Cas9 повышают скорость приобретения последовательностей вирусных спейсеров во время иммунного ответа CRISPR-Cas. Мол. Клетка
65, 168–175
10.1016/j.molcel.2016.11.031 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]47. Li M., Gong L., Zhao D., Zhou J. and Xiang H. (2017) Размер спейсера IB CRISPR модулируется терминальной последовательностью протоспейсера. Нуклеиновые Кислоты Res.
45, 4642–4654
10.1093/nar/gkx229 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]48.
Наука
351, ад4234
10.1126/science.aad4234 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]45. Стаалс Р.Х.Дж., Джексон С.А., Бисвас А., Браунс С.Дж.Дж., Браун С.М. и Финеран П.К. (2016) Приобретение спейсеров, управляемое интерференцией, доминирует над наивной и подготовленной адаптацией в нативной системе CRISPR-Cas. Нац. коммун.
7, 12853
10.1038/ncomms12853 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]46.Хелер Р., Райт А.В., Вучеля М., Бикард Д., Дудна Дж.А. и Marraffini L.A. (2017) Мутации в Cas9 повышают скорость приобретения последовательностей вирусных спейсеров во время иммунного ответа CRISPR-Cas. Мол. Клетка
65, 168–175
10.1016/j.molcel.2016.11.031 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]47. Li M., Gong L., Zhao D., Zhou J. and Xiang H. (2017) Размер спейсера IB CRISPR модулируется терминальной последовательностью протоспейсера. Нуклеиновые Кислоты Res.
45, 4642–4654
10.1093/nar/gkx229 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]48. Рао К., Чин Д. и Энсмингер А.В. (2017) Прайминг в разрешительной системе CRISPR-Cas типа I-C выявляет отчетливую динамику приобретения и потери спейсеров. РНК
23, 1525–1538 гг.
10.1261/rna.062083.117 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]49. Мушарова О., Выговский Д., Медведева С., Гузина Ю., Житнюк Ю., Джорджевич М. и др. (2018) Избегание тринуклеотида, соответствующего мотиву, соседнему с консенсусным протоспейсером, контролирует эффективность отбора преспейсера во время адаптации с праймированием. МБио
9, 02169-18
10.1128/mBio.02169-18 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]50. Кипер С.Н., Альмендрос С., Белер Дж., Маккензи Р.Э., Нобрега Ф.Л., Хаагсма А.С. и соавт. (2018) Cas4 облегчает выбор PAM-совместимого спейсера во время адаптации CRISPR. Сотовый представитель
22, 3377–3384
10.1016/j.celrep.2018.02.103 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]51. Шиимори М., Гаррет С.С., Гравли Б.Р. и Тернс М.П. (2018)Нуклеазы Cas4 определяют PAM, длину и ориентацию фрагментов ДНК, интегрированных в локусы CRISPR.
Рао К., Чин Д. и Энсмингер А.В. (2017) Прайминг в разрешительной системе CRISPR-Cas типа I-C выявляет отчетливую динамику приобретения и потери спейсеров. РНК
23, 1525–1538 гг.
10.1261/rna.062083.117 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]49. Мушарова О., Выговский Д., Медведева С., Гузина Ю., Житнюк Ю., Джорджевич М. и др. (2018) Избегание тринуклеотида, соответствующего мотиву, соседнему с консенсусным протоспейсером, контролирует эффективность отбора преспейсера во время адаптации с праймированием. МБио
9, 02169-18
10.1128/mBio.02169-18 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]50. Кипер С.Н., Альмендрос С., Белер Дж., Маккензи Р.Э., Нобрега Ф.Л., Хаагсма А.С. и соавт. (2018) Cas4 облегчает выбор PAM-совместимого спейсера во время адаптации CRISPR. Сотовый представитель
22, 3377–3384
10.1016/j.celrep.2018.02.103 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]51. Шиимори М., Гаррет С.С., Гравли Б.Р. и Тернс М.П. (2018)Нуклеазы Cas4 определяют PAM, длину и ориентацию фрагментов ДНК, интегрированных в локусы CRISPR. Мол. Клетка
70, 814–824
10.1016/j.molcel.2018.05.002 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]52. Альмендрос К., Нобрега Ф.Л., Маккензи Р.Э. и Браунс С.Дж.Дж. (2019) Слияния Cas4-Cas1 обеспечивают эффективный выбор PAM и контролируют адаптацию CRISPR. Нуклеиновые Кислоты Res.
47, 5223–5230
10.1093/nar/gkz217 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]53. Паес-Эспино Д., Шарон И., Морович В., Шталь Б., Томас Б.С., Баррангоу Р. и др. (2015) Иммунитет CRISPR стимулирует быструю эволюцию фагового генома у Streptococcus thermophilus .МБио
6, 00262-15
10.1128/mBio.00262-15 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]54. Нивала Дж., Шипман С.Л. и Черч Г.М. (2018)Спонтанная генерация локусов CRISPR in vivo путем неканонической интеграции спейсера. Нац. микробиол.
3, 310–318
10.1038/s41564-017-0097-z [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]55. Джексон С.А., Биркхольц Н., Мэлоун Л.М. и Финеран П.К. (2019) Неточное приобретение спейсера создает иммунное разнообразие CRISPR-Cas за счет праймированной адаптации.
Мол. Клетка
70, 814–824
10.1016/j.molcel.2018.05.002 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]52. Альмендрос К., Нобрега Ф.Л., Маккензи Р.Э. и Браунс С.Дж.Дж. (2019) Слияния Cas4-Cas1 обеспечивают эффективный выбор PAM и контролируют адаптацию CRISPR. Нуклеиновые Кислоты Res.
47, 5223–5230
10.1093/nar/gkz217 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]53. Паес-Эспино Д., Шарон И., Морович В., Шталь Б., Томас Б.С., Баррангоу Р. и др. (2015) Иммунитет CRISPR стимулирует быструю эволюцию фагового генома у Streptococcus thermophilus .МБио
6, 00262-15
10.1128/mBio.00262-15 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]54. Нивала Дж., Шипман С.Л. и Черч Г.М. (2018)Спонтанная генерация локусов CRISPR in vivo путем неканонической интеграции спейсера. Нац. микробиол.
3, 310–318
10.1038/s41564-017-0097-z [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]55. Джексон С.А., Биркхольц Н., Мэлоун Л.М. и Финеран П.К. (2019) Неточное приобретение спейсера создает иммунное разнообразие CRISPR-Cas за счет праймированной адаптации. Клеточный микроб-хозяин
25, 250–260
10.1016/j.chom.2018.12.014 [PubMed] [CrossRef] [Google Scholar]56. Кривой А., Руткаускас М., Кузнеделов К., Мушарова О., Руйон К., Северинов К. и др. (2018) Адаптация праймированного CRISPR в клетках Escherichia coli не зависит от конформационных изменений каскадного эффекторного комплекса, обнаруженных in vitro. Нуклеиновые Кислоты Res.
46, 4087–4098
10.1093/nar/gky219 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]57. Шипман С.Л., Нивала Дж., Маклис Дж.Д. и Черч Г.М. (2016) Молекулярные записи путем направленного приобретения спейсера CRISPR.Наука
353, ааф1175
10.1126/science.aaf1175 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]58. Маккензи Р.Э., Альмендрос С., Винк Дж.Н.А. и Браунс С.Дж.Дж. (2019) Использование CAPTURE для обнаружения приобретения спейсера в исходных массивах CRISPR. Нац. протокол
14, 976–990
10.1038/s41596-018-0123-5 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]59. Шмидт Ф., Черепкова М.
Клеточный микроб-хозяин
25, 250–260
10.1016/j.chom.2018.12.014 [PubMed] [CrossRef] [Google Scholar]56. Кривой А., Руткаускас М., Кузнеделов К., Мушарова О., Руйон К., Северинов К. и др. (2018) Адаптация праймированного CRISPR в клетках Escherichia coli не зависит от конформационных изменений каскадного эффекторного комплекса, обнаруженных in vitro. Нуклеиновые Кислоты Res.
46, 4087–4098
10.1093/nar/gky219 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]57. Шипман С.Л., Нивала Дж., Маклис Дж.Д. и Черч Г.М. (2016) Молекулярные записи путем направленного приобретения спейсера CRISPR.Наука
353, ааф1175
10.1126/science.aaf1175 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]58. Маккензи Р.Э., Альмендрос С., Винк Дж.Н.А. и Браунс С.Дж.Дж. (2019) Использование CAPTURE для обнаружения приобретения спейсера в исходных массивах CRISPR. Нац. протокол
14, 976–990
10.1038/s41596-018-0123-5 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]59. Шмидт Ф., Черепкова М. Ю. и Платт Р.Дж. (2018) Запись транскрипции путем получения спейсера CRISPR из РНК. Природа
562, 380–385
10.1038/s41586-018-0569-1 [PubMed] [CrossRef] [Google Scholar]60.Семенова Е., Кузнеделов К., Даценко К.А., Будри П.М., Савицкая Е.Е., Медведева С. и др. (2015) Рибонуклеаза Cas6e не требуется для интерференции и адаптации системой E. coli типа I-E CRISPR-Cas. Нуклеиновые Кислоты Res.
43, 6049–6061
10.1093/nar/gkv546 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]61. Семенова Е., Савицкая Е., Мушарова О., Строцкая А., Воронцова Д., Даценко К.А. и другие. (2016) Высокоэффективное получение первичных спейсеров из целей, уничтоженных интерферирующим комплексом Escherichia coli типа IE CRISPR-Cas.проц. Натл акад. науч. США.
113, 7626–7631
10.1073/pnas.1602639113 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]62. Кузнеделов К., Меклер В., Лемак С., Токмина-Лукашевская М., Даценко К.А., Джайн И. и др. (2016) Измененная стехиометрия Каскадные комплексы Escherichia coli с укороченными спейсерами CRISPR РНК способны к интерференции и первичной адаптации.
Ю. и Платт Р.Дж. (2018) Запись транскрипции путем получения спейсера CRISPR из РНК. Природа
562, 380–385
10.1038/s41586-018-0569-1 [PubMed] [CrossRef] [Google Scholar]60.Семенова Е., Кузнеделов К., Даценко К.А., Будри П.М., Савицкая Е.Е., Медведева С. и др. (2015) Рибонуклеаза Cas6e не требуется для интерференции и адаптации системой E. coli типа I-E CRISPR-Cas. Нуклеиновые Кислоты Res.
43, 6049–6061
10.1093/nar/gkv546 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]61. Семенова Е., Савицкая Е., Мушарова О., Строцкая А., Воронцова Д., Даценко К.А. и другие. (2016) Высокоэффективное получение первичных спейсеров из целей, уничтоженных интерферирующим комплексом Escherichia coli типа IE CRISPR-Cas.проц. Натл акад. науч. США.
113, 7626–7631
10.1073/pnas.1602639113 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]62. Кузнеделов К., Меклер В., Лемак С., Токмина-Лукашевская М., Даценко К.А., Джайн И. и др. (2016) Измененная стехиометрия Каскадные комплексы Escherichia coli с укороченными спейсерами CRISPR РНК способны к интерференции и первичной адаптации. Нуклеиновые Кислоты Res.
44, 10849–10861
10.1093/nar/gkw914 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]63. Мушарова О., Климук Е., Даценко К.А., Метлицкая А., Логачева М., Семенова Е. и др. (2017) Промежуточные соединения ДНК длиной спейсера связаны с Cas1 в клетках, подвергающихся первичной адаптации CRISPR. Нуклеиновые Кислоты Res.
45, 3297–3307
10.1093/nar/gkx097 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]64. Мушарова О., Ситник В., Влот М., Савицкая Е., Даценко К.А., Кривой А. и др. (2019)Систематический анализ способности последовательностей CRISPR-Cas PAM типа IE Escherichia coli способствовать интерференции и первичной адаптации.Мол. микробиол.
111, 1558–1570 гг.
10.1111/mmi.14237 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]65. Строцкая А., Семенова Е., Савицкая Е. и Северинов К. (2015) Быстрое мультиплексное создание штаммов Escherichia coli , способных препятствовать заражению фагом через CRISPR. Методы Мол.
Нуклеиновые Кислоты Res.
44, 10849–10861
10.1093/nar/gkw914 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]63. Мушарова О., Климук Е., Даценко К.А., Метлицкая А., Логачева М., Семенова Е. и др. (2017) Промежуточные соединения ДНК длиной спейсера связаны с Cas1 в клетках, подвергающихся первичной адаптации CRISPR. Нуклеиновые Кислоты Res.
45, 3297–3307
10.1093/nar/gkx097 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]64. Мушарова О., Ситник В., Влот М., Савицкая Е., Даценко К.А., Кривой А. и др. (2019)Систематический анализ способности последовательностей CRISPR-Cas PAM типа IE Escherichia coli способствовать интерференции и первичной адаптации.Мол. микробиол.
111, 1558–1570 гг.
10.1111/mmi.14237 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]65. Строцкая А., Семенова Е., Савицкая Е. и Северинов К. (2015) Быстрое мультиплексное создание штаммов Escherichia coli , способных препятствовать заражению фагом через CRISPR. Методы Мол. биол.
1311, 147–159
10.1007/978-1-4939-2687-9_9 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]66. Зиберт П.Д., Ченчик А., Келлог Д.Э., Лукьянов К.А. и Лукьянов С.А. (1995) Усовершенствованный метод ПЦР для хождения по неклонированной геномной ДНК.Нуклеиновые Кислоты Res.
23, 1087–1088 гг.
10.1093/nar/23.6.1087 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]67. Дьяченко Л., Лау Ю.Ф., Кэмпбелл А.П., Ченчик А., Мокадам Ф., Хуанг Б. и др. (1996) Подавление субтрактивной гибридизации: метод создания дифференциально регулируемых или тканеспецифичных зондов и библиотек кДНК. проц. Натл акад. науч. США.
93, 6025–6030
10.1073/pnas.93.12.6025 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]68. Шагин Д.А., Лукьянов К.А., Вагнер Л.Л. и Мац М.В. (1999) Регулирование средней длины сложного продукта ПЦР. Нуклеиновые Кислоты Res.
27, е23
10.1093/nar/27.18.e23 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]69. Иванчич-Баче И., Касс С.Д., Верн С.Дж. и Болт Э.
биол.
1311, 147–159
10.1007/978-1-4939-2687-9_9 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]66. Зиберт П.Д., Ченчик А., Келлог Д.Э., Лукьянов К.А. и Лукьянов С.А. (1995) Усовершенствованный метод ПЦР для хождения по неклонированной геномной ДНК.Нуклеиновые Кислоты Res.
23, 1087–1088 гг.
10.1093/nar/23.6.1087 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]67. Дьяченко Л., Лау Ю.Ф., Кэмпбелл А.П., Ченчик А., Мокадам Ф., Хуанг Б. и др. (1996) Подавление субтрактивной гибридизации: метод создания дифференциально регулируемых или тканеспецифичных зондов и библиотек кДНК. проц. Натл акад. науч. США.
93, 6025–6030
10.1073/pnas.93.12.6025 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]68. Шагин Д.А., Лукьянов К.А., Вагнер Л.Л. и Мац М.В. (1999) Регулирование средней длины сложного продукта ПЦР. Нуклеиновые Кислоты Res.
27, е23
10.1093/nar/27.18.e23 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]69. Иванчич-Баче И., Касс С.Д., Верн С.Дж. и Болт Э. Л. (2015) Различные белки стабильности генома лежат в основе примированной и наивной адаптации в иммунитете E. coli CRISPR-Cas. Нуклеиновые Кислоты Res.
43, 10821–10830
10.1093/nar/gkv1213 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]70. Фагерлунд Р.Д., Уилкинсон М.Е., Клыков О., Барендрегт А., Пирс Ф.Г., Кипер С.Н. и другие. (2017)Захват и интеграция спейсера с помощью адаптационного комплекса CRISPR типа IF Cas1-Cas2-3. проц. Натл акад. науч. США.
114, Е5122–Е5128
10.1073/pnas.1618421114 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]71. Райт А.В., Лю Дж.-Дж., Нотт Г.Дж., Доксзен К.В., Ногалес Э. и Дудна Дж.А. (2017) Структуры комплекса интеграции генома CRISPR. Наука
357, 1113–1118
10.1126/science.aao0679 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]72.Ли Х., Чжоу Ю., Тейлор Д.В. и Сашитал Д.Г. (2018) Cas4-зависимая обработка предспейсеров обеспечивает высокоточное программирование массивов CRISPR. Мол. Клетка
70, 48–59
10.1016/j.molcel.2018.
Л. (2015) Различные белки стабильности генома лежат в основе примированной и наивной адаптации в иммунитете E. coli CRISPR-Cas. Нуклеиновые Кислоты Res.
43, 10821–10830
10.1093/nar/gkv1213 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]70. Фагерлунд Р.Д., Уилкинсон М.Е., Клыков О., Барендрегт А., Пирс Ф.Г., Кипер С.Н. и другие. (2017)Захват и интеграция спейсера с помощью адаптационного комплекса CRISPR типа IF Cas1-Cas2-3. проц. Натл акад. науч. США.
114, Е5122–Е5128
10.1073/pnas.1618421114 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]71. Райт А.В., Лю Дж.-Дж., Нотт Г.Дж., Доксзен К.В., Ногалес Э. и Дудна Дж.А. (2017) Структуры комплекса интеграции генома CRISPR. Наука
357, 1113–1118
10.1126/science.aao0679 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]72.Ли Х., Чжоу Ю., Тейлор Д.В. и Сашитал Д.Г. (2018) Cas4-зависимая обработка предспейсеров обеспечивает высокоточное программирование массивов CRISPR. Мол. Клетка
70, 48–59
10.1016/j.molcel.2018. 03.003 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]73. Драбавичюс Г., Синкунас Т., Силанскас А., Гасюнас Г., Венцловас Ч. и Siksnys V. (2018) Экзонуклеазоподобный домен Dnaq Cas2 способствует интеграции спейсера в систему CRISPR-Cas типа I-E. Представитель ЕМБО
19, е45543
10.15252/embr.201745543 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]74.Радовчич М., Киллелеа Т., Савицкая Э., Веттштейн Л., Болт Э.Л. и Иванчич-Баче И. (2018) Адаптация CRISPR-cas в Escherichia coli требует активности хеликазы RecBCD, но не нуклеазной активности, не зависит от гомологичной рекомбинации и противодействует экзонуклеазам 5′-оцДНК. Нуклеиновые Кислоты Res.
46, 10173–10183
10.1093/nar/gky799 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]75. Уилкинсон М., Драбавичюс Г., Силанскас А., Гасюнас Г., Сикснис В. и Вигли Д.Б. (2019)Структура ДНК-связанного комплекса захвата спейсера системы CRISPR-Cas типа II.Мол. Клетка
75, 90–101
10.1016/j.molcel.2019.04.020 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]76.
03.003 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]73. Драбавичюс Г., Синкунас Т., Силанскас А., Гасюнас Г., Венцловас Ч. и Siksnys V. (2018) Экзонуклеазоподобный домен Dnaq Cas2 способствует интеграции спейсера в систему CRISPR-Cas типа I-E. Представитель ЕМБО
19, е45543
10.15252/embr.201745543 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]74.Радовчич М., Киллелеа Т., Савицкая Э., Веттштейн Л., Болт Э.Л. и Иванчич-Баче И. (2018) Адаптация CRISPR-cas в Escherichia coli требует активности хеликазы RecBCD, но не нуклеазной активности, не зависит от гомологичной рекомбинации и противодействует экзонуклеазам 5′-оцДНК. Нуклеиновые Кислоты Res.
46, 10173–10183
10.1093/nar/gky799 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]75. Уилкинсон М., Драбавичюс Г., Силанскас А., Гасюнас Г., Сикснис В. и Вигли Д.Б. (2019)Структура ДНК-связанного комплекса захвата спейсера системы CRISPR-Cas типа II.Мол. Клетка
75, 90–101
10.1016/j.molcel.2019.04.020 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]76. Ван Дж., Ли Дж., Чжао Х., Шэн Г., Ван М., Инь М. и др. (2015) Структурная и механистическая основа приобретения PAM-зависимого спейсера в системах CRISPR-Cas. Клетка
163, 840–853
10.1016/j.cell.2015.10.008 [PubMed] [CrossRef] [Google Scholar]77. Ролли С., Грэм С., Руйон С. и Уайт М.Ф. (2018) Обработка предспейсеров и конкретная интеграция в систему CRISPR типа IA. Нуклеиновые Кислоты Res.46, 1007–1020 гг.
10.1093/nar/gkx1232 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]78. Moch C., Fromant M., Blanquet S. and Plateau P. (2017) Специфика связывания ДНК интегразы Cas1-Cas2 Escherichia coli определяет ее рекрутирование в локусе CRISPR. Нуклеиновые Кислоты Res.
45, 2714–2723
10.1093/nar/gkw1309 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]79. Ширяева А.А., Савицкая Е., Даценко К.А., Введенская И.О., Федорова И., Морозова Н. и др. (2019)Обнаружение предшественников спейсеров, образующихся in vivo во время адаптации CRISPR с праймером.Нац. коммун.
10, 4603
10.
Ван Дж., Ли Дж., Чжао Х., Шэн Г., Ван М., Инь М. и др. (2015) Структурная и механистическая основа приобретения PAM-зависимого спейсера в системах CRISPR-Cas. Клетка
163, 840–853
10.1016/j.cell.2015.10.008 [PubMed] [CrossRef] [Google Scholar]77. Ролли С., Грэм С., Руйон С. и Уайт М.Ф. (2018) Обработка предспейсеров и конкретная интеграция в систему CRISPR типа IA. Нуклеиновые Кислоты Res.46, 1007–1020 гг.
10.1093/nar/gkx1232 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]78. Moch C., Fromant M., Blanquet S. and Plateau P. (2017) Специфика связывания ДНК интегразы Cas1-Cas2 Escherichia coli определяет ее рекрутирование в локусе CRISPR. Нуклеиновые Кислоты Res.
45, 2714–2723
10.1093/nar/gkw1309 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]79. Ширяева А.А., Савицкая Е., Даценко К.А., Введенская И.О., Федорова И., Морозова Н. и др. (2019)Обнаружение предшественников спейсеров, образующихся in vivo во время адаптации CRISPR с праймером.Нац. коммун.
10, 4603
10.